Embed Size (px)
Citation preview

Seediscussions,stats,andauthorprofilesforthispublicationat:https://www.researchgate.net/publication/240380219
Recruitmentofcerithiidgastropods(Rhinoclavisspp.)insedimentsatOneTreeReef,GreatBarrierReef
ARTICLEinJOURNALOFEXPERIMENTALMARINEBIOLOGYANDECOLOGY·APRIL1992
ImpactFactor:1.87·DOI:10.1016/0022-0981(92)90013-Z
CITATIONS
9
READS
23
1AUTHOR:
GregAlanSkilleter
UniversityofQueensland
58PUBLICATIONS1,378CITATIONS
SEEPROFILE
Allin-textreferencesunderlinedinbluearelinkedtopublicationsonResearchGate,
lettingyouaccessandreadthemimmediately.
Availablefrom:GregAlanSkilleter
Retrievedon:05February2016

J. Exp. Mar. Biol. Ecol., 156 (1992) 1-21 © 1992 Elsevier Science Publishers BV. All rights reserved 0022-0981/92/$05.00
JEMBE 01714
Recruitment of cerithiid gastropods (Rhinoclavis spp°) in sediments at One Tree Reef, Great Barrier Reef
G.A. Skilleter bastitute of Marine Ecology, University of Sydney, Sydney, New South Wales, Australia
(Received 10 July 1991; revision received 1 October 1991; accepted 21 October 1991)
Abstract: The abundance of three species of deposit-feeding cerithiid gastropods, Rhinoclavis aspera (Linnaeus, 1758), R.fasciata (Brugui6re, 1792) and R. vertagus (Linnaeus, 1758), was monitored seasonally from May 1986 to January 1987 in sediments at One Tree Reef, southern Great Barrier Reef. All three species were most abundant in the coarse sandy sediments of a shallow sandflat, rather than in the finer sediments found in deeper channels within the lagoon. In contrast deposit-feeding temperate zone infauna are typically more abundant in finer sediments. To evaluate whether this pattern of greater abundance in sandy sediments was a result ofgreater recruitment to the sandflat compared with the channels, recruitment of each species was determined in each of two successive summers, January 1987 and January 1988. R. aspera recruited mostly to the sandflat compared with other habitats in both of 1987 and 1988, but for R.fasciata and R. vertagus this was only the case in 1988. These results are consistent with a model oflarval depletion in the water column because larvae are always carried across the sandflat by water currents before reaching the lagoonal habitats so there are fewer larvae left in the water to settle into the finer sediments of the lagoon. The effect of removing all adult macrofaunal molluscs from the sediment on the recruitment of each species was also experimentally determined at 10 locations. The results varied among locations supporting alternative models of adult enhancement of recruitment, reduction of recruitment due to adults, or no effect of adults on recruitment. The results of this experiment, however, provide evidence against the previous larval depletion model because in some cases recruitment was increased in the lagoonal habitats where adults were removed indicating that there were still larvae available in the water over the fine sediments.
Key words: Cerithiid; Coral reef; Infauna; Recruitment; Rhinoclavis
INTRODUCTION
One generalization to arise from the many studies characterizing soft-sediment com- munities is that deposit-feeding infauna typically are found in greater abundances in habitats dominated by fine sediments (Sanders, 1958, 1960; Rhoads & Young, 1970; Levinton, 1972; Holland & Polgar, 1976; Peterson, 1977; Whitlatch, 1977, 1981; Murphy, 1985). This relationship is believed to reflect differences in food availability: presumably, finer sediments contain more organic matter than sandier sediments (Sanders, 1958; Newell, 1965; Dale, 1974; Tunnicliffe & Risk, 1977). Habitats with sandy sediments, in contrast, are dominated by suspension-feeding infauna, especially
Correspondence address: G.A. Skilleter, University of North Carolina at Chapel Hill, Institute of Marine Sciences, Morehead City, NC 28557, USA.
This paper is a contribution from the University of Sydney, One Tree Island Field Station.

2 G.A. SKILLETER
bivalves (Sanders, 1958; Rhoads & Young, 1970; Levinton, 1972; Peterson, 1977; Peterson & Black, 1987).
Although there are examples where this apparent pattern of deposit-feeders in fine sediments and suspension-feeders in sandy sediments does not apply (e.g. Sanders et al., 1962; Rhoads & Young, 1970; Young & Rhoads, 1971; Aller & Dodge, 1974; Maurer et ai., 1979), these "exceptions" have been of little interest to researchers (but see Dobbs & Whitlatch, 1982; Bianchi, 1988; Kemp, 1988). Placing emphasis on the study of such a diverse group of animals as the "deposit-feeders" from only one of several possible habitat types in which they occur may bias our perceptions of the factors that are important to the animals when we try to make generalisations about their ecology. More studies in a greater variety of habitats will provide further insight into the factors which determine the distribution and abundance of deposit-feeding species.
Initial surveys of sediments in the lagoon at One Tree Reef, southern Great Barrier Reef, in October 1985, showed that the deposit-feeding cerithiid gastropods Rhinoclavis aspera (Linnaeus, 1758), R. fasciata (Brugui6re, 1792) and R. vertagus (Linnaeus, 1758) were among the most abundant infaunal molluscs present in the lagoon but that these snails reached their greatest abundances in the coarse sandy sediments on a shallow back-reef sandflat on the southern side of the lagoon (Jones et al., 1990; SkiUeter, 1991). 1~he sediments on the back-reef sandflat were significantly coarser than those in other lagoonal habitats (Johnstone et al., 1990; Jones et al., 1990). Here, I extend these initial surveys, which were only done on a single occasion, to determine whether this pattern of greater densities ofRhinoclavis spp. in the coarse sediments on the sandflat, compared with the finer sediments elsewhere, was maintained through time.
I estimated the intensity of recruitment of Rhinoclavis spp. at different spatial scales on the sandflat and in the lagoon at One Tree Reef in two successive summers in order to determine the relationship between recruitment and the differences in density in the lagoon and sandflat respectively. Simultaneously, I also investigated the patterns of recruitment of Rhinoclavis across the lagoon in the absence of established macrofauna, to determine if resident int'auna have significant effects on the recruitment of other organisms in sediments (Williams, 1980; Peterson, 1982; Crowe et al., 1987; Hunt et al., 1987; Commito & Boncavage, 1989; Hines et al., 1989).
MATERIALS AND METHODS
This study was done in the main lagoon of One Tree Reef (23 ° 30' S, 152 ° 06' E), on the eastern edge of the Capricorn Group of reefs on the southern Great Barrier Reef (Fig. 1). The main lagoon is bounded on the eastern and southern sides by subtidal sandflats and at its centre has a maze of reticulated and linear patch reefs (Marshall & Davies, 1982). These sandflats will be referred to as the "southern sandflat" and "eastern sandflat", respectively. Deep channels occur down the southern and north- eastern sections of the lagoon (Davies et al., 1976). These are referred to as the "south channel" and "'north channel", respectively.

RECRUITMENT OF CERITHIID GASTROPODS 3
6@ ~7
Group Great Reef
N l )NE TREE
REEF
channel N~,..K,. La "~ '~~ ,~ ,~d~ .J'
I I 1 km
23 ° 30' S
Leastern / sandflat
' c °n"n"o,
Fig. 1. Map of Australia showing location of One Tree Reef at southern end of Great Barrier Reef; map of One Tree Reef showing ma:n features of reef.
To determine whether the patterns of abundance observed in October 1985 (Jones et al., 1990; Skilleter, 1991) were maintained through time, the densities of Rhinoclavis aspera, R. fasciata and R. vertaguswere estimated at different locations on the southern sandflat and in the lagoor~ seasonally from May 1986 to January 1987. Four locations were sampled on the southern sandflat and two locations were sampled in each of the south channel, north channel and eastern sandflat habitats. On each occasion, at each location, four replicate 225-cm 2 samples were collected from each of six haphazardly positioned sites.
Samples were collected with a diver-operated compressed-air-powered suction lift, which deposited the sediment contained within a 225-cm 2 core into a mesh bag. The bags acted to sieve, in situ, the sediment through a 1-mm mesh. Samples were collected to a depth of 7 cm, then transferred to the laboratory, and kept in running seawater until sorted. All molluscs were identified and then returned to the location from which they were collected.

4 G.'A. SKILLETER
Densities of recruits were estimated at each location in January 1987 and January 1988, from samples taken weekly at a number of haphazardly located places on the sandflat. The air-lift did not operate efficiently with bags of mesh size < 1 mm so samples were collected by hand-scooping the sediment from within a 225-cm 2 core into plastic bags in order to retain any recruits < 1 mm in length. These samples were sieved through a 0.5-mm mesh and fixed in 10% formalin/seawater and then stained with Biebricht Scarlet. The fraction of the sample retained in the 0.5-mm sieve was then examined for the presence of recruits of Rhinoclavis. Recruits of all three species of Rhinoclavis stained bright pink and were easily sorted from the white carbonate sands. The first recruits were detected in the third week of January 1987 and in the second week in 1988. Animals were already > 750 #m in size (maximal width of shell). At this time, I then co~lected four replicate 225-cm 2 samples at each of six haphazardly positioned sites from all locations previously sampled for adults. These samples were also collected by hand-scooping the sediment from within the corer into plastic bags. Each sample was then sieved through a 750-#m mesh sieve and the remaining fraction of the sample was fixed in 10% formalin/seawater, stained with Biebricht Scarlet, and the molluscs removed.
The total number of R. aspera, R. fasciata and R. vertagus was separated into recruits and adults. Size of recruits was determined by plotting size-frequency histograms for each species. A definite year class (representing the 1987 and 1988 0-year class recruits) was apparent for each species and the upper size limit of this cohort was used to define the sizes of recruits (Fig. 2).
To determine the intensity of recruitment of Rhinoclavis across the lagoon in the absence of resident infauna, I established experimental enclosures filled with defaunated sediment at 12 locations on the southern sandflat and in the lagoon. Six locations were on the southern sandflat and two were in each of the other three habitats. All macro- fauna > 1 mm in size were sieved from sediment collected in situ at each location. The sediment was sieved directly into small plastic tubs (internal dimensions 394 x 284 x 108 mm) which were used as the enclosures. The tubs had an out-turned lip around the top edges. 10 large holes (7 cm diameter) were drilled at equally spaced intervals around the midline of the sides and covered with 1-mm nylon mesh glued on with contact adhesive to allow water to flow through the tubs.
At each location, sediments were excavated to a depth of ~ 60-70 mm and the tubs placed so that the sides protruded 38-48 mm above the surface of the substratum forming a fence enclosing the sediment contained within the tubs. Usually several centimetres of the hard coral base under the sediment were removed with a cold-chisel and lump hammer allowing the tubs to be securely wedged into the hole. The tops of the tubs were left open.
Statistical analyses. Spatial and temporal variation in the total abundance and the abundance of recruits of each species were analysed using three factor, mixed model, nested ANOVA with factors Time (fixed), Locations (fixed) and Sites (nested within

RECRUITMENT OF CERITHIID GASTROPODS 5
1987 R_as_..~.e~ 1988
°°tl '°° , 200- 2 0 0 k ~ a j B I ~ , ~
2 4 6 8 10 12 14 2 4 6 8 10 12 14 R_ fasciata
Z u.I 0 uJ ri- LL
203040 110 .,, .,,ill 2 4 6 8 10 12 14
18o] 1 0 0 - ~
50-
2 4 6 8 10 12 14 R. vertagus
'°° ,, i= m m E ==mlm
2 4 6 8 10 12 14
200]
li 1 O01
% L . . . . . . . . 2 4 6 8 10 12 14
MAXIMAL WIDTH OF SHELL (mm) Fig. 2. Size-frequency distributions for R. aspera, R.fasciata and R. vertagus in January 1987 and January 1988 showing presence of two well-defined size classes in each year. Animals within smaller size class were
defined as recruits. Data were pooled from all samples collected on each occasion.
Times x Locationsmrandom). Data were transformed to ln(x + 1) before analysis to conform to assumptions of homoscedasticity (Cochran's test, 0c = 0.05, Underwood, 1979). Ryan's Q test (Day & Quinn, 1989) was used to compare means found to be significantly different in the preceding ANOVA. x _+ SE values were backtransformed for presentation in the figures.
The number of Rhinoclavis spp. recruits in the "defaunated" sediment was compared with the number of recruits in undisturbed control sediment surrounding the tubs. Abundances of recruits in the control sediment were estimated using the data from the 225-cm 2 cores. Estimates of densities of recruits were, therefore, based on 0.0225 m 2 for the controls, and 0.119 m 2 for the defaunated sediments" the former were multiplied by 5.29 to standardize the area for comparison. Means for the two treatments were compared using a two-tailed t test after homogeneity of variances was tested with an F ratio test.
6 G.A. SKILLETER
RESULTS
SPATIAL AND TEMPORAL VARIATION IN ABUNDANCE
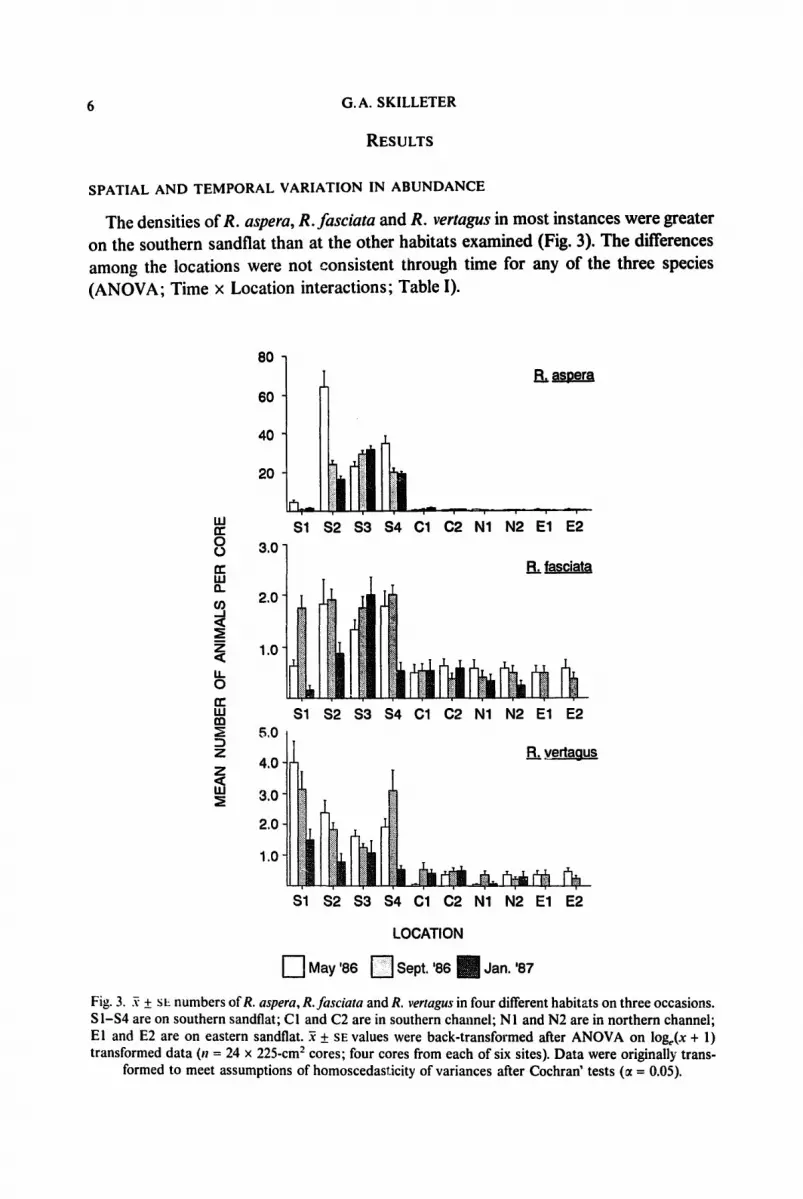
The densities of R. aspera, R. fasciata and R. vertagus in most instances were greater on the southern sandflat than at the other habitats examined (Fig. 3). The differences among the locations were not consistent through time for any of the three species (ANOVA; Time x Location interactions; Table I).
80
60
40
20
LU n" O O 3.0 n" uJ n
2,0 _1 <l:
z 1.0 < LL 0 n" LU m
5,0 Z z 4.0 < LU 3.0
2.0
1.0
R. asoera
$1 $2 $3 $4 C1 C2 N1 - -
, , ,
N2 E1 E2
$1 $2 $3
R. fasciata
$4 C1 C2 N1 N2 E1 E2
R__ yertagus
$1 $2 $3 $4 C1 C2 N1 N2 E1 E2
LOCATION
1-7 May '86 ~ Sept. '86 ~ Jan. '87
Fig. 3. ~ + sL numbers ofR. aspera, R.fasciata and R. vertagus in four different habitats on three occasions. S I - $ 4 are on southern sandflat; CI and C2 are in southern channel; NI and N2 are in northern channel; El and E2 are on eastern sandflat. ~ + SE values were back-transformed after ANOVA on loge(x + 1) transformed data (!1 = 24 x 225-cm 2 cores; four cores from each of six sites). Data were originally trans-
formed to meet assumptions of homoscedasT.icity of variances after Cochran' tests (~ = 0.05).

RECRUITMENT OF CERITHIID GASTROPODS 7
TABLE I
Results of three-factor ANOVA comparing densities of R. aspera, R. fasciata and R. vertagus for three different times at different locations and sites within locations. Time and Location were fixed factors; Sites were nested within Years x Locations and were random. Data were transformed to ln(x + 1) to meet assumptions of homoscedasticity after Cochran's tests (a = 0.05). F values are shown. In this and all
subsequent tables: * significant, P < 0.05; ** significant, P < 0.01.
Source df R. aspera R. fasciata R. vertagus
Time: T 2 5.13"* 15.68"* 9.65"* Location: L 9 275.78"* 21.53"* 34.71 ** T x L 18 4.00"* 3.78 ** 2.02" Sites (T x L) 150 3.08** 1.35"* 2.19"* Residual 540
In May 1986 all four locations on the southern sandflat had significantly greater densities ofR. asperathan at the other habitats but in September 1986 and January 1987 only three of the four locations on the southern sandflat had greater densities (Ryan's Q tests; Table II). In May 1986, three out of four locations on the southern sandflat had greater densities of R. fasciata than elsewhere and in September 1986 all four locations had greater densities. In January 1987, only one of the four locations on the southern sandflat had greater densities than at other habitats (Ryan's Q tests; Table II). For R. vertagus in May and September 1986, all four locations on the southern sandflat had greater densities than elsewhere but in January 1987 there were no significant differences among any of the habitats (Ryan's Q tests; Table II). The densities of each of the three species also varied significantly at the smaller spatial scale of sites (ANOVA; Table I).
SPATIAL AND TEMPORAL VARIATION IN RECRUITMENT
The number of recruits of R. aspera varied among the different locations sampled but differences were not the same for the two summers examined (ANOVA, Time × Loca- tion interaction; Table 111). In January 1986, two of the four locations on the sandflat had significantly more recruits ofR. aspera than the other three habitats (Ryan's Q tests; Table IV; Fig. 4). The third location on the sandflat also had more recruits than elsewhere except for one location in the other habitats, whereas the number of recruits at the fourth location on the sandflat was not significantly different from the other habitats (Table IV). In January 1987, however, all four locations on the sandflat had greater numbers of recruits than the other three habitats (Ryan's Q tests; Table IV). The number of recruits of R. aspera also varied significantly at the smaller spatial scale of sites within each location (Table III).
For R. fasciata and R. vertagus the results of the analyses were very similar. Each species showed a significant Time x Location interaction (ANOVA; Table lll) indicating that differences among locations were not the same between the 2 yr. In January 1987, there were no significant differences in the number ofrecruits ofR. fasciata

8 G.A. SKILLETER
TABLE I I
Results of Ryan's Q tests for differences in mean number of adults at each location at each time. Means are for data (mean number of snails per 225-cm 2 core) transformed to ln(x + 1). Means underlined by same line are not significantly different (P > 0.05). CI and C2 are locations in southern channel, NI and N2 are
in northern channel, E1 and E2 are on eastern sandflat and SI-$4 are on southern sandflat.
R. aspera May 1986 CI C2 El N2 NI E2 SI $3 $4 $2
0.23 0.26 0.29 0.30 0.34 0.36 1.30 3.09 3.45 3.93
Sep 1986 E2 El NI N2 C2 CI SI $4 $2 $3
0.20 0.26 0.28 0.28 0.41 0.42 0.44 2.91 3.12 3.36
Jan 1987 N2 N! El E2 C2 CI SI $2 $4 $3
0.07 0.09 0.09 0.10 0.43 0.71 0.83 2.77 2.92 3.41
R. fasciata May 1986
Sep 1986
Jan 1987
R. vertagus May 1986
Sep 1986
Jan 1987
C 1 E 1 N 1 E2 N2 C2 S I $3 $2 $4 0.31 0.32 0.35 0.36 0.39 0.41 0.42 0.76 0.83 0.92
E2 C2 NI N2 El CI SI $3 S2 $4 0.25 0.26 0.28 0.32 0.32 0.35 0.93 0.93 1.02 i.05
El E2 SI N2 NI CI $4 C2 S2 S3 0.00 0.00 0. ! 2 0.17 0.2 i 0.29 0.33 0.37 0.50 0.92
CI NI El N2 C2 E2 $3 $4 S2 Sl 0.03 0.03 0.25 0.26 0.26 0.30 0.91 0.97 i.08 1.43
N2 E2 El NI C! C2 $3 $2 S4 Sl 0.14 0.17 0.24 0.25 0.29 0.30 0.77 0.98 1.15 1.27
~ e , .
El E2 NI N2 CI C2 $4 S2 $3 SI 0.00 0.00 0.06 0.15 0.26 0.31 0.36 0.42 0.47 0.72
or R. ~ertagus anaong any of the locations (Ryan's Q tests; Table IV; Fig. 4). In January 1988, for each of the species, there were significantly more recruits at three of the four locations on the sandflat compared with the locations in the other h~bitats. There were no significant differences in the number of recruits of R..fasciata or R. vertag,q',; among

RECRUITMENT OF CERITHIID GASTROPODS 9
TABLE I11
Results of three-factor ANOVA comparing densities of R. aspera, R. fasc&ta and R. vertagus recruits in 225-cm 2 cores in 1987 and 1988. Time and Location were fixed factors; Sites were nested within Years x Lo- cations and were random. Data were transformed to loge(x + 1) to meet assumptions of homoscedasticity
after Cochran's tests (oc = 0.05). F values are shown. Significance symbols as in Table I.
Source df R. aspera R. fasciata R. vertagus
Time: T 1 50.04"* 24.93"* 5.93" Location: L 9 61.69 ** 8.93 ** 11.14"* T x L 9 22.12"* 6.95** 3.59** Sites (T x L) 100 2.46** 0.79Ns 2.85** Residual 360
TABLE IV
Results of Ryan's Q tests for differences in mean number of recruits at each location at each time. Means joined by same line are not significantly different (P > 0.05). C1 and C2 are locations in southern channel, Nl and N2 are in northern channel, El and E2 are on eastern sandflat and S1-$4 are on southern
sandflat.
R. aspera Jan 1987 E2 Nl El C1 N2 Sl C2 $4 $2 $3
0.06 0.07 0.09 0.24 0.30 0.40 0.48 0.96 1.29 2.07
Jan 1988 E2 El Nl Cl C2 N2 $3 Sl $4 $2
0.06 0.09 0.09 0.13 0.20 0.39 1.44 2.75 2.82 2.97
R.fasciata Jan 1987 El E2 N2 Sl $4 $3 N1 Cl $2 C2
0.00 0.00 0.00 0.00 0.06 0.09 0.12 0.12 0.14 0.17
Jan 1988 El E2 N2 C2 Nl Cl $3 $4 Sl $2
0.06 0.09 0.09 0.13 0.20 0.39 1.44 2.75 2.82 2.97
R. vertagus Jan 1987 E2 E2 $3 Nl Cl Sl N2 C2 $4 $2
0.00 0.00 0.00 0.03 0.03 0.03 0.09 0.10 0.34 0.50
Jan 1988 El E2 NI N2 CI C2 $3 $4 $2 SI
0.00 0.00 0.00 0.00 0.00 0.00 0.26 0.42 0.69 0.73

10 G.A. SKILLETER
20
15
10
R_ s~..~p.~._~ [ ] 1987 [ ] 1988
n- O
$1 $2 $3 $4 Cl C2 N1 N2 E1 E2 1.0
°" t i ~ 08 R. fasciata
" 0,6
~ 0.4
~ 0.2 .= :::} $1 $2 $3 $4 Cl C2 N1 N2 E1 E2 z 1.5 z 1 < R . ~ W
1.0. i
'i 1 0.5
E~I $2 $3 S'4 C1 C2 N1 N2 E1 E2
LOCATION Fig. 4. ~ + SE number of recruits ofR. aspera, R.fasciata and R. vertagus in four different habitats in January 1987 and January 1988. *Indicates that samples were collected but no recruits were found. Other details
as in Fig. 3.
the locations in the other three habitats (Ryan's Q tests; Table IV). The number of recruits of R.fasciata did not vary significantly at the smaller spatial scale within locations, but there was significant variation in the abundance of recruits ofR. vertagus at this scale (ANOVA; Table III).
The abundance of recruits of each of the species was positively correlated with the abundance of the recruits of the other two species in 1987 and 1988 (Spearman's rank correlations; Table Va) indicating a significant congruence in the patterns of recruitment of Rhinoclavis spp. among the three habitats examined: samples which had greater numbers of one species generally had greater numbers of the other two. For each of the three species, the number of recruits was also positively correlated with the abundance of adults in 1987 and 1988 (Spearman's rank correlations; Table Vb). Variation in the

RECRUITMENT OF CERITHIID GASTROPODS 11
TABLE V
(a) Spearman's rank correlations of number of recruits of R. aspera, R.fasciata and R. vertagus in each sample in 1987 and 1988. Each correlation was tested at P = 0.02 probability level to maintain experiment- wise Type I error rate at P = 0.05 (Bonferroni correction- Snedecor & Cochran, 1980). (b) Spearman's rank correlations of number of recruits vs. number of adults for R. aspera, R.fasciata, R. vertagus in each sample in 1987 and 1988. Values for correlation coefficient rs and associated probability level are shown (n = 240
for each correlation).
a r s P
R. aspera vs. R. fasciata 1987 0.487 <0.01 1988 0.616 <0.01
R. aspera vs. R. vertagus 1987 0.556 <0.01 1988 0.721 <0.01
R. fasciata vs. R. vertagus 1987 0.738 <0.01 1988 0.697 <0.01
b rs P
R. aspera 1987 0.650 <0.01 1988 0.647 <0.01
R. fasciata 1987 0.616 <0.01 1988 0.417 <0.01
R. vertagus 1987 0.616 <0.01 1988 0.542 < 0.01
number of adults of a species was related to 22-42% of the numerical variation of recruits of that species.
RECRUITMENT IN ABSENCE OF OTHER MOLLUSCS
Recruitment of R. aspera to the "defaunated" sediment in the experimental tubs varied among the different locations, but the differences were not consistent between the years (ANOVA; Y x L interaction; Table VI; Fig. 5A). In 1987, only two of the six locations on the sandflat had significantly more recruits in the tubs than in other habitats, whereas in 1988, tubs at five ofthe six locations on the sandflat had significantly more recruits than tubs at the other locations (Ryan's Q tests; Table VII). There were significant differences in the number of recruits in tubs at the smaller spatial scale (Table VI).
Recruitment of R. fasciata to the experimental tubs was sparse in both years (Fig. 5B). There were significant differences among locations (ANOVA; Table Vl) although

12 G.A. SKILLETER
TABLE VI
Results of three-factor ANOVA comparing numbers of recruits of R. aspera, R. fasciata, R. vertagus and Cerithium zonatum in experimental tubs at two sites within each of 10 locations in each of 2 yr. Year and Location were fixed factors; Sites were nested within Years x Locations and were random. Data were transformed to loge(x + 1) to meet assumptions of homoscedasticity aRer Cochran's tests (= = 0.05). F
values are shown. Significance symbols as in Table I.
Source df R. aspera R. fasciata R. vertagus C. zonatum
Year: Y 1 1.69 Ns 7.57 * 2.'/1 NS 6.33 * Location: L 9 14.34"* 2.86" 4.58"* 128.96"* Y x L 9 4.56** 2.08Ns 3.95** 3.76** Sites (Y x L) 20 3.11 ** 0.72NS 1.04NS 0.96NS Residual 40
El
1 0 0
751
r r 50. u.I n
25. n- O W
tL 0 r r 10' uJ m :~ 8 :3 Z 6 Z <I:
4'
2'
(A) 2 . 0
1,0
1 2 3 4 5 6 C1 C2 N1 N2
(c)
1 2 3 4 5 6 C1 C2 N1 N2
1,0'
0.8
0 .6
0 .4
1 2 3 4 5 6 C l C 2 N I N 2 1987 1988
2so (D) 200
150
!~:.-','~ ~,~,:. , ~ . . . . ~ .,~_~.'..',- ~ : - 1 2 3 4 5 6 C1 C2 N1 N2
LOCATION
Fig. 5. ~ + SE numbers of recruits of Rhinoclavis spp. in experimental tubs at 10 locations in three habitats in January 1987 and January 1988. (A) R. aspera; (B) R.fasciata; (C) R. vertagus; (D) C. zonatum. For R. aspera, R. vertagus and C. zonatum, n = four tubs (two tubs from each of two sites). For R.fasciata, (i) n = eight tubs (two tubs from each of two sites in 2 yr), (ii) n = 40 tubs (two tubs from each of two sites
at 10 locations). Other details as in Fig. 3.

RECRUITMENT OF CERITHIID GASTROPODS 13
TABLE VII
Results of Ryan's Q tests for differences in mean number of :ec;ruits in experimental tubs excluding molluscan macrofauna. Means are for data transformed to logflx + 1). Means underl:,a:d by same line are not significantly different (P > 0.05). C 1 and C2 are locations in southern channel, N 1 and N2 are in northern channel, and 1-6 are on southern sandflat (1 and 2 are close to reef crest, 3 and 4 are midway across sandflat,
5 and 6 are close to edge of sandflat).
R. aspera 1987 N ! N2 2 6 C 1 C2 1 4 3 5
0.68 0.75 1.67 1.68 3.04 3.07 3.35 3.87 4.43 4.61
1988 NI CI N2 C2 1 3 2 4 6 5 0.69 !.25 1.37 !.49 1.64 3.12 3.19 3.61 3.67 4.51
R. fasciata Locations N2 2 1 C2 NI 6 4 3 5 CI
0.17 0.17 0.20 0.31 0.35 0.48 0.51 0.62 0.76 0.91
Years 1988 1987 0.32 0.58
R. vertagus 1987 NI N2 2 5 1 C2 6 CI 4 3
0.00 0.28 0.52 0.87 1.14 !.24 1.31 1.37 1.49 1.56
1988 NI C2 CI 4 1 N2 3 6 2 5 0.17 0.35 0.97 1.18 1.25 1.27 1.31 1.37 1.67 2.35
C. 2oHa!um 1987 N 1 2 1 N2 C2 3 CI 4 6 5
0.00 0.00 0.00 0. ! 7 0.35 0.45 0.90 1.94 2.08 5.34
1988 N2 2 1 NI CI 4 3 C2 6 5 0.00 0.00 0.00 0.17 0.17 0.52 0.72 0.75 1.69 4.98
Ryan's Q tests could not detect which locations were different from each other (Table VII). Recruitment was significantly greater in 1987 than in 1988 (Table VII; Fig. 5B). There were no significant differences among different sites within a location (Table VI).
Recruitment of R. vertagus to the defaunated sediment in the experimental tubs varied among different locations, but the differences were not consistent between the 2 yr (ANOVA; Y x L interaction; Table VI; Fig. 5C). Tubs at locations on the sandflat did not receive greater numbers of recruits than tubs in other habitats of the lagoon (Ryan's Q tests; Table VIII). There were no significant differences in the number of recruits among sites within a location (Table VI).

14 G.A. SKILLETER
Another species of cerithiid gastropod, Cerithium zonatum (Wood, 1828), occurred in large numbers at a few of the locations (Fig. 5D). The densities at one location on the sandflat were significantly greater (up to 225 times) than at any other location on the sandflat or in the other habitats examined in 1987 and 1988 (ANOVA; Table VII; Ryan's Q tests; Table VII). There were no significant differences in the number of recruits among the sites within the locations (Table VI).
Striking differences were evident when the numbers of recruits recorded at each location were compared between the defaunated sediments from the experimental tubs and the undisturbed (control) sediments surrounding the tubs. For R. aspera in 1987, there were significantly more recruits per unit area in the defaunated sediments compared with the control sediments at five of the 10 locations (t tests; P < 0.05; Table VIII). At one location on the sandflat, however, the reverse was true with significantly more recruits per unit area occurring in the control sediment than in the defaunated sediment (Location 6; t test; Table VIII). In 1988, three out of the
TABLE Vlll
Results oft tests comparing number of recruits ofR. aspera in defaunated sediment from experimental tubs and undisturbed "control" sediment surrounding tubs. Means are for a standardized area of0.119 m 2 (area
of an experimental tub).
Location Year Control (2) Tubs (2) t P
1 1987 3.97 36.75" 5.09 P < 0.001 1988 10.36 4.25 1.23 0.20 < P < 0.40
2 1987 3.53 9.00 1.80 0.05 < P < 0.10 1988 98.07 24.75 1.91 0.05 < P < 0.10
3 1987 19.17 87.00* 6.98 P < 0.001 1988 173.66" 23.75 4.14 P < 0.001
4 1987 10.80 58.50* 5.87 P < 0.001 1988 101.59" 36.25 2.12 P < 0.05
5 1987 ! 1.90 102.25" 10.80 P < 0.001 1988 30.63" 90.75 4.27 P < 0.001
6 1987 55.54* 8.75 3.45 P < 0.005 1988 28.43 39.75 0.72 0.40 < P < 0.50
C! 1987 1.98 20.25 9.67 P < 0.001 1988 i.10 2.75 !.18 0.20 < P < 0.40
C2 1987 1.98 20.25" 9.67 P < 0.001 1988 1.10 2.75 1.18 0.20 < P < 0.40
NI 1987 0.66 1.50 0.67 P > 0.50 1988 0.66 1.00 0.37 P > 0.50
N2 1987 2.87 1.50 0.67 P > 0.50 1988 3.53 4.75 0.28 P > 0.50
* Denotes mean for that treatment significantly greater than other treatment.

RECRUITMENT OF CERITHIID GASTROPODS 15
10 locations had significantly more recruits in the control sediments compared with the defaunated sediments (t tests; Table VIII). There were no significant differences among the other seven locations (Table VIII).
Removal of macrofauna from the sediments had little effect on the recruitment of R. fasciata. In 1987, three out of the 10 locations had significantly more recruits in the defaunated sediment than in the control sediment (ttests; Table IX) but these
TABLE IX
Results oft tests comparing number of recruits of R.fasciata in defaunated sediment from experimental tubs and undisturbed "control" sediment surrounding tubs. Other details as in Table IX.
Location Year Control (.~) Tubs (~) t P
1 1987 0.00 ! .00" 3.85 P < 0.001 1988 !.98 0.00 1.14 0.10 ~ P < 0.20
2 1987 0.00 0.75" 2.73 P < 0.025 1988 4.19 0.00 1.33 0.10 < P < 0.20
3 1987 1.10 2.00 0.76 P > 0.50 1988 5.51 0.50 1.66 0.10 < P < 0.20
4 1987 0.44 i.75 1.60 0.10 < P < 0.20 1988 2.64 1.50 0.65 P > 0.50
5 1987 1.76 1.50 0.86 0.20 < P < 0.40 1988 1.32 1.50 0.12 P > 0.50
6 1987 0.66 0.25 0.45 P > 0.50 1988 2.20 1.25 0.49 P > 0.50
CI 1987 0.88 2.75 1.71 0.05 < P < 0.10 1988 0.66 1.00 0.37 P > 0.05
C2 1987 1.32 0.75 0.48 P > 0.50 1988 0.44 0 25 0.25 P > 0.50
NI 1987 0.88 1.00 0.12 P > 0.50 1988 0.66 0.00 0.51 P > 0.50
N2 1987 0.00 0.25" 2.72 P > 0.50 1988 0.44 0.25 0.25 P > 0.50
differences were based on small numbers of recruits in either treatment. In 1988, there were no significant differences between treatments at any of the locations. The numbers of recruits of R. vertagus in the tubs (defaunated sediment) and control sediment were similar at most locations but where there were significant differences between the two treatments more recruits occurred in the defaunated sediment than in the control sediments (t tests; Table X). Where there were significant differences in the numbers of recruits of C. zonatum between control and defaunated sediments, their numbers were greater in the defaunated treatment (t tests; Table XI).

16 G.A. SKILLETER
TABLE X
Results of t tests comparing number of recruits of R. vertagus in defaunated sediment from experimental tubs and undisturbed "control" sediment surounding tubs. Other details as in Table IX.
Location Year Control (~) Tubs (R) t P
1 1987 0.22 3.25" 3.69 P < 0.005 1988 4.41 2.75 0.67 P > 0.50
2 1987 0.22 1.00 1.28 0.20 < P < 0.40 1988 9.04 4.50 0.77 0.40 < P < 0.50
3 1987 4.63 4.50 0.05 P > 0.50 1988 6.39 3.00 1.28 0.20 < P < 0.40
4 1987 2.87 4.75 0.93 0.20 < P < 0.40 1988 4.19 2.75 0.45 P > 0.50
5 1987 3.97 2.25 0.61 P > 0.50 1988 7.49 9.75 0.67 P > 0.50
6 1987 0.00 3.25" 6.74 P < 0.001 1988 2.42 3.00 0.23 P > 0.50
CI 1987 0.22 3.00* 4.89 P < 0.001 1988 0.00 1.75" 9.97 P < 0.001
C2 1987 0.88 3.25 1.73 0.05 < P < 0.10 1988 0.00 0.50* 4.72 P < 0.001
N 1 1987 0.22 0.00 0.38 P > 0.50 1988 0.00 0.25" 2.73 P < 0.025
N2 1987 0.66 0.50 0.17 P > 0.50 1988 0.00 2.75* 11.91 P < 0.001
TABI.E XI
Results of t tests comparing number of recruits of C. zonatum in defaunated sediment from experimental tubs and undisturbed "control" sediment surrounding tubs. Only those locations where mean abundance
of recruits in either control sediment or tubs were > 1 are shown. Other details as in Table IX.
Location Year Control (~) Tubs (~) t P
3 1987 0.66 0.75 0.05 > 0.50 1988 3.53 1.25 0.46 > 0.50
4 1987 0.00 6.25 * 15.37 < 0.001 1988 0.00 1.00" 3.86 <0.001
5 1987 0.44 212.50" 22.74 < 0.001 1988 140.60 149.00 0.13 > 0.50
6 1987 1.76 8.5" 2.41 < 0.025 1988 0.00 4.50" 24.53 < 0.001

RECRUITMENT OF CERITHIID GASTROPODS 17
DISCUSSION
Initial surveys in October 1985 of infaunal molluscs in sediments at One Tree Reef showed that deposit-feeding cerithiid gastropods were most abundant in the coarse sandy sediments on a shallow sandflat, rather than in fine sediments of the lagoon (Jones et al., 1990; Skilleter, 1991). The data presented here show that this pattern was maintained through time and that Rhinoclavis aspera, R. fasciata and R. vertagus are found in their greatest densities in sandy sediments on the southern sandflat at One Tree Reef. Densities in the finer sediments of the lagoon never reached the maximal densities observed on the sandflat during the course of this stud).
Deposit-feeding infauna are usually reported to be associated with habitats dominated by fine sediments (e.g. Sanders, 1958, 1960; Rhoads & Young, 1970; Levinton, 1972; Ho!land & Polgar, 1976; Whitlatch, 1977, 198 l). This pattern is usually explained by the greater availability of organic material in finer sediments (Sancters, 1958; Newell, 1965; Dale, 1974; Tunnicliffe & Risk, 1977). The occurrence of Rhino- clavis spp. in an "atypical" habitat provides an exception (sze also Dobbs & Whitlatch, ~ 1982; Bianchi, 1988; Kemp, 1988) and illustrates the problems of unjustifiably "fittiing" existing paradigms to new systems without due caution (Underwood & Denley, 1984).
Rhinoclavis aspera, R. fasciata and R. vertagus each showed considerable variation in the intensity of recruitment a~ spatial scales from a few metres to hundreds of metres, and between the 2 yr examined. Spatial and temporal variability in recruitment is a common feature of the demography of many benthic marine organisms from habitats in temperate regions (rocky intertidal habitats: Underwood, 1979; Underwood & Denley, 1984; Connell, 1985; Caffey, 1985; Gaines et al., 1985; subtidal hard substrata: Sutherland & Karlson, 1977; Dean & Hurd, 1980; Keough, 1984; Butler, 1986; soft sediments: Eagle, 1975; Beukema, 1982; Vahl, 1982; Levin, 1984; Bachelet, 1986; Holland et al., 1987) but there have been few studies which have attempted to document such patterns in the tropics (but see Doherty & Williams, 1988, for a thorough review of patterns of recruitment of fishes on coral reefs).
The greatest densities of recruits of Rhinoclavis spp. were found at locations on the southern sandflat, the habitat which supported greater densities of adults. This was evident for R. aspera where differences in recruitment were tenfold among habitats in each of the years examined, but was also noticeable for R. fasciata and R. vertagus in the second year. The smaller densities of Rhinoclavis spp. in the lagoonal sediments compared with the southern sandflat may be partly explained by reduced recruitment to these habitats.
One factor which can lead to variability in recruitment of infauna among different habitats is variation in the availability of larvae for settlement (Hannah, 1981; Levin, i 984, 1986). The direction of water flow on the southern side of One Tree Reef at all stages of the tide is away from the carbonate reef crest and towards the lagoon (Davies & West, 1981). Larvae entrained in the water entering the reef from the southern side, therefore, first pass over the sandflat before reaching the finer sediments of the lagoon.

18 G.A. SKILLETER
If most of these larvae settle on the sandflat then fewer larvae will be available to settle in the lagoon and recruitment will be lower as was observed. This was the pattern observed. Models based on "larval depletion" within the water column have been used to explain patterns of recruitment in other benthic habitats (Gaines et al., 1985; Gaines & Roughgarden, 1985; Jackson, 1986).
Removing adult molluscs from the sediment had unpredictable results on the recruit- ment of the different species of Rhinoclavis. At some locations, there were more recruits of R. aspera in the "defaunated" sediment than the unmanipulated control sediments, but at other locations the reverse was true or there were no differences between the two treatments. Moreover, at some locations the opposite pattern occurred during the second year and more recruits occurred in the control than in the defaunated sediments. In contrast, for R. fasciata and R. vertagus there was either no difference between the defaunated and control sediments or recruitment was greater in the defaunated sedi- ments. The cerithiid C. zonatum also showed significant increases in recruitment in defaunated sediment at some locations but not at others.
The comparisons of the defa-anated sediments with the control sediments are confounded by the presence of the tubs and possibly also by the disruption of the sediments during the removal of macrofauna. These artifacts should have been constant among locations and could not explain the highly variable effects of removing macro- fauna on the recruitment of Rhinoclavis spp.
The results of this experiment gave no conclusive evidence that the presence of adults can significantly affect the recruitment of Rhinoclavis spp.: the effects were variable and complicated. If only some of the locations had been used, or the experiment had only been done in 1 yr, then the conclusions could have supported models of: (i) no effects of adult molluscs on recruitment; (ii) enhancement of recruitment due to the presence of adults or; (iii) reduction of recruitment due to the presence of adults. This obviously makes generalisations about the effects of adults on recruitment of Rhinoclavis impossi- ble at this time, but highlights the necessity of testing how ecological processes vary among a number ofdifferent sites (Dayton, 1971; Underwood et al., 1983; Caffey, 1985; Peterson & Beai, 1989).
One possible explanation for the variability in the effect of removing macrofauna on the recruitment of Rhinoclavis lies in the fact that the "control" sediments among the different locations varied in the density of resident molluscs. If the recruitment of Rhinoclavis to the control sediment was dependent on the density of resident macro- fauna, but there were nonlinearities in the intensity of the interactions among recruits and adults (Underwood et al., 1983), then one might expect different results for compa- risons made at different locations between defaunated sediments (which were equivalent at all locations) and control sediments (which varied among the locations). For this reason experiments examining interactions among recruits and resident fauna should be done at a range of natural starting densities rather than using the "all or none" approach used here (Underwood & Denley, 1984; Underwood & Fairweather, 1989).
The results of this experiment provide evidence against the model of "larval de-

RECRUITMENT OF CERITHIID GASTROPODS 19
pletion" put forward earlier to explain reduced patterns of recruitment in the lagoon. For R. aspera at one location in the south channel, recruitment was significantly increased with the removal of adults from the sediment. Even if the increase in the number of recruits were not due to the absence of the adult molluscs but due to an artifact of the experiment, the fact remains that there were still abundant larvae of R. aspera left in the water over the sediments of the south channel as evidenced by the increase in the number of recruits during the experiment in that habitat. Similarly, recruitment of R. vertagus was increased at several locations in the lagoon by the removal of adults, although the increase in density of recruits was not as marked as for R. aspera. Clearly, models based on larval depletion and interactions between recruits and resident adults are insufficient by themselves to explain the patterns of reduced recruitment of Rhinoclavis in the finer lagoonal sediments.
In conclusion, the patterns of recruitment of Rhinoclavis can be used to explain the greater abundance of these gastropods in coarse sandy sediments on a shallow sandflat at One Tree Reef, a habitat usually considered atypical for deposit-feeding infauna. The patterns of recruitment, however, could not be fully explained by models of larval depletion as water passed first over the sandflat or interactions between recruits and resident adult macrofauna. Other factors, such as the effects of habitat composition on recruitment, are also probably responsible for the greater abundance of recruits on the sandflat compared with the lagoon.
ACKNOWLEDGEMENTS
This study was supported by a Commonwealth Postgraduate Research Award and grants from the Australian Museum Postgraduate Assistance Scheme, Australian Coral Reef Society, Linnean Society of new South Wales, and the Institute of Marine Ecology, University of Sydney I thank J. Delporte and D. Cable for assistance in the field and E.A. Irlandi, C.H. Peterson, K.L. Skilleter, A.J. Underwood and two anonymous reviewers for helpful comments on an early draft of this manuscript.
REFERENCES
Aller, R.C. & R.E. Dodge, 1974. Animal-sediment relations in a tropical lagoon, Discovery Bay, Jamaica. J. Mar. Res., Vol. 32, pp. 209-232.
Bachelet, G., 1986. Recruitment and year-to-year variability in a population of Macoma balthica (L.). Hydrobiologia, Vol. 142, pp. 233-248.
Beukema, J.J., 1982. Annual variation in reproductive success and biomass of the major macrozoobenthic species living in a tidal flat a~ea of the Wadden Sea. Neth. J. Sea Res., Vol. 16, pp. 37-45.
Bianchi, T.S., 1988. Feeding ecology of subsurface deposit-feeder Leitoscoloplos fragilis Verrill. I. Mechanisms affecting particle availability on intertidal sandflat. J. Exp. Mar. Biol. Ecol., Vol. 115, pp. 79-97.
Butler, A.J., 1986. Recruitment of sessile invertebrates at five sites in Gulf St. Vincent, South Australia. J. Exp. Mar. Biol. Ecol., Vol. 97, pp. 13-36.

20 G.A. SKILLETER
Caffey, H.M., 1985. Spatial and temporal variation in settlement and recruitment of intertidal barnacles. Ecol. Monogr., Vol. 55, pp. 313-332.
Commito, J.A. & E.M. Boncavage, 1989. Suspension-feeders and coexisting infauna: an enhancement counterexample. J. Exp. Mar. Biol. Ecol., Vol. 125, pp. 33-42.
Connell, J. H., 1985. The consequences of variation in initial settlement vs. post-settlement mortality in rocky intertidal communities. J. Exp. Mar. Biol. Ecol., Vol. 93, pp. 11-45.
Crowe, W. A., A. B. Josefson & I. Svane, 1987. Influence of adult density on recruitment into soft sediments: a short term in situ experiment. Mar. Ecol. Prog. Set., Vol. 4, pp. 61-69.
Dale, N.G., 1974. Bacteria in intertidal sediments: factors related to their distribution. Limnol. Oceanogr., Vol. 19, pp. 509-518.
Davies, P.J., B.M. Radke & C. R. Robison, 19"/6. The evolution of One Tree Reef, Southern Great Barrier Reef, Queensland. BMR J. Aust. Geol. Geophys., Vol. i, pp. 231-240.
Davies, P.J. & B.G. West, 1981. Suspended-sediment transport and water movement at One Tree Reef, southern Great Barrier Reef. BMR J. Aust. Geol. Geophys., Vol. 6, pp. 187-195.
Day, R.W. & G.P. Quinn, 1989. Comparisons of treatments after an analysis ofvariance in ecology. Ecol. Monogr., Vol. 59, pp. 433-463.
Dayton, P.K., 1971. Competition, disturbance and community organization: the provision and utilization of space in rocky intertidal community. Ecol. Monogr., Vol. 41, pp. 351-389.
Dean, T.A. & L.E. Hurd, 1980. Development in an estuarine fouling community: the influence of early colonists on later arrivals. Oecologia, Vol. 46, pp. 295-301.
Dobbs, F.C. & R.B. Whitlatch, 1982. Aspects of deposit-feeding by the polychaete Clymenella torquata. Ophelia, Vol. 21, pp. 159-166.
Doherty, P.J. & D. McB Williams, 1988. The replenishment of coral reeffish populations. Oceanogr. Mar. Biol. Atom. Rev., Vol. 26, pp. 487-551.
Eagle, R.A., 1975. Natural fluctuations in a soft-bottom benthic community. J. Mar. Biol. Assoc. UK., Vol. 55, pp. 865-878.
Gaines, S.D., S. Brown & J. Roughgarden, 1985. Spatial variation in larval concentrations as a cause of spatial variation in settlement of the barnacle, Balanus glandula. Oecologia, Vol. 67, pp. 267-272.
Gaines, S. & J. Roughgarden, 1985. Larval settlement rate: a leading determinant of structure in an ecological community of the marine intertidal zone. Proc. Natl. Acad. Sci. U.S.A., Vol. 82, pp. 3707-3711.
Hannan, C.A., 198 !. Polychaete larval settlement: correspondence of patterns in suspended jar collectors and in the adjacent natural habitat in Monterey Bay, California. Limnol. Oceanogr., Vol. 26, pp. 159-171.
Hines, A. H., M. H. Posey & P.J. Haddon, 1989. Effects ofadult suspension-feeding bivalves on recruitment of estuarine infauna. Veliger, Vol. 32, pp. 109-119.
Holland, A. F. & T.T. Polgar, 1976. Seasonal changes in the structure of an intertidal community. Mar. Biol., Vol. 37, pp. 341-348.
Holland, A.F., A.T. Shaughnessy & M.H. Hiegel, 1987. Long-term variation in mesohaline Chesapeake Bay macrobenthos - spatial and temporal patterns. Estuaries, Vol. 10, pp. 227-245.
Hunt, J. H., W.G. Ambrose & C.H. Peterson, 1987. Effects of the gastropod, llyanassa obsoleta (Say), and the bivalve, Mercenaria mercenaria (L.), on larval settlement and juvenile recruitment of infauna. J. Exp. Mar. Biol. Ecol., Vol. 108, pp. 229-240.
Jackson, G.A., 1986. Interaction of physical and biological processes in the settlement ofplanktonic larvae. Bull. Mar. Sci., Vol. 39, pp. 202-2 ! 2.
Johnstone, R.W., K. Koop & A.W.D. Larkum, 1990. Physical aspects of coral reef lagoon sediments in relation to detritus processing and primary production. Mar. Ecol Prog. Ser., Vol. 66, pp. 273-284.
Jones, G.P., D.J. Ferrell & P.F. Sale, 1990. Spatial pattern in the abundance and structure of mollusc populations in the soft sediments of a coral reef lagoon. Mar. EcoL Prog. Set., Vol. 62, pp. 109-120.
Kemp, P.F., 1988. Production and life history of a deposit-feeding polychaete in an atypical environment. Estuarine Coastal Shelf Sci., Vol. 26, pp. 437-446.
Keough, M.J., 1984. Dynamics of the epifauna of the bivalve Pinna bicolor: interactions among recruitment, predation and competition. Ecology, Vol. 65, pp. 677-688.
Levin, L. A., 1984. Life history and dispersal patterns in a dense infaunal polycllaete assemblage: community structure and response to disturbance. Ecology, Vol. 65, pp. 1185-1200.
Levin, L.A., 1986. The influence of tides on larval availability in shallow waters overlying a mudflat. Bull. Mar. Sci., Vol. 39, pp. 224-233.

RECRUITMENT OF CERITHIID GASTROPODS 21
Levinton, J. S., 1972. Stability and trophic structure in deposit-feeding and suspension-feeding communities. Am. Nat., Vol. 106, pp. 472-486.
Marshall, T. F. & P.J. Davies, 1982. Internal structure and holocene evolution of One Tree Reef, southern Great Barrier Reef. Coral Reefs, Vol. i, pp. 21-28.
Maurer, D., W. Leatham, P. Kinner & J. Tinsman, 1979. Seasonal fluctuations in coastal benthic invertebrate assemblages. Estuarine Coastal Mar. Sci., Vol. 8, pp. 181-193.
Murphy, R.C., 1985. Factors affecting the distribution of the introduced bivalve, Mercenaria mercenaria, in a California lagoon - the importance of bioturbation. J. Mar. Res., Vol. 43, ppo 673-692.
Newell, R., 1965. The role of detritus in the nutrition of two marine deposit feeders, the prosobranch Hydrobia ulvae and the bivalve Macoma balthica. Proc. Zool. Soc. London, Vol. 144, pp. 25-45.
Peterson, C. H., 1977. Competitive organisation of the soft bottom macrobenthic communities of Southern California Lagoons. Mar. Biol., Vol. 43, pp. 343-359.
Peterson, C.H., 1982. The importance of predation and intra- and inter-specific competition in the population biology of two infaunal suspension-feeding bivalves Protothaca staminea and Chione undatella. Ecol. Monogr., Vol. 52, pp. 437-475.
Peterson, C.H. & B.F. Beal, 1989. Bivalve growth and higher order interactions: importance of density, site and time. Ecology, Vol. 70, pp. 1390-1404.
Peterson, C.H. & R. Black, 1987. Resource depletion by active suspension feeders on tidal flats: influence of local density and tidal elevation. Limnoi. Oceanogr., Vol. 32, pp. 143-166.
Rhoads, D.C. & D.K. Young, 1970. The influence of deposit-feeding organisms on sediment stability and community trophic structure, d. Mar. Res., Vol. 28, pp. 150-178.
Sanders, H. L., 1958. Benthic studies in Buzzard Bay. I. Animal-sediment relationships. Limnol. Oceanogr., Vol. 3, pp. 245-253.
Sanders, H.L., 1960. Benthic studies in Buzzard Bay. III. The structure of the soft-bottom community. Limnol. Oceanogr., Vol. 5, pp. 138-153.
Sanders, H.L., E.M. Goudsmit, E.L. Mills & G.E. Hamson, 1962. A study of the intertidal fauna of Barnstable Harbor, Massachusetts. Linmol. Oceanogr., Voi. 7, pp. 63-79.
Skiileter, G.A., 1991. Distribution, abundance and size-structure of cerithiid gastropods in sediments at One Tree Reef, southern Great Barrier Reef. J. Exp. Mar. Biol. Ecol., Vol. 151, pp. 185-207..
Sutherland, J. P. & R.H. Karlson, 1977. Development and stability of the fouling community at Beaufort, North Carolina. Ecol. Monogr., Vol. 47, pp. 425-446.
Tunnicliffe, V. & M.J. Risk, 1977. Relationships between the bivalve Macoma balthica and bacteria in intertidal sediments: Minas Basin, Bay of Fundy. J. Mar. Res., Vol. 35, pp. 499-507.
Underwood, A.J., 1979. The ecology of intertidal gastropods. Adv. Mar. Biol., Vol. 16, pp. 111-210. Underwood A.J. & E.J. Denley, 1984. Paradigms, explanations and generalisations in models for the
structure of intertidal communities on rocky shores. In, Ecological communities: conceptual issues and the evidence, edited by D.J. Strong et al., Princeton University Press, Princeton, pp. 151-180.
Underwood, A.J., E.J. Denley & M.J. Moran, 1983. Experimental analyses of the structure and dynamics of mid-shore rocky intertidal communities in New South Wales. Oecologia, Vol. 56, pp. 202-219.
Underwood, A.J. & P.G. Fairweather, 1989. Supply-side ecology and benthic marine assemblages. Trends EcoL EvoL, Vol. 4, pp. 16-20.
Vahl, O. 1982. Long-term variations in recruitment of the Iceland scallop, Chlamys islandica, from northern Norway. Neth. J. Sea Res., Vol. 16, pp. 80-87.
Whitlatch, R.B., 1977. Seasonal changes in the community structure of the macrobenthos inhabiting the intertidal sand and mudflats of Barnstable Harbour, Massachusetts. Biol. Bull., Vol. 152, pp. 275-294.
Whitlatch, R.B., 1981. Animal-sediment relationships in intertidal marine benthic habitats: some deter- minants of deposit-feeding species diversity. J. Exp. Mar. Biol. EcoL, Vol. 53, pp. 31-45.
Williams, J.G., 1980. The influence of adults on the settlement of the clam, Tapes japonica. J. Mar. Res., Vol. 38, pp. 729-741.
Young, D.K. & D.C. Rhoads, 1971. Animal-sediment relations in Cape Cod Bay, Massachusetts. I. A transect study. Mar. Biol., Vol. 11, pp. 242-254.























![[Terrestrial gastropods from Misiones Province, Argentina]](https://img.pdfslide.net/doc/110x75/635294cfcd28b2b8c2079dae/terrestrial-gastropods-from-misiones-province-argentina.jpg)





