Embed Size (px)
Citation preview

Regression of Experimental Burkitt's Lymphoma Induced by Epstein-Barr Virus-Immortalized Human B Cells
By Giovanna Tosato, Cecilia Sgadari, Kazuyuki Taga, Karen D. Jones, Sandra E. Pike, Amy Rosenberg, Joan M.G. Sechler, Ian T. Magrath, Lori A. Love. and Kishur Bhatia
Epstein-Barr virus (EBV)-immortalized human B cells sur- vive only transiently when injected subcutaneously into athymic mice, whereas Burkitt's lymphoma cells give rise to progressively growing subcutaneous tumors. In this study, we tested whether these Burkitt's tumors could be induced to regress via a bystander effect induced by EBV- immortalized B cells. Simultaneous inoculation of EBV-im- mortalized B cells and Burkitt's lymphoma cells in the same subcutaneous site resulted in tumors that regressed with necrosis and scarring. Similarly, simultaneous inoculation of EBV-immortalized B cells and Burkitt's lymphoma cells in separate subcutaneous sites resulted in regression
DMINISTRATION of lymphocytes or tumor cells transduced with specific cytokine genes represent
novel approaches for the treatment of cancer. By either pro- viding antitumor effector cells or stimulating antitumor im- munity at the tumor site, these experimental modalities have resulted in decreased tumorigenicity and regression of existing tumors in a number of experimental model^."^ Re- cently, retroviral-mediated transfer in vivo of a herpes-vi- rus-derived thymidine kinase (tk) gene into a proportion of brain glioma tumor cells rendered them sensitive to destruc- tion by an antiviral drug. I t also led to death of other tumor cells, not transduced with the tk gene, through a poorly de- fined bystander effect." Despite these experimental results, the targeting of effector lymphocytes as well as anticancer cytokines and drug sensitivity factors to particular tumor sites has had limited clinical applications because it involves relatively complex procedures not widely available.' I
We considered the possibility that tumor cell killing might be induced by inoculation of the tumor-bearing host with benign cells that are normally destroyed if inoculated into the same host. We reasoned that, by stimulating a local cytotoxic response in vivo, one might achieve killing of tu- mor cells through a bystander effect."
When injected subcutaneously into irradiated athymic (nude) mice, human B cells immortalized with Epstein-Barr virus (EBV) either fail to grow or give rise to small tumors that soon regress through necrosis and scarring.** The mech- anism for this regression is not fully understood but has been
A
From the Division ofHematologic Products. Center.for Bio1ogic.s Ealuation and Research, and the Center,for Food Safety and Ap- plied Nrrtrition. Food and Drug Administration: and [he Pediatric Oncology Branch, National Cancer Institute. National Institutes o f Health, Bethesda. MD.
Submitted June 29, 1993; accepted September 29, 1993. Address reprint requests to Giovanna Tosato, MD, Division o f H e
matologic Producls. Center for Biologics Evaluation and Research, Bldg 29A. Room 2006, 1401 Rockville Pike, Rockville, MD 20852- 1448.
The publication costs ofthis ariicle were defayed in part by page charge payment. This article must therefore be hereby marked "advertisement" in accordance with 18 U.S.C. section I734 solely t o indicate thisjuct.
This is a USgovernment usork. There are no restrictions on it.s use. 0006-4971/94/8303-0005$0.00/0
776
of a proportion of the Burkitt's tumors. Furthermore, most of the established human Burkitt's tumors regressed with necrosis and scarring after intratumor inoculations with EBV-immortalized B cells. The EBV-immortalized B cells continued to exert this antitumor effect even when killed with irradiation. The experimental approach to Burkitt's lymphoma treatment described here exploits the ability of athymic mice to reject EBV-immortalized 5 cells to target an effective antitumor response to malignant cells normally incapable of eliciting it. This is a US government work. There are no restrictions on its use.
attributed to residual host i m m ~ n i t y . ' ~ In the same experi- mental system, human Burkitt's lymphoma cells generally develop into lethal malignant tumors.I4
In the present study, we examined whether inoculation of EBV-immortalized B cells might have an antitumor effect against experimental Burkitt's lymphoma in athymic mice.
MATERIALS AND METHODS
Cells and cell lines. EBV-immortalized cell lines were obtained either by exposure of normal peripheral blood B cells to EBV (B95- -8 strain) or by spontaneous outgrowth in vitro of peripheral blood B cells from normal EBV-seropositive individual^.'^ The Burkitt's lymphoma cell lines CA46 and JD38 were derived by spontaneous in vitro outgrowth of single cell suspended Burkitt's lymphoma tis- sues. l6 All cell lines were maintained in RPMl I640 medium (Bio- fluids, Rockville, MD) supplemented with 10% heat-inactivated fe- tal calf serum (FCS; Reheis, Armour Pharmaceuticals, Kankakee, IL), 2 mmol/L L-glutamine (GIBCO, Grand Island, NY), and 5 ppl mL gentamicin (Sigma Chemical CO, St Louis, MO). All cell lines were mycoplasma-free. Peripheral blood T cells were obtained from normal mononuclear cells by rosetting with 2-aminoethylisothiour- oniumbromide-treated sheep red blood cells.'5
Animal studies. Six- to eight-week-old female BALB/c nu/nu mice (National Cancer Institute [NCI], Bethesda, MD) maintained in pathogen-limited conditions were used throughout. Twenty-four hours before initiation of the experiments, the mice received 400- rad total body irradiation. Exponentially growing human B cell lines with a viability greater than 95% were injected ( lo7 cells in 0.2 mL RPMl 1640 medium containing 10% FCS) subcutaneously in the right abdominal quadrant through a 25-gauge needle on plastic l -mL syringes. In a number of experiments, two different cell lines were coinjected subcutaneously in the right abdominal quadrant ( IO ' cells from each of the cell lines in 0. I mL RPMI 1640 medium containing 10% FCS, total injection volume 0.2 mL). In some ex- periments, two different cell lines were simultaneously injected in distinct subcutaneous sites. the right and the left abdominal quad- rant ( lo7 cells from each cell line in 0.2 mL RPMI 1640 medium containing 10% FCS). lntratumor injections were performed by in- oculation of IO' cells suspended in 0.1 to 0.2 mL RPMI 1640 me- dium with 10% FCS into one site in the center of the tumor mass. All animals were observed twice weekly and tumor size was esti- mated in square millimeters as the product of two-dimensional cal- iper measurements (longest perpendicular length and width).
Reagents. Recombinant murine tumor necrosis factor-a (TNF- a ) was a kind gift of Genentech, Inc (South San Francisco, CA); recombinant human interleukin-6 (IL-6) was a kind gift of Sandoz Pharmaceutical (Basel, Switzerland); lipopolysaccharide (LPS)
Blood, Vol83, No 3 (February l), 1994: pp 776-784

REGRESSION OF EXPERIMENTAL BURKITT’S LYMPHOMA 777
from Escherichia coli 0127:B8 was purchased from Sigma; a puri- fied rabbit antiserum to highly purified natural murine IL-6” was a gift of Dr R. Nordan (NCI); a purified rabbit antiserum to recombi- nant murine TNF-a18 was a gift of Dr J. Weber (NCI).
Expression ofslrrfacedeterminants andofEB Vgenes. Exponen- tially growing human B cell lines and murine splenocytes ( IO7 cells/ mL in phosphate-buffered saline [PBS] containing 5% FCS) were incubated for 30 minutes at 4°C with control reagents or mono- clonal antibodies (MoAbs). These included the murine antihuman MoAbs anti-CD23 (anti-Leu 2 0 Becton Dickinson Immunocy- tometry Systems, San Jose, CA), anti-CD10 (J5: Coulter Electron- ics, Inc, Hialeah, FL), anti-CD1 8 (CLB-CD18; Janssen Biochimica, Beerse, Belgium), anti-CD20 (anti-Leu 16; Becton Dickinson), and antitransfemn receptor (Becton Dickinson); the antimurine rat MoAbs Thy I .2 (clone 30-H 12) and CD45R (clone RA3-6B2), both from Pharmingen (San Diego, CA). When appropriate, the cells were further incubated (30 minutes at 4‘C) with an affinity purified, fluorescein-labeled, goat antimouse reagent (Becton Dickinson). After washing, the cells were analyzed on an FACScan (Becton Dickinson). For each determination, IO4 cells were examined.
The EBV-encoded nuclear proteins EBNAI and EBNA2 were detected by Western blotting of cell lysates, using a human serum with high antibody titers to these antigens, as described.”
Cell cultures. Splenocytes from 6- to 8-week old female BALB/ c nu/nu mice were incubated for 5 days (2 X lo6 cells/mL in 24-well tissue culture plates (Limbro; Flow Laboratories, Inc, McLean, VA) in complete mouse cell culture medium consisting of a 1: 1 mixture of RPMI-1640 medium (GIBCO) and enriched Eagle’s medium (Biofluids, Inc, Rockville, MD) supplemented with 10% FCS (Re- heis) 2 mmol/L L-glutamine (GIBCO), mol/L 2-mercaptoeth- anol (GIBCO), 5 pg/mLgentamicin (Sigma), and IL-2 (500 U/mL: a gift of Cetus Corp. Emeryville, CA), as described.” Tumors were fragmented into single cell suspensions and cultured (2 X IO6 cells/ mL in 24-well tissue culture plates) in complete mouse cell culture medium.
Assaysfor IL-6, TNF, and interferon (IFN) bioactivities. The murine hybridoma cell line B9 (a gift of Dr R. Nordan, National Institutes of Health, Bethesda, MD) was used in a standard assay for IL-6 bioactivity.2’ One unit of IL-6 bioactivity in this assay is de- fined as the activity inducing half-maximal proliferation of B9 cells. An IL-6 concentration of I U/mL corresponds to approximately 20 pg of E. coli-derived human IL-6 used throughout as a laboratory standard (a gift of Sandoz Pharmaceuticals). Murine IL-6 was iden- tified by neutralization assays with a purified rabbit antiserum to highly purified natural murine IL-6I7 (agift of Dr R. Nordan, NCI). The fibroblast murine cell line L92 1 (a gift of Dr T. Gerrard, Center for Biologics Evaluation and Research) was used in a standard assay for TNF bioactivity.22 One unit of TNF bioactivity in this assay is defined as the activity inducing one-half maximal lysis of L921 targets. A TNF concentration of I U/mL corresponds to approxi- mately 100 pg of a recombinant human TNF-a preparation (a gift ofGenentech, Inc). Murine TNF-a was identified by neutralization assays with a purified rabbit antimurine TNF-a antiserum (rabbit IgG antirecombinant murine TNF-a; a gift of Dr J. Weber, NCI).I8 At the concentration of 20 &mL, this antiserum neutralized ap- proximately 50 ng/mL recombinant murine TNF-a, as determined in the L92 I bioassay.
Murine and human interferon (IFN)-induced antiviral activity were quantified by cytopathic effect neutralization assays, as pre- viously described.22 The murine fibroblast L929 cell line was used for measuring murine I F N activity, and the human epithelial am- nion cell line WISH (American Type Culture Collection [ATCC], Rockville, MD) was used for human I F N testing. One unit of I F N is defined as the reciprocal of the dilution that inhibits viral replica- tion by 50%. Murine and human I F N reference standards were ob- tained from the NIH.
Cellprol~Jcration and cytoto.\-icily assa~?r. Cell proliferation was measured essentially as described.20 The Burkitt’s lymphoma cell lines JD38 and CA46 in exponential growth phase were washed and cultured in triplicate at varying cell densities (250 to 2,000 cells/ well, in 96-well flat bottom plates: Costar), with or without recom- binant murine TNF-(U ( I to 100 pg/mL), in RPM1 1640 medium containing 2 mmol/L L-glutamine (GIBCO), 5 pg/mL gentamicin (Sigma), and 10% FCS (Reheis). After 3 and 7 days of incubation, the cultures were pulsed with ’H-thymidine (0.5 &/well; New En- gland Nuclear, Boston, MA) during 4.5 hours. Results were ex- pressed as mean cpm * SD.
Cytotoxicity was measured using a 4.5-hour chromium release assay, as described.” Briefly, murine leukemia YAC- 1 cells (ATCC) as well as the Burkitt’s and EBV-immortalized B-cell lines, de- scribed above, in exponential growth phase were used as 5’C-labeled targets. Fresh or IL-2-activated murine splenocytes were used as effectors at varying concentrations. The percent specific cytotoxic- ity was calculated as: 100 X ([experimental release - spontaneous release]/[total release - spontaneous release]).
Histology evullrution. Representative samples of tumor tissue were fixed in IO% neutral buffered-formalin (Media Kitchen: NIH), blocked in paraffin, sectioned at 4 rm, and stained with hematoxy- lin and eosin.
RESULTS
In initial studies (Table I ) , irradiated athymic mice were injected subcutaneously with EBV-immortalized (VDS) cells alone (IO’ cells in 0.2 mL medium), with Burkitt’s lym- phoma ( JD38 or CA46) cells alone (IO’ cells in 0.2 mL me- dium), or with mixtures of equal numbers of EBV-immor- talized (VDS) and Burkitt’s lymphoma (JD38 or CA46) cells (IO’ cells each in 0.2 mL medium). In control experi- ments, 7 of I O mice who received the EBV-immortalized B cells alone developed tumors reaching sizes of 0.25 to 2.0 cm2 that rapidly regressed, and 16 of 16 mice receiving Bur- kitt’s B cells alone developed progressively growing tumors, none of which regressed. In contrast, 30 of 40 mice injected with mixtures of EBV-immortalized and Burkitt’s cells de- veloped tumors that reached sizes of l .3 to 2.6 cm2 and then all regressed through necrosis and scarring. Ofthe 30 tumor- bearing mice, 7 died of massive tumor-cell lysis and/or ca- chexia during tumor regression, but all others were cured (ie, the mice survived for a t least 50 days without tumor recurrence). These experiments showed that EBV-immor- talized cells induce an ant i tumor effect on Burkitt’s tumors in this model.
Using the same experimental system, we determined that injection of each of 4 additional EBV-immortalized B cell lines (two obtained by in vitro immortalization of normal B cells with EBV, and two by spontaneous immortalization of normal peripheral blood B cells) mixed with Burkitt’s cells (JD38 line) resulted in tumor regression in 100% of cases. In control experiments, no tumor regression occurred when CA46 Burkitt’s cells, normal peripheral blood T cells (IO’ cells in 0.2 mL medium), or 100 ng LPS were coinjected with JD38 Burkitt’s cells (not shown). Thus, induction of Burkitt’s t umor regression appears to be a common prop- erty of EBV-immortalized B cells.
In further experiments, we examined whether the EBV- immortalized cells and the Burkitt’s lymphoma cells had to be injected into the same subcutaneous si te to induce the ant i tumor effect (Table 2). Subcutaneous injection of EBV-
778
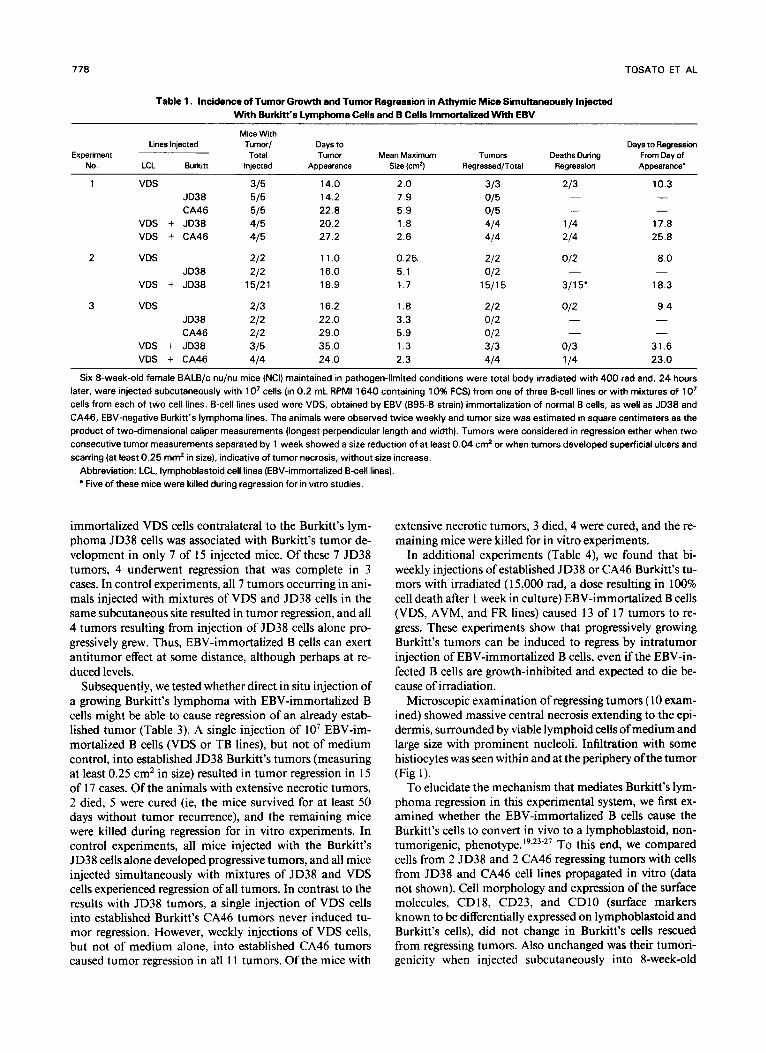
Table 1. Incidence of Tumor Growth and Tumor Regression in Athymic Mice Simultaneously Injected With Burkitt’s Lymphoma Cells and B Cells Immortalized Wittr EBV
TOSATO ET AL
Mice With Lines Injected Tumor/ Days to Days to Regression
Total No. LCL Burkitt
Tumor Mean Maximum Tumors Deaths During From Day of Injected Appearance Size (cm2) Regressed/Total Regression Appearance’
Experiment
1 VDS
VDS VDS
2 VDS
VDS
3 VDS
VDS VDS
JD38 CA46
+ JD38 + CA46
JD38 + JD38
JD38 CA46
+ JD38 + CA46
14.0 14.2 22.8 20.2 27.2
1 1 .o 16.0 18.9
16.2 22.0 29.0 35.0 24.0
2.0 7.9 5.9 1.8 2.6
0.25. 5.1 1.7
1.8 3.3 5.9 1.3 2.3
013 114
10.3 - -
17.8 25.8
8.0
18.3
9.4
-
-
31.6 23.0
-
Six 8-week-old female BALB/c nu/nu mice (NCI) maintained in pathogen-limited conditions were total body irradiated with 400 rad and, 24 hours later, were injected subcutaneously with 1 O7 cells (in 0.2 mL RPM1 1640 containing 10% FCS) from one of three B-cell lines or with mixtures of 1 O7 cells from each of two cell lines. B-cell lines used were VDS, obtained by EBV (895-8 strain) immortalization of normal B cells, as well as JD38 and CA46, EBV-negative Burkitt’s lymphoma lines. The animals were observed twice weekly and tumor size was estimated in square centimeters as the product of two-dimensional caliper measurements (longest perpendicular length and width). Tumors were considered in regression either when two consecutive tumor measurements separated by 1 week showed a size reduction of at least 0.04 cm2 or when tumors developed superficial ulcers and scarring (at least 0.25 mm2 in size), indicative of tumor necrosis, without size increase.
Abbreviation: LCL, lymphoblastoid cell lines (EBV-immortalized B-cell lines). Five of these mice were killed during regression for in vitro studies.
immortalized VDS cells contralateral to the Burkitt’s lym- phoma JD38 cells was associated with Burkitt’s tumor de- velopment in only 7 of 15 injected mice. Of these 7 JD38 tumors, 4 underwent regression that was complete in 3 cases. In control experiments, all 7 tumors occurring in ani- mals injected with mixtures of VDS and JD38 cells in the same subcutaneous site resulted in tumor regression, and all 4 tumors resulting from injection of JD38 cells alone pro- gressively grew. Thus, EBV-immortalized B cells can exert antitumor effect at some distance, although perhaps at re- duced levels.
Subsequently, we tested whether direct in situ injection of a growing Burkitt’s lymphoma with EBV-immortalized B cells might be able to cause regression of an already estab- lished tumor (Table 3). A single injection of lo7 EBV-im- mortalized B cells (VDS or TB lines), but not of medium control, into established JD38 Burkitt’s tumors (measuring at least 0.25 cm2 in size) resulted in tumor regression in 15 of 17 cases. Of the animals with extensive necrotic tumors, 2 died, 5 were cured (ie, the mice survived for at least 50 days without tumor recurrence), and the remaining mice were killed during regression for in vitro experiments. In control experiments, all mice injected with the Burkitt’s JD38 cells alone developed progressive tumors, and all mice injected simultaneously with mixtures of JD38 and VDS cells experienced regression of all tumors. In contrast to the results with JD38 tumors, a single injection of VDS cells into established Burkitt’s CA46 tumors never induced tu- mor regression. However, weekly injections of VDS cells, but not of medium alone, into established CA46 tumors caused tumor regression in all l 1 tumors. Of the mice with
extensive necrotic tumors, 3 died, 4 were cured, and the re- maining mice were killed for in vitro experiments.
In additional experiments (Table 4), we found that bi- weekly injections of established JD38 or CA46 Burkitt’s tu- mors with irradiated ( 1 5,000 rad, a dose resulting in 100% cell death after 1 week in culture) EBV-immortalized B cells (VDS, AVM, and FR lines) caused 13 of 17 tumors to re- gress. These experiments show that progressively growing Burkitt’s tumors can be induced to regress by intratumor injection of EBV-immortalized B cells, even if the EBV-in- fected B cells are growth-inhibited and expected to die be- cause of irradiation.
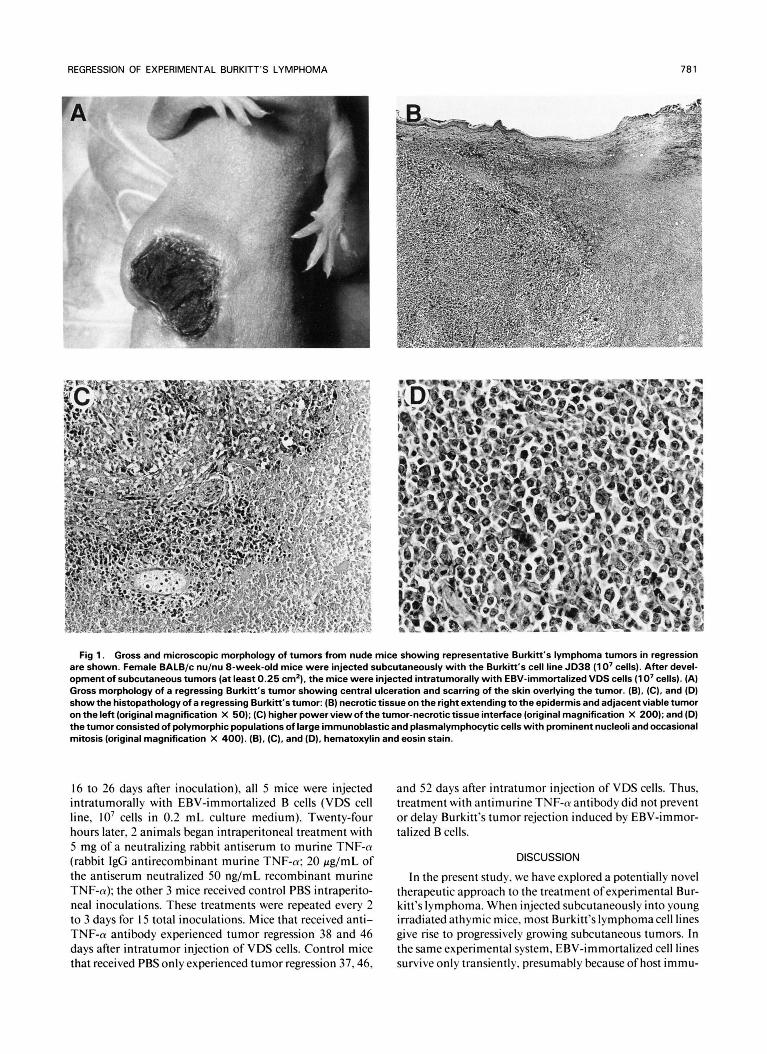
Microscopic examination of regressing tumors ( 10 exam- ined) showed massive central necrosis extending to the epi- dermis, surrounded by viable lymphoid cells of medium and large size with prominent nucleoli. Infiltration with some histiocytes was seen within and at the periphery ofthe tumor (Fig 1).
To elucidate the mechanism that mediates Burkitt’s lym- phoma regression in this experimental system, we first ex- amined whether the EBV-immortalized B cells cause the Burkitt’s cells to convert in vivo to a lymphoblastoid, non- tumorigenic, phenotype.’9923-27 To this end, we compared cells from 2 JD38 and 2 CA46 regressing tumors with cells from JD38 and CA46 cell lines propagated in vitro (data not shown). Cell morphology and expression of the surface molecules, CD 18, CD23, and CD10 (surface markers known to be differentially expressed on lymphoblastoid and Burkitt’s cells), did not change in Burkitt’s cells rescued from regressing tumors. Also unchanged was their tumori- genicity when injected subcutaneously into 8-week-old
REGRESSION OF EXPERIMENTAL BURKITT‘S LYMPHOMA 779
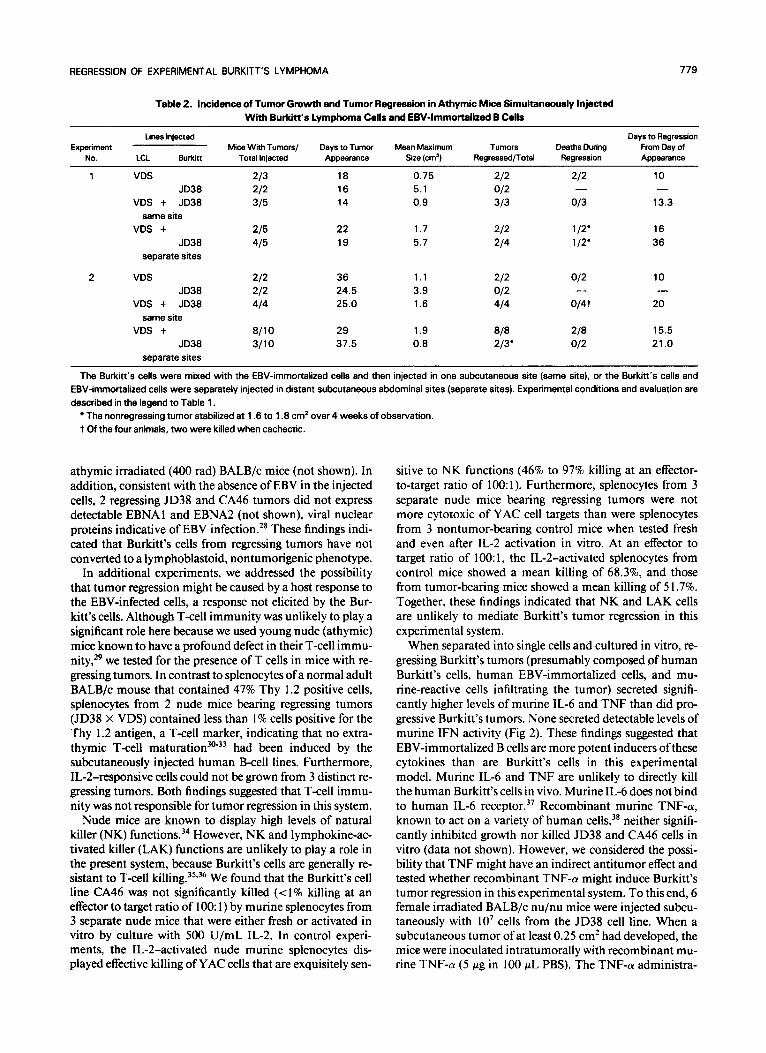
Table 2. Incidence of Tumor Growth and Tumor Regression in Athymic Mice Simultaneously Injected With Burkitt’s Lymphoma Cells and EBV-Immortalized B Cells
tines Injected Days to Regression Experiment Mice With Tumors/ Days to Tumor Mean Maximum Tumors Deaths During From Day of
NO. LCL Burk i Total Injected Appearance Size (cm’) Regrassedflotal Regression Appearance
1 VDS JD38
VDS + JD38 same site
VDS + JD38
separate sites
2 VDS JD38
VDS + JD38 same site
VDS + JD38
separate sites
18 0.75 16 5.1 14 0.9
22 1.7 19 5.7
36 1.1 24.5 3.9 25.0 1.6
29 1.9 37.5 0.8
10 -
13.3
16 36
10
20 -
15.5 21 .o
The Burkitt’s cells were mixed with the EBV-immortalized cells and then injected in one subcutaneous site (same site). or the Burkitt’s cells and EBV-immortalized cells were separately injected in distant subcutaneous abdominal sites (separate sites). Experimental conditions and evaluation are described in the legend to Table 1.
The nonregressing tumor stabilized at 1.6 to 1.8 cm2 over 4 weeks of observation. t Of the four animals, two were killed when cachectic.
athymic irradiated (400 rad) BALB/c mice (not shown). In addition, consistent with the absence of EBV in the injected cells, 2 regressing JD38 and CA46 tumors did not express detectable EBNAl and EBNA2 (not shown), viral nuclear proteins indicative of EBV infection.” These findings indi- cated that Burkitt’s cells from regressing tumors have not converted to a lymphoblastoid, nontumorigenic phenotype.
In additional experiments, we addressed the possibility that tumor regression might be caused by a host response to the EBV-infected cells, a response not elicited by the Bur- kitt’s cells. Although T-cell immunity was unlikely to play a significant role here because we used young nude (athymic) mice known to have a profound defect in their T-cell immu- nit^,'^ we tested for the presence of T cells in mice with re- gressing tumors. In contrast to splenocytes of a normal adult BALB/c mouse that contained 47% Thy 1.2 positive cells, splenocytes from 2 nude mice bearing regressing tumors (JD38 X VDS) contained less than 1% cells positive for the Thy 1.2 antigen, a T-cell marker, indicating that no extra- thymic T-cell had been induced by the subcutaneously injected human B-cell lines. Furthermore, IL-2-responsive cells could not be grown from 3 distinct re- gressing tumors. Both findings suggested that T-cell immu- nity was not responsible for tumor regression in this system.
Nude mice are known to display high levels of natural killer (NK) functions.34 However, NK and lymphokine-ac- tivated killer (LAK) functions are unlikely to play a role in the present system, because Burkitt’s cells are generally re- sistant to T-cell killing.35,36 We found that the Burkitt’s cell line CA46 was not significantly killed (< l% killing at an effector to target ratio of 100: 1) by murine splenocytes from 3 separate nude mice that were either fresh or activated in vitro by culture with 500 U/mL IL-2. In control experi- ments, the IL-2-activated nude murine splenocytes dis- played effective killing of YAC cells that are exquisitely sen-
sitive to NK functions (46% to 97% killing at an effector- to-target ratio of lO0:l). Furthermore, splenocytes from 3 separate nude mice bearing regressing tumors were not more cytotoxic of YAC cell targets than were splenocytes from 3 nontumor-bearing control mice when tested fresh and even after IL-2 activation in vitro. At an effector to target ratio of 100: 1, the IL-2-activated splenocytes from control mice showed a mean killing of 68.3%, and those from tumor-bearing mice showed a mean killing of 5 1.7%. Together, these findings indicated that NK and LAK cells are unlikely to mediate Burkitt’s tumor regression in this experimental system.
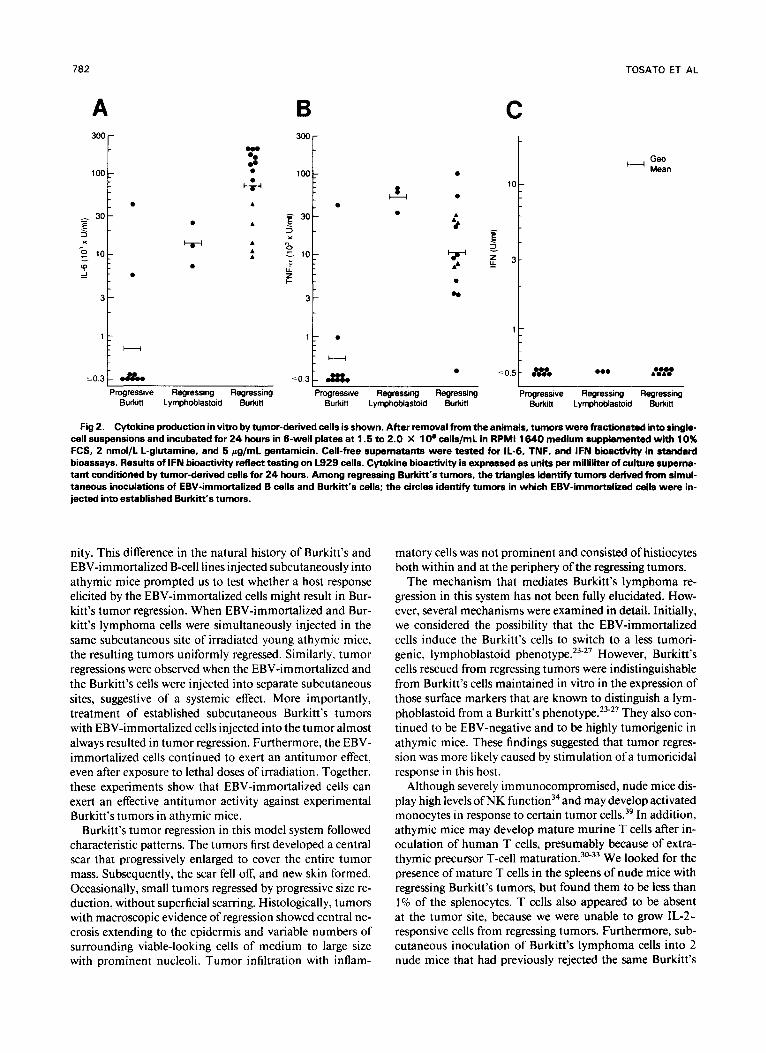
When separated into single cells and cultured in vitro, re- gressing Burkitt’s tumors (presumably composed of human Burkitt’s cells, human EBV-immortalized cells, and mu- rine-reactive cells infiltrating the tumor) secreted signifi- cantly higher levels of murine IL-6 and TNF than did pro- gressive Burkitt’s tumors. None secreted detectable levels of murine IFN activity (Fig 2). These findings suggested that EBV-immortalized B cells are more potent inducers ofthese cytokines than are Burkitt’s cells in this experimental model. Murine IL-6 and TNF are unlikely to directly kill the human Burkitt’s cells in vivo. Murine IL-6 does not bind to human IL-6 Recombinant murine TNF-a, known to act on a variety of human cells,38 neither signifi- cantly inhibited growth nor killed JD38 and CA46 cells in vitro (data not shown). However, we considered the possi- bility that TNF might have an indirect antitumor effect and tested whether recombinant TNF-a might induce Burkitt’s tumor regression in this experimental system. To this end, 6 female irradiated BALB/c nu/nu mice were injected subcu- taneously with IO7 cells from the JD38 cell line. When a subcutaneous tumor of at least 0.25 cm2 had developed, the mice were inoculated intratumorally with recombinant mu- rine TNF-a (5 pg in 100 pL PBS). The TNF-a administra-
780 TOSATO ET AL
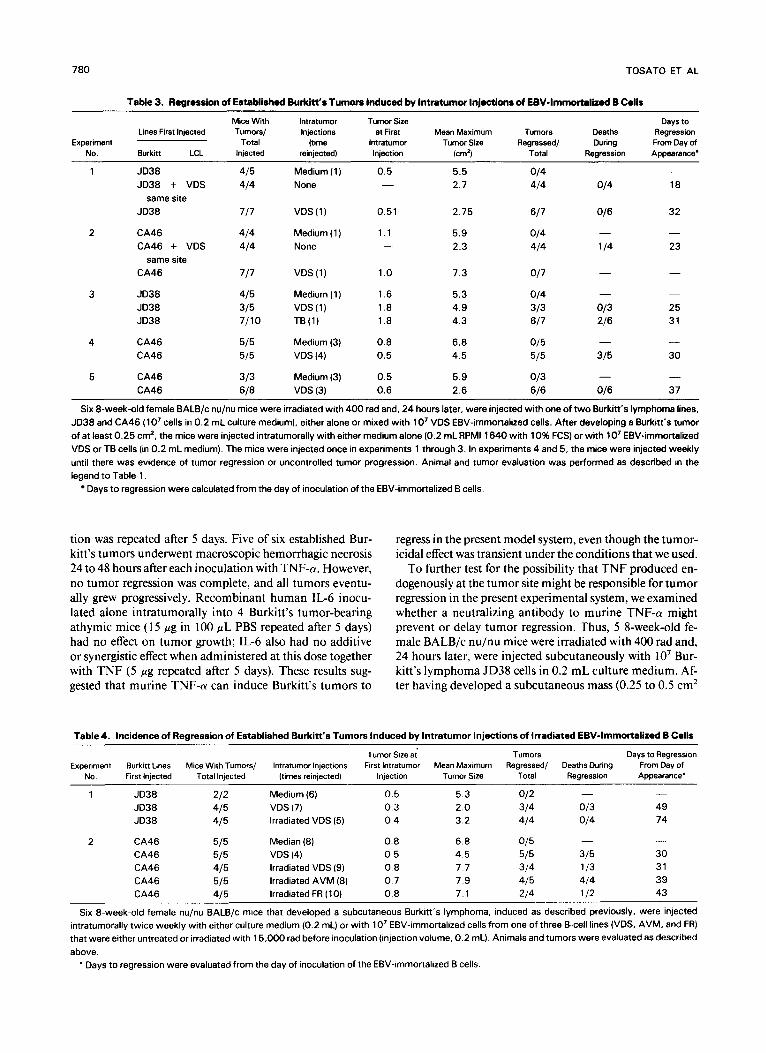
Table 3. Regression of EstaMihd Burkii’s Tumors Induced by lntratumor Inioctions of EBV-Immortal id B Cells
Mice With lntratumor Tumor Size Lines First Injected Tumors/
Days to
Experiment Total Injections at First
(time lntratumor Mean Maximum Tumors Deaths
Tumor Size Regressed/ Regression
No. Burkitt LCL Injected reinjected) Injection (Cm2) Total Regression Appearance’ During From Day of
1 JD38 JD38 + VDS
same site JD38
2 CA46 CA46 + VDS
same site CA46
3 JD38 JD38 JD38
4 CA46 CA46
5 CA46 CA46
Medium (1 ) None
0.5 -
5.5 2.7
- 18
VDS(1)
Medium (1 ) None
0.51
1.1 -
2.75
5.9 2.3
32
23
VDS (1)
Medium (1) VDS (1) TB(1)
Medium (3) VDS (4)
Medium (3) VDS (3)
1 .o
1.6 1.8 1.8
0.8 0.5
0.5 0.6
7.3
5.3 4.9 4.3
6.8 4.5
5.9 2.6
- 25 31
315
- - 30
- 37
Six 8-week-old female BALB/c nu/nu mice were irradiated with 400 rad and, 24 hours later, were injected with one of two Burkitt’s lymphoma lines, JD38 and CA46 ( lo7 cells in 0.2 mL culture medium), either alone or mixed with 1 O7 VDS EBV-immortalized cells. After developing a Burkitt‘s tumor of at least 0.25 cm2, the mice were injected intratumorally with either medium alone (0.2 mL RPM1 1640 with 10% FCS) or with lo7 EBV-immortalized VDS or TB cells (in 0.2 mL medium). The mice were injected once in experiments 1 through 3. In experiments 4 and 5, the mice were injected weekly until there was evidence of tumor regression or uncontrolled tumor progression. Animal and tumor evaluation was performed as described in the legend to Table l .
Days to regression were calculated from the day of inoculation of the EBV-immortalized B cells.
tion was repeated after 5 days. Five of six established Bur- kitt’s tumors underwent macroscopic hemorrhagic necrosis 24 to 48 hours after each inoculation with TNF-a. However, no tumor regression was complete, and all tumors eventu- ally grew progressively. Recombinant human IL-6 inocu- lated alone intratumorally into 4 Burkitt’s tumor-bearing athymic mice ( 15 p g in 100 pL PBS repeated after 5 days) had no effect on tumor growth; IL-6 also had no additive or synergistic effect when administered at this dose together with TNF (5 pg repeated after 5 days). These results sug- gested that murine TNF-a can induce Burkitt’s tumors to
regress in the present model system, even though the tumor- icidal effect was transient under the conditions that we used.
To further test for the possibility that TNF produced en- dogenously at the tumor site might be responsible for tumor regression in the present experimental system, we examined whether a neutralizing antibody to murine TNF-a might prevent or delay tumor regression. Thus, 5 8-week-old fe- male BALB/c nu/nu mice were irradiated with 400 rad and, 24 hours later, were injected subcutaneously with IO’ Bur- kitt’s lymphoma JD38 cells in 0.2 mL culture medium. Af- ter having developed a subcutaneous mass (0.25 to 0.5 cmz
Table 4. Incidence of Regression of Established Burkitt’s Tumors Induced by lntratumor Injections of Irradiated EBV-lmmortaliied B Cells
Tumor Size at Expermant Burkitt Lines Mice With Tumors/ lntratumor Injections First lntratumor Mean Maxlmum Regressed/ Deaths During From Day of
No. First Injected Total Injected (times reinjected) Injection Tumor Sire Total Regression Appearance’
Tumors Days to Regression
1 JD38 JD38 JD38
2 CA46 CA46 CA46 CA46 CA46
Medium (6) VDS (7) Irradiated VDS (5)
Median (8) VDS (4) Irradiated VDS (9) Irradiated AVM (8) Irradiated FR ( l 0)
0.5 0.3 0.4
0.8 0.5 0.8 0.7 0.8
5.3 2.0 3.2
6.8 4.5 7.7 7.9 7.1
- 49 74
- 30 31 39 43
Six 8-week-old female nu/nu BALB/c mice that developed a subcutaneous Burkitt‘s lymphoma, induced as described previously. were injected intratumorally twice weekly with either culture medium (0.2 mL) or with 10’ EBV-immortalized cells from one of three B-cell lines (VDS, AVM, and FR) that were either untreated or irradiated with 15,000 rad before inoculation (injection volume, 0.2 mL). Animals and tumors were evaluated as described above.
* Days to regression were evaluated from the day of inoculation of the EBV-immortalized 8 cells.
REGRESSION OF EXPERIMENTAL 8URKITT’S LYMPHOMA 781
Fig 1. Gross and microscopic morphology of tumors from nude mice showing representative Burkitt‘s lymphoma tumors in regression are shown. Female BALB/c nu/nu 8-week-old mice were injected subcutaneously with the Burkitt’s cell line JD38 (1 0’ cells). After devel- opment of subcutaneous tumors (at least 0.25 cm’), the mice were injected intratumorally with EBV-immortalized VDS cells (1 0’ cells). (A) Gross morphology of a regressing Burkitt’s tumor showing central ulceration and scarring of the skin overlying the tumor. (B), (C), and (D) show the histopathology of a regressing Burkitt’s tumor: (B) necrotic tissue on the right extending to the epidermis and adjacent viable tumor on the left (original magnification X 50); (C) higher power view of the tumor-necrotic tissue interface (original magnification X 200); and (D) the tumor consisted of polymorphic populations of large immunoblastic and plasmalymphocytic cells with prominent nucleoli and occasional mitosis (original magnification X 400). (B), (C), and (D), hematoxylin and eosin stain.
16 to 26 days after inoculation), all 5 mice were injected intratumorally with EBV-immortalized B cells (VDS cell line, IO’ cells in 0.2 mL culture medium). Twenty-four hours later, 2 animals began intraperitoneal treatment with 5 mg of a neutralizing rabbit antiserum to murine TNF-a (rabbit IgG antirecombinant murine TNF-a: 20 g / m L of the antiserum neutralized 50 ng/mL recombinant murine TNF-a); the other 3 mice received control PBS intraperito- neal inoculations. These treatments were repeated every 2 to 3 days for I5 total inoculations. Mice that received anti- TNF-a antibody experienced tumor regression 38 and 46 days after intratumor injection of VDS cells. Control mice that received PBS only experienced tumor regression 37.46.
and 52 days after intratumor injection of VDS cells. Thus, treatment with antimurine TNF-a antibody did not prevent or delay Burkitt’s tumor rejection induced by EBV-immor- talized B cells.
DISCUSSION
In the present study, we have explored a potentially novel therapeutic approach to the treatment ofexperimental Bur- kitt’s lymphoma. When injected subcutaneously into young irradiated athymic mice, most Burkitt’s lymphomacell lines give rise to progressively growing subcutaneous tumors. In the same experimental system, EBV-immortalized cell lines survive only transiently, presumably because of host immu-
782
B
TOSATO ET AL
C A 300 -
100:
- 30 3
- E -
5 10:
2 :
x
-
3 -
1 -
m
W
m
t
H Geo Mean
H
m
t
1 H [ ;o,S[ tHm *mm ..t..t Progressive Regressing Regressing Progresswe Regressing Regressing Progressive Regressing Regressing
Burkin Lyrnphoblastold Burkitt Burkin Lyrnphoblastoid Burkin
10.3 &mm 50.3 a& Burkitl Lymphoblastoid Burkin
Fig 2. Cytokine production in vitro by tumor-derived cells is shown. After removal from the animals, tumors were fractionated into single- cell suspensions and incubated for 24 hours in 6-well plates at 1.5 to 2.0 X 10’ cells/mL in RPM1 1640 medium supplemented with 10% FCS, 2 nmol/L L-glutamine, and 5 rg/mL gentamicin. Cell-free supernatants were tested for 11-6, TNF, and IFN bioactivity in standard bioassays. Results of IFN bioactivity reflect testing on L929 cells. Cytokine bioactivity is expressed as units per milliliter of culture superna- tant conditioned by tumor-derived cells for 24 hours. Among regressing Burkii‘s tumors, the triangles identify tumors derived from simul- taneous inoculations of EBV-immortalized B cells and Burkitt‘s cells; the circles identify tumors in which EBV-immortalized cells were in- jected into established Burkitt’s tumors.
nity. This difference in the natural history of Burkitt’s and EBV-immortalized B-cell lines injected subcutaneously into athymic mice prompted us to test whether a host response elicited by the EBV-immortalized cells might result in Bur- kitt’s tumor regression. When EBV-immortalized and Bur- kitt’s lymphoma cells were simultaneously injected in the same subcutaneous site of irradiated young athymic mice, the resulting tumors uniformly regressed. Similarly, tumor regressions were observed when the EBV-immortalized and the Burkitt’s cells were injected into separate subcutaneous sites, suggestive of a systemic effect. More importantly, treatment of established subcutaneous Burkitt’s tumors with EBV-immortalized cells injected into the tumor almost always resulted in tumor regression. Furthermore, the EBV- immortalized cells continued to exert an antitumor effect, even after exposure to lethal doses of irradiation. Together, these experiments show that EBV-immortalized cells can exert an effective antitumor activity against experimental Burkitt’s tumors in athymic mice.
Burkitt’s tumor regression in this model system followed characteristic patterns. The tumors first developed a central scar that progressively enlarged to cover the entire tumor mass. Subsequently, the scar fell off, and new skin formed. Occasionally, small tumors regressed by progressive size re- duction, without superficial scarring. Histologically, tumors with macroscopic evidence of regression showed central ne- crosis extending to the epidermis and variable numbers of surrounding viable-looking cells of medium to large size with prominent nucleoli. Tumor infiltration with inflam-
matory cells was not prominent and consisted of histiocytes both within and at the periphery of the regressing tumors.
The mechanism that mediates Burkitt’s lymphoma re- gression in this system has not been fully elucidated. How- ever, several mechanisms were examined in detail. Initially, we considered the possibility that the EBV-immortalized cells induce the Burkitt’s cells to switch to a less tumori- genic, lymphoblastoid p h e n ~ t y p e . ~ ~ - ’ ~ However, Burkitt’s cells rescued from regressing tumors were indistinguishable from Burkitt’s cells maintained in vitro in the expression of those surface markers that are known to distinguish a lym- phoblastoid from a Burkitt’s phen~type.‘~-*~ They also con- tinued to be EBV-negative and to be highly tumorigenic in athymic mice. These findings suggested that tumor regres- sion was more likely caused by stimulation of a tumoricidal response in this host.
Although severely immunocompromised, nude mice dis- play high levels of NK function34 and may develop activated monocytes in response to certain tumor cells.39 In addition, athymic mice may develop mature murine T cells after in- oculation of human T cells, presumably because of extra- thymic precursor T-cell mat~ration.~’”~ We looked for the presence of mature T cells in the spleens of nude mice with regressing Burkitt’s tumors, but found them to be less than 1% of the splenocytes. T cells also appeared to be absent at the tumor site, because we were unable to grow IL-2- responsive cells from regressing tumors. Furthermore, sub- cutaneous inoculation of Burkitt’s lymphoma cells into 2 nude mice that had previously rejected the same Burkitt’s

REGRESSION OF EXPERIMENTAL BURKITT’S LYMPHOMA 783
tumor after intratumor injections with EBV-immortalized cells resulted in the appearance of progressively growing tu- mors (preliminary results not shown). This suggests that tu- mor regression in this system is not associated with T-cell memory.
In subsequent experiments, we explored the role of NK cells in Burkitt’s tumor regression. However, we found that one of the Burkitt’s cell lines used here, CA46, was highly resistant to killing from fresh nude splenocytes. This cell line was also resistant to killing by IL-2-activated nude spleno- cytes. In addition, mice with regressing Burkitt’s tumors did not show increased splenocyte killing of NK-sensitive targets. Thus, it appeared unlikely that either T cells or NK cells were involved in tumor regression in the present sys- tem.
Cell suspensions derived from regressing Burkitt’s tumors were found to secrete significantly higher levels of murine IL-6 and TNF-a compared with those from progressive tu- mors. This suggested that EBV-immortalized cells are more potent inducers of inflammatory cytokines than are Bur- kitt’s cells, and prompted us to examine whether IL-6 and/ or TNF-a might be involved in tumor regression. A direct effect of the murine cytokines on the human Burkitt’s cells appeared unlikely because murine IL-6 does not bind to the human IL-6 re~eptor,~’ and recombinant murine TNF-a neither inhibited the growth nor killed Burkitt’s cells in vi- tro. However, TNF-a is known to cause intravascular thrombosis and, secondarily, tumor tissue All the regressing tumors studied here had central necrosis oc- casionally surrounding a blood vessel, suggesting that tumor necrosis may be related to tissue ischemia. In addition, we observed that 5 of 5 established Burkitt’s tumors underwent macroscopic necrosis after 1 or 2 intratumor inoculations of 5 p g recombinant murine TNF-a. In contrast, IL-6 was ineffective alone and appeared not to increase the tumorici- dal effect of exogenous TNF-a. Although the effects of exog- enous TNF-a injection were transient, it was possible that endogenously produced TNF at the tumor site, over a pro- longed period of time, might have been responsible for Bur- kitt’s tumor regression. However, multiple intraperitoneal injections of a neutralizing antiserum to murine TNF-a failed to appreciably affect tumor regression. Although TNF may be one of several factors involved, a role for TNF in Burkitt’s tumor regression induced by EBV-immortalized B cells remains uncertain.
As discussed above, initial necrosis characterizes tumor regression in this system, suggesting that tissue ischemia may be central to this process. We now believe that factors regulating vascular endothelium growth and survival at the tumor site may be responsible for Burkitt’s lymphoma’s progressive growth and regression in this system. Our pres- ent understanding ofthe intricacies of vascular endothelium biology is still limited, but this is an increasingly active area of investigation. Elucidation of these processes is essential for understanding Burkitt’s tumor growth and regression in this system.
With improved chemotherapeutic and adjuvant thera- pies, Burkitt’s lymphoma is cured in 70% to 80% of all case^.^',^^ However, B-cell lymphomas in acquired immu- nodeficiency syndrome, including Burkitt’s lymphoma,
have a median survival of 2.5 months despite the rap^.^' Al- though further study will be necessary to clarify the mecha- nisms for the observed antitumor effects, these data support the potential cIinical application of this approach for treat- ment of Burkitt’s tumors not amendable to other treat- ments.
ACKNOWLEDGMENT
We thank L. Diaz for technical support; Dr K. Zoon for help with interferon assays; Drs J. Weber and J. Mule for providing antimu- rine TNF-a antiserum; Cenentech for providing recombinant mu- rine TNF-a; Sandoz Pharmaceuticals for providing recombinant human IL-6; Drs M. Blaese. R. Yarchoan, and A. Angiolillo for helpful discussions; and E. Caruso for editing the manuscript.
REFERENCES
I . Tepper RI, Pattengate PK, Leder P: Murine interleukin-4 dis- plays potent anti-tumor activity in vitro. Cell 57503, 1989
2. Watanabe Y, Kuribayashi K, Miyatake S , Nishihara K, Na- kayama E, Taniyama T, Sakata T: Exogenous expression of mouse interferon y cDNA in mouse neuroblastoma Cl 300 cells results in reduced tumorigenicity by augmented anti-tumor immunity. Proc Natl Acad Sci USA 86:9456, 1989
3. Fearon ER, Pardoll DM, ltaya T: Interleukin-2 production by tumor cells bypasses T helper function in the generation ofan anti- tumor response. Cell 60:397, 1990
4. Gansbacher B, Bonnerji R, Daniels B, Zier K, Cronin K, Gil- boa E: Retroviral vector-mediated y interferon gene transfer in tu- mor cells generates potent and long lasting anti-tumor immunity. Cancer Res 50:7820, 1990
5. Gansbacher B, Zier K, Daniels B, Cronin K, Bannerji R, Gil- boa E: Interleukin-2 gene transfer into tumor cells abrogates tumor- igenicity and induces protective immunity. J Exp Med 172: 12 17, 1990
6. Lay V, Roth C, Langlade-Demoyen P, Larsson-Sciard EL, Kourilsky P A novel approach to the induction of specific cytolytic Tcells in vivo. ResImmunol 141:855, 1990
7. Columbo MP, Ferrari G, Stopaccino A, Parenza M, Rodolfo M, Mavilio F, Parmiani G: Granulocyte colony-stimulating factor gene transfer suppressed tumorigenicity of a murine adenocarci- noma in vivo. J Exp Med I73:889, I991
8. Vanhaesebroeck B, Mareel M, Van Roy F, Grooten J, Fiers W: Expression of the tumor necrosis factor gene in tumor cells cor- relates with reduced tumorigenicity and reduced invasiveness in vi- fro. Cancer Res 51:2229, 1991
9. Blankenshein T, Gin Z, Uberla K, Muller W, Rosen H, Volk H-D, Diamantstein T: Tumor suppression after tumor-cell targeted tumor necrosis factor alpha gene transfer. J Exp Med 173: 1047, 1991
IO. Culver KW, Ram Z , Wallbridge S, Ishii H, Oldfield EH, Blaese RM: In vivo gene transfer with retroviral vector-producer cells for treatment ofexperimental brain tumors. Science 256: 1550, I992
1 1 . Gutierrez AA, Lemoione NR, Sikora K: Gene therapy of cancer. Lancet 339:715, 1992
12. Nilsson K, Giovanella BC, Stehlin JS, Klein G: Tumorige- nicity of human hematopoietic cell lines in athymic nude mice. Int J Cancer 19:337. 1977
13. Walter J, Miller P, Moldenhauer G, Schirrmacher V, Pawlita M, Wolf J: Local growth of a Burkitt’s lymphoma versus dissemi- nated invasive growth of the autologous EBV-immortalized lym- phoblastoid cells and their somatic cell hybrids in SCID mice. Int J Cancer 50:265, 1992
14. Gursevitch VE. OConnor GT. Lenoir GM: Burkitt’s lym-

784 TOSATO ET AL
phoma cell lines reveal different degrees of tumorigenicity in nude mice. Int J Cancer 41537, 1987
15. Tosato G, Blaese RM, Yarchoan R: Relationship between immunoglobulin production and immortalization by Epstein-Barr virus. J Immunol 135:959, 1985
16. Shiramizu B, Barriga F, Neequaye J, Jafri A, Dalla-Favera R. Nieri A, Guttierez M, Levine P, Magrath I: Patterns of chromo- somal breakpoint locations in Burkitt’s lymphoma: Relevance to geography and Epstein-Barr virus association. Blood 77: I5 16. 199 1
17. Hilbert DM, Cancro MP, Scherie PA, Nordan RP, Van Snick J, Gerhard W, Rudikoff S: T-cell-derived IL-6 is differentially required for antigen-specific antibody secretion by primary and sec- ondary Bcells. J Immunol 143:4019, 1989
18. Doherty GM, Lange JR, Langstein HN, Alexander HR, Bu- resh CM, Norton JA: Evidence for IFN-r as a mediator of the le- thality of endotoxin and tumor necrosis factor-tu. J lmmunol 149: 1666, 1992
19. Rowe M, Rowe DT, Gregory CD, Young LS, Farrell PJ, Ru- pani H, Rickinson AB: Differences in B cell growth phenotype re- flect novel patterns of Epstein-Barr virus latent gene expression in Burkitt’s lymphoma. EMBO J 6:2743, 1987
20. Tanner J, Tosato G: Impairment of natural killer functions by interleukin-6 increases lymphoblastoid cell tumorigenicity in athymic mice. J Clin Invest 88:239, 1991
21. Aarden LA, DeGroot ER, Schaap DL, Lansdorp PM: Pro- duction of hybridoma growth factor by human monocytes. Eur J lmmunol 17:141 I . 1987
22. Vogel SN, Friedman RM: Measurement ofantiviral activity induced by interferons a, p, and y. in Coligan JE, Kruisbeek AM, Margulies DH, Shevach EM, Strober W (eds): Current Protocols in Immunology (ed I ). New York. NY, Greene and Wiley, 1991. p 6.9.1
23. Nilsson K, Klein G: Phenotypic and cytogenetic characteris- tics of human B lymphoid cell lines and their relevance for the eti- ology of Burkitt‘s lymphoma. Adv Cancer Res 37:3 19, 1982
24. Rowe M, Ronney CM, Edwards CF. Lenoir GM, Rickinson AB: Epstein-Barr virus status and tumor cell phenotype in sporadic Burkitt’s lymphoma. Int J Cancer 37:367, 1985
25. Ehlin-Hendriksson B, Manneborg-Sondlund A, Klein G: Expression of B-cell specific markers in different Burkitt lymphoma subgroups. Int J Cancer 39:21 1. 1987
26. Rooney CM, Gregory CD, Rowe M, Finerty S, Edwards C. Rupani H, Rickinson AB: Endemic Burkitt’s lymphoma: Pheno- typic analysis of tumor biopsy cells and of derived tumor cell lines. J Natl Cancer Inst 77:68 I , 1986
27. Patarroyo M, Prieto J, Ernberg 1, Gahmberg CC: Absence or low expression of leukocyte-adhesion molecules CD1 1 and CD18 on Burkitt’s lymphoma cells. Int J Cancer 4 I :90 l , 1988
28. Kieff E, Leibowitz D: The Epstein-Barr virus, in Fields S, Knipe DM (eds): Fields Virology (ed 2). New York, NY, Raven, 1990, p 1889
29. Pantelouris EM: Observations on the immunobiology of nude mice. Immunology 20:247, 197 1
30. Scheid MP, Goldstein G, Boyse EA: Antigen specificity of cytotoxic T cells from athymic mice. Science 190: 12 1 I , 1975
3 I . Humig T. Bevan MJ: Antigen specificity of cytotoxic T cells from athymic mice. J Exp Med 152:688, 1980
32. LeesA, Glasebrook L, Sordat B: Interleukin-2 production by
lymphoid cells from congenitally athymic mice. J Immunol 129: 521, 1982
33. Dosch H-M, White D, Grant C: Reconstitution of nude mouse T cell function in vivo; IL-2 independent effects of human T cells. J Immunol 134:336, 1985
34. Kiessling R, Klein G, Pross H, Wigzell H: Natural killer cells in the mouse. 11. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Characteristics of the killer cells. Eur J lmmunol 5 : 117, 1975
35. McCormick KJ, Giovanella BC, Klein G, Nilsson K, Stehlin JS: Diploid human lymphoblastoid and Burkitt’s lymphoma cell lines: Susceptibility to murine NK cells and heterotransplantation to nude mice. Int J Cancer 28:455, 1981
36. Gregory CD, Murray RM, Edwards CF, Rickinson AB: Downregulation of cell adhesion molecules LFA-3 and ICAM- I in Epstein-Barr virus positive Burkitt’s lymphoma underlies tumor cell escape from virus-specific T cell surveillance. J Exp Med 167: 1811, 1988
37. Van Snick J: Interleukin-6: An overview. Annu Rev Immu- no1 8:253, 1990
38. Helson L, Green S, Carswell EA, Old LJ: Effect of tumor necrosis factor on cultured human melanoma cells. Nature 258: 731. 1975
39. Jacubovich R, Cabrillat H, Gerlier D, Bailly M, Dore JF: Tu- morigenic phenotypes of human melanoma cell lines in nude mice determined by an active antitumor mechanism. Br J Cancer 5 l : 335, 1985
40. Gamble JR, Harlan JM. Klebanoff SJ, Vadas MA: Stimula- tion of the adherence of neutrophils to umbilical vein endothelium by human recombinant tumor necrosis factor. Proc Natl Acad Sci USA 82:8667. 1985
4 I . Bevilacqua MP, Rober JS, Majeau GR, Fiers W, Cotran R, Gimbrone MA Jr: Recombinant tumor necrosis factor induces pro- coagulant activity in cultured human vascular endothelium: Char- acterization and comparison with the actions of interleukin- I . Proc Natl Acad Sci USA 83:4533, 1986
42. Nawroth PP. Stern DM: Modulation ofendothelium cell he- mostatic properties by tumor necrosis factor. J Exp Med 163:740. I986
43. Shimomura K, Manda T, Mukumoto S, Kobayashi K, Na- kano K. Mori J: Recombinant human tumor necrosis factor-a: Thrombus formation is a cause of anti-tumor activity. Int J Cancer 41 :243, I988
44. Robaye B, Mosselmans R, Fiers W, Dumont JE, Galand P: Tumor necrosis factor induces apoptosis (programmed cell death) in normal endothelial cells in vitro. Am J Pathol 138:447, 199 1
45. Patte C, Philip T, Rodary C, Zucker J-M, Behrendt H, Gen- tet J-C. Lamagnere J-P, Otten J, Dufillot D, Pein F, Caillou B, Lemerle J: High survival rate in advanced-stage B-cell lympho- mas and leukemias without CNS involvement with a short inten- sive polychemotherapy: Results from the French Pediatric On- cology Society ofa randomized trial of2 16 children. J Clin Oncol 9:123, 1991
46. Schwenn MR, Blattner SR, Lynch E, Weinstein HJ: HiC- COM: A 2-month intensive chemotherapy regimen for children with stage I11 and IV Burkitt’s lymphoma and B-cell acute lympho- blastic leukemia. J Clin Oncol 9: 133, 1991
47. Levine A: Acquired immunodeficiency syndrome-related lymphoma. Blood 80:8, 1991





























