Embed Size (px)
Citation preview

BASIC NEUROSCIENCES, GENETICS AND IMMUNOLOGY - ORIGINAL ARTICLE
Regulation of the immediate-early genes arc and zif268 in a mouseoperant model of cocaine seeking reinstatement
Barbara Ziołkowska • Michał Kiełbinski •
Agnieszka Gieryk • Guadalupe Soria •
Rafael Maldonado • Ryszard Przewłocki
Received: 7 September 2010 / Accepted: 9 January 2011
� Springer-Verlag 2011
Abstract Reinstatement of extinguished operant res-
ponding for drug is an appropriate model of relapse to drug
abuse. Due to the difficulty of implementing in mice the
procedure of instrumental intravenous self-administration,
mechanisms of reinstatement have so far been studied
almost exclusively in rats. A mouse model of reinstatement
of cocaine seeking has recently been characterized (Soria
et al. 2008). The aim of the present study was to assess
regional brain activation, as measured by induction of the
immediate early genes (IEG) arc and zif268, during
priming- or cue-elicited reinstatement of cocaine seeking
using this new mouse model and the in situ hybridization
technique. We have demonstrated that cue-elicited rein-
statement of cocaine seeking was associated with induction
of the IEG in the medial prefrontal cortex (prelimbic and
infralimbic) and basolateral amygdala. Priming-induced
reinstatement produced a more widespread up-regulation
of those genes in forebrain regions including medial pre-
frontal, orbitofrontal and motor cortex, dorsal striatum and
basolateral amygdala. These patterns of IEG expression are
in agreement with previous results obtained in rats and thus
indicate that the new mouse model of reinstatement is
functionally equivalent to rat models. That comparability
adds to the usefulness of the mouse model as a tool for
addressing neurobiological mechanisms of addiction.
Keywords Cocaine � Drug seeking � Reinstatement �Immediate early gene � Brain activity mapping
Introduction
Drug addiction is a chronic and relapsing brain disorder.
Cessation of chronic taking of a drug of abuse is usually
accompanied by episodes of craving for the drug, which
often leads to relapse into the drug use even after a long
abstinence (Anton 1999; Franken 2003; O’Brien et al.
1998). Craving may be elicited by several factors, which
include stress, as well as re-exposure to a priming dose of
the drug or to environmental stimuli previously associated
with drug taking (O’Brien et al. 1998; Shaham et al. 2003;
Shalev et al. 2002). From both theoretical and therapeutical
standpoints, understanding neural mechanisms underlying
craving is essential because they actually represent the
main driving force of addiction.
Intravenous operant self-administration is considered
the most relevant, reliable and useful animal model to
study these processes (Shaham et al. 2003; Shalev et al.
2002). In addition to its similarity to human scenarios (i.e.
the fact that the drug use is controlled by the animal rather
than the experimenter), the self-administration model per-
mits the separate study of several aspects and stages of
addiction: acquisition and maintenance of operant
responding for the drug, as well as extinction and rein-
statement of the drug-directed behaviour. Similar instru-
mental self-administration paradigms (e.g. oral) can also be
B. Ziołkowska � M. Kiełbinski � A. Gieryk � R. Przewłocki (&)
Department of Molecular Neuropharmacology,
Institute of Pharmacology, Polish Academy of Sciences,
Smetna 12, 31-343 Krakow, Poland
e-mail: [email protected]
G. Soria � R. Maldonado
Laboratorio de Neurofarmacologıa, Universitat Pompeu Fabra,
Parc de Recerca Biomedica de Barcelona, C/Dr. Aiguader 88,
08003 Barcelona, Spain
Present Address:G. Soria
Institute for Biomedical Research (IIBB-CSIC), Experimental
MRI Unit (IDIBAPS), Rossello 161, 08036 Barcelona, Spain
123
J Neural Transm
DOI 10.1007/s00702-011-0583-z

used to study effects of natural reinforcers, such as palatable
and/or energy-dense food, which may as well produce
addiction-like behavioural disturbances and recruit partly the
same neural mechanisms as drugs of abuse (Nair et al. 2009).
In order to fully elucidate the mechanisms of addiction,
it is necessary to combine behavioural studies with com-
plementary molecular and genomic approaches. Recent
advances in generation of genetically modified mouse lines
have yielded powerful tools for studying multiple genes
and pathways that could be involved in drug addiction
(Changeux 2010; Kieffer and Gaveriaux-Ruff 2002; Sora
et al. 2010; Stephens et al. 2002). However, due to the
technical difficulty of implementing the self-administration
procedure in mice (Thomsen and Caine 2007), most studies
in operant models of drug self-administration have so far
been conducted in rats (Shaham et al. 2003; Shalev et al.
2002; Schmidt et al. 2005). It is therefore necessary to
develop reliable mouse models in order to take advantage
of genetic modification technologies in addiction research.
Such an operant mouse model of acquisition, extinction
and reinstatement of cocaine seeking behaviour, in which all
factors known to precipitate relapse in humans (i.e. drug
priming dose, discrete drug-associated cues and stress) rein-
stated extinguished responding for the drug, has recently been
described for the first time by Soria et al. (2008). Whereas the
model has been characterized in behavioural terms, where its
similarity to the existing rat models has been demonstrated, it
still remains to be established if neural mechanisms underly-
ing reinstatement of operant responding for cocaine after
extinction in mice are the same as in rats. This can be primarily
assessed by comparing patterns of brain activation during
reinstatement of operant responding in the different models.
Although such an approach does not give an insight into
intracellular processes, it does enable to locate brain regions
and neuronal circuits, activation of which is associated with
behavioural responses leading to reinstatement of drug seek-
ing. Thereby, brain regions and circuits can be identified, in
which some specific changes had occurred because of the
previous self-administration training.
Immediate early genes (IEG), such as c-fos, arc and
zif268 (also known as egr-1, krox-24, and NGFI-A), have
been commonly used as markers of brain activation in
laboratory animals (Herdegen and Leah 1998; Kaczmarek
and Chaudhuri 1997; Ziołkowska and Przewłocki 2002).
Their suitability results from the fact that IEG are rapidly
upregulated in neurons in a highly localized, cell-specific,
activity- and time-dependent manner in response to various
cell-excitatory stimuli (Herdegen and Leah 1998; Morgan
et al. 1987; Sagar et al. 1988). IEG expression in several
experimental models of drug addiction has been studied
extensively and has been characterized in rat models of
reinstatement of operant responding for psychostimulants
(Ciccocioppo et al. 2001; Harlan and Garcia 1998; Hearing
et al. 2008a; Kufahl et al. 2009; Neisewander et al. 2000;
Thomas et al. 2003). Several neural systems can be con-
sidered critical to the processes of reinstatement of cocaine
seeking behaviour based on IEGs being consistently
upregulated in these brain regions in the rat paradigms.
These include prefrontal cortical regions (anterior cingulate
and prelimbic cortex) and basolateral amygdala (Ciccocioppo
et al. 2001; Hearing et al. 2008a; Kufahl et al. 2009; Neise-
wander et al. 2000; Thomas et al. 2003). Some studies also
demonstrated IEG induction in additional regions, e.g.
nucleus accumbens, dorsal striatum, hippocampus and ventral
tegmental area (Hearing et al. 2008b; Kufahl et al. 2009;
Neisewander et al. 2000; Thomas et al. 2003).
In order to get insight into specific patterns of brain
activation during reinstatement of cocaine seeking in mice,
we studied regional expression of two IEG, zif268 and arc,
using in situ hybridization, in the behavioural paradigm
recently designed by Soria et al. (2008). The effect of two
relapse-triggering factors was assessed, a drug-associated
cue and drug priming, which may produce reinstatement of
responding by activation of different neuronal pathways
(Shaham et al. 2003; Shalev et al. 2002).
Materials and methods
Animals
The experiment was carried out on male CD1 mice (Charles
River, France) weighing 20–25 g upon arrival at the labora-
tory. The animals were housed individually in plastic cages,
kept on a 12-h light/dark cycle, at constant temperature
(21 ± 1�C) and air humidity (55 ± 10%). Food and water
were provided ad libitum, except during the experimental
sessions. Mice were handled daily in order to minimize stress
during the experiments. Animal care and experimental pro-
cedures were in accordance with institutional and interna-
tional standards (the European Communities Council
Directive 86/609/EEC, 24 November 1986) and were
approved by the local Ethics Committee (CEEA-PRBB).
Control injections of cocaine and saline
Control experiments were carried out in intact, behavio-
urally non-trained animals in order to assess the pharma-
cological effect of cocaine and the impact of injection
stress on arc and zif268 gene expression. Mice were
injected i. p. with saline or cocaine (10 mg/kg), while naive
animals served as a control. First, the time-course of IEG
induction due to cocaine and stress was estimated quali-
tatively, the animals being sacrificed at 30 min or 2 h after
the injection (n = 3 to 4). Quantitative measurements of
IEG expression were then performed in an independent
B. Ziołkowska et al.
123

experiment, in mice decapitated 2 h post-injection, i.e. the
time equivalent to the duration of a reinstatement session
(saline: n = 7; cocaine: n = 7; naive: n = 5).
Behavioural training and reinstatement procedures
The present in situ hybridization experiments were performed
on the brains of the mice which had been behaviourally
characterized in the previous paper by Soria et al. (2008).
Thus, all details concerning the animal treatment and the
behavioural procedure are to be found in the aforementioned
paper. Briefly, mice were implanted with intravenous silastic
catheters and, after a recovery period, trained to self-admin-
ister cocaine hydrochloride (Ministerio de Sanidad y Con-
sumo, Spain) at the dose of 1 mg/kg per infusion. The training
consisted of daily 2-h sessions and was conducted in MED
Associates (Georgia, VT, USA) operant chambers equipped
with two retractable levers. Whenever the animal pressed one
of the levers (designated as ‘active’), it obtained one infusion
of cocaine and was presented a light cue. The training was
carried out until the mice met the criteria of self-administra-
tion acquisition (cf Soria et al. 2008). They were then sub-
jected to extinction procedure which consisted of two daily
2-h sessions in the Skinner box, during which lever presses
were not reinforced. The extinction training took place 6 days
per week until the mice reached the extinction criteria (cf Soria
et al. 2008).
One or two days after the extinction training was com-
pleted, the animals were tested for reinstatement of lever
pressing behaviour elicited either by an injection of a
priming cocaine dose (10 mg/kg, i.p.) given immediately
before placement of the animal in the operant chamber or
by presentation of the conditioned light cue previously
associated with cocaine infusions. During reinstatement
sessions, cocaine was not available, and responding was
reinforced by presentation of the drug-associated cue light
only in the cue (but not priming) reinstatement group. A
control group for the priming cocaine dose effect was
injected i.p. with saline and tested for responding in the
Skinner box. All the reinstatement sessions lasted 2 h. An
animal was considered a responder in the reinstatement test
if its behaviour met the following criteria: (1) the number
of responses on the ‘active’ lever was at least double of that
during the last extinction session and (2) the number of
responses on the ‘active’ lever equalled ten or more.
Animal groups analyzed by in situ hybridization
Four animal groups were used for the biochemical analysis
described herein: (a) sham-operated animals, which were
placed in Skinner boxes for 20 daily 2-h sessions without
receiving any infusions and without receiving any other
type of reinforcers for pressing the lever (sham; n = 5);
(b) mice subjected to the self-administration and then to the
extinction training which met the extinction criteria
(extinction; n = 5); (c) mice after self-administration and
extinction training which then underwent testing for rein-
statement triggered by the drug priming dose (priming-
induced reinstatement; n = 10) or (d) by the conditioned
stimulus (cue-induced reinstatement; n = 5). All the mice
were sacrificed by decapitation immediately after com-
pletion of the last 2-h session in the operant chamber.
In situ hybridization
After sacrifice, the brains were removed and frozen on dry
ice. They were then cut into 12 lm thick coronal sections
on a cryostat microtome (Leica Microsystems, Nussloch,
Germany), the sections were thaw-mounted on gelatin-
chrom-alum-coated slides and processed for in situ
hybridization according to the method of Young et al.
(1986). Briefly, the sections were fixed with 4% parafor-
maldehyde, washed with PBS and acetylated by incubation
with 0.25% acetic anhydrite (in 0.1 M triethanolamine and
0.9% sodium chloride). The sections were then dehydrated
using increasing concentrations of ethanol (70–100%),
treated with chloroform for 5 min, and rehydrated with
decreasing concentrations of ethanol.
The sections were hybridized for approximately 15 h at
37�C with arc and zif268 oligonucleotide probes. The arc
probe 50 TGC CCA CCG ACC TGT GCA ACC CTT TCA
GCT CTC GCT CCA 30 (accession No. NM_018790;
probe complementary to nucleotides 368–406) had been
designed by Ammon et al. (2003). The zif268 probe 50
CCG TTG CTC AGC AGC ATC ATC TCC TCC AGT
TTG GGG TAG TTG TCC 30 (accession No. NM_007913;
probe complementary to nucleotides 369–413) had been
designed by Wisden et al. (1990). The probes were labeled
with 35S-dATP by the 30-tailing reaction using terminal
transferase (MBI Fermentas, Vilnius, Lithuania).
After hybridization, the slices were washed three times
for 20 min with 1xSSC/50% formamide at 40�C, and twice
for 50 min with 19 SSC at room temperature. Then, the
slices were dried and exposed to Fujifilm phosphorimager
imaging plates for 5 days. The hybridization signal was
digitized using the Fujifilm BAS-5000 phosphorimager and
the ImageReader software.
Image analysis
In situ hybridization signal was analyzed using the MCID
Elite system (Imaging Research, St. Catharines, ON,
Canada). Signal density was measured bilaterally in
selected forebrain regions in the Fujifilm BAS-5000 ima-
ges. The regions were chosen based on their reported
activation in rat models and included: the anterior
Regulation of the immediate-early genes arc and zif268
123

cingulate, primary motor cortex, ventrolateral orbital cor-
tex, prelimbic and infralimbic cortical areas, dorsal stria-
tum, nucleus accumbens, hippocampus and the basolateral
amygdala (Fig. 1). In addition, the pharmacological effect
of cocaine injection was assessed in the medial dorsal
striatum (Fig. 1), which had previously been demonstrated
to be the main site of cocaine-elicited IEG induction
(Berke et al. 1998; Fosnaugh et al. 1995; Graybiel et al.
1990; Moratalla et al. 1992).
In those regions of interest, the arc and zif268 hybrid-
ization signal was distributed heterogeneously, i.e. patches
or layers of very intense signal were interspersed with areas
of very low expression, both in basal and stimulated condi-
tions. In order to exclude areas of very low (near background)
expression from the analysis, a thresholding method was
used, in which only those pixels were quantified whose
signal density exceeded a preselected threshold value. For
the spots of the signal exceeding the threshold, area and mean
signal density were measured. Tissue background signal,
measured over the corpus callosum, was subtracted from all
density measurements. The final result was calculated
according to the following formula:
Expression ¼ ðD� bgÞ � AS
AT;
where D represents mean optical density of the above-threshold
pixels, bg represents tissue background (corpus callosum), AS
represents area occupied by pixels with above-threshold den-
sity, and AT represents total sampled area. The data were then
expressed as percentage of the mean in the extinction group.
Data analysis
Data are presented as group means, with error bars showing
the standard error of the mean. The effects of treatment on
IEG mRNA levels were analyzed with one-way analysis of
variance (ANOVA) followed by the post-hoc Newman–
Keuls test to detect differences between groups.
Results
Regulation of IEG expression by cocaine and injection
stress (Figs. 2, 3)
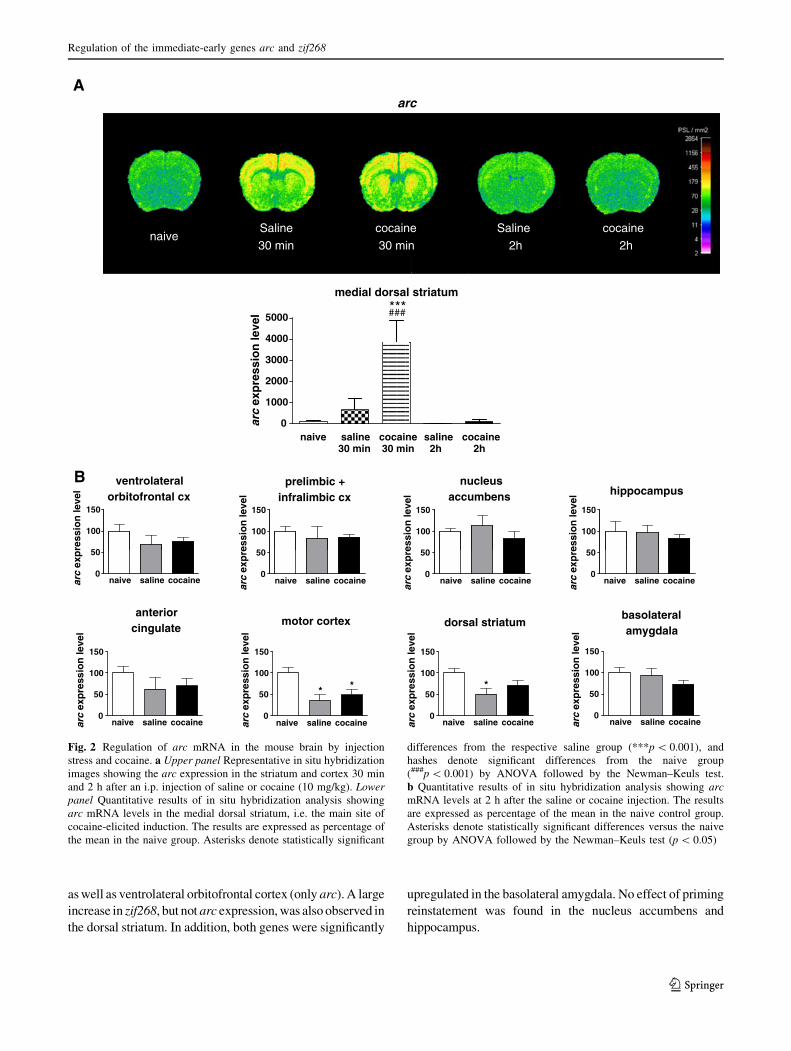
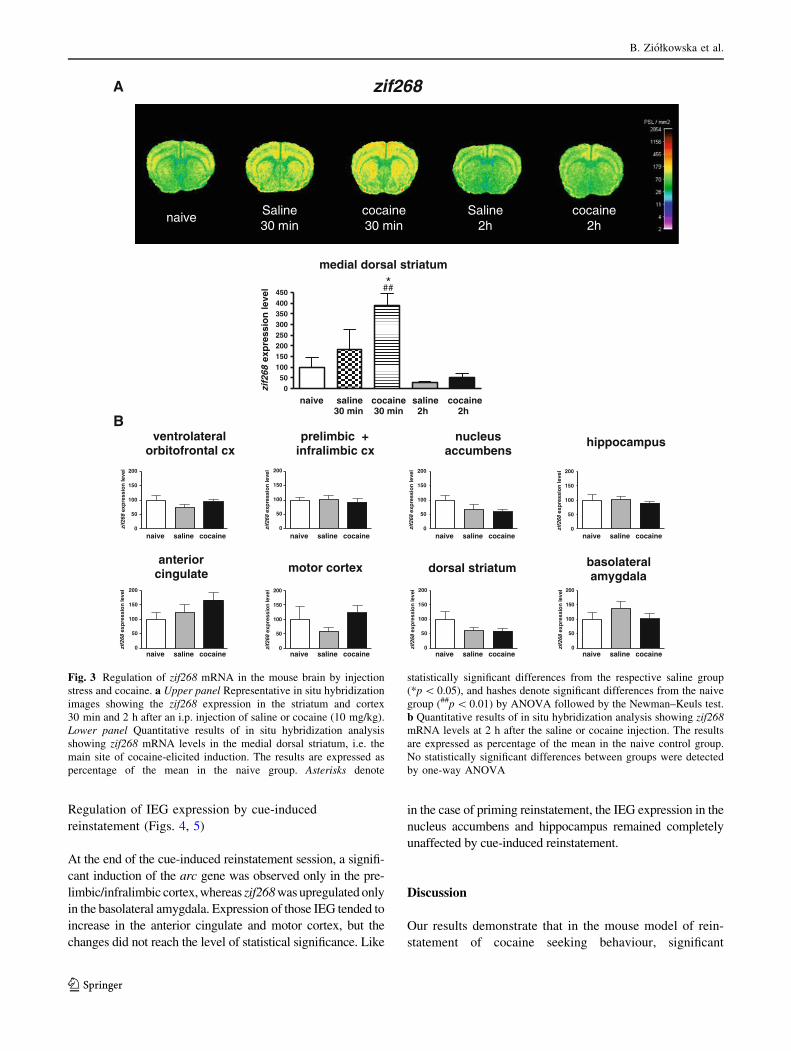
Saline injection produced a widespread induction of the arc
and zif268 genes at 30 min post-injection in the cortex,
striatum and hippocampus, which may be attributed to the
stress of injection and animal handling (Figs. 2a, 3a).
Administration of cocaine (10 mg/kg, i.p.), in addition to
the stress-related IEG activation, up-regulated arc and
zif268 genes in the medial dorsal striatum at 30 min, which
probably reflects the direct pharmacological action of the
drug (Figs. 2a, 3a). Such a cocaine effect has long been
described in the literature (Berke et al. 1998; Fosnaugh
et al. 1995; Graybiel et al. 1990; Moratalla et al. 1992).
Both stress- and cocaine-related IEG inductions dissipated
by 2 h post-injection (Figs. 2, 3). A quantitative analysis of
arc and zif268 mRNA hybridization signal in eight brain
regions (selected for analysis in self-administration rein-
statement paradigms) demonstrated that the IEG mRNA
levels were not significantly elevated in any region at 2 h.
Actually, they were decreased in some areas, especially in
the striatum and motor cortex, as compared to naive ani-
mals, which might be a rebound effect following the earlier
IEG induction in the same brain regions (Figs. 2b, 3b).
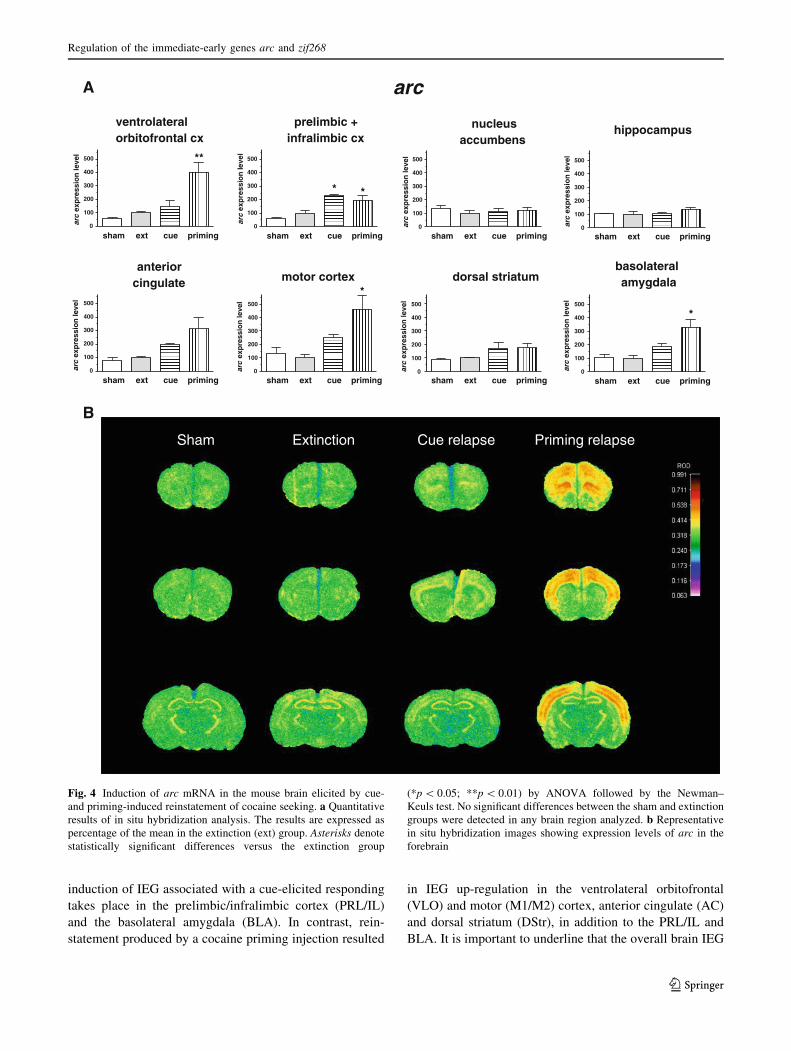
Regulation of IEG expression by priming-induced
reinstatement (Figs. 4, 5)
Of the two reinstatement paradigms studied, cocaine priming
produced greater changes in arc and zif268 gene expression.
Induction of both genes during priming reinstatement was
most pronounced in cortical regions, especially the anterior
cingulate and motor cortex, but significant increases were also
detected in the prelimbic/infralimbic (for both arc and zif268),
Fig. 1 Outlines of the regions of interest in which optical density
measurements were performed. AC anterior cingulated, BLA basolat-
eral amygdale, DStr dorsal striatum, HIP hippocampus, m medial
dorsal striatum; M1/2 primary motor cortex, NAc nucleus accumbens,
PRL ? IL prelimbic and infralimbic cortex, VLO ventrolateral orbital
cortex. The section schemes were taken from the atlas by Paxinos and
Franklin (2001)
B. Ziołkowska et al.
123
as well as ventrolateral orbitofrontal cortex (only arc). A large
increase in zif268, but not arc expression, was also observed in
the dorsal striatum. In addition, both genes were significantly
upregulated in the basolateral amygdala. No effect of priming
reinstatement was found in the nucleus accumbens and
hippocampus.
A
B
arc
naiveSaline
30 min
Saline
2h
cocaine
30 min
cocaine
2h
medial dorsal striatum
3000
4000
5000***###
0
1000
2000
naive saline30 min
saline 2h
cocaine 30 min
cocaine 2h
arc
exp
ress
ion
leve
l
100
150
100
150
ventrolateralorbitofrontal cx
prelimbic +infralimbic cx
nucleusaccumbens hippocampus
naive saline cocaine
naive saline cocaine naive saline cocaine naive saline cocaine naive saline cocaine
naive saline cocaine naive saline cocaine naive saline cocaine0
50
arc
exp
ress
ion
leve
l
100
150
0
50
arc
exp
ress
ion
leve
l
100
150
0
50
arc
exp
ress
ion
leve
l
100
150
0
50
arc
exp
ress
ion
leve
l
100
150
0
50
arc
exp
ress
ion
leve
l
arc
exp
ress
ion
leve
l
0
50
100
150
arc
exp
ress
ion
leve
l
0
50
100
150
arc
exp
ress
ion
leve
l0
50
anteriorcingulate
motor cortex dorsal striatumbasolateralamygdala
* * *
Fig. 2 Regulation of arc mRNA in the mouse brain by injection
stress and cocaine. a Upper panel Representative in situ hybridization
images showing the arc expression in the striatum and cortex 30 min
and 2 h after an i.p. injection of saline or cocaine (10 mg/kg). Lowerpanel Quantitative results of in situ hybridization analysis showing
arc mRNA levels in the medial dorsal striatum, i.e. the main site of
cocaine-elicited induction. The results are expressed as percentage of
the mean in the naive group. Asterisks denote statistically significant
differences from the respective saline group (***p \ 0.001), and
hashes denote significant differences from the naive group
(###p \ 0.001) by ANOVA followed by the Newman–Keuls test.
b Quantitative results of in situ hybridization analysis showing arcmRNA levels at 2 h after the saline or cocaine injection. The results
are expressed as percentage of the mean in the naive control group.
Asterisks denote statistically significant differences versus the naive
group by ANOVA followed by the Newman–Keuls test (p \ 0.05)
Regulation of the immediate-early genes arc and zif268
123
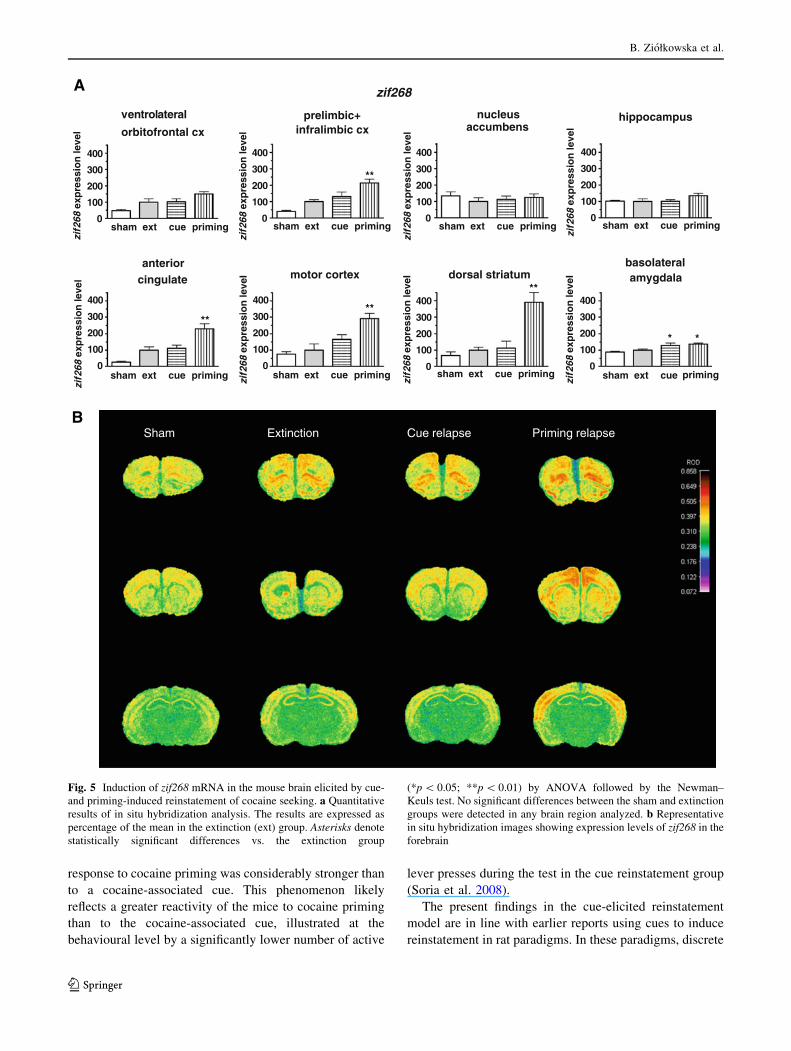
Regulation of IEG expression by cue-induced
reinstatement (Figs. 4, 5)
At the end of the cue-induced reinstatement session, a signifi-
cant induction of the arc gene was observed only in the pre-
limbic/infralimbic cortex, whereas zif268 was upregulated only
in the basolateral amygdala. Expression of those IEG tended to
increase in the anterior cingulate and motor cortex, but the
changes did not reach the level of statistical significance. Like
in the case of priming reinstatement, the IEG expression in the
nucleus accumbens and hippocampus remained completely
unaffected by cue-induced reinstatement.
Discussion
Our results demonstrate that in the mouse model of rein-
statement of cocaine seeking behaviour, significant
naive saline cocaine0
50
100
150
200
zif2
68ex
pre
ssio
n le
vel
naive saline cocaine0
50
100
150
200
zif2
68ex
pre
ssio
n le
vel
naive saline cocaine0
50
100
150
200
zif2
68ex
pre
ssio
n le
vel
naive saline cocaine0
50
100
150
200
zif2
68ex
pre
ssio
n le
vel
naive saline cocaine0
50
100
150
200
zif2
68ex
pre
ssio
n le
vel
naive saline cocaine0
50
100
150
200
zif2
68ex
pre
ssio
n le
vel
naive saline cocaine0
50
100
150
200
zif2
68ex
pre
ssio
n le
vel
naive saline cocaine0
50
100
150
200
zif2
68ex
pre
ssio
n le
vel
ventrolateralorbitofrontal cx
prelimbic +infralimbic cx
nucleusaccumbens
hippocampus
anteriorcingulate motor cortex dorsal striatum basolateral
amygdala
B
A
naive Saline30 min
Saline2h
cocaine30 min
cocaine2h
zif268
medial dorsal striatum
050
100150200250300350400450
*##
naive saline30 min
saline 2h
cocaine 30 min
cocaine 2h
zif2
68 e
xpre
ssio
n le
vel
Fig. 3 Regulation of zif268 mRNA in the mouse brain by injection
stress and cocaine. a Upper panel Representative in situ hybridization
images showing the zif268 expression in the striatum and cortex
30 min and 2 h after an i.p. injection of saline or cocaine (10 mg/kg).
Lower panel Quantitative results of in situ hybridization analysis
showing zif268 mRNA levels in the medial dorsal striatum, i.e. the
main site of cocaine-elicited induction. The results are expressed as
percentage of the mean in the naive group. Asterisks denote
statistically significant differences from the respective saline group
(*p \ 0.05), and hashes denote significant differences from the naive
group (##p \ 0.01) by ANOVA followed by the Newman–Keuls test.
b Quantitative results of in situ hybridization analysis showing zif268mRNA levels at 2 h after the saline or cocaine injection. The results
are expressed as percentage of the mean in the naive control group.
No statistically significant differences between groups were detected
by one-way ANOVA
B. Ziołkowska et al.
123
induction of IEG associated with a cue-elicited responding
takes place in the prelimbic/infralimbic cortex (PRL/IL)
and the basolateral amygdala (BLA). In contrast, rein-
statement produced by a cocaine priming injection resulted
in IEG up-regulation in the ventrolateral orbitofrontal
(VLO) and motor (M1/M2) cortex, anterior cingulate (AC)
and dorsal striatum (DStr), in addition to the PRL/IL and
BLA. It is important to underline that the overall brain IEG
arc
0
100
200
300
400
500
sham ext cue priming
**
arc
exp
ress
ion
leve
l
0
100
200
300
400
500
sham ext cue priming
* *
arc
exp
ress
ion
leve
l
0
100
200
300
400
500
sham ext cue priming
arc
exp
ress
ion
leve
l
0
100
200
300
400
500
sham ext cue priming
arc
exp
ress
ion
leve
l
0
100
200
300
400
500
sham ext cue priming
arc
exp
ress
ion
leve
l
0
100
200
300
400
500
sham ext cue priming
*
arc
exp
ress
ion
leve
l
0
100
200
300
400
500
sham ext cue priming
arc
exp
ress
ion
leve
l
0
100
200
300
400
500
sham ext cue priming
*
arc
exp
ress
ion
leve
l
ventrolateralorbitofrontal cx
prelimbic +infralimbic cx
motor cortexanterior
cingulate dorsal striatum
nucleusaccumbens
hippocampus
basolateral amygdala
A
B
Sham Extinction Cue relapse Priming relapse
Fig. 4 Induction of arc mRNA in the mouse brain elicited by cue-
and priming-induced reinstatement of cocaine seeking. a Quantitative
results of in situ hybridization analysis. The results are expressed as
percentage of the mean in the extinction (ext) group. Asterisks denote
statistically significant differences versus the extinction group
(*p \ 0.05; **p \ 0.01) by ANOVA followed by the Newman–
Keuls test. No significant differences between the sham and extinction
groups were detected in any brain region analyzed. b Representative
in situ hybridization images showing expression levels of arc in the
forebrain
Regulation of the immediate-early genes arc and zif268
123
response to cocaine priming was considerably stronger than
to a cocaine-associated cue. This phenomenon likely
reflects a greater reactivity of the mice to cocaine priming
than to the cocaine-associated cue, illustrated at the
behavioural level by a significantly lower number of active
lever presses during the test in the cue reinstatement group
(Soria et al. 2008).
The present findings in the cue-elicited reinstatement
model are in line with earlier reports using cues to induce
reinstatement in rat paradigms. In these paradigms, discrete
zif268
0
100
200
300
400
sham ext cue priming
sham ext cue priming sham ext cue priming sham ext cue priming sham ext cue priming
sham ext cue priming sham ext cue priming sham ext cue priming
zif2
68 e
xpre
ssio
n le
vel
zif2
68 e
xpre
ssio
n le
vel
zif2
68 e
xpre
ssio
n le
vel
zif2
68 e
xpre
ssio
n le
vel
zif2
68 e
xpre
ssio
n le
vel
zif2
68 e
xpre
ssio
n le
vel
zif2
68 e
xpre
ssio
n le
vel
zif2
68 e
xpre
ssio
n le
vel
0
100
200
300
400
**
0
100
200
300
400
0
100
200
300
400
200
300
400
**200
300
400**
200
300
400**
200
300
400
* *
motor cortexanterior
cingulate dorsal striatumbasolateralamygdala
0
100
0
100
0
100
0
100
Sham Extinction Cue relapse Priming relapse
ventrolateral prelimbic+ nucleus
orbitofrontal cx
infralimbic cx accumbenshippocampus
A
B
Fig. 5 Induction of zif268 mRNA in the mouse brain elicited by cue-
and priming-induced reinstatement of cocaine seeking. a Quantitative
results of in situ hybridization analysis. The results are expressed as
percentage of the mean in the extinction (ext) group. Asterisks denote
statistically significant differences vs. the extinction group
(*p \ 0.05; **p \ 0.01) by ANOVA followed by the Newman–
Keuls test. No significant differences between the sham and extinction
groups were detected in any brain region analyzed. b Representative
in situ hybridization images showing expression levels of zif268 in the
forebrain
B. Ziołkowska et al.
123

cues, previously associated with cocaine infusions or pre-
dictive of the drug availability were shown to induce c-fos
mRNA and protein in the rat prelimbic and infralimbic
cortex, as well as in the BLA (Ciccocioppo et al. 2001;
Kufahl et al. 2009). zif268 mRNA was induced in the BLA,
and arc mRNA in the prelimbic cortex of rats undergoing
cue-elicited reinstatement of cocaine seeking (Thomas
et al. 2003; Zavala et al. 2008), in agreement with the
results obtained in the present mouse model. The rat
studies also reported arc and c-fos induction in the AC
(Cg1 or Cg2) and VLO, where only slight increase ten-
dencies in arc mRNA levels were observed in our exper-
iment. Kufahl et al. (2009) described c-fos activation in
several additional brain regions, which was not consistent
with previous observations in the rat or our mouse model
(Ciccocioppo et al. 2001; Thomas et al. 2003; Zavala et al.
2008), perhaps due to minor procedural differences and
variability in signal detection sensitivity.
Thus, while spatiotemporal patterns of induction may
slightly differ for specific IEG (see below), both rat and
mouse IEG studies provide concurring evidence for
activation of medial prefrontal cortical subregions and
BLA during cue-elicited reinstatement of cocaine seek-
ing. The predictive value of such mapping is indicated by
the fact that activation of the corresponding human brain
regions was demonstrated by in vivo imaging techniques
in drug addicts undergoing craving after presentation of
cocaine-associated stimuli (Bonson et al. 2002; Childress
et al. 1999; Grant et al. 1996; Kilts et al. 2001). The
causative role of dorsomedial prefrontal cortex (PRL and
AC) and BLA in cue-induced reinstatement of cocaine
seeking was confirmed in experimental rat models, where
their lesion or temporary functional inactivation pre-
vented reinstatement of instrumental responding for
cocaine (Kantak et al. 2002; McLaughlin and See 2003;
Meil and See 1997).
The influence of priming-induced reinstatement of
cocaine seeking on IEG expression has so far been assessed
in only one study in rats to our knowledge (Neisewander
et al. 2000). The authors reported a specific effect of
cocaine priming on Fos protein expression only in the AC,
while, in addition to AC, we found arc and/or zif268
mRNA induction in many other brain regions (VLO, PRL/
IL, M1/M2, DStr and BLA). Our control experiments
excluded a contribution of IEG inductions related to
injection stress and pharmacological cocaine actions to the
arc and zif268 activation observed at the end of the rein-
statement session, i.e. 2 h after the priming injection, thus
indicating that the observed gene up-regulation was spe-
cifically associated with animals’ behaviour during the
reinstatement test. Both Neisewander et al. (2000) and our
study are consistent in showing the lack of IEG expression
changes in the nucleus accumbens and hippocampus.
Whereas the prefrontal cortical subregions VLO and
PRL/IL were not subjected to examination by Neisewander
et al. (2000), there is substantial evidence from functional
inactivation and pharmacological studies indicating that
activation of the rat PRL/IL (along with the adjacent AC) is
absolutely necessary for a cocaine prime to trigger rein-
statement responding (Capriles et al. 2003; McFarland and
Kalivas 2001; Schmidt et al. 2005). Involvement of the
VLO has also been suggested by effects of its lesioning
(Fuchs et al. 2004), although not confirmed by other
methods (cf. Schmidt et al. 2005). Thus, our finding in the
mouse model of IEG activation in all prefrontal cortical areas,
i.e. VLO, PRL/IL and AC provides new information about the
neurobiological substrate involved in priming-induced rein-
statement of cocaine seeking and is in agreement with previ-
ous pharmacological and lesion studies in rats.
The discrepancy between our data and those by Neise-
wander et al. (2000) consists in the finding of IEG induc-
tion in the DStr, M1/M2 and BLA of mice but not rats.
Whereas species-specific mechanisms may account for
some of these discrepancies, they may just as well result
from differences in the design of both studies, taking into
consideration the fact that different genes were assessed
(Fos vs. arc and zif) and that inductions were detected
either at the mRNA level (in our study) or on protein level
(by Neisewander et al. 2000), which have largely non-
overlapping time-courses. In addition, detection of a rein-
statement-related induction in the DStr in the Neisewander
et al. study might have been confounded by pharmaco-
logical effect of cocaine priming (which was excluded in
our experimental setting) (Berretta et al. 1992; Graybiel
et al. 1990). On the other hand, although Neisewander et al.
(2000) did not detect Fos induction in the BLA during
priming-induced reintatement in rats, both lesion and
pharmacological rat studies (Alleweireldt et al. 2003; Yun
and Fields 2003) suggest an involvement of BLA in
priming-triggered cocaine seeking.
Activation of the IEG transcription in the motor cortex
during the reinstatement session could reflect brain activity
associated with the motor act of instrumental responding.
However, IEG were not induced in the motor cortex in
several rat studies in which animals were allowed to lever-
press during the session of reinstatement induced by
priming, cocaine-associated cues or context (Ciccocioppo
et al. 2001; Kufahl et al. 2009; Neisewander et al. 2000;
Zavala et al. 2008). On the other hand, Hearing et al.
(2008b) reported up-regulation of c-fos, arc and zif268
genes in the motor cortex during context-elicited cocaine
seeking, which did not depend on whether the rats actually
performed the instrumental reaction or not since levers
were retracted during the test session. Thus, it seems that
both in mice and in rats, motor cortex IEG activation does
occur only in some reinstatement paradigms and may be
Regulation of the immediate-early genes arc and zif268
123

related to general motor activation during the test session
rather than being specifically associated with instrumental
responding.
Interestingly, our experiment demonstrated differences
between induction patterns of arc and zif268 genes. While
some of these differences seemed to be of quantitative nature
(one gene being significantly induced and the other one dis-
playing a non-significant tendency to up-regulation in the
same brain region), others seemed rather qualitative. In par-
ticular, only zif268 but not arc was induced in the DStr,
whereas only arc but not zif268 was induced in the VLO, both
in the priming reinstatement group. These apparently some-
what distinct spatiotemporal patterns of arc and zif268 mRNA
induction may result from (i) different intracellular mecha-
nisms of transcriptional activation for each gene (cf. e.g.
Lemberger et al. 2008; Rodriguez Parkitna et al. 2010; Worley
et al. 1993) and (ii) differences between brain regions in decay
rates of specific mRNA species (Kelly and Deadwyler 2003).
Therefore, due to the partial complementarity of IEG
expression patterns, the use of more than one IEG marker for
brain activity mapping may yield a more complete picture.
In conclusion, we have demonstrated that in the mouse
model of relapse, cue-elicited reinstatement of cocaine
seeking is associated with induction of the IEG arc and
zif268 in the medial prefrontal cortex (PRL/IL) and BLA.
Priming-induced reinstatement produced a more wide-
spread up-regulation of those genes in forebrain regions.
These patterns of IEG mRNA expression are mostly in
agreement with those described in previous studies in rats,
suggesting that no large-scale functional differences exist
between the mouse and rat models. Thus, future conclu-
sions derived from the present mouse model can be com-
bined with the existing abundant data obtained in rat
models to address further issues concerning brain mecha-
nisms of drug craving and relapse.
Acknowledgments This work was supported by the EU grant
LSHM-CT-2007-037669 (PHECOMP), the Polish Ministry of Sci-
ence and Higher Education subsidiary grant No. 478/6. PR UE/2007/
7, statutory funds from the Institute of Pharmacology, Polish Acad-
emy of Sciences, the Spanish Ministerio de Ciencia e Innovacion
(#SAF 2007-64062), the Catalan Government (S6R 2009-00131) and
the ICREA Foundation (ICREA-Academia 2008).
References
Alleweireldt AT, Kirschner KF, Blake CB, Neisewander JL (2003)
D1-receptor drugs and cocaine-seeking behavior: investigation
of receptor mediation and behavioral disruption in rats. Psycho-
pharmacology (Berl) 168:109–117
Ammon S, Mayer P, Riechert U, Tischmeyer H, Hollt V (2003)
Microarray analysis of genes expressed in the frontal cortex of
rats chronically treated with morphine and after naloxone
precipitated withdrawal. Mol Brain Res 112:113–125
Anton RF (1999) What is craving? Models and implications for
treatment. Alcohol Res Health 23:165–173
Berke JD, Paletzki RF, Aronson GJ, Hyman SE, Gerfen CR (1998) A
complex program of striatal gene expression induced by
dopaminergic stimulation. J Neurosci 8:5301–5310
Berretta S, Robertson HA, Graybiel AM (1992) Dopamine and
glutamate agonists stimulate neuron-specific expression of Fos-
like protein in the striatum. J Neurophysiol 68:767–777
Bonson KR, Grant SJ, Contoreggi CS, Links JM, Metcalfe J, Weyl
HL et al (2002) Neural systems and cue-induced cocaine
craving. Neuropsychopharmacology 26:376–386
Capriles N, Rodaros D, Sorge RE, Stewart J (2003) A role for the
prefrontal cortex in stress- and cocaine-induced reinstatement of
cocaine seeking in rats. Psychopharmacology (Berl) 168:66–74
Changeux JP (2010) Nicotine addiction and nicotinic receptors:
lessons from genetically modified mice. Nat Rev Neurosci
11:389–401
Childress AR, Mozley PD, McElgin W, Fitzgerald J, Reivich M,
O’Brien CP (1999) Limbic activation during cue-induced
cocaine craving. Am J Psychiatry 156:11–18
Ciccocioppo R, Sanna PP, Weiss F (2001) Cocaine-predictive
stimulus induces drug-seeking behavior and neural activation
in limbic brain regions after multiple months of abstinence:
reversal by D(1) antagonists. Proc Natl Acad Sci USA
98:1976–1981
Fosnaugh JS, Bhat RV, Yamagata K, Worley PF, Baraban JM (1995)
Activation of arc, a putative ‘‘effector’’ immediate early gene,
by cocaine in rat brain. J Neurochem 64:2377–2380
Franken IH (2003) Drug craving and addiction: integrating psycho-
logical and neuropsychopharmacological approaches. Prog Neu-
ropsychopharmacol Biol Psychiatry 27:563–579
Fuchs RA, Evans KA, Parker MP, See RE (2004) Differential
involvement of orbitofrontal cortex subregions in conditioned
cue-induced and cocaine-primed reinstatement of cocaine seek-
ing in rats. J Neurosci 24:6600–6610
Grant S, London ED, Newlin DB, Villemagne VL, Liu X, Contoreggi
C et al (1996) Activation of memory circuits during cue-elicited
cocaine craving. Proc Natl Acad Sci USA 93:12040–12045
Graybiel AM, Moratalla R, Robertson HA (1990) Amphetamine and
cocaine induce drug-specific activation of the c-fos gene in
striosome-matrix compartments and limbic subdivisions of the
striatum. Proc Natl Acad Sci USA 87:6912–6916
Harlan RE, Garcia MM (1998) Drugs of abuse and immediate-early
genes in the forebrain. Mol Neurobiol 16:221–267
Hearing MC, Miller SW, See RE, McGinty JF (2008a) Relapse to
cocaine seeking increases activity-regulated gene expression
differentially in the prefrontal cortex of abstinent rats. Psycho-
pharmacology (Berl) 198:77–91
Hearing MC, See RE, McGinty JF (2008b) Relapse to cocaine-
seeking increases activity-regulated gene expression differen-
tially in the striatum and cerebral cortex of rats following
short or long periods of abstinence. Brain Struct Funct 213:
215–227
Herdegen T, Leah JD (1998) Inducible and constitutive transcription
factors in the mammalian nervous system: control of gene
expression by Jun, Fos and Krox, and CREB/ATF proteins.
Brain Res Rev 28:370–490
Kaczmarek L, Chaudhuri A (1997) Sensory regulation of immediate-
early gene expression in mammalian visual cortex: implications
for functional mapping and neural plasticity. Brain Res Rev
23:237–256
Kantak KM, Black Y, Valencia E, Green-Jordan K, Eichenbaum HB
(2002) Dissociable effects of lidocaine inactivation of the rostral
and caudal basolateral amygdala on the maintenance and reinstate-
ment of cocaine-seeking behavior in rats. J Neurosci 22:1126–1136
Kelly MP, Deadwyler SA (2003) Experience-dependent regulation of
the immediate-early gene arc differs across brain regions.
J Neurosci 23:6443–6451
B. Ziołkowska et al.
123

Kieffer BL, Gaveriaux-Ruff C (2002) Exploring the opioid system by
gene knockout. Prog Neurobiol 66:285–306
Kilts CD, Schweitzer JB, Quinn CK, Gross RE, Faber TL, Muham-
mad F et al (2001) Neural activity related to drug craving in
cocaine addiction. Arch Gen Psychiatry 58:334–341
Kufahl PR, Zavala AR, Singh A, Thiel KJ, Dickey ED, Joyce JN,
Neisewander JL (2009) c-Fos expression associated with rein-
statement of cocaine-seeking behavior by response-contingent
conditioned cues. Synapse 63:823–835
Lemberger T, Parkitna JR, Chai M, Schutz G, Engblom D (2008)
CREB has a context-dependent role in activity-regulated tran-
scription and maintains neuronal cholesterol homeostasis.
FASEB J 22:2872–2879
McFarland K, Kalivas PW (2001) The circuitry mediating cocaine-
induced reinstatement of drug-seeking behavior. J Neurosci
21:8655–8663
McLaughlin J, See RE (2003) Selective inactivation of the dorsome-
dial prefrontal cortex and the basolateral amygdala attenuates
conditioned-cued reinstatement of extinguished cocaine-seeking
behavior in rats. Psychopharmacology (Berl) 168:57–65
Meil WM, See RE (1997) Lesions of the basolateral amygdala abolish
the ability of drug associated cues to reinstate responding during
withdrawal from self-administered cocaine. Behav Brain Res
87:139–148
Moratalla R, Robertson HA, Graybiel AM (1992) Dynamic regulation
of NGFI-A (zif268, egr1) gene expression in the striatum.
J Neurosci 12:2609–2622
Morgan JI, Cohen DR, Hempstead JL, Curran T (1987) Mapping
patterns of c-fos expression in the central nervous system after
seizure. Science 237:192–197
Nair SG, Adams-Deutsch T, Epstein DH, Shaham Y (2009) The
neuropharmacology of relapse to food seeking: methodology,
main findings, and comparison with relapse to drug seeking.
Prog Neurobiol 89:18–45
Neisewander JL, Baker DA, Fuchs RA, Tran-Nguyen LT, Palmer A,
Marshall JF (2000) Fos protein expression and cocaine-seeking
behavior in rats after exposure to a cocaine self-administration
environment. J Neurosci 20:798–805
O’Brien CP, Childress AR, Ehrman R, Robbins SJ (1998) Condi-
tioning factors in drug abuse: can they explain compulsion?
J Psychopharmacol 12:15–22
Paxinos G, Franklin KBJ (2001) The mouse brain in stereotaxic
coordinates, 2nd edn. Academic Press, CD-ROM version
Rodriguez Parkitna J, Bilbao A, Rieker C, Engblom D, Piechota M,
Nordheim A, Spanagel R, Schutz G (2010) Loss of the serum
response factor in the dopamine system leads to hyperactivity.
FASEB J 24(7):2427–2435
Sagar SM, Sharp FR, Curran T (1988) Expression of c-fos protein in brain:
metabolic mapping at the cellular level. Science 240:1328–1331
Schmidt HD, Anderson SM, Famous KR, Kumaresan V, Pierce RC
(2005) Anatomy and pharmacology of cocaine priming-induced
reinstatement of drug seeking. Eur J Pharmacol 526:65–76
Shaham Y, Shalev U, Lu L, De Wit H, Stewart J (2003) The
reinstatement model of drug relapse: history, methodology and
major findings. Psychopharmacology (Berl) 168:3–20
Shalev U, Grimm JW, Shaham Y (2002) Neurobiology of relapse to
heroin and cocaine seeking: a review. Pharmacol Rev 54:1–42
Sora I, Li B, Igari M, Hall FS, Ikeda K (2010) Transgenic mice in the
study of drug addiction and the effects of psychostimulant drugs.
Ann N Y Acad Sci 1187:218–246
Soria G, Barbano MF, Maldonado R, Valverde O (2008) A reliable
method to study cue-, priming-, and stress-induced reinstatement
of cocaine self-administration in mice. Psychopharmacology
(Berl) 199:593–603
Stephens DN, Mead AN, Ripley TL (2002) Studying the neurobiol-
ogy of stimulant and alcohol abuse and dependence in genet-
ically manipulated mice. Behav Pharmacol 13:327–345
Thomas KL, Arroyo M, Everitt BJ (2003) Induction of the learning
and plasticity-associated gene Zif268 following exposure to a
discrete cocaine-associated stimulus. Eur J Neurosci 17:1964–
1972
Thomsen M, Caine SB (2007) Intravenous drug self-administration in
mice: practical considerations. Behav Genet 37:101–118
Wisden W, Errington ML, Williams S, Dunnett SB, Waters C,
Hitchcock D et al (1990) Differential expression of immediate
early genes in the hippocampus and spinal cord. Neuron
4:603–614
Worley PF, Bhat RV, Baraban JM, Erickson CA, McNaughton BL,
Barnes CA (1993) Thresholds for synaptic activation of
transcription factors in hippocampus: correlation with long-term
enhancement. J Neurosci 13:4776–4786
Young WS 3rd, Bonner TI, Brann MR (1986) Mesencephalic
dopamine neurons regulate the expression of neuropeptide
mRNAs in the rat forebrain. Proc Natl Acad Sci USA 83:
9827–9831
Yun IA, Fields HL (2003) Basolateral amygdala lesions impair both
cue- and cocaine-induced reinstatement in animals trained on a
discriminative stimulus task. Neuroscience 121:747–757
Zavala AR, Osredkar T, Joyce JN, Neisewander JL (2008) Upreg-
ulation of Arc mRNA expression in the prefrontal cortex
following cue-induced reinstatement of extinguished cocaine-
seeking behavior. Synapse 62:421–431
Ziołkowska B, Przewłocki R (2002) Methods used in inducible
transcription factor studies: focus on mRNA. In: Kaczmarek L,
Robertson HA (eds) Handbook of Chemical Neuroanatomy, vol
19. Immediate early genes and inducible transcription factors in
mapping of the central nervous system function and dysfunction.
Elsevier, Amsterdam, pp 1–38
Regulation of the immediate-early genes arc and zif268
123





























