Embed Size (px)
Citation preview

E. Kebreab, J. Dijkstra, A. Bannink and J. France
Recent advances in modeling nutrient utilization in ruminants
doi: 10.2527/jas.2008-1313 originally published online Sep 26, 2008; 2009.87:E111-E122. J Anim Sci
http://jas.fass.org/cgi/content/full/87/14_suppl/E111the World Wide Web at:
The online version of this article, along with updated information and services, is located on
www.asas.org
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from

ABSTRACT: Mathematical modeling techniques have been applied to study various aspects of the rumi-nant, such as rumen function, postabsorptive metabo-lism, and product composition. This review focuses on advances made in modeling rumen fermentation and its associated rumen disorders, and energy and nutrient utilization and excretion with respect to environmental issues. Accurate prediction of fermentation stoichiom-etry has an impact on estimating the type of energy-yielding substrate available to the animal, and the ratio of lipogenic to glucogenic VFA is an important determi-nant of methanogenesis. Recent advances in modeling VFA stoichiometry offer ways for dietary manipulation to shift the fermentation in favor of glucogenic VFA. Increasing energy to the animal by supplementing with starch can lead to health problems such as sub-acute rumen acidosis caused by rumen pH depression. Mathematical models have been developed to describe changes in rumen pH and rumen fermentation. Models that relate rumen temperature to rumen pH have also been developed and have the potential to aid in the diagnosis of subacute rumen acidosis. The effect of pH has been studied mechanistically, and in such models,
fractional passage rate has a large impact on substrate degradation and microbial efficiency in the rumen and should be an important theme in future studies. The efficiency with which energy is utilized by ruminants has been updated in recent studies. Mechanistic models of N utilization indicate that reducing dietary protein concentration, matching protein degradability to the microbial requirement, and increasing the energy sta-tus of the animal will reduce the output of N as waste. Recent mechanistic P models calculate the P require-ment by taking into account P recycled through saliva and endogenous losses. Mechanistic P models suggest reducing current P amounts for lactating dairy cattle to at least 0.35% P in the diet, with a potential reduc-tion of up to 1.3 kt/yr. A model that integrates nutri-ent utilization and health has great potential benefit for ruminant nutrition research. Finally, whole-animal or farm level models are discussed. An example that used a multiple-criteria decision-making framework is reviewed, and the approach is considered to be appro-priate in dealing with the multidimensional nature of agricultural systems and can be applied to assist the decision process in cattle operations.
Key words: modeling, nutrient utilization, ruminant
©2009 American Society of Animal Science. All rights reserved. J. Anim. Sci. 2009. 87(E. Suppl.):E111–E122 doi:10.2527/jas.2008-1313
INTRODUCTION
Ruminants account for almost all of the milk and one-third of the meat production worldwide (FAO,
2007). The widespread application of advanced breed-ing and feeding technologies had a major effect on the improved production of ruminants. Additionally, data are being generated at a rapidly increasing rate because of advances in technology, computing, and engineer-ing. As qualitative knowledge increased, it became pos-sible to develop quantitative approaches to further our understanding of biological systems and to integrate various aspects of nutrient utilization. Initially, this was achieved by complex statistical analyses, but in recent years, dynamic mathematical models have been used not only to summarize existing data, but also to show where gaps in knowledge exist and where further re-
Recent advances in modeling nutrient utilization in ruminants1
E. Kebreab,*2 J. Dijkstra,† A. Bannink,‡ and J. France§
*Department of Animal Science, University of Manitoba, Winnipeg, Manitoba R3T 2N2, Canada; †Wageningen Institute of Animal Sciences, Animal Nutrition Group, Wageningen University, Marijkeweg 40,
6709 PG Wageningen, the Netherlands; ‡Animal Sciences Group, Division Animal Production, Wageningen University and Research Centre, PO Box 65, 8200 AB Lelystad, the Netherlands;
§Centre for Nutrition Modelling, Department of Animal and Poultry Science, University of Guelph, Guelph, Ontario N1G 2W1, Canada
1 Supported in part by the Canada Research Chairs Program (Ot-tawa, Ontario, Canada). Presented at the Early Career Achievement Awards during the joint annual meeting of the American Society of Animal Science and American Dairy Science Association in India-napolis, IN, July 7 to 11, 2008.
2 Corresponding author: [email protected] July 21, 2008.Accepted September 6, 2008.
E111
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from
search should focus (Dijkstra et al., 2005a). A search of all manuscripts published in the Journal of Animal Science with the key word of “modeling” revealed that nearly 50% of the 616 papers were published after the year 2000. No modeling manuscripts were found before 1970, and the proportion of total modeling papers to date increased from 3.2% in the 1970s to 17.0% in the 1980s and to 31.5% in the 1990s.
Various modeling approaches have been applied to study nutrient utilization in ruminants. These range from statistics, and in particular, meta-analysis, to op-erational research for the study of complex decision-making problems concerned with the best utilization of limited resources, and to applied mathematics us-ing theories of mass conservation and thermodynamics (France and Kebreab, 2008). The primary objective of this review is to summarize advances made through mathematical methods in understanding the underly-ing concepts of ruminant nutrient utilization at various levels of organization. Advances in rumen fermentation, including rumen health issues, and in energy and nutri-ent utilization and excretion in ruminants using studies that have utilized empirical and mechanistic approach-es are discussed.
RUMEN FERMENTATION
Fermentation of Substrates for Energy
Carbohydrate and protein digestion in the rumen, and starch and fat digestion in the intestine deliver most of the energy used by the ruminant. Dietary carbohy-drates, such as cellulose, hemicelluloses, pectin, starch, and soluble sugars, are the main sources of energy and are degraded in the rumen by microorganisms to hexo-ses and pentoses before being fermented to VFA. In ruminants, VFA can contribute up to 70% of the caloric requirement (Bergman, 1990). Acetate, propionate, and butyrate account for more than 95% of VFA produced, and the molar proportions of these VFA have implica-tions for energy utilization. Therefore, the fermentation stoichiometry of distinct substrates must be known. Murphy et al. (1982) made an important attempt to derive the coefficients that describe this stoichiometry by using a modeling approach. These coefficient values were considered generally applicable (Baldwin, 1995) and have been used in several mechanistic rumen mod-els (Baldwin et al., 1987; Dijkstra et al., 1992). Howev-er, it has been shown that the prediction of VFA molar proportions in rumen fluid of high-yielding dairy cows by these models is inaccurate (Bannink et al., 1997c). Subsequent simulation studies (Bannink et al., 1997b) demonstrated that this inaccuracy is most likely caused by an inadequate representation of the stoichiometry of VFA production or the rate of VFA absorption. In an attempt to improve predictability, Bannink et al. (2006) used meta-analysis of data from lactating cows only to determine coefficient values that define the con-
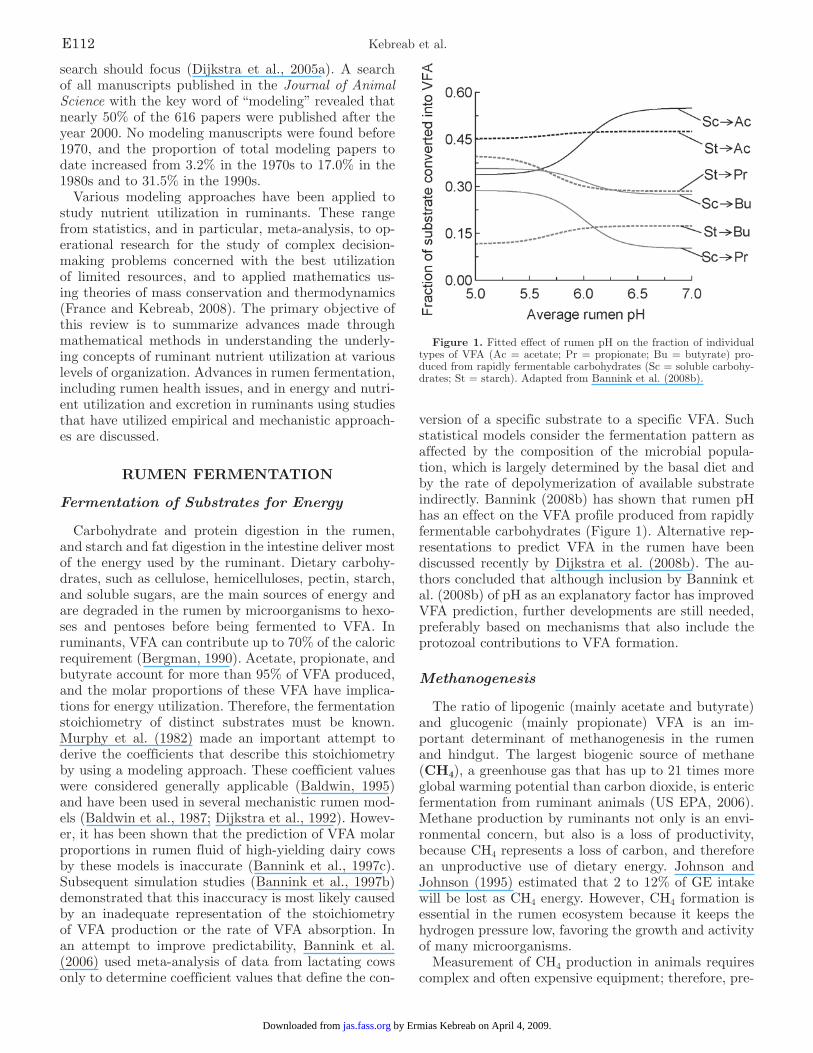
version of a specific substrate to a specific VFA. Such statistical models consider the fermentation pattern as affected by the composition of the microbial popula-tion, which is largely determined by the basal diet and by the rate of depolymerization of available substrate indirectly. Bannink (2008b) has shown that rumen pH has an effect on the VFA profile produced from rapidly fermentable carbohydrates (Figure 1). Alternative rep-resentations to predict VFA in the rumen have been discussed recently by Dijkstra et al. (2008b). The au-thors concluded that although inclusion by Bannink et al. (2008b) of pH as an explanatory factor has improved VFA prediction, further developments are still needed, preferably based on mechanisms that also include the protozoal contributions to VFA formation.
Methanogenesis
The ratio of lipogenic (mainly acetate and butyrate) and glucogenic (mainly propionate) VFA is an im-portant determinant of methanogenesis in the rumen and hindgut. The largest biogenic source of methane (CH4), a greenhouse gas that has up to 21 times more global warming potential than carbon dioxide, is enteric fermentation from ruminant animals (US EPA, 2006). Methane production by ruminants not only is an envi-ronmental concern, but also is a loss of productivity, because CH4 represents a loss of carbon, and therefore an unproductive use of dietary energy. Johnson and Johnson (1995) estimated that 2 to 12% of GE intake will be lost as CH4 energy. However, CH4 formation is essential in the rumen ecosystem because it keeps the hydrogen pressure low, favoring the growth and activity of many microorganisms.
Measurement of CH4 production in animals requires complex and often expensive equipment; therefore, pre-
Figure 1. Fitted effect of rumen pH on the fraction of individual types of VFA (Ac = acetate; Pr = propionate; Bu = butyrate) pro-duced from rapidly fermentable carbohydrates (Sc = soluble carbohy-drates; St = starch). Adapted from Bannink et al. (2008b).
Kebreab et al.E112
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from

diction equations are widely used to estimate CH4 emis-sions. Some models have been developed specifically to predict CH4 emissions from animals (Blaxter and Clap-perton, 1965; Moe and Tyrrell, 1979; Mills et al., 2003; Ellis et al., 2007) and others have been either modified or adapted to estimate CH4 emission from rumen fer-mentation (Dijkstra et al., 1992; Baldwin, 1995). At present, mathematical models are used to estimate CH4 emissions from enteric fermentation at a national and global level (see review by Kebreab et al., 2006a). The Intergovernmental Panel on Climate Change publishes guidelines (IPCC, 2006) that are used for official esti-mates of CH4 emissions. However, the accuracy of these models has been challenged (Kebreab et al., 2006b).
Using the rumen model of Dijkstra et al. (1992), Mills et al. (2001) added a mechanistic model of methanogen-esis that also included CH4 emissions from the hindgut. A comparison of models performed by Kebreab et al. (2008a) showed that mechanistic models were better than empirical ones in predicting CH4 emissions from US cattle and were more suited to assessing the effec-tiveness of mitigation options implemented at a farm or national level. Bannink et al. (2005) developed an adapted version of the model of Mills et al. (2001) and evaluated the impact of rumen fermentation conditions (i.e., pH) on type of VFA and amount of CH4 produced in the rumen of lactating cows. This version of the model has been used for estimating CH4 emissions from dairy cows in the Dutch Inventory Report since the year 1990 (Van der Maas et al., 2008). Empirical models are applicable only within the range in which they were developed; therefore, further advances in modeling CH4 emissions will probably result from a deepening of the mechanistic description of the underlying fermentation biology in the gastrointestinal tract of ruminants. To this end, Ellis et al. (2008) have reviewed and identified key aspects of rumen microbiology that could be incor-porated or that could improve representation within a model of ruminant digestion and environmental emis-sions. Descriptions of rumen pH dynamics (discussed subsequently) and of protozoal dynamics are the main key elements, but explicit representation of protozoa in rumen models is scarce (Dijkstra, 1994).
Acid Load in the Rumen
Rumen models (and experiments) have shown that feeding starch instead of structural carbohydrates fa-vors the production of glucogenic VFA and reduces CH4 emissions. However, overfeeding of starchy sub-strates, such as corn or wheat grains, to meet increased energy demands (e.g., during early lactation) may lead to a digestive disorder that affects health and produc-tivity, commonly known as subacute ruminal acidosis (SARA). Various authors (e.g., Garrett et al., 1999) have described SARA as the accumulation of organic acids within the rumen, causing depressed ruminal pH below a critical range (i.e., pH 5.0 to 5.8) for several hours.
Empirical Modeling of Rumen Acid Load. AlZahal et al. (2007) used meta-analysis to evaluate various equations to describe pH curves collected from different experiments that used continuous rumen pH measurement. The authors reported that the logistic equation (Eq. 1) described curves constructed from pH data:
y xy y
y y y
f
fkx
( ) ,=+ -( )é
ëêùûú
-
0
0 0 e [1]
where y(x) is the time spent below cutoff point x (min/d); x is pH −5; and y0, yf, and k (all >0) are parameters that define the scale and shape of the curve. AlZahal et al. (2007) suggested that the degree of decline in pH can be described by a shift of the pH curve position to-ward the decreased pH range, hence, by greater values of the predicted time and area below most critical pH cutoff points. The authors also suggested that the shift can be identified by a decrease in the curve inflection point (x*, y*) and curve slope, calculated as follows:
x
k
y y
y
y y
f
f
* ln ,
* .
=-æ
è
ççççç
ö
ø
÷÷÷÷÷÷
=
1
12
0
0 [2]
AlZahal et al. (2007) compared 3 concentrations of nonfiber carbohydrate in the diet (low, <37%; moder-ate, 37 to 39%; and high, ≥40% of DM). The authors reported that the inflection points for moderate and low nonfiber carbohydrate concentrations in the diet were 16 and 27% greater, respectively, compared with the diet with high concentrations of nonfiber carbo-hydrate. The authors argue that the approach allows comparison of pH data across studies and helps quanti-fy dietary effects on rumen pH. Similarly, using a meta-analysis, Zebeli et al. (2008) showed that an increase in physically effective NDF (peNDF) up to 31.2% of dairy cattle dietary DM increased rumen pH to a pla-teau value of 6.27, after which no further increase in pH was achieved. However, the concentration of peNDF required to stabilize rumen pH and maintain milk fat content without compromising milk energy efficiency varied. The required peNDF increased quadratically with increases in concentration of rumen degradable starch from grains and with increases of DMI.
Subsequently, AlZahal et al. (2008) developed a predictive equation that might aid in the diagnosis of SARA by investigating the relationship between rumen pH and rumen temperature. Rumen pH and tempera-ture were linearly associated within the range of 39 to 41°C, but the authors expected it to be nonlinear over extreme ranges of temperature and pH. The linear equation developed was
Modeling nutrient utilization E113
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from

pH = 16.9 − 0.29 temperature,
r2 = 0.77, n = 22, P = 0.002. [3]
Although rumen temperature can predict minimum rumen pH and has the potential to be developed into a noninvasive diagnostic technique, its use in field situa-tions depends on future development of a practical and cost-effective intraruminal wireless telemetry tempera-ture-sensing device compared with pH sensors.
Mechanistic Modeling of Rumen Acid Load. Bannink (2007) and Bannink et al. (2008b) developed a mechanistic model that represents the mechanisms involved with the formation of VFA in the lumen of the reticulorumen, absorption of VFA across the reticulo-rumen wall, and the intraepithelial metabolism of VFA by reticulorumen epithelia. The effects of pH on VFA yields from some substrates, including starch, were in-cluded in the model and were based on in vivo rumen observations. The model assesses the effect of nutri-tional strategies on VFA metabolism and acid load of the rumen. Simulations showed that a reduction of pH from 6.3 to 5.7 had moderate effects on the absorption rates of VFA from the rumen and that the relative con-tribution of facilitated transport of VFA was reduced drastically with reduced pH. Bannink et al. (2008b) considered the rumen wall to play a key regulatory role by reacting to exposure of the rumen epithelia to acid load. A dynamic model was constructed that combines the representation of the absorption of VFA by the ru-men wall and rumen wall adaptation to feeding strat-egy. They hypothesized that epithelial adaptation is evoked by the integration of the history of epithelia ex-posure to rumen VFA load. This memory function and the goal-directed adaptation of rumen epithelial tissue were represented by a dynamic modeling approach and a mechanism of negative feedback control. Simulations showed that a doubling of VFA production rate from 65 to 130 mol of VFA/d within 11 d postpartum resulted in 0 and 12% increases of rumen VFA concentrations after 10 and 20 d postpartum, respectively, similar to observed values. They concluded that the role of the rumen wall in preventing rumen contents from becom-ing too acidic during early lactation was more impor-tant than rumen pH in affecting VFA clearance. With respect to investigating the functionality of the rumen wall, Bannink et al. (2008a) recommended the use of genomics techniques in collecting more specific infor-mation on the genetic control and limits of the molecu-lar mechanisms involved with epithelial adaptation and function. They argued that a systems biology approach is needed to analyze the meaning of such data and to identify the precise mechanisms involved with rumen adaptation. Such an effort should include a representa-tion of changes in the rumen wall in response to nutri-tional strategies and the physiological state of the cow, as well as a representation of the intraluminal fermen-tation processes and conditions.
The pH of rumen fluid is an important indicator of rumen fermentation conditions. Simulating the effects of rumen pH on the diverse microbial groups present in the rumen remains a challenge, but perhaps an even greater task is to model adequately the diurnal fluctua-tions in pH itself. This would require the representation of all causal factors, which include all relevant buffer-ing mechanisms such as VFA production with microbial growth, VFA clearance with fluid outflow and absorp-tion by the rumen wall (passive and facilitated trans-port), and saliva production.
Rumen Feed Degradability
Feed evaluation systems and mechanistic rumen mod-els rely on degradation and passage kinetics. Therefore, accurate estimates of variables that drive degradation and passage are required. Several in vitro and in situ techniques have been developed to estimate the degrad-ability of feedstuffs in the rumen and their digestion in the whole gastrointestinal tract, including batch culture digestibility with rumen microbial inocula or added en-zymes and in situ methods. Regression equations are applied to predict in vivo digestibility from these in vitro or in situ methods (Dijkstra et al., 2005b). Fathi Nasri et al. (2006) evaluated linear, negative exponen-tial, and inverse polynomial models to describe DM and CP degradation kinetics. The authors reported that al-though there was no significant difference in predict-ing the extent of degradation, the linear and negative exponential models fit the data better when describing degradation kinetics. Lopez et al. (1999) showed that for analyzing in situ disappearance curves, sigmoidal-type models provided useful alternatives to the most commonly used negative exponential model. These sig-moidal models have the advantage of being versatile and able to cope with the changing shapes of disap-pearance profiles. They also represent the biological process in a more rational way than do nonsigmoidal models. However, the predicted extent of degradation was very similar among all models evaluated by Lopez et al. (1999). Thus, in choice of the model, the ability to obtain good estimates of degradation parameters and the feasibility of each model of being incorporated into the feeding system of choice are likely more important than the accuracy of fitting.
More recently, rate of degradation of feeds has been studied by using gas production profiles obtained from manual or automated systems of in vitro fermentation of feeds, and their application to ruminant feed eval-uation systems has been discussed by Dijkstra et al. (2005b). Dhanoa et al. (2004) proposed a combination of statistical and mechanistic models to determine ru-minal feed degradation based on a degradation profile using feces as the inoculum. France et al. (2005) devel-oped a 3-pool general compartmental model for inter-preting gas production profiles. The equations in the model permit the extent of ruminal degradation, and
Kebreab et al.E114
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from

the supply of microbial protein to the duodenum, to be evaluated, thereby linking the gas production tech-nique to animal production. Evaluation of the extent of degradation and the microbial protein supply relies on assumptions regarding fractional passage rate and microbial efficiency per unit of fermented substrate. These factors are unlikely to be constant, and calcula-tions suffer from the same drawbacks as empirical sys-tems. The equations are derived from first principles, and the potentially degradable feed fraction, undegrad-able feed fraction, and accumulated gases represent the underlying pools. In most cases, the models derived assume that substrate degradation follows first-order kinetics, resulting in nonlinear functions representing either diminishing returns (constant fractional degra-dation rate) or sigmoidal (fractional degradation rate that varies with time) behavior (France et al., 2005). Lopez et al. (2007) derived an alternative approach as-suming that degradation can follow zero-order kinetics, and evaluated the resulting piecewise linear model with other nonlinear models. Although the model showed acceptable behavior, nonlinear models showed some su-periority in terms of statistical goodness-of-fit, and the authors questioned the validity of the assumption of a constant degradation rate implied in the zero-order kinetics model. Ellis et al. (2005) considered various models for describing ruminal digestion of NDF based on the gamma distribution. The 2-pool, age-dependent, and gamma-distributed models were inferred to be su-perior when quality data were available, and the gam-ma-distributed age-constant model was superior when data quality was limited. However, gamma-distributed events can themselves be interpreted as simple com-partmental schemes, as demonstrated by France et al. (1985).
Much less recent attention has been given to modeling passage kinetics, although issues still remain, such as the nonavailability of models capable of resolving bimodal fecal marker excretion patterns (S. Lopez, University of Leon, Spain, personal communication). Experimental data on ruminal retention times are relatively scarce, yet accounting for retention time is a prerequisite for many feed evaluation systems (e.g., AFRC, 1993; NRC, 2001; Thomas, 2004), and fractional passage rates are also key parameters in mechanistic models of rumen fermentation (e.g., Dijkstra et al., 1992). Moreover, the fractional passage rate is a major determinant of mi-crobial protein efficiency, as indicated by the classical microbial growth equations derived by Pirt (1975) and observed experimentally (e.g., Oba and Allen, 2003). Fractional rate of growth of microbes is related to their fractional passage rate. A greater fractional rate of growth decreases the relative amount of substrate for maintenance purposes and increases efficiency. Even though the partitioning of energy into microbial growth and nongrowth functions has been mathematically well established, these are applied surprisingly little in empirical protein evaluation systems. Often external markers are used to estimate the passage dynamics of
feed particles. However, external markers may restrict the digestibility of the particles they are linked to, they may bind preferentially to small particles, and they may migrate to rumen fluid. Internal markers based on stable isotopes do not suffer from these disadvantages (Dijkstra et al., 2007). Fecal excretion of both external and internal markers can be described by using vari-ous models. Of these, the multicompartmental model of Dhanoa et al. (1985) has often been applied, and the results of this model based on fecal excretion pat-terns will yield estimates of the fractional passage rate from the slowest and second-slowest compartments. Fractional passage rates as applied in rumen models are usually based on estimates from regression equa-tions, in which DMI and proportion of concentrate or roughage of the diet are independent variables. Recent-ly, Seo et al. (2007) developed a mechanistic model to describe the physiological aspects of liquid dynamics in the rumen and to predict liquid flow out of the rumen quantitatively. These authors assumed the liquid flow through the reticuloomasal orifice to be coordinated with the frequency, duration, and amplitude of reticu-lar contractions during eating, ruminating, or resting. Such a model helps to understand the factors that af-fect the liquid outflow dynamics. Given the impact of passage on ruminal degradation and microbial growth, quantitative knowledge of rates of nutrient passage out of the rumen should be an important theme in future research.
ENERGY AND NUTRIENT UTILIZATION AND EXCRETION
Energy Utilization
Determination of energy requirements in ruminants and the efficiency with which energy is utilized have traditionally been described by the NE and ME sys-tems in North America and the United Kingdom, re-spectively (AFRC, 1993; NRC 2000, 2001). The values of the key parameters in the NE and ME systems were determined largely by using linear regression methods to analyze energy balance data from calorimetry ex-periments. Kebreab et al. (2003) analyzed an exten-sive data set by using a meta-analytical approach and nonlinear regression modeling. Based on the best fit models, the average efficiencies of utilization of ME in-take for milk production, BW gain, efficiency of uti-lization of tissue energy for milk production, and the maintenance energy requirement were 0.55, 0.83, 0.66, and 0.59 MJ/(kg0.75∙d) compared with 0.64, 0.75, 0.82, and 0.51 MJ/(kg0.75∙d) in the NE system (NRC, 2001) and 0.62, 0.60, 0.84, and 0.49 MJ/(kg0.75∙d) in the ME system, assuming the diet contains an ME:GE value of 0.6 (AFRC, 1993). The results of the analysis now make up the core values for the Feed into Milk system in the United Kingdom (Thomas, 2004). However, in an independent evaluation of various energy evaluation systems in grass-based diets fed to dairy cattle, the
Modeling nutrient utilization E115
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from

Feed into Milk system, like the Agricultural and Food Research Council system, still predicted energy require-ments for maintenance and milk production to be con-sistently less than actual energy supply (Dijkstra et al., 2008a). A mechanistic model based on those of Dijkstra et al. (1992, 1996a,b) predicted glucogenic nutrients to limit the performance of dairy cattle on grass-based di-ets. The mechanistic model proved to be more accurate and precise than the energy systems evaluated.
Current NE and ME systems do not consider the fact that animals have a certain requirement for particular nutrients per se, rather than just a requirement for their energetic potential. If diets are balanced for the same amount of ME or NE but the supply of glucogenic, ami-nogenic, or lipogenic nutrients differs, the response of the animal can vary. In a series of studies, Van Knegsel et al. (2007a,c) evaluated the effect of isocaloric (NE basis, Dutch VEM system) diets with different concen-trations of glucogenic or ketogenic nutrients in early-lactating dairy cattle (1 to 9 wk postpartum) by using indirect calorimetry in climate-respiration chambers. Cows fed the lipogenic diet had a greater milk fat pro-duction, and thus partitioned more energy to milk, than cows fed the glucogenic diet. Energy mobilized from body fat tended to be greater in the lipogenic group. In the glucogenic group, energy retention of body fat was positive from wk 8 postpartum, whereas in the li-pogenic group energy retention of body fat was still negative at wk 9 postpartum. Cows fed the lipogenic diet had greater plasma NEFA and decreased insulin concentrations than cows fed the glucogenic diet. The effects of isocaloric lipogenic or glucogenic diets on milk yield, milk composition, and plasma concentrations were confirmed in a large-scale trial (i.e., 114 cows; Van Knegsel et al., 2007b). Thus, cows fed the glucogenic diets evaluated have a reduced risk of metabolic disor-ders compared with cows fed lipogenic diets, although NE intake does not differ. Such results highlight vari-ous weaknesses of the current feed evaluation systems. Current energy and protein systems do not predict how production level and product composition will change in response to deliberate changes in feeding strategy. The amount of aggregation of these systems hampers the inclusion of new concepts and data that arise from experiments conducted at the organ, tissue, or cellular levels. To improve on current prediction schemes, new feeding systems need to be based on mechanisms that govern the response of animals to nutrients, by quan-titatively describing metabolite supply in more detail than the current aggregated components (Dijkstra et al., 2007).
N Utilization and Excretion
Nutritionists aim to maximize protein output in ani-mal products, such as milk and meat. The overall ef-ficiency of utilization of dietary N varied from 13 to 31% in various experiments measured by Kebreab et al. (2001b), but 40 to 45% efficiency values are theoretical-
ly possible (Hvelplund and Madsen, 1995). Increasing the efficiency of use of protein N by ruminants, leading to less N excretion, is becoming an environmental im-perative in many countries (Castillo et al., 2001). There have been several attempts to formulate mathematical models to predict N utilization from cattle. The models can be classified into 2 principal groups: empirical (sta-tistical) models that relate protein intake to N output directly and mechanistic models that attempt to simu-late N utilization based on a mathematical description of rumen and postrumen fermentation and digestion biochemistry.
Castillo et al. (2000) collated data from the literature and, through regression analysis, described the relation-ships between N intake and output in feces, urine, and milk. The authors reported that 72% of dietary N is ex-creted in manure. Kebreab et al. (2001b) further stud-ied the relationship between N intake and the form in which N is excreted because urinary N has been shown to be a much more important determinant of ammonia and N2O emissions and of nitrate leaching than fecal N. The authors reported a nonlinear correlation between N intake and urinary N, and the model predicted that approximately 80% of dietary N consumed above 500 g of N/d would be excreted in urine. The efficiency of N utilization for milk protein synthesis decreased from 31.5% at 400 g of N/d to 26.1% at 550 g of N/d for a 600-kg lactating dairy cow.
Nitrogen utilization is affected by many factors, such as dietary N concentration, degradability, microbial community, and their interaction with other nutrients (Firkins and Reynolds, 2005). Therefore, a mechanistic approach is necessary to improve the prediction of N utilization in cattle. Kebreab et al. (2002) developed a dynamic, process-based model that can predict the amount and form of N excreted by dairy cattle under various nutritional strategies, and they fitted predic-tions to observed values (Figure 2). The 4-pool model took into account several variables that determine N utilization, and they reported that urine N was sig-nificantly affected by protein degradability and energy status of the animal. The authors showed that increas-ing the energy concentration from 8 to 11 MJ of fer-mentable ME/kg of DM could potentially reduce urine N excretion and, by extension, ammonia emissions by up to 25%. Similarly, the authors argued that reducing dietary protein concentration from 19% of DM to ap-proximately 16% of DM could reduce ammonia emis-sions by 20% and reducing the degradability of protein to match the microbial requirement by 19%. Kebreab et al. (2004) went further by incorporating an N utili-zation module into the rumen model of Dijkstra et al. (1992). The integrated model was able to take advan-tage of detailed microbial representation in the rumen, which improved the prediction of N utilization by a lactating dairy cow. This representation allowed the as-sessment of diet manipulation to improve N utilization and, as a consequence, to reduce environmental pollu-tion caused by ammonia emissions and excess N excre-
Kebreab et al.E116
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from
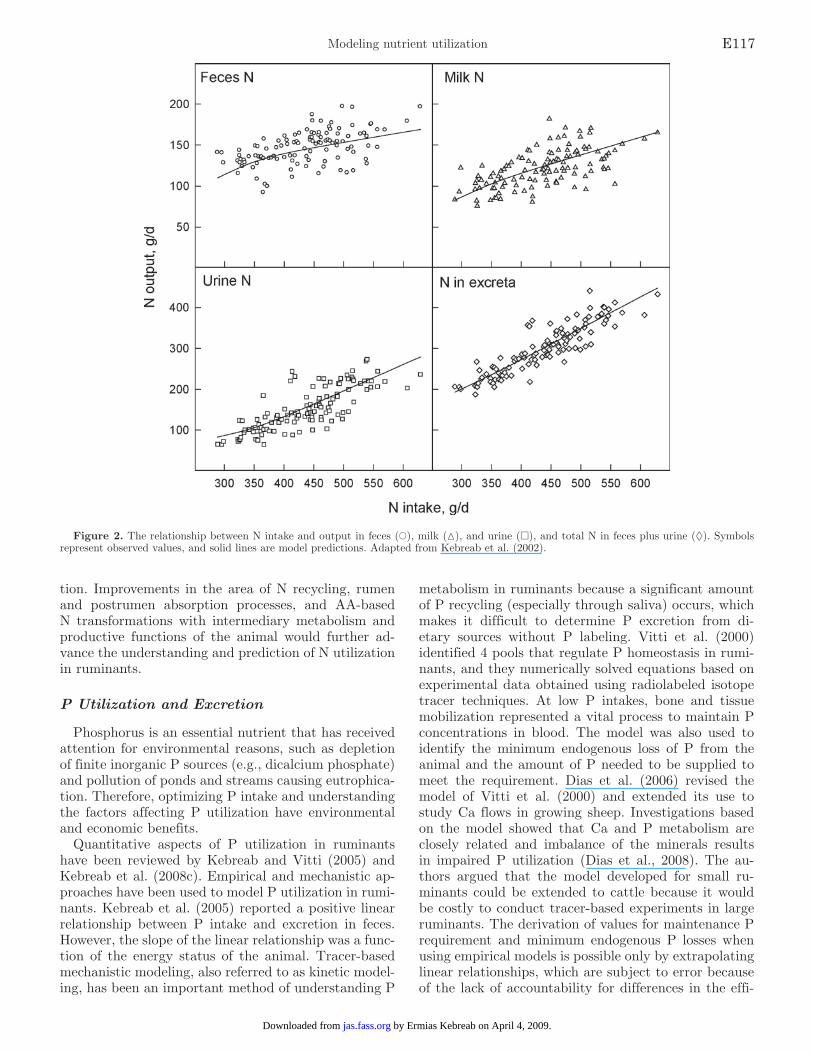
tion. Improvements in the area of N recycling, rumen and postrumen absorption processes, and AA-based N transformations with intermediary metabolism and productive functions of the animal would further ad-vance the understanding and prediction of N utilization in ruminants.
P Utilization and Excretion
Phosphorus is an essential nutrient that has received attention for environmental reasons, such as depletion of finite inorganic P sources (e.g., dicalcium phosphate) and pollution of ponds and streams causing eutrophica-tion. Therefore, optimizing P intake and understanding the factors affecting P utilization have environmental and economic benefits.
Quantitative aspects of P utilization in ruminants have been reviewed by Kebreab and Vitti (2005) and Kebreab et al. (2008c). Empirical and mechanistic ap-proaches have been used to model P utilization in rumi-nants. Kebreab et al. (2005) reported a positive linear relationship between P intake and excretion in feces. However, the slope of the linear relationship was a func-tion of the energy status of the animal. Tracer-based mechanistic modeling, also referred to as kinetic model-ing, has been an important method of understanding P
metabolism in ruminants because a significant amount of P recycling (especially through saliva) occurs, which makes it difficult to determine P excretion from di-etary sources without P labeling. Vitti et al. (2000) identified 4 pools that regulate P homeostasis in rumi-nants, and they numerically solved equations based on experimental data obtained using radiolabeled isotope tracer techniques. At low P intakes, bone and tissue mobilization represented a vital process to maintain P concentrations in blood. The model was also used to identify the minimum endogenous loss of P from the animal and the amount of P needed to be supplied to meet the requirement. Dias et al. (2006) revised the model of Vitti et al. (2000) and extended its use to study Ca flows in growing sheep. Investigations based on the model showed that Ca and P metabolism are closely related and imbalance of the minerals results in impaired P utilization (Dias et al., 2008). The au-thors argued that the model developed for small ru-minants could be extended to cattle because it would be costly to conduct tracer-based experiments in large ruminants. The derivation of values for maintenance P requirement and minimum endogenous P losses when using empirical models is possible only by extrapolating linear relationships, which are subject to error because of the lack of accountability for differences in the effi-
Figure 2. The relationship between N intake and output in feces (○), milk (▵), and urine (□), and total N in feces plus urine (◊). Symbols represent observed values, and solid lines are model predictions. Adapted from Kebreab et al. (2002).
Modeling nutrient utilization E117
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from

ciency of utilization of P fed at low concentrations and the availability of other nutrients and energy.
Mechanistic models of P utilization have been either developed as a stand-alone (Hill et al., 2008) or in-corporated into a larger model (Kebreab et al., 2004). The latter researchers developed a model containing 10 state variables representing P pools in the rumen, small intestine, large intestine, blood, and saliva, and incorporated it into the rumen model of Dijkstra et al. (1992). Comparison of model predictions with mea-sured values showed that mean fecal P excretion was within 8.7% of the observed mean. Inclusion of Ca me-tabolism and its interaction with P might increase the accuracy of prediction. Kebreab et al. (2008b) used the integrated model to assess the environmental and eco-nomic implications of reducing P pollution from dairy cows in Canada. Using the mechanistic model, the au-thors showed that producers could save up to CAN $20/cow per year by feeding 0.35% P, which, in many cases, means not supplementing the diet with inorganic P sources.
In constructing a dynamic, mechanistic model of P digestion and metabolism, Hill et al. (2008) took a slightly different approach to the model of Kebreab et al. (2004) because of different objectives. Hill et al. (2008) designed a model to investigate the composition of P in excreta, in particular the amounts of organic, inorganic, and phytate P. The model contained 5 state variables and 4 compartments, and a comparison of predicted against measured values showed that model prediction errors were within 20% of observed values. The authors indicated the need for additional data to derive parameters associated with the regulation of P absorption and bone P turnover because absorption ap-peared to be an important site of regulation. Repre-sentation of active and passive P transport across the intestinal wall would also advance understanding and prediction of P utilization.
Nutrient Utilization at the Animal and Whole-Farm Levels
Thornley and France (2007) described different levels of organization in representing biological systems. Ad-vances in whole-animal or farm level modeling include addition of methanogenesis, N, and P transactions into the mechanistic model of rumen function by Dijkstra et al. (1992), which has been described earlier (Kebreab et al., 2004). Usually, models of fermentation and diges-tion assume a constant feed intake. However, cows do not eat continuously but in discrete bouts, both dur-ing grazing (e.g., Taweel et al., 2004) and in stable-fed situations (e.g., Abrahamse et al., 2008). This intake behavior may have pronounced effects on the fermen-tation of substrates in the rumen. Among the major effects of a meal is the instantaneous decline in ru-men pH, in particular if the diet is rich in rapidly fer-mentable carbohydrates. Baldwin (1995) integrated the physical concept of particle dynamics with the chemical
transactions in his rumen model. He assumed that large particles are not subject to degradation or passage and are subject only to comminution. Thus, this represen-tation will greatly affect the diurnal pattern of rumen pool sizes and flows to the duodenum, but its effect on the simulated average extent of degradation compared with continuous feed intake is likely to be small (Ban-nink et al., 1997a). Collao-Saenz et al. (2005) adopted this representation as developed by Baldwin (1995) but included a representation of increased microbial death rate when substrate availability was decreased several hours after feeding. Moreover, rumen volume was as-sumed to be related to size of the meals and DM con-tent. Chilibroste et al. (2008) included a representation of effect of chewing efficiency on the release rate of intracellular constituents in their rumen model. This release rate appears to be an important determinant of instantaneous substrate availability to rumen microor-ganisms, and therefore also determines the rumen pH and VFA diurnal patterns. However, quantitative data on release rates are scarce. Moreover, insufficient quan-titative information on variation in fractional passage rates in meal-fed animals hampers further development of these models.
Nutrient digestion and utilization models for tropi-cal cattle have been developed and adapted for region-specific nutrient availability. Dijkstra et al. (1996a,b) constructed a model to predict nutrient supply to the animal from predefined dietary inputs, as a means of indicating pre-experimentally which combination of lo-cally available ingredients was most likely to enhance animal performance. The mechanistic model comprised 11 state variables representing N-containing, carbohy-drate, fatty acid, and microbial fractions, and 4 zero pools representing the nutrients available for absorp-tion. The model was optimized for cattle fed sugarcane-based diets, and Kebreab et al. (2001a) compared mod-el simulations with data observed in feeding trials. The authors were satisfied that the model predictions were close to observed values and, based on simulations, rec-ommended supplementing the basal diet with Leucaena and rice bran to increase milk production. Subsequent-ly, Chilibroste et al. (2001) and Rodrigues et al. (2002) adapted the model to evaluate supplements to enhance the nutrient supply and milk production of cattle fed diets based on ryegrass and elephant grass, respectively. Behera et al. (2005) also used the model to simulate milk production by dairy cows fed sugarcane top-based diets and other local supplements in India. Assis et al. (2008) reviewed advances in modeling sugarcane utili-zation by dairy cows in the tropics, describing the ad-dition of a submodel to represent endogenous protein N and large intestine digestion, discontinuous feed intake, and extension to other forage diets.
Various linear programming (LP) approaches were used to develop a whole-farm level model to optimize resource utilization in small-scale dairy operations in several stages. First, Val-Arreola et al. (2004c) used LP and partial budgeting methods to optimize land use
Kebreab et al.E118
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from

for forage production and nutrient availability, and to evaluate the economic impact of such changes in small-scale dairy systems. The model provided a solution for a better nutrient supply and for maintaining a greater stocking rate than the traditional strategy, and includ-ed utilizing a combination of forages such as alfalfa hay, corn silage, fresh ryegrass, and oat hay. Second, LP and Markov chain components were built to study the effect of calving interval on calving pattern and herd structure (Val-Arreola et al., 2004b). The Markov chain model suggested adopting different reproduction strategies according to period of the year in which the cow was expected to calve. Third, they conducted an analysis of feeding strategies for small-scale dairy systems by using LP (Val-Arreola et al., 2005). The study recommended an improvement in forage quality to increase efficiency, rather than increasing concentrate supplementation, as a viable strategy.
As part of optimizing nutrient utilization in the cow, Val-Arreola et al. (2004a) also considered 5 equations to describe the lactation curve. The authors used an extensive data set that was divided into small-scale and intensive systems and first, second, and third or greater parity to fit the equations. The equations described by Dijkstra et al. (1997; Eq. 4) consistently gave better predictions and because of its more mechanistic nature, the parameters lend themselves to physiological inter-pretations. Therefore, differences between systems and parity could be explained by variations in theoretical initial milk production at parturition (a), specific rates of secretory cell proliferation (b) and death (d), and specific rate of decay (c):
Y a b c dtct= - --exp[ ( ) / ],1 e [4]
where Y is milk yield (kg/d) and t is time of lactation (d).
Finally, a multiple-criteria decision-making frame-work was considered to be appropriate in dealing with the multidimensional nature of agricultural systems and was applied to assist the decision process in small-scale dairy farms (Val-Arreola et al., 2006). Goal pro-gramming and compromise programming were used to examine whether a single optimal solution or a set of nearly optimal solutions could better achieve the ob-jectives. Both models provided information concerning how a forage shortage might be alleviated by modi-fying the calving period to match nutrient availabil-ity with requirements at each lactation stage, with a minimum reduction in milk production. The use of a multiple-criteria decision-making framework and dy-namic programming techniques does allow more realis-tic representations than ordinary LP and offers better prediction in simulating whole-farm model interactions. Recommendations of management options or an evalu-ation of nutritional strategies under specific farming conditions requires realistic coefficients that are derived in an integrated manner.
SUMMARY AND CONCLUSIONS
In conclusion, empirical and mechanistic models have important applications in understanding rumen fermentation, and energy and nutrient utilization and excretion. In describing the relationships between nu-trient intake and excretion, empirical models can be useful, particularly if input data are limited. However, as Dijkstra et al. (2007) pointed out, empirical models lack the biological basis necessary to evaluate mitiga-tion strategies to reduce the excretion of waste, in-cluding N, P, and CH4, in an integrated manner. Such models may have little predictive value when compar-ing various feeding strategies. Examples include the Intergovernmental Panel on Climate Change (IPCC, 2006) Tier II models to quantify CH4 emissions and the current protein evaluation systems to evaluate low-protein diets to reduce N losses to the environment. Nutrient-based mechanistic models can address such issues (e.g., the Tier III model used by Van der Maas et al., 2008). These models still need to be extended to include prediction of rumen health issues such as SARA and to predict and understand the effects of nutritional strategy on pH and, subsequently, on rumen function. Finally, as advances in mathematical and computing techniques offer powerful data-processing capabilities, construction of mechanistic models to understand and predict biological phenomena at the organ, animal, and farm levels should be actively encouraged.
LITERATURE CITED
Abrahamse, P. A., B. Vlaeminck, S. Tamminga, and J. Dijkstra. 2008. The effect of silage and concentrate type on intake behav-ior, rumen function and milk production in dairy cows in early and late lactation. J. Dairy Sci. 91:4778–4792.
Agricultural and Food Research Council (AFRC). 1993. Energy and Protein Requirements of Ruminants. CAB International, Wall-ingford, United Kingdom.
AlZahal, O., E. Kebreab, J. France, M. Froetschel, and B. W. McBride. 2008. Ruminal temperature may aid in the detection of subacute ruminal acidosis. J. Dairy Sci. 91:202–207.
AlZahal, O., E. Kebreab, J. France, and B. W. McBride. 2007. A mathematical approach to predict biological values from rumen pH measurements. J. Dairy Sci. 90:3777–3785.
Assis, A. G., O. F. Campos, J. Dijkstra, E. Kebreab, and J. France. 2008. Modelling sugarcane utilization by dairy cows in the tropics. Pages 526–543 in Mathematical Modelling in Animal Nutrition. J. France and E. Kebreab, ed. CAB International, Wallingford, UK.
Baldwin, R. L. 1995. Modeling Ruminant Digestion and Metabo-lism. Chapman and Hall, London, UK.
Baldwin, R. L., J. H. M. Thornley, and D. E. Beever. 1987. Me-tabolism of the lactating dairy cow. II. Digestive elements of a mechanistic model. J. Dairy Res. 54:107–131.
Bannink, A. 2007. Modelling volatile fatty acid dynamics and ru-men function in lactating cows. PhD Diss. Wageningen Univ., Wageningen, the Netherlands.
Bannink, A., H. De Visser, J. Dijkstra, and J. France. 1997a. Impact of diet-specific input parameters on simulated rumen function. J. Theor. Biol. 184:371–384.
Bannink, A., H. De Visser, A. Klop, J. Dijkstra, and J. France. 1997b. Causes of inaccurate prediction of volatile fatty acids
Modeling nutrient utilization E119
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from

by simulation models of rumen function in lactating cows. J. Theor. Biol. 189:353–366.
Bannink, A., H. De Visser, and A. M. Van Vuuren. 1997c. Compari-son and evaluation of mechanistic rumen models. Br. J. Nutr. 78:563–581.
Bannink, A., J. Dijkstra, J. A. N. Mills, E. Kebreab, and J. France. 2005. Nutritional strategies to reduce enteric methane forma-tion in dairy cows. Pages 367–376 in Emissions from European Agriculture. T. Kuczynski, U. Dämmgen, J. Webb, and A. My-czko, ed. Wageningen Academic Publishers, Wageningen, the Netherlands.
Bannink, A., J. France, E. Kebreab, and J. Dijkstra. 2008a. Model-ling rumen wall functionality and potential for a systems biol-ogy approach. Can. J. Anim. Sci. In press.
Bannink, A., J. France, S. Lopez, W. J. J. Gerrits, E. Kebreab, S. Tamminga, and J. Dijkstra. 2008b. Modelling the implications of feeding strategy on rumen fermentation and functioning of the rumen wall. Anim. Feed Sci. Technol. 143:3–26.
Bannink, A., J. Kogut, J. Dijkstra, J. France, E. Kebreab, A. M. Van Vuuren, and S. Tamminga. 2006. Estimation of the stoichi-ometry of volatile fatty acid production in the rumen of lactat-ing cows. J. Theor. Biol. 238:36–51.
Behera, U. K., E. Kebreab, J. Dijkstra, A. G. Assis, and J. France. 2005. Simulation of milk production in dairy cows fed sugar-cane top based diets with locally available supplements under Indian conditions. J. Agric. Sci. 143:217–229.
Bergman, E. N. 1990. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 70:567–590.
Blaxter, K. L., and J. L. Clapperton. 1965. Prediction of the amount of methane produced by ruminants. Br. J. Nutr. 19:511–522.
Castillo, A. R., E. Kebreab, D. E. Beever, J. H. Barbi, J. D. Sutton, H. C. Kirby, and J. France. 2001. The effect of energy supple-mentation on nitrogen utilization in lactating dairy cows fed grass silage diets. J. Anim. Sci. 79:240–246.
Castillo, A. R., E. Kebreab, D. E. Beever, and J. France. 2000. A review of efficiency of nitrogen utilisation in dairy cows and its relationship with environmental pollution. J. Anim. Feed Sci. 9:1–32.
Chilibroste, P., J. Dijkstra, P. H. Robinson, and S. Tamminga. 2008. A simulation model “CTR Dairy” to predict the supply of nu-trients in dairy cows managed under discontinuous feeding pat-terns. Anim. Feed Sci. Technol. 143:148–173.
Chilibroste, P., J. Dijkstra, and S. Tamminga. 2001. Design and evaluation of a non-steady state rumen model. Neth. J. Agric. Sci. 49:297–312.
Collao-Saenz, E. A., J. Dijkstra, P. C. Aguiar Paiva, A. Bannink, P. B. Arcuri, J. C. Teixera, J. R. O. Perez, and F. M. David. 2005. Simulation model for particle dynamics in rumen of cattle fed sugarcane diet. Sci. Agric. 62:102–110.
Dhanoa, M. S., J. France, L. A. Crompton, R. M. Mauricio, E. Ke-breab, J. A. N. Mills, R. Sanderson, J. Dijkstra, and S. Lopez. 2004. A proposed method to determine the extent of degrada-tion of a feed in the rumen from the degradation profile ob-tained with the in vitro gas production technique using feces as the inoculum. J. Anim. Sci. 82:733–746.
Dhanoa, M. S., R. C. Siddons, J. France, and D. L. Gale. 1985. A multi-compartmental model to describe marker excretion pat-terns in ruminant faeces. Br. J. Nutr. 53:663–671.
Dias, R. S., E. Kebreab, D. M. S. S. Vitti, A. P. Roque, I. C. S. Bueno, and J. France. 2006. A revised model for studying phos-phorus and calcium kinetics in growing sheep. J. Anim. Sci. 84:2787–2794.
Dias, R. S., E. Kebreab, D. M. S. S. Vitti, A. P. Roque, and J. France. 2008. Application and comparison of two models to study the effects of calcium sources in sheep. Anim. Feed Sci. Technol. 143:89–103.
Dijkstra, J. 1994. Simulation of the dynamics of protozoa in the ru-men. Br. J. Nutr. 72:679–699.
Dijkstra, J., J. M. Forbes, and J. France. 2005a. Quantitative As-pects of Ruminant Digestion and Metabolism. 2nd ed. CAB International, Wallingford, UK.
Dijkstra, J., J. France, A. G. Assis, H. D. Neal, O. F. Campos, and L. J. M. Aroeira. 1996a. Simulation of digestion in cattle fed sugar cane: Prediction of nutrient supply for milk production with lo-cally available supplements. J. Agric. Sci. 127:247–260.
Dijkstra, J., J. France, M. S. Dhanoa, J. A. Maas, M. D. Hanigan, A. J. Rook, and D. E. Beever. 1997. A model to describe growth patterns of the mammary gland during pregnancy and lacta-tion. J. Dairy Sci. 80:2340–2354.
Dijkstra, J., J. France, H. D. St. C. Neal, A. G. Assis, L. J. M. Aroeira, and O. F. Campos. 1996b. Simulation of digestion in cattle fed sugarcane: Model development. J. Agric. Sci. 127:231–246.
Dijkstra, J., E. Kebreab, A. Bannink, L. A. Crompton, S. López, P. A. Abrahamse, P. Chilibroste, J. A. N. Mills, and J. France. 2008a. Comparison of energy evaluation systems and a mecha-nistic model for milk production by dairy cattle offered fresh grass-based diets. Anim. Feed Sci. Technol. 143:203–219.
Dijkstra, J., E. Kebreab, A. Bannink, J. France, and S. López. 2005b. Application of the gas production technique in feed evaluation systems for ruminants. Anim. Feed Sci. Technol. 123–124:561–578.
Dijkstra, J., E. Kebreab, J. France, and A. Bannink. 2008b. Model-ling protozoal metabolism and volatile fatty acid production in the rumen. Pages 170–188 in Mathematical Modelling in Animal Nutrition. J. France and E. Kebreab, ed. CAB Interna-tional, Wallingford, UK.
Dijkstra, J., E. Kebreab, J. A. N. Mills, W. F. Pellikaan, S. Lopez, A. Bannink, and J. France. 2007. Predicting the profile of nutri-ents available for absorption: From nutrient requirement to ani-mal response and environmental impact. Animal 1:99–111.
Dijkstra, J., H. D. St. C. Neal, D. E. Beever, and J. France. 1992. Simulation of nutrient digestion, absorption and outflow in the rumen: Model description. J. Nutr. 122:2239–2256.
Ellis, J. L., J. Dijkstra, E. Kebreab, A. Bannink, N. E. Odongo, B. W. McBride, and J. France. 2008. Aspects of rumen microbiol-ogy central to mechanistic modelling of methane production in cattle. J. Agric. Sci. 146:213–233.
Ellis, J. L., E. Kebreab, N. E. Odongo, B. W. McBride, E. K. Okine, and J. France. 2007. Prediction of methane production from dairy and beef cattle. J. Dairy Sci. 90:3456–3467.
Ellis, W. C., M. Mahlooji, and J. H. Matis. 2005. Models for esti-mating parameters of neutral detergent fiber digestion by rumi-nal microorganisms. J. Anim. Sci. 83:1591–1601.
Fathi Nasri, M. H., M. Danesh Mesgaran, J. France, J. P. Cant, and E. Kebreab. 2006. Evaluation of models to describe ruminal degradation kinetics from in situ ruminal incubation of whole soybeans. J. Dairy Sci. 89:3087–3095.
Firkins, J. L., and C. K. Reynolds. 2005. Whole animal nitrogen balance in cattle. Pages 167–185 in Nitrogen and Phosphorus Nutrition of Cattle and Environment. E. Pfeffer and A. Hristov, ed. CAB International, Cambridge, MA.
Food and Agriculture Organization of the United Nations (FAO). 2007. The State of Food and Agriculture. FAO, Rome, Italy.
France, J., and E. Kebreab. 2008. Mathematical Modelling in Ani-mal Nutrition. CAB International, Wallingford, UK.
France, J., S. López, E. Kebreab, A. Bannink, M. S. Dhanoa, and J. Dijkstra. 2005. A general compartmental model for interpret-ing gas production profiles. Anim. Feed Sci. Technol. 123–124:473–485.
France, J., J. H. M. Thornley, M. S. Dhanoa, and R. C. Siddons. 1985. On the mathematics of digesta flow kinetics. J. Theor. Biol. 113:743–758.
Garrett, E. F., M. N. Pereira, K. V. Nordlund, L. E. Armentano, W. J. Goodger, and G. R. Oetzel. 1999. Diagnostic methods for the detection of subacute ruminal acidosis in dairy cows. J. Dairy Sci. 82:1170–1178.
Kebreab et al.E120
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from

Hill, S. R., K. F. Knowlton, E. Kebreab, J. France, and M. D. Hani-gan. 2008. A model of phosphorus digestion and metabolism in the lactating dairy cow. J. Dairy Sci. 91:2021–2032.
Hvelplund, T., and J. Madsen. 1995. Protein utilization in rumi-nants. Pages 83–89 in Proc. 7th Int. Symp. Protein Metabolism and Nutrition. A. F. Nunes, A. V. Portugal, J. P. Costa, and J. R. Ribeiro ed. Estacoa Zootechnia National Santerem, Valde Santare, Portugal.
Intergovernmental Panel on Climate Change (IPCC). 2006. 2006 IPCC Guidelines for National Greenhouse Gas Inventories. IGES, Hayama, Kanagawa, Japan. http://www.ipcc-nggip.iges.or.jp. Accessed Jan. 20, 2008.
Johnson, K. A., and D. E. Johnson. 1995. Methane emissions from cattle. J. Anim. Sci. 73:2483–2492.
Kebreab, E., A. G. Assis, J. Dijkstra, and J. France. 2001a. Evalu-ating sugarcane diets for dairy cows using a digestion model. Trop. Anim. Health Prod. 33:127–139.
Kebreab, E., K. Clarke, C. Wagner-Riddle, and J. France. 2006a. Methane and nitrous oxide emissions from Canadian animal agriculture—A review. Can. J. Anim. Sci. 86:135–158.
Kebreab, E., J. France, R. E. Agnew, T. Yan, M. S. Dhanoa, J. Dijkstra, D. E. Beever, and C. K. Reynolds. 2003. Alternatives to linear analysis of energy balance data from lactating dairy cows. J. Dairy Sci. 86:2904–2913.
Kebreab, E., J. France, D. E. Beever, and A. R. Castillo. 2001b. Ni-trogen pollution by dairy cows and its mitigation. Nutr. Cycl. Agroecosyst. 60:275–285.
Kebreab, E., J. France, B. W. McBride, N. E. Odongo, A. Bannink, J. A. N. Mills, and J. Dijkstra. 2006b. Evaluation of models to predict methane emissions from enteric fermentation in North American dairy cattle. Pages 299–313 in Nutrient Utilization in Farm Animals: Modelling Approach. E. Kebreab, J. Dijkstra, W. J. J. Gerrits, A. Bannink, and J. France, ed. CAB Interna-tional, Wallingford, UK.
Kebreab, E., J. France, J. A. N. Mills, R. Allison, and J. Dijkstra. 2002. A dynamic model of N metabolism in the lactating dairy cow and an assessment of impact of N excretion on the environ-ment. J. Anim. Sci. 80:248–259.
Kebreab, E., J. France, J. D. Sutton, L. A. Crompton, and D. E. Beever. 2005. Effect of energy and protein supplementation on phosphorus utilization in lactating dairy cows. J. Anim. Feed Sci. 14:63–77.
Kebreab, E., K. A. Johnson, S. L. Archibeque, D. Pape, and T. Wirth. 2008a. Model for estimating enteric methane emissions from US cattle. J. Anim. Sci. 86:2738–2748.
Kebreab, E., J. A. N. Mills, L. A. Crompton, A. Bannink, J. Di-jkstra, W. J. J. Gerrits, and J. France. 2004. An integrated mathematical model to evaluate nutrient partition in dairy cattle between the animal and its environment. Anim. Feed Sci. Technol. 112:131–154.
Kebreab, E., N. E. Odongo, B. W. McBride, M. D. Hanigan, and J. France. 2008b. Phosphorus utilization and environmental and economic implications of reducing phosphorus pollution from Ontario dairy cows. J. Dairy Sci. 91:241–246.
Kebreab, E., and D. M. S. S. Vitti. 2005. Mineral metabolism. Pages 469–486 in Quantitative Aspects of Ruminant Digestion and Metabolism. J. Dijkstra, J. M. Forbes, and J. France, ed. CAB International, Wallingford, UK.
Kebreab, E., D. M. S. S. Vitti, N. E. Odongo, L. A. Crompton, and J. France. 2008c. Modelling phosphorus metabolism. Pages 353–369 in Mathematical Modelling in Animal Nutrition. J. France and E. Kebreab, ed. CAB International, Wallingford, UK.
Lopez, S., M. S. Dhanoa, J. Dijkstra, A. Bannink, E. Kebreab, and J. France. 2007. Some methodological and analytical consider-ations regarding application of the gas production technique. Anim. Feed Sci. Technol. 135:139–156.
Lopez, S., J. France, M. S. Dhanoa, F. Mould, and J. Dijkstra. 1999. Comparison of mathematical models to describe disappearance curves obtained using the polyester bag technique for incubat-ing feeds in the rumen. J. Anim. Sci. 77:1875–1888.
Mills, J. A. N., J. Dijkstra, A. Bannink, S. B. Cammell, E. Ke-breab, and J. France. 2001. A mechanistic model of whole-tract digestion and methanogenesis in the lactating dairy cow: Model development, evaluation, and application. J. Anim. Sci. 79:1584–1597.
Mills, J. A. N., E. Kebreab, C. Yates, L. A. Crompton, S. B. Cam-mell, M. S. Dhanoa, R. E. Agnew, and J. France. 2003. Alter-native approaches to predicting methane emissions from dairy cows. J. Anim. Sci. 81:3141–3150.
Moe, P. W., and H. F. Tyrrell. 1979. Methane production in dairy cows. J. Dairy Sci. 62:1583–1586.
Murphy, M. R., R. L. Baldwin, and L. J. Koong. 1982. Estimation of stoichiometric parameters for rumen fermentation of roughage and concentrate diets. J. Anim. Sci. 55:279–296.
NRC. 2000. Nutrient Requirements of Beef Cattle. 7th rev. ed. Up-date 2000. Natl. Acad. Press., Washington, DC.
NRC. 2001. Nutrient Requirements of Dairy Cattle. 7th rev. ed. Natl. Acad. Press., Washington, DC.
Oba, M., and M. S. Allen. 2003. Effect of diet fermentability on effi-ciency of microbial nitrogen production in lactating dairy cows. J. Dairy Sci. 86:195–207.
Pirt, S. J. 1975. Principles of Microbe and Cell Cultivation. Black-well Scientific Publication, Oxford, UK.
Rodrigues, S. I. F. C, J. Dijkstra, A. G. Assis, J. France, and S. Tamminga. 2002. Evaluation of supplements to enhance nutri-ent supply and milk production of cattle fed diets based on elephant grass (Pennisetum purpureum) using a mathematical model. Livest. Prod. Sci. 77:273–286.
Seo, S., C. Lanzas, L. O. Tedeschi, and D. G. Fox. 2007. Develop-ment of a mechanistic model to represent the dynamics of liquid flow out of the rumen and to predict the rate of passage of liquid in dairy cattle. J. Dairy Sci. 90:840–855.
Taweel, H. Z., B. M. Tas, J. Dijkstra, and S. Tamminga. 2004. Intake regulation and grazing behavior of dairy cows under continuous stocking. J. Dairy Sci. 87:3417–3427.
Thomas, C. 2004. Feed into Milk: A New Applied Feeding System for Dairy Cows. Nottingham University Press, Nottingham, UK.
Thornley, J. H. M., and J. France. 2007. Mathematical Models in Agriculture: A Quantitative Approach to Problems in Agricul-ture and Related Sciences. 2nd ed. CAB International, Wall-ingford, UK.
US Environmental Protection Agency (US EPA). 2006. Global An-thropogenic Non-CO2 Greenhouse Gas Emissions: 1990–2020. EPA Report 430-R-06-003, June 2006. http://www.epa.gov/methane/pdfs/GreenhouseGasReport.pdf Accessed Jun. 3, 2008.
Val-Arreola, D., E. Kebreab, J. Dijkstra, and J. France. 2004a. Study of the lactation curve in dairy cattle on farms in central Mexico. J. Dairy Sci. 87:3789–3799.
Val-Arreola, D., E. Kebreab, and J. France. 2006. Modeling small-scale dairy farms in central Mexico using multi criteria pro-gramming. J. Dairy Sci. 89:1662–1672.
Val-Arreola, D., E. Kebreab, J. A. N. Mills, and J. France. 2004b. Analysis of the calving pattern of herds in small-scale dairy systems in central Mexico. J. Anim. Feed Sci. 13:557–574.
Val-Arreola, D., E. Kebreab, J. A. N. Mills, and J. France. 2005. Analysis of feeding strategies for small-scale dairy systems in central Mexico using linear programming. J. Anim. Feed Sci. 14:607–624.
Val-Arreola, D., E. Kebreab, S. L. Wiggins, and J. France. 2004c. A study of forage production and nutrient availability in small-scale production systems in central Mexico. Nutr. Cycl. Agro-ecosyst. 69:191–201.
Van der Maas, C. W. M., P. W. H. G. Coenen, P. G. Ruyssenaars, H. H. J. Vreuls, L. J. Brandes, K. Baas, G. van den Berghe, G. J. van den Born, B. Guis, A. Hoen, R. te Molder, D. S. Nijdam, J. G. J. Olivier, C. J. Peek, and M. W. van Schijn-del. 2008. Greenhouse Gas Emissions in the Netherlands 1990–2006. National Inventory Report 2008. MNP report 500080009. Netherlands Environmental Assessment Agency, Bilthoven, the Netherlands.
Modeling nutrient utilization E121
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from

Van Knegsel, A. T. M., H. Van den Brand, J. Dijkstra, W. M. Van Straalen, M. J. W. Heetkamp, S. Tamminga, and B. Kemp. 2007a. Dietary energy source in dairy cows in early lacta-tion: Energy partitioning and milk composition. J. Dairy Sci. 90:1467–1476.
Van Knegsel, A. T. M., H. Van den Brand, J. Dijkstra, W. M. Van Straalen, R. Jorritsma, S. Tamminga, and B. Kemp. 2007b. Ef-fect of glucogenic vs. lipogenic diets on energy balance, blood metabolites, and reproduction in primiparous and multiparous dairy cows in early lactation. J. Dairy Sci. 90:3397–3409.
Van Knegsel, A. T. M., H. Van den Brand, E. A. M. Graat, J. Dijk-stra, R. Jorritsma, E. Decuypere, S. Tamminga, and B. Kemp.
2007c. Dietary energy source in dairy cows in early lactation: Metabolites and metabolic hormones. J. Dairy Sci. 90:1477–1485.
Vitti, D. M. S. S., E. Kebreab, J. B. Lopes, A. L. Abdalla, F. F. R. De Carvalho, K. T. De Resende, L. A. Crompton, and J. France. 2000. A kinetic model of phosphorus metabolism in growing goats. J. Anim. Sci. 78:2706–2712.
Zebeli, Q., J. Dijkstra, M. Tafaj, H. Steingass, B. N. Ametaj, and W. Drochner. 2008. Modeling the adequacy of dietary fiber in dairy cows based on the responses of ruminal pH and milk fat produc-tion to composition of the diet. J. Dairy Sci. 91:2046–2066.
Kebreab et al.E122
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from

References http://jas.fass.org/cgi/content/full/87/14_suppl/E111#BIBL
This article cites 67 articles, 33 of which you can access for free at:
by Ermias Kebreab on April 4, 2009. jas.fass.orgDownloaded from





























