Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Forest Ecology and Management 261 (2011) 949–957
Contents lists available at ScienceDirect
Forest Ecology and Management
journa l homepage: www.e lsev ier .com/ locate / foreco
Plant communities, species diversity, richness, and regeneration of a traditionallymanaged coastal forest, Kenya
Staline Kibet ∗
Coastal Forest Conservation Unit, National Museums of Kenya, P.O. Box 596, Kilifi, Kenya
a r t i c l e i n f o
Article history:Received 12 July 2010Received in revised form22 November 2010Accepted 26 November 2010Available online 17 January 2011
Keywords:Kaya forestsRegenerationBiodiversitySpecies associationCultural sitesConservation
a b s t r a c t
The Kenyan coastal forests make up one of the World 25 Biodiversity Hotspots. They consist of over 140fragments (the majority with areas less than 0.5 km2) of the once extensive Zanzibar–Inhambane lowlandmoist forest. The over 60 known Mijikenda sacred Kaya forests and groves scattered along the coastalhinterland form the greater part of this ecosystem. The forests are of biological and cultural significance,and this has been recognized nationally and internationally, with some now listed as World Heritage Sites.The forests are protected by councils of Kaya elders who regulate use of their resources. Increasing humanpopulation and subsequent rise in demand for forest products and land for settlement has put a strain onthese relic forests. Farm encroachment and extraction of forest products in different Kaya forests haveaffected the vegetation ecology at varying levels. This study investigated the spatial species distribution,association and regeneration potential of commonly utilized plants in one of these traditionally managedecosystems. A modified nested plot method was used to collect data in the field.
Using TWINSPAN multivariate, and indicator species analysis, two plant communities (Asteranthe andBridelia) and an undifferentiated vegetation type were identified. Species association in Asteranthe con-sisted largely of forest dependant species, with a significant presence of woody climbers. It was comprisedof two sub-communities namely Manilkara and Scorodophloeos. In contrast the second plant community,Bridelia, was dominated by light demanding species. It comprised one sub-community (Catunaregam)and a seral stage (Keetia). The species diversity and richness was higher in the Asteranthe communitycompared to Bridelia. Some of the forest species commonly utilized by the local people were observedto regenerate both in open and closed forest habitats while others had seedling recruitment confined toclosed forest.
Despite some coastal forests showing physiognomic similarity, detailed study shows intra-variationlinked to topography, exposition, type and intensity of human perturbation both currently and in thedistant past. Clearly, vegetation patterns of coastal forests of eastern Africa change at fairly short intervals.
Recruitment of forest specialists is likely to decline if closed forests are opened up by farm encroach-ment, however their less specialized counterparts can pioneer in re-colonization of disturbed sites ifconservation is strengthened. There is need to invigorate traditional management systems of forestswith cultural significance by recognizing and giving increased legal mandates to the local custodians.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
The Kenya coastal forest, which constitutes part of the onceextensive Zanzibar–Inhambane lowland moist forests, is one exam-ple of an ecosystem under threat in eastern Africa (Janzen, 1988;White, 1983). It is estimated at 660 km2 (Burgess et al., 2000;Waiyaki, 1995) with a significant number of the fragments havingan area less than 0.5 km2. Many of these fragments are the Mijik-
∗ Current address: East Africa Herbarium/Botany Department, National Museumsof Kenya, P.O. Box 40658-00100, Nairobi, Kenya.Tel.: +254 20 37412131/2/3/4x2205; fax: +254 20 3741424.
E-mail addresses: [email protected], [email protected]
enda sacred Kaya forests: there are over 60 Kaya forest patches.They range in area from as little as 2 ha to over 500 ha and are scat-tered over a distance of 200 km in the administrative Counties ofKwale in the south to Kilifi in the North. They owe their existenceto the culture and beliefs of nine closely related ethnic groups col-lectively called the Mijikenda. Historically, the Kaya forests wereused by these communities as a refuge against aggression fromunfriendly neighbors, so cutting of trees and other form of vegeta-tion destruction inside and around the sites was strictly prohibited(Spear, 1978).
The Kaya forests have gain recognition since the early 1990safter the first comprehensive inventory of coastal vegetation wascarried out (Robertson, 1987; Robertson and Luke, 1993). They havebeen recognized nationally and internationally as important natu-
0378-1127/$ – see front matter © 2010 Elsevier B.V. All rights reserved.doi:10.1016/j.foreco.2010.11.027

Author's personal copy
950 S. Kibet / Forest Ecology and Management 261 (2011) 949–957
ral and cultural landscapes as exemplified by their registration asNational Monuments under Antiquities and Monuments Act (Cap215) laws of Kenya and most recently when several were listed asWorld Heritage Sites (UNESCO, 2008).
Many vegetation types along the Kenyan Coast have beendescribed in varying degrees of details; however few studies havetargeted the sacred Kaya forests (Clarke and Robertson, 2000). Stud-ies done in the Kaya forests and surrounding areas (Kibet andNyamweru, 2008; Pakia, 2000; Nyamweru et al., 2008; Tengeza,1999; Waiyaki, 1995) indicate that these forest patches are biolog-ically and culturally diverse. Unfortunately, several of these sitesdespite their high conservation value and cultural significance havecontinued to suffer destruction and degradation. Pressure fromincreased demand for firewood, timber, mining, and more landfor farming and settlement linked to increasing human popula-tion, and the development of the tourism industry are threateningthese forest patches (Kibet and Nyamweru, 2008; Nyamweru et al.,2008). Fear of divine retribution played a significant role in theenforcement of traditional rules. Unfortunately, the decline in thetraditional values and the authority of elders who are the custodi-ans of the forests have made the system less effective.
It is of utmost important that these forest patches are effec-tively protected. To design appropriate and focused conservationmeasures, there is a need for quantitative data that can be used toempirically define the vegetation communities and identify theiruniqueness. Currently these data are inadequate in some areas andcompletely lacking in other coastal forests (Clarke and Robertson,2000).
In view of increasing threats faced by Kaya forests, this studywas carried out to understand spatial species distribution, plantassociation, and regeneration potential of commonly utilized plantin traditionally managed ecosystem. The specific objectives were to(1) identify plant communities, (2) identify species diversity, rich-ness and associations, (3) assess recruitment and regeneration ofcommonly utilized species and conservation options.
2. Materials and methods
2.1. Study area
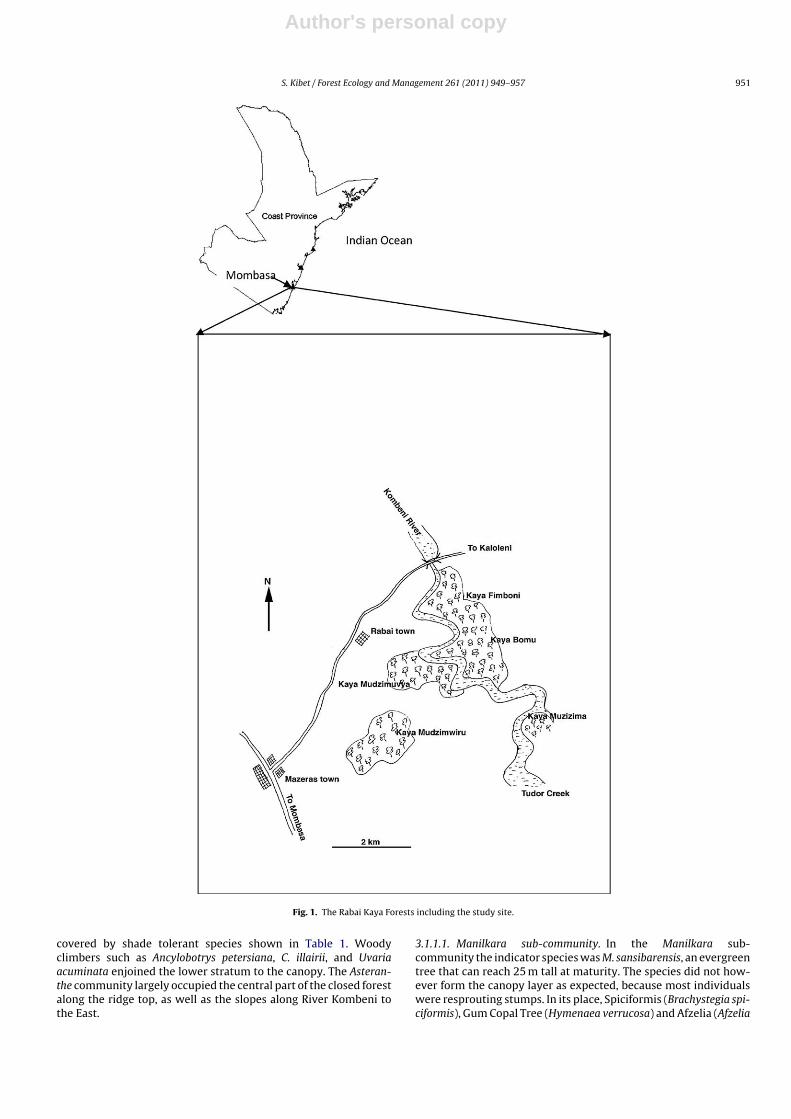
The study was conducted in a Rabai sacred forest named KayaMudzimuvya, located between 39◦34′E to 39◦36′E and 3◦56′S to3◦57′S in Rabai location of Kilifi County of coastal Kenya (seeFig. 1).Topographically, the area is gently undulating to undu-lating, with altitudes ranging between 20 and 200 m above sealevel.
The area experiences bimodal rainfall patterns with longrains (Mwaka) between March and July and short rains (Vuri)October–December with a dry period between January and March.Average annual rainfall ranges between 1100 and 2000 mm (Boxemet al., 1986). The minimum and maximum temperatures range22.5–24.5◦C and 26.5–34◦C respectively.
Kaya Mudzimuvya forest is one of the five remaining sacredforests of the Rabai community; the other four are Kaya Mudz-imwiru, Kaya Bomu, Kaya Fimboni, and Kaya Mzizima. These forestsare under the protection of Kaya elders who are the custodians ofthe sacred forests in the area. Regrettably, the authority of the eldershas been declining over the years coupled with social, economic,and political dynamics that has allowed some of the sites (e.g. KayaBendeje) to be encroached upon.
The population density of Rabai area based on 1999 census was493 persons per square kilometer (Republic of Kenya, 2000). Subsis-tence agriculture and animal husbandry are the main land activitiesin areas neighboring the Kaya forests. Despite the enormous influ-ence of new religion and western civilization, a significant portion
of the population remains committed to their culture and traditionswhich are hallmark on the Kaya-based rituals and ceremonies.
In 1998, 171.3 ha of Kaya Mudzimuvya forest was registered asa National Monument (NM) under the Antiquities and MonumentAct (Cap 215), Laws of Kenya as a measure to protect it.
2.2. Sampling design
A random nested plot method modified from Hall and Bawa(1993) was used for sampling. The modifications made includedthe use of randomised plots instead of plots placed along prede-termined transects, reducing sampling area for trees from 1.0 hato 0.4 ha and using four 2 m × 2 m plots for seedlings instead ofone. The plots measuring 20 m × 20 m were used to enumeratetrees (individuals with diameter at breast height – DBH > 10 cm),10 m × 10 m for saplings/shrubs (with DBH < 10 cm and >1 cm) and2 m × 2 m for herbs and seedlings. One 10 × 10 m plot was placedwithin one quarter of the 20 m × 20 m plot while four 2 m × 2 mplots were placed at each of the corners of the 20 m × 20 m plots.Random plots were generated using a random number generatorand a global positioning system (GPS) receiver was used to locatethem on the ground.
All individual species were identified following Beentje (1994)procedure. Heights of all individuals from 1 m and above wereestimated, basal diameter (BD), DBH, and their densities perplot recorded. Cover for three strata, trees, shrubs, and herbs,was estimated. In each sampling plot, indicators of disturbances(e.g. grazing), species harvested, and presence of bundles of fire-wood/poles of harvested species were recorded.
2.3. Analysis of data
The raw data initially recorded in Excel sheet were subjectedto Systat program version 8.0 (SPSS Software Inc., 1998), wherebyvolumes, basal area and density were calculated per hectare. Thefigures generated were keyed into statistical software; PC-ORD ver-sion 4 program whereby TWINSPAN multivariate analysis (McCuneand Mefford, 1999) was used to analyze phytosociological associ-ation. Basal area figures were used as cover-abundance values inthe analysis. Shannon diversity index (H′) was calculated using theformula H′ = −∑
{(ni/N) log (ni/N)} (Misra, 1989).
3. Results
Two hundred and three plots, covering 8.12 ha and representing4.7% of the study area, were sampled. A total of 355 woody specieswere identified and 320 of them run in TWINSPAN multivariateanalysis as described by Hill (1979) to determine species associa-tion. Some species were, however, expunged to reduce ‘noise’. Atwo-way ordered matrix table was used to identify plant clustersthat were then named based on dominant species.
3.1. Plant communities
Two plant communities (Asteranthe and Bridelia) and anundifferentiated vegetation type were identified. The Asteranthecommunity consisted of two sub-communities namely Manilkaraand Scorodophloeos, while the Bridelia community had one sub-community and one seral stage namely Catunaregam and Keetiarespectively (see Fig. 2).
3.1.1. Asteranthe communityThe diagnostic species for the Asteranthe community were
Asteranthe asterias and Combretum illairii. Tree layer (canopy)preferential species included Scorodophloeos fischeri, Manilkarasansibarensis and Haplocoelum inoploeum. The other layers were
Author's personal copy
S. Kibet / Forest Ecology and Management 261 (2011) 949–957 951
Fig. 1. The Rabai Kaya Forests including the study site.
covered by shade tolerant species shown in Table 1. Woodyclimbers such as Ancylobotrys petersiana, C. illairii, and Uvariaacuminata enjoined the lower stratum to the canopy. The Asteran-the community largely occupied the central part of the closed forestalong the ridge top, as well as the slopes along River Kombeni tothe East.
3.1.1.1. Manilkara sub-community. In the Manilkara sub-community the indicator species was M. sansibarensis, an evergreentree that can reach 25 m tall at maturity. The species did not how-ever form the canopy layer as expected, because most individualswere resprouting stumps. In its place, Spiciformis (Brachystegia spi-ciformis), Gum Copal Tree (Hymenaea verrucosa) and Afzelia (Afzelia

Author's personal copy
952 S. Kibet / Forest Ecology and Management 261 (2011) 949–957
Kaya Mudzimuvya Forest vegetation
B
Asteranthe community
Closed forest
N= 65
Bridelia community Open Wooded Bushland/Shrubland/Grassland
N= 138
Scorodophloeossub-community
N=37
Manilkarasub-community
N=19
Keetia seral stage
N=23
Catunaregamsub-community
N= 63
Transitional vegetation (ecotone)
N=9
Undifferentiated vegetation type
N= 52
Fig. 2. The plants community clusters identified in study site.
Table 1Species that defined plant communities’ strata.
Communities Preferential/indicator species
Tree layer Shrub layer Herb layer
Asteranthe Haplocoelum inoploeum, Manilkara sansibarensis,Scorodophloeos fischeri
Asteranthe asterias, Combretum illairii, Vepristrichocarpa, Acalypha fruticosa
Psilotrichum sclerethum
Bridelia Bridelia carthatica, Cocos nucifera, Anacardiumoccidentale, Mangifera indica
Allophyllus rubifolius, Aspilia mossambicensis, Premnachrysoclada
Vernonia hildebrandtii,Agathisanthemum bojeri
quanzensis) constituted the canopy layer. Preferential species thatformed the understory layers are listed in Table 2. Common woodyclimbers of the sub-community included Synaptolepis kirkii, Hugo-nia castaneifolia, Apodostigma pallens, and Artabotrys modestusamong others.
3.1.1.2. Scorodophloeos sub-community. The Scorodophloeos sub-community was characterized by dominant presence of dryevergreen forest species namely; Scorodophloeos fischeri, Mildbrae-dia carprinifolia and Commiphora eminii. Preferential tree and shrublayer species that dominated this sub-community are listed inTable 2. Unlike the Manilkara sub-community where climbers weresignificant, only a few species such as Grewia forbesii, G. holstii andHibiscus faulkenarae were observed in the sub-community.
3.1.2. Bridelia communityThe Bridelia community was dominated by Bridelia cathartica, a
forest margin or bush land thicket species. Preferential species thatdefined the tree, shrub and herb layers are indicated in Table 1.Bridelia community occurred in open woody shrub land and thick-ets, largely in sites previously disturbed by grazing and cultivationthat have been undergoing regeneration in the last 40 years.
3.1.2.1. Catunaregam sub-community. The species Catunaregamnilotica characterized the Catunaregam sub-community. Despitethe species being common in several habitats such as woodedgrassland, several coastal bush lands, and palm woodland, itwas not represented in the Asteranthe community at all and
Table 2Indicator species in the vegetation sub-communities.
Sub-communities Preferential/indicator species
Tree layer Shrub layer Herb layer
Manilkara Manilkara sansibarensis, Brachystegia spiciformis,Hymenaea verrucosa, Afzelia quanzensis
Phyllanthus welwitschianus, Suregada zanzibarensis,Heinsia crinita, Ritchiea capparoides
Chazaliella abrupta var. abrupta,paveta stenosepala
Scorodophloeos Scorodophloeos fischeri, Commiphora eminii, Gyrocarpusamericanus, Manilkara sulcata, Combretum schumannii,Dobera loranthifolia, Lecaniodiscus fraxinifolius
Mildbraedia carprinifolia, Tricalysia ovalifolia,Memecylon fragrans, Croton pseudopulchellus,Uvariodendron kirkii
Vitellariopsis kirkii,Scorodophloeos fischeri
Catunaregam Vitex payos, Acacia nilotica, A. etbaica and A. mellifera,Strychnos madagascariensis, S. spinosa
Dichrostachys cinerea, Ormocarpum kirkii, Harrisonaabyssinica
Aganthisanthemum bojeri,Vernonia hildebrandtii

Author's personal copy
S. Kibet / Forest Ecology and Management 261 (2011) 949–957 953
Table 3Some recorded species that were confined to specific identified sub-communities.
Manilkara Scorodophloeos Catunaregam
Drypetes natalensis Memecylon fragrans Vitex payosPleiocarpa pycnantha Craibia brevicaudata Catunaregam niloticaChazaliella abrupta Pycnocoma littoralis Acacia etbaicaAllophylus pervillei Uvaria faulknerae Acacia niloticaArtabotrys modestus Cola minor Acacia melliferaLudia mauritiana Croton pseudopulchellus Dichrostachys cinereaDiospyros consolatae Ricinodendron heudelotii Omorcarpum kirkiiXylopia parviflora Grewia stulhmanniiPseudobersama
mossambicensisSynadenium pereskiifolium
Clerodendrum incisum Hibiscus altissimaStrophanthus kombe Sterculia schliebeniiErythrina sacleuxii Combretum tenuipetiolatumOphrypetalum odoratum Scorodophloeos fischeriMemecylon sansibaricum Cynometra suaheliensis
only once within the Keetia seral stage. Other species unique tothis sub-community included wooded grassland species such asDichrostachys cinerea and Vitex payos. The vegetation type was char-acterized by several thorny species, especially in the shrub layer.
3.1.2.2. Keetia seral stage. Unlike the earlier mentioned plant com-munities, no diagnostic species was identified for Keetia seral stagehowever; forest margins species such as Keetia zanzibarica andRytigynia celastroides as well as weedy Stachytarpheta jamaicensiswere common. Moreover, short-lived perennials such as Waltheriaindica, Agathesanthemum bojeri, Tinnea aethiopica and some agri-cultural tree crops were well represented.
Some species occurred in more than one sub-community whilesome were exclusive to one sub-community or community. Table 3indicates some of the species that occurred exclusively in some ofthe identified sub-communities.
3.2. Species diversity and richness
Statistically, the Asteranthe community recorded higher speciesdiversity and richness than the Bridelia community with values of1.83 and 38 compared to 1.77 and 28 respectively. There was adecline of woody species from 260 in the Asteranthe to 233 in theBridelia community.
On average the Asteranthe community recorded 6 trees and 21understory species per plot while the Bridelia community recordedan average of 3 trees and 18 understory species. Trees species gen-eral diversity declined from 1.65 to 1.61 while understory speciesdeclined from 2.01 to 1.93 in Asteranthe to Bridelia respectively.
The most species rich plot was found in the Bridelia communitywith 57 species. There were 7 other plots with more than 50 specieseach. Of all the 8 plots with over 50 species each, 6 of them werefound in the Asteranthe community majority within ecotone zone.The 3 plots with the least species richness (less than 10 specieseach) occurred in the Bridelia community.
In general, closed forest sub-communities (Manilkara andScorodophloeos) had on average the highest species richness. TheManilkara sub-community with 28 plots was the richest, withspecies ranging from 21 to 56 (mean = 43, S.D. = 8.1) while theScorodophloeos sub-community, represented by 37 plots, was sec-ond with species ranging from 19 to 49 (mean = 34, S.D. = 7.1). TheCatunaregam sub-community represented by 63 plots was speciespoor with a range of 11–57 (mean = 27, S.D. = 9.5).
3.3. Recruitment and regeneration of species
Commonly harvested species for building houses such asVitellariopsis kirkii, Scorodophloeos fischeri, H. verrucosa, Julbernar-dia magnistipulata and Millettia usaramensis were represented at
seedling stage in more than 10 plots sampled; other speciesoccurred in lower frequencies.
Some of the taxa had their seedlings exclusively confined tothe closed forest and others in the open woody bush land/shrubland. Cynometra suaheliensis, C. webberi, M. sansibarensis and J.magnistipulata seedlings occurred solely within the closed forestunder parent plants. In contrast, the seedlings of forest marginspecies such as Strychnos madagascariensis, Premna chrysoclada,and Uvaria lucida occurred exclusively in the open woody bushland. A. quanzensis, Combretum schumannii, H. verrucosa and V. kirkiiseedlings occurred in both the closed forest and open woody shrubland.
Over 40% of H. verrucosa and all Parkia filicoidea seedlingsrecorded in sampling plots occurred in the open woody shrubland under cashew (Anacardium occidentale) trees. Few H. verrucosaseedlings grew under their parent plants, irrespective of whethersuch mother plant occurred in the closed forest or open shrub land.The majority of P. filicoidea seedlings were found in closed forestunder the mother plant outside sample plots. In comparison, under-story and climber species had more seedlings than canopy speciesboth in the Asteranthe and Bridelia communities.
A majority of commonly utilized species found in the Brideliaand Asteranthe communities regenerated more as resprouters thanas reseeders.
4. Discussion
Although the lowland coastal dry forest of Kaya Mudzimuvya iscurrently protected from indiscriminate human destruction, vege-tation continues to be impacted by the extraction of forest productsas well as by livestock grazing. The past and present extraction offirewood, poles, withers, fibers, and weaving materials has shapedthe phytosociological associations, structure and composition ofspecies and this has a bearing on the choice of possible conservationoptions.
Previous studies in the region revealed that variation in vegeta-tion is attributed to ecological factors such as soil characteristics,topography, wind direction, and level of forest disturbance (Clarkeand Robertson, 2000; Schmidt, 1991). In this study major veg-etation types seemed shaped largely by anthropogenic factors,whereas intra community variation point to other ecologicaldynamics. Fig. 3 shows a hypothetical diagrammatic illustration ofvegetation dynamics as driven by both biotic and abiotic factors.
As envisaged in the above diagram, any extensive clearing offorests, coupled with consistent grazing alters Asteranthe closedforest into Bridelia open woodland vegetation type and eventu-ally into grassland. The reverse option through natural successionis possible; however its success rate depends on the frequency,duration and intensity of perturbation instigated. The dynamicsare less systemic if human perturbations vary in space and timeand this is demonstrated by observed plant communities’ variabil-ity. The forest clearing and farm encroachment in the past causeddegeneration of Manilkara forest type to a Keetia-Rytigynia forma-tion/vegetation type. Continued grazing and extraction of firewood,poles, withers and other non timber forest products has not onlyfacilitated the establishment of short-lived perennials and otherweedy species but also made the formation unstable. Further evo-lution of this vegetation type is described under Sections 4.1 and4.3.
4.1. Plants communities
The Bridelia community largely covered sites used in thepast as cultivation and grazing fields by the Rabai people.These sites stretched from down slope at the riverine habitat

Author's personal copy
954 S. Kibet / Forest Ecology and Management 261 (2011) 949–957
Cultivation, NTFPs extraction
Slash and burn cultivation, livestock grazing
Human induce vegetation changes
Natural vegetation dynamics (succession and natural disturbances).
NTFPs – Non Timber Forest Products
ManilkaraSub-community
Keetia seral stage
Scorodophloeossub-community
Catunaregamsub-community
Asteranthecommunity
Brideliacommunity
Open grassland Vegetation
Low past perturbations & abiotic factors
High present perturbations & abiotic factors
Low present perturbations
High past perturbations (logging) & abiotic factors
Fig. 3. Vegetation dynamics due to biotic and abiotic variability.
to as high as the ridge-top adjacent to Asteranthe closed forest.It is interpreted therefore that persistent anthropogenic pres-sures over many years could have created Bridelia communityfrom Asteranthe. The presence of isolated huge (>50 cm DBH)forest canopy species as well as shade tolerant species in dom-inantly open shrub land vegetation supports this assertion. Thediagnostic as well as preferential species found in the Brideliacommunity were either generalists such as Allophylus rubifolius,Premna chrysoclada and Tinnea aethiopica, and therefore found inseveral habitats, and/or bushland–grassland specialists (Beentje,1994).
The lack of well defined species association within Keetia seralstage could be ascribed to its unstable state. The Keetia seral stageoccurs within close proximity to human settlement and thus suffersfrom frequent disturbance from grazing and wood harvesting. It isdominated by short-lived perennials and exotic species creatinga human-induce vegetation formation whose ecological evolutionis less predictable. Dominance of short-lived perennials with fewseedlings of climax forest species corroborates this position.
Although the exotic species were numerically few, some weresignificantly huge and dominant so that they greatly influencedtheir immediate ecological unit. Occasionally, cashew (A. occiden-tale) trees “hosted” Rytigynia celastroides, Trichilia emetica, Keetiazanzibarica, P. filicoidea and H. verrucosa underneath them. Themutual relationship between the species is not clear. Howeverthere are two likely scenarios; first the A. occidentale may havebeen acting as a ‘nurse plant’ for the associating species, thuspromoting their development. Secondly, it may be strategicallyproviding good resting points for dispersers of the seeds of theassociating species, thus promoting their propagation in the pro-cess. Notably all associating species were juvenile (either saplingsor seedlings) as compared to their ‘host plant’ thus indicating laterarrival. Whitmore (1991) reported a similar observation in Mexicowhereby tree relics on the farms left after cultivation provided
perches for birds, some of them seed dispersers, predicted to helpthe reinvasion of climax species. It is predicted therefore that withenhanced conservation, Keetia seral stage will evolve into a Keetia-Rytigynia “sub-community”. Over the years H. verrucosa and P.filicoidea will take over, creating a Manilkara-like type of vegetation.
Unlike Keetia seral stage, species association in Catunaregamsub-community is better defined, with assemblages of decidu-ous thorny species. The dominant presence of Vitex payos andCatunaregam nilotica, both wooded grassland tree species (Beentje,1994), indicates some level of plant community stability. It mighttake many years for forest species to take over if natural succes-sion progresses unperturbed. The sub-community can be comparedwith the Kaya Mtswakara thickets and wooded vegetation (Pakia,2000) and Acacia thorn-bushland (Moomaw, 1960). The sub-community has significant population of Acacia spp and otherthorny species such as Strychnos spp., Harrisonia abyssinica andDichrostachys cinerea and could best be described as the “thorn inthe flesh – functional group” or association. They are largely decid-uous species and sometimes form impenetrable thickets that arehighly susceptible to fire during the dry season. The herbaceouslayer commonly consists of annuals. Unlike the Keetia seral stage,where anthropogenic influence is conspicuously displayed by thepresence of huge exotic fruit trees, less than 10 such individualsoccurred in the Catunaregam sub-community. The Rabai peopleused tree crops to mark and lay claim of ownership to a piece of land,which were planted soon after one has cleared the forests and culti-vated it for a couple of seasons, to thwart potential ‘squatters’ giventhat the land was communally owned (Kibet, 2002). The presenceof fewer exotic plants in this secondary vegetation could mean thatthe Catunaregam sub-community is a creation of a relatively “morerecent” human disturbance later than 1970, when most of the west-ern part of forest had already been cleared for cultivation. Despiteearlier perturbations the Catunaregam sub-community seemed tohave steadily regenerated from the early 1980s, after individu-

Author's personal copy
S. Kibet / Forest Ecology and Management 261 (2011) 949–957 955
als who had encroached on the forest were forcefully evicted bythe elders with the help of the government (Nyamweru et al.,2008). The close proximity of Catunaregam and Scorodophloeos sub-communities and thus the possibility of shared edaphic factorspoints to a possible succession toward later sub-community typeof vegetation if future perturbations are curtailed. The presence ofsecondary forest species such as Commiphora eminii and Gyrocarpusamericanus in the two sub-communities supports this argument. Italso presupposes that any form of cultivation, grazing or extractionof non-timber forest products from Scorodophloeos will cause thesub-community to degenerate into a Catunaregam type of vegeta-tion dominated by thorny generalists species but devoid of forestdependant species, and ultimately into a grassland if the frequencyand intensity of perturbation persist.
In contrast, the Asteranthe community occupied areas with cul-tural sites where plant extraction, grazing, and cultivation areprohibited, meaning that the vegetation has remained ‘relatively’undisturbed for more than 100 years compared to other sites.Taxa associations showed concentrations of forest, dense bushland, or thicket specialists. This observation agrees with Pakia(2000) description of dense canopy forest of Asteranthe consisting ofScorodophloeos and Hugonia communities in Kaya Mtswakara for-est, 25 km away from the study site. Given that most of the taxaare trees, any extraction could greatly alter the microclimate andsubsequently floristic composition and structure.
The association of species in the Scorodophloeos sub-communityclosely relates to legume-dominated dry forest (Clarke andRobertson, 2000) due to fair representation of Caesalpiniaceae(S. fischeri, Cynometra webberi, C. suaheliensis and Julbernardiamagnistipulata), Manilkara sulcata and M. sansibarensis. The sub-community is stable, as indicated by dominance of S. fischeri at thetree as well as the shrub layer (Clarke and Robertson, 2000). Othermembers of the shrub layer included M. carprinifolia, Memecylonfragrans and Grandidiera boivinii, which form a thick understory.The sub-community occurred on the eastern side of the foreststretching from the mid slope downwards and forming the river-ine ecosystem along Kombeni River. Contrary to expectation thissub-community had a low population of woody climber species,unlike the situation in the nearby Kaya Kambe forest, where selec-tive logging of huge trees had triggered massive invasion by lianas(Hawthorne, 1984).
The Manilkara sub-community was largely confined within theridge top at the middle of the Kaya but with a small patch sur-rounded by the Scorodophloeos sub-community at the lower slopes.In terms of anthropogenic disturbance, the sub-community has notexperienced significant extraction of large trees due to the pres-ence of prayer shrines and burial grounds – cultural sites highlyrevered by the local community (Nyamweru et al., 2008). However,occasional natural tree fall would occur especially among the oldBrachystegia trees, creating tree gaps. Unlike in the Scorodophloeos,where lianas were scarce, Manilkara sub-community had strongpresence of S. kirkii, H. castaneifolia, A. pallens and A. modestus forestwoody climbers. As earlier conceptualized in Fig. 3 and expoundedin Section 4.1, Manilkara sub-community is by no means static,natural tree fall and selective pole harvesting instigates ecologi-cal cascade within the ‘equilibrium’, unless greater perturbationswhere huge trees are removed and seedlings destroyed to causesignificant change in species composition and structure.
Physiognomically, Manilkara and Scorodophloeos sub-communities appeared similar and thus without detailed samplingit is easy to miss out species unique to each vegetation type.Despite sharing several species, each support species that arestrictly confined to either of the sub-communities (see Table 3).The presence of species exclusive to one sub-community thoughadjacent to each other is subject to debate given that the altitudinalrange and extend of forest is fairly small therefore segregation
due to elevation is unlikely. Possible explanations include: (1)ecological differences such as edaphic factors and the presenceof micro-habitats; some of the species are highly specialized forcertain niches such as riverine; (2) some preferred species couldhave been selectively harvested in one sub-community; (3) theinfluence of topography and site exposure might bear on speciesassemblages and distribution. This phenomenon indicates thatcoastal forest vegetation changes in species composition andassociation in fairly short time intervals, and this could be a featurethat has contributed to making coastal forests biologically diversewith high species endemism.
4.2. Species diversity and richness
In terms of species diversity and richness, the Asteranthe andBridelia communities varied significantly, a scenario best explainedby variability in abiotic factors as well as type, frequency and inten-sity of human perturbation both in the recent and the distantpast. The general tree layer species diversity differed significantlybetween Asteranthe and Bridelia communities. The wider life formsof the Asteranthe community presuppose the presence of diverseecological niches to support high diversity of species. The primaryspecies germinates below closed canopies and get established intoseedling banks to await favorable conditions to be ‘released’, whilegaps are invaded by pioneers and woody climbers (Whitmore,1991). The Asteranthe community is comprised of tall forest withinterlocking canopies, with occasional tree gaps created by natu-ral tree fall. The gaps together with forest edges provided ecotonemicroclimates suitable for both forest dependent and forest marginspecies to thrive.
The two closed forest sub-communities (Manilkara andScorodophloeos) differed in species diversity and richness. Speciesrichness of woody climbers and understory specialists declinedfrom Manilkara to Scorodophloeos. The comparatively higher treespecies diversity in Manilkara than Scorodophloeos probably hassomething to do with past human perturbation that targeted thelatter sub-community. Past logging was evident in Scorodophloeossub-community where old stumps were recorded in several plots.Increased light penetration to ground level may have led to thedevelopment of a dense understorey, consisting of light demand-ing Acalypha fruticosa and M. carprinifolia (Hawthorne, 1984)at the expense of shade tolerant species. Most of the reveredcultural sites and burial grounds located within the Manilkara sub-community and the fear of the unknown for transgressing againstthe spirits kept potential culprits at bay (Nyamweru et al., 2008).The high number of species (57 species) in one plot within theCatunaregam sub-community occurred close to the forest edge,showing that light demanding, forest margin and a few forestdependant species were beginning to expand into the secondaryvegetation in response to conservation measures currently in place.
4.3. Recruitment and regeneration
Broadly, the seedlings of canopy species were few in numbercompared to those of shrub layer species. This could be attributedto the phenomenon where seedlings of several canopy speciesin Asteranthe were only confined to close forest. These closedforests occur on only a third of the total area sampled. Secondly,some species showed rather clumped distribution and thus thosewith large seeds may have had all of their seedlings concentratedunderneath mother plants. This was the case with Caesalpinioids;Julbernadia magnistipulata, C. webberi and C. suaheliensis. In termsof conservation these species are seriously threatened by habitatdestruction as their seedlings prefer closed forest to thrive. Fur-thermore, if such species are commonly preferred for use by thelocal people, they are prone to over exploitation as all the popula-

Author's personal copy
956 S. Kibet / Forest Ecology and Management 261 (2011) 949–957
tion are concentrated in a few areas thus making them easy targetsfor harvesting and transporting.
Species whose seedlings grow both in the closed and openvegetation have higher chances of establishing themselves, andare likely to take over degraded habitat if human perturbation isremoved. A. quanzensis, C. schumannii and H. verrucosa could eas-ily constitute the first climax species to re-establish themselves indisturbed sites if parent plants are in close proximity to supply thegermplasm. Species whose seedlings are exclusively confined to theopen shrub land (S. madagascariensis, P. chrysoclada and U. lucida)are likely to gradually decline as forest species re-invade adjacentopen shrub lands. The opposite is the case if wood extraction andopening up of forest recur.
The seedlings of Parkia filicoidea and H. verrucosa consistentlyoccurred underneath a number of A. occidentale individuals thatwere closer to forest edge. The agricultural tree crops formed themajority of the huge evergreen individuals in open shrub land, mak-ing them obvious stopover points for birds and mammals. The fruitsof A. occidentale attract frugivores and small mammals like mon-keys and bats, all of whom are major seed dispersers. This scenarioneeds further investigation as it may provide insight on the role ofnurse plants and/or forest relics in degraded ecosystems.
The forest gaps in the Asteranthe community were dominatedby light requiring species that could have been from ‘seed-rain’,trans-located by mammals, borne by birds or wind, or from seedbanks, thus increasing its diversity.
Some species identified during the inventory work are of conser-vation concern. Bauhinia mombassae, Combretum tenuipetiolatum,Holarrhena pubescens and Euphorbia wakefieldii are classified ascritically endangered in 1997 IUCN Red List of Threatened plants(Walter and Gillet, 1998) though they are currently undergoingreview by plant experts with experience in the region. One suchreview took place in April 2010 at IUCN Eastern Africa RegionalOffice in Nairobi. All the species seems to have a specialised habitat.B. mombassae and C. tenuipetiolatum were common along KombeniRiver while H. pubescens and E. wakefieldii were common on rockyareas and on forest edges. Due to their specificity to certain eco-logical niches, these species are in danger of becoming extinct ifhabitat destruction is not stopped. Other species requiring closemonitoring include Encephalartos hildebrandtii and V. kirkii bothrecorded in Appendix 1 in the IUCN Red List data although in thiscurrent inventory their frequencies were fairly high compared toother species. Similarly Milicia excelsa, Terminalia sambesiaca andBrachylaena huillensis occurred in extremely low numbers. Singleresprouting individuals of each species were noted and may needspecial attention.
5. Conclusions
Physiognomic similarities among the coastal forests of easternAfrica do not necessarily mean similarity in plant communitiesand/or species diversity and richness. The findings of this studyindicate that it is easy to overlook the uniqueness of plan t com-munities and presence of exceptional species that may requirespecialized attention. The anthropogenic factor is the single mostimportant influence in shaping plant communities, species com-position, recruitment/regeneration and grand pattern, reflecting ofpeoples’ culture and historical influence within the study site.
The high species diversity within areas having cultural sitessupports the need to promote and strengthen traditional manage-ment system within natural and cultural landscapes, so as to takeadvantage of cultural values of biodiversity and the local ecologicalknowledge. Recruitment and regeneration of some commonly har-vested species is affected by their spatial distribution pattern, habi-tat suitability, and availability of mature individuals to provide the
needed germplasm. Human input in the restoration could enhancethe process if sound interventions are put in place. Introducing afew relatively fast growing non-invasive tree species (such as fruittrees) in strategic locations within degraded sites may hasten re-colonisation by attracting pollinators and/or seed dispersal agents.
Acknowledgements
This study was part of an MSc research financially supported byUNESCO – People and Plants Initiative for which I will forever beindebted. I sincerely appreciate support from my supervisors; Dr.Robert Hoft, Dr. Enoch Mrabu and Dr. Elizabeth Omino. This workwould not have been possible without the commitment of Drs.Robert Hoft and Martina Hoft who squeeze their free time to assistwith data analysis. The Coastal Forest Conservation Unit projectpersonnel help with field logistics for which I am truly grateful. Iappreciate the assistance I received from Quentin Luke with speciesidentification and John Charo who helped me during my fieldwork.I thank Celia Nyamweru for editing the paper. Lastly, I am indebtedto the two reviewers for their edits, comments and critiques on themanuscript.
References
Beentje, H.J., 1994. Kenya Trees Shrubs and Lianas. National Museums of Kenya,Nairobi.
Boxem, H.W., Meester, T., Smaling, E.M.A., 1986. Soils of the Kilifi Area. Reconnais-sance Soil Survey Report No. R111987. Kenya Soil Survey. Ministry of AgricultureKenya.
Burgess, N.D., Clarke, G.P., Madgwick, J., Robertson, S.A., Dickinson, A., 2000. Distri-bution and status. In: Burgess, N.D., Clarke, G.P. (Eds.), Coastal Forests of EasternAfrica. IUCN, Gland, Switzerland and Cambridge, UK.
Clarke, G.P., Robertson, S.A., 2000. Vegetation communities. In: Burgess, N.D., Clarke,G.P. (Eds.), Coastal Forests of Eastern Africa. IUCN, Gland, Switzerland and Cam-bridge, UK.
Hall, P., Bawa, K., 1993. Methods to assess the impact of extraction of non-timbertropical forest products on plant population. Economic Botany 47 (3), 234–247.
Hawthorne, W.D., 1984. Ecological and biogeographical patterns in the coastal forestof East Africa, DPhil Thesis, University of Oxford.
Hill, M.O., 1979. TWINSPAN, a FORTRAN programs for arranging multivariate data inan ordered two-way table by classification of individuals and attributes. Ecologyand Systematics Cornell University, Ithaca, NY.
Janzen, D.H., 1988. Management of Habitat Fragments in Tropical Dry Forests,Growth. Annals, Missouri Botanical Garden, USA.
Kibet, S., 2002. Human Disturbance and its Impact on Vegetation Structure, Com-position and Regeneration of Kenya Coastal Forests (a case study of KayaMudzimuvya Forest). Unpublished MSc Thesis submitted to Jomo Kenyatta Uni-versity of Agriculture and Technology, Nairobi, Kenya.
Kibet, S., Nyamweru, C., 2008. Cultural and biological heritage at risk; the case of theRabai Kaya forests in Coastal Kenya. Journal of Human Ecology 24 (4), 287–295.
McCune, B., Mefford, M.J., 1999. PC-ORD multivariate analysis of ecological data,version 4. MJM software designs. Gleneden Beach, Oregon, USA.
Misra, K.C., 1989. Manual of Plant Ecology, third ed. Oxford and IBH, New Delhi,India.
Moomaw, J.C., 1960. A study of the plant Ecology of the Coast Region of Kenya, EastAfrica, Nairobi. Government Printer.
Nyamweru, C., Kibet, S., Pakia, M., Cooke, A.J., 2008. The Kaya forest of Coastal Kenya;remnant patches or dynamic entities? In: Sheridan, M.J., Nyamweru, C. (Eds.),African Sacred Groves: Ecological Dynamic & Social Change. James Currey, UK,pp. 62–86.
Pakia, M., 2000. Plant ecology and ethno botany of two sacred forests (Kayas) ofKenyan Coast. Unpublished MSc thesis for School of Life and EnvironmentalScience, University of Natal, South Africa.
Republic of Kenya, 2000. Population Census – Central Bureau of Statistics. Ministryof Planning and National Development, Nairobi.
Robertson, S.A., 1987. Preliminary floristic survey of Kaya forests of coastal KenyaUnpublished reports to the Director NMK and to WWF, Nairobi, Kenya.
Robertson, S.A., Luke, W.R.Q., 1993. The Vegetation and Conservation Status of KayaCoastal Forests in Kenya. WWF, Nairobi.
Schmidt, R., 1991. Ecology of a Tropical Lowland Rain Forest plant communities: soilcharacteristics and nutrients relations of the forests of the Shimba Hills NationalReserve, Kenya, Band 179. Dissertationes Botanicae, Berlin-Stuttgart.
Spear, T.T., 1978. The Kaya Complex. Kenya Literature Bureau, Nairobi, Kenya.SPSS Inc., 1998. Systat Software. Statistical Package for Social Sciences, USA.Tengeza, A.H., 1999. The Mijikenda Grave markers: A report of a study on timber and
live tree grave markers and associated traditions in Kaya forests. In: Jianchu, X.(Ed.) Links Between Cultures and Biodiversity: Proceedings of the Cultures andBiodiversity Congress, 2000, Yunnan, China.
UNESCO, 2008. http://whc.unesco.org/en/list/1231.

Author's personal copy
S. Kibet / Forest Ecology and Management 261 (2011) 949–957 957
Walter, K.S., Gillet, H.J., 1998. 1997 IUCN Red List of Threatened Plants. Compiledby the World Conservation Monitoring Centre. IUCN – The World ConservationUnion, Gland, Switzerland and Cambridge, UK.
Waiyaki, E.M., 1995. Effects of forest fragmentation, isolation and structure,on the richness and abundance of bird communities in major coastalforests of south coast, Kenya. Unpublished MSc Thesis in Conservation
Biology. Durrell Institute of Conservation and Ecology, University of Kent,UK.
Whitmore, T.C., 1991. Tropical rain forest dynamics and its implications for man-agement. In: Gomez-Pompa, A., Whitmore, T.C. (Eds.), Rainforest Regenerationand Management MAB Series, vol. 6. UNESCO, Paris.
White, F., 1983. The Vegetation of Africa. UNESCO, Paris.





























