Embed Size (px)
Citation preview

Ther Adv Respir Dis
(2013) 7(3) 175 –200
DOI: 10.1177/ 1753465812472940
© The Author(s), 2013. Reprints and permissions: http://www.sagepub.co.uk/ journalsPermissions.nav
Therapeutic Advances in Respiratory Disease Review
http://tar.sagepub.com 175
IntroductionPulmonary hypertension (PH) is a rare and severe disease, with a progressive elevation of pulmonary arterial pressure (PAP) often leading to right heart failure. Therapeutic strategies currently available, mostly designed to induce vasodilation, alleviate PH symptoms but do not afford a cure. Clinical worsening leads to a necessary heart/lung graft with reduced life expectancy. PH cellular and molecular mechanisms are therefore under investigation in order to define new therapeutic targets. Among the new potential strategies that have emerged recently, targeting of reactive
oxygen species (ROS) appears as an interesting approach in PH treatment. Indeed, ROS play important physiological roles in the pulmonary vasculature, and recent studies have also evi-denced a role for ROS in PH pathophysiology.
After a brief summary of PH pathophysiology and treatments currently available, this review will provide information about ROS production and metabolism in the pulmonary vasculature, ROS-dependent signalling pathways in pulmonary vas-cular cells and will summarize the data showing a role for ROS in the pathogenesis of PH.
Reactive oxygen species as therapeutic targets in pulmonary hypertensionVéronique Freund-Michel, Christelle Guibert, Mathilde Dubois, Arnaud Courtois, Roger Marthan, Jean-Pierre Savineau and Bernard Muller
Abstract: Pulmonary hypertension (PH) is characterized by a progressive elevation of pulmonary arterial pressure due to alterations of both pulmonary vascular structure and function. This disease is rare but life-threatening, leading to the development of right heart failure. Current PH treatments, designed to target altered pulmonary vascular reactivity, include vasodilating prostanoids, phosphodiesterase-5 inhibitors and endothelin-1 receptor antagonists. Although managing to slow the progression of the disease, these molecules still do not cure PH. More effective treatments need to be developed, and novel therapeutic strategies, targeting in particular vascular remodelling, are currently under investigation. Reactive oxygen species (ROS) are important physiological messengers in vascular cells. In addition to atherosclerosis and other systemic vascular diseases, emerging evidence also support a role of ROS in PH pathogenesis. ROS production is increased in animal models of PH, associated with NADPH oxidases increased expression, in particular of several Nox enzymes thought to be the major source of ROS in the pulmonary vasculature. These increases have also been observed in vitro and in vivo in humans. Moreover, several studies have shown either the deleterious effect of agents promoting ROS generation on pulmonary vasculature or, conversely, the beneficial effect of antioxidant agents in animal models of PH. In these studies, ROS production has been directly linked to pulmonary vascular remodelling, endothelial dysfunction, altered vasoconstrictive responses, inflammation and modifications of the extracellular matrix, all important features of PH pathophysiology. Altogether, these findings indicate that ROS are interesting therapeutic targets in PH. Blockade of ROS-dependent signalling pathways, or disruption of sources of ROS in the pulmonary vasculature, targeting in particular Nox enzymes, represent promising new therapeutic strategies in this disease.
Keywords: endothelial dysfunction, inflammation, NAPDH oxidases, Nox enzymes, pulmonary hypertension (PH), pulmonary vasculature, reactive oxygen species (ROS), vascular hyperreactivity, vascular remodelling.
Correspondence to: Véronique Freund-Michel, PhD Laboratoire de Pharmacologie - INSERM U1045, UFR des Sciences Pharmaceutiques, Université Bordeaux Segalen, Case 83, 146 Rue Léo Saignat, 33076 Bordeaux Cedex, France [email protected]
Christelle Guibert, PhD Mathilde Dubois, PhD Arnaud Courtois, PhD Roger Marthan, MD Jean-Pierre Savineau, PhD Bernard Muller, PhD INSERM U1045 ‘Centre de Recherche Cardio-thoracique de Bordeaux’, Bordeaux, France and Université Bordeaux Segalen, Bordeaux, France
472940 TAR731753465812472940Therapeutic Advances in Respiratory DiseaseV Freund-Michel, C Guibert2013

Therapeutic Advances in Respiratory Disease 7 (3)
176 http://tar.sagepub.com
Pathophysiology of PHPH is defined as an elevation of mean PAP above 25 mmHg at rest, compared with normal mean PAP comprised between 10 and 15 mmHg [Simonneau et al. 2009]. PH is characterized by both functional and structural modifications of pulmonary arteries, leading to altered pulmonary vascular reactivity and remodelling of the pulmo-nary arterial wall [Schermuly et al. 2011]. These mechanisms are associated with thrombotic lesions of pulmonary arteries [Humbert et al. 2004; Voelkel et al. 2012]. Increased pulmonary vascular resistance leads to increase in right heart work to maintain cardiac output, ultimately lead-ing to right heart failure and death. PH classifica-tion has been updated recently and PH forms are now divided into five categories: pulmonary arte-rial hypertension belongs to group 1, being idio-pathic, familial or associated with other diseases including for example connective tissue diseases, HIV infection, congenital heart disease or portal hypertension. Group 2 includes PH caused by left-sided heart disease, such as for example cardiomyopathy, diastolic dysfunction or aortic stenosis. Secondary PH forms associated with chronic respiratory diseases such as chronic obstructive pulmonary diseases or pulmonary fibrosis are often named ‘hypoxic’ PH and belong to group 3. Group 4 includes PH due to pulmo-nary embolus or pulmonary thrombosis. Group 5 includes PH due to other miscellaneous causes, which do not fit into the other four categories [Simonneau et al. 2009].
Altered pulmonary vascular reactivity evidenced in PH is characterized by an increased contrac-tion associated with a decreased endothelium-dependent relaxation of pulmonary arteries. This phenomenon is due in particular to an endothe-lial dysfunction leading to an imbalance in the production/bioactivity of both vasoconstrictive mediators such as serotonin or endothelin-1 and vasodilators such as nitric oxide (NO) or prosta-cyclin (PGI2) [Humbert et al. 2004]. The func-tion of the pulmonary arterial smooth muscle is also modified, with altered contractile signalling pathways, such as the RhoA/ROCK pathway [Connolly and Aaronson, 2011], and decreased expression/activity of ions channels, such as volt-age-gated potassium channels [Guibert et al. 2007; Rhodes et al. 2009], leading to excessive contrac-tion of pulmonary arteries.
Structural changes observed in pulmonary arter-ies, also called pulmonary vascular remodelling,
include medial and adventitial thickening in prox-imal arteries, due to decreased apoptosis, hyper-plasia and hypertrophy of smooth muscle cells and of fibroblasts associated with excessive accu-mulation of matrix proteins [Morrell et al. 2009]. Formation of a neointima is also observed, due to accumulation of smooth muscle cells and myofi-broblasts, together with the formation of plexiform lesions resulting from excessive and disorganized smooth muscle and endothelial cell proliferation. In normally nonmuscular distal arteries, forma-tion of a media de novo is observed, resulting from recruitment, differentiation and proliferation of fibroblasts and progenitor cells [Hassoun et al. 2009; Schermuly et al. 2011]. All of these struc-tural changes are therefore leading to arterial wall thickening, thus contributing to the increased pulmonary vascular resistance and elevation of PAP observed in PH.
Current treatments of PHThe current therapeutic strategy developed for PH is to associate a ‘specific’ treatment, against excessive pulmonary vascular contraction and remodelling, to a more ‘conventional’ treatment dedicated to alleviate symptoms, effort and right heart failure [Frumkin, 2012; O’Callaghan et al. 2011]. Conventional treatments comprise lifestyle modification, diuretics to alleviate right heart fail-ure, anticoagulant therapies to reduce risks of thrombotic lesions and oxygen therapy to treat hypoxemia [Galie et al. 2004; Simonneau et al. 2009]. Whereas nonselective systemic vasodila-tors such as calcium channel blockers are effective only in a few groups of responding patients [Montani et al. 2010; Rubin, 1985; Tonelli et al. 2010], more specific vasodilator treatments have been developed for the treatment of PH. Three families of molecules are currently used: vasodilat-ing prostanoids, phosphodiesterase-5 inhibitors and endothelin-1 receptor antagonists [Frumkin, 2012].
Prostacyclin, also named prostaglandin I2 (PGI2), is a potent vasodilator whose endothelial produc-tion is decreased in PH. This decrease therefore contributes to excessive pulmonary arterial con-traction, but also to pulmonary vascular remod-elling, since PGI2 normally also inhibits smooth muscle proliferation, platelet aggregation and exerts anti-inflammatory actions [Gomberg-Maitland and Olschewski, 2008]. Prostacyclin therapy has therefore been employed for over 25 years to treat PH and showed efficiency in alleviating

V Freund-Michel, C Guibert et al.
http://tar.sagepub.com 177
symptoms, and even in increasing patient survival for epoprostenol [Sitbon et al. 2002]. However, because of its complex intravenous administra-tion and its numerous adverse effects, more stable prostacyclin analogues have been developed with other routes of administration, in particular orally, subcutaneously or inhaled active compounds [Yildiz, 2009]. Therapy strategies combining oral prostacyclin analogues with phosphodiesterase-5 inhibitors and/or endothelin-1 receptor antago-nists have also been developed to enhance thera-peutic efficiency.
Endothelin-1, activating ETA and ETB receptors, is a potent vasoconstrictor and also participates in smooth muscle proliferation, fibrosis and inflam-mation [Kim and Rubin, 2002]. Activation of ETA receptors, mainly expressed on smooth mus-cle cells, induces vasoconstriction whereas acti-vation of ETB receptors, also expressed on endothelial cells, induces NO release and vasodi-lation [Dupuis, 2001]. However, both ETA selec-tive antagonists such as ambrisentan and dual ETA/ETB antagonists such as bosentan are used in clinics and have all been shown to alleviate PH symptoms and to improve exercise capacity [Galie et al. 2008a, 2008b; O’Callaghan et al. 2011; Rubin et al. 2002].
NO-mediated relaxation of the pulmonary vas-cular smooth muscle involves synthesis of cyclic guanosine monophosphate (cGMP), which is then degraded by phosphodiesterase-5 (PDE5). Inhibitors of PDE5 such as sildenafil and tadalafil are therefore also used in PH treatment. PDE5 is abundantly expressed in the pulmonary vascula-ture [Hanson et al. 1998; Pauvert et al. 2003] and PDE5 inhibitors induce beneficial pulmonary vasodilation and antiproliferative effects in PH patients [Archer and Michelakis, 2009; Montani et al. 2009].
In summary, the great progress made in the com-prehension of PH pathophysiological mecha-nisms has led to the development of these specific treatments and greatly improved patient’s clinical course and life expectancy. However, although being able to alleviate PH symptoms, these treat-ments still do not afford a cure, and this shows the need to persevere in the comprehension of PH cellular and molecular mechanisms in order to define new therapeutic strategies. Among the var-ious targets under investigation at the moment, targeting of ROS appears as an interesting new approach in PH treatment.
Targeting ROS as a novel therapeutic strategy in PHROS are cellular products derived from oxygen metabolism. Depending on their local concentra-tion, ROS can play physiological but also patho-physiological roles, becoming harmful to cells. Indeed, at low concentrations, ROS are intracel-lular signalling messengers, and ROS production is counterbalanced by antioxidant systems con-tributing to ROS metabolism [Touyz, 2004]. However, an imbalance between ROS production and ROS metabolism can lead to increased ROS levels, a situation named oxidative stress, with alteration by oxidation of essential cellular compo-nents such as lipids, proteins, mitochondria and even DNA, which can ultimately lead to cell death.
The vasculature is an important source of ROS [Touyz, 2004], and many studies have shown a role for ROS in vascular physiology and pathophysiol-ogy. Indeed, oxidative stress is a mediator of vas-cular injury in cardiovascular diseases such as systemic hypertension [Touyz, 2004], atheroscle-rosis [Victor et al. 2009] or ischemia–reperfusion [Murphy and Steenbergen, 2008]. A growing body of evidence, summarized in the following sections, also demonstrates a role for ROS and for oxidative stress in various aspects of PH pathophysiology.
Production and metabolism of ROS in the pulmonary vasculatureBiology of ROS. Many products are generated in normal cell respiration, derived from the reduc-tion of molecular oxygen and all termed ROS. However, ROS have different chemical proper-ties, being either free or nonfree radicals. Indeed, some ROS have an unpaired electron on their outer orbital, such as superoxide (O2
) or the hydroxyl radical (HO), leading to a high chemi-cal reactivity. Other ROS, such as hydrogen perox-ide (H2O2), peroxynitrites (ONOO-), hypochlorous acid (HOCl), ozone (O3), singlet oxygen (1O2) or lipid peroxides (LOOH), although lacking an unpaired electron, are still chemically reactive by exchanging some electrons with other molecules [Guzik and Harrison, 2006].
ROS generation often begins with one- or two-electron reduction of molecular oxygen to form O2
or H2O2 (Figure 1). Under ambient condi-tions, superoxide then rapidly dismutates to H2O2, either spontaneously, or catalysed by superox-ide dismutase (SOD). H2O2 is then converted into water by catalase, extracellular glutathione

Therapeutic Advances in Respiratory Disease 7 (3)
178 http://tar.sagepub.com
peroxidase or thiols. When H2O2 is bound to per-oxidases such as catalase, it also forms a metabo-lite named compound I which can oxidize NO to nitrogen dioxide anion NO2
- . Compound I can then react with NO2
- to form nitrogen dioxide radical NO2
which can induce nitrosylation reac-tions on protein tyrosine residues or on lipids [Rubbo et al. 2009]. Finally, H2O2 can also undergo a Fenton reaction in the presence of heavy metals to form the highly reactive hydroxyl radical HO. H2O2 and HO can then oxidize lipids, proteins and DNA, and can lead to patho-logical cell response when being produced in excess [Cai, 2005].
Sources of ROS in the pulmonary vasculature. In mammalian cells, enzymatic sources of ROS include NADPH oxidases, uncoupled nitric oxide synthase (NOS), cytochromes P450, xanthine oxidase (XO), the mitochondrial electron trans-port chain, and other enzymes such as lipoxygen-ases, cyclooxygenases or peroxidases as well as
hemoproteins such as heme and hematin [Griendling et al. 2000]. All vascular cell types are able to pro-duce ROS either from metabolic or from enzy-matic sources. However, in the pulmonary vasculature, ROS main sources are NADPH oxi-dases, uncoupled endothelial NOS (eNOS), XO as well as the mitochondrial electron transport chain [Li and Shah, 2004; Lyle and Griendling, 2006].
NAPDH oxidases. NADPH oxidases are enzymes composed of multiple subunits that catalyse O2
production after O2 reduction, using NADPH (reduced form of the nicotinamide adenine dinucleotide phosphate NADP) as the electron donor [Sedeek et al. 2009]. NADPH oxi-dases were initially identified in phagocytes of the innate immune response, where they produce large amounts of O2
and contribute to the extracellular oxidative burst dedicated to kill invading patho-gens [Quinn and Gauss, 2004]. These NADPH oxidases were therefore termed phox (standing for
Figure 1. Generation and metabolism of ROS.Molecular oxygen can undergo electronic rearrangement to form singlet oxygen (1O2). Molecular oxygen can also form superoxide (O2
) or hydrogen peroxide (H2O2) after one or two electron reduction, respectively. O2 can
rapidly react with nitric oxide (NO) to form peroxynitrites (ONOO-). H2O2 is also produced after O2 dismutation,
either spontaneously, or through action of superoxide dismutase (SOD). H2O2 can then produce the hydroxyl radical (HO), after one electron reduction, through the Fenton reaction, or indirectly through generation of hypochlorous acid (HOCl). HO is then degraded to water. H2O2 can also be directly converted to water by catalase. In addition, H2O2 can also oxidize Fe3+-containing enzymes to produce compound I which can oxidize NO to form nitrogen dioxide anion (NO2
-), therefore leading to formation of nitrogen dioxide radical (NO2).
All of these highly reactive species (shown in blue) can then alter lipids, proteins and DNA function through oxidation, nitrosation and nitration reactions.

V Freund-Michel, C Guibert et al.
http://tar.sagepub.com 179
phagocyte oxidase). However, NADPH oxidases were later also identified in nonphagocytic cells, and the term Nox (standing for NAPDH oxidase) was introduced to designate these nonphagocytic NADPH oxidases [Brandes et al. 2010].
The mammalian Nox family comprises seven members: Nox1, Nox2, Nox3, Nox4, Nox5, Duox1 and Duox2 [Brandes et al. 2010]. They all differ in their mode of activation, expression, and/or interaction with other proteins. In the cardio-vascular system, Nox1, Nox2, Nox4 and Nox5 have been detected [Brandes et al. 2010]. The prototypical phagocytic NADPH oxidase (Nox2) is composed of five subunits (Figure 2): the two membrane-bound subunits gp91phox (the cata-lytic subunit, also termed Nox2) and p22phox, forming a heterodimeric complex named cytochrome b558, and the three cytosolic subu-nits p40phox, p47phox and p67phox. In stimu-lated cells, p47phox phosphorylation leads to
assembly of the three cytosolic subunits to form a complex that translocates to the membrane and associates with cytochrome b558 to assemble the active oxidase [Touyz and Briones, 2010]. This activation also requires the small G proteins Rac (Rac1 or Rac2) and Rap1A [Bokoch and Knaus, 2003]. Nox1 is the closest homolog of Nox2, and requires the membrane subunit p22phox as well as at least two other cytosolic subunits termed Noxo1 (Nox organizer 1, which is a p47phox ana-logue) and Noxa1 (Nox activator 1, which is a p67phox analogue) (Figure 2). Nox4 was initially termed Renox (for renal oxidase, as its expression is abundant in the kidney). Nox4 differs from Nox1 and Nox2 in the fact that it mainly pro-duces H2O2 directly, whereas Nox1 and Nox2 first produce O2
that is then converted to H2O2. Moreover, although Nox4 also requires the mem-brane subunit p22phox for its activity, it does not seem to require recruitment of the cytosolic subu-nits p40phox, p47phox, p67phox or of any small
Figure 2. Structure of NADPH oxidases expressed in pulmonary arteries.Nox core structure consists of six transmembrane domains (shown in blue) and a cytosolic C-terminal end. Nox1, 2 and 4 are constitutively associated to the membrane-bound subunit p22phox (shown in yellow), that acts as a maturation and stabilization partner. Regulatory cytosolic subunits are necessary for Nox1 and Nox2 structure and activity. Nox1 associates with Noxo1 and Noxa1. Nox2 associates with p40phox, p47phox and p67phox. Both Nox1 and Nox2 associate with the small G protein Rac1 and/or Rac2. Noxa1, p40phox and p47phox are cytosolic organizers (shown in orange), whereas Noxa1 and p67phox (shown in green) as well as Rac proteins (shown in purple) participate to Nox1 and Nox2 activity. By contrast, Nox4 does not require any cytosolic subunit for its structure or activity.

Therapeutic Advances in Respiratory Disease 7 (3)
180 http://tar.sagepub.com
G protein (Figure 2). Nox4 is therefore the only NADPH oxidase being able to produce ROS con-stitutively. In contrast to the other Nox isoforms, Nox5 is expressed in humans but not in rodents. Moreover, Nox5 does not require the membrane subunit p22phox or any other cytosolic subunit for its activity. It is the only Nox isoform that is directly activated by Ca2+, due to the presence of four Ca2+ binding sites (EF hands) in its amino terminus [Jagnandan et al. 2007].
In the lung, expression of the membrane-bound subunits Nox1, Nox2, Nox4 and p22phox have been shown in pulmonary vessels, together with expression of the cytosolic subunits Noxa1, Noxo1, p40phox, p47phox and p67phox [Perez-Vizcaino et al. 2010] (Figures 2 and 3). In cul-tured cells, expression of Nox2 and Nox4 has been reported in pulmonary arterial endothelial cells (PAECs) [Bayraktutan et al. 2000; Griffith et al. 2009], in adventitial fibroblasts [Haurani and Pagano, 2007], and in pulmonary arterial smooth muscle cells (PASMCs) [Brandes and Kreuzer, 2005; Mittal et al. 2007]. In all of these cells, Nox4 is the predominant Nox isoform expressed [Perez-Vizcaino et al. 2010]. Nox1 expression seems to be restricted to PASMCs and is present in low concentrations in basal states of these cells [Mittal et al. 2007] (Figure 3).
Uncoupled endothelial NOS. NOS is the enzyme responsible for NO synthesis. Three iso-forms have been identified: NOS1 (neuronal NOS or nNOS), NOS2 (inducible NOS or iNOS) and NOS3 (endothelial NOS or eNOS) [Zucker-braun et al. 2011]. Active eNOS is a homodimer and catalyses the formation of NO from L-arginine. Each monomer contains a carboxy-terminal reduc-tase domain and an amino-terminal oxygenase domain. The reductase domain is composed of two flavinic cofactors (flavin mononucleotide [FMN] and flavin adenine dinucleotide [FAD]) binding sites and also contains a NADPH bind-ing site. The oxygenase domain contains binding sites for heme, for the substrate L-arginine and for the cofactor tetrahydrobiopterin (BH4). Oxy-genase and reductase domains are linked with a small sequence of amino acids that contains a calmodulin binding site [Alderton et al. 2001]. Heme groups are essential for dimerization of both monomers and for promoting electron transfer from the flavin to the heme of the other monomer. Finally, a zinc ion binds at both mono-mers to stabilize the dimeric structure [Weseler and Bast, 2010]. An electron transfer is catalysed
within the reductase domain via the flavins FMN and FAD to the heme in the oxygenase domain. Binding of the calcium-calmodulin complex to the reductase domain facilitates this electron transfer. O2 is then reduced at the heme of the oxygenase domain and L-arginine is oxidized to L-citrulline and NO. Many signals can regulate eNOS activity, in particular intracellular calcium concentration or phosphorylation in the reduc-tase and calmodulin-binding domains through the action of kinases such as protein-kinase B (PKB, also named Akt), protein-kinase A (PKA) or calmodulin-dependent kinase II (CamKII) [Fleming, 2010; Forstermann, 2006].
Under some conditions, eNOS generates O2
rather than NO, a phenomenon that is known as NOS uncoupling: electrons flowing from the reductase domain to the oxygenase domain are diverted to molecular oxygen rather than to L-arginine, with production of O2
from the oxy-genase by dissociation of the ferrous–dioxygen complex [Belik et al. 2009; Konduri et al. 2007] (Figure 3). The limited availability of the cofactor BH4 seems to play a major role in eNOS uncou-pling. Indeed during normal catalytic function of eNOS, BH4 acts as an electron donor and BH4 bioavailability depends on a balance between de novo synthesis by GTP cyclohydrolase-1 (GTPCH-1, the first and limiting step for BH4 biosynthesis from GTP), loss of BH4 (due to oxi-dation of BH4 to form dihydrobiopterin BH2) and recycling of BH2 into BH4 by dihydrofolate reduc-tase (DHFR) [Crabtree and Channon, 2011]. However, when ROS levels are increased (follow-ing activation of NADPH oxidases, for example), O2
can react with NO to the highly reactive ONOO- that is able to oxidize BH4 to BH2. When BH2 is not fully recycled, it can compete with BH4 for binding to eNOS, resulting in eNOS uncou-pling [Crabtree and Channon, 2011; Gielis et al. 2011; Noguchi et al. 2011; Weseler and Bast, 2010]. Rather than absolute BH4 concentrations, the eNOS/BH4 stoichiometry and the biopterin redox status now appear as key determinants of eNOS uncoupling [Bendall et al. 2005; Crabtree and Channon, 2011]. Other potential and biopterin-independent mechanisms of eNOS uncoupling include oxidation of the zinc-thiolate cluster of eNOS with subsequent release of zinc [Zou et al. 2002], L-arginine depletion [Forstermann, 2006], accumulation of the endogenous eNOS inhibitors methylarginines [Druhan et al. 2008; Forstermann, 2006], defect in heat shock protein 90/eNOS interaction [Sud et al. 2007],
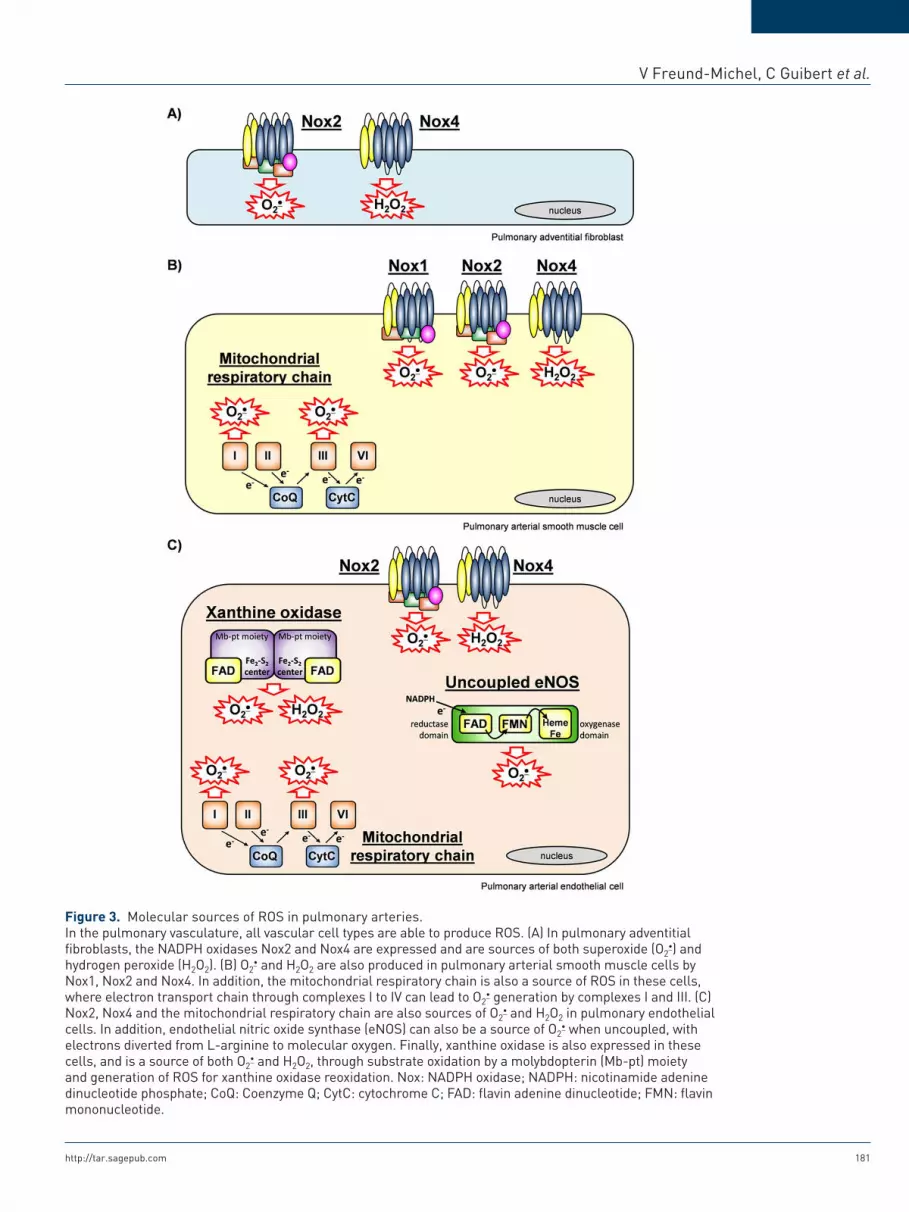
V Freund-Michel, C Guibert et al.
http://tar.sagepub.com 181
Figure 3. Molecular sources of ROS in pulmonary arteries.In the pulmonary vasculature, all vascular cell types are able to produce ROS. (A) In pulmonary adventitial fibroblasts, the NADPH oxidases Nox2 and Nox4 are expressed and are sources of both superoxide (O2
) and hydrogen peroxide (H2O2). (B) O2
and H2O2 are also produced in pulmonary arterial smooth muscle cells by Nox1, Nox2 and Nox4. In addition, the mitochondrial respiratory chain is also a source of ROS in these cells, where electron transport chain through complexes I to IV can lead to O2
generation by complexes I and III. (C) Nox2, Nox4 and the mitochondrial respiratory chain are also sources of O2
and H2O2 in pulmonary endothelial cells. In addition, endothelial nitric oxide synthase (eNOS) can also be a source of O2
when uncoupled, with electrons diverted from L-arginine to molecular oxygen. Finally, xanthine oxidase is also expressed in these cells, and is a source of both O2
and H2O2, through substrate oxidation by a molybdopterin (Mb-pt) moiety and generation of ROS for xanthine oxidase reoxidation. Nox: NADPH oxidase; NADPH: nicotinamide adenine dinucleotide phosphate; CoQ: Coenzyme Q; CytC: cytochrome C; FAD: flavin adenine dinucleotide; FMN: flavin mononucleotide.

Therapeutic Advances in Respiratory Disease 7 (3)
182 http://tar.sagepub.com
S-glutathionylation of eNOS [Hu et al. 2010] or thiyl radical formation on the enzyme [Chen et al. 2011].
Xanthine oxidase. XO and xanthine dehy-drogenase (XDH) are xanthine oxidoreductases (XORs), and are involved in the last steps of purine metabolism, catalysing the transforma-tion of hypoxanthine and xanthine to uric acid [Jarasch et al. 1981]. XO and XDH are encoded by the same gene, and can be interconverted in vivo [Nishino et al. 2008]. XDH uses NAD+ as an electron acceptor, thus producing NADH, whereas XO prefers O2 as an electron acceptor, therefore generating O2
and H2O2. The ratio of XO to xanthine dehydrogenase in pulmonary vascular cells is therefore critical to determine the amount of ROS produced by this enzyme system in the pulmonary vasculature [Borges et al. 2002].
XO is composed of two monomers, each contain-ing a molybdopterin moiety that catalyses oxida-tion of a substrate. The accepted electrons are then rapidly transported via a Fe2–S2 centre to the cofactor FAD, which then reduces the oxidant substrate O2 [Harrison, 2002]. Reoxidation of XO needs the transfer of six electrons to O2, therefore generating two molecules of H2O2 and two mole-cules of O2
[Weseler and Bast, 2010] (Figure 3). XO expression has been evidenced in the pulmo-nary arterial endothelium in vitro and in vivo [Partridge et al. 1992; Perez-Vizcaino et al. 2010].
Mitochondrial electron transport chain. Mito-chondria are also a major source of ROS in the pulmonary vasculature. The mitochondrial matrix contains enzymes that participate in the metabo-lism of pyruvate and fatty acids to produce acetyl CoA. The matrix also contains enzymes of the cit-ric acid cycle which can then oxidize acetyl CoA to CO2, generating three molecules of NADH and one molecule of FADH2 during this process. NADH and FADH2 are then used as electron donors by the mitochondrial electron transport chain to generate the proton gradient necessary to produce ATP.
The mitochondrial respiratory chain is the main energy source for the cell. Indeed, more than 95% of O2 consumed by cells is reduced by four elec-trons to yield two molecules of H2O. This occurs at the inner mitochondrial membrane, via mito-chondrial electron transport chain complexes I to IV [Birukov, 2009; Fuchs et al. 2010] (Figure 3). NADH is oxidized by complex I, and FADH2 is
oxidized by complex II, leading to reduction of ubiquinone to ubiquinol. Two electrons are car-ried by ubiquinol from complex I to complex III (coenzyme Q: cytochromes c-oxidoreductase). Another pair of electrons is transferred to ubiqui-none at complex II, and is then also carried by ubiquinol to complex III. In complex III, one electron is transferred at the Q0 site to the Rieske iron–sulfur protein and then to cytochrome c, while the other electron is transferred to cytochrome b, followed by a transfer to ubiqui-none at the Q0 site. Cytochrome c then donates electrons to O2 in complex IV, thus producing H2O [Shimoda and Undem, 2010].
Under physiological conditions, approximately 2–3% of the oxygen consumed by mitochondria is incompletely reduced, and mitochondria there-fore also produce O2
as a byproduct. Production of superoxide can occur at the complex I by auto-oxidation of flavins, with O2
formed being able to enter the matrix. Production of superoxide can also occur at the complex III at both Q0 and Qi sites, with O2
formed being able to enter either the matrix or the mitochondrial intermembrane space [Perez-Vizcaino et al. 2010; Stowe and Camara, 2009] (Figure 3). In the matrix, O2
is converted into H2O2 by mitochondrial Mn SOD, and then to water by glutathione peroxidase [Shimoda and Undem, 2010]. In the intermembrane space, O2
can be degraded by the cytosolic CuZn superoxide dismutase, can be scavenged by cytochrome c, or can enter the cytosol via voltage-dependent anion channels [Han et al. 2003; Waypa et al. 2002].
In the pulmonary vasculature, production of ROS by mitochondria is regulated by modulation of mitochondria biogenesis and bioenergetics [Erusalimsky and Moncada, 2007]. Modulation of these processes are dependent of multiple factors, including for example the availability of electron donors or the activity of cytokines and vasoactive mediators such as NO, angiotensin II or thrombox-ane A2 [Bailey et al. 2005; Rathore et al. 2006; Waypa et al. 2010]. In addition, a ROS-induced ROS release mechanism may also occur in the pul-monary vasculature, with for example angiotensin II and hypoxia triggering an increased ROS pro-duction after Nox activation in PASMC, leading to a subsequent increase in mitochondrial ROS pro-duction [Rathore et al. 2008]. Finally, the concentra-tion of oxygen in pulmonary tissues is also particularly important in the modulation of ROS production by mitochondria, with ROS levels being elevated dur-ing hypoxia [Stowe and Camara, 2009].

V Freund-Michel, C Guibert et al.
http://tar.sagepub.com 183
Antioxidant systems in the pulmonary vasculature. Enzymatic and nonenzymatic antioxidant sys-tems are present in vascular tissues to protect against deleterious oxidative stress. Pulmonary vascular nonenzymatic antioxidants can scavenge HO and other free radicals and include ascor-bate, glutathione, bilirubin, tocopherols and uric acid [Tajima et al. 2009]. Major pulmonary vas-cular enzymatic antioxidants include SOD, gluta-thione peroxidases, catalases, thioredoxin and peroxiredoxin [Touyz and Briones, 2010]. The major vascular SOD is the extracellular SOD that is produced and secreted by vascular smooth muscle cells and then binds to glycosaminogly-cans in the vascular extracellular matrix [Gongora et al. 2006].
ROS-dependent signalling pathways in pulmonary vascular cellsROS serve as important intracellular and intercel-lular messengers to regulate a variety of signalling pathways in pulmonary vascular cells. This redox signalling occurs mainly through reactions with protein residues, either with cysteine residues and formation of disulfide bridges, or through protein carbonylation [Perez-Vizcaino et al. 2010; Wong et al. 2008]. The main ROS targets in the pulmonary vasculature are described below and in Figure 4.
NO/guanylyl cyclase pathway. When superoxide production is increased, it can rapidly interact with NO and therefore attenuate its capacity to stimu-late soluble guanylyl cyclase (sGC) in the systemic and pulmonary vasculatures [Perez-Vizcaino et al. 2010]. Moreover, oxidative stress can oxidize the ferrous (Fe2+)–heme and thiol sites on sGC which control its activation by NO, and therefore contrib-ute to decreased cGMP production and smooth muscle relaxation [Wolin et al. 2010].
In addition, recent data obtained in our group show that ROS production in PASMC reduces endothe-lial NO-dependent control of pulmonary vasoreac-tivity, by targeting PAEC via myoendothelial gap junctions [Billaud et al. 2009]. This is the first direct evidence of a smooth muscle negative feedback on NO-dependent endothelial vasodilatation, through O2 generation and decreased NO-induced cGMP
production [Billaud et al. 2009].
Redox-sensitive enzymesReceptor and nonreceptor tyrosine kinases.
Classical activation of growth factor receptors by their agonist can be redox sensitive. For example,
activation of its receptor by the platelet-derived growth factor (PDGF) is ROS-dependent [Sund-aresan et al. 1995]. In addition, many studies have also focused on transactivation of growth factor receptors, which is a way of activating the recep-tor tyrosine kinase in the absence of any extra-cellular agonist [Knock and Ward, 2011]. This mechanism has been evidenced in particular in vascular smooth muscle cells for the epidermal growth factor receptor (EGFR) whose activation is induced by hydrogen peroxide or peroxynitrites in the absence of EGF [Knock and Ward, 2011; Ushio-Fukai et al. 2001]. Although these mech-anisms have been evidenced in systemic vessels only, growth factors such as EGF or PDGF play an important role in particular in vascular remod-elling observed in pulmonary arteries [Thomas, 2010]. Such ROS-dependent mechanism may also occur in the pulmonary vasculature and may therefore contribute to PH pathophysiology.
Among nonreceptor tyrosine kinases, the src fam-ily is of particular interest in the pulmonary vas-culature [Knock and Ward, 2011]. Indeed, activation of Src kinase, in relation to activation of transcription factors such as the signal transducer and activator of transcription (STAT) 3 and the nuclear factor of activated T cells (NFAT), seems to play a pathophysiological role in PH [Courboulin et al. 2011; Paulin et al. 2011]. Since Src kinases have been implicated in regulation of hypoxia-induced contraction of pulmonary arteries [Knock et al. 2008], and since this mechanism seems to be, at least in part, mediated by ROS generation [Knock et al. 2008], the src kinase family may play a pivotal role in ROS-dependent signalling path-ways in the pulmonary vasculature [Griendling et al. 2000; Knock and Ward, 2011], in both smooth muscle and endothelial cells. In the latter, the stim-ulatory effect of hydrogen peroxide on eNOS activ-ity is src-dependent [Rafikov et al. 2011].
Protein tyrosine phosphatases. Protein tyros-ine phosphorylation is one of the major post-tran-scriptional mechanisms and plays a critical role in many cell functions such as for example pro-liferation, differentiation and migration. The level of tyrosine phosphorylation is tightly regulated with a balance between protein tyrosine kinases (PTK) and protein tyrosine phosphatases (PTP) [Paravicini and Touyz, 2006]. All PTP possess a reactive and redox-regulated cysteine, necessary for protein dephosphorylation. ROS-induced oxi-dation of this cysteine residue renders PTP totally inactive [Tonks, 2005]. In particular, studies have

Therapeutic Advances in Respiratory Disease 7 (3)
184 http://tar.sagepub.com
shown ability of hydrogen peroxide, superoxide and peroxynitrites to inhibit PTP rapidly and irreversibly [Lee and Esselman, 2002; Paravicini and Touyz, 2006]. Since a role of PTP inhibition has recently been shown in PH, in particular in signalling pathways activated by growth factors such as PDGF [ten Freyhaus et al. 2011], redox-dependent inhibition of PTP by ROS might con-tribute to these mechanisms.
Serine/threonine kinases. Mitogen-activated protein kinases (MAPK) are a family of serine/
threonine kinases that control many cellular effects such a proliferation, survival, differentiation, apop-tosis and stress signals [Griendling et al. 2000]. Activation of MAPK depends on many upstream phosphorylations, leading to activation of mul-tiple signalling pathways. MAPK kinase redox sensitivity has been known for a long time, since exogenous ROS such as hydrogen peroxide and superoxide are able to activate MAPK in endothelial [Huot et al. 1997, 1998] and smooth muscle cells [Ushio-Fukai et al. 1998; Yoshizumi et al. 2000]. In the vasculature, all of the major
Figure 4. Redox-sensitive signalling pathways in pulmonary vascular cells and roles of ROS in pulmonary hypertension.Increased ROS production in the pulmonary vasculature influences redox-sensitive signalling pathways in pulmonary vascular cells, leading to activation of redox-sensitive transcription factors and increased expression of many proteins involved in pulmonary vascular remodelling, inflammation and altered reactivity (both endothelium and smooth muscle dysfunctions).EGF: epidermal growth factor; EGFR: epidermal growth factor receptor; PDGF: platelet-derived growth factor; PDGFR: platelet-derived growth factor receptor; MAPK: mitogen-activated protein kinase; ROCK: Rho kinase; PKC: protein kinase C; JAK: janus kinase; Pyk: prolin-rich tyrosine kinase; PTP: protein tyrosine phosphatases; SHP-2: SH2 domain-containing protein tyrosine phosphatase-2; PLD: phospholipase D; sGC: soluble guanylyl cyclase; MMPs: matrix metalloproteinases; COX: cyclo-oxygenases; PECAM: platelet/endothelial cell adhesion molecule; ICAM: intercellular adhesion molecule; VCAM: vascular cell adhesion molecule; HIF: hypoxia inducible factor; Egr-1: early growth response-1 transcription factor; STAT: signal transducer and activator of transcription; NF-κB: nuclear factor-κB; AP-1: activator protein-1; TGF-β1: transforming growth factor-β1; FGF-2: fibroblast growth factor-2; VEGF: vascular endothelial growth factor; TNF-α: tumor necrosis factor-α; IL: interleukin; eNOS: endothelial nitric oxide synthase; TRP: transient receptor potential channels; KATP: ATP-sensitive potassium channels; KCa: calcium-activated potassium channels; Kv: voltage-gated potassium channels.

V Freund-Michel, C Guibert et al.
http://tar.sagepub.com 185
MAPK (extracellular-regulated protein kinase or ERK, p38 and c-jun N-terminal kinase [JNK]) are activated by growth factors such as angio-tensin II or PDGF [Sundaresan et al. 1995; Ushio-Fukai et al. 1998]. More recently, stud-ies have shown that intracellular ROS, mainly derived from NADPH oxidases, are involved in these signalling pathways [Meloche et al. 2000; Torrecillas et al. 2001; Viedt et al. 2000]. Less is known about redox-induced regulation of MAPK in the pulmonary vasculature. How-ever, ROS have been shown to activate ERK and p38 pathways in PAEC [Usatyuk et al. 2003]. Inversely, ROS synthesis in PAEC by NADPH oxidases is dependent upon activa-tion of ERK and p38 pathways [Parinandi et al. 2003]. These results therefore suggest an interrelation between ROS and MAPK in pulmonary endothelial cells after activation by growth factors.
Rho kinases (ROCK) are serine/threonine kinases involved in the regulation of vascular tone, and in the development of several cardiovascular dis-eases [Perez-Vizcaino et al. 2010]. Two isoforms have been identified and named ROCK-1 and ROCK-2, and both are expressed in the vascular smooth muscle [Loirand et al. 2006]. ROCKs are activated by the small G protein RhoA and then phosphorylate the myosin light chain phosphatase (MLCP), which becomes inactive. Myosin phos-phorylation is therefore increased, thus increasing contraction of the smooth muscle, independently of any increase in cytosolic calcium concentra-tions, a mechanism termed calcium sensitization [Resta et al. 2010]. Such calcium sensitization through activation of the RhoA/ROCK pathway may contribute to the smooth muscle dysfunction observed in PH [Connolly and Aaronson, 2011; Oka et al. 2008], and a relationship between ROS and the RhoA/ROCK pathway has been described in pulmonary arteries [Connolly and Aaronson, 2011; Oka et al. 2008]. Indeed, in chronic hypoxia, RhoA activity is enhanced and is related to increased ROS production [Broughton et al. 2010; Jernigan et al. 2008]. Moreover, a super-oxide generator causes ROCK-2 translocation to the nucleus in rat pulmonary arteries and induces a vasoconstriction sensitive to a ROCK inhibitor [Knock et al. 2009]. Hydrogen peroxide may also be able to increase RhoA activity in PAECs and PASMCs [Chi et al. 2010] although controversial results have been observed in other studies [Knock and Ward, 2011; Perez-Vizcaino et al. 2010].
The protein kinase C family contains several iso-forms belonging either to conventional PKC or cPKC (α, β1, β2 and γ), to novel PKC or nPKC (δ, ε, η, θ, υ, and µ) or to atypical PKC or aPKC (ζ and λ/ι) [Reyland, 2009]. In pulmonary arter-ies, various PKC isoforms are expressed in fibro-blasts, endothelial and smooth muscle cells [Dempsey et al. 2007]. Hydrogen peroxide and peroxynitrites can activate PKC in PASMCs [Chakraborti and Michael, 1993]. In addition, hydrogen-peroxide-induced calcium sensitization is associated with PKCα activation, and is blocked by PKC inhibitors [Pourmahram et al. 2008]. Moreover, hypoxia-induced ROS production is PKC-dependent in PASMCs [Rathore et al. 2008] and in PAECs [Chatterjee et al. 2011]. These results therefore suggest a role of ROS in PKC-dependent signalling pathways in pulmo-nary vascular cells.
The serine/threonine kinase Akt, also called pro-tein kinase B or PKB, is a downstream target of phosphatidylinositol-3 kinase, and plays a major role in various cellular processes, including cell proliferation and survival, as well as in protein synthesis [Coffer et al. 1998]. A relation between Akt and ROS has been evidenced in systemic vas-cular smooth muscle cells from various origins, since exogenous hydrogen peroxide induces Akt activation in these cells [Ushio-Fukai et al. 1999]. In addition, angiotensin-II-induced activation of Akt is inhibited by catalase, suggesting a role for ROS in agonist-induced Akt activation [Ushio-Fukai et al. 1999]. In the pulmonary vascula-ture, ROS generation by NADPH oxidase in PASMC leads to redox-sensitive activation of Akt [Djordjevic et al. 2005]. Moreover, in the same cells, expression of the fibroblast growth factor-2 (FGF-2) is mediated by an increase in superoxide levels via NADPH oxidase activation, a mecha-nism that is blocked by a pharmacological inhibi-tor of Akt [Black et al. 2008]. In addition, chronic intermittent hypoxia causes PH and is associated with increased lung levels of NADPH oxidases as well as increased activity of PDGFR and Akt [Nisbet et al. 2009]. These mechanisms are reduced in Nox2 knockout mice [Nisbet et al. 2009], there-fore suggesting a role for NADPH oxidase-derived ROS in Akt activation associated with PH.
Other candidate enzymes. Many other enzymes appear to be redox sensitive and are activated by addition of exogenous ROS [Griendling et al. 2000]. For example, phospholipase D is acti-vated by hydrogen peroxide in endothelial cells

Therapeutic Advances in Respiratory Disease 7 (3)
186 http://tar.sagepub.com
[Natarajan et al. 1993], and PDGF-induced acti-vation of STAT 1 is inhibited by antioxidants [Simon et al. 1998]. In addition, activation of many other kinases such as Fyn kinase, Proline-rich tyrosine kinase (Pyk) or JAK, seem to be ROS-dependent in vascular tissues from other origins than the lung and may therefore also con-tribute to the regulation of pulmonary vascular tone in physiological and pathophysiological con-ditions [Knock and Ward, 2011].
Ion channels. Voltage-gated potassium channels (Kv) play an important role in the regulation of pulmonary vascular tone, and their inhibition seems to be involved in PH pathophysiological mechanisms [Moudgil et al. 2006]. Effects of ROS on Kv channels have been evidenced in pul-monary arteries. Indeed, in PASMC, hypoxia induces ROS production and leads to inhibition of Kv channels [Bonnet and Archer, 2007]. More-over, hydrogen peroxide homologues also block Kv channel activity, whereas hypoxia-induced Kv channel inhibition is prevented by catalase [Cogolludo et al. 2006].
Transient receptor potential (TRP) channels are composed of several subfamilies and contribute to cation entry in pulmonary arteries [Dietrich and Gudermann, 2011; Guibert et al. 2011]. In particular, expression of TRP channel subtypes belonging to the canonical (TRPC), melastatin-related (TRPM) and vanilloid-related (TRPV) subfamilies have been identified in pulmonary arteries [Yang et al. 2010]. Although direct evi-dence of TRP modulation by ROS in pulmonary arteries is lacking, ROS-induced regulation of TRP channels has been observed in other vascu-lar tissues [Song et al. 2011], and may therefore contribute to regulation of calcium entry, in par-ticular in PASMC.
Redox-sensitive gene expression. Many cardio-vascular genes are redox sensitive, with ROS reg-ulating expression of adhesion molecules, chemotactic factors, antioxidant enzymes and vasoactive substances [Griendling et al. 2000]. This regulation occurs at various levels. Redox sensitivity of some genes can lead to activation of their transcription by ROS-dependent signalling pathways [Wung et al. 1999]. ROS can also modify the activity of specific transcription factors, modu-lating their turnover, expression, translocation or affinity to their cognate DNA-binding sites. This has been shown in systemic vascular cells in par-ticular for the nuclear factor (NF)-κB [Morgan
and Liu, 2011] and the activator protein-1 (AP-1) [Barchowsky et al. 1995; Remacle et al. 1995], but also for the hypoxia-inducible factor (HIF)-1, known to play an important role in hypoxia-induced PH [Huang et al. 1996].
Most of the redox-sensitive genes identified so far are responsive to externally applied oxidant stress, but some of them have also been demon-strated to be downstream of an endogenous source of ROS, such as genes for angiotensin II, PDGF or tumor necrosis factor-α (TNF-α) [Griendling et al. 2000], or genes for many other inflammatory cytokines and adhesion molecules [Paravicini and Touyz, 2006]. Since all of these molecules play an important role in PH development and persistence, these data suggest that regulation of gene expression by oxidative stress may be important in pulmonary vascular cells.
Important role of ROS in the pathogenesis of PHSources of ROS in PH. Oxidative stress in the pulmonary vasculature has been evidenced in many PH animal models. In chronic hypoxia-induced PH, an increase in ROS production has been shown in pulmonary arteries [Fresquet et al. 2006; Hoshikawa et al. 2001; Jankov et al. 2008; Liu et al. 2006]. In addition, many of the patho-logical changes in the pulmonary vasculature observed in this model can be reversed by the administration of antioxidants [Elmedal et al. 2004; Hoshikawa et al. 2001; Lai et al. 1998]. In the rat model of PH induced by monocrotaline (an alkaloid extracted from the plant Crotalaria spectabilis that, once metabolized in the liver, induces pulmonary endothelial cell necrosis and leads to severe PH), elevated levels of superoxide have also been reported in pulmonary arteries [Kamezaki et al. 2008; Mathew et al. 2002]. In this model as well, antioxidant therapies attenuate PH development and decrease right ventricular hypertrophy [Kamezaki et al. 2008; Redout et al. 2010]. In addition, elevated superoxide levels have been reported in other PH models [Brennan et al. 2003; Grobe et al. 2006]. These results are in accordance with data observed in humans where increased ROS levels associated with decreased activity of antioxidant enzymes such as Mn SOD have been reported in the lungs of PH patients [Bowers et al. 2004; Demarco et al. 2010; Spieker-mann et al. 2009]. These data therefore suggest that chronic oxidative stress is present in lungs of PH patients.

V Freund-Michel, C Guibert et al.
http://tar.sagepub.com 187
In accordance with increased ROS levels detected in the pulmonary vasculature during PH, expres-sion and/or activity of various sources of ROS is increased in pulmonary arteries. Several studies have reported an important role of NADPH oxi-dases, and in particular of several Nox subunits, in various PH animal models [Dennis et al. 2009; Grobe et al. 2006; Redout et al. 2007]. Indeed, Nox2 knockout mice fail to develop hypoxia-induced PH [Liu et al. 2006], and these mice were in particular protected against the development of endothelial dysfunction in intrapulmonary arteries [Fresquet et al. 2006]. Hypoxia-induced PH in mice was also associated with increased lung levels of Nox4 [Mittal et al. 2007; Nisbet et al. 2009]. In lungs from patients with idiopathic pulmonary arterial hypertension, expression levels of Nox4 were upregulated [Mittal et al. 2007]. In vitro, a significant increase of Nox4 expression has been observed under hypoxic conditions in human pul-monary artery adventitial fibroblasts [Li et al. 2008] and in PASMC [Mittal et al. 2007]. Nox4 expression was further increased under hypoxic conditions in pulmonary artery adventitial fibro-blasts isolated from the lungs of patients with idi-opathic pulmonary arterial hypertension compared with control donors [Li et al. 2008]. Altogether, these results therefore suggest a pre-ponderant role for NAPDH oxidase containing Nox2 and/or Nox4 in oxidative stress in PH.
Some data also provide evidence of eNOS uncou-pling in pulmonary arteries, contributing to the generation of elevated levels of ROS during PH. Data obtained in mutant models underline that local BH4 availability is crucial in maintaining pulmonary vascular homeostasis and plays a piv-otal role in the pathogenesis of PH [Khoo et al. 2005; Nandi et al. 2005]. Indeed, increased BH4 synthesis prevents PH development. Conversely, BH4 deficiency elevates pulmonary vascular tone by decreasing eNOS activity and NO bioactivity, and promotes pulmonary vascular remodelling, PH and right ventricular hypertrophy [Khoo et al. 2005; Nandi et al. 2005]. These data were obtained in genetically modified mice models, in which tissue BH4 levels are low because of reduced expression of GTPCH-1, or in which endothelial BH4 synthesis is augmented by tar-geted overexpression of GTPCH-1. However, only few studies have investigated whether altera-tions of biopterin metabolism and eNOS uncou-pling occur in pathophysiological relevant PH models: in lamb models of persistent PH of the newborn, increases in oxidative stress due to
uncoupled eNOS and elevated levels of BH2 likely contribute to impaired NO-dependent pulmonary vasodilation [Grobe et al. 2006; Konduri et al. 2007]. Other studies also provide evidence of eNOS uncoupling in pulmonary arteries, contributing to generate elevated levels of ROS during PH [Gielis et al. 2011; Jerkic et al. 2011; Konduri et al. 2007; Lakshminrusimha et al. 2007; Toporsian et al. 2010; Wunderlich et al. 2008].
Altered expression and/or activity of other ROS sources in pulmonary arteries have also been reported. For example, activity of XO is increased, leading to elevated ROS levels in vitro in cultured pulmonary vascular cells [Terada et al. 1992], and in vivo as well with increased nitrotyrosine forma-tion in pulmonary arteries [Jankov et al. 2008].
ROS and altered pulmonary vascular reactivity. Many studies have shown evidence of a role of ROS in altered pulmonary vascular reactivity observed in PH (Figure 4). Indeed, both increased reactivity to contractile agents and endothelial dysfunction can be prevented or reversed by anti-oxidant treatments [Elmedal et al. 2004; Lai et al. 1998; Liu et al. 2006]. An important role for NADPH oxidase-derived ROS has been evi-denced in some studies. Indeed, hypoxia-induced amplification of the responses of pulmonary arteries to various vasoconstrictors is decreased in Nox2 knockout mice [Liu et al. 2006]. As demon-strated by our group, these Nox2 knockout mice are also protected against development of endo-thelial dysfunction in intrapulmonary arteries [Fresquet et al. 2006]. Therefore, these data show a role for Nox2-derived ROS in exacerbated vaso-constriction and endothelial dysfunction induced by chronic hypoxia in pulmonary arteries. Nox4 also plays a role in hypoxia-induced contraction of mice [Rathore et al. 2008] or of bovine pulmo-nary arteries [Ahmad et al. 2010]. Indeed, Nox4 is activated by hypoxia and increases intracellular ROS and calcium levels to participate to PASMC contraction [Rathore et al. 2008].
Data obtained in our group reveal an endothelial dysfunction occurring within the intrapulmonary arteries of hypoxic mice, despite marked upregu-lation of eNOS expression (Figure 5). Moreover, these data also show that, even though hypoxic Nox2 knockout mice did not develop endothelial dysfunction, endothelium-dependent relaxation to acetylcholine was not improved in hypoxic wild-type mice in the presence of ROS scavengers

Therapeutic Advances in Respiratory Disease 7 (3)
188 http://tar.sagepub.com
such as cell-permeant SOD or catalase [Fresquet et al. 2006] (Figure 5). These results therefore suggest that impairment of endothelial NO-dependent relaxation in pulmonary arteries from hypoxic mice is not due to inactivation of NO by ROS. This is further supported by the finding that β2-adrenergic receptor-induced relaxation is not altered in intrapulmonary arteries from hypoxic mice, even if this relaxation, similarly to the one induced by acetylcholine, is mediated by endothe-lial NO [Leblais et al. 2008] (Figure 5). Our data even show that relaxation induced through activa-tion of β2-adrenergic receptors or through the exogenous NO donor sodium nitroprusside may rather be increased in intrapulmonary arteries from hypoxic mice [Leblais et al. 2008] (Figure 5). Altogether, these data indicate that NADPH-oxidase derived ROS may act as a trigger rather than a direct mediator of endothelial dysfunction
in intrapulmonary arteries during hypoxic PH. Moreover, since activation of pulmonary β2-adrenergic and muscarinic receptors both lead to eNOS activation, but through activation of differ-ent signalling pathways [Banquet et al. 2011; Leblais et al. 2008], mechanisms underlying eNOS activation might therefore be key deter-minants of pulmonary endothelial dysfunction occurring in PH.
In addition to endothelial dysfunction, pulmo-nary arteries often exhibit hyperreactivity to con-tractile agents in PH. Isoprostanes (products from ROS-mediated peroxidation of fatty acids) may participate in these vasomotor alterations. Indeed, as recently demonstrated by our group, release of isoprostanes by pulmonary arteries is increased during chronic hypoxia [Delannoy et al. 2010]. Moreover, pulmonary arterial hyperreactivity to
Figure 5. ROS are triggers rather than direct mediators of endothelial dysfunction in hypoxic pulmonary hypertension.(A) Effect of chronic hypoxia on endothelial nitric oxide synthase (eNOS) expression in lungs from mice exposed to normoxia (NX) or 21 days of hypoxia (HX 21d). Results are expressed as mean ± SEM of quantified data normalised to β-actin and blots are representative of n = 5 independent experiments. ***p < 0.001 compared with NX. These results show that eNOS expression is significantly increased in mice total lungs after chronic hypoxia. (B) Relaxant responses to acetylcholine (ACh, 10 µM), to the exogenous nitric oxide donor sodium nitroprusside (SNP, 1 µM) or to the β2-adernergic receptor agonist procaterol (Proc, 0.1 µM) in intrapulmonary arteries from mice exposed to normoxia (NX, white bars) or 21 days of hypoxia (HX 21d, black bars). Relaxant responses to ACh were also evaluated in the presence of ROS scavengers: either catalase (Cat, 250 U/ml) or the cell-permeant superoxide dismutase (SOD, 200 U/ml). Results are expressed as mean ± SEM of n = 5–12 independent experiments. NS: not significant, **p < 0.01 and ****p < 0.001. These results show that endothelium-dependent relaxation to acetylcholine is altered by chronic hypoxia, but this effect is not modified in the presence of ROS scavengers such as catalase or SOD. Conversely, endothelium-dependent relaxation to procaterol is rather increased by chronic hypoxia, and so is the relaxation induced by the exogenous NO donor SNP. Altogether, these results show that, although eNOS expression is increased after chronic hypoxia, acetylcholine-induced relaxation is impaired in intrapulmonary arteries. This impairment is not due to NO inactivation by ROS. Moreover, such impairment is not observed after chronic hypoxia when NO-dependent relaxation of intrapulmonary arteries is induced by other stimuli through activation of other signalling pathways. Mechanisms underlying eNOS activation may therefore be of major importance in pulmonary endothelial dysfunction induced by chronic hypoxia.

V Freund-Michel, C Guibert et al.
http://tar.sagepub.com 189
phenylephrine in hypoxic mice is decreased by catalase and by TP receptor antagonists. These results therefore show that ROS contribute to pulmonary arterial hyperreactivity to contractile agents in PH via isoprostane production and fur-ther activation of TP receptors [Delannoy et al. 2010]. Such mechanism has also been recently observed in newborn piglets with chronic hypoxia-induced PH [Fike et al. 2011].
ROS and pulmonary vascular remodelling. Stud-ies have evidenced a role of ROS in pulmonary vascular remodelling observed in PH, with in par-ticular an important role of NADPH oxidase-derived ROS (Figure 4). Indeed, Nox2 knockout mice are protected against development of vascu-lar remodelling in chronic hypoxia-induced PH [Liu et al. 2006], and Nox4 participates in hypoxia-induced pulmonary vascular remodelling [Mittal et al. 2007]. ROS generation from other sources may also contribute to pulmonary vascu-lar remodelling. For example, the increased pul-monary vascular remodelling in a mouse model of familial PH is associated with eNOS-dependent ROS production [Jerkic et al. 2011].
Mechanisms activated by ROS and participating in pulmonary vascular remodelling are various and involve all pulmonary vascular cell types [Frazziano et al. 2012]. Medial thickening observed in pulmonary arterial remodelling is characterized in particular by increased proliferation and decreased apoptosis of PASMCs [Hassoun et al. 2009]. PASMC proliferation can be directly stim-ulated by ROS such as superoxide and hydrogen peroxide [Wedgwood et al. 2001]. Conversely, antioxidants such as superoxide dismutase or cat-alase can inhibit proliferation of PASMC, and even induce their apoptosis in the long term [Wedgwood and Black, 2003, 2004; Wedgwood et al. 2001]. In addition, ROS contribute to PASMC proliferation triggered by hypoxia, by directly involving activation of Nox4 [Mittal et al. 2007]. Hypoxia-induced ROS generation in PASMC also contributes to increased expression and activ-ity of the hypoxia-inducible factors (HIF)-1α and HIF-2α, involved in PASMC proliferation [Diebold et al. 2010a, 2010c]. Interestingly, acti-vation of HIF-1α by hypoxia in PASMC increases ROS production in these cells through activation of Nox4 [Diebold et al. 2010c] and Rac-1 tran-scription [Diebold et al. 2010b; Patil et al. 2004]. These data therefore suggest the exist-ence of a positive feedback loop that may con-tribute to increase PASMC proliferation in
hypoxic conditions. This positive feedback loop has also been demonstrated for HIF-2α in PASMC [Diebold et al. 2010a]. Finally, ROS-induced activation of HIF-1α in PASMC also increases expression of several factors, such as FGF-2, that can exert an autocrine proliferative effect and contribute to increased PASMC prolif-eration in hypoxic conditions [Black et al. 2008]. Moreover, hypoxia-induced PASMC prolifera-tion also involves increased mitochondrial ROS production with opening of KATP channels and mitochondrial depolarization [Hu et al. 2010]. Finally, ROS can activate other transcription fac-tors, such as for example the redox-sensitive transcription factor early growth response-1 (Egr-1), involved in hypoxia-induced PASMC proliferation [Hartney et al. 2011]. ROS also con-tribute to PASMC proliferation induced by sev-eral vasoactive factors. For example, endothelin-1 stimulates PASMC proliferation via NADPH oxidase-catalysed ROS production [Wedgwood et al. 2001]. Activation of NADPH oxidase by serotonin stimulates the proliferation of bovine PASMC via a pathway involving the MAP kinases ERK1/2 [Lee et al. 1999, 2001]. Several growth factors have also been shown to activate NADPH oxidase in vascular smooth muscle cells, includ-ing angiotensin II [Griendling et al. 1994] and PDGF [Marumo et al. 1997]. In PASMCs, the transforming growth factor-β1 (TGF-β1) acti-vates Nox4 and increases ROS production that contributes to TGF-β1-induced cell proliferation [Ismail et al. 2009; Sturrock et al. 2006]. TGF-β1-induced ROS production in PASMC also contrib-utes to upregulated expression of other growth factors, such as the vascular endothelium growth factor (VEGF) [Mata-Greenwood et al. 2005]. These results therefore suggest that ROS-mediated signalling pathways contribute to increase PASMC proliferation stimulated by various contractile and growth factors whose expression is increased in PH. As described recently, iron may play an important role in these ROS-dependent signalling pathways for PASMC growth [Wong et al. 2012].
ROS may also be involved in pulmonary vascular remodelling through participating to the excessive proliferation of endothelial cells. Indeed, ROS directly stimulates PAEC proliferation [Browning et al. 2012; Milovanova et al. 2004, 2006], and triggers angiogenesis [Nijmeh et al. 2010]. Conversely, antioxidants can inhibit PAEC prolif-eration [Han et al. 2010; Milovanova et al. 2006]. ROS may also contribute to PAEC survival induced by other factors, such as the endogenous

Therapeutic Advances in Respiratory Disease 7 (3)
190 http://tar.sagepub.com
cytochrome P-450 product 20-hydroxyeicosa-tetraenoic acid (20-HETE) [Dhanasekaran et al. 2009; Medhora et al. 2008]. Finally, oxidative stress can also activate HIF-1α in PAEC and therefore increase expression of factors such as PDGF that participate in pulmonary vascular remodelling [Mermis et al. 2011].
ROS have been shown to participate to many sig-nalling pathways in adventitial fibroblasts from various origins [Haurani and Pagano, 2007]. The role of the adventitial layer in vascular inflamma-tion and remodelling is now largely studied, in particular in PH [Stenmark et al. 2012]. ROS stimulate proliferation and inhibit apoptosis of pulmonary artery adventitial fibroblasts [Li et al. 2008], and may therefore contribute to adventi-tial remodelling observed in PH [Stenmark et al. 2012].
Finally, modulation of the extracellular matrix by matrix metalloproteinases (MMPs) is also an important component in vascular remodelling of the pulmonary vasculature [Chelladurai et al. 2012]. MMPs are a family of enzymes capable of degrading components of the extracellular matrix, and their activity is tightly regulated at transcrip-tional and protein synthesis levels. MMP-2 and MMP-9 expression and activity are increased in PH [Cantini-Salignac et al. 2006; Frisdal et al. 2001; Lepetit et al. 2005; Umar et al. 2007], and MMP-9 overexpression in transgenic mice aug-ments PH [George and D’Armiento, 2011]. NAPDH oxidase-dependent generation of ROS in PAEC leads to MMP activation [Cook-Mills, 2006]. Moreover, a role for ROS has recently been established in right ventricular failure observed in PH, through induction of MMP activity [Qipshidze et al. 2011]. These results therefore suggest that redox-sensitive mechanisms occur in the pulmonary vasculature for MMPs, and may contribute to pulmonary arterial remodelling.
ROS and pulmonary vascular inflammation. Inflammation is a common feature in remodelling vessels in PH, characterized by an accumulation of perivascular inflammatory cells, including macrophages, dendritic cells, T and B lympho-cytes, and mast cells, together with elevated circu-lating levels of various cytokines and chemokines [Price et al. 2012]. In the lung, ROS activate redox-sensitive transcription factors such as AP-1, HIF-1 of NF-κB to initiate inflammatory responses [Lee and Yang, 2012; Park et al. 2009]. For example, in response to oxidative stress, lung
cells release pro-inflammatory cytokines such as interleukin-1β (IL-1β), IL-6, IL-8 and TNF-α [Rahman and MacNee, 2000]. Moreover, expres-sion of many inflammatory target proteins trig-gered by pro-inflammatory cytokines such as IL-1β or TNF-α occurs through increased ROS production [Lee et al. 2011; Phalitakul et al. 2011]. Increased ROS levels in the lung also augment vascular leak by stimulating endothelial permeability, thus exacerbating inflammation [Pearse et al. 2003; Usatyuk et al. 2003]. ROS also participate in hypoxia-induced pulmonary vascular leak [Wojciak-Stothard et al. 2005]. This effect involves increased expression and activation of the transient receptor potential melastatin (TRPM)2 [Frey et al. 2009; Hecquet et al. 2008], as well as increased VEGF expres-sion, triggered by ROS-induced HIF-1α activa-tion [Irwin et al. 2009; Lee et al. 2006]. Finally, a direct role of pro-inflammatory cytokines such as IL-1β or TNF-α has been recently shown in endothelial dysfunction, involving activation of NADPH oxidase and ROS production in endo-thelial cells [Barbieri et al. 2011]. These results therefore suggest that oxidative stress, by regu-lating inflammatory signalling pathways and expression of inflammatory targets in the lung, may participate in vascular inflammation in PH (Figure 4).
Future directions and conclusionsOver recent years, a large body of evidence has shown an increased oxidative stress in PH, with ROS playing multiple roles in the pathogenesis of this disease. Current PH treatments, mainly designed to target altered pulmonary arterial reactivity, manage to slow the progression of the disease but do not afford a cure. In this context, targeting ROS seems to be an interesting thera-peutic approach. Indeed, ROS participate in pul-monary vascular altered reactivity, but are also involved in pulmonary vascular inflammation and remodelling, two other major PH features. Several therapeutic strategies have therefore been devel-oped to reduce ROS levels in PH [Frazziano et al. 2012; Tabima et al. 2012].
A first strategy developed is to use ROS scaven-gers. For example, catalase and SOD mimetics decrease ROS levels and exert preventive effects in experimental PH [Archer et al. 2010; Delannoy et al. 2010]. In addition, use of recombinant human SOD [Farrow et al. 2008] and gene trans-fer of extracellular SOD [Kamezaki et al. 2008;

V Freund-Michel, C Guibert et al.
http://tar.sagepub.com 191
Nozik-Grayck et al. 2008] have shown beneficial effects in PH animal models.
Another strategy is the disruption of sources of ROS in the pulmonary vasculature. Use of several pharmacological inhibitors have shown beneficial effects in experimental PH, such as inhibition of uncoupled eNOS with L-NAME [Wunderlich et al. 2008], or inhibition of XO with allopuri-nol [Jankov et al. 2008]. Therapies targeting the altered mitochondrial metabolism are also investi-gated and constitute a promising approach, as shown by dichloroacetate, an inhibitor of mito-chondrial pyruvate dehydrogenase kinase that dis-plays curative effects in PH animal models [Bonnet et al. 2006; McMurtry et al. 2004]. Another approach consists of targeting the fatty acid oxi-dation pathway, through activation of the peroxi-some proliferator-activated receptor (PPAR)γ [Rabinovitch, 2010]. The PPARγ agonist rosigli-tazone attenuates hypoxia-induced PH in the rat [Kim et al. 2010], through regulation of NF-κB and HIF-1α activity [Kang et al. 2011], and lead-ing in particular to decreased Nox4 expression and decreased ROS production [Lu et al. 2010]. These results therefore suggest PPARγ as a poten-tial PH therapeutic target. However, adverse cardiovascular and cancer events detected in diabetic patients for rosiglitazone and pioglita-zone, respectively, emphasize the need for caution before employing these agents in PH patients [Green et al. 2011].
Finally, particular attention is given to NADPH oxidases. Indeed, Nox enzymes are involved in many pulmonary vascular mechanisms, and recent findings suggest their pivotal role in PH. NADPH oxidase inhibitors prevent and reverse PH in ani-mal models [Dennis et al. 2009; Liu et al. 2006]. Further evidence comes from genetically modi-fied mice lacking NAPDH oxidase and therefore protected from experimental PH [Fresquet et al. 2006; Lee et al. 2006]. Development of specific Nox inhibitors designed to modulate Nox iso-forms in pulmonary vascular cells may therefore be of interest in PH patients [Frazziano et al. 2012].
In conclusion, many studies have shown enhanced oxidative stress associated with PH, and evi-denced an important role of ROS in various pathophysiological aspects of this disease. New therapeutic approaches in PH targeting ROS through blockade of ROS-dependent signalling pathways, or through disruption of sources of
ROS in the pulmonary vasculature, targeting in particular Nox enzymes, remain therefore promising.
FundingThis research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Conflict of interest statementThe authors declare no conflicts of interest in preparing this article.
ReferencesAhmad, M., Kelly, M., Zhao, X., Kandhi, S. and Wolin, M. (2010) Roles for Nox4 in the contractile response of bovine pulmonary arteries to hypoxia. Am J Physiol Heart Circ Physiol 298: H1879-H1888.
Alderton, W., Cooper, C. and Knowles, R. (2001) Nitric oxide synthases: structure, function and inhibition. Biochem J 357: 593-615.
Archer, S., Marsboom, G., Kim, G., Zhang, H., Toth, P., Svensson, E. et al. (2010) Epigenetic attenuation of mitochondrial superoxide dismutase 2 in pulmonary arterial hypertension: a basis for excessive cell proliferation and a new therapeutic target. Circulation 121: 2661-2671.
Archer, S. and Michelakis, E. (2009) Phosphodiesterase type 5 inhibitors for pulmonary arterial hypertension. N Engl J Med 361: 1864-1871.
Bailey, S., Mitra, S., Flavahan, S. and Flavahan, N. (2005) Reactive oxygen species from smooth muscle mitochondria initiate cold-induced constriction of cutaneous arteries. Am J Physiol Heart Circ Physiol 289: H243-H250.
Banquet, S., Delannoy, E., Agouni, A., Dessy, C., Lacomme, S., Hubert, F. et al. (2011) Role of G(i/o)-Src kinase-PI3K/Akt pathway and caveolin-1 in beta(2)-adrenoceptor coupling to endothelial NO synthase in mouse pulmonary artery. Cell Signal 23: 1136-1143.
Barbieri, S., Zacchi, E., Amadio, P., Gianellini, S., Mussoni, L., Weksler, B. et al. (2011) Cytokines present in smokers’ serum interact with smoke components to enhance endothelial dysfunction. Cardiovasc Res 90: 475-483.
Barchowsky, A., Munro, S., Morana, S., Vincenti, M. and Treadwell, M. (1995) Oxidant-sensitive and phosphorylation-dependent activation of NF-kappa B and AP-1 in endothelial cells. Am J Physiol 269: L829-L836.

Therapeutic Advances in Respiratory Disease 7 (3)
192 http://tar.sagepub.com
Bayraktutan, U., Blayney, L. and Shah, A. (2000) Molecular characterization and localization of the NAD(P)H oxidase components gp91-phox and p22-phox in endothelial cells. Arterioscler Thromb Vasc Biol 20: 1903-1911.
Belik, J., Jerkic, M., McIntyre, B., Pan, J., Leen, J., Yu, L. et al. (2009) Age-dependent endothelial nitric oxide synthase uncoupling in pulmonary arteries of endoglin heterozygous mice. Am J Physiol Lung Cell Mol Physiol 297: L1170-L1178.
Bendall, J., Alp, N., Warrick, N., Cai, S., Adlam, D., Rockett, K. et al. (2005) Stoichiometric relationships between endothelial tetrahydrobiopterin, endothelial NO synthase (eNOS) activity, and eNOS coupling in vivo: insights from transgenic mice with endothelial-targeted GTP cyclohydrolase 1 and eNOS overexpression. Circ Res 97: 864-871.
Billaud, M., Marthan, R., Savineau, J. and Guibert, C. (2009) Vascular smooth muscle modulates endothelial control of vasoreactivity via reactive oxygen species production through myoendothelial communications. PLoS One 4(7): e6432.
Birukov, K. (2009) Cyclic stretch, reactive oxygen species, and vascular remodeling. Antioxid Redox Signal 11: 1651-1667.
Black, S., DeVol, J. and Wedgwood, S. (2008) Regulation of fibroblast growth factor-2 expression in pulmonary arterial smooth muscle cells involves increased reactive oxygen species generation. Am J Physiol Cell Physiol 294: C345-C354.
Bokoch, G. and Knaus, U. (2003) NADPH oxidases: not just for leukocytes anymore! Trends Biochem Sci 28: 502-508.
Bonnet, S. and Archer, S. (2007) Potassium channel diversity in the pulmonary arteries and pulmonary veins: implications for regulation of the pulmonary vasculature in health and during pulmonary hypertension. Pharmacol Ther 115: 56-69.
Bonnet, S., Michelakis, E., Porter, C., Andrade-Navarro, M., Thebaud, B., Bonnet, S. et al. (2006) An abnormal mitochondrial-hypoxia inducible factor-1alpha-Kv channel pathway disrupts oxygen sensing and triggers pulmonary arterial hypertension in fawn hooded rats: similarities to human pulmonary arterial hypertension. Circulation 113: 2630-2641.
Borges, F., Fernandes, E. and Roleira, F. (2002) Progress towards the discovery of xanthine oxidase inhibitors. Curr Med Chem 9: 195-217.
Bowers, R., Cool, C., Murphy, R., Tuder, R., Hopken, M., Flores, S. et al. (2004) Oxidative stress in severe pulmonary hypertension. Am J Respir Crit Care Med 169: 764-769.
Brandes, R. and Kreuzer, J. (2005) Vascular NADPH oxidases: molecular mechanisms of activation. Cardiovasc Res 65: 16-27.
Brandes, R., Weissmann, N. and Schroder, K. (2010) NADPH oxidases in cardiovascular disease. Free Radic Biol Med 49: 687-706.
Brennan, L., Steinhorn, R., Wedgwood, S., Mata-Greenwood, E., Roark, E., Russell, J. et al. (2003) Increased superoxide generation is associated with pulmonary hypertension in fetal lambs: a role for NADPH oxidase. Circ Res 92: 683-691.
Broughton, B., Jernigan, N., Norton, C., Walker, B. and Resta, T. (2010) Chronic hypoxia augments depolarization-induced Ca2+ sensitization in pulmonary vascular smooth muscle through superoxide-dependent stimulation of RhoA. Am J Physiol Lung Cell Mol Physiol 298: L232-L242.
Browning, E., Chatterjee, S. and Fisher, A. (2012) Stop the flow: a paradigm for cell signaling mediated by reactive oxygen species in the pulmonary endothelium. Annu Rev Physiol 74: 403-424.
Cai, H. (2005) Hydrogen peroxide regulation of endothelial function: origins, mechanisms, and consequences. Cardiovasc Res 68: 26-36.
Cantini-Salignac, C., Lartaud, I., Schrijen, F., Atkinson, J. and Chabot, F. (2006) Metalloproteinase-9 in circulating monocytes in pulmonary hypertension. Fundam Clin Pharmacol 20: 405-410.
Chakraborti, S. and Michael, J. (1993) Role of protein kinase C in oxidant-mediated activation of phospholipase A2 in rabbit pulmonary arterial smooth muscle cells. Mol Cell Biochem 122: 9-15.
Chatterjee, S., Feinstein, S., Dodia, C., Sorokina, E., Lien, Y., Nguyen, S. et al. (2011) Peroxiredoxin 6 phosphorylation and subsequent phospholipase A2 activity are required for agonist-mediated activation of NADPH oxidase in mouse pulmonary microvascular endothelium and alveolar macrophages. J Biol Chem 286: 11696-11706.
Chelladurai, P., Seeger, W. and Pullamsetti, S. (2012) Matrix metalloproteinases and their inhibitors in pulmonary hypertension. Eur Respir J 40: 766-782.
Chen, C., Lin, C., Druhan, L., Wang, T., Chen, Y. and Zweier, J. (2011) Superoxide induces endothelial nitric-oxide synthase protein thiyl radical formation, a novel mechanism regulating eNOS function and coupling. J Biol Chem 286: 29098-29107.
Chi, A., Waypa, G., Mungai, P. and Schumacker, P. (2010) Prolonged hypoxia increases ROS signaling and RhoA activation in pulmonary artery smooth muscle and endothelial cells. Antioxid Redox Signal 12: 603-610.

V Freund-Michel, C Guibert et al.
http://tar.sagepub.com 193
Coffer, P., Jin, J. and Woodgett, J. (1998) Protein kinase B (c-Akt): a multifunctional mediator of phosphatidylinositol 3-kinase activation. Biochem J 335: 1-13.
Cogolludo, A., Frazziano, G., Cobeno, L., Moreno, L., Lodi, F., Villamor, E. et al. (2006) Role of reactive oxygen species in Kv channel inhibition and vasoconstriction induced by TP receptor activation in rat pulmonary arteries. Ann N Y Acad Sci 1091: 41-51.
Connolly, M. and Aaronson, P. (2011) Key role of the RhoA/Rho kinase system in pulmonary hypertension. Pulm Pharmacol Ther 24: 1-14.
Cook-Mills, J. (2006) Hydrogen peroxide activation of endothelial cell-associated MMPs during VCAM-1-dependent leukocyte migration. Cell Mol Biol (Noisy-le-grand) 52(4): 8-16.
Courboulin, A., Paulin, R., Giguere, N., Saksouk, N., Perreault, T., Meloche, J. et al. (2011) Role for miR-204 in human pulmonary arterial hypertension. J Exp Med 208: 535-548
Crabtree, M. and Channon, K. (2011) Synthesis and recycling of tetrahydrobiopterin in endothelial function and vascular disease. Nitric Oxide 25(2): 81-88.
Delannoy, E., Courtois, A., Freund-Michel, V., Leblais, V., Marthan, R. and Muller, B. (2010) Hypoxia-induced hyperreactivity of pulmonary arteries: role of cyclooxygenase-2, isoprostanes, and thromboxane receptors. Cardiovasc Res 85: 582-592.
Demarco, V., Whaley-Connell, A., Sowers, J., Habibi, J. and Dellsperger, K. (2010) Contribution of oxidative stress to pulmonary arterial hypertension. World J Cardiol 2: 316-324.
Dempsey, E., Cool, C. and Littler, C. (2007) Lung disease and PKCs. Pharmacol Res 55: 545-559.
Dennis, K., Aschner, J., Milatovic, D., Schmidt, J., Aschner, M., Kaplowitz, M. et al. (2009) NADPH oxidases and reactive oxygen species at different stages of chronic hypoxia-induced pulmonary hypertension in newborn piglets. Am J Physiol Lung Cell Mol Physiol 297: L596-L607.
Dhanasekaran, A., Bodiga, S., Gruenloh, S., Gao, Y., Dunn, L., Falck, J. et al. (2009) 20-HETE increases survival and decreases apoptosis in pulmonary arteries and pulmonary artery endothelial cells. Am J Physiol Heart Circ Physiol 296: H777-H786.
Diebold, I., Flugel, D., Becht, S., Belaiba, R., Bonello, S., Hess, J. et al. (2010a) The hypoxia-inducible factor-2alpha is stabilized by oxidative stress involving NOX4. Antioxid Redox Signal 13: 425-436.
Diebold, I., Petry, A., Djordjevic, T., Belaiba, R., Fineman, J., Black, S. et al. (2010b) Reciprocal regulation of Rac1 and PAK-1 by HIF-1alpha: a positive-feedback loop promoting pulmonary
vascular remodeling. Antioxid Redox Signal 13: 399-412.
Diebold, I., Petry, A., Hess, J. and Gorlach, A. (2010c) The NADPH oxidase subunit NOX4 is a new target gene of the hypoxia-inducible factor-1. Mol Biol Cell 21: 2087-2096.
Dietrich, A. and Gudermann, T. (2011) TRP channels in the cardiopulmonary vasculature. Adv Exp Med Biol 704: 781-810.
Djordjevic, T., BelAiba, R., Bonello, S., Pfeilschifter, J., Hess, J. and Gorlach, A. (2005) Human urotensin II is a novel activator of NADPH oxidase in human pulmonary artery smooth muscle cells. Arterioscler Thromb Vasc Biol 25: 519-525.
Druhan, L., Forbes, S., Pope, A., Chen, C., Zweier, J. and Cardounel, A. (2008) Regulation of eNOS-derived superoxide by endogenous methylarginines. Biochemistry 47: 7256-7263.
Dupuis, J. (2001) Endothelin-receptor antagonists in pulmonary hypertension. Lancet 358: 1113-1114.
Elmedal, B., de Dam, M., Mulvany, M. and Simonsen, U. (2004) The superoxide dismutase mimetic, tempol, blunts right ventricular hypertrophy in chronic hypoxic rats. Br J Pharmacol 141: 105-113.
Erusalimsky, J. and Moncada, S. (2007) Nitric oxide and mitochondrial signaling: from physiology to pathophysiology. Arterioscler Thromb Vasc Biol 27: 2524-2531.
Farrow, K., Lakshminrusimha, S., Reda, W., Wedgwood, S., Czech, L., Gugino, S. et al. (2008) Superoxide dismutase restores eNOS expression and function in resistance pulmonary arteries from neonatal lambs with persistent pulmonary hypertension. Am J Physiol Lung Cell Mol Physiol 295: L979-L987.
Fike, C., Aschner, J., Slaughter, J., Kaplowitz, M., Zhang, Y. and Pfister, S. (2011) Pulmonary arterial responses to reactive oxygen species are altered in newborn piglets with chronic hypoxia-induced pulmonary hypertension. Pediatr Res 70: 136-141.
Fleming, I. (2010) Molecular mechanisms underlying the activation of eNOS. Pflugers Arch 459: 793-806.
Forstermann, U. (2006) Janus-faced role of endothelial NO synthase in vascular disease: uncoupling of oxygen reduction from NO synthesis and its pharmacological reversal. Biol Chem 387: 1521-1533.
Frazziano, G., Champion, H. and Pagano, P. (2012) NADPH oxidase-derived ROS and the regulation of pulmonary vessel tone. Am J Physiol Heart Circ Physiol 302: H2166-H2177.
Fresquet, F., Pourageaud, F., Leblais, V., Brandes, R., Savineau, J., Marthan, R. et al. (2006) Role of

Therapeutic Advances in Respiratory Disease 7 (3)
194 http://tar.sagepub.com
reactive oxygen species and gp91phox in endothelial dysfunction of pulmonary arteries induced by chronic hypoxia. Br J Pharmacol 148: 714-723.
Frey, R., Ushio-Fukai, M. and Malik, A. (2009) NADPH oxidase-dependent signaling in endothelial cells: role in physiology and pathophysiology. Antioxid Redox Signal 11: 791-810.
Frisdal, E., Gest, V., Vieillard-Baron, A., Levame, M., Lepetit, H., Eddahibi, S. et al. (2001) Gelatinase expression in pulmonary arteries during experimental pulmonary hypertension. Eur Respir J 18: 838-845.
Frumkin, L. (2012) The pharmacological treatment of pulmonary arterial hypertension. Pharmacol Rev 64: 583-620.
Fuchs, B., Sommer, N., Dietrich, A., Schermuly, R., Ghofrani, H., Grimminger, F. et al. (2010) Redox signaling and reactive oxygen species in hypoxic pulmonary vasoconstriction. Respir Physiol Neurobiol 174: 282-291.
Galie, N., Olschewski, H., Oudiz, R., Torres, F., Frost, A., Ghofrani, H. et al. (2008a) Ambrisentan for the treatment of pulmonary arterial hypertension: results of the ambrisentan in pulmonary arterial hypertension, randomized, double-blind, placebo-controlled, multicenter, efficacy (ARIES) study 1 and 2. Circulation 117: 3010-3019.
Galie, N., Rubin, L., Hoeper, M., Jansa, P., Al-Hiti, H., Meyer, G. et al. (2008b) Treatment of patients with mildly symptomatic pulmonary arterial hypertension with bosentan (EARLY study): a double-blind, randomised controlled trial. Lancet 371: 2093-2100.
Galie, N., Seeger, W., Naeije, R., Simonneau, G. and Rubin, L. (2004) Comparative analysis of clinical trials and evidence-based treatment algorithm in pulmonary arterial hypertension. J Am Coll Cardiol 43(12 Suppl. S): 81S-88S.
George, J. and D’Armiento, J. (2011) Transgenic expression of human matrix metalloproteinase-9 augments monocrotaline-induced pulmonary arterial hypertension in mice. J Hypertens 29: 299-308.
Gielis, J., Lin, J., Wingler, K., Van Schil, P., Schmidt, H. and Moens, A. (2011) Pathogenetic role of eNOS uncoupling in cardiopulmonary disorders. Free Radic Biol Med 50: 765-776.
Gomberg-Maitland, M. and Olschewski, H. (2008) Prostacyclin therapies for the treatment of pulmonary arterial hypertension. Eur Respir J 31: 891-901.
Gongora, M., Qin, Z., Laude, K., Kim, H., McCann, L., Folz, J. et al. (2006) Role of extracellular superoxide dismutase in hypertension. Hypertension 48: 473-481.
Green, D., Sutliff, R. and Hart, C. (2011) Is peroxisome proliferator-activated receptor gamma (PPARgamma) a therapeutic target for the treatment of pulmonary hypertension? Pulm Circ 1: 33-47.
Griendling, K., Minieri, C., Ollerenshaw, J. and Alexander, R. (1994) Angiotensin II stimulates NADH and NADPH oxidase activity in cultured vascular smooth muscle cells. Circ Res 74: 1141-1148.
Griendling, K., Sorescu, D., Lassegue, B. and Ushio-Fukai, M. (2000) Modulation of protein kinase activity and gene expression by reactive oxygen species and their role in vascular physiology and pathophysiology. Arterioscler Thromb Vasc Biol 20: 2175-2183.
Griffith, B., Pendyala, S., Hecker, L., Lee, P., Natarajan, V. and Thannickal, V. (2009) NOX enzymes and pulmonary disease. Antioxid Redox Signal 11: 2505-2516.
Grobe, A., Wells, S., Benavidez, E., Oishi, P., Azakie, A., Fineman, J. et al. (2006) Increased oxidative stress in lambs with increased pulmonary blood flow and pulmonary hypertension: role of NADPH oxidase and endothelial NO synthase. Am J Physiol Lung Cell Mol Physiol 290: L1069-L1077.
Guibert, C., Ducret, T. and Savineau, J. (2011) Expression and physiological roles of TRP channels in smooth muscle cells. Adv Exp Med Biol 704: 687-706.
Guibert, C., Marthan, R. and Savineau, J. (2007) Modulation of ion channels in pulmonary arterial hypertension. Curr Pharm Des 13: 2443-2455.
Guzik, T. and Harrison, D. (2006) Vascular NADPH oxidases as drug targets for novel antioxidant strategies. Drug Discov Today 11: 524-533.
Han, D., Canali, R., Rettori, D. and Kaplowitz, N. (2003) Effect of glutathione depletion on sites and topology of superoxide and hydrogen peroxide production in mitochondria. Mol Pharmacol 64: 1136-1144.
Han, Y., Moon, H., You, B., Kim, S., Kim, S. and Park, W. (2010) Pyrogallol-induced endothelial cell death is related to GSH depletion rather than ROS level changes. Oncol Rep 23: 287-292.
Hanson, K., Ziegler, J., Rybalkin, S., Miller, J., Abman, S. and Clarke, W. (1998) Chronic pulmonary hypertension increases fetal lung cGMP phosphodiesterase activity. Am J Physiol 275: L931-L941.
Harrison, R. (2002) Structure and function of xanthine oxidoreductase: where are we now? Free Radic Biol Med 33: 774-797.
Hartney, T., Birari, R., Venkataraman, S., Villegas, L., Martinez, M., Black, S. et al. (2011) Xanthine oxidase-derived ROS upregulate Egr-1 via ERK1/2

V Freund-Michel, C Guibert et al.
http://tar.sagepub.com 195
in PA smooth muscle cells; model to test impact of extracellular ROS in chronic hypoxia. PLoS One 6(11): e27531.
Hassoun, P., Mouthon, L., Barbera, J., Eddahibi, S., Flores, S., Grimminger, F. et al. (2009) Inflammation, growth factors, and pulmonary vascular remodeling. J Am Coll Cardiol 54(1 Suppl.): S10-S19.
Haurani, M. and Pagano, P. (2007) Adventitial fibroblast reactive oxygen species as autocrine and paracrine mediators of remodeling: bellwether for vascular disease? Cardiovasc Res 75: 679-689.
Hecquet, C., Ahmmed, G., Vogel, S. and Malik, A. (2008) Role of TRPM2 channel in mediating H2O2-induced Ca2+ entry and endothelial hyperpermeability. Circ Res 102: 347-355.
Hoshikawa, Y., Ono, S., Suzuki, S., Tanita, T., Chida, M., Song, C. et al. (2001) Generation of oxidative stress contributes to the development of pulmonary hypertension induced by hypoxia. J Appl Physiol 90: 1299-1306.
Hu, H., Zhang, Z., Chen, C., Cai, C., Zhao, J. and Wang, X. (2010) Effects of mitochondrial potassium channel and membrane potential on hypoxic human pulmonary artery smooth muscle cells. Am J Respir Cell Mol Biol 42: 661-666.
Huang, L., Arany, Z., Livingston, D. and Bunn, H. (1996) Activation of hypoxia-inducible transcription factor depends primarily upon redox-sensitive stabilization of its alpha subunit. J Biol Chem 271: 32253-32259.
Humbert, M., Morrell, N., Archer, S., Stenmark, K., MacLean, M., Lang, I. et al. (2004) Cellular and molecular pathobiology of pulmonary arterial hypertension. J Am Coll Cardiol 43(12 Suppl. S): 13S-24S.
Huot, J., Houle, F., Marceau, F. and Landry, J. (1997) Oxidative stress-induced actin reorganization mediated by the p38 mitogen-activated protein kinase/heat shock protein 27 pathway in vascular endothelial cells. Circ Res 80: 383-392.
Huot, J., Houle, F., Rousseau, S., Deschesnes, R., Shah, G. and Landry, J. (1998) SAPK2/p38-dependent F-actin reorganization regulates early membrane blebbing during stress-induced apoptosis. J Cell Biol 143: 1361-1373.
Irwin, D., McCord, J., Nozik-Grayck, E., Beckly, G., Foreman, B., Sullivan, T. et al. (2009) A potential role for reactive oxygen species and the HIF-1alpha-VEGF pathway in hypoxia-induced pulmonary vascular leak. Free Radic Biol Med 47: 55-61.
Ismail, S., Sturrock, A., Wu, P., Cahill, B., Norman, K., Huecksteadt, T. et al. (2009) NOX4 mediates hypoxia-induced proliferation of human pulmonary artery smooth muscle cells: the role of autocrine
production of transforming growth factor-{beta}1 and insulin-like growth factor binding protein-3. Am J Physiol Lung Cell Mol Physiol 296: L489-L499.
Jagnandan, D., Church, J., Banfi, B., Stuehr, D., Marrero, M. and Fulton, D. (2007) Novel mechanism of activation of NADPH oxidase 5. calcium sensitization via phosphorylation. J Biol Chem 282: 6494-6507.
Jankov, R., Kantores, C., Pan, J. and Belik, J. (2008) Contribution of xanthine oxidase-derived superoxide to chronic hypoxic pulmonary hypertension in neonatal rats. Am J Physiol Lung Cell Mol Physiol 294: L233-L245.
Jarasch, E., Grund, C., Bruder, G., Heid, H., Keenan, T. and Franke, W. (1981) Localization of xanthine oxidase in mammary-gland epithelium and capillary endothelium. Cell 25: 67-82.
Jerkic, M., Kabir, M., Davies, A., Yu, L., McIntyre, B., Husain, N. et al. (2011) Pulmonary hypertension in adult Alk1 heterozygous mice due to oxidative stress. Cardiovasc Res 92: 375-384.
Jernigan, N., Walker, B. and Resta, T. (2008) Reactive oxygen species mediate RhoA/Rho kinase-induced Ca2+ sensitization in pulmonary vascular smooth muscle following chronic hypoxia. Am J Physiol Lung Cell Mol Physiol 295: L515-L529.
Kamezaki, F., Tasaki, H., Yamashita, K., Tsutsui, M., Koide, S., Nakata, S. et al. (2008) Gene transfer of extracellular superoxide dismutase ameliorates pulmonary hypertension in rats. Am J Respir Crit Care Med 177: 219-226.
Kang, B., Kleinhenz, J., Murphy, T. and Hart, C. (2011) The PPARgamma ligand rosiglitazone attenuates hypoxia-induced endothelin signaling in vitro and in vivo. Am J Physiol Lung Cell Mol Physiol 301: L881-L891.
Khoo, J., Zhao, L., Alp, N., Bendall, J., Nicoli, T., Rockett, K. et al. (2005) Pivotal role for endothelial tetrahydrobiopterin in pulmonary hypertension. Circulation 111: 2126-2133.
Kim, E., Lee, J., Oh, Y., Lee, Y. and Lee, S. (2010) Rosiglitazone attenuates hypoxia-induced pulmonary arterial hypertension in rats. Respirology 15: 659-668.
Kim, N. and Rubin, L. (2002) Endothelin in health and disease: endothelin receptor antagonists in the management of pulmonary artery hypertension. J Cardiovasc Pharmacol Ther 7: 9-19.
Knock, G., Snetkov, V., Shaifta, Y., Connolly, M., Drndarski, S., Noah, A. et al. (2009) Superoxide constricts rat pulmonary arteries via Rho-kinase-mediated Ca(2+) sensitization. Free Radic Biol Med 46: 633-642.

Therapeutic Advances in Respiratory Disease 7 (3)
196 http://tar.sagepub.com
Knock, G., Snetkov, V., Shaifta, Y., Drndarski, S., Ward, J. and Aaronson, P. (2008) Role of src-family kinases in hypoxic vasoconstriction of rat pulmonary artery. Cardiovasc Res 80: 453-462.
Knock, G. and Ward, J. (2011) Redox regulation of protein kinases as a modulator of vascular function. Antioxid Redox Signal 15: 1531-1547.
Konduri, G., Bakhutashvili, I., Eis, A. and Pritchard, K., Jr (2007) Oxidant stress from uncoupled nitric oxide synthase impairs vasodilation in fetal lambs with persistent pulmonary hypertension. Am J Physiol Heart Circ Physiol 292: H1812-H1820.
Lai, Y., Wu, H. and Chen, C. (1998) Antioxidants attenuate chronic hypoxic pulmonary hypertension. J Cardiovasc Pharmacol 32: 714-720.
Lakshminrusimha, S., Wiseman, D., Black, S., Russell, J., Gugino, S., Oishi, P. et al. (2007) The role of nitric oxide synthase-derived reactive oxygen species in the altered relaxation of pulmonary arteries from lambs with increased pulmonary blood flow. Am J Physiol Heart Circ Physiol 293: H1491-H1497.
Leblais, V., Delannoy, E., Fresquet, F., Begueret, H., Bellance, N., Banquet, S. et al. (2008) beta-adrenergic relaxation in pulmonary arteries: preservation of the endothelial nitric oxide-dependent beta2 component in pulmonary hypertension. Cardiovasc Res 77: 202-210.
Lee, I. and Yang, C. (2012) Role of NADPH oxidase/ROS in pro-inflammatory mediators-induced airway and pulmonary diseases. Biochem Pharmacol 84: 581-590.
Lee, J., Park, S., Lee, E., Park, C., Chung, H., Rha, S. et al. (2006) Activation of hypoxia-inducible factor-1alpha is necessary for lysophosphatidic acid-induced vascular endothelial growth factor expression. Clin Cancer Res 12: 6351-6358.
Lee, K. and Esselman, W. (2002) Inhibition of PTPs by H(2)O(2) regulates the activation of distinct MAPK pathways. Free Radic Biol Med 33: 1121-1132.
Lee, S., Simon, A., Wang, W. and Fanburg, B. (2001) H(2)O(2) signals 5-HT-induced ERK MAP kinase activation and mitogenesis of smooth muscle cells. Am J Physiol Lung Cell Mol Physiol 281: L646-L652.
Lee, S., Wang, W., Finlay, G. and Fanburg, B. (1999) Serotonin stimulates mitogen-activated protein kinase activity through the formation of superoxide anion. Am J Physiol 277: L282-L291.
Lee, Y., Lee, W. and Kim, P. (2011) Role of NADPH oxidase in interleukin-4-induced monocyte chemoattractant protein-1 expression in vascular endothelium. Inflamm Res 59: 755-765.
Lepetit, H., Eddahibi, S., Fadel, E., Frisdal, E., Munaut, C., Noel, A. et al. (2005) Smooth muscle
cell matrix metalloproteinases in idiopathic pulmonary arterial hypertension. Eur Respir J 25: 834-842.
Li, J. and Shah, A. (2004) Endothelial cell superoxide generation: regulation and relevance for cardiovascular pathophysiology. Am J Physiol Regul Integr Comp Physiol 287: R1014-R1030.
Li, S., Tabar, S., Malec, V., Eul, B., Klepetko, W., Weissmann, N. et al. (2008) NOX4 regulates ROS levels under normoxic and hypoxic conditions, triggers proliferation, and inhibits apoptosis in pulmonary artery adventitial fibroblasts. Antioxid Redox Signal 10: 1687-1698.
Liu, J., Zelko, I., Erbynn, E., Sham, J. and Folz, R. (2006) Hypoxic pulmonary hypertension: role of superoxide and NADPH oxidase (gp91phox). Am J Physiol Lung Cell Mol Physiol 290: L2-L10.
Loirand, G., Guerin, P. and Pacaud, P. (2006) Rho kinases in cardiovascular physiology and pathophysiology. Circ Res 98: 322-334.
Lu, X., Murphy, T., Nanes, M. and Hart, C. (2010) PPAR{gamma} regulates hypoxia-induced Nox4 expression in human pulmonary artery smooth muscle cells through NF-{kappa}B. Am J Physiol Lung Cell Mol Physiol 299: L559-L566.
Lyle, A. and Griendling, K. (2006) Modulation of vascular smooth muscle signaling by reactive oxygen species. Physiology (Bethesda) 21: 269-280.
Marumo, T., Schini-Kerth, V., Fisslthaler, B. and Busse, R. (1997) Platelet-derived growth factor-stimulated superoxide anion production modulates activation of transcription factor NF-kappaB and expression of monocyte chemoattractant protein 1 in human aortic smooth muscle cells. Circulation 96: 2361-2367.
Mata-Greenwood, E., Grobe, A., Kumar, S., Noskina, Y. and Black, S. (2005) Cyclic stretch increases VEGF expression in pulmonary arterial smooth muscle cells via TGF-beta1 and reactive oxygen species: a requirement for NAD(P)H oxidase. Am J Physiol Lung Cell Mol Physiol 289: L288-L289.
Mathew, R., Yuan, N., Rosenfeld, L., Gewitz, M. and Kumar, A. (2002) Effects of monocrotaline on endothelial nitric oxide synthase expression and sulfhydryl levels in rat lungs. Heart Dis 4: 152-158.
McMurtry, M., Bonnet, S., Wu, X., Dyck, J., Haromy, A., Hashimoto, K. et al. (2004) Dichloroacetate prevents and reverses pulmonary hypertension by inducing pulmonary artery smooth muscle cell apoptosis. Circ Res 95: 830-840.
Medhora, M., Chen, Y., Gruenloh, S., Harland, D., Bodiga, S., Zielonka, J. et al. (2008) 20-HETE increases superoxide production and activates NAPDH oxidase in pulmonary artery endothelial

V Freund-Michel, C Guibert et al.
http://tar.sagepub.com 197
cells. Am J Physiol Lung Cell Mol Physiol 294: L902-L911.
Meloche, S., Landry, J., Huot, J., Houle, F., Marceau, F. and Giasson, E. (2000) p38 MAP kinase pathway regulates angiotensin II-induced contraction of rat vascular smooth muscle. Am J Physiol Heart Circ Physiol 279: H741-H751.
Mermis, J., Gu, H., Xue, B., Li, F., Tawfik, O., Buch, S. et al. (2011) Hypoxia-inducible factor-1 alpha/platelet derived growth factor axis in HIV-associated pulmonary vascular remodeling. Respir Res 12: 103.
Milovanova, T., Chatterjee, S., Manevich, Y., Kotelnikova, I., Debolt, K., Madesh, M. et al. (2006) Lung endothelial cell proliferation with decreased shear stress is mediated by reactive oxygen species. Am J Physiol Cell Physiol 290: C66-C76.
Milovanova, T., Manevich, Y., Haddad, A., Chatterjee, S., Moore, J. and Fisher, A. (2004) Endothelial cell proliferation associated with abrupt reduction in shear stress is dependent on reactive oxygen species. Antioxid Redox Signal 6: 245-258.
Mittal, M., Roth, M., Konig, P., Hofmann, S., Dony, E., Goyal, P. et al. (2007) Hypoxia-dependent regulation of nonphagocytic NADPH oxidase subunit NOX4 in the pulmonary vasculature. Circ Res 101: 258-267.
Montani, D., Chaumais, M., Savale, L., Natali, D., Price, L., Jais, X. et al. (2009) Phosphodiesterase type 5 inhibitors in pulmonary arterial hypertension. Adv Ther 26: 813-825.
Montani, D., Savale, L., Natali, D., Jais, X., Herve, P., Garcia, G. et al. (2010) Long-term response to calcium-channel blockers in non-idiopathic pulmonary arterial hypertension. Eur Heart J 31: 1898-1907.
Morgan, M. and Liu, Z. (2011) Crosstalk of reactive oxygen species and NF-kappaB signaling. Cell Res 21: 103-115.
Morrell, N., Adnot, S., Archer, S., Dupuis, J., Jones, P., MacLean, M. et al. (2009) Cellular and molecular basis of pulmonary arterial hypertension. J Am Coll Cardiol 54(1 Suppl.): S20-S31.
Moudgil, R., Michelakis, E. and Archer, S. (2006) The role of k+ channels in determining pulmonary vascular tone, oxygen sensing, cell proliferation, and apoptosis: implications in hypoxic pulmonary vasoconstriction and pulmonary arterial hypertension. Microcirculation 13: 615-632.
Murphy, E. and Steenbergen, C. (2008) Mechanisms underlying acute protection from cardiac ischemia-reperfusion injury. Physiol Rev 88: 581-609.
Nandi, M., Miller, A., Stidwill, R., Jacques, T., Lam, A., Haworth, S. et al. (2005) Pulmonary
hypertension in a GTP-cyclohydrolase 1-deficient mouse. Circulation 111: 2086-2090.
Natarajan, V., Taher, M., Roehm, B., Parinandi, N., Schmid, H., Kiss, Z. et al. (1993) Activation of endothelial cell phospholipase D by hydrogen peroxide and fatty acid hydroperoxide. J Biol Chem 268: 930-937.
Nijmeh, J., Moldobaeva, A. and Wagner, E. (2010) Role of ROS in ischemia-induced lung angiogenesis. Am J Physiol Lung Cell Mol Physiol 299: L535-L541.
Nisbet, R., Graves, A., Kleinhenz, D., Rupnow, H., Reed, A., Fan, T. et al. (2009) The role of NADPH oxidase in chronic intermittent hypoxia-induced pulmonary hypertension in mice. Am J Respir Cell Mol Biol 40: 601-609.
Nishino, T., Okamoto, K., Eger, B., Pai, E. and Nishino, T. (2008) Mammalian xanthine oxidoreductase - mechanism of transition from xanthine dehydrogenase to xanthine oxidase. FEBS J 275: 3278-3289.
Noguchi, K., Hamadate, N., Matsuzaki, T., Sakanashi, M., Nakasone, J., Uchida, T. et al. (2011) Increasing dihydrobiopterin causes dysfunction of endothelial nitric oxide synthase in rats in vivo. Am J Physiol Heart Circ Physiol 301: H721-H729.
Nozik-Grayck, E., Suliman, H., Majka, S., Albietz, J., Van Rheen, Z., Roush, K. et al. (2008) Lung EC-SOD overexpression attenuates hypoxic induction of Egr-1 and chronic hypoxic pulmonary vascular remodeling. Am J Physiol Lung Cell Mol Physiol 295: L422-L430.
O’Callaghan, D., Savale, L., Montani, D., Jais, X., Sitbon, O., Simonneau, G. et al. (2011) Treatment of pulmonary arterial hypertension with targeted therapies. Nat Rev Cardiol 8: 526-538.
Oka, M., Fagan, K., Jones, P. and McMurtry, I. (2008) Therapeutic potential of RhoA/Rho kinase inhibitors in pulmonary hypertension. Br J Pharmacol 155: 444-454.
Paravicini, T. and Touyz, R. (2006) Redox signaling in hypertension. Cardiovasc Res 71: 247-258.
Parinandi, N., Kleinberg, M., Usatyuk, P., Cummings, R., Pennathur, A., Cardounel, A. et al. (2003) Hyperoxia-induced NAD(P)H oxidase activation and regulation by MAP kinases in human lung endothelial cells. Am J Physiol Lung Cell Mol Physiol 284: L26-L38.
Park, H., Kim, S. and Lee, Y. (2009) Impact of oxidative stress on lung diseases. Respirology 14: 27-38.
Partridge, C., Blumenstock, F. and Malik, A. (1992) Pulmonary microvascular endothelial cells constitutively release xanthine oxidase. Arch Biochem Biophys 294: 184-187.

Therapeutic Advances in Respiratory Disease 7 (3)
198 http://tar.sagepub.com
Patil, S., Bunderson, M., Wilham, J. and Black, S. (2004) Important role for Rac1 in regulating reactive oxygen species generation and pulmonary arterial smooth muscle cell growth. Am J Physiol Lung Cell Mol Physiol 287: L1314-L1322.
Paulin, R., Meloche, J., Jacob, M., Bisserier, M., Courboulin, A. and Bonnet, S. (2011) Dehydroepiandrosterone inhibits the Src/STAT3 constitutive activation in pulmonary arterial hypertension. Am J Physiol Heart Circ Physiol 301: H1798-H1809.
Pauvert, O., Lugnier, C., Keravis, T., Marthan, R., Rousseau, E. and Savineau, J. (2003) Effect of sildenafil on cyclic nucleotide phosphodiesterase activity, vascular tone and calcium signaling in rat pulmonary artery. Br J Pharmacol 139: 513-522.
Pearse, D., Shimoda, L., Verin, A., Bogatcheva, N., Moon, C., Ronnett, G. et al. (2003) Effect of cGMP on lung microvascular endothelial barrier dysfunction following hydrogen peroxide. Endothelium 10: 309-317.
Perez-Vizcaino, F., Cogolludo, A. and Moreno, L. (2010) Reactive oxygen species signaling in pulmonary vascular smooth muscle. Respir Physiol Neurobiol 174: 212-220.
Phalitakul, S., Okada, M., Hara, Y. and Yamawaki, H. (2011) Vaspin prevents TNF-alpha-induced intracellular adhesion molecule-1 via inhibiting reactive oxygen species-dependent NF-kappaB and PKCtheta activation in cultured rat vascular smooth muscle cells. Pharmacol Res 64: 493-500.
Pourmahram, G., Snetkov, V., Shaifta, Y., Drndarski, S., Knock, G., Aaronson, P. et al. (2008) Constriction of pulmonary artery by peroxide: role of Ca2+ release and PKC. Free Radic Biol Med 45: 1468-1476.
Price, L., Wort, S., Perros, F., Dorfmuller, P., Huertas, A., Montani, D. et al. (2012) Inflammation in pulmonary arterial hypertension. Chest 141: 210-221.
Qipshidze, N., Tyagi, N., Metreveli, N., Lominadze, D. and Tyagi, S. (2011) Autophagy mechanism of right ventricular remodeling in murine model of pulmonary artery constriction. Am J Physiol Heart Circ Physiol 302: H688-H696.
Quinn, M. and Gauss, K. (2004) Structure and regulation of the neutrophil respiratory burst oxidase: comparison with nonphagocyte oxidases. J Leukoc Biol 76: 760-781.
Rabinovitch, M. (2010) PPARgamma and the pathobiology of pulmonary arterial hypertension. Adv Exp Med Biol 661: 447-458.
Rafikov, R., Fonseca, F., Kumar, S., Pardo, D., Darragh, C., Elms, S. et al. (2011) eNOS activation and NO function: structural motifs responsible for the
posttranslational control of endothelial nitric oxide synthase activity. J Endocrinol 210: 271-284.
Rahman, I. and MacNee, W. (2000) Oxidative stress and regulation of glutathione in lung inflammation. Eur Respir J 16: 534-554.
Rathore, R., Zheng, Y., Li, X., Wang, Q., Liu, Q., Ginnan, R. et al. (2006) Mitochondrial ROS-PKCepsilon signaling axis is uniquely involved in hypoxic increase in [Ca2+]i in pulmonary artery smooth muscle cells. Biochem Biophys Res Commun 351: 784-790.
Rathore, R., Zheng, Y., Niu, C., Liu, Q., Korde, A., Ho, Y. et al. (2008) Hypoxia activates NADPH oxidase to increase [ROS]i and [Ca2+]i through the mitochondrial ROS-PKCepsilon signaling axis in pulmonary artery smooth muscle cells. Free Radic Biol Med 45: 1223-1231.
Redout, E., van der Toorn, A., Zuidwijk, M., van de Kolk, C., van Echteld, C., Musters, R. et al. (2010) Antioxidant treatment attenuates pulmonary arterial hypertension-induced heart failure. Am J Physiol Heart Circ Physiol 298: H1038-H1047.
Redout, E., Wagner, M., Zuidwijk, M., Boer, C., Musters, R., van Hardeveld, C. et al. (2007) Right-ventricular failure is associated with increased mitochondrial complex II activity and production of reactive oxygen species. Cardiovasc Res 75: 770-781.
Remacle, J., Raes, M., Toussaint, O., Renard, P. and Rao, G. (1995) Low levels of reactive oxygen species as modulators of cell function. Mutat Res 316: 103-122.
Resta, T., Broughton, B. and Jernigan, N. (2010) Reactive oxygen species and RhoA signaling in vascular smooth muscle: role in chronic hypoxia-induced pulmonary hypertension. Adv Exp Med Biol 661: 355-373.
Reyland, M. (2009) Protein kinase C isoforms: Multi-functional regulators of cell life and death. Front Biosci 14: 2386-2399.
Rhodes, C., Davidson, A., Gibbs, J., Wharton, J. and Wilkins, M. (2009) Therapeutic targets in pulmonary arterial hypertension. Pharmacol Ther 121: 69-88.
Rubbo, H., Trostchansky, A. and O’Donnell, V. (2009) Peroxynitrite-mediated lipid oxidation and nitration: mechanisms and consequences. Arch Biochem Biophys 484: 167-172.
Rubin, L. (1985) Calcium channel blockers in primary pulmonary hypertension. Chest 88(4 Suppl.): 257S-260S.
Rubin, L., Badesch, D., Barst, R., Galie, N., Black, C., Keogh, A. et al. (2002) Bosentan therapy for pulmonary arterial hypertension. N Engl J Med 346: 896-903.

V Freund-Michel, C Guibert et al.
http://tar.sagepub.com 199
Schermuly, R., Ghofrani, H., Wilkins, M. and Grimminger, F. (2011) Mechanisms of disease: pulmonary arterial hypertension. Nat Rev Cardiol 8: 443-455.
Sedeek, M., Hebert, R., Kennedy, C., Burns, K. and Touyz, R. (2009) Molecular mechanisms of hypertension: role of Nox family NADPH oxidases. Curr Opin Nephrol Hypertens 18: 122-127.
Shimoda, L. and Undem, C. (2010) Interactions between calcium and reactive oxygen species in pulmonary arterial smooth muscle responses to hypoxia. Respir Physiol Neurobiol 174: 221-229.
Simon, A., Rai, U., Fanburg, B. and Cochran, B. (1998) Activation of the JAK-STAT pathway by reactive oxygen species. Am J Physiol 275: C1640-C1652.
Simonneau, G., Robbins, I., Beghetti, M., Channick, R., Delcroix, M., Denton, C. et al. (2009) Updated clinical classification of pulmonary hypertension. J Am Coll Cardiol 54(1 Suppl.): S43-S54.
Sitbon, O., Humbert, M., Nunes, H., Parent, F., Garcia, G., Herve, P. et al. (2002) Long-term intravenous epoprostenol infusion in primary pulmonary hypertension: prognostic factors and survival. J Am Coll Cardiol 40: 780-788.
Song, M., Makino, A. and Yuan, J. (2011) Role of reactive oxygen species and redox in regulating the function of transient receptor potential channels. Antioxid Redox Signal 15: 1549-1565.
Spiekermann, S., Schenk, K. and Hoeper, M. (2009) Increased xanthine oxidase activity in idiopathic pulmonary arterial hypertension. Eur Respir J 34: 276.
Stenmark, K., Frid, M., Yeager, M., Li, M., Riddle, S., McKinsey, T. et al. (2012) Targeting the adventitial microenvironment in pulmonary hypertension: A potential approach to therapy that considers epigenetic change. Pulm Circ 2: 3-14.
Stowe, D. and Camara, A. (2009) Mitochondrial reactive oxygen species production in excitable cells: modulators of mitochondrial and cell function. Antioxid Redox Signal 11: 1373-1414.
Sturrock, A., Cahill, B., Norman, K., Huecksteadt, T., Hill, K., Sanders, K. et al. (2006) Transforming growth factor-beta1 induces Nox4 NAD(P)H oxidase and reactive oxygen species-dependent proliferation in human pulmonary artery smooth muscle cells. Am J Physiol Lung Cell Mol Physiol 290: L661-L673.
Sud, N., Sharma, S., Wiseman, D., Harmon, C., Kumar, S., Venema, R. et al. (2007) Nitric oxide and superoxide generation from endothelial NOS: modulation by HSP90. Am J Physiol Lung Cell Mol Physiol 293: L1444-L1453.
Sundaresan, M., Yu, Z., Ferrans, V., Irani, K. and Finkel, T. (1995) Requirement for generation of H2O2 for platelet-derived growth factor signal transduction. Science 270: 296-299.
Tabima, D., Frizzell, S. and Gladwin, M. (2012) Reactive oxygen and nitrogen species in pulmonary hypertension. Free Radic Biol Med 52: 1970-1986.
Tajima, M., Kurashima, Y., Sugiyama, K., Ogura, T. and Sakagami, H. (2009) The redox state of glutathione regulates the hypoxic induction of HIF-1. Eur J Pharmacol 606: 45-49.
ten Freyhaus, H., Dagnell, M., Leuchs, M., Vantler, M., Berghausen, E., Caglayan, E. et al. (2011) Hypoxia enhances platelet-derived growth factor signaling in the pulmonary vasculature by down-regulation of protein tyrosine phosphatases. Am J Respir Crit Care Med 183: 1092-1102.
Terada, L., Guidot, D., Leff, J., Willingham, I., Hanley, M., Piermattei, D. et al. (1992) Hypoxia injures endothelial cells by increasing endogenous xanthine oxidase activity. Proc Natl Acad Sci U S A 89: 3362-3366.
Thomas, M. (2010) Pharmacological targets for pulmonary vascular disease: vasodilation versus anti-remodelling. Adv Exp Med Biol 661: 475-490.
Tonelli, A., Alnuaimat, H. and Mubarak, K. (2010) Pulmonary vasodilator testing and use of calcium channel blockers in pulmonary arterial hypertension. Respir Med 104: 481-496.
Tonks, N. (2005) Redox redux: revisiting PTPs and the control of cell signaling. Cell 121: 667-670.
Toporsian, M., Jerkic, M., Zhou, Y., Kabir, M., Yu, L., McIntyre, B. et al. (2010) Spontaneous adult-onset pulmonary arterial hypertension attributable to increased endothelial oxidative stress in a murine model of hereditary hemorrhagic telangiectasia. Arterioscler Thromb Vasc Biol 30: 509-517.
Torrecillas, G., Boyano-Adanez, M., Medina, J., Parra, T., Griera, M., Lopez-Ongil, S. et al. (2001) The role of hydrogen peroxide in the contractile response to angiotensin II. Mol Pharmacol 59: 104-112.
Touyz, R. (2004) Reactive oxygen species, vascular oxidative stress, and redox signaling in hypertension: what is the clinical significance? Hypertension 44: 248-252.
Touyz, R. and Briones, A. (2010) Reactive oxygen species and vascular biology: implications in human hypertension. Hypertens Res 34: 5-14.
Umar, S., Hessel, M., Steendijk, P., Bax, W., Schutte, C., Schalij, M. et al. (2007) Activation of signaling molecules and matrix metalloproteinases in right ventricular myocardium of rats with pulmonary hypertension. Pathol Res Pract 203: 863-872.

Therapeutic Advances in Respiratory Disease 7 (3)
200 http://tar.sagepub.com
Usatyuk, P., Vepa, S., Watkins, T., He, D., Parinandi, N. and Natarajan, V. (2003) Redox regulation of reactive oxygen species-induced p38 MAP kinase activation and barrier dysfunction in lung microvascular endothelial cells. Antioxid Redox Signal 5: 723-730.
Ushio-Fukai, M., Alexander, R., Akers, M. and Griendling, K. (1998) p38 Mitogen-activated protein kinase is a critical component of the redox-sensitive signaling pathways activated by angiotensin II. Role in vascular smooth muscle cell hypertrophy. J Biol Chem 273: 15022-15029.
Ushio-Fukai, M., Alexander, R., Akers, M., Yin, Q., Fujio, Y., Walsh, K. et al. (1999) Reactive oxygen species mediate the activation of Akt/protein kinase B by angiotensin II in vascular smooth muscle cells. J Biol Chem 274: 22699-22704.
Ushio-Fukai, M., Griendling, K., Becker, P., Hilenski, L., Halleran, S. and Alexander, R. (2001) Epidermal growth factor receptor transactivation by angiotensin II requires reactive oxygen species in vascular smooth muscle cells. Arterioscler Thromb Vasc Biol 21: 489-495.
Victor, V., Rocha, M., Sola, E., Banuls, C., Garcia-Malpartida, K. and Hernandez-Mijares, A. (2009) Oxidative stress, endothelial dysfunction and atherosclerosis. Curr Pharm Des 15: 2988-3002.
Viedt, C., Soto, U., Krieger-Brauer, H., Fei, J., Elsing, C., Kubler, W. et al. (2000) Differential activation of mitogen-activated protein kinases in smooth muscle cells by angiotensin II: involvement of p22phox and reactive oxygen species. Arterioscler Thromb Vasc Biol 20: 940-948.
Voelkel, N., Gomez-Arroyo, J., Abbate, A., Bogaard, H. and Nicolls, M. (2012) Pathobiology of pulmonary arterial hypertension and right ventricular failure. Eur Respir J 2012; in press.
Waypa, G., Marks, J., Guzy, R., Mungai, P., Schriewer, J., Dokic, D. et al. (2010) Hypoxia triggers subcellular compartmental redox signaling in vascular smooth muscle cells. Circ Res 106: 526-535.
Waypa, G., Marks, J., Mack, M., Boriboun, C., Mungai, P. and Schumacker, P. (2002) Mitochondrial reactive oxygen species trigger calcium increases during hypoxia in pulmonary arterial myocytes. Circ Res 91: 719-726.
Wedgwood, S. and Black, S. (2003) Role of reactive oxygen species in vascular remodeling associated with pulmonary hypertension. Antioxid Redox Signal 5: 759-769.
Wedgwood, S. and Black, S. (2004) Combined superoxide dismutase/catalase mimetics alter fetal pulmonary arterial smooth muscle cell growth. Antioxid Redox Signal 6: 191-197.
Wedgwood, S., Dettman, R. and Black, S. (2001) ET-1 stimulates pulmonary arterial smooth muscle cell proliferation via induction of reactive oxygen species. Am J Physiol Lung Cell Mol Physiol 281: L1058-1067.
Weseler, A. and Bast, A. (2010) Oxidative stress and vascular function: implications for pharmacologic treatments. Curr Hypertens Rep 12: 154-161.
Wojciak-Stothard, B., Tsang, L. and Haworth, S. (2005) Rac and Rho play opposing roles in the regulation of hypoxia/reoxygenation-induced permeability changes in pulmonary artery endothelial cells. Am J Physiol Lung Cell Mol Physiol 288: L749-L760.
Wolin, M., Gupte, S., Neo, B., Gao, Q. and Ahmad, M. (2010) Oxidant-redox regulation of pulmonary vascular responses to hypoxia and nitric oxide-cGMP signaling. Cardiol Rev 18: 89-93.
Wong, C., Cheema, A., Zhang, L. and Suzuki, Y. (2008) Protein carbonylation as a novel mechanism in redox signaling. Circ Res 102: 310-318.
Wong, C., Preston, I., Hill, N. and Suzuki, Y. (2012) Iron chelation inhibits the development of pulmonary vascular remodeling. Free Radic Biol Med 53: 1738-1747.
Wunderlich, C., Schmeisser, A., Heerwagen, C., Ebner, B., Schober, K., Braun-Dullaeus, R. et al. (2008) Chronic NOS inhibition prevents adverse lung remodeling and pulmonary arterial hypertension in caveolin-1 knockout mice. Pulm Pharmacol Ther 21: 507-515.
Wung, B., Cheng, J., Chao, Y., Hsieh, H. and Wang, D. (1999) Modulation of Ras/Raf/extracellular signal-regulated kinase pathway by reactive oxygen species is involved in cyclic strain-induced early growth response-1 gene expression in endothelial cells. Circ Res 84: 804-812.
Yang, X., Lin, M. and Sham, J. (2010) Physiological functions of transient receptor potential channels in pulmonary arterial smooth muscle cells. Adv Exp Med Biol 661: 109-122.
Yildiz, P. (2009) Molecular mechanisms of pulmonary hypertension. Clin Chim Acta 403: 9-16.
Yoshizumi, M., Abe, J., Haendeler, J., Huang, Q. and Berk, B. (2000) Src and Cas mediate JNK activation but not ERK1/2 and p38 kinases by reactive oxygen species. J Biol Chem 275: 11706-11712.
Zou, M., Shi, C. and Cohen, R. (2002) Oxidation of the zinc-thiolate complex and uncoupling of endothelial nitric oxide synthase by peroxynitrite. J Clin Invest 109: 817-826.
Zuckerbraun, B., George, P. and Gladwin, M. (2011) Nitrite in pulmonary arterial hypertension: therapeutic avenues in the setting of dysregulated arginine/nitric oxide synthase signalling. Cardiovasc Res 89: 542-552.
Visit SAGE journals online http://tar.sagepub.com
SAGE journals





























