Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Proteomic analysis of NME1/NDPK A null mouse liver:evidence for a post-translational regulation of annexin IVand EF-1Bα
Arnaud Bruneel & Dominique Wendum & Valérie Labas & Odile Mulner-Lorillon &
Joelle Vinh & Nelly Bosselut & Eric Ballot & Bruno Baudin & Chantal Housset &Sandrine Dabernat & Marie-Lise Lacombe & Mathieu Boissan
Received: 5 February 2011 /Accepted: 7 April 2011 /Published online: 4 May 2011# Springer-Verlag 2011
Abstract NME/NDPK family proteins are involved in thecontrol of intracellular nucleotide homeostasis as well as inboth physiological and pathological cellular processes, suchas proliferation, differentiation, development, apoptosis,and metastasis dissemination, through mechanisms stilllargely unknown. One family member, NME1/NDPK-A,is a metastasis suppressor, yet the primary physiologicalfunctions of this protein are still missing. The purpose ofthis study was to identify new NME1/NDPK-A-dependent
biological functions and pathways regulated by this gene inthe liver. We analyzed the proteomes of wild-type andtransgenic NME1-null mouse livers by combining two-dimensional gel electrophoresis and mass spectrometry(matrix-assisted laser desorption/ionization time of flightand liquid chromatography–tandem mass spectrometry).We found that the levels of three proteins, namely,phenylalanine hydroxylase, annexin IV, and elongationfactor 1 Bα (EF-1Bα), were strongly reduced in the
Electronic supplementary material The online version of this article(doi:10.1007/s00210-011-0639-5) contains supplementary material,which is available to authorized users.
A. Bruneel :N. Bosselut :B. BaudinService de Biochimie A, Hôpital Saint-Antoine, AP-HP,75012 Paris, France
A. Bruneel :B. BaudinUPRES EA 4530, Université Paris-Sud 11,Châtenay-Malabry, France
D. Wendum :O. Mulner-Lorillon : C. Housset :M.-L. Lacombe :M. Boissan (*)UPMC Université Paris 06,75005 Paris, Francee-mail: [email protected]
D. Wendum :C. Housset :M.-L. Lacombe :M. BoissanINSERM UMR_S938, Centre de Recherches Saint-Antoine,75571 Paris Cedex 12, France
D. WendumLaboratoire d’Anatomie Pathologique, Hôpital Saint-Antoine,75012 Paris, France
V. Labas : J. VinhCNRS USR 3149, Spectrométrie de Masse Biologiqueet Protéomique, ESPCI-ParisTech,Paris, France
O. Mulner-LorillonUMR 7150 CNRS, Station Biologique de Roscoff,29682 Roscoff Cedex, France
E. BallotService d’Immunologie et Hématologie Biologique,Hôpital Saint-Antoine,75012 Paris, France
S. DabernatEA DRED 3674, Laboratoire de Biologie de la Différenciationet du Développement, Université de Bordeaux 2,Bordeaux, France
M. BoissanService de Biochimie et Hormonologie, AP-HP, Hôpital Tenon,75020 Paris, France
Present Address:M. BoissanDynamique de la Membrane et du Cytosquelette, Institut Curie,CNRS UMR 144,Paris, France
Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419DOI 10.1007/s00210-011-0639-5

cytosolic fraction of NME1−/− mouse livers when comparedto the wild type. This was confirmed by immunoblottinganalysis. No concomitant reduction in the correspondingmessenger RNAs or of total protein level was observed,however, suggesting that NME1 controls annexin IV andEF-1Bα amounts by post-translational mechanisms. NME1deletion induced a change in the subcellular location ofannexin IV in hepatocytes resulting in enrichment of thisprotein at the plasma membrane. We also observed aredistribution of EF-1Bα in NME1−/− hepatocytes to anintracytoplasmic compartment that colocalized with a markerof the reticulum endoplasmic. Finally, we found reducedexpression of annexin IV coincident with decreased NME1expression in a panel of different carcinoma cell lines. Takentogether, our data suggest for the first time that NME1 mightregulate the subcellular trafficking of annexin IVand EF-1Bα.The potential role of these proteins in metastatic disseminationis discussed.
Keywords NM23 . Transgenic mice . Liver . Proteome .
Annexin IV. EF-1Bα
AbbreviationsNDPK Nucleoside diphosphate kinase2-DE Two-dimensional gel electrophoresisCCB Colloidal Coomassie bluePMF Peptide mass fingerprintingLC-MS/MS Liquid chromatography–tandem
mass spectrometryER Endoplasmic reticulum
Introduction
Ten genes have been identified in the NME/NM23 familyin humans. The two most abundantly expressed NMEgenes, and by far the most studied, are NME1 and NME2,which encode, respectively, the A and B isoforms ofnucleoside diphosphate kinases (NDPK). These enzymescatalyze phosphorylation of nucleoside diphosphates toform the corresponding nucleoside triphosphates. BothNDPK A and B isoforms, which are 88% identical, aremainly cytosolic, but they can be found also, at leasttransiently, associated with membranes and in nuclei. Themouse orthologs of NME1 and NME2 encode proteins thatare respectively 94% and 98% identical to their humancounterparts. Besides their known function in controllingintracellular nucleotide homeostasis, NME/NDPK proteinsare involved in both physiological and pathological cellularprocesses, such as proliferation, differentiation, develop-ment, apoptosis, and metastasis dissemination, throughmechanisms still largely unknown (Boissan et al. 2009).
Notably, NME1 is a metastasis suppressor (Steeg et al.1988; Boissan et al. 2005, 2010).
A variety of regulatory activities have been attributed toNME/NDPK proteins, including DNA binding and regulationof gene expression, and cleavage of DNA (Fan et al. 2003;Postel 2003; Kaetzel et al. 2006). The phosphorylatedintermediate of these proteins is also reported to havephosphotransferase activity towards a number of proteinssuch as ATP citrate lyase, aldolase, and succinyl thiokinase(Engel et al. 1995; Steeg et al. 2003). Over the past twodecades, many proteins have been found that interact withthe NME/NDPK family members, providing new insightsinto the NDPK biological functions. Among the multipleproteins interacting with NME, many are related directly orindirectly to G proteins, to their exchange factors or theirregulatory factors, for example, Rad (Zhu et al. 1999; Tsenget al. 2001), Tiam1 (Otsuki et al. 2001), KSR (Hartsough etal. 2002), (Salerno et al. 2005), Lbc (Iwashita et al. 2004),and Dbl-1 (Murakami et al. 2008a,b). Due to their NDPKactivity, NME1/NME2 were proposed to play a nucleotide-charging role for tubulin during the process of microtubuleassembly. Several studies reported that NME co-purifies withmicrotubules, although no direct interaction with NME wasobserved (Melki et al. 1992; Gallagher et al. 2003). Severaldata point to a role of NME in the regulation of vesiculartrafficking. NME2 interacts with the cytosolic coat proteincomplex II (COPII), which mediates vesicle formation fromthe endoplasmic reticulum (ER) and is essential fortrafficking from the ER to the Golgi apparatus (Kapetanovichet al. 2005). In addition, studies of Drosophila andmammalian cells have revealed a function of NME in theendocytic pathway, in particular, in regulating synapticvesicle endocytosis (Krishnan et al. 2001) and vesicletransport-mediated turnover of membrane receptors (Dammaiet al. 2003; Nallamothu et al. 2008). NME2 was also foundto bind to ICAP1-α, a regulator of cell–matrix adhesionmediated by β1 integrin (Fournier et al. 2002). Anotherinteresting finding is the direct binding of cytosolic NDPK Aand B to membrane lipids, reinforcing a possible role ofNDPK at the membrane (Baughman et al. 2008; Mitchellet al. 2009).
To identify potential new biological functions of NME1/NDPK A, we have used a differential proteomic approachcombining two-dimensional gel electrophoresis (2-DE) andmass spectrometry (MS) to compare the proteomes ofnormal liver from wild-type and transgenic NME1-nullmice. We have focused on liver cytosolic proteins firstbecause NME1 is mainly and abundantly expressed in thecytosol, second because the cytosol comprises at least 50%of all liver proteins, and third because the separation andfurther identification of membrane proteins by 2-DE andMS remains technically challenging. In this study, wereport that the steady-state levels of annexin IV and EF-
408 Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419

1Bα proteins are reduced in the liver cytosol of NME1-nullmice when compared to wild type. There was no change inthe corresponding messenger RNA (mRNA) levels forthese two proteins or in total protein levels in the livers,indicating that NME1 controls annexin IV and EF-1Bαprotein levels by post-translational mechanisms regulatingsubcellular localization. Our data suggest for the first timethat NME1 might control annexin IV and EF-1Bαsubcellular trafficking.
Materials and methods
Animals
Wild-type (NME1+/+) and transgenic NME1-null male mice(NME1−/−) of the 129 Sv genetic background were used at4–5 months of age. Three mice were used for eachexperimental condition. The livers were removed, snap-frozen in liquid nitrogen, and stored at −80°C until use. Allanimals were handled in compliance with French ethicalguidelines for the care and use of laboratory animals.
Protein preparation
Total liver extracts were prepared by homogenizing liversamples with a Dounce homogenizer in cold RIPA buffer[150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate,0.1% sodium dodecyl sulfate (SDS), 50 mM Tris–HCl,pH 7.5, 1 mM phenylmethanesulfonylfluoride (PMSF),1 mM Na3VO4, and 10 mg/ml leupeptin] and centrifuged at10,000×g at 4°C for 10 min. To prepare cytosolic extracts,liver tissues were homogenized on ice with a Potter–Elvehjem apparatus in 10 mM Tris–HCl (pH 7.4), 0.25 Msucrose, 1 mM EDTA, 1 mM PMSF, and 10 mg/mlleupeptin. The cytosolic fractions were collected byultracentrifugation (100,000×g at 4°C for 1 h).
Two-dimensional gel electrophoresis of cytosolicliver proteins
Proteins of liver cytosolic extracts were precipitated in coldacetone for 3 h on ice. After centrifugation, the pellets weredissolved in 6 M urea, 2 M thiourea, 2% (w/v) CHAPS,0.3% (w/v) dithiothreitol (DTT) (all from Sigma, St. Louis,MO, USA), 0.5% IPG buffer (Amersham Biosciences,Uppsala Sweden), 5 mM N-ethylmaleinimide, 2 mM PMSFand 1 mg of Orange G (Sigma). 2-DE was performed withimmobilized pH gradient (IPG) gels (180-mm ImmobilineDryStrips, pH 3–10 NL or 4–7, Amersham Biosciences) forthe first dimension, and with SDS polyacrylamide gelelectrophoresis (SDS-PAGE) 12% for the second dimen-sion. Approximately 1.5 mg of protein was applied to IPG
gels, and isoelectric focusing was carried out in an IPGphorapparatus (Amersham Biosciences). Voltage started at 30 Vand was gradually increased to 8,000 until 50,000 Vh.Strips were equilibrated by two 15-min incubation steps,the first in equilibration buffer containing 6 M urea, 30%(v/v) glycerol (Sigma), 2% (w/v) SDS (Bio-Rad) in 50 mMTris–HCl buffer, pH 6.8, supplemented with 5 mg/mL DTT,and the second in 45 mg/mL iodoacetamide (Sigma)containing a trace of bromophenol blue. For the seconddimension, the strips were applied to the cathodic end ofSDS-PAGE gels, which were run for 4 h at 80 mA. Afterstaining with colloidal Coomassie blue (CCB) as describedby Neuhoff et al. (1988), 2-DE gels were analyzed with theImage Master 4.01 software package from AmershamBiosciences.
Comparative analyses
Each spot volume (optical density × spot area) was normalizedagainst the total volume of all spots present in the gel.Normalized spot volumes in gels prepared with cytosolicextracts of NME1+/+ mice (three gels per mouse) werecompared with the matched spot volumes in thecorresponding gels prepared with extracts from NME1−/−
mice (three gels per mouse). Differential spots were selectedif considering at least 100% of normalized volume variation(volume ratios set at 0.5 and 2.0) between matching spots inall three comparative experiments. AWilcoxon test was usedto determine spots that have a significant variation in thenormalized volume between NME1+/+ and NME1−/−. AP<0.05 was considered significant.
In-gel trypsin digestion
Spots of interest were excised from 2-DE gels and digestedwith bovine trypsin (sequencing grade, EC 3.4.21.4, Roche,Mannheim, Germany) as described by Shevchenko et al.(1996) overnight at 37°C. The resulting peptide wereextracted by incubation in 1% formic acid (FA) for15 min, followed by two extractions with 1% FA/acetonitrile (ACN) 1:1 (v/v) and ACN, respectively. Peptideextracts were pooled and dried in a vacuum centrifugebefore solubilization in 10 μL of 1% FA, then desalted withZipTip® C18 from Millipore (Bedford, MA, USA). Thedesalted peptides were dried and stored at −20°C until use.
MALDI-TOF MS and peptide mass fingerprinting
The matrix was a saturated solution of α-cyano-4-hydroxycin-namic acid (Sigma) in 0.1% aqueous trifluoroacetic acid/ACN(1:1, v/v). After dissolution in 3 μL of 1% FA, the peptidesand matrix were mixed on the target (2:1, v/v) and air-dried.Matrix-assisted laser desorption/ionization time of flight
Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419 409

(MALDI-TOF) spectra were acquired with a Voyager-DESTR biospectrometry workstation mass spectrometer (AppleraApplied Biosystems Inc., Framingham, MA, USA) aspreviously described (Bruneel et al. 2005). Data mining wasperformed with ProFound software (http://129.85.19.192/profound_bin/), with the following parameters: mouse spe-cies, molecular weight ranges within ±25% of measuredvalues, pI range from 0 to 14.0, monoisotopic peptide masses,two missed cleavages by trypsin, and mass deviation of50 ppm. Partial chemical modifications such as methionineoxidation and cysteine carbamidomethylation were taken intoaccount for the queries. The peptide mass fingerprinting(PMF) analysis-based identifications were validated whenobtaining at least four peptide matches and coverage of 20%.
Liquid chromatography–tandem mass spectrometry
Nanoscale capillary liquid chromatography–tandem massspectrometry (LC-MS/MS) was used to analyze trypsindigests. After dilution in 6 μL of buffer (H2O/ACN/FA, 96/4/0.1, v/v), the trypsin digests were analysed with a capillaryliquid chromatography system (Famos-Switchos-UltiMateDionex, LC Packings, Amsterdam, The Netherlands) coupledwith a hybrid nanoESI Quadrupole-time of flight massspectrometer (Q-Tof2, Waters, Micromass, Manchester, UK)as previously described (Bruneel et al. 2005). Data acquisitionwas in automatic mode, switching between the surveyacquisition in MS mode and fragmentation acquisition inMS–MS mode on the four most intense ions detected in theformer survey scan. Mass data were converted into a PKLfile by using Masslynx™ software (Micromass, Manchester,UK), before being submitted to Mascot search software(http://www.matrixscience.com/). Proteins were identified bycomparing the experimental data to the NCBI non-redundantmouse database and were validated when obtaining at leasttwo peptide sequences per protein.
Western blotting
Proteins were separated by SDS-PAGE and transferred topolyvinylidene fluoride for EF-1Bα and NME1 detectionor to nitrocellulose membranes for the other proteins. Afterblocking with 5% non-fat milk, the membranes wereprobed with the following antibodies: rabbit polyclonalanti-EF-1Bα (1/2,500, gift from Dr. J. Dijk), goatpolyclonal anti-PAH (1/100, Santa Cruz BiotechnologyInc., Santa Cruz, CA, USA), sheep polyclonal anti-annexinIV (1/3,000, gift from Dr. M. Kaetzel), rabbit polyclonalanti-14-3-3 epsilon protein (1/200, Santa Cruz Biotechnol-ogy Inc.), rabbit polyclonal anti-NME1 (Boissan et al.2005) and mouse monoclonal anti-β-actin (1/5,000,Sigma). Immune complexes were visualized by enhanced
chemiluminescence (Amersham Biosciences, Saclay,France) after secondary reaction with peroxidase-conjugated anti-goat (Santa Cruz Biotechnology Inc.),anti-rabbit (Cell Signaling Technology Inc., Beverly, MA,USA), anti-mouse (Cell Signaling Technology Inc.), oranti-sheep antibodies (gift from Dr. M. Kaetzel).
Histology and immunohistochemistry
Mouse liver samples were fixed in 4% formaldehydesolution and embedded in paraffin. Four-micrometer tissuesections were stained with hematoxylin–phloxine and siriusred for histological examination. Immunohistochemicalstaining for EF-1Bα and calnexin was performed onparaffin sections. Staining for annexin IV was performedon frozen sections. Before immunohistochemistry, anavidin/biotin kit (Vector Laboratories, Burlingame, CA,USA) was used to prevent nonspecific labeling. Immuno-labeling was performed using a SuperSensitive link-labelimmunohistochemistry detection system (Biogenex, SanRamon, CA, USA) according to the manufacturer’sprotocol with specific antibodies against EF-1Bα (1:750,gift from Dr. J. Dijk), calnexin (1:100, Abcam Inc., SanFrancisco, CA, USA), and annexin IV (1:250, AbcamInc.). Microwave antigen retrieval in 10 mM citrate buffer(pH 6) was performed before EF-1Bα and calnexinimmunostaining. Peroxidase activity was revealed with3-amino-9-ethyl-carbazole. Slides were counterstainedwith hematoxylin.
Blood assays
Serum concentrations of aspartate aminotransferase, alanineaminotransferase, alkaline phosphatase, total bilirubin, andalbumin were measured on an Olympus AU 640 analyserusing Bio-Mérieux reagents.
Reverse transcription and real-time polymerasechain reaction
Total RNAwas extracted from liver samples using RNA Pluslysis solution (Quantum, Montreuil-sous-Bois, France), and2 μg of the extracts was reverse-transcribed by extension ofrandom decamers with MMLV reverse transcriptase (RET-ROscript™, Ambion Inc.). Real-time PCR was performedwith the TaqMan system (Applied Biosystem, Foster City,CA, USA) and the SYBR green master mix (AppliedBiosystem). The following primers were designed fromGenBank murine complementary DNA (cDNA) sequenceswith Primer Express software v1.5 (PE Applied Biosystem):EF-1Bα forward, 5′- ACGATTACCTGGCGGACAAG -3′and reverse 5′- CAGGTCAGCAGGTGGTGGA -3′; PAH
410 Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419

forward 5′- TATGGAGCCGAACTGGATGC-3′ and reverse5′- CCATGGCGGTAGTTGTAGGC-3′; annexin IV forward5′- TAGAGCCTTTCTCCGCACCA -3′ and reverse 5′-GAAGCAGCTTTCACGGTTCC -3′. An 18S rRNATaqManreagent was used as internal control. One-step RT-PCR wasused for both the target genes and the endogenous controls.Duplicate CT values were analyzed with Microsoft Excel,using the comparative CT (CT) method as described by themanufacturer (Applied Biosystem). The amount of target(2−CT) was normalized to 18S.
Phenylalanine catabolism
Animals were injected intraperitoneally with 1 mg ofL-phenylalanine per gram of body weight, using an aqueousstock solution of 25 mg/mL phenylalanine (Sigma). Plasmawas obtained by centrifugation of whole blood at varioustimes after phenylalanine challenge. After deproteinizationwith sulfosalicylic acid, phenylalanine concentrationswere measured with a JLC-500/V amino acid analyzer(JEOL Ltd, Tokyo, Japan) based on ion-exchange chroma-tography followed by post-column detection withninhydrin.
Cell lines
MDA-MB-435-derived cell lines C-100 and H1-177 (giftfrom Dr. P. Steeg), liver (HepG2), colon (HCT8/S11), andbreast (MDA-MB-231) cancer cell lines were maintained inDulbecco’s modified Eagle’s medium supplemented with10% fetal bovine serum (FBS). Hepatoma cell lines PLC/PRF/5 and Mahlavu were maintained in MEM containingEarle’s salts, 1% nonessential amino acids, 1 mM sodiumpyruvate, and 10% FBS. Breast cancer cell lines SKBr3,BT474, and MCF7 were maintained as described byAmerican Type Culture Collection.
Results
2-DE liver cytosolic protein patterns of wild-typeand NME1-null mice
Figure 1 shows representative 2-DE patterns of CCB-stainedliver cytosolic proteins from NME1+/+ and NME1−/− mice.These patterns contained more than 360 protein spotsbetween pI 4.0 and 7.0 with a high level of reproducibility(i.e., percentages of matched spots between triplicate gelsfrom the two genotypes were always above 97%). Thenormalized volume of six spots was significantly lowerin 2-DE gels of liver cytosol from NME1−/− mice thanfrom NME1+/+ mice. Differential 2-DE experiments using abroad pI range (3.0–10.0) revealed no additional significantdifferences (Supplementary Figure 1). As detailed in Fig. 2and in Supplementary Figures 2a and 2b, the pI/Mw values(pH units/kDa) of these six spots were 6.8/17 (spot 1), 4.5/25(spot 2), 5.4/36 (spot 3), 5.87/52 (spot 4), 5.93/52 (spot 5),and 6.0/52 (spot 6). Spots 1, 4, and 5 were undetectable ingels of NME1−/− samples, whereas mean values of normalizedvolume ratios for spot 2, 3, and 6 (NME1+/+ vs. NME1−/−)were 4.77, 3.03, and 7.0, respectively (P<0.05).
MS-based identification of modulated protein spots
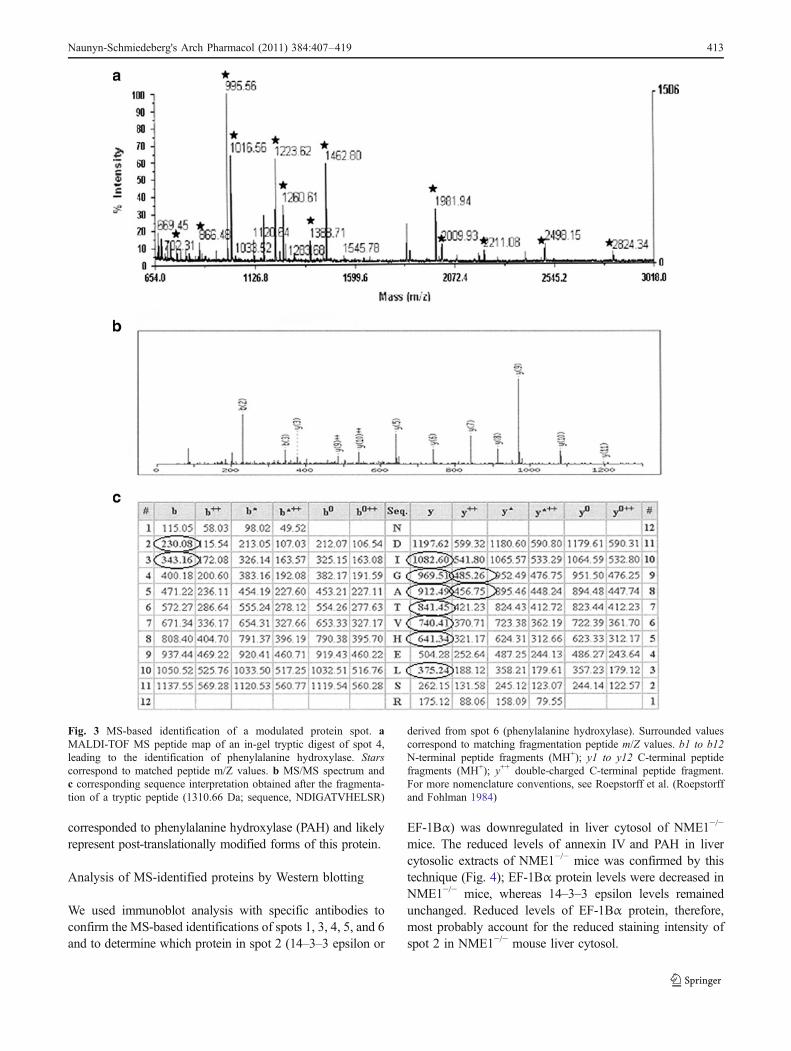
The proteins in the six spots of interest were identified byMALDI-TOF peptide mass fingerprinting and LC-MS/MSsequencing (Table 1). Five spots were unambiguouslyidentified by the two MS-based techniques, whereas spot 2corresponded to a mix of proteins that could only bedistinguished by MS/MS peptide sequencing (Fig. 3 andSupplementary Figure 3). Spot 1 was NME1/NDPK-A,absent, as expected, from the NME1−/− liver extracts. Spot2 was a mix of 14–3–3 epsilon protein and elongation factor1 Bα (EF-1Bα). Spot 3 was annexin IV. Spots 4, 5, and 6 all
Fig. 1 2-DE gels of wild-typeand NME1−/− mouse liver cyto-solic fractions. 2D patterns(pH 4–7) from NME1+/+ andNME1−/− mouse liver cytosolicextracts (1.5 mg of protein) weresimilar, with the exception of sixmodulated spots (arrows)
Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419 411
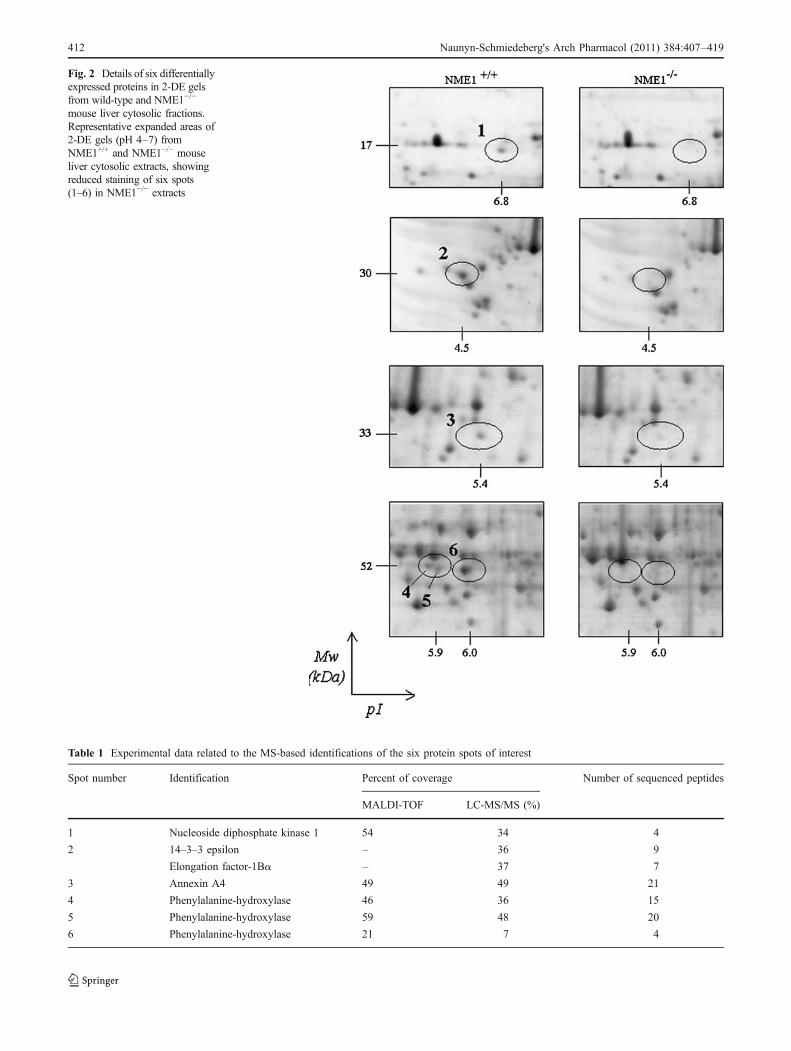
Fig. 2 Details of six differentiallyexpressed proteins in 2-DE gelsfrom wild-type and NME1−/−
mouse liver cytosolic fractions.Representative expanded areas of2-DE gels (pH 4–7) fromNME1+/+ and NME1−/− mouseliver cytosolic extracts, showingreduced staining of six spots(1–6) in NME1−/− extracts
Table 1 Experimental data related to the MS-based identifications of the six protein spots of interest
Spot number Identification Percent of coverage Number of sequenced peptides
MALDI-TOF LC-MS/MS (%)
1 Nucleoside diphosphate kinase 1 54 34 4
2 14–3–3 epsilon – 36 9
Elongation factor-1Bα – 37 7
3 Annexin A4 49 49 21
4 Phenylalanine-hydroxylase 46 36 15
5 Phenylalanine-hydroxylase 59 48 20
6 Phenylalanine-hydroxylase 21 7 4
412 Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419
corresponded to phenylalanine hydroxylase (PAH) and likelyrepresent post-translationally modified forms of this protein.
Analysis of MS-identified proteins by Western blotting
We used immunoblot analysis with specific antibodies toconfirm the MS-based identifications of spots 1, 3, 4, 5, and 6and to determine which protein in spot 2 (14–3–3 epsilon or
EF-1Bα) was downregulated in liver cytosol of NME1−/−
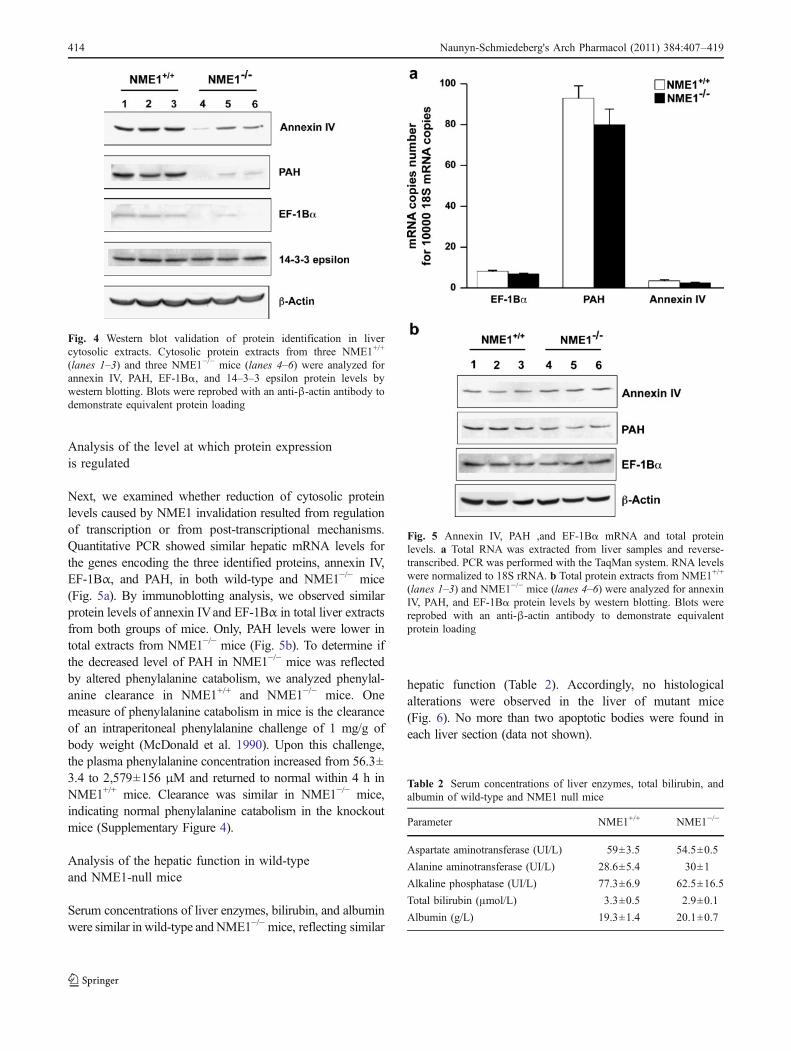
mice. The reduced levels of annexin IV and PAH in livercytosolic extracts of NME1−/− mice was confirmed by thistechnique (Fig. 4); EF-1Bα protein levels were decreased inNME1−/− mice, whereas 14–3–3 epsilon levels remainedunchanged. Reduced levels of EF-1Bα protein, therefore,most probably account for the reduced staining intensity ofspot 2 in NME1−/− mouse liver cytosol.
Fig. 3 MS-based identification of a modulated protein spot. aMALDI-TOF MS peptide map of an in-gel tryptic digest of spot 4,leading to the identification of phenylalanine hydroxylase. Starscorrespond to matched peptide m/Z values. b MS/MS spectrum andc corresponding sequence interpretation obtained after the fragmenta-tion of a tryptic peptide (1310.66 Da; sequence, NDIGATVHELSR)
derived from spot 6 (phenylalanine hydroxylase). Surrounded valuescorrespond to matching fragmentation peptide m/Z values. b1 to b12N-terminal peptide fragments (MH+); y1 to y12 C-terminal peptidefragments (MH+); y++ double-charged C-terminal peptide fragment.For more nomenclature conventions, see Roepstorff et al. (Roepstorffand Fohlman 1984)
Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419 413
Analysis of the level at which protein expressionis regulated
Next, we examined whether reduction of cytosolic proteinlevels caused by NME1 invalidation resulted from regulationof transcription or from post-transcriptional mechanisms.Quantitative PCR showed similar hepatic mRNA levels forthe genes encoding the three identified proteins, annexin IV,EF-1Bα, and PAH, in both wild-type and NME1−/− mice(Fig. 5a). By immunoblotting analysis, we observed similarprotein levels of annexin IVand EF-1Bα in total liver extractsfrom both groups of mice. Only, PAH levels were lower intotal extracts from NME1−/− mice (Fig. 5b). To determine ifthe decreased level of PAH in NME1−/− mice was reflectedby altered phenylalanine catabolism, we analyzed phenylal-anine clearance in NME1+/+ and NME1−/− mice. Onemeasure of phenylalanine catabolism in mice is the clearanceof an intraperitoneal phenylalanine challenge of 1 mg/g ofbody weight (McDonald et al. 1990). Upon this challenge,the plasma phenylalanine concentration increased from 56.3±3.4 to 2,579±156 μM and returned to normal within 4 h inNME1+/+ mice. Clearance was similar in NME1−/− mice,indicating normal phenylalanine catabolism in the knockoutmice (Supplementary Figure 4).
Analysis of the hepatic function in wild-typeand NME1-null mice
Serum concentrations of liver enzymes, bilirubin, and albuminwere similar in wild-type andNME1−/− mice, reflecting similar
hepatic function (Table 2). Accordingly, no histologicalalterations were observed in the liver of mutant mice(Fig. 6). No more than two apoptotic bodies were found ineach liver section (data not shown).
Fig. 5 Annexin IV, PAH ,and EF-1Bα mRNA and total proteinlevels. a Total RNA was extracted from liver samples and reverse-transcribed. PCR was performed with the TaqMan system. RNA levelswere normalized to 18S rRNA. b Total protein extracts from NME1+/+
(lanes 1–3) and NME1−/− mice (lanes 4–6) were analyzed for annexinIV, PAH, and EF-1Bα protein levels by western blotting. Blots werereprobed with an anti-β-actin antibody to demonstrate equivalentprotein loading
Fig. 4 Western blot validation of protein identification in livercytosolic extracts. Cytosolic protein extracts from three NME1+/+
(lanes 1–3) and three NME1−/− mice (lanes 4–6) were analyzed forannexin IV, PAH, EF-1Bα, and 14–3–3 epsilon protein levels bywestern blotting. Blots were reprobed with an anti-β-actin antibody todemonstrate equivalent protein loading
Table 2 Serum concentrations of liver enzymes, total bilirubin, andalbumin of wild-type and NME1 null mice
Parameter NME1+/+ NME1−/−
Aspartate aminotransferase (UI/L) 59±3.5 54.5±0.5
Alanine aminotransferase (UI/L) 28.6±5.4 30±1
Alkaline phosphatase (UI/L) 77.3±6.9 62.5±16.5
Total bilirubin (μmol/L) 3.3±0.5 2.9±0.1
Albumin (g/L) 19.3±1.4 20.1±0.7
414 Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419
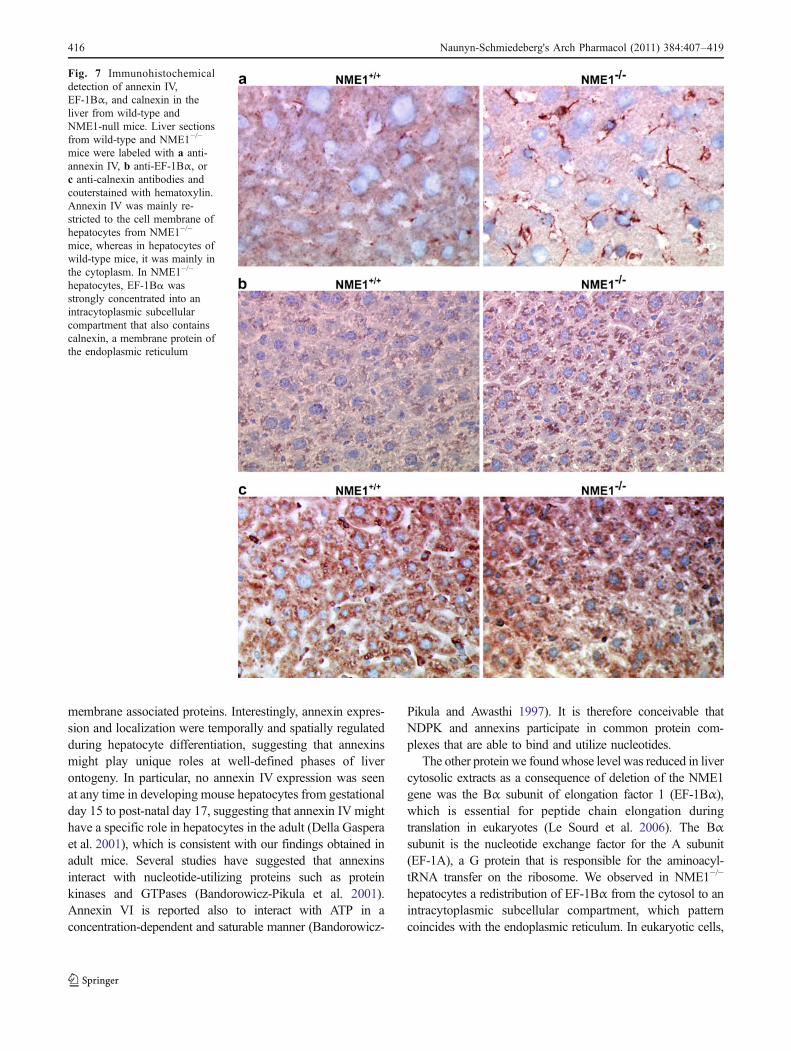
Immunohistochemical detection of annexin IV and EF-1Bαin the liver
Immunolabeling of liver sections showed that annexin IVappeared mainly in the cytoplasm with only faint, barelyperceptible membrane staining in wild-type hepatocytes. Insharp contrast, annexin IV immunostaining of hepatocytesfrom NME1−/− mice showed moderate to strong labeling ofthe plasma membrane (Fig. 7a). No nuclear annexin IVlabeling was observed. EF-1Bα immunostaining of hepato-cytes revealed differences between wild-type and null mice.Indeed, we observed that EF-1Bα labeling in NME1−/−
hepatocytes was concentrated in an intracytoplasmic subcel-lular compartment that also immunostained for calnexin, amembrane protein of the endoplasmic reticulum (Fig. 7b, c).
Correlation of NME1-regulated proteins with metastaticpotential and NME1 expression in a panel of carcinomacell lines
The expression of NME1 and annexin IV was assessed byimmunoblot analysis in a panel of human carcinoma cell lineswith various metastatic capacities. The H1-177 cell lineexpressed much higher levels of NME1 and exhibited reducedmotility in vitro and metastasis in vivo compared with vectortransfectant, C-100. Immunoblot analysis of these cell linesshowed that the NME1 transfectants retained higher levels ofannexin IV protein relative to the C-100 cell line (Supple-mentary Figure 5A). As shown in Supplementary Figure 5B,the non-invasive digestive carcinoma cell lines HCT8/S11,HepG2, and PLC/PRF/5 had higher levels of NME1 proteincompared with the highly invasive Mahlavu cell line. Therewas a positive correlation between levels of NME1 andannexin IV and thus an inverse correlation of both of theseproteins with the invasive potential of all the cell lines tested.Similar data were observed with breast carcinoma cell lines(Supplementary Figure 5C).
Discussion
In this study, we report that the steady-state levels ofannexin IV and EF-1Bα proteins are reduced in livercytosol in the absence of NME1. There was no change incorresponding mRNA levels or in the total protein levels inthe livers of NME1−/− mice, suggesting that NME1 controlsannexin IV and EF-1Bα protein levels by post-translationalmechanisms regulating subcellular location. Interestingly,annexin IV was concentrated mainly at the plasmamembrane of hepatocytes from NME1-null mice, whereasit appeared essentially cytoplasmic in wild-type hepatocytesand only rarely at the membrane. Taken together, these datasuggest that loss of NME1 induces a change in thesubcellular location of annexin IV in hepatocytes resultingin enrichment at plasma membrane domains. Annexin IV isfound predominantly in epithelial cells at both cytoplasmicand cell surface locations (Massey et al. 1991a,b; Mayran etal. 1996). A common property of annexin family proteins istheir Ca2+-dependent binding to various cellular mem-branes, with the plasma membrane being the preferredtarget (Gerke et al. 2005). These observations stronglysuggest that annexins play a role in membrane-relatedevents, particularly in membrane domain organization,exocytosis, and endocytosis (Rescher and Gerke 2004;Gerke et al. 2005). By forming networks on the membranesurface, annexins can function as organizers of membranedomains and membrane-recruitment platforms for proteinswith which they interact. It has been suggested that annexinnetworks might function in stabilizing certain plasmamembrane structures and/or changing membrane curvatureand therefore cell shape. The work of Piljic et al. providesmechanistic insights into how annexin IV may regulateplasma membrane protein function (Piljic and Schultz2006; Skrahina et al. 2008). These studies revealed theimmobile nature of annexin IV aggregates on the membranesurface, which, in turn, reduced the mobility of plasma
Fig. 6 Liver histology of wild-type and NME1-null mice. Livertissue samples were fixed in 4%formaldehyde solution andembedded in paraffin. Four-micrometer tissue sections werestained with hematoxylin andphloxin before histological ex-amination. No liver histologicalalterations were observed inmutant mice
Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419 415
membrane associated proteins. Interestingly, annexin expres-sion and localization were temporally and spatially regulatedduring hepatocyte differentiation, suggesting that annexinsmight play unique roles at well-defined phases of liverontogeny. In particular, no annexin IV expression was seenat any time in developing mouse hepatocytes from gestationalday 15 to post-natal day 17, suggesting that annexin IV mighthave a specific role in hepatocytes in the adult (Della Gasperaet al. 2001), which is consistent with our findings obtained inadult mice. Several studies have suggested that annexinsinteract with nucleotide-utilizing proteins such as proteinkinases and GTPases (Bandorowicz-Pikula et al. 2001).Annexin VI is reported also to interact with ATP in aconcentration-dependent and saturable manner (Bandorowicz-
Pikula and Awasthi 1997). It is therefore conceivable thatNDPK and annexins participate in common protein com-plexes that are able to bind and utilize nucleotides.
The other protein we found whose level was reduced in livercytosolic extracts as a consequence of deletion of the NME1gene was the Bα subunit of elongation factor 1 (EF-1Bα),which is essential for peptide chain elongation duringtranslation in eukaryotes (Le Sourd et al. 2006). The Bαsubunit is the nucleotide exchange factor for the A subunit(EF-1A), a G protein that is responsible for the aminoacyl-tRNA transfer on the ribosome. We observed in NME1−/−
hepatocytes a redistribution of EF-1Bα from the cytosol to anintracytoplasmic subcellular compartment, which patterncoincides with the endoplasmic reticulum. In eukaryotic cells,
Fig. 7 Immunohistochemicaldetection of annexin IV,EF-1Bα, and calnexin in theliver from wild-type andNME1-null mice. Liver sectionsfrom wild-type and NME1−/−
mice were labeled with a anti-annexin IV, b anti-EF-1Bα, orc anti-calnexin antibodies andcouterstained with hematoxylin.Annexin IV was mainly re-stricted to the cell membrane ofhepatocytes from NME1−/−
mice, whereas in hepatocytes ofwild-type mice, it was mainly inthe cytoplasm. In NME1−/−
hepatocytes, EF-1Bα wasstrongly concentrated into anintracytoplasmic subcellularcompartment that also containscalnexin, a membrane protein ofthe endoplasmic reticulum
416 Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419

it is generally accepted that protein synthesis is compartmen-talized. Soluble proteins are synthesized on free ribosomes,whereas secretory and membrane proteins are synthesized onER-bound ribosomes (Diehn et al. 2000; Nicchitta et al.2005). Accordingly, enrichment of EF-1Bα on the ER ofNME1-null hepatocytes may be related to an increase inmembrane protein synthesis, which cannot be appreciated byconventional two-dimensional gel electrophoresis. Futurestudies aimed at the determination of membrane proteincomposition in hepatocytes from wild-type and knockoutmice are therefore required. Assembly of EF-1 into ER isimportant for regulating protein synthesis in eukaryotic cells.Kinectin is the major membrane anchor for EF-1 complexinto ER (Ong et al. 2006). Indeed, the kinectin-dependentanchorage of EF-1 into ER regulates the efficiency ofmembrane and cytosolic protein synthesis. The disruption ofthe EF-1/kinectin interaction inhibited expression of mem-brane proteins but enhanced synthesis of cytosolic proteins.
PAH is the enzyme responsible for phenylalanine conver-sion to tyrosine in the presence of tetrahydrobiopterin (Huftonet al. 1995). Mutations that reduce the activity of PAH or thatof the enzymes involved in tetrahydrobiopterin synthesisresult in congenital disorders of phenylalanine catabolismsuch as phenylketonuria and hyperphenylalaninemia (Scriver1995). We found that PAH was less abundant in both thetotal and cytosolic fractions of livers from NME1−/− micewhen compared with NME1+/+ mice. As RT-PCR showed nocorresponding loss at the mRNA level, we suspect an effectof NME1 on PAH protein synthesis and/or stability.Interestingly, phenylalanine catabolism was normal inNME1-null mice, possibly because residual PAH levels weresufficient to ensure tyrosine synthesis.
Our proteomic approach identified three proteins that werenot previously known to be linked to NME1, two of which,annexin IV and EF-1Bα, are involved in nucleotide bindingand/or exchange on specific targets and in regulating cellgrowth and differentiation. Previous studies have indicatedthat NME1 is a suppressor of metastasis. Might annexin IVand EF-1Bα also play a role in metastasis? Several changes inannexin expression have been reported during tumorigenesis;however, very little data exist on the role of annexin IV incancer. Annexin IV is upregulated in renal cell carcinoma andcould play a role in tumor dissemination by promoting cellmigration (Zimmermann et al. 2004). In a panel of differentcarcinoma cell line extracts, we found a positive correlationbetween NME1 and annexin IV expression, while in normalliver, we observed only a change of the subcellular locationof annexin IV depending on the NME1 status. Thus, anotherlevel of regulation is introduced by the tumoral state. Thisdiscrepancy and the correlative expression of NME1 andannexin IV in tumor cohorts deserve to be further investi-gated. Several studies reveal that annexin IV may play a role
in resistance to chemotherapy. When annexin IV cDNAwas transfected into 293T cells, the transfected cells weremore resistant to the chemotherapy drug paclitaxel thancontrol cells (Han et al. 2000). Similarly, annexin IVconferred resistance to carboplatin in ovarian cancer cells(Kim et al. 2009). Accordingly, colorectal cancer patientswith low annexin expression had a better survival thanpatients with high annexin IV expression (Duncan et al.2008). Paradoxically, annexin IV was less expressed in thecisplatin-resistant ovarian cell line IGROV1-R10 in compari-son with the cisplatin-sensitive counterpart (Le Moguen et al.2006). This last result was consistent with the fact thatannexin IV was downregulated in hormone refractory prostatecancer (Xin et al. 2003). Interestingly, annexin IV is regulatedby Tiam1, the specific exchange nucleotide factor of Rac1,which has been shown to bind directly to NME1 (Otsuki et al.2001; Liu et al. 2008). This new link could provide insightinto the mechanism of NME1 in metastasis. The geneencoding annexin IV is a direct transcriptional target of p53;its expression is enhanced by wild-type p53 in ovarianadenocarcinoma cells (Masuishi et al. 2011). Interestingly,NME1 and one of its interacting partners STRAP, a TGF-βreceptor-interacting protein, physically interact with p53 andpositively regulate its functions, including p53-inducedapoptosis and cell cycle arrest. Consistent with this, silencingof NME1 induced down-regulation of p53-mediated tran-scription (Seong et al. 2007). The same group, also throughNME1-silencing experiments, identifies an interactionbetween NME1 and the macrophage migration inhibitoryfactor (MIF) to alleviate theMIF-mediated suppression of p53activity (Jung et al. 2008). Some EF-1 subunits appear tohave oncogenic potential, but the time course of EF-1expression during metastatic dissemination is poorly docu-mented (Anand et al. 2002; Joseph et al. 2002). It isnoteworthy that EF-1A overexpression is associated with themetastatic potential of mammary adenocarcinoma (Edmondset al. 1996). The cancer-related role of the Bα subunit ispoorly documented. Recent data indicate that the individualsubunits can exist separately from the EF-1B complex incancer tissues and that disintegration of EF-1B could be animportant sign of cancer development (Veremieva et al.2011).
Our observations indicate that NME1 might participateregulating the subcellular location of annexin IV and EF-1Bα in hepatocytes, thus conferring on NME1 a newfunction in the liver. Further studies of the proteins that aremodulated by NME1 may provide new insights into themolecular mechanisms by which NME1 acts as a metastasissuppressor. In particular, it will be interesting to see if theexpression of these proteins is also altered in mouse andhuman tumors (especially liver tumors) and if they areinvolved in metastatic dissemination.
Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419 417

Acknowledgments We are very grateful to Drs. J. Dijk and M.Kaetzel for the gift of anti-EF-1Bα and anti-annexin IV antibodies,respectively, to Dr. V. Barbu for advice on real-time PCR, and to Dr.N. Chignard for helpful comments. This work was supported by theInstitut National de la Santé et de la Recherche Médicale (INSERM),the Université Pierre et Marie Curie (UPMC), and grants (to MLL)from the Groupement des Entreprises Françaises contre le Cancer(GEFLUC) and from the Association pour la Recherche contre leCancer (ARC).
References
Anand N, Murthy S, Amann G, Wernick M, Porter LA, Cukier IH,Collins C, Gray JW, Diebold J, Demetrick DJ, Lee JM (2002)Protein elongation factor EEF1A2 is a putative oncogene inovarian cancer. Nat Genet 31:301–305
Bandorowicz-Pikula J, Awasthi YC (1997) Interaction of annexins IVand VI with ATP. An alternative mechanism by which a cellularfunction of these calcium- and membrane-binding proteins isregulated. FEBS Lett 409:300–306
Bandorowicz-Pikula J, Buchet R, Pikula S (2001) Annexins asnucleotide-binding proteins: facts and speculations. Bioessays23:170–178
Baughman C, Morin-Leisk J, Lee T (2008) Nucleoside diphosphatekinase B (NDKB) scaffolds endoplasmic reticulum membranes invitro. Exp Cell Res 314:2702–2714
Boissan M, Wendum D, Arnaud-Dabernat S, Munier A, Debray M,Lascu I, Daniel JY, Lacombe ML (2005) Increased lungmetastasis in transgenic NM23-Null/SV40 mice with hepatocel-lular carcinoma. J Natl Cancer Inst 97:836–845
Boissan M, Dabernat S, Peuchant E, Schlattner U, Lascu I, LacombeML (2009) The mammalian Nm23/NDPK family: from metas-tasis control to cilia movement. Mol Cell Biochem 329:51–62
Boissan M, De Wever O, Lizarraga F, Wendum D, Poincloux R,Chignard N, Desbois-Mouthon C, Dufour S, Nawrocki-Raby B,Birembaut P, Bracke M, Chavrier P, Gespach C, Lacombe ML(2010) Implication of metastasis suppressor NM23-H1 in main-taining adherens junctions and limiting the invasive potential ofhuman cancer cells. Cancer Res 70:7710–7722
Bruneel A, Labas V, Mailloux A, Sharma S, Royer N, Vinh J, PernetP, Vaubourdolle M, Baudin B (2005) Proteomics of humanumbilical vein endothelial cells applied to etoposide-inducedapoptosis. Proteomics 5:3876–3884
Dammai V, Adryan B, Lavenburg KR, Hsu T (2003) Drosophila awd,the homolog of human nm23, regulates FGF receptor levels andfunctions synergistically with shi/dynamin during tracheal develop-ment. Genes Dev 17:2812–2824
Della Gaspera B, Braut-Boucher F, Bomsel M, Chatelet F, Guguen-Guillouzo C, Font J, Weinman J, Weinman S (2001) Annexinexpressions are temporally and spatially regulated during rathepatocyte differentiation. Dev Dyn 222:206–217
Diehn M, Eisen MB, Botstein D, Brown PO (2000) Large-scaleidentification of secreted and membrane-associated gene productsusing DNA microarrays. Nat Genet 25:58–62
Duncan R, Carpenter B, Main LC, Telfer C, Murray GI (2008)Characterisation and protein expression profiling of annexins incolorectal cancer. Br J Cancer 98:426–433
Edmonds BT, Wyckoff J, Yeung YG, Wang Y, Stanley ER, Jones J,Segall J, Condeelis J (1996) Elongation factor-1 alpha is anoverexpressed actin binding protein in metastatic rat mammaryadenocarcinoma. J Cell Sci 109(Pt 11):2705–2714
Engel M, Veron M, Theisinger B, Lacombe ML, Seib T, Dooley S,Welter C (1995) A novel serine/threonine-specific protein
phosphotransferase activity of Nm23/nucleoside-diphosphatekinase. Eur J Biochem 234:200–207
Fan Z, Beresford PJ, Oh DY, Zhang D, Lieberman J (2003) Tumorsuppressor NM23-H1 is a granzyme A-activated DNase duringCTL-mediated apoptosis, and the nucleosome assembly proteinSET is its inhibitor. Cell 112:659–672
Fournier HN, Dupe-Manet S, Bouvard D, Lacombe ML, Marie C,Block MR, Albiges-Rizo C (2002) Integrin cytoplasmic domain-associated protein 1alpha (ICAP-1alpha ) interacts directly withthe metastasis suppressor nm23-H2, and both proteins aretargeted to newly formed cell adhesion sites upon integrinengagement. J Biol Chem 277:20895–20902
Gallagher BC, Parrott KA, Szabo G, de S Otero A (2003) Receptoractivation regulates cortical, but not vesicular localization ofNDP kinase. J Cell Sci 116:3239–3250
Gerke V, Creutz CE, Moss SE (2005) Annexins: linking Ca2+
signalling to membrane dynamics. Nat Rev Mol Cell Biol6:449–461
Han EK, Tahir SK, Cherian SP, Collins N, Ng SC (2000) Modulationof paclitaxel resistance by annexin IV in human cancer cell lines.Br J Cancer 83:83–88
Hartsough MT, Morrison DK, Salerno M, Palmieri D, Ouatas T, MairM, Patrick J, Steeg PS (2002) Nm23-H1 metastasis suppressorphosphorylation of kinase suppressor of Ras via a histidineprotein kinase pathway. J Biol Chem 277:32389–32399
Hufton SE, Jennings IG, Cotton RG (1995) Structure and function of thearomatic amino acid hydroxylases. Biochem J 311(Pt 2):353–366
Iwashita S, Fujii M, Mukai H, Ono Y, Miyamoto M (2004) Lbc proto-oncogene product binds to and could be negatively regulated bymetastasis suppressor nm23-H2. Biochem Biophys Res Commun320:1063–1068
Joseph P, Lei YX, Whong WZ, Ong TM (2002) Oncogenic potentialof mouse translation elongation factor-1 delta, a novel cadmium-responsive proto-oncogene. J Biol Chem 277:6131–6136
Jung H, Seong HA, Ha H (2008) Direct interaction between NM23-H1 and macrophage migration inhibitory factor (MIF) is criticalfor alleviation of MIF-mediated suppression of p53 activity. JBiol Chem 283:32669–32679
Kaetzel DM, Zhang Q, Yang M, McCorkle JR, Ma D, Craven RJ(2006) Potential roles of 3′–5′ exonuclease activity of NM23-H1in DNA repair and malignant progression. J Bioenerg Biomembr38:163–167
Kapetanovich L, Baughman C, Lee TH (2005) Nm23H2 facilitatescoat protein complex II assembly and endoplasmic reticulumexport in mammalian cells. Mol Biol Cell 16:835–848
Kim A, Enomoto T, Serada S, Ueda Y, Takahashi T, Ripley B,Miyatake T, Fujita M, Lee CM, Morimoto K, Fujimoto M,Kimura T, Naka T (2009) Enhanced expression of Annexin A4 inclear cell carcinoma of the ovary and its association withchemoresistance to carboplatin. Int J Cancer 125:2316–2322
Krishnan KS, Rikhy R, Rao S, Shivalkar M, Mosko M, Narayanan R,Etter P, Estes PS, Ramaswami M (2001) Nucleoside diphosphatekinase, a source of GTP, is required for dynamin-dependentsynaptic vesicle recycling. Neuron 30:197–210
Le Moguen K, Lincet H, Deslandes E, Hubert-Roux M, Lange C,Poulain L, Gauduchon P, Baudin B (2006) Comparativeproteomic analysis of cisplatin sensitive IGROV1 ovariancarcinoma cell line and its resistant counterpart IGROV1-R10.Proteomics 6:5183–5192
Le Sourd F, Boulben S, Le Bouffant R, Cormier P, Morales J, Belle R,Mulner-Lorillon O (2006) eEF1B: At the dawn of the 21stcentury. Biochim Biophys Acta 1759:13–31
Liu L, Wang S, Zhang Q, Ding Y (2008) Identification of potentialgenes/proteins regulated by Tiam1 in colorectal cancer bymicroarray analysis and proteome analysis. Cell Biol Int32:1215–1222
418 Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419

Massey D, Traverso V, Maroux S (1991a) Lipocortin IV is abasolateral cytoskeleton constituent of rabbit enterocytes. J BiolChem 266:3125–3130
Massey D, Traverso V, Rigal A, Maroux S (1991b) Cellular andsubcellular localization of annexin IV in rabbit intestinalepithelium, pancreas and liver. Biol Cell 73:151–156
Masuishi Y, Arakawa N, Kawasaki H, Miyagi E, Hirahara F, Hirano H(2011) Wild-type p53 enhances annexin IV gene expression inovarian clear cell adenocarcinoma. Febs J. doi:10.1111/j.1742-4658.2011.08059.x.
Mayran N, Traverso V, Maroux S, Massey-Harroche D (1996)Cellular and subcellular localizations of annexins I, IV, and VIin lung epithelia. Am J Physiol 270:L863–L871
McDonald JD, Bode VC, Dove WF, Shedlovsky A (1990) Pahhph-5:a mouse mutant deficient in phenylalanine hydroxylase. ProcNatl Acad Sci USA 87:1965–1967
Melki R, Lascu I, Carlier MF, Veron M (1992) Nucleosidediphosphate kinase does not directly interact with tubulin normicrotubules. Biochem Biophys Res Commun 187:65–72
Mitchell KA, Szabo G, de S Otero A (2009) Direct binding ofcytosolic NDP kinases to membrane lipids is regulated bynucleotides. Biochim Biophys Acta 1793:469–476
Murakami M, Meneses PI, Knight JS, Lan K, Kaul R, Verma SC,Robertson ES (2008a) Nm23-H1 modulates the activity of theguanine exchange factor Dbl-1. Int J Cancer 123:500–510
Murakami M, Meneses PI, Lan K, Robertson ES (2008b) Thesuppressor of metastasis Nm23-H1 interacts with the Cdc42Rho family member and the pleckstrin homology domain ofoncoprotein Dbl-1 to suppress cell migration. Cancer Biol Ther7:677–688
Nallamothu G, Woolworth JA, Dammai V, Hsu T (2008) Awd, thehomolog of metastasis suppressor gene Nm23, regulates Dro-sophila epithelial cell invasion. Mol Cell Biol 28:1964–1973
Neuhoff V, Arold N, Taube D, Ehrhardt W (1988) Improved stainingof proteins in polyacrylamide gels including isoelectric focusinggels with clear background at nanogram sensitivity usingCoomassie Brilliant Blue G-250 and R-250. Electrophoresis9:255–262
Nicchitta CV, Lerner RS, Stephens SB, Dodd RD, Pyhtila B (2005)Pathways for compartmentalizing protein synthesis in eukaryoticcells: the template-partitioning model. Biochem Cell Biol83:687–695
Ong LL, Lin PC, Zhang X, Chia SM, Yu H (2006) Kinectin-dependent assembly of translation elongation factor-1 complexon endoplasmic reticulum regulates protein synthesis. J BiolChem 281:33621–33634
Otsuki Y, Tanaka M, Yoshii S, Kawazoe N, Nakaya K, Sugimura H(2001) Tumor metastasis suppressor nm23H1 regulates Rac1GTPase by interaction with Tiam1. Proc Natl Acad Sci USA98:4385–4390
Piljic A, Schultz C (2006) Annexin A4 self-association modulatesgeneral membrane protein mobility in living cells. Mol Biol Cell17:3318–3328
Postel EH (2003) Multiple biochemical activities of NM23/NDPkinase in gene regulation. J Bioenerg Biomembr 35:31–40
Rescher U, Gerke V (2004) Annexins—unique membrane bindingproteins with diverse functions. J Cell Sci 117:2631–2639
Roepstorff P, Fohlman J (1984) Proposal for a common nomenclaturefor sequence ions in mass spectra of peptides. Biomed MassSpectrom 11:601
Salerno M, Palmieri D, Bouadis A, Halverson D, Steeg PS (2005)Nm23-H1 metastasis suppressor expression level influences thebinding properties, stability, and function of the kinase suppres-sor of Ras1 (KSR1) Erk scaffold in breast carcinoma cells. MolCell Biol 25:1379–1388
Scriver CR (1995) Whatever happened to PKU? Clin Biochem28:137–144
Seong HA, Jung H, Ha H (2007) NM23-H1 tumor suppressorphysically interacts with serine-threonine kinase receptor-associated protein, a transforming growth factor-beta (TGF-beta)receptor-interacting protein, and negatively regulates TGF-betasignaling. J Biol Chem 282:12075–12096
Shevchenko A, Wilm M, Vorm O, Mann M (1996) Mass spectrometricsequencing of proteins silver-stained polyacrylamide gels. AnalChem 68:850–858
Skrahina T, Piljic A, Schultz C (2008) Heterogeneity and timing oftranslocation and membrane-mediated assembly of differentannexins. Exp Cell Res 314:1039–1047
Steeg PS, Bevilacqua G, Kopper L, Thorgeirsson UP, Talmadge JE,Liotta LA, Sobel ME (1988) Evidence for a novel geneassociated with low tumor metastatic potential. J Natl CancerInst 80:200–204
Steeg PS, Palmieri D, Ouatas T, Salerno M (2003) Histidine kinasesand histidine phosphorylated proteins in mammalian cell biology,signal transduction and cancer. Cancer Lett 190:1–12
Tseng YH, Vicent D, Zhu J, Niu Y, Adeyinka A, Moyers JS, WatsonPH, Kahn CR (2001) Regulation of growth and tumorigenicity ofbreast cancer cells by the low molecular weight GTPase Rad andnm23. Cancer Res 61:2071–2079
Veremieva M, Khoruzhenko A, Zaicev S, Negrutskii B, El"skaya A(2011) Unbalanced expression of the translation complex eEF1subunits in human cardioesophageal carcinoma. Eur J Clin Invest41:269–276
Xin W, Rhodes DR, Ingold C, Chinnaiyan AM, Rubin MA (2003)Dysregulation of the annexin family protein family is associatedwith prostate cancer progression. Am J Pathol 162:255–261
Zhu J, Tseng YH, Kantor JD, Rhodes CJ, Zetter BR, Moyers JS, KahnCR (1999) Interaction of the Ras-related protein associated withdiabetes rad and the putative tumor metastasis suppressor NM23provides a novel mechanism of GTPase regulation. Proc NatlAcad Sci USA 96:14911–14918
Zimmermann U, Balabanov S, Giebel J, Teller S, Junker H, SchmollD, Protzel C, Scharf C, Kleist B, Walther R (2004) Increasedexpression and altered location of annexin IV in renal clear cellcarcinoma: a possible role in tumour dissemination. Cancer Lett209:111–118
Naunyn-Schmiedeberg"s Arch Pharmacol (2011) 384:407–419 419





























