Embed Size (px)
Citation preview

BIOLOGY OF REPRODUCTION 58, 670-677 (1998)
Protein C Inhibitor May Modulate Human Sperm-Oocyte Interactions'
Marc G.L.M. Elisen, 2,3,6 Roelof J. van Kooij,4 Martijn A. Nolte,3 J. Arnoud Marquart, 3
Tycho M.T.W.T. Lock,5 Bonno N. Bouma, 3 ,6 and Joost C.M. Meijers3 ,6
Department of Haematology, 3 Obstetrics and Gyneacology,4 and Urology,5 University Hospital, andInstitute of Biomembranes,6 Utrecht University, Utrecht, The Netherlands
ABSTRACT
Protein C inhibitor (PCI) is a heparin-binding plasma serineprotease inhibitor that was originally identified as an inhibitorof activated protein C. PCI has a broad protease specificity, in-hibiting several proteases in hemostasis and fibrinolysis by act-ing as a suicide substrate. Recently it has been reported thatproteases of the reproductive system, such as acrosin, prostate-specific antigen, and tissue kallikrein, can also be effectively in-hibited by PCI. However, a direct relation between PCI andphysiological events during fertilization has not yet been estab-lished. An attempt was made to monitor and localize the inhi-bition of the sperm protease acrosin by PCI. Localization ex-periments for PCI on epididymal spermatozoa showed that PCIis present on the acrosomal cap of human spermatozoa, whichdemonstrates the early presence of PCI in the male reproductivetract. Induction of the acrosome reaction in ejaculated humanspermatozoa resulted in the disappearance of PCI from the plas-ma membrane overlying the acrosomal head and the appearanceof a strict distribution at the equatorial segment of human sper-matozoa. The activity of acrosin in sperm extracts could be ef-fectively inhibited by PCI. Zona-binding assays showed that ac-tive PCI is able to block sperm-egg binding in a concentration-dependent manner. The combination of the potent inhibition ofacrosin and sperm-egg binding by PCI and the localization stud-ies suggested that PCI may protect spermatozoa against pre-mature acrosome reaction and degradation, thereby modulatingthe acrosin activity so that it can coincide with binding to theoocyte.
INTRODUCTION
Protein C inhibitor (PCI) is a plasma glycoprotein be-longing to the serpin superfamily of serine protease inhib-itors, of which otl-protease inhibitor is the prototype [1, 2].PCI was initially identified in blood plasma by Marlar andGriffin [3] and isolated by Suzuki et al. [4] as a majorregulator of the anticoagulant protease activated protein C.In addition to activated protein C, PCI can inhibit severalother proteases involved in blood coagulation, fibrinolysis,and reproduction.
The occurrence of PCI in body fluids other than bloodplasma suggested that PCI may have a function outside theprotein C system. Laurell et al. [5] observed that the con-centration of PCI in seminal plasma was 40 times higherthan the blood plasma level. Hermans et al. [6] describeda very potent inhibition of the sperm protease acrosin byPCI. This created an apparent paradox because it had beendemonstrated that the main source of PCI is the seminal
Accepted October 10, 1997.Received December 30, 1996.'This work was supported in part by grant 92.306 from the Nether-
lands Heart Association and a fellowship from the Royal NetherlandsAcademy for Arts and Sciences. J.C.M.M. is an Established Investigator ofthe Netherlands Heart Foundation.
2Correspondence: Marc G.L.M. Elisen, University Hospital, Depart-ment of Haematology G03.647, P.O. Box 85.500, 3508 GA Utrecht, theNetherlands. FAX: 31-30-251.18.93.
vesicles [5] and that PCI is very rapidly inactivated in sem-inal plasma by proteinases in the reproductive tract such asprostate-specific antigen (PSA) and tissue kallikrein [7-9].Acrosin-PCI complexes could not be detected in seminalplasma, most likely because proacrosin is activated and re-leased only as a consequence of the acrosome reaction dur-ing the fertilization process. On the other hand, it was ob-served that nonphysiological inhibitors of serine proteasessuch as soybean trypsin inhibitor and p-nitrophenyl p'-guanidinobenzoate can inhibit sperm binding to and pene-tration in the zona pellucida, thereby suggesting a regula-tory role for serine proteases and serpins in human fertil-ization [10-13]. Alternatively, this inhibitory effect canalso be placed in another perspective, since the presence ofan inhibitor of proteases in the male reproductive systemcan block premature activation of proacrosin, thereby pro-tecting the spermatozoa and other tissues in the male re-productive system from the destructive action of acrosin.Complex formation of free inhibitor after ejaculation wouldprevent further protection of the spermatozoa by this inhib-itor, and thus at this time the spermatozoa are no longerprotected against degradation by proteases.
The role of PCI in sperm-zona binding was monitoredin combination with localization experiments on humanspermatozoa. This paper provides evidence that PCI is pres-ent on (epididymal) spermatozoa where it can act as animportant regulator of acrosin activity, protecting the sper-matozoa against degradation and premature acrosome re-action without interfering during fertilization.
MATERIALS AND METHODS
All reagents used were analytical grade. All chromogen-ic substrates were purchased from Chromogenix (Milndal,Sweden). Heparin (Thromboliquine) was from OrganonTechnika (Oss, The Netherlands). Human protein C waspurified and activated as previously described [14]. PCI wasimmunopurified from human blood plasma as describedpreviously [15]. Heat-inactivated PCI was obtained by in-cubating PCI for 10 min at 100°C. Functional activity wastested, and the results showed that the heat-inactivated PCIwas no longer able to inhibit activated protein C and throm-bin. Human thrombin was a generous gift of Dr. W. Kisiel(University of New Mexico, Albuquerque, NM).
Rabbit anti-PCI IgG was obtained after immunization ofa New Zealand White rabbit according to standard proto-cols. The antiserum was judged monospecific by immu-nodiffusion [16]. IgG was purified from the serum usingprotein G Sepharose (Pharmacia LKB Biotechnology, Upp-sala, Sweden). The resulting IgG preparation was subse-quently immunopurified using PCI-Sepharose. Immuno-blotting after SDS-PAGE of blood plasma gave a singleband that comigrated with purified PCI. Biotinylated swineanti-rabbit IgG and avidin-fluorescein isothiocyanate(FITC) were purchased from Vector (Burlingame, CA). The
670

PROTEIN C INHIBITOR AND SPERM-EGG BINDING
calcium ionophore A23187 was obtained from Boehringer(Mannheim, Germany).
GPO (pasteurized plasma-protein solution) was obtainedfrom the Central Laboratory of the Red Cross Blood Trans-fusion Services (Amsterdam, The Netherlands). Human tu-bular fluid (HTF) consisted of 100 mM NaC1, 4.7 mM KC1,0.2 mM MgSO 4, 0.37 mM KH 2PO 4, 2.0 mM CaC12, 2.8mM glucose, 21.4 mM sodium lactate, 30 U/ml penicillinG, 10 mg/L streptomycin, 0.0005% phenol red, 24 mMNaHCO 3, 37 mg/L sodium pyruvate, and 10% GPO; pHwas adjusted to 7.4 and the osmolarity to 280 mOsmol/kg.HTF washing medium was almost identical to HTF, withthe following modifications: 4 mM NaHCO 3, 20 mM Hepesbuffer, no phenol red, and no GPO.
Purification and Activation of Boar Proacrosin
Proacrosin was purified from freshly ejaculated boar se-men according to Polakoski and Parrish [17] with someminor modifications. After S-Sepharose chromatography,fractions containing proacrosin from a linear salt gradientfrom 0 to 1.5 M NaCl in 50 mM NaAc, pH 3.0, werepooled and dialyzed against 0.1% trifluoroacetic acid. Fur-ther purification was achieved using a ProRPC HR 10/16reversed-phase column (Pharmacia) with a linear gradientof 0-60% (v:v) acetonitrile in 0.1% trifluoroacetic acid ata flow rate of 1 ml/min using a fast protein liquid chro-matography (FPLC) system. Fractions were pooled and di-alyzed overnight against 7 M urea, 1 M formic acid. FPLCwas further used to subject aliquots of the dialyzed ProRPCpools to gel filtration on a Superose 12 HR 10/30 (Phar-macia) column equilibrated in 7 M urea, 1 M formic acidat a flow rate of 0.45 ml/min. Fractions containing(pro)acrosin were pooled, dialyzed against 50 mM NaAc,pH 4.0, and stored at -70°C until use. Acrosin was acti-vated at pH 8.0 as described previously [17], resulting in asingle band of 37 kDa as analyzed by SDS-PAGE.
Kinetics
All enzymatic reactions were performed at 25°C in 0.05M Tris-HCl (pH 7.4), 125 mM NaCl, 0.1% polyethyleneglycol (Mr 6000), 2 mM CaC12, and 0.5% BSA. Theamount of p-nitroaniline formed was followed at 405 nmusing a kinetic microplate reader (Molecular Devices Cor-poration, Menlo Park, CA). The inhibition of activated pro-tein C, thrombin, and acrosin by PCI was monitored bymeasuring the hydrolysis of the chromogenic substrates S-2366, S-2238, and S-2444, respectively. The reactions werestarted by adding activated protein C (final concentration 1nM), thrombin (final concentration 100 pM), or acrosin (fi-nal concentration 1 nM) to the reaction mixture containingPCI and the chromogenic substrate. Control assays showedthat the enzymes were stable during the time of the exper-iments. Slow binding kinetics were used to study the in-hibition reactions, and second-order rate inactivation con-stants were determined as previously described [15]. Timecourses were obtained by measuring absorbances every 15sec for 45-60 min. For this mechanism, the progress curveof product formation is given by P = vst + ((v0 - v) / k')x (1 - e kt) where P is the amount of product formed attime t, k' is an apparent first-order rate constant, and vo andv, are the initial and steady-state velocities [6]. The realpseudo first-order inactivation constant can subsequently beobtained by nonlinear regression of the progress-curve dataaccording to the equation and correction for the presence
of chromogenic substrate S using the equation kas, = k' x(1 + S/Km,).
Spermatozoa
Human ejaculated spermatozoa were obtained fromhealthy volunteers who were normospermic according toWHO criteria [18]. Ejaculates were collected by mastur-bation into a wide-mouth plastic jar. Semen was liquefiedfor 30 min and spermatozoa were harvested using a 70%Percoll cushion (Pharmacia). Isotonic Percoll (90%) wasprepared by adding one volume of 10-strength concentratedEarle's Balanced Salt Solution (Serva, Heidelberg, Ger-many) to 9 volumes of Percoll. Dilution to 70% was donewith HTF, 0.3% HSA. Liquefied semen was layered on topof 70% Percoll, and the cells were fractionated by centrif-ugation at 300 x g for 15 min. The pellet was washed withHTF and resuspended to the desired concentration. The mo-tility of the spermatozoa was monitored using a Hamilton-Thorn (Beverly, MA) motility analyzer.
Epididymal spermatozoa were obtained from healthymen who had had their vasectomy reversed. The patientsand donors used in this study had given informed consentfor the use of their gametes.
Western Blotting of Sperm Extracts
Liquefied semen samples from healthy volunteers werepooled and centrifuged as described above. The pellet waswashed twice with HTF and centrifuged. Thereafter the pel-let was resuspended in hypotonic HTF medium (200mOsm), cooled to 40C, and vortexed to remove acrosomalcaps as described by Moore et al. [13]. The acrosomal capswere separated from spermatozoa by centrifugation at 300x g for 5 min and subsequently purified on a 10-40%Percoll density gradient formed in isotonic medium andcentrifuged at 1500 x g for 15 min [13]. Acrosomes wererecovered by centrifugation at 14 000 x g for 15 min. Thepellet was treated with 0.1% (v:v) Nonidet P-40 and 8 Murea and then subjected to heat denaturation. The extractwas dialyzed against 1 mM HCI to remove urea.
PCI and acrosome extract were subjected to SDS-PAGE,followed by Western blotting to polyvinylidene fluoridemembrane. The blots were blocked in Tris-HCI, pH 7.4,150 mM NaCl (TBS) containing 5% skimmed milk powderand were incubated with a polyclonal antibody against PCI.After washing with TBS, the blots were incubated withperoxidase-conjugated anti-rabbit IgG (Dako, Glastrup,Denmark) for 2 h. Protein was visualized using chemilu-minescence (SuperSignal Substrate, Western blotting;Pierce, Rockford, IL) according to the manufacturer's in-structions.
Inhibition of Sperm Extract Activity by PCI
Acrosomes were removed from spermatozoa as de-scribed by Moore et al. [13]; this was followed by acidextraction with 1 mM HCI and 0.1% (v:v) Nonidet P-40for 24 h [17]. After centrifugation for 10 min at 1500 X g,the supernatant was used as sperm extract. Protein concen-tration was determined by BCA (Pierce) and found to be0.7 mg/ml. The sperm extract was incubated for 30 min atpH 8 as previously described [17] to allow for activationof proacrosin. Activated sperm extracts (0.14 mg/ml) wereincubated with various concentrations of PCI (0-250 nM)in the presence of heparin (0.1 U/ml) in Tris-HCI (pH 7.4),125 mM NaCl, 0.1% polyethylene glycol 6000, 2 mM
671

ELISEN ET AL.
FIG. 1. Immunofluorescent localizationof PCI antigen on ejaculated spermatozoa.Spermatozoa were stained with rabbitanti-PCI IgG and fluorescein-labeled swineanti-rabbit IgG (B). A depicts a phase-con-trast image of B.
CaCl 2, and 0.5% BSA. At various time points, the proteo-lytic activity was measured by following the hydrolysis ofthe chromogenic substrate S2444 at 405 nm using a kineticmicroplate reader.
Immunocytochemical Detection of PCI on HumanSpermatozoa
Spermatozoa were washed, centrifuged for 3 min at 600x g, and finally resuspended in PBS to a concentration of107 spermatozoa/ml. A 25-lxl sample was put on a glassslide and air dried. For induction of the acrosome reaction,spermatozoa were preincubated for 10 min at 37C with 5mM CaCI 2 in HTF and then incubated for 60 min at 37°Cwith 10 RIM of the calcium ionophore A23187 in dimethylsulfoxide (final concentration 0.5% [v:v]). The sperm werethen centrifuged for 3 min at 600 x g, and the supernatantwas removed carefully and replaced by the same amountof HTF Spermatozoa were fixed by submerging the samplein methanol:acetone (1:1) for 30 min followed by triplewashing with PBS. In addition, two other fixatives wereused for localization experiments: 3% p-formaldehyde/0.025% glutaraldehyde in PBS, and a 3% glutaraldehydesolution in PBS. When these fixatives were used, the fix-ation was followed by incubation with 0.1 M glycine inPBS for 15 min.
The slides were incubated for 60 min with 3% BSA inPBS, followed by a triple wash with PBS. Slides were cov-ered with a drop of rabbit anti-PCI antibodies (10 pxg/ml)in PBS containing 1% BSA and incubated for 60 min in amoist petri dish. After washing of the slides with PBS, bio-tinylated anti-rabbit IgG was added, and incubation wascarried out for 60 min at room temperature. Avidin-FITCwas added after extensive washing with PBS, and incuba-tion was performed for 30 min in the dark. After extensivewashing, the slides were studied using fluorescence mi-croscopy (Leica TCS 4D; Leica laser technik GmbH, Hei-delberg, Germany).
Zona-Binding Assay
Human oocytes that had failed to fertilize during routinein vitro fertilization procedures were collected within 24 h
of culture and washed free of medium through several pas-sages with PBS; they were then stored in a salt solutioncontaining 2 M (NH4 )2S0 4 , 40 mM Hepes, and 0.5% dex-tran at pH 7.0. Before use, these oocytes were extensivelywashed by means of 5 passages with Biggers, Whitten, andWhittingham (BWW) medium [19] containing 4 mg/ml hu-man serum albumin. Spermatozoa from fertile men werewashed twice in HTF and suspended to the desired con-centration. Spermatozoa were incubated for 6 h to allowcapacitation before oocytes and protein were added.
Alternatively, a population of acrosome-reacted spermwas obtained as follows. A 4-h capacitation period wasfollowed by incubation with 3 mM CaC12 for 10 min at37°C, and the acrosome reaction was initiated by additionof A23187 as described above. The acrosome reaction wasallowed to proceed for 120 min at 37°C. The incubationwas stopped by centrifugation for 5 min at 300 g fol-lowed by triple washing with BWW medium.
Aliquots of 250 pl1 of spermatozoa solution (1.5 x 106motile spermatozoa/ml, for both acrosome-intact and ac-rosome-reacted sperm preparations) were added to the oo-cytes together with PCI, heat-inactivated PCI, or rabbit anti-PCI. After a 4-h incubation period, the oocytes were aspi-rated in and out of a narrow-bore pipette several times toremove the loosely attached spermatozoa. After multiplewashing steps, the oocytes were examined for sperm bind-ing using phase-contrast microscopy.
RESULTS
Immunofluorescent Localization of PCI Antigen onHuman Spermatozoa
A polyclonal antiserum against PCI bound selectively tothe acrosomal region of intact human spermatozoa asshown by indirect immunofluorescent staining. This distri-bution was observed in ejaculated sperm (Fig. 1) as wellas on epididymal spermatozoa (Fig. 2). No staining at themidsection and tail was observed. The same result was ob-tained after fixation of the spermatozoa with 3% p-form-aldehyde or 0.5% glutaraldehyde instead of methanol:ace-tone (1:1). Also, incubation of live sperm with the antibodyagainst PCI followed by posterior fixation with 3% p-form-
672

PROTEIN C INHIBITOR AND SPERM-EGG BINDING
FIG. 2. Immunofluorescent localizationof PCI antigen on epididymal spermato-zoa. PCI was visualized by indirect immu-nofluorescent staining of intact spermato-zoa (B). A depicts a phase-contrast imageof epididymal spermatozoa of B.
aldehyde or 0.5% glutaraldehyde did not alter the stainingpattern. Preabsorption of the antiserum with purified PCIquenched the staining of the spermatozoa completely (datanot shown).
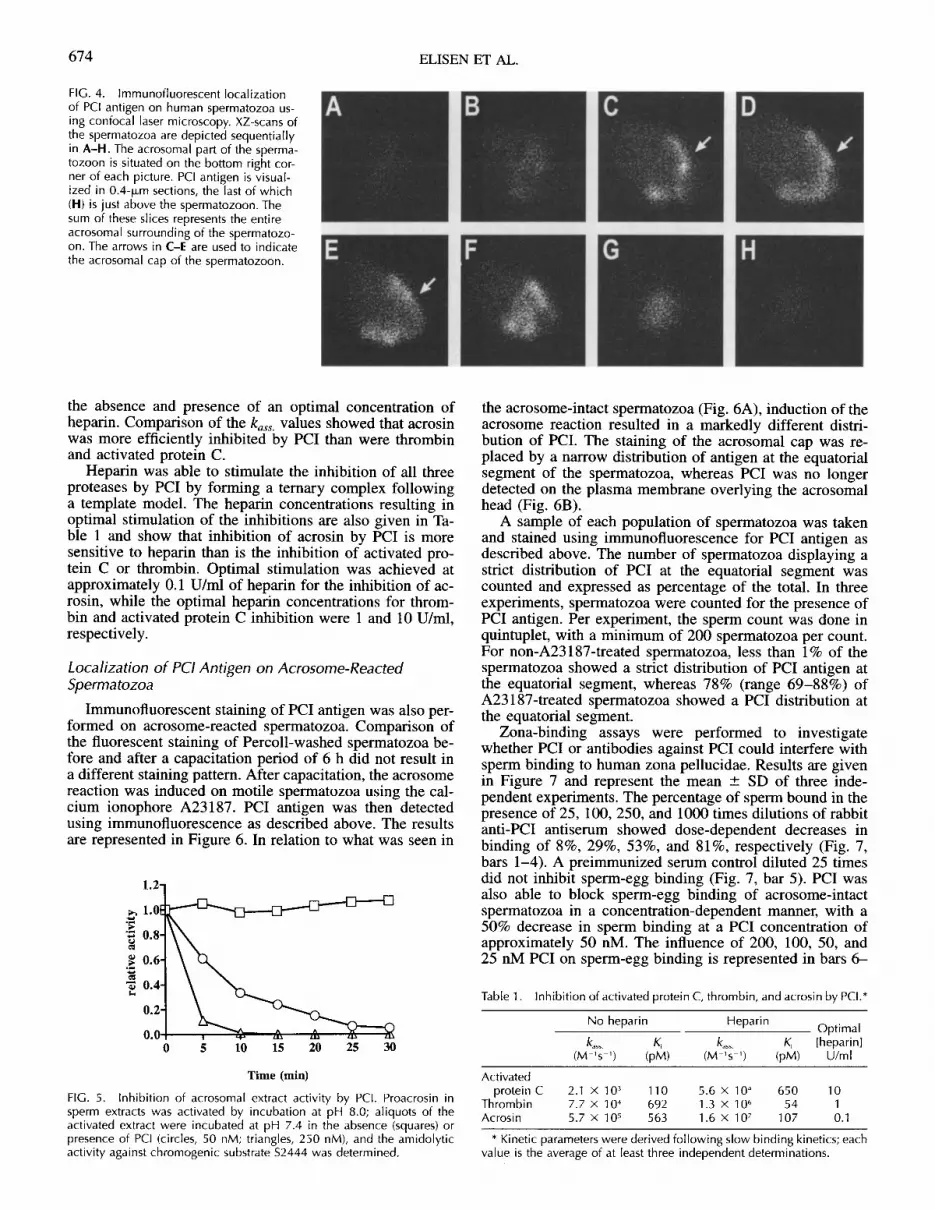
The specificity of the antiserum was further checked bySDS-PAGE and Western blotting of a sperm extract and ofPCI derived from blood plasma (Fig. 3). In both samples,a protein was visualized that had the molecular weight ofPCI. Confocal laser microscopy was used to monitor thedistribution of PCI antigen. A scan of a spermatozoon, us-ing 0.4-jlm horizontal intervals, is represented in Figure 4.PCI antigen is visualized in the various sections starting atpanel A, which is just below the spermatozoon. The spermhead is on the bottom right corner of each picture. SectionB represents a 0.4-jLm layer above layer A, and the sum ofthe consecutive sections (A-H) represents the entire spermhead. Section B shows the presence of PCI antigen acrossthe 0.4-jlm layer of the spermatozoon. Panels C-E displaya horseshoe-like distribution of PCI antigen (arrows) andno PCI antigen detected in the center of the spermatozoalhead. The horseshoe-like distribution of PCI antigenchanges into a staining pattern across the acrosomal cap ofthe spermatozoon (sections F-H). The changing pattern ofthe 0.4-1m sections is suggestive of a surface staining ofthe acrosomal cap of human spermatozoa.
Inhibition of Sperm Extract Activity by PCI
After incubation of the sperm extract at pH 8, which isa specific way to activate proacrosin, an increase in theproteolytic activity of the extract toward the chromogenicsubstrate S2444 was measured. The sperm extracts wereused as source for acrosin activity and incubated with PCI.At various time intervals, the remaining amidolytic activitywas measured. Figure 5 shows that the proteolytic activityof the extract was inhibited by PCI in a concentration-de-pendent manner. Aliquots of sperm extract that had notbeen incubated at pH 8 did not exhibit measurable activitytoward the chromogenic substrate either in the absence orpresence of PCI (not shown).
Inhibition of Serine Proteases by PCI
Slow binding kinetics were used to determine the kineticconstants for the inhibition of activated protein C, throm-bin, and acrosin by PCI. The inhibition reactions were alsomonitored at optimal concentrations of heparin, since it isknown that heparin alters the kinetic constants for all threeprotease-serpin reactions. The kinetic constants Ki and ka,,are shown in Table 1. The kinetic parameters indicate thatacrosin could be very effectively inhibited by PCI with as-sociation constants of 5.7 x 105 and 1.6 x 107 M-Is - in
FIG. 3. Western blot showing PCI purified from blood plasma (lane A)and a sperm extract (lane B). The blot was incubated with rabbit anti-PCIantibodies followed by incubation with peroxidase-conjugated anti-rabbitIgG.
673
ELISEN ET AL.
FIG. 4. Immunofluorescent localizationof PCI antigen on human spermatozoa us-ing confocal laser microscopy. XZ-scans ofthe spermatozoa are depicted sequentiallyin A-H. The acrosomal part of the sperma-tozoon is situated on the bottom right cor-ner of each picture. PCI antigen is visual-ized in 0.4-Lm sections, the last of which(H) is just above the spermatozoon. Thesum of these slices represents the entireacrosomal surrounding of the spermatozo-on. The arrows in C-E are used to indicatethe acrosomal cap of the spermatozoon.
the absence and presence of an optimal concentration ofheparin. Comparison of the ka,, values showed that acrosinwas more efficiently inhibited by PCI than were thrombinand activated protein C.
Heparin was able to stimulate the inhibition of all threeproteases by PCI by forming a ternary complex followinga template model. The heparin concentrations resulting inoptimal stimulation of the inhibitions are also given in Ta-ble 1 and show that inhibition of acrosin by PCI is moresensitive to heparin than is the inhibition of activated pro-tein C or thrombin. Optimal stimulation was achieved atapproximately 0.1 U/ml of heparin for the inhibition of ac-rosin, while the optimal heparin concentrations for throm-bin and activated protein C inhibition were 1 and 10 U/ml,respectively.
Localization of PCI Antigen on Acrosome-ReactedSpermatozoa
Immunofluorescent staining of PCI antigen was also per-formed on acrosome-reacted spermatozoa. Comparison ofthe fluorescent staining of Percoll-washed spermatozoa be-fore and after a capacitation period of 6 h did not result ina different staining pattern. After capacitation, the acrosomereaction was induced on motile spermatozoa using the cal-cium ionophore A23187. PCI antigen was then detectedusing immunofluorescence as described above. The resultsare represented in Figure 6. In relation to what was seen in
eZ:
U
5.'U
I-
0 5 10 15 20 25 30
Time (min)
FIG. 5. Inhibition of acrosomal extract activity by PCI. Proacrosin insperm extracts was activated by incubation at pH 8.0; aliquots of theactivated extract were incubated at pH 7.4 in the absence (squares) orpresence of PCI (circles, 50 nM; triangles, 250 nM), and the amidolyticactivity against chromogenic substrate S2444 was determined.
the acrosome-intact spermatozoa (Fig. 6A), induction of theacrosome reaction resulted in a markedly different distri-bution of PCI. The staining of the acrosomal cap was re-placed by a narrow distribution of antigen at the equatorialsegment of the spermatozoa, whereas PCI was no longerdetected on the plasma membrane overlying the acrosomalhead (Fig. 6B).
A sample of each population of spermatozoa was takenand stained using immunofluorescence for PCI antigen asdescribed above. The number of spermatozoa displaying astrict distribution of PCI at the equatorial segment wascounted and expressed as percentage of the total. In threeexperiments, spermatozoa were counted for the presence ofPCI antigen. Per experiment, the sperm count was done inquintuplet, with a minimum of 200 spermatozoa per count.For non-A23187-treated spermatozoa, less than 1% of thespermatozoa showed a strict distribution of PCI antigen atthe equatorial segment, whereas 78% (range 69-88%) ofA23187-treated spermatozoa showed a PCI distribution atthe equatorial segment.
Zona-binding assays were performed to investigatewhether PCI or antibodies against PCI could interfere withsperm binding to human zona pellucidae. Results are givenin Figure 7 and represent the mean SD of three inde-pendent experiments. The percentage of sperm bound in thepresence of 25, 100, 250, and 1000 times dilutions of rabbitanti-PCI antiserum showed dose-dependent decreases inbinding of 8%, 29%, 53%, and 81%, respectively (Fig. 7,bars 1-4). A preimmunized serum control diluted 25 timesdid not inhibit sperm-egg binding (Fig. 7, bar 5). PCI wasalso able to block sperm-egg binding of acrosome-intactspermatozoa in a concentration-dependent manner, with a50% decrease in sperm binding at a PCI concentration ofapproximately 50 nM. The influence of 200, 100, 50, and25 nM PCI on sperm-egg binding is represented in bars 6-
Table 1. Inhibition of activated protein C, thrombin, and acrosin by PCI.*
No heparin HeparinOptimal
ka,, Ki kas K [heparin](M- s -) (pM) (M-s ') (pM) U/ml
Activatedprotein C 2.1 x 103 110 5.6 x 104 650 10
Thrombin 7.7 x 104 692 1.3 x 106 54 1Acrosin 5.7 x 105 563 1.6 x 107 107 0.1
* Kinetic parameters were derived following slow binding kinetics; eachvalue is the average of at least three independent determinations.
674

PROTEIN C INHIBITOR AND SPERM-EGG BINDING
FIG. 6. Immunofluorescent localization of PCI antigen on capacitated(A) and acrosome-reacted (B) human spermatozoa. Sperm were stainedwith rabbit anti-PCI IgG and fluorescein-labeled swine anti-rabbit IgG.
9, giving 13%, 27%, 55%, and 81% sperm-egg binding,respectively, as compared to the value for a buffer control(bar 10). In contrast, 200 nM heat-inactivated PCI did notaffect sperm-egg binding (bar 11). Control experiments ver-ified that the heat-inactivated PCI preparation was no lon-ger able to inhibit activated protein C, thrombin, or acrosin.
Zona-binding studies were also performed with A23187-treated and acrosome-intact spermatozoa (Fig. 8). To ruleout a possible effect of usage of different donors, the spermpreparation was divided in two parts of which one half wastreated with ionophore. The values for zona binding of ac-rosome-intact and acrosome-reacted spermatozoa per donorwere compared and expressed as a percentage of the buffercontrol value. The results, representing experiments with atleast three donors, were averaged and are depicted in Figure8. Minor differences in percentages of buffer and serumcontrols between Figures 7 and 8 are due to differencesbetween donors. Similar to what was seen in acrosome-intact sperm, the binding of acrosome-reacted spermatozoato zona pellucidae was inhibited by addition of 200 nMPCI (Fig. 8, bars 1-2). Comparison of the zona binding ofthe two populations of spermatozoa in the presence of abuffer control showed no significant differences.
When acrosome-reacted spermatozoa were incubatedwith rabbit anti-PCI antiserum, a 40% ( 12%) decreasein sperm binding to the zona pellucida was observed inrelation to the value for the preimmunized serum control,whereas the zona binding of acrosome-intact cells wasblocked by 86% (± 8%) (Fig. 8). A motility analysis ofboth acrosome-intact and acrosome-reacted cells excludedthe possibility that differences in motility were responsiblefor decreased binding in the presence of PCI or antibodies.Acrosome-intact cells revealed a progressive motility of81% (range: 76-82%) and a mean track speed of 85.9 jim/sec. A23187-treated cells had a motility of 63% (range 49-71%) and a mean track speed of 65.6 jim/sec. Both acro-some-intact and acrosome-reacted populations had astraightness varying between 85% and 92%.
DISCUSSION
The aim of this study was to investigate the apparentparadox between the potent inhibition of the sperm proteaseacrosin by PCI [6] and the rapid complex formation andinactivation of PCI by proteases other than acrosin, such asPSA and tissue kallikrein, after ejaculation [5-7]. A regu-latory role of PCI on acrosin activity was therefore ques-tioned the more so because this protease was not expectedto be accessible to PCI after ejaculation.
125-
N 100'
75
25
1234 5 6 7 8 9 1011
FIG. 7. Effect of PCI and rabbit anti-PCI antibodies on binding of sper-matozoa to human zona pellucida. In each experiment, 15 zonae pellu-cidae were used per reaction condition. Spermatozoa and human zonapellucidae were incubated in the presence of PCI and rabbit anti-PCIantiserum for 4 h. After incubation, the unbound spermatozoa were re-moved and the bound spermatozoa were counted. Bars 1-4 denote thenumber of sperm bound in the presence of 25, 100, 250, and 1000 timesdilutions of rabbit anti-PCI antiserum, respectively. Bar 5 represents a 25times-diluted preimmunized serum control. The effect of 200, 100, 50,and 25 nM PCI on sperm binding is represented in bars 6-9, respectively,and compared to a buffer control (bar 10). Bar 11 represents the effect of200 nM heat-inactivated PCI on sperm-egg binding. Results are the meant SD of three independent experiments.
However, PCI may also function as an inhibitor of ac-rosin during storage of spermatozoa. A premature activa-tion of proacrosin, possibly occurring during removal ofdead spermatozoa, could lead to damage in the male repro-ductive tissues because of the matrix-degrading propertiesof acrosin [20]. A premature activation of proacrosin mayalso initiate massive autoactivation of proacrosin, resultingin a population of acrosome-reacted spermatozoa in themale reproductive tract.
Immunofluorescence data on human spermatozoa isolat-ed from the epididymis showed the presence of PCI, ex-clusively localized at the acrosomal region of human sper-matozoa. Localization of PCI using confocal laser micros-copy revealed a horseshoe-like distribution of PCI on theplasma membrane overlying the acrosomal head, whereasno staining inside the spermatozoa was observed. This in-dicated that PCI was expressed and secreted in testis orepididymis followed by association or complex formationto the plasma membrane or membrane-associated proteinsof the spermatozoon. The presence of PCI on epididymalspermatozoa suggests a function during storage of sper-matozoa in the epididymis. The presence of PCI in the ep-ididymal glands had already been described by Laurell andcoworkers [5], who studied the presence of PCI in the malereproductive system and found PCI in the secretory epithe-lium and secretory content in the glandular lumina of the
125-
o 100-
75-
50-
25
IiI M. ".
1 2 34 5 6 7 8
FIG. 8. Zona-binding experiments with acrosome-intact (hatched bars)and acrosome-reacted (open bars) spermatozoa. Spermatozoa and humanzona pellucidae were coincubated with 200 nM PCI (1-2), buffer control(3-4), rabbit anti-PCI antiserum (25 times dilution, 5-6), or a 25 times-diluted preimmunized serum control (7-8) for 4 h. After incubation, theunbound spermatozoa were removed and the bound spermatozoa werecounted. Results are the mean SD of three individual experiments.
-· ·· ~ - ------ -r .I _
675

ELISEN ET AL.
epididymal glands. Kirchhoff et al. [21] described the pres-ence in the epithelium of an mRNA encoding for an extra-cellular protease inhibitor related to PCI. One of the func-tions of epididymal fluid is to prevent lytic enzymes, re-leased from the acrosomes of degenerating spermatozoa,from acting upon other cells. The presence of PCI can resultin an effective inhibition of acrosin. In agreement with Her-mans and coworkers [6], we showed that PCI is a potentinhibitor of acrosin. Our data also indicate that acrosin is amuch better target protease for PCI than are activated pro-tein C and thrombin, two target proteases for PCI in bloodcoagulation. The stimulating effect of heparin on the ac-rosin inhibition by PCI suggests that this reaction can occuron a glycosaminoglycan-containing surface.
Our study does not explain whether the association/com-plex formation of PCI to the spermatozoa is due to leakageof acrosin followed by complex formation with PCI or as-sociation of PCI to heparin-like molecules on the plasmamembrane, but the distribution of PCI on the entire acro-somal region suggests the latter. The effect of the rabbitanti-PCI antibodies in the zona-binding assay suggests thatthe interaction of the antibody with the spermatozoa occurson the outer surface of the acrosomal caps of human sper-matozoa.
This agrees in part with observations made by Zheng etal. [22], who have localized PCI in the vicinity of disruptedacrosomal membranes of abnormal sperm heads after ejac-ulation. However, Zheng et al. did not observe PCI on ep-ididymal or morphologically normal sperm heads, nor didthey show whether PCI-containing spermatozoa were ableto bind to human zona pellucidae. We observed PCI onpopulations of living spermatozoa containing probably bothnormal and abnormal sperm heads. Crude ejaculates, aswell as spermatozoa after swim-up procedures used for invitro fertilization, expressed PCI on the plasma membraneoverlying the acrosomal head of all spermatozoa. Zona-binding assays demonstrated that the spermatozoa positivefor PCI had fertilizing capacity. The addition of exogenousactive PCI blocked sperm-egg binding, whereas exogenousheat-inactivated PCI did not. Therefore, it appears that thepresence of this potent acrosin inhibitor on spermatozoadoes not have to interfere with sperm-egg binding, whichmay indicate that PCI is no longer reactive when the sper-matozoon encounters the oocyte. After ejaculation, sper-matozoa encounter kallikrein-like proteases such as PSAthat subsequently can digest the insoluble coagula semen-ogelin I+II (Sg I+II) but also damage spermatozoa. It hasbeen demonstrated that PCI inhibits PSA-catalyzed degra-dation of Sg I+II by forming PCI-PSA complexes [23].The occurrence of active PCI on the outside of the acro-somal cap may protect the acrosomal content against deg-radation by PSA, thereby maintaining the fertilizing abilityof the spermatozoon. Since PCI has been shown to formcomplexes with PSA and tissue kallikrein in seminal plas-ma after ejaculation [7], the presence of high levels of com-plexed PCI does not necessarily have to interfere withsperm-egg binding.
Another finding showing that the presence of PCI on theplasma membrane overlying the acrosomal head does nothave to interfere with fertilization was that acrosome-re-acted spermatozoa lacked PCI on the acrosomal head. In-duction of the acrosome reaction by calcium and A23187resulted in spermatozoa in which PCI was present only atthe equatorial segment. So even if active PCI were presenton the acrosomal head, the disappearance or shift of PCIfrom the acrosomal to the equatorial region shows that PCI
does not have to interfere with zona binding as the initialstep in fertilization. The disappearance of a potent acrosininhibitor (PCI) from the plasma membrane overlying theacrosomal head coincides with the activation of proacrosin.
We observed that addition of exogenous active PCI hadan inhibitory effect on sperm binding to the zona pellucidaof both acrosome-intact and acrosome-reacted spermatozoa.Both types of spermatozoa were able to bind to the zonapellucida; this confirmed a possible involvement of acrosinin sperm binding to and penetration through the zona pel-lucida [24, 25]. Other groups have found that nonphysiol-ogic serine protease inhibitors such as p-nitrophenyl p'-guanidinobenzoate, soybean trypsin inhibitor, and 4-ami-nobenzamidine are able to inhibit sperm-egg binding andhave related this to the inhibition of acrosin [10-13].
When rabbit anti-PCI antibodies were added to acro-some-intact spermatozoa, a dose-dependent inhibitory ef-fect was observed in sperm binding to the zona pellucida.The inhibitory effect in this experimental setting is mostlikely caused by coverage of the acrosomal cap by the anti-PCI antibodies. This coverage may disable binding of zonapellucida proteins to intact spermatozoa and/or stimulatethe acrosome reaction.
No PCI could be observed on the plasma membraneoverlying the acrosomal head of A23187-treated sperma-tozoa. Therefore, when A23187-treated spermatozoa wereused instead of acrosome-intact spermatozoa, an inhibitoryeffect on sperm-egg binding due to coverage of the plasmamembrane overlying the acrosomal head by anti-PCI anti-bodies was not expected or observed. The presence of PCIexclusively on the equatorial segment of the spermatozoacan be explained by either the remainder of the plasmamembrane or a redistribution of PCI.
The observation that the population of A23187-treatedspermatozoa cells included acrosome-intact as well as ac-rosome-reacted spermatozoa agrees with the results ob-tained for the zona-binding assay. Incubation of theA23187-treated spermatozoa with rabbit anti-PCI antiserumresulted in a small but significant decrease in binding ofspermatozoa, probably caused by the remaining -20% ac-rosome-intact spermatozoa.
Inhibition of acrosin by PCI in the vicinity of an oocyteappears to be a disadvantage for successful fertilization.Furthermore, PCI will be complexed to seminal enzymesafter ejaculation. Therefore, the potent inhibition of acro-somal extract activity by PCI and the presence of PCI onepididymal spermatozoa suggest a role for PCI as acrosininhibitor during storage of spermatozoa rather than duringsperm-egg binding. In addition, our findings may explainwhy human epididymal spermatozoa have a rather low fer-tilization potential [26]. Zona-binding assays have demon-strated that active PCI inhibits sperm-egg binding whereasinactive PCI does not. The presence of inactivated PCI onejaculated spermatozoa due to cleavage by prostatic pro-teases does not interfere with sperm-egg binding. Epidid-ymal spermatozoa that have not encountered proteases ofprostatic/seminal vesicle origin may possess active PCI.The presence of active PCI on the sperm heads may inter-fere with efficient sperm-egg contact when epididymalspermatozoa are used for in vitro fertilization.
Studies by Tesarik and coworkers [24] have demonstrat-ed that human spermatozoa may release acrosin during anearly stage of the acrosome reaction, before definitive mem-brane vesiculization, with acrosin exposed on the spermsurface overlying the acrosome. It is not unlikely that theseearly stages of acrosome reactions also occur in the male
676

PROTEIN C INHIBITOR AND SPERM-EGG BINDING
reproductive tract and that they have to be inhibited despitea low occurrence.
The presence of PCI in the male reproductive tract maymodulate the activity of acrosin, thereby protecting the ac-rosomal content of spermatozoa and possibly also surround-ing tissue against premature degradation by acrosin. In ad-dition, PCI located on the sperm head will protect the ac-rosomal content against proteolytic attack in seminal plas-ma after ejaculation. PCI may therefore be regarded as aprotector of human spermatozoa that modulates the acrosinactivity so that activation and release of acrosin coincideonly with binding to the oocyte.
ACKNOWLEDGMENTS
The authors are highly indebted to the personnel of the fertility labo-ratory of the University Hospital Utrecht for helpful advice. We also thankthe personnel of the Red Cross Blood Bank for providing us with thedonor plasma used for the isolation of PCI.
REFERENCES
1. Huber R, Carrell RW. Implications of the three-dimensional structureof al-antitrypsin for structure and function of serpins. Biochemistry1989; 28:8951-8966.
2. Suzuki K, Deyashiki Y, Nishioka J, Kurachi K, Akira M, YamamotoS, Hashimoto S. Characterization of a cDNA for human protein Cinhibitor. A new member of the plasma serine protease inhibitor su-perfamily. J Biol Chem 1987; 262:611-616.
3. Marlar RA, Griffin JH. Deficiency of protein C inhibitor in combinedfactor V/VIII disease. J Clin Invest 1980; 6:1186-1189.
4. Suzuki K, Nishioka J, Hashimoto S. Protein C inhibitor. Purificationfrom human plasma and characterization. J Biol Chem 1983; 258:163-168.
5. Laurell M, Christensson A, Abrahamsson P-A, Stenflo J, Lilja H. Pro-tein C inhibitor in human body fluids. J Clin Invest 1992; 89:1094-1101.
6. Hermans JM, Jones R, Stone SR. Rapid inhibition of the sperm pro-tease acrosin by protein C inhibitor. Biochemistry 1994; 33:5440-5444.
7. Christensson A, Lilja H. Complex formation between protein C in-hibitor and prostate-specific antigen in vitro and in human semen. EurJ Biochem 1994; 220:45-53.
8. Espafia F, Fink E, Sanchez-Cuenca J, Gilabert J, Estelles A, WitzgallK-H. Complexes of tissue kallikrein with protein C inhibitor in humansemen and urine. Eur J Biochem 1995; 234:641-649.
9. Deperthes D, Chapdelaine P, Tremblay RR, Brunet C, Berton J, H6bertJ, Lazure C, Dub6 JY. Isolation of prostatic kallikrein hK2, alsoknown as hGK-1, in human seminal plasma. Biochim Biophys Acta1995; 1245:311-316.
10. Van der Ven HH, Kaminski J, Bauer L, Zaneveld LJD. Inhibition of
human sperm penetration into zona-free hamster oocytes by proteinaseinhibitors. Fertil Steril 1985; 43:609-616.
11. Llanos M, Vigil P, Salgado AM, Morales P. Inhibition of the acrosomereaction by trypsin inhibitors and prevention of penetration of sper-matozoa through the zona pellucida. J Reprod Fertil 1993; 97:173-178.
12. Benau DA, Storey BT. Characterization of the mouse sperm plasmamembrane zona-binding site sensitive to trypsin inhibitors. Biol Re-prod 1987; 36:282-292.
13. Moore A, Penfold LM, Johnson JL, Latchman DS, Moore HDM. Hu-man sperm-egg binding is inhibited by peptides corresponding to coreregion of an acrosomal serine protease inhibitor. Mol Reprod Dev1993; 34:280-291.
14. Hackeng TM, Hessing M, van't Veer C, Meijer-Huizinga F, MeijersJCM, de Groot PG, van Mourik JA, Bouma BN. Protein S binding isrequired for expression of cofactor activity for activated protein C. JBiol Chem 1993; 268:3993-4000.
15. Elisen MGLM, Maseland MHH, Church FC, Bouma BN, MeijersJCM. Role of the A+ helix in heparin binding to protein C inhibitor.Thromb Hemostasis 1996; 75:760-766.
16. Ouchterlony 6. Antigen antibody reactions in gels. Ark Mineral Geol1949; 26:1-16.
17. Polakoski KL, Parrish RE Boar proacrosin: purification and prelimi-nary activation studies of proacrosin isolated from ejaculated boarsperm. J Biol Chem 1977; 252:1888-1894.
18. World Health Organisation. WHO Laboratory Manual for the Exam-ination of Human Semen and Semen-Cervical Mucus Interaction.Cambridge: The Press Syndicate of the University of Cambridge;1987: 55-58.
19. Biggers JD, Whitten WK, Whittingham DG. The culture of mouseembryos in vitro. In: Daniel JC Jr. (ed.), Methods in Mammalian Em-bryology. San Francisco, CA: WH Freeman and Co.; 1971: 86-116.
20. Planchenault T, echovd D, Keil-Dlouha V. Matrix degrading prop-erties of sperm serine protease acrosin. FEBS Lett 1991; 294:279-281.
21. Kirchhoff C, Habben I, Ivell R, Krull N. A major human epididymis-specific cDNA encodes a protein with sequence homology to extra-cellular proteinase inhibitors. Biol Reprod 1991; 45:350-357.
22. Zheng X, Geiger M, Ecke S, Bielek E, Donner E, Eberspicher U,Schleuning W-D, Binder BR. Inhibition of acrosin by protein C in-hibitor and localization of protein C inhibitor to spermatozoa. Am JPhysiol 1994; 267:C466-C472.
23. Kise H, Nishioka J, Kawamura J, Suzuki K. Characterization of se-menogelin and its molecular interaction with prostate-specific antigenand protein C inhibitor. Fur J Biochem 1996; 238:88-96.
24. Tesarik J, Drahorad J, Peknicova J. Subcellular immunochemical lo-calization of acrosin in human spermatozoa during the acrosome re-action and zona pellucida penetration. Fertil Steril 1988; 50:133-144.
25. De Jonge CJ, Mack SR, Zaneveld LJD. Inhibition of human acrosomereaction by proteinase inhibitors. Gamete Res 1989; 23:387-397.
26. Silber SJ, Nagy ZP, Liu J, Godoy H, Devroey P, Van Steirtegem AC.Conventional in-vitro fertilization versus intracytoplasmic sperm in-jection for patients requiring microsurgical sperm aspiration. Hum Re-prod 1994; 9:1705-1709.
677





























