Embed Size (px)
Citation preview

Research Journal of Biotechnology Vol. 8 (9) September (2013) Res. J. Biotech
(56)
Purification and Characterization of Laccase from newly isolated Thermophilic Brevibacillus sp. (Z1) and its
applications in removal of Textile Dyes Ceyda Bozoglu1, Ahmet Adiguzel1*, Hayrunnisa Nadaroglu2*, Derya Yanmis3 and Medine Gulluce3
1. Faculty of Science, Department of Molecular Biology and Genetic, Ataturk University, 25240, Erzurum, TURKEY 2. Department of Food Technology, Erzurum Vocational Training School, Ataturk University, 25240, Erzurum, TURKEY
3. Faculty of Science, Department of Biology, Ataturk University, 25240, Erzurum, TURKEY *[email protected], *[email protected]
Abstract Laccases are multicopper-containing oxidases (EC 1.10.3.2.) and they are widely found in fungi, higher plants and bacteria. Because of their high pH stability and thermostability, laccases from this strain have a potential for the treatment of textile bleaching effluents. In this research, the thermophilic test strain isolated from the Agri-Diyadin thermal springs, in the east of Turkey, were subjected to analysis of 16S rRNA sequence. The obtained sequence data were compared with data from the database in the Genebank and as a result it has been identified that this organism is a member of Brevibacillus genus (GenBank number KC292196). The laccase from Brevibacillus sp. (Z1) was purified by using precipitate of saturation (NH4)2SO4, DEAE-cellulose and Sephacryl S200. The purification degree and the molecular mass of the enzyme (93 and 110 kDa) were determined by SDS-PAGE and gel filtration chromatography. In order to determine the enzyme's resistance against the metal ions, the effect of metal ions such as Ca2+, Ba2+, Mn2+, Ni2+, Co2+, Cu2+, Zn2+, Fe2+, K+ and certain chemical substances such as EDTA and ascorbic acid on the pure laccase enzyme activity have been investigated and no significant change was observed. Vmax and Km values were calculated using Lineweaver Burk plots for ABTS, 2,6-dimetoxyphenol and guaiacol. Purified laccase enzyme was also used to remove some textile dyes which are used in textile industries and known to cause major environmental problems such as Reactive Black 5, Fuchsine, Allura Red and Acid Red 37 from waste water. Consequently, it is concluded that the purified laccase from Brevibacillus sp. (Z1) can successfully be used for the removal of the textile dyes from wastewater. Keywords: Laccase, Brevibacillus, Purification, Charac-terization, Removal textile dyes. Introduction The discovery of life at high temperatures and the isolation of Thermus aquaticus from the Yellowstone National park
were major steps in the field of biotechnology. Especially the success of T. aquaticus achieved in the commercial area has directed scientists towards studies on isolation and identification of thermophilic microorganisms in geothermal areas, not identified or designated yet1. In the identification and characterization of unknown microorganisms of different ecosystems, the classical methods (morphological, physiological and biochemical) had been used until the molecular methods become available. Today, however, it has been understood that the identification and characterization of the microorganisms by classical methods is not enough and this paved the way for rapid developments in molecular biology2. After the thermostable properties of the thermophilic microorganisms have been identified, in line with the identification and characterization studies, researches on revealing the potential industrial uses of theses enzymes which were purified from these organisms and having economic importance, were increased significantly2. The dyes, which are important industrially and used in diverse applications in different industrial fields such as cosmetics, food, paper and textile, have negative impacts on the photosynthetic activity, due to the decrease in luminous transmittance and this has toxic effects on living organisms3,4. A large portion of these synthetic dyes are azo dyes which are toxic, mutagenic and carcinogenic5. These dyes have the potential to accumulate in the environment at higher levels since they are resistant to degradation due to their azo bonds. Treatment of these dyes is difficult with the classical treatment methods, since they present a stable structure in acidic and alkaline conditions and resistant to aerobic degradation, heat and light6. Industrially important laccase enzyme (EC 1.1.3.2, p-diphenol, dioxygen oxidoreductase) belongs to a gene family named as the blue-copper proteins of the oxidases7 and is produced by 4 type of living organisms including bacteria, insects, higher plants and fungi8. So far, very few bacterial origin laccase enzymes have been purified and characterized. The first study on this subject is the prokaryotic laccase which was derived from the Azospirillum lipoferum, a rizospheric bacterium. It was determined in this bacterium that the enzyme was

Research Journal of Biotechnology Vol. 8 (9) September (2013) Res. J. Biotech
(57)
composed of a catalytic subunit and is a multimeric enzyme, consisting of 1 or 2 larger chains.9
Laccase enzyme was purified from the Marinomonas mediterranea, which is a melanogenic marine bacterium. It has also been determined that these bacteria produce two different polyphenol oxidase, capable of oxidizing the substrates of both laccases and tyrosinases9. In a study conducted recently, Ruijssenaars and Hartmans10 have identified a protein of Bacillus halodurans encoded by ORF bh2082 as a potential bacterial laccase. There are very few studies performed with thermophilic bacteria. Recent studies have been conducted with thermophilic organisms including Laccase derived from Streptomyces lavendula REN-711, laccase-like protein derived from Pyrobaculum aerophilum IM2, which is a thermophilic archaea12, laccase 31-like protein derived from Aquifex aeolicus VF5 which is a hyperthermophilic bacterium13 and laccase derived from Thermus thermophilus9,14 and Bacillus pumilus. Laccase mediated catalysis occurs with reduction of oxygen to water, accompanied by the oxidation of substrate. Laccases thus oxidize polyphenols, methoxy-substituted phenols, aromatic diamines and a range of other compounds15. Consequently, it is possible to use the laccase enzyme in many industrial areas such as the removal of textile dyes, phenols removal and waste detoxification, since it does not have the substrate specificity. For this reason, purification and characterization of the laccase enzyme produced intracellularly by Brevibacillus sp. (Z1) bacteria (Genbank number KC292196) which were isolated in the Agri-Diyadin thermal springs and investigating the usability of this enzyme in removing textile dyes in wastewater were aimed in this study. Material and Methods Materials: Bovine serum albumin (BSA), ABTS [2,2’-azino-bis(3-ethylbenzylthiazoline-6-sulfonic acid)], DEAE-sephedex, Sephacryl S-200, ethylene diamine tetra acetic acid (EDTA), dithioerythritol, ß-mercaptoethanol, tris(hydroxymethyl)aminomethane, hydrogen peroxide (H2O2), 4-methylcatechol, guaiacol, pyrogallol, hydroquinone, Reactive Black 5 (RB5), fuchsine (C20H19N3.HCl), allura red (C18H14N2Na2O8S2), Acid red 37 diammonium salt (C18H16N4O8S2.2H3N) and agents for SDS-PAGE were purchased from Sigma (USA). Sodium acetate (CH3COONa), ammonium sulphate [(NH4)2SO4], sodium chloride (NaCl), BaCl2, CuCl2, MnCl2, CoCl2, ZnCl2, FeCl2, CaCl2, NiCl2 HgCl2, ethylenediam-inetetraacetic acid (EDTA), sodium bicarbonate (NaHCO3), sodium acetate (CH3COONa), ascorbic acid (C6H8O6) and sodium hydrogen phosphate monohydrate (Na2HPO4.H2O) were purchased from Merck (Darmstadt, Germany). All other chemicals were of analytical grade. Isolation of strain: The thermophilic bacterial strain used
in this study was isolated from water and sludge samples of Diyadin Hotspring in provinces of Agri in Turkey. The water temperature of this hot spring was around 70 °C. The samples were streaked on Nutrient Agar (NA) plates and then incubated in aerobic and anaerobic incubators at 55-60 °C for 24-48 hours. After incubation, different colony developed in the media were selected and purified by subculturing. The purity of the test isolate was assessed by using colony morphology and microscopy. Isolated and purified bacterial strains were stored in Nutrient Broth containing 15% glycerol at -86 °C for further studies16. Morphological, physiological and biochemical characterizations of isolate: The temperature range for growth was determined by incubating strain from 35 to 80 °C with 1 °C of intervals. The pH dependence of growth was tested from pH 4.0 to pH 12.0. Cell morphology of isolates was investigated by light microscopy. The effect of NaCl on the thermophilic bacterial growth was studied in NB medium containing 2.0, 3.0, 4.0, 5.0, 8.0, 10.0% (w/v) NaCl. Cell and colony morphology, gram reaction, the presence of catalase, oxidase and amylase were investigated according to the methods described by Harley and Prescott17. DNA extraction from pure culture: Total genomic DNA was extracted from bacteria samples using a modified method previously described by Adiguzel2. PCR amplification and cloning of 16S rRNA gene fragment: The 16S rRNA gene was selectively amplified from purified genomic DNA by using oligonucleotide primers designed to anneal to conserved positions in the 3' and 5' regions of bacterial 16S rRNA genes. The forward primer, UNI16S-L (5'-ATTCTAGAGTTTGATCATGGCT CA-3'), corresponded to positions 11 to 26 of Escherichia coli 16S rRNA and the reverse primer, UNI16S-R (5'-AT GGTACCGTGTGACGGGCGGTGTGTA-3'), correspond-ed to the complement of positions 1411 to 1393 of Escherichia coli 16S rRNA18. PCR reaction conditions were carried out according to Beffa et al19 and the PCR product was cloned to a pGEM-T vector system (Promega, UK). Sequencing analysis: Following PCR amplification and cloning of the 16S rRNA gene of the strain, the 16S rRNA gene sequence with an Applied Biosystems model 373A DNA sequence was determined by using the ABI PRISM cycle sequencing kit (Macrogen, Korea). The sequence consisting of about 1425 nucleotides (nt) of the 16S rRNA gene was determined. This sequence was compared with those contained within GenBank20 by using a BLAST search21. The 16S rRNA gene sequences of the species most closely related to our strain were retrieved from the database16. Purification of Laccase from Brevibacillus sp. (Z1): The crude enzyme extract was filtered and centrifuged for 15

Research Journal of Biotechnology Vol. 8 (9) September (2013) Res. J. Biotech
(58)
min at 5000 rpm and then the ammonium sulphate enzyme homogenate was precipitated at 0-20%, 20-40%, 40-60%, 60-80% and 80-100% intervals. Then ammonium sulphate was used to saturate the obtained supernatant at 60-80%. After dissolving the precipitate in 20 mM of phosphate buffer (pH 5.0), it was dialyzed against the same buffer above. Anion exchange chromatography: After the suspension obtained by ammonium sulphate, precipitation was dialyzed, the 20 mM of phosphate buffer (pH 5.0) and previously equilibrated DEAE-cellulose ion exchange column (2.5 x 30) were studied. The column was washed until the eluate protein detection fails with the same buffer. Following this, NaCl gradient from 0 up to 1 M was applied to elute the proteins attached to the column. The flow rates of the collected fractions were 3 ml with a 3 mL/min. The absorbance of protein elution was spectrophotometrically measured at 280 nm using ABTS substrate to measure the activity in the fractions. The active fractions were pooled (combined) and allowed to stand at 4 °C22. Gel filtration: Using an Amicon membrane concentrator with a 10 kDa cut off, the active fractions which have been obtained from anion exchange column were combined, dialyzed, desalted and concentrated. Obtained enzyme solution was applied to Sephacryl S-200 column (120 cm × 1 cm), which has been pre-equilibrated with 20 mM of phosphate buffer (pH:5.0) having 0.5 M NaCl and later the enzyme has been obtained with the very same buffer. And all of fractions have been analyzed as stated above. And for later use, the active fractions have been combined, concentrated and allowed to stand at 4 °C. Protein concentration: The protein concentration was determined spectrophotometrically with an absorbance at 280 nm in addition to the Bradford's method23 using Bovine Serum Albumin (BSA) as a standard. Determination of laccase activity: The laccase activity has been determined using ABTS substrate24,25. The reaction mixture was prepared by adding 0.5 mL of the enzyme solution on top of the ABTS (3 mM) substrate dissolved in 0.5 mL of 0.1 M acetate buffer (pH = 4.5) and then it was incubated at 55 °C. The oxidation of ABTS was determined by monitoring the increase in absorbance at 420 nm. And the one unit of a laccase activity was defined as the required amount of enzyme to oxidize 1 μmol of ABTS/min (ε420 = 36,000 M-1 cm-1)26. In order to find out the Km and Vmax values, the laccase activity was determined by measuring at 420 nm, 470 and 468 nm respectively by making use of 3 mM ABTS, guaiacol and 2,6-dimetoksifenol substrates. The blind was prepared by adding the enzyme-containing buffer, instead of enzyme solution, which was used as the sample27.
SDS polyacrylamide gel electrophoresis: Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) was performed through the 10-30% discontinued electrophoresis method which was defined by Laemmli28. 20 µg of protein was applied to each of the samples. Electrophoresis in a gelcasting apparatus (Bio-RAD) execution buffer (0.25 M Tris, 1.92 M Glycine, 1% SDS (pH 8.3) was applied at 4 °C. The gel was dyed with 0.1% Coomassie Brilliant Blue R-250 in 50% methanol, 10% acetic acid and 40% distilled water for 1.5 h. Then the gel was washed with 50% methanol, 10% acetic acid and 40% distilled water until its surface becomes limpid. The electrophoretic pattern was photographed. Molecular weight determination by gel filtration: Molecular weight of the laccase enzyme was performed by using column (3x70 cm) of Sephadex G100. The column was equilibrated until reaching zero absorbance, by using 0.05 M Na2HPO4, 1 mM dithioerythretol, pH 7.0 buffer. Standard protein solution (bovine serum albumin, 66 kDa, egg ovalbumin, 45 kDa, pepsin, 34 kDa, trypsinogen, 24 kDa, ß-lactoglobulin and lysozyme, 14 kDa) was applied to the column and was eluted from the column with the same buffer. Then, the pure laccase enzyme was applied to the column and was eluted under the same conditions. The flow-rate through the column was 20 mL/h. The elution volume was compared with standard proteins29. Studies on certain in vitro chemicals and metal ions: The impact of the metal ions of Fe2+, Cu2+, Zn2+, Hg2+, Ba2+, Ca2+, Co2+, Mn2+, Ni2+, EDTA and ascorbic acid in different inhibitory concentrations on laccase enzyme was investigated. All metal ions were tested in three runs for each concentration. Laccase activity was measured in different concentrations of the inhibitor. I50 values were calculated for those that present inhibition30. Remediation of some textile dyes: In this study, it was investigated that whether the purified laccase enzyme can be used in the removal of some dyes used in industry or not. For this purpose, Reactive black 5, Fuchsine, Allura Red and Acid red 37 diammonium salt were used as 4 different dyes. The dye solutions were prepared at 50 mg/L concentration and by taking 50 mL from these solutions 1 mL (5 µg protein), the purified laccase enzyme was added. The reaction medium was set to pH 4.0 and 55 °C, which is the optimum pH and temperature for the enzyme and samples were taken at 15th, 30th, 60th, 90th, 120th, 150th and 180th minutes from the medium and the absorbance was measured spectrophotometrically for Reactive Black 5, Fuchsine, Allura Red, Acid Red 37 Diammonium Salt, at 597 nm, 547 nm, 513 nm and 504 nm respectively. The results obtained are shown in the chart as Absorbance vs. mg dye plots. Statistical analysis: All of the tests were conducted in
Research Journal of Biotechnology Vol. 8 (9) September (2013) Res. J. Biotech
(59)
three runs in order to determine the laccase activities of the samples. Data were expressed in terms of mean ± standard errors. Statistical analyses were performed using SPSS version 10.0 software (SPSS Inc., Chicago, IL., USA) and the significant differences were determined with a 95% confidence interval (p<0.05) using Tukey's test. Results and Discussion In this present study, the bacterial strain was subjected to a range of physiological and biochemical tests. It was revealed in the results that test strain was gram-positive, amylase negative, catalase and oxidase positive and also is of type endospore forming and mobile rod. Optimum temperature was 59 ± 1 °C and pH was 6.5-8.5 for this strain. Test strain was capable of growing in a 2%-8% salt concentration. These findings comply with the criteria of thermophilic bacteria which were living at temperatures higher than 50 °C18. 16S rRNA Gene Sequence Analysis: 1425 nucleotides of 16SrRNA from the test isolate (Z1) were aligned to compare with the sequences of the related bacteria. Stackebrandt and Goebel31 have concluded that the strains belonging to the same genus, which were exhibiting less than 97% 16S rRNA gene sequence similarity, should be considered as the members of different species. The isolate (Z1) has exhibited 99% resemblance to Brevibacillus, according to 16S rRNA gene sequence analysis18. The strain Z1's GenBank accession number is KC292196. 16S rRNA sequence analysis method was found inadequate to reveal the distinction between the species which are in a very close relationship with each other, however, it was found adequate for identification at the genus level again in
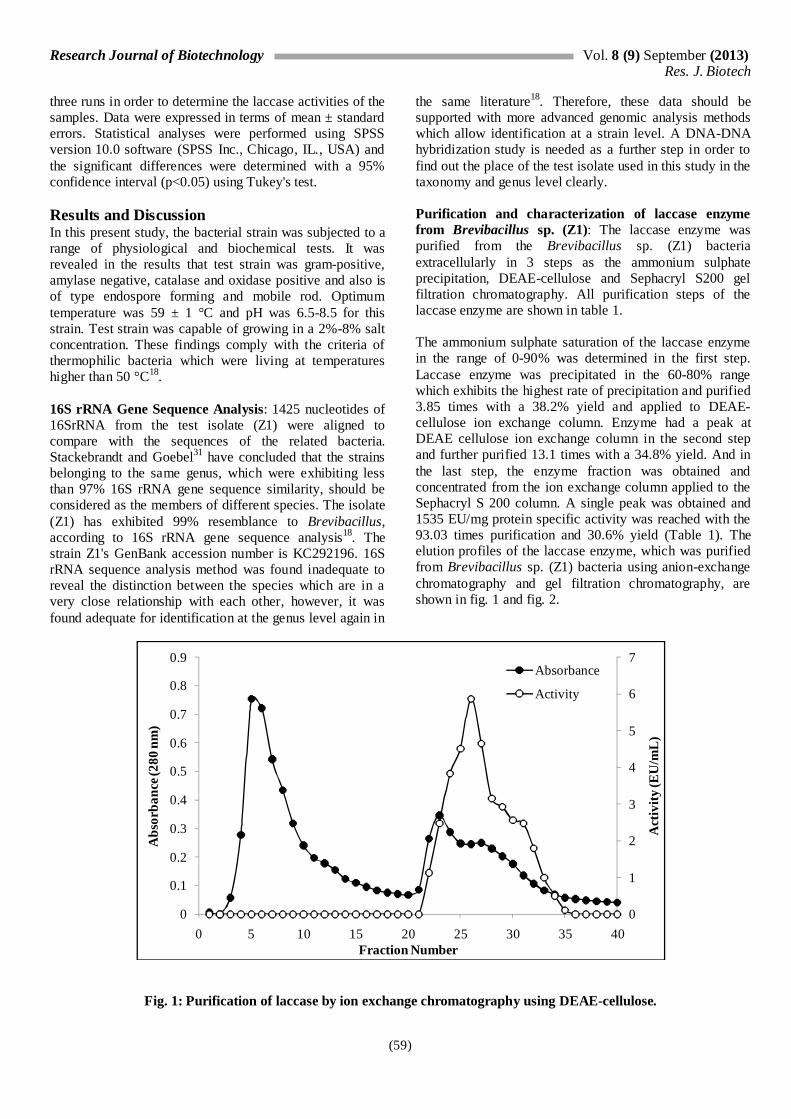
the same literature18. Therefore, these data should be supported with more advanced genomic analysis methods which allow identification at a strain level. A DNA-DNA hybridization study is needed as a further step in order to find out the place of the test isolate used in this study in the taxonomy and genus level clearly. Purification and characterization of laccase enzyme from Brevibacillus sp. (Z1): The laccase enzyme was purified from the Brevibacillus sp. (Z1) bacteria extracellularly in 3 steps as the ammonium sulphate precipitation, DEAE-cellulose and Sephacryl S200 gel filtration chromatography. All purification steps of the laccase enzyme are shown in table 1. The ammonium sulphate saturation of the laccase enzyme in the range of 0-90% was determined in the first step. Laccase enzyme was precipitated in the 60-80% range which exhibits the highest rate of precipitation and purified 3.85 times with a 38.2% yield and applied to DEAE-cellulose ion exchange column. Enzyme had a peak at DEAE cellulose ion exchange column in the second step and further purified 13.1 times with a 34.8% yield. And in the last step, the enzyme fraction was obtained and concentrated from the ion exchange column applied to the Sephacryl S 200 column. A single peak was obtained and 1535 EU/mg protein specific activity was reached with the 93.03 times purification and 30.6% yield (Table 1). The elution profiles of the laccase enzyme, which was purified from Brevibacillus sp. (Z1) bacteria using anion-exchange chromatography and gel filtration chromatography, are shown in fig. 1 and fig. 2.
Fig. 1: Purification of laccase by ion exchange chromatography using DEAE-cellulose.
0
1
2
3
4
5
6
7
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
0 5 10 15 20 25 30 35 40
Act
ivity
(EU
/mL
)
Abs
orba
nce (
280 n
m)
Fraction Number
Absorbance
Activity

Research Journal of Biotechnology Vol. 8 (9) September (2013) Res. J. Biotech
(60)
Fig. 2: Elution profile of protein and laccase activity on Sephacryl S200.
Table 1 The purification process of laccase from Brevibacillus sp. (Z1)
Enzyme Fraction Volume Activity Total Activity Protein Specific Activity
Purification Fold
Crude extract 50 55.86 2793 100 3.39 16.5 -
(NH4)2SO4 (%60-80) 25 42,63 1065.8 38.2 0.67 63.6 3.85
DEAE-cellulose 25 38,89 972,25 34.8 0.18 216.1 13.1
Sephacryl S 200 25 33,77 844.3 30.2 0.022 1535.0 93.03
The sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE) analysis revealed that the laccase enzyme consists of 3 subunits of sizes 122.000, 93.756 and 73.790 kDa (Fig. 3). The Sephadex G-100 gel filtration chromatography was used to find the molecular weight of the active form of the enzyme, log MA-Kav was plotted and the same results were obtained. The molecular weight of the laccase enzyme derived from Streptomyces griseus (EpoA) bacteria was found as 114 kDa and it was homotrimer32,33. In addition, the laccase enzyme derived from Bacillus sp (mnxg) bacteria had been found to have a multimeric structure consisting of 3 subunits of different molecular weights34 as 48.9, 97.8 and 179.3 kDa. In order to find the optimum pH of the laccase enzyme purified and derived from Brevibacillus sp. (Z1), seven different pH medium were studied ranging from pH 3 to 9. The buffers used were: acetate for pH 3 to 5, phosphate for
pH 6 to 7 and Tris/HCl for pH 8 to 9. Activity measurements were made using ABTS substrate, the activity was calculated by using the obtained results. Activity-pH was plotted and the results are shown in fig. 4. The optimum pH value was found as 4.0 for the ABTS substrate in the laccase enzyme purified and derived from Brevibacillus sp. (Z1).
The laccase enzyme was incubated for 5 days at +4 °C by using the buffers of acetate for pH 3 to 5, phosphate for pH 6 to 7 and Tris/HCl for pH 8 to 9, activity measurements were performed periodically and the findings are shown in fig. 5. As can be seen in the plots, it was found that the laccase enzyme maintains 55.6% of its activity after 5 days and has a stable pH of 5.0 for ABTS. The laccase enzymes derived from C. hirsutus, T. atroviride, Chalara (syn. Thielaviopsis) paradoxa CH32 and Paraconiothyrium variable have presented optimum pH values in the range of pH 4-635.
0
0.1
0.2
0.3
0.4
0.5
0.6
0
5
10
15
20
25
0 5 10 15 20 25 30 35
Act
ivity
(EU
/mL
)
Fraction Number
Abs
orba
nce (
280 n
m)
ActivityAbsorbance

Research Journal of Biotechnology Vol. 8 (9) September (2013) Res. J. Biotech
(61)
Fig. 3: SDS-PAGE Electrophoretic pattern of laccase [(I) standart protein; (II) homogenate; (III) purified
laccase enzyme from Brevibacillus sp. (Z1)].
Fig. 4: Effect of pH on the activity of the purified
laccase. Enzymes and substrate were dissolved either in 10 mM buffers of various pH. Other conditions were as
given for the standard assay method
Fig. 5: Effect of the pH on the stability of the purified laccase. Enzymes were incubated at 55 ◦C for 1 h at
different pH and the residual activity was measured the standard assay method
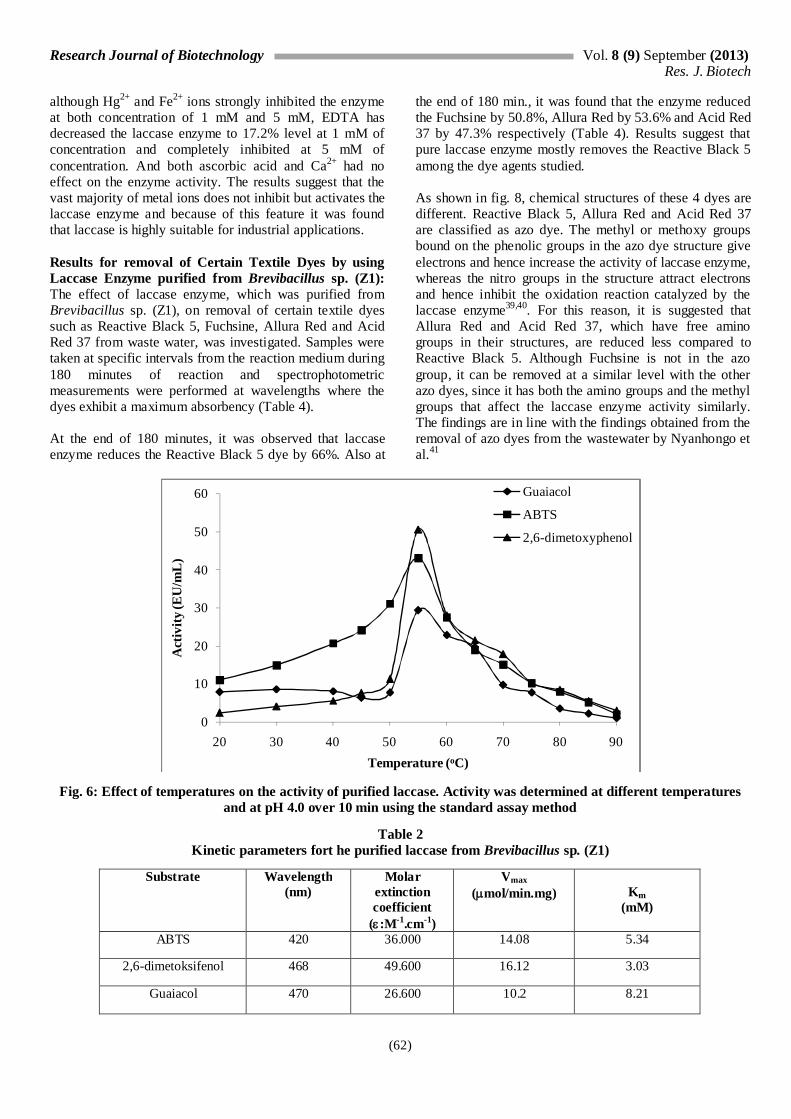
Optimum Temperature: In order to find the optimum pH of the purified laccase enzyme, the activity was measured for each substrate in the temperature range of 20 to 90 °C, by keeping it for 5 minutes at each temperature. As can be seen in fig. 6, it was found that the laccase enzyme was highly active between 30 to 80 °C for ABTS and DMP substrates and has the maximum activity at 55 °C. Similarly, it was also found in a study conducted by Frootanfar et al36 that the laccase enzyme derived from Paraconiothyrium variable had the optimum temperature of 50 °C. Stable Temperature: Enzyme was incubated for 1 week at 30-90 °C in order to determine its stable temperature. Activity measurements have been performed by standing the enzyme at the specified temperatures for 15, 30, 45 and 60 minutes and adding the substrate later. This study has been performed using ABTS substrate. It was observed that the enzyme maintains most of its activity after 1 hour at all temperatures. It was found that it exhibits a 71.7% activity even at 80 °C after 1 hour (Fig. 7). In general, bacterial laccases are stable at 70 °C for half an hour and at 80 °C for 10 minutes15,35. However, it appears that the laccase enzyme we have purified from Brevibacillus sp. (Z1) is more resistant to heat than the bacterial laccases purified previously. The stability of Brevibacillus sp. (Z1) laccase enzyme at high temperatures increases the use of this enzyme in different industrial areas. Results of the Vmax and Km Values of the Laccase Enzyme Purified from Brevibacillus sp. (Z1): The studies on Vmax and Km values of the laccase enzyme were performed with 5 mM of ABTS, Guaiacol (phenolic substrates) and 2.6 dimetoxiphenol (non-phenolic substrate). The kinetic parameters obtained by using different substrates are summarized in table 2. It was found that, Vmax: 16.12 μmol/min.mg and Km: 3.03 mM of the pure Laccase enzyme in the presence of 2,6-DMP and the highest affinity and the most effective oxidation of the enzyme were against 2.6-DMP in comparison to other substrates. Vmax values for the ABTS and guaiacal substrates were 14.08 μmol/min.mg, 10.2 μmol/min.mg and the Km values were 5.34 mM and 8.21 mM respectively. When the DMP substrate was used, the Km values were found as 10.23 and 2.66 mM for the Laccase enzyme purified from Fusarium solani MAS237 and T. atroviride38. Impact of Ca2+, Mg2+, Co2+, Fe3+, Zn2+, Cu2+, Hg2+, EDTA and Ascorbic Acid on the Activity of the Laccase Enzyme Purified from Brevibacillus sp. (Z1): Impacts of Ca2+, Mg2+, Co2+, Fe3+, Zn2+, Cu2+, Hg2+, EDTA and ascorbic acid on the activity of the Laccase enzyme purified from Brevibacillus sp. (Z1) have been studied. The obtained values on inhibitory effects and concentration vs. activity values are shown in table 3. As shown in table 3, Ca2+, Mn2+, Zn2+, Cu2+, Ba2+, Ni2+ and Co2+ metal ions have increased the activity of the laccase enzyme. However,
0
15
30
45
60
75
90
3 5 7 9
Act
ivity
(EU
/mL
)
pH
0
10
20
30
40
50
60
0 1 2 3 4 5 6
Act
ivity
(EU
/mL
)
Time (Min)
3 4 5 6 7 8 9
I II II III
110.000 kDa 93.000 kDa
50.000 kDa
30.000 kDa
20.000 kDa
70.000 kDa
150.000 kDa 100.000 kDa
Research Journal of Biotechnology Vol. 8 (9) September (2013) Res. J. Biotech
(62)
although Hg2+ and Fe2+ ions strongly inhibited the enzyme at both concentration of 1 mM and 5 mM, EDTA has decreased the laccase enzyme to 17.2% level at 1 mM of concentration and completely inhibited at 5 mM of concentration. And both ascorbic acid and Ca2+ had no effect on the enzyme activity. The results suggest that the vast majority of metal ions does not inhibit but activates the laccase enzyme and because of this feature it was found that laccase is highly suitable for industrial applications. Results for removal of Certain Textile Dyes by using Laccase Enzyme purified from Brevibacillus sp. (Z1): The effect of laccase enzyme, which was purified from Brevibacillus sp. (Z1), on removal of certain textile dyes such as Reactive Black 5, Fuchsine, Allura Red and Acid Red 37 from waste water, was investigated. Samples were taken at specific intervals from the reaction medium during 180 minutes of reaction and spectrophotometric measurements were performed at wavelengths where the dyes exhibit a maximum absorbency (Table 4). At the end of 180 minutes, it was observed that laccase enzyme reduces the Reactive Black 5 dye by 66%. Also at
the end of 180 min., it was found that the enzyme reduced the Fuchsine by 50.8%, Allura Red by 53.6% and Acid Red 37 by 47.3% respectively (Table 4). Results suggest that pure laccase enzyme mostly removes the Reactive Black 5 among the dye agents studied. As shown in fig. 8, chemical structures of these 4 dyes are different. Reactive Black 5, Allura Red and Acid Red 37 are classified as azo dye. The methyl or methoxy groups bound on the phenolic groups in the azo dye structure give electrons and hence increase the activity of laccase enzyme, whereas the nitro groups in the structure attract electrons and hence inhibit the oxidation reaction catalyzed by the laccase enzyme39,40. For this reason, it is suggested that Allura Red and Acid Red 37, which have free amino groups in their structures, are reduced less compared to Reactive Black 5. Although Fuchsine is not in the azo group, it can be removed at a similar level with the other azo dyes, since it has both the amino groups and the methyl groups that affect the laccase enzyme activity similarly. The findings are in line with the findings obtained from the removal of azo dyes from the wastewater by Nyanhongo et al.41
Fig. 6: Effect of temperatures on the activity of purified laccase. Activity was determined at different temperatures and at pH 4.0 over 10 min using the standard assay method
Table 2 Kinetic parameters fort he purified laccase from Brevibacillus sp. (Z1)
Substrate Wavelength
(nm) Molar
extinction coefficient
(:M-1.cm-1)
Vmax (mol/min.mg)
Km
(mM)
ABTS 420 36.000 14.08 5.34
2,6-dimetoksifenol 468 49.600 16.12 3.03
Guaiacol 470 26.600 10.2 8.21
0
10
20
30
40
50
60
20 30 40 50 60 70 80 90
Act
ivity
(EU
/mL
)
Temperature (oC)
Guaiacol
ABTS
2,6-dimetoxyphenol

Research Journal of Biotechnology Vol. 8 (9) September (2013) Res. J. Biotech
(63)
Fig. 7: Effect of the temperature on the stability of the purified laccase. Enzymes were incubated at pH:4 for 1 h at different different temperature and the residual activity was measured the standard assay method
Reactive Black 5
Fuchsin
Allura Red
Acid Red 37
Fig. 8: Structure of used commercial dyes, Color Index (CI) names given in table 4
0
10
20
30
40
50
60
0 10 20 30 40 50 60
Act
ivity
(EU
/mL
)
Time (Min)
30 40 50 60 70 80 90

Research Journal of Biotechnology Vol. 8 (9) September (2013) Res. J. Biotech
(64)
Table 3 The effect of some metal ions and some chemicals on laccase activity from Brevibacillus sp. (Z1)
Chemical Compounds
Concentration (mM)
Laccase Activity (%)
Concentration (mM)
Laccase Activity
(%)
None - 100 - 100
Ascorbic acid 1 100 5 100
EDTA 1 17.2 5 0
Ca2+
1 100 5 100
Hg2+
1 33.8 5 19.2
Mn2+
1 200 5 300
Zn2+
1 166.7 5 181.6
Cu2+
1 120 5 182.2
Fe2+
1 82.2 5 46.7
Ba2+
1 153.3 5 181.1
Ni2+ 1 122.7 5 327.3
Co2+ 1 126.7 5 189.3
Table 4 Dye decolorization rates of the laccase from Brevibacillus sp. (Z1)
Dyes (C.I. names) Dye
class Absorbance
max. Concentration (mg/L)
Time (Min) 0 15 30 60 90 120 150 180
Reactive Black 5 Azo 597 50 48.6 48.5 48.4 47.1 44.25 37.65 33.0 Fuchsin Magenta 547 50 41.2 40.35 38.4 36.6 35.45 30.8 25.4
Allura Red Azo 513 50 48.8 48.55 44.1 40.9 38.75 33.05 26.8 Acid Red 37 Azo 504 50 39.95 34.05 31.05 29.15 27.55 25.1 23.65
Conclusion The results suggest that the laccase enzyme purified from Brevibacillus sp. (Z1) is of very higher catalytic activity, durable against the metal ions, highly stable at different temperatures and pH levels and are economic. Thanks to these features that it can be used widely and effectively in the removal of the dyes that cause environmental pollution from waste water. Acknowledgement This research was performed under the project numbered 2011/380 and supported by the Research Development Center of Ataturk University. The authors acknowledge the support of Ataturk University, Turkey for this work. References 1. Lindstrom R.F., Ramaley R.F. and Bizzoco R.L.W., Invisible invasion: Potential contamination of yellowstone hot springs by human activity, West N. Am. Naturalist, 62(1), 44-58 (2002)
2. Adiguzel A. Molecular Characterization of thermophilic Bacteria Isolated from Water Samples Taken from Various thermal Plants, Ph.D. thesis, Atatürk University, Graduate School at Natural and Applied Sciences, Erzurum, Turkey, 130 (2006) 3. Aksu Z., Application of biosorption for the removal of organic pollutants: a review, Process Biochem., 40, 997-1026 (2005)
4. Aksu Z. and Donmez G., Combined effects of molasses sucrose and reactive dye on the growth and dye bioaccumulation properties of Candida tropicalis, Process Biochem., 40, 437-444 (2005) 5. Seesuriyachana P., Takenakab S., Kuntiyaa A., Klayraungc S., Murakamib S. and Aokib K., Metabolism of azo dyes by Lactobacillus casei TISTR 1500 and effects of various factors on decolorization, Water Res., 41, 985-992 (2007) 6. Lucas M.S., Amaral C., Sampaio A., Peres J.A. and Dias A.A., Biodegradation of the diazo dye Reactive Black 5 by a wild isolate of Candida oleophila, Enzyme Microb. Tech., 39, 51-55 (2006)

Research Journal of Biotechnology Vol. 8 (9) September (2013) Res. J. Biotech
(65)
7. Thurston C.F., The structure and function of fungal laccases, Microbiol., 140, 19-26 (1994) 8. Delanoy G., Li Q. and Yu J., Activity and stability of laccase in conjugation with chitosan, Int. J. Biol Macromol, 35, 89 (2005) 9. Sharma P., Goel R. and Capalash N., Bacterial laccases, World J. Microbiol. Biotechnol, 23, 823-832 (2007) 10. Ruijssenaars H.J. and Hartmans S., A cloned Bacillus halodurans multicopper oxidase exhibiting alkaline laccase activity, Appl. Microbiol. Biotechnol., 65, 177-182 (2004) 11. Suzuki T., Endo K., Ito M., Tsujibo H., Miyamoto K. and Inamori Y., A thermostable laccase from Streptomyces lavendulae REN-7: Purification, characterization, nucleotide sequence and expression, Biosci. Biotechnol. Biochem., 67, 2167-2175 (2003) 12. Fitz-Gibbon S.T., Ladner H., Kim U.J., Stetter K.O., Simon M.I. and Miller J.H., Genome sequence of the hyperthermophilic crenarchaeon Pyrobaculum aerophilum, Proc. Natl. Acad. Sci., USA, 99, 984-989 (2002) 13. Deckert G., Warren P.V., Gaasterland T., Young W.G., Lenox A.L., Graham D.E., Overbeek R., Snead M.A., Keller M., Aujay M., Huber R., Feldman R.A., Short J.M., Olson G.J. and Swanson R.V., The complete genome of the hyperthermophilic bacterium Aquifex aeolicus, Nature, 393, 353-358 (1998) 14. Miyazaki K., A hyperthermophilic laccase from Thermus thermophilus HB27, Extremophiles, 9, 415-425 (2005) 15. Baldrian P., Fungal laccases-occurrence and properties, FEMS Microbiol Rev., 30(2), 215-242 (2006) 16. Sukumar M., Sivasamy A. and Swaminathan G.. Biosorption of Azo Dye (AR 66) by Hypnea valentiae and Ulva fasciate, Res. J. Biotech., 1(1), 30-40 (2006) 17. Harley J.P. and Prescott L.M., Laboratory Exercises in Microbiology, Fifth Edition, New York, The McGraw-Hill Companies (2002) 18. Adiguzel A., Ozkan H., Baris O., Inan K. and Gulluce M., Identification and characterization of thermophilic bacteria isolated from hot springs in Turkey, J. Microbiol. Meth., 79, 321-328 (2009)
19. Beffa T., Blanc M., Lyon P.F., Vogt G., Marchiani M., Fischer J.L. and Aragno M., Isolation of Th ermus strains from hot composts (60 to 80 °C), Appl. Environ. Microbiol., 62, 1723-1727 (1996) 20. Benson D.A., Boguski D.S., Lipman D.J., Ostell J., Ouellette B.F.F., Rapp B.A. and Wheeler D.L., GenBank, Nucleic Acids Res., 27, 12-17 (1999) 21. Altschul S.F., Gish W., Miller W., Myers E.W. and Lipman D.J., Basic local alignment search tool, J. Mol. Biol., 215, 403-410 (1990) 22. Zeng X., Cai Y., Liao X., Zeng X., Luo S. and Dabing Z., Anthraquinone dye assisted the decolorization of azo dyes by a
novel Trametes trogii laccase, Process Biochem., 47(1), 160-163 (2012) 23. Bradford M.M., A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding, Annal Biochem., 7, 248-254 (1976) 24. Alberts J.F., Gelderblom W.C.A., Botha A. and Vanzyl W.H., Degradation of aflatoxin B1 by fungal laccase enzymes, Int J Food Microbiol, 135, 47-52 (2009) 25. Faramarzi M.A. and Forootanfar H., Biosynthesis and characterization of gold nanoparticles produced by laccase from Paraconiothyrium variabile, Colloid Surf. B, 87, 23-27 (2011) 26. Lu L., Zhao M., Zhang B.B., Yu S.Y., Bian X.J., Wang W. and Wang Y., Purification and characterization of laccase from Pycnoporus sanguineus and decolorization of an anthraquinone dye by the enzyme, Appl. Microbiol. Biotechnol., 74, 1232-1239 (2006) 27. Arias M.E., Arenas M., Rodríguez J., Soliveri J., Ball A.S. and Hernández M., Kraft pulp biobleaching and mediated oxidation of a nonphenolic substrate by laccase from Streptomyces cyaneus CECT 3335, Appl Environ Microbiol., 69(4), 1953 (2003) 28. Laemmli D.K., Cleavage of structural proteins during in assembly of the heat of bacteriophage T4, Nature, 227, 680-685 (1970) 29. Whitaker J.R., Determination of molecular weight of proteins by gel filtration on sephadex, Anal. Chem., 35, 1950-1953 (1963) 30. Nadaroglu H., Celebi N., Kalkan E. and Tozsin G., Water purification of textile dye Acid red 37 by adsorption on laccase-modified silica fume, Jökull J., 63, 87-118 (2013) 31. Stackebrandt E. and Goebel B.M., Taxonomic note: a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology, Int. J. Syst. Bacteriol., 44, 846-849 (1994) 32. Endo K., Hayashi Y., Hibi T., Hosono K., Beppu T. and Ueda K., Enzymological characterization of EpoA, a laccase-like phenol oxidase produced by Streptomyces griseus, J. Biochem., 133, 671-677 (2003) 33. Endo K., Hosono K., Beppu T. and Ueda K., A novel extracytoplasmic phenol oxidase of Streptomyces: its possible involvement in the onset of morphogenesis, Microbiol., 148, 1767–1776 (2002) 34. Van Waasbergen L.G., Hildebrand M. and Tebo B.M., Identification and characterization of a gene cluster involved in manganese oxidation by spores of the marine Bacillus sp. strain SG-1, J. Bacteriol., 178(12), 3517-3530 (1996) 35. Sadhasivam S., Savitha S., Swaminathan K. and Lin F.H., Production, purification and characterization of mid-redox potential laccase from a newly isolated Trichoderma harzianum WL1, Process Biochem., 43, 736-742 (2008)

Research Journal of Biotechnology Vol. 8 (9) September (2013) Res. J. Biotech
(66)
36. Forootanfar H., Faramarzi M.A., Shahverdi A.R. and Yazdi M.T., Purification and biochemical characterization of extracellular laccase from the ascomycete Paraconiothyrium variabile, Bioresour Technol., 102(2), 1808-14 (2011) 37. Wu Y.R., Luo Z.H., Chow R.K.K. and Vrijmoed L.L.P., Purification and characterization of an extracellular laccase from the anthracene degrading fungus Fusarium solani MAS2, Biores. Technol., 101(24), 9772-9777 (2010) 38. Chakroun H., Mechichi T., Martinez M.J., Dhouib A. and Sayadi S., Purification and characterization of a novel laccase from the ascomycete Trichoderma atroviride: application on bioremediation of phenolic compounds, Process Biochem., 507-513 (2010)
39. Xu F., Oxidation of phenols, anilines and benzenethiols by fungal laccases: correlation between activity and redox potentials as well as halide inhibition, Biochem., 35, 7608-7614 (1996) 40. Chivukula M. and Renganathan V., Phenolic azo dye oxidation by laccase from Pycularia oryzae, Appl. Environ. Microbiol., 61, 4374-4377 (1995) 41. Nyanhongo G.S., Gomes J., Gubitz G.M., Zvauya R., Read J. and Steiner W., Decolorization of textile dyes by laccases froma newly isolated strain of Trametes modesta, Water Res., 36, 1449-1456 (2002). (Received 30th May 2013, accepted 26th July 2013)





























