Embed Size (px)
Citation preview

Downloaded from www.microbiologyresearch.org by
IP: 54.205.18.105
On: Thu, 23 Jun 2016 06:26:26
Journal of General Virology (1990), 71, 3061-3066. Printed in Great Britain 3061
Purification and initial characterization of human placental trophoblast interferon induced by polyriboinosinic, polyribocytidylic acid
George Aboagye-Mathiesen, 1 Ferenc D. T6th, 1,2 Claus Juhl, 1 Nieis Norskov-Lauritsen, ~ Peter M. Petersen 1 and Peter Ebbesen 1.
I The Danish Cancer Society, Department o f Virus and Cancer, Gustav Wieds Vej 10, DK-8000 Aarhus C, Denmark and 2 Institute o f Microbiology, Medical University, H-4012 Debrecen, Hungary
Human placental trophoblast interferon (tro-IFN), induced in trophoblast cultures by a superinduction procedure, was purified to a homogeneous product with retention of biological activity. The problems associated with isolation from serum-containing medi- um were overcome by a combination of Blue Sepharose affinity chromatography and reversed-phase HPLC (RP-HPLC) on Separon SGX C-18. This two-step
purification procedure yielded tro-IFN with a specific activity of 3.4 × 107 international units/mg of protein. The overall recovery of interferon activity was 66-7 %. The purified tro-IFN was shown to be a glycoprotein with an Mr of 24K on native and SDS-PAGE. Its antiviral activity was stable at pH 2.0 at 37 °C but was sensitive to heat at 56 °C for 1 h and was neutralized by antibodies to human IFN-fl.
Interferons (IFNs) are a class of proteins secreted by animal cells in response to various inducers, and exert a non-specific cellular resistance towards a wide variety of DNA and RNA viruses (Fisher & Grant, 1985). In the human species, three classes of IFNs have been distinguished biochemically and immunologically and are designated as ~t, t , y. However, it is also recognized that there are a variety of different IFNs, and probably there are others that have not yet been discovered (Kirchner, 1986).
IFNs have been produced and characterized from various animal cells and it is now clear that the quantity and type produced vary considerably from one cell and inducer to another (Ng & Vil~ek, 1972). Cesario et al. (1977) observed that IFNs induced by the same inducer in different cells were different in terms of their charge, heat and chemical stabilities. They also found that the cell induced and the circumstances of the induction exerted an important influence on the properties of the IFNs produced. Human placental trophoplast IFN (tro- IFN) has recently been produced in our laboratory from primary cultures of human placental trophoblast, in- duced with" polyriboinosinic-polyribocytidylic acid [poly(rI)-poly(rC)], as a secretion product in the medium (T6th et al., 1990a). In preparation for a study of the regulation, expression and the structure-function rela- tions of human placental tro-IFNs and the role of each in vivo in the protection of the foetus against virus infections, we report here the purification from serum- containing medium of tro-IFN, superinduced by
poly(rI)-poly(rC) together with actinomycin D and cycloheximide, to a homogeneous protein by Blue Sepharose affinity chromatography and reverse-phase (RP-HPLC) on Separon SGX C-18. In addition, the initial characterization of the protein is also reported.
Term placental trophoblasts were separated on a Percoll gradient (Kliman et al., 1987). Cultures used for IFN induction were composed of 97 % trophoblasts, with fibroblasts and macrophages in roughly equal amounts. Tro-IFN was induced by a modification of the super- induction method reported by Tan & Berthold (1977) using poly(rI), poly(rC), cycloheximide and actinomycin D (T6th et al., 1990b).
Tro-IFN was assayed using inhibition of vesicular stomatitis virus plaque formation in WISH cells (Tbth et al., 1990b). IFN titres, expressed as international IFN units (IU), were standardized by comparison with the NIH standard for human IFN-fl (G-023-902-527). Antibody neutralization of IFN antiviral activity was performed by incubating IFN samples in different dilutions (from 100 to 1 IU) for 1 h at 37 °C with a 10-fold excess of horse immune serum to human IFN-~, IFN-fl and IFN-y, and then assaying the samples for the presence of IFN antiviral activity.
The first purification step of the tro-IFN was by Blue Sepharose affinity chromatography, using a modifica- tion of that used by Knight & Fahey (1980) for human fibroblast IFN produced in serum-free medium. IFN- containing culture supernatant (220 ml; total activity of 1.80 × 106 IU, 146.7 mg protein) was made to
0000-9651 © 1990 SGM
Downloaded from www.microbiologyresearch.org by
IP: 54.205.18.105
On: Thu, 23 Jun 2016 06:26:26
3062 Short communication
l-5 M-NaC1 by directly adding solid NaC1. The sample (culture supernatant with 1.5 M-NaC1) was filtered through a 0.22 gm Millipore filter and pumped onto a column (16 mm x 25 mm, packed with Blue Sepharose CL-6B; Pharmacia LKB) equilibrated with column buffer (0.02 M-sodium phosphate buffer pH 7-2, contain- ing 1-5 M-NaC1) at a flow rate of 1 ml/min. The column was washed with 20 ml of the column buffer and the bound proteins were eluted with increasing concentra- tions of ethylene glycol in the same buffer at the same flow rate. The elution steps were 0 to 20~ for 40 min, 20 to 50~ for 15 min, and 50~ for 37 min. The elution from the column was monitored continuously at an absor- bance of 280 nm (1.28 AUS) and fractions of 2 ml were collected into 1 ml of column buffer. All fractions were then assayed for IFN antiviral activity.
The second purification step consisted of RP-HPLC on Separon SGX C-18 (octadecasilyl silica material with a particle size of 5 gin, Tessek A/S). The IFN-containing fractions from the affinity chromatography were diluted twice with 1 M-pyridine/acetic acid pH 5.0 and immedi- ately applied to an RP-HPLC column (3 x 150 mm). The column was washed with 10 ml of 25~ propan-l-ol in 1 M-pyridine/acetic acid buffer pH 5.0, at a flow rate of 0.20 ml/min (7.5 MPa). The column was then eluted with a linear gradient of 25 to 60~ propan-l-ol in the same buffer for 100 min at a flow rate of 0-20 ml/min. The eluate from the column was monitored continuously at an absorbance of 280 nm (0.32 AUS) and fractions of 0.4 ml/tube were collected. All fractions were again assayed for IFN antiviral activity. IFN-containing fractions were pooled and propan-l-ol was removed by two extractions with an equal volume of hexane. Finally, traces of hexane were removed under a stream of nitrogen and the IFN preparation was stored at - 20 °C.
The thermal stability of IFN samples was determined at 37 °C and 56°C. Tro-IFN samples (0-1 ml) were heated to 37 °C or 56 °C for various times. After heating, samples were allowed to cool to room temperature or were stored on ice for 1 h until they were assayed for antiviral activity. To determine the pH stability, tro- IFN-containing fluids were dialysed against glycine- HC1 buffer pH 2.0 for 24 h at 4 °C, followed by dialysis against Tris-HCl buffer pH 7-2 for a further 24 h at 4 °C to restore the pH to neutrality. The samples were then tested for IFN antiviral activity.
Protein concentration of the original culture superna- tant was measured by the dye binding assay of Bradford (1976) using bovine serum albumin (BSA) as a standard. The concentration of the purified IFN was measured by absorbance at 280 nm or by derivatization with fluorescamine and then injecting the samples into a fluorescence HPLC monitor and using known concen- trations of BSA as a standard.
2.0
1.5
"~ 1.o
I ' l ' I I I
j -
!
0.5
"_22__ ¸ "6 t5°_ 1o
• q30
_, lO , o
30
25 X
20 "~
15 ~
z
0 l l0 120 130 140 150 160 170 180 Fraction number
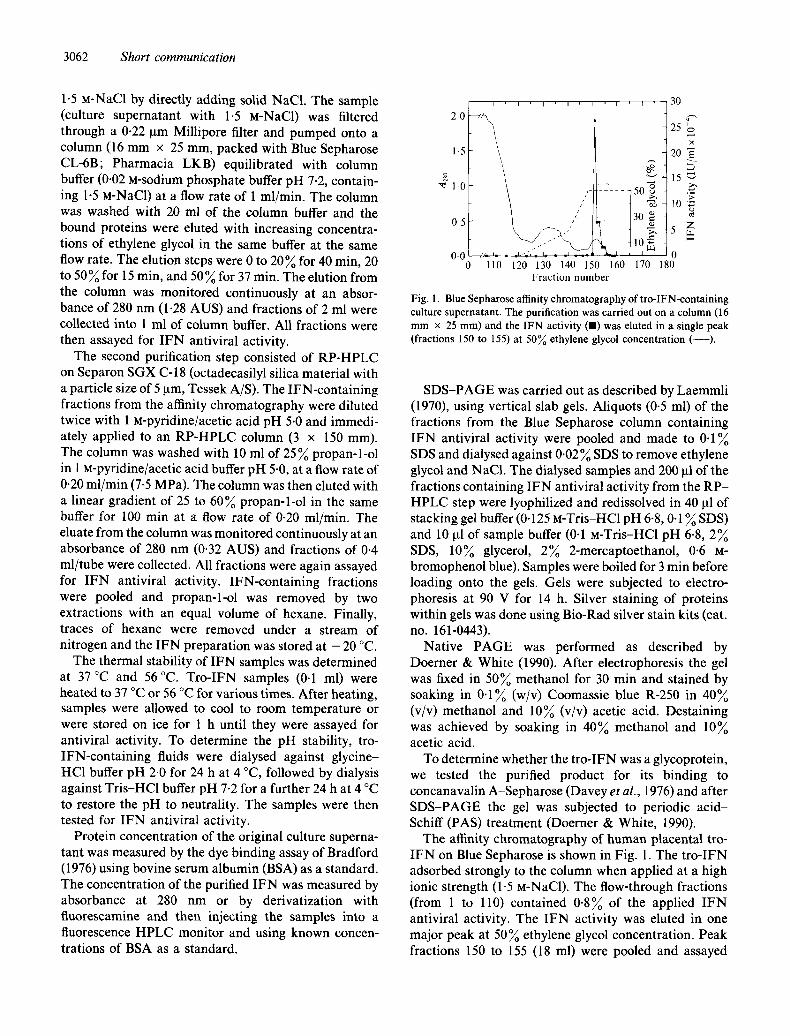
Fig. 1. Blue Sepharose affinity chromatography of t ro-IFN-containing culture supernatant. The purification was carried out on a column (16 m m x 25 mm) and the IFN activity (11) was eluted in a single peak (fractions 150 to 155) at 5 0 ~ ethylene glycol concentration ( - - - ) .
SDS-PAGE was carried out as described by Laemmli (1970), using vertical slab gels. Aliquots (0-5 ml) of the fractions from the Blue Sepharose column containing IFN antiviral activity were pooled and made to 0.1~ SDS and dialysed against 0.02 ~ SDS to remove ethylene glycol and NaC1. The dialysed samples and 200 gl of the fractions containing IFN antiviral activity from the RP- HPLC step were lyophilized and redissolved in 40 p.1 of stacking gel buffer (0-125 M-Tris-HC1 pH 6-8, 0.1 ~ SDS) and 10 gl of sample buffer (0.1 M-Tris-HCI pH 6-8, 2 ~ SDS, 10~ glycerol, 2~ 2-mercaptoethanol, 0.6 M- bromophenol blue). Samples were boiled for 3 min before loading onto the gels. Gels were subjected to electro- phoresis at 90 V for 14 h. Silver staining of proteins within gels was done using Bio-Rad silver stain kits (cat. no. 161-0443).
Native PAGE was performed as described by Doerner & White (1990). After electrophoresis the gel was fixed in 50~ methanol for 30 min and stained by soaking in 0"1~o (w/v) Coomassie blue R-250 in 40~ (v/v) methanol and 10~ (v/v) acetic acid. Destaining was achieved by soaking in 40~ methanol and 10~ acetic acid.
To determine whether the tro-IFN was a glycoprotein, we tested the purified product for its binding to concanavalin A-Sepharose (Davey et al., 1976) and after SDS-PAGE the gel was subjected to periodic acid- Schiff (PAS) treatment (Doerner & White, 1990).
The affinity chromatography of human placental tro- IFN on Blue Sepharose is shown in Fig. 1. The tro-IFN adsorbed strongly to the column when applied at a high ionic strength (1-5 M-NaC1). The flow-through fractions (from 1 to 110) contained 0.8~ of the applied IFN antiviral activity. The IFN activity was eluted in one major peak at 50~ ethylene glycol concentration. Peak fractions 150 to 155 (18 ml) were pooled and assayed
Downloaded from www.microbiologyresearch.org by
IP: 54.205.18.105
On: Thu, 23 Jun 2016 06:26:26
Short communication 3063
1 2 3 4 5 6
67K--
43K--
30K-- 25K-- ~---tro-IFN
20.1K--
14.4K --
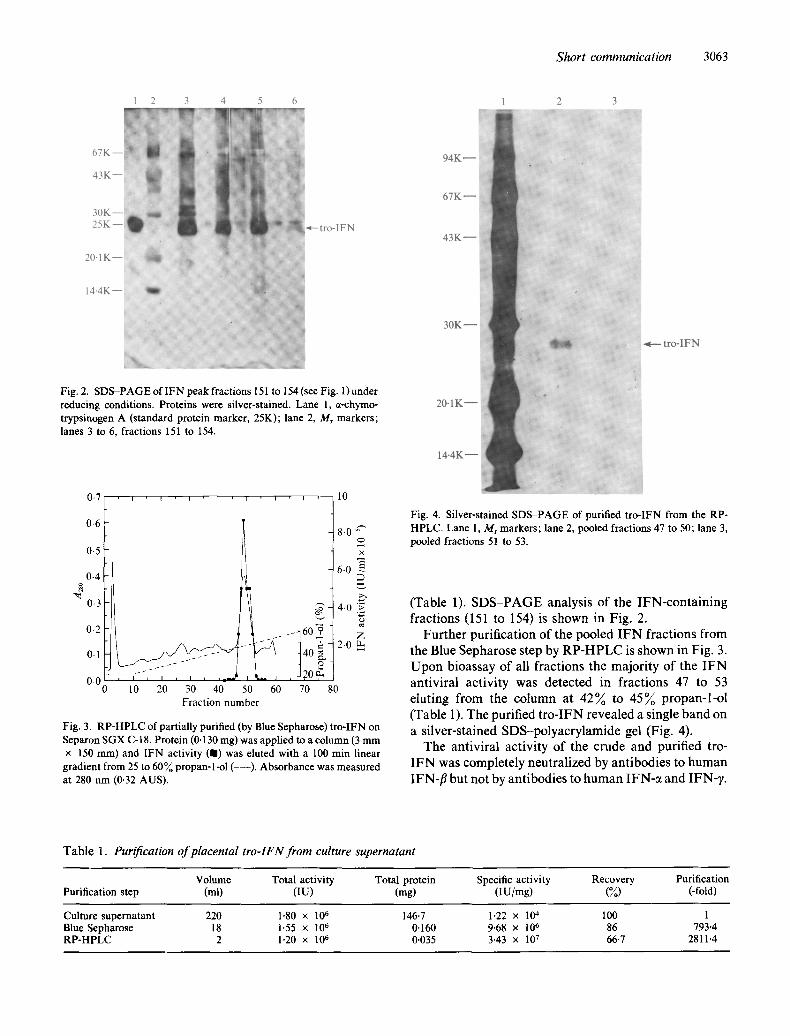
Fig. 2. SDS-PAGE of lFN peak fractions 151 to 154 (see Fig. 1)under reducing conditions. Proteins were silver-stained. Lane 1, e-chymo- trypsinogen A (standard protein marker, 25K); lane 2, Mr markers; lanes 3 to 6, fractions 151 to 154.
0.7 i , , , i , ,
06 I 0.5
0-4
"~ 0.3 , ~~,_,
0.2 _- 60"?-
0.1 40 ~.
0-0 10 20 30 40 50 60 70 80
Fraction number
10
8.0 ~"
X
6-0
4.0 IE
2.0 Z
Fig. 3. RP-HPLC of partially purified (by Blue Sepharose) tro-IFN on Separon SGX C-18. Protein (0.130 mg) was applied to a column (3 mm x 150 mm) and IFN activity (11) was eluted with a 100 min linear
gradient from 25 to 60% propan-l-ol (---). Absorbance was measured at 280 nm (0-32 AUS).
94K
67K
43K--
30K--
20.1K--
2 3
14.4K--
Fig. 4. Silver-stained SDS-PAGE of purified tro-IFN from the RP- HPLC. Lane 1, M r markers; lane 2, pooled fractions 47 to 50; lane 3, pooled fractions 51 to 53.
(Table 1). S D S - P A G E analysis of the IFN-con ta in ing fractions (151 to 154) is shown in Fig. 2.
Fur ther purification of the pooled I F N fractions f rom the Blue Sepharose step by R P - H P L C is shown in Fig. 3. U p o n bioassay of all fractions the majori ty of the I F N antiviral activity was detected in fractions 47 to 53 eluting f rom the column at 42% to 45% propan- l -o t (Table 1). The purified t ro - IFN revealed a single band on a silver-stained SDS-polyacry lamide gel (Fig. 4).
The antiviral activity o f the crude and purified tro- I F N was completely neutralized by antibodies to h u m a n IFN-f l but not by antibodies to h u m a n IFN-~ and IFN-~.
Table 1. Purification of placental tro-IFN from culture supernatant
Volume Total activity Total protein Specific activity Recovery Purification Purification step (ml) (IU) (mg) (IU/mg) (%) (-fold)
Culture supernatant 220 1.80 x 106 146.7 1.22 x 104 100 1 Blue Sepharose 18 1.55 × 106 0-160 9.68 x 106 86 793-4 RP-HPLC 2 1.20 x 106 0.035 3.43 x 107 66-7 2811.4

Downloaded from www.microbiologyresearch.org by
IP: 54.205.18.105
On: Thu, 23 Jun 2016 06:26:26
3064 Short communication
These results showed that pure cultures of human placental trophoblasts exposed to poly(rI)-poly(rC), cycloheximide and actinomycin D produced exclusively an IFN-fl type.
The tro-IFN retained its antiviral activity after 24 h treatment at pH 2-0 (initially 5000 IU/ml, 4860 after treatment). From the results the antiviral activity after 24 h at pH 2.0 (4820 IU/ml) was similar to the control incubated under the same conditions except at pH 7.2. The stability of tro-IFN at low pH is similar to that of human fibroblast IFN reported by other workers (Sedmak & Grossberg, 1977; Fantes, 1973).
The antiviral activity of the tro-IFN was stable at 37 °C but was temperature-sensitive at 56 °C. The tro- IFN retained 55 ~ of its antiviral activity in 3 h at 37 °C. However, it lost 50, 75 and 97yo of its antiviral activity in 10, 15 and 60 min respectively at 56 °C. These results differed from the half life values of 3 to 7 min reported by Marshall et al. (1972) for human fibroblast IFN induced by poly(rI)-poly(rC) or virus. Cesario et aL (1977) have also reported the half-life or crude virus and poly(rI). poly(rC)-induced fibroblast IFNs to be 2 and 3 min.
The tro-IFN bound to concanavalin A-Sepharose at high ionic strength (1.0 M-NaC1) and was partially eluted from the column with 0-1 M-~-methyl-D-mannopyrano- side (15 ~ of the amount applied) or 50 ~o ethylene glycol (61~ of the amount applied). However, both 0.1 M-Ct- methyl-D-mannopyranoside and 50~ ethylene glycol resulted in complete recovery of tro-IFN from the concanavalin A-Sepharose column (Fig. 5). The partial displacement of the tro-IFN by the sugar suggested that the binding was partly due to carbohydrate recognition. The complete recovery of the IFN activity with combined sugar and ethylene glycol further suggested that the recognition by concanavalin A was due not only to carbohydrate but also to hydrophobic interaction. Furthermore, following PAS treatment after SDS- PAGE of RP-purified tro-IFN and the IFN fraction from the concanavalin A-Sepharose (from Fig. 5c) gave reddish-pink protein bands typical of glycoproteins (Fig. 5d).
The analysis of the purified tro-IFN by native and SDS-PAGE showed a single protein band with an Mr of 24K (Fig. 6). The Mr of the t ro - IFN, however, differed from the 20K value reported by others (Knight & Fahey, 1980; Knight, 1976; Tan et al., 1979) for IFN-fl induced in other human diploid cells but it was the same of the value obtained by Otto et al. (1980) for IFN-fl induced in the FS-4 strain of human foreskin fibroblast cells by poly(rI).poly(rC). Other workers (Reynolds & Pitha, 1975; Heine et al. 1981; Duc-Goiran et al., 1983) have also reported the Mr of human IFN-fl to be in the range of 20K to 25K and have claimed the size differences to be due to the varying carbohydrate content of the proteins
.~ o.1
o . O • _
0 . 1
(a) . . . . . ~ 2
' , ' I ~ 2
_ _ _ - - ,
2 4 6 8 10 12 Fraction number
1 2 (d) -
Fig. 5. Affinity chromatography of tro-IFN on concanavalin A- Sepharose. (a) RP-purified tro-IFN (12000 IU) was dialysed against column buffer (0.02 M-sodium phosphate buffer pH 7.2, containing 1 M- NaC1 and 1 mM-MnCI2, CaC12 and MgCI2) and was applied onto a concanavalin A Sepharose column (7-5 mmx 50 ram). The column was washed with the column buffer and the IFN activity was eluted with elution buffer B, (0.1 M-~-methyl-~mannopyranoside in column buffer). A bioassay of all fractions showed a recovery of 15 ~ of the total IFN antiviral activity (11) applied. (b) Elution profile of tro-IFN with 50% ethylene glycol in column buffer (B2). Tro-IFN (8500 IU) was applied to the column. Elution of the column with buffer B2 resulted in the recovery of 61 ~ of the total IFN antiviral activity (11) applied. (c) Elution of tro-IFN with both 0-1 M-a-methyl-D-mannopyranoside and 50% ethylene glycol in column buffer (B3). Tro-IFN (20000 IU) was applied to the column. After washing with the column buffer IFN activity was eluted with buffer B 3. A bioassay of all fractions showed complete recovery (100 ~) of the applied tro-tFN antiviral activity (11). The arrows (~) indicate the start of the elution with buffers BI, Bz and B 3. (d) PAS stain of tro-IFN. Lane 1, RP-HPLC-purified tro-IFN; lane 2, tro-IFN eluted from the concanavalin A-Sepharose column in (c).
which may have resulted in the differences in their secondary structures. In view of the differences between the tro-IFN and IFN-fl species from different human diploid cells in heat stability and size (all properties dependent on conformation), they may have structural and functional differences. Detailed structural compari- son of the tro-IFN with other IFN-fl species will have to await elucidation of the primary and secondary struc- tures. This information will enable us to compare the amino acid sequences, the active sites and the three- dimensional structures. The differences between these IFNs, although they have similar antigenic properties, are important especially for identifying a human IFN species which is superior for protection of the human placenta against viral infections.

Downloaded from www.microbiologyresearch.org by
IP: 54.205.18.105
On: Thu, 23 Jun 2016 06:26:26
Short communicat ion 3065
(a) (b)
1 2 1 2
.q--~tro-IFN
Fig. 6. Coomassie blue-stained (a) native PAGE and (b) SDS-PAGE (under reduced conditions) of RP-HPLC-purified tro-IFN. Lanes 1, M, markers; lanes 2, RP-HPLC-purified tro-IFN.
We have explored the use of Blue Sepharose affinity chromatography and RP-HPLC on Separon SGX C-18 to isolate a homogeneous tro-IFN of high quality suitable for structural analysis, monoclonal antibody production and biochemical and clinical studies. The purification method described here has several advantages over a number already reported (Knight, 1976; Rubinstein et al., 1978; Okamura et al., 1980) for purifying human IFNs. The method is simple in that it involves only two steps, it yields a homogeneous product, it does not involve extremes of pH and therefore can be used successfully to purify a number of different IFNs (including acid-labile IFNs) with retention of biological activity, and can easily be scaled-up to meet the growing demand for large amounts of pure IFNs.
The demonstration in our laboratory that the human trophoblast produces large amounts of IFN when exposed in vitro to poly(rI)- poly(rC) is interesting because the trophoblast layer of the human placenta functions as an anatomical and immunological interface between mother and foetus, and also acts as a barrier to the transmission of infection from mother to foetus. It is well documented that maternal infection may spread to the placenta but fail to progress to the foetus (Yamauchi et al., 1974; Klein et al., 1976; Remington & Desmonts, 1976). The mechanisms that determine the ability of the placenta to prevent virus transmission to the foetus are, however, not known, although IFN is known to play a major role with viruses. Since, in addition to its antiviral
activity, IFN influences a variety of other processes ranging from cell surface alterations and growth control to immunoregulation we feel that continued elucidation of the structure of tro-IFNs may contribute to under- standing how they contribute to the specific physiology of the placenta.
The authors wish to thank the Snedkermester Sophus Jacobsens og Hustru Astrid Jacobsens Fond and the Danish Automobile Auctions' Fond for supporting the project.
References
BRADFORD, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72, 248- 254.
CESARIO, T. C., SCHRYER, P. J. & TILLES, J. G. (1977). Relationship between the physicochemical nature of human interferon, the cell induced, and the inducing agent. Antimicrohial Agents and Chemo- therapy 11, 291-298.
DAVEY, M. W., SULKOWSKI, E. & CAgTER, W. A. (1976). Binding of human fibroblast interferon to concanavalin A-agarose. Involve- ment of carbohydrate recognition and hydrophobic interaction. Biochemistry 15, 704-713.
DOERNER, K. C. & WHITE, B. A. (1990). Detection of glycoproteins separated by nondenaturating polyacrylamide gel electropboresis using the periodic acid-Schiffstain. AnalyticalBiochemistry 187, 147- 150.
Doc-GoIRAN, P., ROBERT-GALLIOT, B., CHUDZIO, T. & CrL~NY, C. (1983). Unusual human interferons produced by virus-infected amniotic membranes. Proceedings of the National Academy of Sciences, U.S.A. 80, 2628-2631.
FANTES, K. H. (1973). Purification and physico-chemical properties of interferons. In Interferons and Interferon Inducers, pp. 171-200. Edited by N. B. Finter. New York: North-Holland.
FlSl-mR, P. B. & GRANT, S. (1985). Effects of interferon on differentiation of normal and tumor cells. Pharmacology and Therapy 27, 143-166.
HEINE, J. W., VAN DAMME, J., DE LEY, i . , BILLIAU, A. & DE SOMER, P. (1981). Purification of human fibroblast interferon by zinc chelate chromatography. Journal of General Virology 54, 47-56.
KIRCHNER, H. (1986). The interferon system as an integral part of the defence system against infections. Antiviral Research 6, 1-17.
KLEIN, J. O., REMINGTON, J. S. & MARCY, S. M. (1976). An introduction to infections of the foetus and newborn infant. In Infectious Diseases of the Foetus and Newborn Infants, pp. 1-32. Edited by J. S. Remington & J. O. Klein. Philadelphia: W. B. Saunders.
KLIMAN, H. J., FEINMAN, M. A. & STRAUSS, J. F. (1987). Differenti- ation of human cytotrophoblast into syncytrophoblast in culture. Trophoblast Research 2, 407-421.
KNIGHT, E., JR (1976). Interferon: purification and initial characteri- zation from diploid cells. Proceedings of the National Academy of Sciences, U.S.A. 73, 520-523.
KNIGHT, E., JR & FAHEY, D. (1980). Human fibroblast interferon: an improved purification. Journal of Biological Chemistry 256, 3609- 3611.
LAEMMLI, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, London 227, 680- 685.
MARSSALL, L. W., PITHA, P. M. & CARTER, W. A. (1972). Inactivation of interferon: the effect of protonation. Virology 48, 607-611.
NG, M. H. & VIL~EK, J. (1972). Interferons: physicochemical properties and control of cellular synthesis. Advances in Protein Chemistry, 173-234.

Downloaded from www.microbiologyresearch.org by
IP: 54.205.18.105
On: Thu, 23 Jun 2016 06:26:26
3066 Short communication
OKAMURA, H., BERTHOLD, W., HOOD, L., HUNKAPILLER, M., INOVE, M., SMITH-JoHANNSEN, H. & TAN, Y. H. (1980). Human fibroblas- told interferon: immunosorbent column chromatography and N- terminal amino acid sequence. Biochemistry 19, 3831-3835.
OTTO, M. J., SEDMAK, J. J. & GROSSBERG, S. E. (1980). Enzymatic modifications of human fibroblast and leukocyte interferons. Journal of Virology 35, 390-399.
REMINGTON, J. S. & DESMONTS, G. (1976). Toxoplasmosis. In Infectious Diseases of the Foetus and Newborn Infants. pp. 191-332. Edited by J. S. Remington & J. O. Klein. Philadelphia: W. B. Saunders.
REYNOLDS, F. H., JR & PITHA, P. M. (1975). Molecular weight study of human fibroblast interferon. Biochemical and Biophysical Research Communications 65, 107-111.
RUBINSTEIN, M., RUBINSTEIN, S., FAMILLETI, P. C., GROSS, M. S., MILLER, R. S., WALOMAN, A. A. & PESTKA, S. (1978). Human leukocyte interferon purified to homogeneity. Science 202, 1289- 1290.
SEDMAK, J. J. & GROSSBERG, S. E. (1977). Stabilization of interferons. In Texas Reports on Biology and Medicine, vol. 35, pp. 198-203. Edited by S. Baron & F. Dianzani. Galveston: University of Texas Medical Branch.
TAN, Y. H. & BERTHOLD, W. (1977). A mechanism for the induction and regulation of human fibroblastoid interferon genetic expression. Journal of General Virology 34, 401-411.
TAN, Y. H., BARAKAT, F., BERTHOLD, W., SMITH-JOHANN'SEN, H. & TAN, C. (1979). The isolation and amino acid/sugar composition of human fibroblastoid interferon. Journal of Biological Chemistry 254, 8067-8073.
T6TH, F. D., JurIL, C., NOILSKOV-LAURITSEN, N., PETEKSEN, P. M. & EBBESEN, P. (1990a). Interferon production by cultured human trophoblast induced with double stranded polyribonucleotide. Journal of Reproductive Immunology 17, 217-227.
TbTH, F. D., JUnL, C., NORSKOV-LAURITSEN, N., PETERSEN, P. M. & EBBESEN, P. (1990b). Human trophoblast interferon: pattern of response to priming and superinduction of purified term trophob- lasts and choriocarcinoma cells. Journal of Reproductive Immunology (in press).
Y ~ o c m , T., WILSON, C. & ST GEME, J. W., JR (1974). Transmission of live, attenuated mumps virus to the human placenta. New England Journal of Medicine 290, 710-712.
(Received 2 May 1990; Accepted 17 August 1990)





























