Embed Size (px)
Citation preview

Neuropharmacology 50 (2006) 1030e1040www.elsevier.com/locate/neuropharm
Reinforcing effects of nicotine microinjections into the ventraltegmental area of mice: Dependence on cholinergic nicotinic
and dopaminergic D1 receptors
Vincent David a, Morgane Besson b, Jean-Pierre Changeux b, Sylvie Granon b,*, Pierre Cazala a
a Laboratoire de Neurosciences Cognitives, CNRS UMR 5106, Universite de Bordeaux I, Talence, Franceb Unite ‘‘Recepteurs & Cognition’’, Departement de Neuroscience, CNRS 2182, Institut Pasteur, 25 rue du Dr. Roux, 75015 Paris, France
Received 8 July 2005; received in revised form 3 February 2006; accepted 3 February 2006
Abstract
We used an intracranial self-administration (ICSA) procedure to assess the involvement of the ventral tegmental area (VTA) nicotinic recep-tors in the rewarding effects of nicotine. We then challenged intra-VTA nicotine self-administration via systemic or local injections of dopamine(DA)-D1 and nicotinic receptor antagonists. C57BL/6J mice were stereotaxically implanted unilaterally with a guide cannula above the VTA.After 1 week of recovery, mice were allowed to discriminate between two arms of a Y-maze over seven daily sessions, one arm being reinforcedby intracranial nicotine microinjection. Mice exhibited nicotine self-administration at both doses tested, i.e. 10 ng (21.6 pmol) and 100 ng(216 pmol)/50-nl injection. In contrast, mice receiving a 216-pmol nicotine dose 0.8 mm above VTA performed at chance level. Once theICSA response was acquired, systemic pretreatment with the DA-D1 receptor antagonist SCH 23390 (25 mg/kg i.p.) or co-infusion of the nAChRantagonist DHbE with nicotine disrupted ICSA. Replacement of SCH 23390 by vehicle, or withdrawal of DHbE from nicotine/DHbE mixedsolutions led to recovery of intra-VTA nicotine self-administration. We conclude that nicotinic receptors in the VTA, presumably a4b2 nAChRsare critically to mediate the rewarding effects of nicotine and that DA-D1 receptors are also directly implicated.� 2006 Elsevier Ltd. All rights reserved.
Keywords: Nicotine; Addiction; Reward; Dopamine; Nicotinic receptors
1. Introduction
Nicotine affects several cognitive and psychomotorprocesses in various animal species (Sahakian et al., 1989;Nomikos et al., 2000; Shim et al., 2001; Picciotto andCorrigall, 2002). Under particular conditions, cognitive perfor-mance can be improved (Levin et al., 1996) but motivationaland decisional processes may also be altered, which can even-tually lead to compulsory behaviours and pathological statesof dependence. Indeed, rodents self-administer nicotine andits withdrawal produces both dose- and time-dependent with-drawal effects as classically observed for other abused drugs(Rose and Corrigall, 1997; Skjei and Markou, 2003).
* Corresponding author. Tel.: þ33 1 40 61 3777; fax: þ33 1 45 68 8836.
E-mail address: [email protected] (S. Granon).
0028-3908/$ - see front matter � 2006 Elsevier Ltd. All rights reserved.
doi:10.1016/j.neuropharm.2006.02.003
Intravenous nicotine self-administration using both fixed andprogressive ratio schedules has been demonstrated in naiverats (Donny et al., 1999; Paterson and Markou, 2002; Parkeret al., 2004) and in animals previously trained to respond forfood (Bruijnzeel and Markou, 2003). In mice, intravenousnicotine induces dose-dependent self-administration usinga nose-poke operant response (Rasmussen and Swedberg,1998; Semenova et al., 2003) or drug discrimination usinga lever press paradigm (Stolerman et al., 1999). Moreover,mutant mice lacking the b2 nicotinic receptor subunit are im-paired in drug discrimination and nicotine self-administration,revealing a crucial role of the b2 subunit in nicotine rewardmechanisms (Picciotto et al., 1998; Shoaib et al., 2002;Maskos et al., 2005).
Several lines of evidence suggest the involvement of multi-ple neurotransmitter systems in nicotine’s effects, and there-fore, potentially, in nicotine addiction. Indeed, acute nicotine

1031V. David et al. / Neuropharmacology 50 (2006) 1030e1040
administration induces the release of glutamate, GABA andDA in the nucleus accumbens (Picciotto and Corrigall, 2002;Imperato et al., 1986) and in the prefrontal cortex (seePicciotto and Corrigall, 2002, for review). The mesocortico-limbic DA pathway originating in the VTA and projecting tothe nucleus accumbens (NAc), amygdala and prefrontal cortexis clearly involved in nicotine reward (Picciotto and Corrigall,2002; Berridge and Robinson, 1998; Mansvelder et al., 2003).Moreover, the integrity of this pathway has been shown to becrucial for both the rewarding (Corrigall and Coen, 1991) andthe aversive effects of nicotine (Jorenby et al., 1990;Laviolette and van der Kooy, 2003b). The VTA should thusplay a pivotal role in these effects, since injection of nicotineinto the VTA mimics its systemic effects on DA release and onc-fos expression in the NAc (Nisell et al., 1994; Schilstromet al., 2000). Moreover, nicotine enhances synaptic plasticityin the glutamatergic inputs to the VTA (Mansvelder andMcGehee, 2000). Therefore, increasing evidence shows thatVTA is critically involved in nicotine’s rewarding effects.However, VTA involvement in nicotine self-administration be-haviour has never been directly investigated, and the underly-ing neuronal mechanisms remain to be elucidated (Nomikoset al., 2000).
Here we used an intracranial self-administration (ICSA)paradigm based on a spatial discrimination task in a Y-mazeto investigate whether nicotine microinjections into the VTAwould elicit rewarding effects in C57BL/6J mice, this inbredstrain being the common genetic background of knock-outand knock-in strains relevant to nicotine research. Discrimina-tion tasks based on choice behaviour provides one of the mostrelevant methods for the assessment of reward in animals(Mason et al., 1985). The present self-administration modelcombines drug-induced spatial discrimination with voluntarydrug intake, therefore adding face validity to the predictivevalidity of classical drug-induced place conditioning studies(Bardo and Bevins, 2000). The neuropharmacological basesof intra-VTA nicotine self-administration were further ex-plored by assessing the involvement of DA-D1 and VTA nic-otinic receptors in this behaviour using systemic or localinjection, respectively, of specific antagonists.
2. Methods
2.1. Ethical statement
All surgical and experimental procedures were conducted in accordance
with the European Communities Council Directive of 24 November 1986
(86/609/EEC).
2.2. Animals and surgery
The present experiments used male C57BL/6JiCo mice (Janvier, Lyon,
France). At 12 weeks of age, they were housed individually with ad libitum
access to food and water in a temperature-controlled room (23 �C) maintained
on a light/dark cycle (12 h/12 h, light on at 08:00 h) and sawdust bedding
changed weekly. Surgery started at least 2 weeks later. The subjects were
deeply anaesthetized with tribromoethanol (Avertin�, 300 mg/kg, i.p.). More-
over, lidocaine HCl (Xylocaine�, 5%) was applied locally both before opening
the scalp and trepanation. The animals were unilaterally implanted in
a counterbalanced, left and right order, and since it has previously been dem-
onstrated that the magnitude of the motivational effects of unilaterally applied
opioids are equivalent to those observed when bilateral injections are used
(Phillips and Le Piane, 1980; Bozarth, 1987). The tip of the guide-cannula
(outer diameter 0.460 mm; inner diameter 0.255 mm) was positioned either
1.5 mm (VTA groups) or 2.3 mm (dorsal control or D-VTA group) above
the VTA. The stereotaxic coordinates used were: 0.40 mm anterior to the in-
teraural line; �0.30 mm lateral to the sagittal line; 2.5 or 3.30 mm vertically
below the surface of the skull. The incisor bar was kept level with the interau-
ral line. Mice were allowed to recover from surgery for 1 week.
2.3. Materials and experimental protocol
2.3.1. Intracranial self-injection procedure
Self-administration behaviour was studied in a grey Plexiglas Y-maze, the
two arms of which were separated by a 90 � angle. The stem and the arms were
31-cm long and 12-cm high. The starting box (14 � 8 cm) was separated from
the stem by a sliding door. A photoelectric cell was situated 6 cm from the end
of each arm. On each day of the experimental period, a stainless-steel injection
cannula (outer diameter 0.229 mm, inner diameter 0.127 mm) was inserted
into the VTA and held in a fixed position by means of a small connector.
The tip of the injection cannula projected beyond the guide cannula by
1.5 mm. It was connected by flexible polyethylene tubing to the microinjection
system, which housed a 5-ml Hamilton syringe. By interrupting the photocell
beam in one of the two target arms, mice could trigger an infusion of nicotine.
The other arm was neutral (no injection). Intracranial injections were carried
out using an automatic computer-controlled apparatus, which provided, via
a microvernier system, a precise and highly reproducible descent of the micro-
syringe piston. Each self-injection (50 nl) lasted 4 s. Normal drug flow was
verified visually both before and after each ICSA session for each animal.
The movements of the animals in the Y-maze were detected using an optical
system. This information was transmitted to a computer, which rotated in
turn the injector in the same direction as each animal’s movement. This
process avoided the twisting of the flexible tubing. Automatic equipment trig-
gered the opening of the door to the stem. The number of self-administrations
per daily session and the latency to enter either the reinforced arm (injection
latency) or the neutral arm were recorded for each subject.
2.3.2. Drugs
(�)-Nicotine di(þ)-tartrate (Sigma-Aldrich�, Saint Quentin Fallavier,
France) was dissolved in polyionic Ringer (Aguettant�, Lyon, France). The so-
lution obtained was very acidic (pH 3.0), thus a small volume of NaOH 1 N
was added in order to attain a pH corresponding to that of the vehicle (pH
6.5). Two doses were tested: 10 ng and 100 ng/50-nl injection or 21.6 pmol
and 216 pmol (as referred to the salt), resulting respectively in nicotine con-
centrations of 0.4 mM and 4 mM (referred to the base). Choice of the initial
dose was based on previous studies of intra-VTA nicotine injections in rats
showing that place preference was obtained with the dose of 2 nmol/0.5 ml/
side (Laviolette and van der Kooy, 2004). The D1 receptor antagonist (þ)-
SCH 23390 HCl ((R)-(þ)-7-chloro-8-hydroxy-3-methyl-1-phenyl-2,3,4,-5-tet-
rahydro-1H-3-benzazepine, Sigma-Aldrich�) was dissolved in isotonic NaCl
0.9% to the required final concentration (25 mg/kg i.p.). Injections were admin-
istered 10 min prior to the self-administration session in a volume of 0.1 ml/
10 g of body weight. The competitive nicotinic cholinergic receptor antagonist
dihydro-b-erythroidine hydrobromide (DHbE, Sigma-Aldrich�) was co-in-
jected with nicotine using an equimolar concentration for nicotine and
DHbE (4 mM). In a different group of mice, the DHbE concentration was
raised to 8 mM.
2.3.3. Behavioural protocol
Four groups were constituted as follows: VTA nicotine 10 ng (n ¼ 6); VTA
nicotine 100 ng (n ¼ 18); D-VTA nicotine 100 ng (n¼ 7), and controls
(n¼ 16) having only vehicle (Ringer) available. Controls were subdivided
into three groups corresponding to Ringer (n ¼ 6), Ringer þ SCH23390
(n ¼ 5) and Ringer þ DHbE (n ¼ 5). The D-VTA nicotine group refers to
animals implanted with the cannula guide 0.8 mm above the VTA nicotine
group.

1032 V. David et al. / Neuropharmacology 50 (2006) 1030e1040
The protocol consisted of three phases:
(i) Acquisition phase. This phase lasted for seven daily sessions. Each ses-
sion comprised the following steps. To begin a trial, a mouse was placed
in the start box and, after 1 min, the door to the stem was opened. In each
group, half of the animals were assigned to the right arm to trigger the
injection of nicotine, whereas the other half of the subjects were assigned
to the left arm. Each daily session was composed of 10 trials each sep-
arated by a 1-min inter-trial interval. Therefore, a maximum of 10 injec-
tions could be obtained by each subject per daily session. During the first
four trials of the first session only, if an animal made an error in choosing
the neutral arm, it was immediately allowed to access to the arm enabling
an injection of nicotine. From the fifth trial onward and during the fol-
lowing sessions, if an animal entered into the neutral arm, it was replaced
into the start box for the following trial.
(ii) Challenge with either DA or nicotinic cholinergic antagonists. Mice which
had previously acquired nicotine self-administration ICSA with the 10-ng
dose were then pre-injected with the DA D1-receptor antagonist (þ)-SCH
23390 HCl (25 mg/kg, i.p.) for five consecutive sessions. Furthermore
a first subgroup (n ¼ 6) of subjects that exhibited nicotine self-administra-
tion with the 100-ng dose received the same pretreatment. The remaining
mice of the 100-ng nicotine ICSA group continued to receive nicotine into
the VTA, but were also co-injected with the nicotinic receptor antagonist
DHbE at either 4 mM (equimolar, n ¼ 6) or 8 mM concentrations (n ¼ 6).
(iii) Replacement of antagonists with vehicle (recovery test). Following the last
antagonist treatment session, SCH 23390 was then replaced by pre-trial
injections of vehicle alone for two consecutive daily sessions. Concur-
rently, mice which had previously received DHbE had access to nico-
tine-only infusions.
2.4. Histology
At the end of the experimental period, animals were sacrificed with an
overdose of Avertin�. The head was removed, with the guide cannula attached,
and placed into 10% formol for a 72-h period. The guide cannula was then
withdrawn, the brain dissected and placed in a solution of formol containing
30% sucrose for an additional week. Brains were then frozen and cut in a mi-
crotome to provide 60-mm frontal sections, which were stained using 0.1% thi-
onine to identify the injection site.
2.5. Statistical analyses
To analyse acquisition of nicotine self-administration, ANOVAs for the
number of nicotine self-injections and self-injection latencies were conducted
with group (drug or dose) as a between-subjects factor and session (training
day) as a within-subjects repeated measure. The same parameters were analysed
following pharmacological challenge of intra-VTA nicotine self-administration
using antagonists (SCH 23390 or DHbE) as a between-subjects factor and ses-
sion as a within-subjects repeated measure. Significant main effects were fur-
ther analysed using Fisher’s PLSD tests. Paired Student’s t tests were used to
compare either the number of self-administrations or time to trigger the injec-
tion between two sessions in the same group (e.g. session E5 versus R1). A min-
imum significance level of p < 0.05 was required for all statistical analyses.
3. Results
3.1. Acquisition of nicotine ICSA
3.1.1. Spatial discriminationNo discrimination between the two arms of the Y-maze was
observed in subjects having access only to vehicle (Ringer) forintra-VTA self-injections, demonstrating the absence of anynon-specific (chemical or mechanical) stimulating effects ofthe microinjection system and confirming previous observa-tions in C57BL/6 mice (David et al., 2004). In contrast micehaving access to the 10-ng dose of nicotine (0.4 mM)
exhibited significant preference for the nicotine-associatedarm of the Y-maze. Accuracy of spatial discrimination furtherincreased significantly with the nicotine dose of 100 ng(4 mM). These observations are supported by a two-wayANOVA, revealing main effects of nicotine on choice perfor-mance [main effect of drug F(2,15) ¼ 20.44 p < 0.0001; maineffect of session: F(6,90) ¼ 12.05 p < 0.001; drug � sessioninteraction: F(12,90) ¼ 3.60 p < 0.001; post-hoc comparisonof doses: p < 0.05] (Fig. 1A, left panel). When nicotine wasinjected 0.8 mm dorsally to the VTA (D-VTA group), self-administration was no longer observed. This demonstratesthat neurons located within the VTA or contained withina 0.8 mm radius sphere around the injection cannula tip areresponsible for this behaviour [main effect of injection site(dose of 100 ng): F(1,19) ¼ 79.68 p < 0.0001; main effect ofsession: F(6,114) ¼ 6.27 p < 0.001; site � session interaction:F(6,114) ¼ 8.15 p < 0.001] (Fig. 1C, left panel).
3.1.2. Self-injection latenciesAnalyses of self-injection latencies revealed that this
parameter increased over sessions in the vehicle-injected group,whereas a significant decrease was observed in all nicotine-injected mice [main effect of drug: F(2,15) ¼ 23.16 p <0.0001; main effect of session: F(6,90) ¼ 1.15 ns; drug �session interaction F(12,90) ¼ 2.25 p < 0.01] (Fig. 1B, leftpanel). Mice self-administering 100 ng-nicotine dose tendedto be faster to trigger the injections than subjects of the 10-nggroup, although this trend did not reach statistical significance(Fig. 1B; left panel). When infused 0.8 mm dorsally to theVTA, nicotine produced an effect similar to vehicle injectionson the latency, i.e. increasing instead of decreasing this measure[main effect of injection site (dose of 100 ng): F(1,19) ¼ 164.34p < 0.0001; main effect of session: F(6,114) ¼ 2.21 p < 0.05;site � session interaction: F(6,114) ¼ 9.82 p < 0.0001](Fig. 1D, left panel).
3.2. Effects of the DA-D1 receptor antagonist((þ)-SCH 23390)
3.2.1. Spatial discriminationPre-treatment with SCH 23390 (25 mg/kg, i.p., n ¼ 5)
10 min before each session in mice self-injecting either the10-ng or 100-ng nicotine dose clearly altered nicotine ICSA.A significant reduction in the number of nicotine-reinforcedarm choices was observed from the first pretreatment session(E1) in the 100-ng group, and from the second treatmentsession (E2) in the 10-ng group [comparison between thelast acquisition session A7 and the first extinction session E1,respectively: t(5) ¼ 5.96 p < 0.01); t(5) ¼ 3.46 p < 0.02]. Dis-crimination performance decreased, reaching chance levelduring the third session (E3: Fig. 1A, mid panel). SCH23390 treatment did not have any significant effects on thechance level performance of subjects having only the vehicleavailable. Because of this rapid extinction, there was no differ-ence in the number of self-injections between the three groupstested over the five extinction sessions but a main effect ofsession, thus confirming the extinction of self-administration
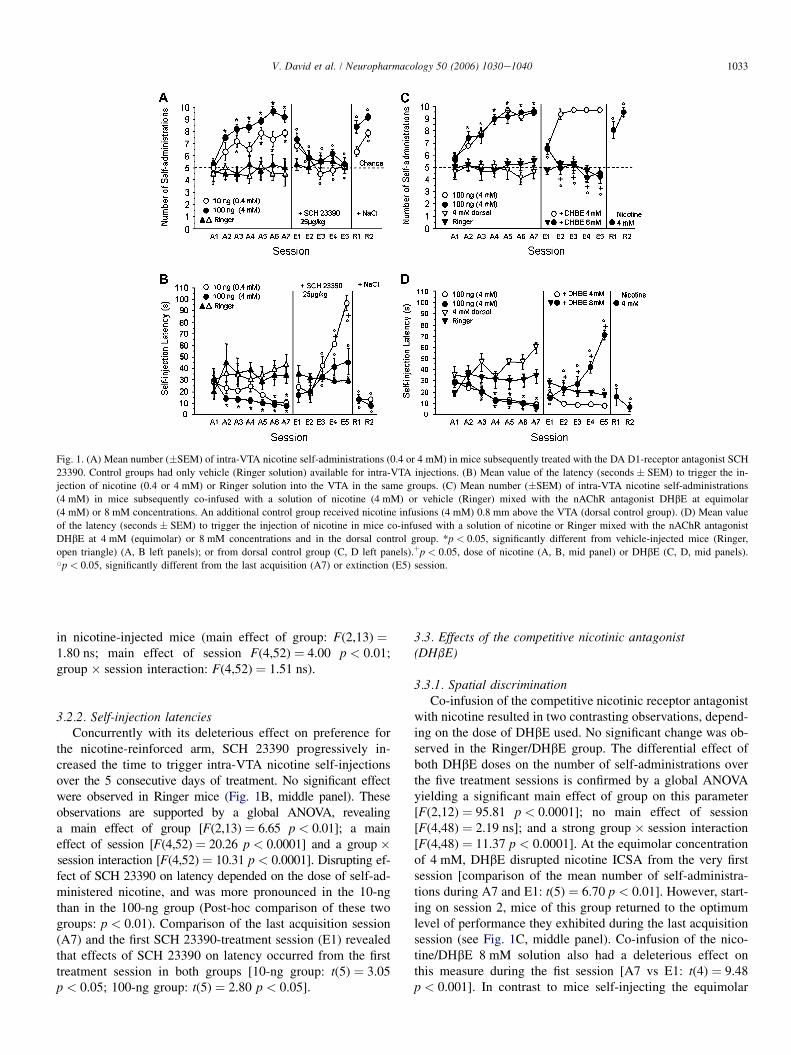
1033V. David et al. / Neuropharmacology 50 (2006) 1030e1040
Fig. 1. (A) Mean number (�SEM) of intra-VTA nicotine self-administrations (0.4 or 4 mM) in mice subsequently treated with the DA D1-receptor antagonist SCH
23390. Control groups had only vehicle (Ringer solution) available for intra-VTA injections. (B) Mean value of the latency (seconds � SEM) to trigger the in-
jection of nicotine (0.4 or 4 mM) or Ringer solution into the VTA in the same groups. (C) Mean number (�SEM) of intra-VTA nicotine self-administrations
(4 mM) in mice subsequently co-infused with a solution of nicotine (4 mM) or vehicle (Ringer) mixed with the nAChR antagonist DHbE at equimolar
(4 mM) or 8 mM concentrations. An additional control group received nicotine infusions (4 mM) 0.8 mm above the VTA (dorsal control group). (D) Mean value
of the latency (seconds � SEM) to trigger the injection of nicotine in mice co-infused with a solution of nicotine or Ringer mixed with the nAChR antagonist
DHbE at 4 mM (equimolar) or 8 mM concentrations and in the dorsal control group. *p < 0.05, significantly different from vehicle-injected mice (Ringer,
open triangle) (A, B left panels); or from dorsal control group (C, D left panels).þp < 0.05, dose of nicotine (A, B, mid panel) or DHbE (C, D, mid panels).Bp < 0.05, significantly different from the last acquisition (A7) or extinction (E5) session.
in nicotine-injected mice (main effect of group: F(2,13) ¼1.80 ns; main effect of session F(4,52) ¼ 4.00 p < 0.01;group � session interaction: F(4,52) ¼ 1.51 ns).
3.2.2. Self-injection latenciesConcurrently with its deleterious effect on preference for
the nicotine-reinforced arm, SCH 23390 progressively in-creased the time to trigger intra-VTA nicotine self-injectionsover the 5 consecutive days of treatment. No significant effectwere observed in Ringer mice (Fig. 1B, middle panel). Theseobservations are supported by a global ANOVA, revealinga main effect of group [F(2,13)¼ 6.65 p < 0.01]; a maineffect of session [F(4,52)¼ 20.26 p < 0.0001] and a group �session interaction [F(4,52) ¼ 10.31 p < 0.0001]. Disrupting ef-fect of SCH 23390 on latency depended on the dose of self-ad-ministered nicotine, and was more pronounced in the 10-ngthan in the 100-ng group (Post-hoc comparison of these twogroups: p < 0.01). Comparison of the last acquisition session(A7) and the first SCH 23390-treatment session (E1) revealedthat effects of SCH 23390 on latency occurred from the firsttreatment session in both groups [10-ng group: t(5) ¼ 3.05p < 0.05; 100-ng group: t(5) ¼ 2.80 p < 0.05].
3.3. Effects of the competitive nicotinic antagonist(DHbE)
3.3.1. Spatial discriminationCo-infusion of the competitive nicotinic receptor antagonist
with nicotine resulted in two contrasting observations, depend-ing on the dose of DHbE used. No significant change was ob-served in the Ringer/DHbE group. The differential effect ofboth DHbE doses on the number of self-administrations overthe five treatment sessions is confirmed by a global ANOVAyielding a significant main effect of group on this parameter[F(2,12) ¼ 95.81 p < 0.0001]; no main effect of session[F(4,48) ¼ 2.19 ns]; and a strong group � session interaction[F(4,48) ¼ 11.37 p < 0.0001]. At the equimolar concentrationof 4 mM, DHbE disrupted nicotine ICSA from the very firstsession [comparison of the mean number of self-administra-tions during A7 and E1: t(5) ¼ 6.70 p < 0.01]. However, start-ing on session 2, mice of this group returned to the optimumlevel of performance they exhibited during the last acquisitionsession (see Fig. 1C, middle panel). Co-infusion of the nico-tine/DHbE 8 mM solution also had a deleterious effect onthis measure during the fist session [A7 vs E1: t(4) ¼ 9.48p < 0.001]. In contrast to mice self-injecting the equimolar
1034 V. David et al. / Neuropharmacology 50 (2006) 1030e1040
nicotine/DHbE solution, this group no longer exhibited anypreference for the nicotine-associated arm, reaching chancelevel within two sessions (Fig. 1C).
3.3.2. Self-injection latencyAs previously observed for the number of self-administrations,
the temporal evolution of this parameter was in marked opposi-tion as a function of the DHbE dose, starting on session 2(Fig. 1D, middle panel). Initial statistical analysis revealed nomain effect of group [F(2,12) ¼ 3.31 ns]; but a main effect ofsession [F(4,48) ¼ 2.66 p < 0.05]; and a strong dose � sessioninteraction [F(4,48)¼ 32.03 p< 0.0001]. Co-injection ofa nicotine/DHbE solution resulted in a similar increase of self-injection la-tency during session 1, whatever the dose of DHbE used [4 mM:t(5) ¼ 3.87 p < 0.01; 8 mM: t(4) ¼ 5.47 p < 0.01]. However,mice co-infused with the equimolar nicotine/DHbE solutionexhibited latencies similar to their baseline (pretreatment) levelfrom session 2 to 5. In contrast, latency increased significantlyand progressively over the four remaining sessions in miceself-injecting the nicotine/DHbE 8 mM solution, whereas thisparameter decreased slightly in the Ringer/DHbE group [A7vs E5: t(4) ¼ 3.80 p < 0.05].
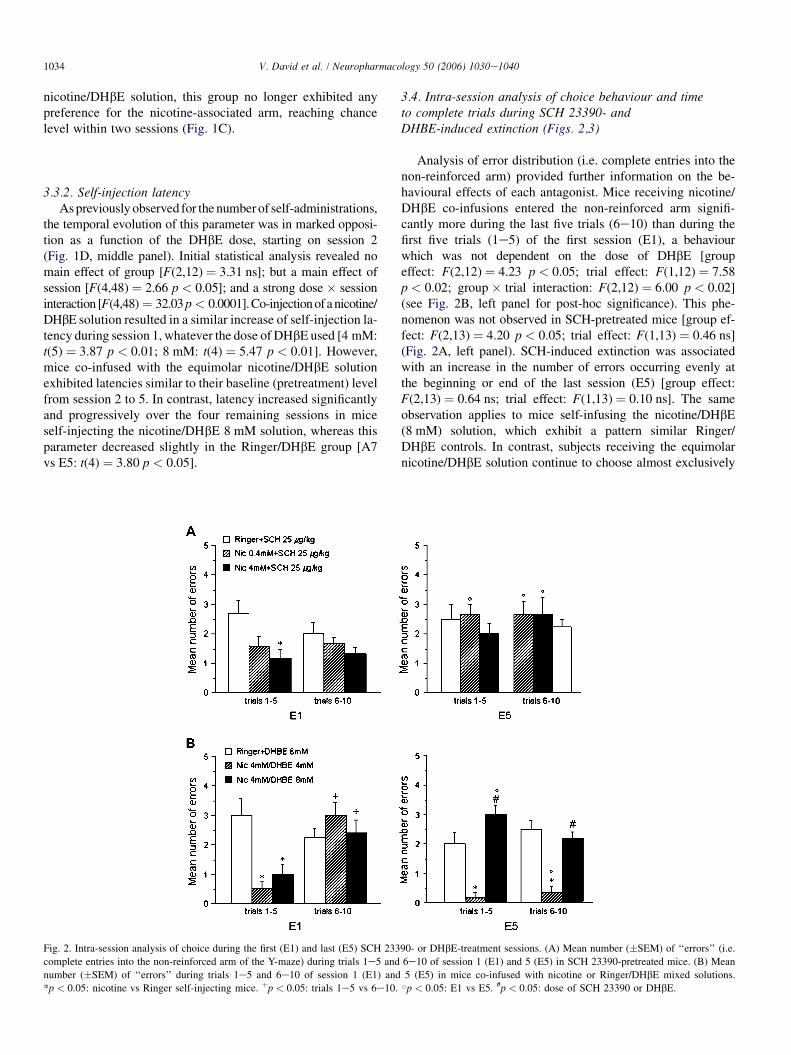
3.4. Intra-session analysis of choice behaviour and timeto complete trials during SCH 23390- andDHBE-induced extinction (Figs. 2,3)
Analysis of error distribution (i.e. complete entries into thenon-reinforced arm) provided further information on the be-havioural effects of each antagonist. Mice receiving nicotine/DHbE co-infusions entered the non-reinforced arm signifi-cantly more during the last five trials (6e10) than during thefirst five trials (1e5) of the first session (E1), a behaviourwhich was not dependent on the dose of DHbE [groupeffect: F(2,12) ¼ 4.23 p < 0.05; trial effect: F(1,12) ¼ 7.58p < 0.02; group � trial interaction: F(2,12) ¼ 6.00 p < 0.02](see Fig. 2B, left panel for post-hoc significance). This phe-nomenon was not observed in SCH-pretreated mice [group ef-fect: F(2,13) ¼ 4.20 p < 0.05; trial effect: F(1,13) ¼ 0.46 ns](Fig. 2A, left panel). SCH-induced extinction was associatedwith an increase in the number of errors occurring evenly atthe beginning or end of the last session (E5) [group effect:F(2,13) ¼ 0.64 ns; trial effect: F(1,13) ¼ 0.10 ns]. The sameobservation applies to mice self-infusing the nicotine/DHbE(8 mM) solution, which exhibit a pattern similar Ringer/DHbE controls. In contrast, subjects receiving the equimolarnicotine/DHbE solution continue to choose almost exclusively
Fig. 2. Intra-session analysis of choice during the first (E1) and last (E5) SCH 23390- or DHbE-treatment sessions. (A) Mean number (�SEM) of ‘‘errors’’ (i.e.
complete entries into the non-reinforced arm of the Y-maze) during trials 1e5 and 6e10 of session 1 (E1) and 5 (E5) in SCH 23390-pretreated mice. (B) Mean
number (�SEM) of ‘‘errors’’ during trials 1e5 and 6e10 of session 1 (E1) and 5 (E5) in mice co-infused with nicotine or Ringer/DHbE mixed solutions.
*p < 0.05: nicotine vs Ringer self-injecting mice. þp < 0.05: trials 1e5 vs 6e10. Bp < 0.05: E1 vs E5. #p < 0.05: dose of SCH 23390 or DHbE.
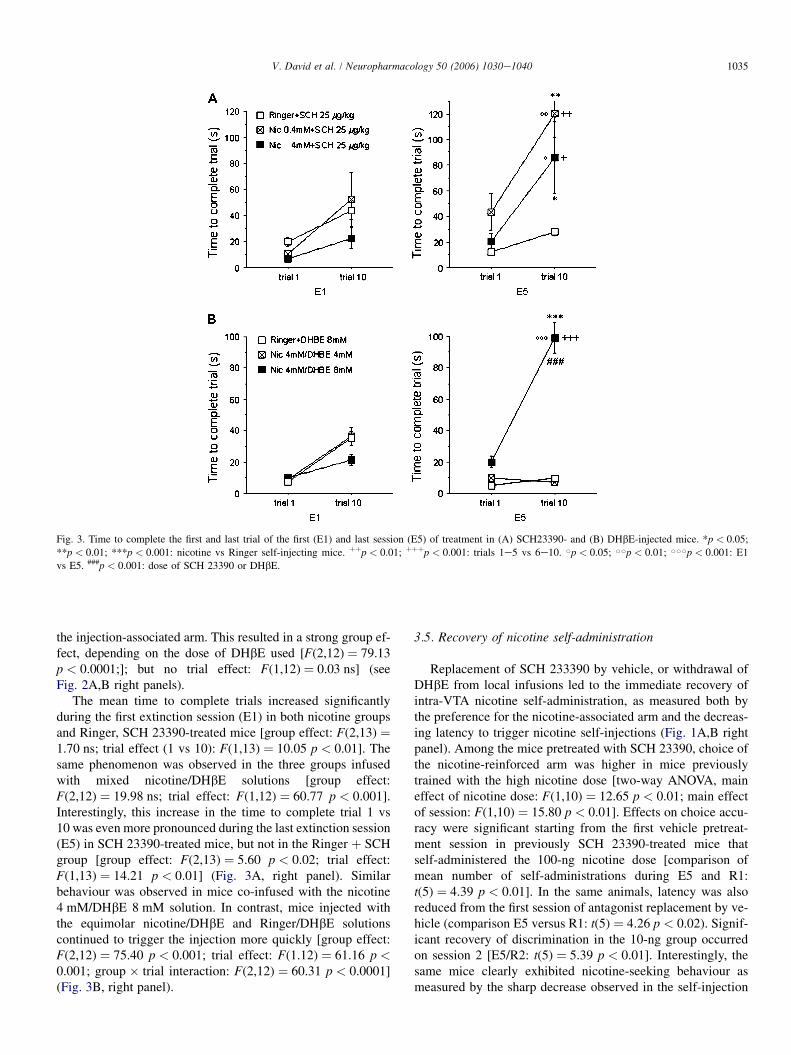
1035V. David et al. / Neuropharmacology 50 (2006) 1030e1040
Fig. 3. Time to complete the first and last trial of the first (E1) and last session (E5) of treatment in (A) SCH23390- and (B) DHbE-injected mice. *p < 0.05;
**p < 0.01; ***p < 0.001: nicotine vs Ringer self-injecting mice. þþp < 0.01; þþþp < 0.001: trials 1e5 vs 6e10. Bp < 0.05; BBp < 0.01; BBBp < 0.001: E1
vs E5. ###p < 0.001: dose of SCH 23390 or DHbE.
the injection-associated arm. This resulted in a strong group ef-fect, depending on the dose of DHbE used [F(2,12) ¼ 79.13p < 0.0001;]; but no trial effect: F(1,12) ¼ 0.03 ns] (seeFig. 2A,B right panels).
The mean time to complete trials increased significantlyduring the first extinction session (E1) in both nicotine groupsand Ringer, SCH 23390-treated mice [group effect: F(2,13) ¼1.70 ns; trial effect (1 vs 10): F(1,13) ¼ 10.05 p < 0.01]. Thesame phenomenon was observed in the three groups infusedwith mixed nicotine/DHbE solutions [group effect:F(2,12) ¼ 19.98 ns; trial effect: F(1,12) ¼ 60.77 p < 0.001].Interestingly, this increase in the time to complete trial 1 vs10 was even more pronounced during the last extinction session(E5) in SCH 23390-treated mice, but not in the Ringer þ SCHgroup [group effect: F(2,13) ¼ 5.60 p < 0.02; trial effect:F(1,13) ¼ 14.21 p < 0.01] (Fig. 3A, right panel). Similarbehaviour was observed in mice co-infused with the nicotine4 mM/DHbE 8 mM solution. In contrast, mice injected withthe equimolar nicotine/DHbE and Ringer/DHbE solutionscontinued to trigger the injection more quickly [group effect:F(2,12) ¼ 75.40 p < 0.001; trial effect: F(1.12) ¼ 61.16 p <0.001; group � trial interaction: F(2,12) ¼ 60.31 p < 0.0001](Fig. 3B, right panel).
3.5. Recovery of nicotine self-administration
Replacement of SCH 233390 by vehicle, or withdrawal ofDHbE from local infusions led to the immediate recovery ofintra-VTA nicotine self-administration, as measured both bythe preference for the nicotine-associated arm and the decreas-ing latency to trigger nicotine self-injections (Fig. 1A,B rightpanel). Among the mice pretreated with SCH 23390, choice ofthe nicotine-reinforced arm was higher in mice previouslytrained with the high nicotine dose [two-way ANOVA, maineffect of nicotine dose: F(1,10) ¼ 12.65 p < 0.01; main effectof session: F(1,10) ¼ 15.80 p < 0.01]. Effects on choice accu-racy were significant starting from the first vehicle pretreat-ment session in previously SCH 23390-treated mice thatself-administered the 100-ng nicotine dose [comparison ofmean number of self-administrations during E5 and R1:t(5) ¼ 4.39 p < 0.01]. In the same animals, latency was alsoreduced from the first session of antagonist replacement by ve-hicle (comparison E5 versus R1: t(5) ¼ 4.26 p < 0.02). Signif-icant recovery of discrimination in the 10-ng group occurredon session 2 [E5/R2: t(5) ¼ 5.39 p < 0.01]. Interestingly, thesame mice clearly exhibited nicotine-seeking behaviour asmeasured by the sharp decrease observed in the self-injection
1036 V. David et al. / Neuropharmacology 50 (2006) 1030e1040
latency on the first recovery session [E5/R1: t(5) ¼ 12.94p < 0.001] (Fig. 1B, right panel).
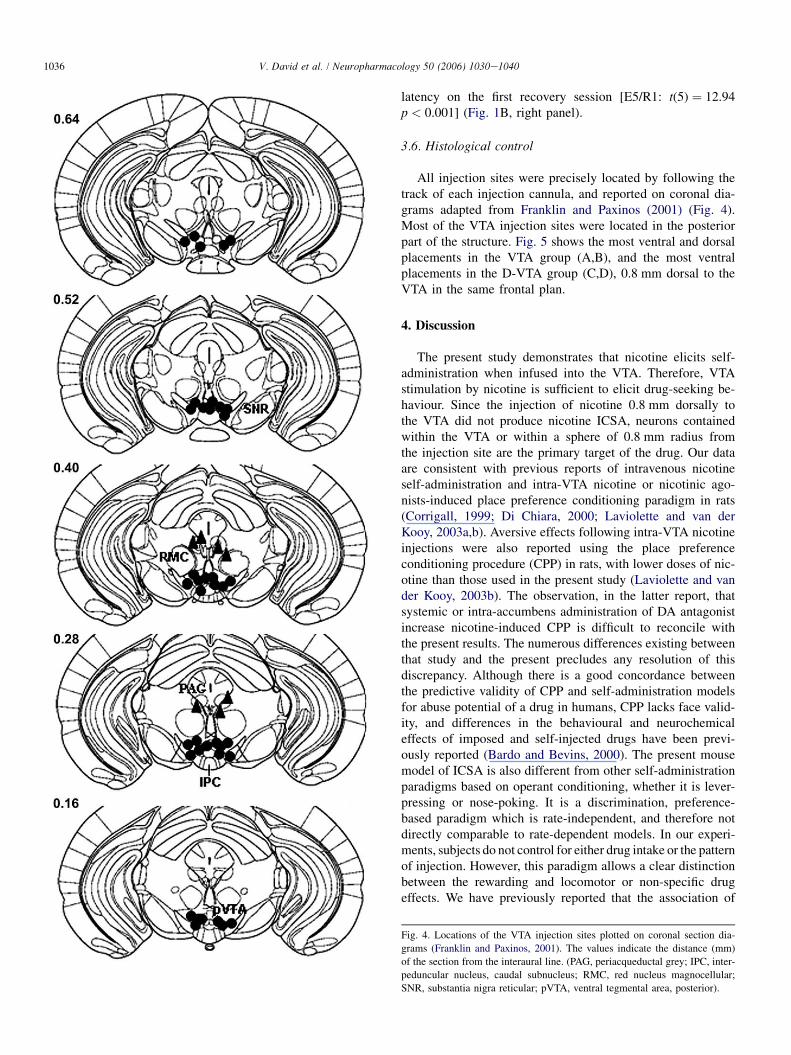
3.6. Histological control
All injection sites were precisely located by following thetrack of each injection cannula, and reported on coronal dia-grams adapted from Franklin and Paxinos (2001) (Fig. 4).Most of the VTA injection sites were located in the posteriorpart of the structure. Fig. 5 shows the most ventral and dorsalplacements in the VTA group (A,B), and the most ventralplacements in the D-VTA group (C,D), 0.8 mm dorsal to theVTA in the same frontal plan.
4. Discussion
The present study demonstrates that nicotine elicits self-administration when infused into the VTA. Therefore, VTAstimulation by nicotine is sufficient to elicit drug-seeking be-haviour. Since the injection of nicotine 0.8 mm dorsally tothe VTA did not produce nicotine ICSA, neurons containedwithin the VTA or within a sphere of 0.8 mm radius fromthe injection site are the primary target of the drug. Our dataare consistent with previous reports of intravenous nicotineself-administration and intra-VTA nicotine or nicotinic ago-nists-induced place preference conditioning paradigm in rats(Corrigall, 1999; Di Chiara, 2000; Laviolette and van derKooy, 2003a,b). Aversive effects following intra-VTA nicotineinjections were also reported using the place preferenceconditioning procedure (CPP) in rats, with lower doses of nic-otine than those used in the present study (Laviolette and vander Kooy, 2003b). The observation, in the latter report, thatsystemic or intra-accumbens administration of DA antagonistincrease nicotine-induced CPP is difficult to reconcile withthe present results. The numerous differences existing betweenthat study and the present precludes any resolution of thisdiscrepancy. Although there is a good concordance betweenthe predictive validity of CPP and self-administration modelsfor abuse potential of a drug in humans, CPP lacks face valid-ity, and differences in the behavioural and neurochemicaleffects of imposed and self-injected drugs have been previ-ously reported (Bardo and Bevins, 2000). The present mousemodel of ICSA is also different from other self-administrationparadigms based on operant conditioning, whether it is lever-pressing or nose-poking. It is a discrimination, preference-based paradigm which is rate-independent, and therefore notdirectly comparable to rate-dependent models. In our experi-ments, subjects do not control for either drug intake or the patternof injection. However, this paradigm allows a clear distinctionbetween the rewarding and locomotor or non-specific drugeffects. We have previously reported that the association of
Fig. 4. Locations of the VTA injection sites plotted on coronal section dia-
grams (Franklin and Paxinos, 2001). The values indicate the distance (mm)
of the section from the interaural line. (PAG, periacqueductal grey; IPC, inter-
peduncular nucleus, caudal subnucleus; RMC, red nucleus magnocellular;
SNR, substantia nigra reticular; pVTA, ventral tegmental area, posterior).

1037V. David et al. / Neuropharmacology 50 (2006) 1030e1040
Fig. 5. Photomicrographs of thionin-stained frontal brain sections (60 mm) through the guide-cannula tracks and the injection site into the VTA (A,B) and 0.8 mm
dorsally to the VTA (C,D; D-VTA group) in the same frontal plane. Scale Bar 0.5 mm.
high discrimination performances with short latencies revealsgreater rewarding effects, whereas locomotor activating effectsare typically associated with a decrease in discrimination per-formance (David et al., 2002). Furthermore, nicotine hasweak activating effects in rats and fails to induce locomotor ac-tivation in mice (Villegier et al., 2005). Therefore, the progres-sive decrease of injection latencies observed in self-injectinganimals likely reveals the learning of nicotine-seeking behav-iour, rather than direct locomotor effects of the drug.
Systemic injection of the D1 DA receptor antagonist SCH23390, or co-infusion of nicotine with DHbE, a nAChR antag-onist, disrupted the nicotine ICSA. DHBE blocked nicotineICSA only when co-infused at a concentration twice that of nic-otine. Despite a slight increase in the choice of the nicotine-as-sociated arm during session 4 of the SCH 23390 treatment in100 ng nicotine-injected mice, there was no significant signof functional recovery of DA receptors. If that was the case, dis-crimination performance should have been increasing over sev-eral consecutive sessions, as previously observed followingchronic DA D2 antagonist treatment (David et al., 2002). De-creased ICSA of nicotine is unlikely to have resulted from me-chanical lesion due to the daily insertion of the cannula or thesystemic injection per se, since ICSA behaviour rapidly reap-peared when both antagonists were replaced by saline. Intracra-nial infusion of either DHBE or SCH 23390 have beenpreviously reported not to produce lesions or long-term block-ade of the receptors (Granon et al., 1995, 2000). Alternatively,the blocking effects of both antagonists on nicotine ICSA mayinvolve non-specific motor, perceptual or memory deficits. Thisis especially critical for the DA D1-receptor antagonist pre-treatment, as SCH 23390 was administered systemically. Theobservation that self-injection latency increased progressivelyover sessions is consistent with motivational effects, whereasmotor impairment should have been observed from the first trialof each treatment session (Beninger and Miller, 1998). In con-trast, intra-session analysis of the time to complete trials
revealed that increase of this parameter occurred at the end ofeither the first (E1) or the last (E5) SCH23390-treatment ses-sions. This behaviour was also observed with the dose ofDHbE inducing extinction. Finally, SCH 23390 did not haveany effect on motor activity in mice self-injecting Ringer.
D1 receptors are involved in acquisition and expression ofappetitive associative or instrumental learning, spatial memoryand early consolidation (Sawaguchi and Goldman-Rakic,1991; Beninger and Miller, 1998; El-Ghundi et al., 1999;Floresco and Phillips, 2001; Mele et al., 2004; Dalley et al.,2005). Coincident activation of D1 and NMDA receptor withinthe nucleus accumbens or the prefrontal cortex is required forappetitive instrumental learning (Smith-Roe and Kelley, 2000;Baldwin et al., 2002). However, several lines of evidence sug-gest that memory-disrupting effects of SCH 23390 may not ac-count for extinction. D1-receptor antagonist treatment disruptsacquisition/consolidation rather than retention/retrieval of ap-petitive conditioning (Smith-Roe and Kelley, 2000; Baldwinet al., 2002; Dalley et al., 2005). In the present study, SCH23390 was administered only during the retention phase ofICSA. Furthermore, systemic SCH 23390 pre-treatment doesnot prevent the acquisition of a Y-maze spatial discriminationtask when this task is reinforced with food (data not shown).Motivational rather than memory effects are indicated sincechoice behaviour and latency were disrupted mostly by theend of each extinction session. Although performance in thehidden platform version of the Morris Water-Maze task isslightly affected in mice lacking D1 receptors, visually cuedtasks, spontaneous alternation and contextual learning are in-tact (El-Ghundi et al., 1999). Finally, systemic administrationof SCH 23390 does not disrupt reference memory (Bushnelland Levin, 1993; Levin et al., 1996). Together, these datasupport the view that neither the memory-disrupting nor thelocomotor effects of SCH 23390 account for the extinctionof nicotine ICSA. Instead, SCH-treatment may produce a de-crease in the reinforcing value of nicotine, as is observed in

1038 V. David et al. / Neuropharmacology 50 (2006) 1030e1040
subjects undergoing classical extinction (i.e. replacement ofthe drug by its vehicle alone, David et al., 2002).
Co-administration of nicotine with an equimolar dose ofDHbE, a nAChR antagonist, induced only a transient decreaseof nicotine ICSA (first session under DHbE treatment)followed by a rapid improvement of the performance. Sincemice re-attained their pre-treatment level of discriminationwithin two sessions, it is likely that nicotine reward was par-tially blocked by the competitive antagonist. Consistentlywith this interpretation, increasing the dose of DHbE eventu-ally resulted in a rapid and significant decrease of the ICSAresponse. It is noteworthy that intra-VTA, but not intra-accumbens, injections of DHbE disrupt intravenous nicotineself-administration (Corrigall et al., 1994). This observationsupports the results of the present study in showing thatVTA nicotinic receptors are crucial for nicotine’s rewardingeffects. DHbE shows a relatively high affinity for hetero-meric-mainly a4b2- nAChR. These subunits are specificallyexpressed in mesencephalic DA and GABA neurons (Le No-vere et al., 1996). Accordingly, mutant mice lacking the b2subunit of nAChRs are impaired in showing both nicotine-in-duce DA release in the NAc and intravenous nicotine self-ad-ministration (Picciotto et al., 1998).
Recent studies have expanded initial observations high-lighting the role of DA systems in the reinforcing effect ofnicotine. Impairment of DA function using 6-hydroxydop-amine lesions or DA receptor antagonists demonstrate thatDA is involved in nicotine-induced facilitation of electricalself-stimulation, intravenous nicotine self-administration andnicotine induced place preference conditioning (Di Chiara,2000). Microdialysis studies reported that systemic, intra-NAcor intra-VTA infusion of nicotine increases DA release inthe NAc (Ferrari et al., 2002; Tizabi et al., 2002). Active im-munization against nicotine prevented the nicotine-induced in-crease in DA release in the NAc shell (de Villiers et al., 2002).Moreover, acute or chronic systemic application of nicotineexerts stimulatory effect on somatodendritic DA release inthe VTA (Rahman et al., 2003). The involvement of restrictedterminal regions of the mesocorticolimbic system in the main-tenance of nicotine self-administration was also demonstratedby mapping changes of c-fos expression (Pagliusi et al., 1996).Various experimental data support the notion that acute effectsof nicotine on mesolimbic DA transmission result primarilyfrom activation of nAChRs in the VTA rather than in theNAc. For instance, local infusion of the nAChR antagonistmecamylamine into the VTA but not into the NAc, antago-nizes the systemic nicotine-induced DA release within theNAc (Nisell et al., 1994). Similarly, a reduction in NAc DArelease in chronically nicotine-treated animals was observedafter selective blockade of nAChR in the VTA (Hildebrandet al., 1999), but not when the same treatment was appliedin the NAc (Hildebrand and Svensson, 2000). Therefore,nAChRs located in the VTA appear to be crucial for the stim-ulatory effects of nicotine on NAc DA release and behaviour(Hildebrand and Svensson, 2000).
In summary, the present report demonstrates that nicotineacts within the VTA to elicit reward in C57BL/6J mice, as
revealed by the rapid acquisition of intra-VTA nicotine self-administration. Nicotine-seeking behaviour relies on localheteropentameric nAChRs, as well as functional DA-D1 recep-tors. The intracranial nicotine self-administration paradigmused in the present study provides a suitable experimentalprocedure allowing to further investigate the neuropharmaco-logical basis of nicotine’s addictive properties. The availabilityof several lines of knock-out mice lacking specific subunits ofnAChRs offers an opportunity to unravel the critical neuronalprocesses underlying nicotine addiction and to dissect therespective role of the various nAChR subunits in theseprocesses.
Acknowledgements
We would like to thank Mrs L. Decorte and D. Panzeri fortheir excellent technical assistance as well as Dr T.P. Durkinfor his help and suggestions for the English text. This investi-gation was supported by the Centre National de la RechercheScientifique (UMR 5106) and the Institut Pasteur. MB is sup-ported by a grant from the Letten F. Saugstad foundation.
References
Baldwin, A.E., Sadeghian, K., Kelley, A.E., 2002. Appetitive instrumental
learning requires coincident activation of NMDA and dopamine D1 recep-
tors within the medial prefrontal cortex. J. Neurosci. 22, 1063e1071.
Bardo, M.T., Bevins, R.A., 2000. Conditioned place preference: what does it
add to our preclinical understanding of drug reward? Psychopharmacology
153, 31e43.
Beninger, R.J., Miller, R., 1998. Dopamine D1-like receptors and reward-
related incentive learning. Neurosci. Biobehav. Rev. 22, 335e345.
Berridge, K.C., Robinson, T.E., 1998. What is the role of dopamine in reward:
hedonic impact, reward learning, or incentive salience? Brain Res. Brain
Res. Rev. 28, 309e369.
Bozarth, M.A., 1987. Neuroanatomical boundaries of the reward relevant
opiate receptors field in the ventral tegmental area as mapped by the
conditioned place preference method in rats. Brain Res. 414, 77e84.
Bruijnzeel, A.W., Markou, A., 2003. Characterization of the effects of bupro-
pion on the reinforcing properties of nicotine and food in rats. Synapse 50,
20e28.
Bushnell, P.J., Levin, E.D., 1993. Effects of dopaminergic drugs on working
and reference memory in rats. Pharmacol. Biochem. Behav. 45, 765e776.
Corrigall, W.A., Coen, K.M., 1991. Selective dopamine antagonists reduce
nicotine self-administration. Psychopharmacology 104, 171e176.
Corrigall, W.A., Coen, K.M., Adamson, K.L., 1994. Self-administered nicotine
activates the mesolimbic dopamine system through the ventral tegmental
area. Brain Res. 653, 278e284.
Corrigall, W.A., 1999. Nicotine self-administration in animals as a dependence
model. Nicotine. Tob. Res. 1, 11e20.
Dalley, J.W., Laane, K., Theobald, D.E., Armstrong, H.C., Corlett, P.R.,
Chudasama, Y., Robbins, T.W., 2005. Time-limited modulation of appeti-
tive Pavlovian memory by D1 and NMDA receptors in the nucleus accum-
bens. Proc. Natl. Acad. Sci. USA. 102, 6189e6194.
David, V., Durkin, T.P., Cazala, P., 2002. Differential effects of the D2/D3
dopaminergic antagonist on self-administration of morphine into the ven-
tral tegmental area or the nucleus accumbens. Psychopharmacology 160,
307e317.
David, V., Segu, L., Buhot, M.-C., Ichaye, M., Cazala, P., 2004. Rewarding
effects elicited by cocaine microinjections into the ventral tegmental
area of C57BL/6 mice: involvement of dopamine D1 and serotonin1B
receptors. Psychopharmacology 174, 367e375.

1039V. David et al. / Neuropharmacology 50 (2006) 1030e1040
Di Chiara, G., 2000. Role of dopamine in the behavioural actions of nicotine
related to addiction. Eur. J. Pharmacol. 393, 295e314.
Donny, E.C., Caggiula, A.R., Mielke, M.M., Booth, S., Gharib, M.A.,
Hoffman, A., Maldovan, V., Shupenko, C., McCallum, S.E., 1999. Nico-
tine self-administration in rats on a progressive ratio schedule of reinforce-
ment. Psychopharmacology 147, 135e142.
El-Ghundi, M., Fletcher, P.J., Drago, J., Sibley, D.R., O’Dowd, B.F.,
George, S.R., 1999. Spatial learning deficit in dopamine D(1) receptor
knockout mice. Eur. J. Pharmacol. 383, 95e106.
Ferrari, R., Le Novere, N., Picciotto, M.R., Changeux, J.-P., Zoli, M., 2002.
Acute and long-term changes in the mesolimbic dopamine pathway
after systemic or local single nicotine injections. Eur. J. Neurosci. 15,
1810e1818.
Floresco, S.B., Phillips, A.G., 2001. Delay-dependent modulation of memory
retrieval by infusion of a dopamine D1 agonist into the rat medial prefron-
tal cortex. Behav. Neurosci. 115, 934e939.
Franklin, K.B.J., Paxinos, G., 2001. The Mouse Brain in Stereotaxic Coordi-
nates, second ed. Academic Press, San Diego.
Granon, S., Poucet, B., Thinus-Blanc, C., Changeux, J.-P., Vidal, C., 1995.
Nicotinic and muscarinic receptors in the rat prefrontal cortex: differential
roles in working memory, response selection and effortful processing.
Pyschopharmacology 119, 139e144.
Granon, S., Passetti, F., Thomas, K.L., Everitt, B.J., Robbins, T.W., 2000.
Enhanced and impaired attentional performance following infusion of
D1 dopaminergic receptor agents into the rat prefrontal cortex. J. Neurosci.
20, 1208e1215.
Hildebrand, B.E., Panagis, G., Svensson, T.H., Nomikos, G.G., 1999. Behav-
ioral and biochemical manifestations of mecamylamine-precipitated
nicotine withdrawal in the rat: role of nicotinic receptors in the ventral teg-
mental area. Neuropsychopharmacology 21, 560e574.
Hildebrand, B.E., Svensson, T.H., 2000. Intraaccumbal mecamylamine infu-
sion does not affect dopamine output in the nucleus accumbens of chron-
ically nicotine-treated rats. J. Neural. Transm. 107, 861e872.
Imperato, A., Mulas, A., Di Chiara, G., 1986. Nicotine preferentially stimu-
lates dopamine release in the limbic system of freely moving rats. Eur.
J. Pharmacol. 132, 337e338.
Jorenby, D.E., Steinpreis, R.E., Sherman, J.E., Baker, T.B., 1990. Aversion
instead of preference learning indicated by nicotine place conditioning
in rats. Psychopharmacology 101, 533e538.
Laviolette, S.R., van der Kooy, D., 2003a. The motivational valence of
nicotine in the rat ventral tegmental area is switched from rewarding to
aversive following blockade of the alpha7-subunit-containing nicotinic
acetylcholine receptor. Psychopharmacology 166, 306e313.
Laviolette, S.R., van der Kooy, D., 2003b. Blockade of mesolimbic dopamine
transmission dramatically increases sensitivity to the rewarding effects of
nicotine in the ventral tegmental area. Mol. Psychiatry. 8, 50e59.
Laviolette, S.R., van der Kooy, D., 2004. The neurobiology of nicotine addiction:
bridging the gap from molecules to behaviour. Nat. Rev. Neurosci. 5, 55e65.
Le Novere, N., Zoli, M., Changeux, J.-P., 1996. Neuronal nicotinic receptor
alpha 6 subunit mRNA is selectively concentrated in catecholaminergic
nuclei of the rat brain. Eur. J. Neurosci. 8, 2428e2439.
Levin, E.D., Kim, P., Meray, R., 1996. Chronic nicotine working and reference
memory effects in the 16-arm radial maze: interactions with D1 agonist
and antagonist drugs. Psychopharmacology 127, 25e30.
Mansvelder, H.D., McGehee, D.S., 2000. Long-term potentiation of excitatory
inputs to brain reward areas by nicotine. Neuron 27, 349e357.
Mansvelder, H.D., De Rover, M., McGehee, D.S., Brussaard, A.B., 2003. Cho-
linergic modulation of dopaminergic reward areas: upstream and down-
stream targets of nicotine addiction. Eur. J. Pharmacol. 480, 117e123.
Mason, P.A., Milner, P.M., Miousse, R., 1985. Preference paradigm: provides
better self-stimulation reward discrimination than a rate-dependent para-
digm. Behav. Neural Biol. 44, 521e529.
Maskos, U., Molles, B.E., Pons, S., Besson, M., Guiard, B.P., Guilloux, J.P.,
Evrard, A., Cazala, P., Cormier, A., Mameli-Engvall, M., Dufour, N.,
Cloez-Tayarani, I., Bemelmans, A.P., Mallet, J., Gardier, A.M.,
David, V., Faure, P., Granon, S., Changeux, J.P., 2005. Nicotine reinforce-
ment and cognition restored by targeted expression of nicotinic receptors.
Nature 436, 103e107.
Mele, A., Avena, M., Roullet, P., De Leonibus, E., Mandillo, S., Sargolini, F.,
Coccurello, R., Oliverio, A., 2004. Nucleus accumbens dopamine
receptors in the consolidation of spatial memory. Behav. Pharmacol. 15,
423e431.
Nisell, M., Nomikos, G.G., Svensson, T.H., 1994. Systemic nicotine-induced
dopamine release in the rat nucleus accumbens is regulated by nicotinic
receptors in the ventral tegmental area. Synapse 16, 36e44.
Nomikos, G.G., Schilstrom, B., Hildebrand, B.E., Panagis, G., Grenhoff, J.,
Svensson, T.H., 2000. Role of alpha7 nicotinic receptors in nicotine
dependence and implications for psychiatric illness. Behav. Brain Res.
113, 97e103.
Pagliusi, S.R., Tessari, M., DeVevey, S., Chiamulera, C., Pich, E.M., 1996. The
reinforcing properties of nicotine are associated with a specific patterning
of c-fos expression in the rat brain. Eur. J. Neurosci. 8, 2247e2256.
Parker, S.L., Fu, Y., McAllen, K., Luo, J., McIntosh, J.M., Lindstrom, J.M.,
Sharp, B.M., 2004. Up-regulation of brain nicotinic acetylcholine receptors
in the rat during long-term self-administration of nicotine: disproportionate
increase of the alpha6 subunit. Mol. Pharmacol. 65, 611e622.
Paterson, N.E., Markou, A., 2002. Increased GABA neurotransmission via ad-
ministration of gamma-vinyl GABA decreased nicotine self-administration
in the rat. Synapse 44, 252e253.
Phillips, A.G., Le Piane, F.G., 1980. Reinforcing effects of morphine micro-
injection into the ventral tegmental area. Pharmacol. Biochem. Behav.
12, 965e968.
Picciotto, M.R., Zoli, M., Rimondini, R., Lena, C., Marubio, L.M., Pich, E.M.,
Fuxe, K., Changeux, J.-P., 1998. Acetylcholine receptors containing the
beta2 subunit are involved in the reinforcing properties of nicotine. Nature
391, 173e177.
Picciotto, M.R., Corrigall, W.A., 2002. Neuronal systems underlying behaviors
related to nicotine addiction: neural circuits and molecular genetics.
J. Neurosci. 22, 3338e3341.
Rahman, S., Zhang, J., Corrigall, W.A., 2003. Effects of acute and chronic
nicotine on somatodendritic dopamine release of the rat ventral tegmental
area: in vivo microdialysis study. Neurosci. Lett. 348, 61e64.
Rasmussen, T., Swedberg, M.D., 1998. Reinforcing effects of nicotinic
compounds: intravenous self-administration in drug-naive mice. Pharma-
col. Biochem. Behav. 60, 567e573.
Rose, J.E., Corrigall, W.A., 1997. Nicotine self-administration in animals and
humans: similarities and differences. Psychopharmacology 130, 28e40.
Sahakian, B., Jones, G., Levy, R., Gray, J., Warburton, D., 1989. The effects of
nicotine on attention, information processing, and short-term memory in pa-
tients with dementia of the Alzheimer type. Br. J. Psychiatry. 154, 797e800.
Sawaguchi, T., Goldman-Rakic, P.S., 1991. D1 dopamine receptors in prefron-
tal cortex: involvement in working memory. Science 251, 947e950.
Schilstrom, B., de Villiers, S., Malmerfelt, A., Svensson, T.H., Nomikos, G.G.,
2000. Nicotine-induced Fos expression in the nucleus accumbens and the
medial prefrontal cortex of the rat: role of nicotinic and NMDA receptors
in the ventral tegmental area. Synapse 36, 314e321.
Semenova, S., Bespalov, A., Markou, A., 2003. Decrease prepulse inhibition
during nicotine withdrawal in DBA/2J mice is reversed by nicotine self-
administration. Eur. J. Pharmacol. 472, 99e110.
Shim, I., Javaid, J.I., Wirtshafter, D., Jang, S.Y., Shin, K.H., Lee, H.J., Chung, Y.C.,
Chun, B.G., 2001. Nicotine-induced behavioral sensitization is associated
with extracellular dopamine release and expression of c-Fos in the striatum
and nucleus accumbens of the rat. Behav. Brain Res. 121, 137e147.
Shoaib, M., Gommans, J., Morley, A., Stolerman, I.P., Grailhe, R.,
Changeux, J.-P., 2002. The role of nicotinic receptor beta-2 subunits in
nicotine discrimination and conditioned taste aversion. Neuropharmacology
42, 530e539.
Skjei, K.L., Markou, A., 2003. Effects of repeated withdrawal episodes,
nicotine dose, and duration of nicotine exposure on the severity and dura-
tion of nicotine withdrawal in rats. Psychopharmacology 168, 280e292.
Smith-Roe, S.L., Kelley, A.E., 2000. Coincident activation of NMDA and
dopamine D1 receptors within the nucleus accumbens core is required
for appetitive instrumental learning. J. Neurosci. 20, 7737e7742.
Stolerman, I.P., Naylor, C., Elmer, G.I., Goldberg, S.R., 1999. Discrimination
and self-administration of nicotine by inbred strains of mice. Psychophar-
macology 141, 297e306.

1040 V. David et al. / Neuropharmacology 50 (2006) 1030e1040
Tizabi, Y., Copeland Jr., R.L., Louis, V.A., Taylor, R.E., 2002. Effects of
combined systemic alcohol and central nicotine administration into ventral
tegmental area on dopamine release in the nucleus accumbens. Alcohol
Clin. Exp. Res. 26, 394e399.
de Villiers, S.H., Lindblom, N., Kalayanov, G., Gordon, S., Malmerfelt, A.,
Johansson, A.M., Svensson, T.H., 2002. Active immunization against
nicotine suppresses nicotine-induced dopamine release in the rat nucleus
accumbens shell. Respiration 69, 247e253.
Villegier, A.S., Salomon, L., Granon, S., Changeux, J.P., Belluzzi, J.D., Leslie,
F.M., Tassin, J.P. Monoamine oxidase inhibitors allow locomotor and
rewarding responses to nicotine. Neuropsychopharmacology, in press
[Epub ahead of print].





























