Embed Size (px)
Citation preview

J. Basic Microbiol. 37 (1997) 6,451 4 6 3
(Department of Molecular Biology, University of Gdansk, Kladki 24, 80-822 Gdahk, Poland)
Replication of plasmids derived from P1, F, R1, R6K and RK2 replicons in amino acid-starved Escherichia coli stringent and relaxed strains BORYS WROBEL and GRZEGORZ WQGRZYN
(Received 24 March 1997lAccepted 16 July 1997)
Replication of mini-plasmids derived from bacteriophage P1 and naturally existing plasmids F, R1, R6K and RK2 in otherwise isogenic relA+ and reLk Escherichia coli strains during amino acid star- vation and limitation was investigated. Since it was previously demonstrated that inhibition of DNA synthesis or amplification of plasmid DNA may depend on the nature of deprived amino acid, we starved bacteria for five different amino acids. We found differential replication of all these plasmids but RK2 (which did not replicate at all in amino acid-starved bacteria) during the stringent and re- laxed response. While in almost all cases plasmid DNA replication was inhibited during the stringent response irrespective of the nature of deprived amino acid, wild-type or copy-up mini-P1, mini-F and mini-R1 plasmids replicated in relA- bacteria depending on the kind of starvation. R6K-derived plasmids harbouring ori /3 and y (but not those containing ori a, /3 and y or only ori y ) were able to replicate in relA- bacteria starved for all tested amino acids. Possible explanations for the mechanisms of regulation of replication of plasmids derived from P1, F, R1, R6K and RK2 during amino acid starvation are discussed. Our results also indicate that, like in the case of some other replicons, ap- propriate amino acid starvation or limitation may be used as a method for efficient amplification of plasmids derived from P1, F, R1 and R6K.
Although in laboratories microorganisms are usually cultivated in optimal growth condi- tions (rich media, good aeration, optimal temperature), in their natural environment, bacte- ria (like all other organisms) are almost permanently endangered by various stresses. One of the biggest problem for bacterial cells living in natural environment is nutritional limitation. In fact, for the most of time these cells are starved (for a review see MATIN 1991). There- fore, organisms had to develop cellular mechanisms which might allow them to survive starvation conditions. Among them, one of the best known is the stringent response (or stringent control). This is the bacterial response to amino acid starvation. The stringent control has been found to exist in many bacterial species but it is best characterised in Escherichia coli (for recent review see CASHEL et aZ. 1996). The first event of the stringent response is production of high amounts of guanosine-S-diphosphate3'-diphosphate (ppGpp). This nucleotide interacts with RNA polymerase (REDDY et al. 1995) resulting in inhibition of transcription of many genes (genes coding for rRNAs and tRNAs are repressed espe- cially strongly) and activation of some promoters. There are two enzymes capable of syn- thesising ppGpp: ppGpp synthetase I, the reZA gene product, and ppGpp synthetase 11, the spoT gene product (HERNANDEZ and BREMER 1991, XIAO et a f . 1991; GENTRY and CASHEL 1996, MURRAY and BREMER 1996). The RelA protein, which is bound to a ribo- some, is activated upon amino acid starvation due to appearance of uncharged tRNA mole- cules, their interaction with codons at the ribosomal A site and so called ribosome idling. On the other hand, the SpoT protein is responsible for ppGpp synthesis during carbon source limitation. The spoT gene product is also the main enzyme which degrades ppGpp; thus it is a bifunctional protein capable of both ppGpp synthesis and hydrolysis (GENTRY
30*

452 B. WR6BEL and G. WQGRZYN
and CASHEL 1996). Mutants in the relA gene do not produce ppGpp during amino acid starvation as they do not contain functional ppGpp synthetase I. Moreover, ppGpp syn- thetase I1 is inactive in these conditions. Therefore, in amino acid-starved relA mutants the level of ppGpp not only does not increase but even decreases. This phenomenon is called the relaxed response (for a review see CASHEL et al. 1996). The existence of the stringent response is very important for bacteria as relA mutants are much less viable upon prolonged starvation relative to wild-type strains (HECKER et al. 1986).
The stringent response is a pleiotropic phenomenon as most of cellular processes are changed in amino acid starved bacteria (CASHEL et al., 1996). This concerns also DNA replication. Despite the fact that DNA replication is one of the fundamental processes in living cells, there are only few examples of replicons investigated in stringent and relaxed conditions. Until now, DNA replication in amino acid-starved E. coli cells was reported only for ColE1, oriC, 1, pSClOl and RK2 replicons (HECKER et al. 1983, LIN-CHAO and BREMER 1986, GUZMAN et al. 1988, 1991, WEjGRZYN et al. 1991a, b, 1995, 1996a, W E p RZYN and TAYLOR, 1992, HERMAN et al. 1994a, b, c, SZALEWSKA-PAtASZ et al. 1994, SZALEWSKA-PALASZ and WE~GRZYN 1994, 1995, HERMAN and WEGRZYN, 1995; SCHREI- BER et al. 1995, WQGRZYN and WQGRZYN 1995). On the other hand, results obtained dur- ing these investigations had two important general implications. First, these reports allowed to understand much better the mechanisms of regulation of replication of ColEl plasmids, plasmids derived from bacteriophage 1, and E. coli chromosome not only in amino acid- starved cells but also under normal growth conditions. Analogous investigations on the stringent control of Bacillus subtilis chromosome replication led to discovery of an inter- esting general mechanism of replication regulation (SEROR et al. 1986, LEVINE et al. 1991). Secondly, appropriate amino acid starvation has been reported as an efficient method for in vivo DNA amplification in the case of ColE1, 1, and pSClOl replicons (HECKER et al., 1985, 1988, SCHROETER et al., 1988, RIETHDORF et al. 1989, HOFMANN et al. 1990, HER- MAN et al. 1994a, b, c, WEGRZYN 1995, NEUBAUER et al. 1996). Therefore, we decided to investigate replication of other replicons during stringent and relaxed response in E. coli. We used replicons derived from bacteriophage P1 and plasmids F, R1, and R6K as they are models in studies on regulation of DNA replication. Since previously reported findings indicated that replication of at least some plasmids may depend on the nature of deprived amino acid, we investigated DNA replication in bacteria starved for five different amino acids. Apart from P1, F, R1 and R6K replicons we investigated also the RK2-derived plas- mid as its replication was previously studied only during amino acid limitation and in cells starved for isoleucine but not for other amino acids.
Materials and methods
Bacterial strains: E. coli relA+ strain CW8 (leu, arg, thr, his, thi) and its otherwise isogenic relA- derivative CP79 (FIIL and FRIESEN 1968) were used. Strain BM80 (relA-) was constructed by P1 transduction using CF1652 (AreL4::kan) (XIAO et al. 1991) as a donor and CP78 as a recipient, and was provided by Michael Cashel.
Plasmids: Plasmids used in this work are listed and characterised in Table 1.
Culture media: Bacteria were cultivated at 37 "C in minimal media 1 and 2, described previously by WQGRZYN et al. (1991 b). These media consist of salts, glucose (l%), thiamine (10 pg/ml) and either Casamino acids (1%) (Medium 1) or appropriate L-amino acids (50 pg/ml each) (Medium 2). Amino acid starvation and limitation: Isoleucine starvation was provoked by addition of L-valine to the medium 2 up to 1 mg/ml as described previously (W~GRZYN et al. 1991a, b). Starvation of auxotrophic bacteria for threonine, arginine, leucine and histidine was achieved by removing of appropriate amino acid from the medium 2. This was performed by centrifugation of the culture, washing the bacterial pellet and resuspending it in the same medium but lacking one of amino acids

Stringent control of plasmid DNA replication 453
Table 1 Plasmids
Plasmid Type of replicon ori region Relevant Copy numbeda) Reference
pAX274 mini-P oriR wild-type 1-2 OGURA et al. (1990) pSP102 mini-P oriR AincA 8 PAL et al. (1986) pMF3 mini-F ori2 wild-type 1-2 MANIS and KLINE (1977) pKV5110 mini-F ori2 AincC 5-10 KAWASAKI et al. (1991) pKN500 mini-R1 oriRl wild-type 1-2 MOLIN et al. (1979) pKN177 mini-Rl oriRl copA 6-10 DIAZ and STAUDEN-
pR6K mini-R6K ori cz p y wild-type 15-20 KOTLER and HELINSKI
pMF26 mini-R6K o r i a y Aoria 15-20 FILUTOWICZ et al. (1985)
pTJS42 mini-RK2
(a) The values represent average plasmid copy number in exponentially growing wild-type E. coli
genotype
BAUER (1982)
(1978)
mini-R6K ori y A o r i a p 15-20 WU et al. (1992)
HELINSKI (1991) oriV wild-type 4 4 LEWIS-KIITELL and
PFW8
cells.
as described previously (HERMAN et al. 1994~). In this paper, the term “amino acid limitation” is used to describe conditions in which concentration of particular amino acid in the medium is low enough to be a limiting factor for bacterial growth (causing an inhibition of growth) but the medium is not com- pletely devoid of this amino acid (as during amino acid starvation), according to the definition pro- vided elsewhere (WROBEL and WQGRZYN 1997). Amino acid limitation was achieved by cultivating bacteria in medium 1 to the stationary phase of growth. At the beginning of this growth phase amino acid limitation starts due to exhaustion or arginine (HOFMANN et al. 1990). This method has been used for similar approach previously (HOFMA” et al. 1990, HERMAN et al. 1994c, W~GRZYN 1995). Measurement of plasmid content: Samples of bacterial culture were withdrawn at indicated times. During investigation of different plasmids, these samples corresponded to 1, 2, or 4 OD,, unit(s) depending on the initial plasmid copy number in cells. Plasmid content in bacterial cells was esti- mated by DNA isolation using alkaline lysis (SAMBROOK et al. 1989), linearization with appropriate restriction endonuclease, agarose gel electrophoresis, staining with ethidium bromide and densitomet- ric analysis of plasmid bands as described previously (HERMAN et al. 1994b, c).
Calculation of the amplification factor: Amplification factor is a value which can be used for esti- mation of plasmid DNA synthesis in amino acid-starved bacteria (W~GZRYN et al. 1991 a, WEGZRYN 1995). This factor was calculated as the ratio of plasmid content per bacterial mass at indicated times after the onset of starvation (in the case of amino acid-starved cultures) or 6 and 18 hours after reaching the OD, value of 0.2 by bacterial culture (in the case of amino acid limitation) to the plas- mid content per % a c t e d mass in the exponentially growing (unstarved) bacteria, according to WQGRZYN et al. (199 1 a) and W~GRZYN (1 995).
Results
Experimental systems
To investigate plasmid DNA replication during stringent and relaxed response, bacteria were grown in minimal medium 2 and amino acid starvation was induced. The samples were withdrawn from the culture of starved bacteria as well as from the culture of unstarved bacteria. Plasmid DNA was isolated and relative plasmid content was estimated. Then, amplification factor was calculated as the ratio of plasmid content per bacterial mass at indicated times after the onset of starvation to the plasmid content per bacterial mass in the
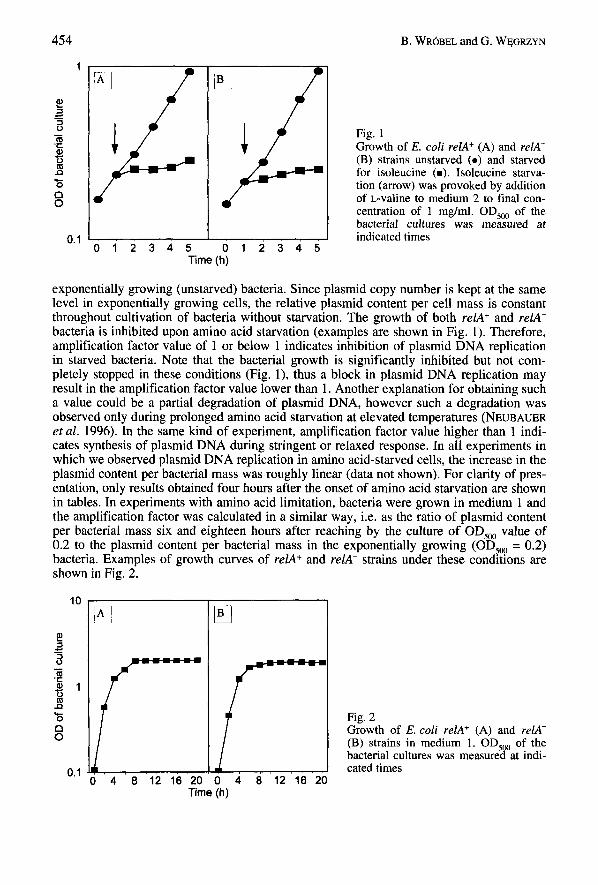
B. WR6BEL and G. W ~ G R Z Y N 454
1
* 5 - a $ 0 n 0 0 0
0 m - .- c
Ic
0.1
El f
Fig. 1 Growth of E. coli relA+ (A) and rel.4- (B) strains unstarved ( 0 ) and starved for isoleucine (m). Isoleucine starva- tion (arrow) was provoked by addition of L-valine to medium 2 to final con- centration of 1 mg/ml. OD,, of the bacterial cultures was measured at indicated times
0 1 2 3 4 5 0 1 2 3 4 5 Time (h)
exponentially growing (unstarved) bacteria. Since plasmid copy number is kept at the same level in exponentially growing cells, the relative plasmid content per cell mass is constant throughout cultivation of bacteria without starvation. The growth of both relA+ and relA- bacteria is inhibited upon amino acid starvation (examples are shown in Fig. 1). Therefore, amplification factor value of 1 or below 1 indicates inhibition of plasmid DNA replication in starved bacteria. Note that the bacterial growth is significantly inhibited but not com- pletely stopped in these conditions (Fig. l), thus a block in plasmid DNA replication may result in the amplification factor value lower than 1. Another explanation for obtaining such a value could be a partial degradation of plasmid DNA, however such a degradation was observed only during prolonged amino acid starvation at elevated temperatures (NEUBAUER et al. 1996). In the same kind of experiment, amplification factor value higher than 1 indi- cates synthesis of plasmid DNA during stringent or relaxed response. In all experiments in which we observed plasmid DNA replication in amino acid-starved cells, the increase in the plasmid content per bacterial mass was roughly linear (data not shown). For clarity of pres- entation, only results obtained four hours after the onset of amino acid starvation are shown in tables. In experiments with amino acid limitation, bacteria were grown in medium 1 and the amplification factor was calculated in a similar way, i.e. as the ratio of plasmid content per bacterial mass six and eighteen hours after reaching by the culture of OD,,, value of 0.2 to the plasmid content per bacterial mass in the exponentially growing (OD,, = 0.2) bacteria. Examples of growth curves of relA+ and relA- strains under these conditions are shown in Fig. 2.
0.1 - 0 4 8 12 16 20
U
Fig. 2 Growth of E. coli relA+ (A) and relA- (B) strains in medium 1. OD, of the bacterial cultures was measurerat indi- cated times
0 4 8 12 16 20 Time (h)
Stringent control of plasmid DNA replication 455
Table 2 Replication of the wild-type (pAX274) and copy-up (pSP102) mini-P1 plasmids in amino acid starved, otherwise isogenic, relA+ and relA- strains
Deprived Amplification factor(a) amino acid
pAX274 (wild-type mini-P1)
relA+ relA- relA+ relA-
pSP102 (copy-up mini-P1)
Isoleucine 1 .1 0.9 2.3 0.7 Threonine 0.9 0.8 1.4 0.8 Arginine 0.8 2.1 1 . 1 0.9 Leucine 1.2 3.8 1.2 1.6 Histidine 0.9 0.8 0.9 0.7
(a) The amplification factor values presented in the table were calculated four hours after the onset of amino acid starvation. The data are average values from three measurements; in each case the standard deviation was not higher than 0.3.
Plasmid DNA replication in amino acid starved relA+ and relA- strains
It was previously reported that starvation for different amino acids may have different ef- fects on plasmid DNA replication at least in the case of some replicons (HERMAN et al. 1994~). Therefore, we investigated replication of plasmids in otherwise isogenic relA+ (CP78) and reU- (CP79 and BM80) strains starved for different amino acids, namely: iso- leucine, threonine, arginine, leucine and histidine.
Mini-PI replicons are plasmids derived from bacteriophage PI. The wild-type P1 plas- mids are maintained in E. coli cells in a copy number of 1-2, similarly to P1 prophages (for a recent review see CHATTORAJ 1995). We found that replication of a wild-type miniP1 plasmid is inhibited during the stringent response (Table 2). The replication proceeds in the relA bacteria starved for arginine and leucine, but not if the bacteria are starved for iso- leucine, threonine and histidine (Table 2). We investigated also replication of the copy-up mutant of mini-P1 plasmid, pSP102. This plasmid is devoid of the incA locus which acts as a negative regulator of P1 replication by sequestering the RepA protein, an initiator of replication. As a consequence, copy number of pSP102 in exponentially growing E. coli cells increases up to 8 in comparison to 1-2 copies found in the case of a wild-type P1 mini-plasmid (PAL et al. 1986). We found that replication of pSP102 was inhibited in al- most all experiments in both relA' and relA- strains starved for different amino acids. The only exception was isoleucine starvation of the relA+ bacteria where some replication was observed, though the amplification factor value of about 2 indicates that only one round of plasmid replication could occur after onset of isoleucine starvation (Table 2).
The mini-F replicons derive from a conjugative plasmid F (called also the F sex factor). F plasmids are of low copy number, 1-2 per cell. We found that replication of the wild-type mini-F plasmid is generaly inhibited during both stringent and relaxed response irrespective of the nature of deprived amino acid but one exception, leucine-starved relA' cells (Table 3) . Plasmid pKV5110 is a derivative of mini-F lacking the incC region. This region contains sequences similar to those found at ori2 (origin of F plasmid replication) and exhibits affin- ity to the initiator protein RepE thus acting as a negative regulator of replication initiation. Plasmids lacking incC (like pKV5llO) have increased copy number in E. coli up to 5-10 copies per cell (KAWASAKI et al. 1991). We found that plasmid pKV5110 does not replicate during the stringent response (Table 3). However, contrary to the wild-type mini-F, the copy-up derivative of mini-F replicon replicates in the relA- mutant starved for arginine, leucine and histidine (Table 3).
456 B. WR6BEL and G. W~GRZYN
Table 3 Replication of the wild-type (pMF3) and copy-up (pKV5 110) mini-F plasmids in amino acid starved, otherwise isogenic, relA+ and relA- strains
Deprived amino acid
Isoleucine Threonine Arginine Leucine Histidine
Amplification factor@)
pMF3 (wild-type mini-F)
relA+ relA- relA+ r e K
1.2 0.6 0.9 1.1 1.3 0.9 1.2 1.6 1.6 0.4 1.4 4.2 6.5 0.3 1.3 3.9 1.3 0.5 1.1 4.4
pKV5 1 10 (copy-up mini-F)
(a) The amplification factor values presented in the table were calculated four hours after the onset of amino acid starvation. The data are average values from three measurements; in each case the standard deviation was not higher than 0.4.
Plasmid R1 is a large (95.8 kb) conjugative plasmid. Mini-R1 plasmids, like the parental plasmid, are of low copy number (1 -2) in E. coli. The results presented in Table 4 indicate that replication of the mini-R1 plasmid (pKN500) is inhibited during the stringent response in all cases but isoleucine starvation. Some pKN500 replication (about one round) was observed in the reZA- mutant starved for threonine and leucine but not in the same bacteria starved for other amino acids (Table4). The copy number of another mini-R1 plasmid, pKN177, is increased about six times relative to the wild-type plasmid due to a single base change in the loop of the antisense RNA coded in the copA locus (DIAZ and STAUDEN- BAUER, 1982; G . WAGNER, personal communication). The decrease in the negative regula- tion of replication by copA RNA results in higher copy number of pKN177. Replication of this plasmid is under stringent control as it is inhibited in amino acid-starved relA+ bacteria and proceeds during starvation of the reZk for most amino acids (Table 4).
Plasmid R6K is a 38 kb resistance transfer plasmid (carrying ampicillin and streptomycin resistance genes) that occurs naturally in E. coli in 15-20 copies per cell (KONTOMICHALOU et al. 1970). This plasmid contains three origins of replication, named a, B and y. Replica- tion from each origin requires the R6K-encoded JC protein and a DNA segment containing seven 22 bp direct repeats. Moreover, each origin requires different cis-acting regions for
Table 4 Replication of the wild-type (pKN500) and copy-up (pKN177) mini-R1 plasmids in amino acid starved, otherwise isogenic, relA+ and relA- strains
Deprived Amplification factor@) amino acid
pKNS00 (wild-type mini-R1)
relA+ relk relA+ relA- pKN177 (copy-up mini-Rl)
Isoleucine 4.0 0.5 0.8 2.8 Threonine 0.5 2.2 1 .o 3.9
Leucine 0.4 2.3 1.1 2.7 Histidine 0.6 0.6 0.9 2.6
Arginine 0.5 1.5 0.9 1 .o
(a) The amplification factor values presented in the table were calculated four hours after the onset of amino acid starvation. The data are average values from three measurements; in each case the standard deviation was not higher than 0.2.
Stringent control of plasmid DNA replication 457
Table 5 Replication of the mini-R6K plasmids containing ori a, B and y (pR6K), ori #? and y (pMF26) or only ori y (pFW8) in amino acid starved, otherwise isogenic, relA+ and relA- strains
Deprived Amplification factor(") amino acid
pR6K (ori a B y )
relA+ re& relA+ relA- relA+ relA- pMF26 (ori B y ) pFW8 (ori y )
Isoleucine 0.8 0.6 0.7 2.6 0.8 0.4 Threonine 0.9 1.6 0.6 2.3 0.8 0.5 Arginine 1.1 0.9 0.5 2.1 0.9 0.4 Leucine 1.2 1.7 0.7 3.7 0.8 0.4 Histidine 1.1 2.0 0.5 6.5 1.1 0.4
(a) The amplification factor values presented in the table were calculated four hours after the onset of amino acid starvation. The data are average values from three measurements; in each case the standard deviation was not higher than 0.2.
activity. Replication of mini-R6K replicons containing all three origins (pR6K), only /3 and y origins (pMF26) and only y origin (pFW8) was investigated during stringent and relaxed response. We found that replication of all these plasmids was stopped in amino acid-starved relA+ strain irrespective of the nature of deprived amino acid (Table 5). However, important differences were observed during the relaxed response as only the plasmid carrying p and y origins (pMF26) was able to replicate in relA- bacteria starved for all tested amino acids (Table 5).
Plasmid RK2 is a 60 kb broad-host-range plasmid that replicates at a copy number of four to seven in E. coli. Although it was previously reported that a derivative of this plasmid (pLAFRx) cannot replicate during both stringent and relaxed responses (HERMAN et al., 1994c), we decided to test another mini-RK2 plasmid (pTJS42) as the previous report con- cerned only amino acid limitation and starvation for isoleucine. We confirmed previously published results and demonstrated that inability of RK2 to replicate during the stringent and relaxed response concerns also starvation for other amino acids (Table 6).
Plasmid DNA replication in relA+ and relA- strains during amino acid limitation Previous reports demonstrated that amino acid limitation, which starts at the time of enter- ing of a culture of auxotrophic bacteria into stationary phase of growth, may result in more significant differences in plasmid DNA replication between stringent and relaxed strains
Table 6 Replication of the mini-RK2 plasmid (pTJS42) in amino acid starved, otherwise isogenic, relA+ and relA- strains
Deprived amino acid Amplification factor(")
Isoleucine Threonine Ar g i n i n e Leucine Histidine
1.4 0.9 1 .o 1 .o 0.8
1 .o 0.9 0.9 1 .o 0.6
(a) The amplification factor values presented in the table were calculated four hours after the onset of amino acid starvation. The data are average values from three measurements; in each case the standard deviation was not higher than 0.2.
45 8 B. WR6BEL and G. WQGRZYN
Table 7 Replication of different plasmids in otherwise isogenic relA+ and relA- strains during amino acid limitation.
Plasmid Type of ori region Relevant Amplification factor") replicon genotype
relA+ relA-
6h 18h 6 h 18h pAX274 mini-P pSP102 mini-P pMF3 mini-F pKV5llO mini-F pKN5OO mini-Rl pKN177 mini-R1 pR6K mini-R6K pMF26 minLR6K pFW8 mini-R6K
oriR wild-type oriR AincA ori2 wild-type ori2 AincC oriRl wild-type oriRl copA ori a p y wild-type or ip y Aor ia o r i y A o r i a p
1 .o 0.9 0.9 0.7 0.6 0.8 1.5 0.3 2.0 1.9 1 .o 0.9 1.3 0.8 1.2 1 .o 2.3 0.6 2.4 1.2 5.8 4.2 1.6 0.9 1.8 1.7 2.5 5.4 1.9 1 .o 0.9 0.9 1.4 0.8 1.2 0.8
(a) Samples for calculation of the amplification factor were withdrawn from the bacterial culture 6 and 18 hours after reaching the OD,, value of 0.2. The data are average values from three meas- urements; in each case the standard deviation was not higher than 0.4.
than starvation for a particular amino acid (HERMAN et al. 1994c, WQGRZYN 1995). There- fore, we investigated replication of all above described plasmids but the RK2 replicon (which was shown previously not to replicate under these conditions, HERMAN et al. 1994c) during amino acid limitation in relA' and relA- strains. Plasmid DNA content was estimated from samples withdrawn from the bacterial culture 6 and 18 hours after reaching the OD,, value of 0.2. The amplification factor was calculated relative to the plasmid content in the sample withdrawn from exponentially growing culture (OD,,, of 0.2). The results summa- rized in Table 7 indicate that intensive replication of DNA during amino acid limitation occurs only in the case of mini-R1 copy-up plasmid (pKN177) in the reZA+ strain and mini- R6K plasmid containing all three origins in relA- bacteria. In several other cases we ob- served some plasmid replication but this process was limited as the value of the amplifica- tion factor was about 2.
Discussion
Although the stringent response provokes changes in whole cell physiology, until recently reports on the regulation of DNA replication, one of fundamental cellular processes, in amino acid-starved bacteria were very rare. On the other hand, recent studies demonstrated that results obtained during investigation of the stringent and relaxed control of DNA repli- cation led to important general conclusions. For example, studies on the regulation of repli- cation of plasmids derived from bacteriophage 1 in amino acid-starved E. coli cells resulted in discovery of the phenomenon called protein inheritance (DNA replication carried out by the heritable replication complex) which has been subsequently demonstrated to occur also in normal growth conditions (WQGFUYN et al. 1996b). Furthermore, investigation of repli- cation of Bacillus subtilis chromosome during stringent and relaxed response resulted in discovery of the interesting phenomenon of premature replication termination, the mecha- nism which seems to have an important regulatory role (SEROR et al. 1986; LEVINE et al. 1991). A role for uncharged tRNA molecules in regulation of replication of ColE1 plasmids has been suggested also on the basis of investigation of the stringent control of DNA repli-

Stringent control of plasmid DNA replication 459
cation (YAVACHEV and IVANOV 1988, HERMAN et al. 1994a, b, c). Since until now DNA replication in amino acid-starved E. coli cells was investigated only for ColE1, oriC, 1 and RK2 replicons, we studied replication of other well-defined model replicons during strin- gent and relaxed response.
Replication of mini-P1 plasmids is generally inhibited during amino acid starvation. How- ever we found quite intensive replication of the wild-type miniP1 plasmid in leucine-starved relA- strain and some replication of the wild-type mini-Pl plasmid in arginine-starved reh- mutant and of copy-up mini-P1 plasmid in isoleucine-starved reZA' bacteria. It is all the more intriguing that both plasmids do not replicate in reL4' and reM- strains during amino acid limitation. Differential replication of ColEl and pSC101-derived plasmids upon starvation for different amino acids has been reported previously (HECKER et al. 1983, LIN-CHAO and BREMER 1986; HERMAN et al. 1994b, c). The mechanism of the stringent control of plasmid pBR322 (ColEl-type replicon) has been proposed (HERMAN et al. 1994b), however, despite interesting speculations, the molecular mechanism of the regulation of pSClOl replicaton in amino acid-starved bacteria remains to be elucidated. The same concerns P1-derived replicons.
With one exception, both wild-type and copy-up mini-F plasmids do not replicate in amino acid-starved relA+ bacteria. The exception is amplification of the wild-type plas- mid in leucine-starved relA' strain. The mechanism of this phenomenon remains unclear. Inhibition of wild-type mini-F plasmid replication was observed during the relaxed response, however the copy-up derivative replicates in the relA- mutant starved for arginine, leucine and histidine. What is a possible mechanism of this phenomenon? The copy- up plasmid (pKV511O) is devoid of the incC region which exhibits a modest affinity to the replication initiator RepE (KAWASAKI et al. 1991). Titration of the RepE protein by incC results in inhibition of plasmid DNA replication. Amino acid starvation causes drastically decreased rate of total protein synthesis, but some translation is possible, mainly due to amino acids which come from intracellular protein degradation. If any RepE syn- thesis occurs in cells harbouring wild-type mini-F plasmid, the low amount of the initiator protein could be titrated out by the incC region resulting in inhibition of replication initia- tion. This would not be a case if incC is deleted, like in copy-up plasmid pKV5 110, and RepE could interact efficiently with the ori2 sequence. Since in amino acid-starved relA' bacteria high levels of ppGpp are produced, transcription of most genes is inhibited. Thus, it is possible that synthesis of low amounts of the RepE protein during amino acid starvation, and thus replication of mini-F plasmid devoid of in&, is possible only in relA- mutants (the exception is leucine starvation of the relA+ bacteria but currently no pu- tative mechanism of this phenomenon can be proposed). If this hypothesis were true, one should assume higher rate of RepE synthesis in reL4- cells starved for arginine, leu- cine or histidine (as under these conditions we observed significant pKV5 1 10 replication) than in the same cells starved for isoleucine and threonine (as under these conditions replication of pKV5110 was negligible). In fact, synthesis of a protein in cells starved for some amino acids in contrast to starvation for other amino acids has recently been reported for another replication protein, the bacteriophage 1 P gene product. This pro- tein was found to be efficiently produced in bacteria starved for isoleucine, threonine and histidine but not during arginine and leucine starvation (OBUCHOWSKI and WQGRZYN 1996).
In almost all cases, replication of wild-type as well as copy-up mini-R1 plasmids was in- hibited during the stringent response. The only exception was found for the wild-type plas- mid in isoleucine-starved cells. On the other hand, in most cases we observed at least some replication of both plasmids (though generally more effective for the copy-up mutant) in amino acid-staved relA- mutant. The copy-up phenotype of plasmid pKN177 results from a single base change in the loop of the antisense copA RNA, a negative regulator of the ex- pression of the repA gene coding for a replication initiator (DIM and STAUDENBAUER 1982, G. WAGNER, personal communication). It seems that interaction of the copA RNA with

460 B. WR6BEL and G. WEGRZYN
repA mRNA together with effects of ppGpp on transcription of both these genes may play a crucial role in the stringent control of R1 plasmid replication. Since it appears that R1 repli- cation is dependent on de novo protein synthesis (NORDSTROM et al. 1984) it is worth not- ing that the replication of both wild-type and copy-up mini-R1 plasmids was observed dur- ing amino acid limitation, though the highest amplification factor were calculated for the copy-up mutant (pKN177) in relA' strain.
Replication of R6K-derived replicons containing either all three origins (a, /3 and y), p and y origins or only y origin was investigated during the stringent and relaxed response. We found that replication of all three plasmids was inhibited in amino acid-starved relA+ bacteria irrespective of the kind of deprived amino acid. During the relaxed response, repli- cation proceeded only in the case of plasmid harbouring /3 and y origins. These results may suggest that in amino acid-starved relA- mutant replication of mini-R6K plasmid starts from the ori /? and that presence of ori a impairs this replication initiation. Such an interpretation is in agreement with the observation that during the relaxed response some replication of the plasmid harbouring all three origins occurs in bacteria starved for leucine and histidine, i.e. under conditions giving the highest amplification factor for the plasmid bearing /3 and y origins.
It was previously reported that RK2-derived plasmid did not replicate in relA' and reZA- strains starved for isoleucine as well as during amino acid limitation (HERMAN etal. 1994~). We confirmed that this is also true for the mini-RK2 plasmid pTJS42. Moreover, we demonstrated that replication of this plasmid is halted in both stringent and relaxed strains starved for arginine, threonine, leucine and histidine. As plasmids derived from RK2 replicate in unstarved cells overexpressing the relA gene (HERMAN and WQGRZYN, 1995), it seems that inhibition of its replication during the stringent and relaxed response is due to significantly decreased protein synthesis in amino acid-starved cells rather than to ppGpp action.
Apart from providing basic information about the mechanisms of regulation of DNA rep- lication, studies on replication of plasmids during stringent and relaxed response led to development of a new method for DNA amplification in vivo (HECKER et al. 1985). It was found that appropriate amino acid starvation may be used as a mean of achieving effective amplification of plasmids derived from ColE1, pSClOl and bacteriophage Iz (HECKER et al., 1985, 1988; SCHROETER et al. 1988, RIETHDORF et al. 1989, HOFMANN et al. 1990; HER- MAN et al. 1994a, b, c, WQGRZYN, 1995, NEUBAUER et al. 1996). Our results indicate that using this method it is possible to achieve significant amplification of plasmids derived from P1, F, R1 and R6K replicons. This concerns in particular: wild-type mini-P1 plasmid in relA- bacteria starved for leucine, wild-type mini-F plasmid in relA+ bacteria starved for leucine, copy-up (incC-) mini-F plasmid in reZA- bacteria starved for arginine, leucine or histidine, wild-type mini-R1 plasmid in relA' bacteria starved for isoleucine, copy-up (copA mutant) mini-R1 plasmid in relA' bacteria during amino acid limitation or in relA- bacteria starved for threonine, R6K-derived plasmid harbouring ori a, /3 and y in relA- bacteria during amino acid limitation, and R6K-derived plasmid harbouring ori /3 and y in relA- bacteria starved for leucine or histidine.
Acknowledgements
We are very grateful to following researchers for providing plasmids and bacteria: ROLF BERNANDER for plasmids pKN177 and pKN500, MICHEAL CASHEL for strain BM80, DHRUBA CHAITORAJ for plasmids pMF3 and pSP102, MARCIN FILUTOWICZ for plasmids pR6K, pMF26 and pFW8, IGOR KONIECZNY for plasmid pTJS42, and CHIEKO WADA for plasmids pAX274 and pKV5 1 10. This work was supported by the University of Gdahsk (DS/ll90-4-0114-7), Medical University of Gdahsk (grant A-20) and Ministry of National Education.

Stringent control of plasmid DNA replication
References
46 1
CASHEL, M., GENTRY, D. R., HERNANDEZ, V. J. and VINELLA, D., 1996. The stringent response. In: Escherichiu coli and Salmonella: Cellular and Molecular Biology (F. C. NEIDHARDT, R. CURTISS 111, J. L. INGRAHAM, E. C. C. LIN, K. B. Low, B. MAGASANIK, W. S. REZNIKOFF, M. RILEY, M. SCHAECHTER and H. E. UMBARGER, Editors), pp. 1458-1496. American Society for Micro- biology, Washington, DC.
CHAITORAJ, D. K., 1995. Role of molecular chaperones in initiation of plasmid DNA replication, In: Genetic Engineering (J. K. SETLOW, Editor), 17, 81 -98. Plenum Press, NY.
DIM, R. and STAUDENBAUER, W. L., 1982. Origin and direction of mini-R1 plasmid DNA replication in cell extracts of Escherichiu coli. J. Bacteriol.. 150. 1077-1084.
FIIL, N., and FRIESEN, J. D., 1968. Isolation of relaxed’mutants of Escherichia coli. J. Bacteriol,, 95, 729-73 1.
m m w c z , M., UHLENHOPP, E. and HELINsKI, D. R., 1985. Binding of purified wild-type and mutant n initation proteins to a replication origin region of plasmid R6K. J. Mol. Biol., 187,225-239.
GENTRY, D. R. and CASHEL, M., 1996. Mutational analysis of the Escherichia coli spoT gene identifies distinct but overlapping regions involved in ppGpp synthesis and degradation. Mol. Microbiol., 19,
GUZMAN, E. C., CARRILLO, F. J. and JIMENEZ-SANCHEZ, A., 1988. Differential inhibition of the initiation of DNA replication in stringent and relaxed strains of Escherichiu coZi. Genet. Res., 51, 173-177.
HECKER, M., SCHROETER, A. and MACH, F., 1983. Replication of pBR322 DNA in stringent and relaxed strains of Escherichia coli. Mol. Gen. Genet., 190, 355-357.
HECKER, M., SCHROETER, A. and MACH, F., 1985. Escherichia coli relA strains as hosts for amplifi- cation od pBR322 plasmid DNA. FEMS Microbiol. Lett., 29, 331 -334.
HECKER, M., SCHROETER, A., TRADER, K. and MACH, F., 1986. Role of relA mutation in the survival of amino acid-starved Escherichia coli. Arch. Microbiol., 143,400402.
HECKER, M., RIETHDORF, S., BAUER, C., SCHROETER, A. and BORRISS, R. (1988): Expression of a cloned B-glucanase gene from Bacillus umyloliquefuciens in an Escherichiu coli relA strain after plasmid amplification. Mol. Gen. Genet., 215, 181 -183.
HERMAN, A. and WQGRZYN, G., 1995. Effect of increased ppGpp concentration on DNA replication of different replicons in Escherichia coli. J. Basic Microbiol., 35, 33-39.
HERMAN, A., WEGRZYN, A. and WQGRZYN, G., 1994a. Combined effect of stringent and relaxed response, temperature and rom function on the replication of pUC plasmids in Escherichiu coli. Acta Biochim. Polon., 41, 122-124.
HERMAN, A., WQGRZYN, A. and WQGRZYN, G., 1994b. Regulation of replication of plasmid pBR322 in amino acid-starved Escherichiu coli strains. Mol. Gen. Genet., 243, 374-378.
HERMAN, A., WQGRZYN, A. and WQGRZYN, G., 1994c. Differential replication of plasmids during stringent and relaxed response of Escherichia coli. Plasmid, 32, 89-94.
HERNANDEZ, V. J. and BREMER, H., 1991. Escherichiu coli ppGpp synthetase I1 activity requires SPOT. J. Biol. Chem., 266,5991 -5999.
HOFMANN, K. H., NEUBAUER, P., RIETHDORF, S. and HECKER, M., 1990. Amplification of pBR322 plasmid DNA in Escherichiu coli relA strains during batch and fed-batch fermentation. J. Basic. Microbiol., 30, 37-41.
KAWASAKI, Y., WADA, C. and YURA, T., 1991. Mini-F plasmid mutants able to replicate in the ab- sence of a? mutations in the RepE coding region producing hyperactive initaitor protein. J. Bacte-
KONTOMICHALOU, P., MITANI, M. and CLOWES, R. C., 1970. Circular R-factor molecules, controlling penicillinase synthesis, replicating in Escherichiu coli under either relaxed or stringent control. J. Bacteriol., 1 0 4 , 3 4 4 .
KOTLER, R. and HELINSKI, D. R., 1978. Construction of R6K plasmid derivatives in vitro: characteri- sation of the R6K replication region. Plasmid, 1,571 -580.
LEVINE, A., VANNIER, F., DEHBI, M., HENCKES, G. and Seror, S. J., 1991. The stringent response blocks DNA replication outside the ori region in Bacillus subtilis and at the origin in Escherichiu coli. J. Mol. Biol., 219,605-613.
LEWIS-KITELL, B. and HELINSKI, D. R., 1991. Iteron inhibition of plasmid RK2 replication in vitro: evidence for intermolecular coupling of replication origins as a mechanism for RK2 replication control. Roc. Natl. Acad. Sci. USA, 88, 1389-1393.
1373 -1384.
n ~ l . , 173, 1064-1072.

462 B. WR6BEL and G. WQGRZYN
LIN-CHAO, S. and BREMER, H., 1986. Effect of relA function on the replication of plasmid pBR322 in Escherichia coli. Mol. Gen. Genet., 203, 150-153.
MANIS, J. J. and KLINE, B. C., 1977. Restriction endonuclease mapping and mutagenesis of the F sex factor replication region. Mol. Gen. Genet., 152, 175-182.
MATIN, A., 199 1. The molecular basis of carbon-starvation-induced general resistance in Escherichia coli. Mol. Microbiol., 5 , 3-10.
MOLIN, S . , STOUGAARD, P., UHLIN, B. E., GUSTAFSSON, P. and NORDSTROM, K., 1979. Clustering of genes involved in replication, copy number control, incompatibility, and stable maintenance of the resistance plasmid Rldrd-19. J. Bacteriol., 138, 70-79.
MURRAY, K. D. and BREMER, H., 1996. Control of spoT-dependent ppGpp synthesis and degradation in Escherichia coli. J. Mol. Biol., 259,41-57.
NEUBAUER, P., WROBEL, B. and WQGRZYN, G., 1996. DNA degradation at elevated temperatures after plasmid amplification in amino acid-starved Escherichia coli cells. Biotechnol. Lett., 18, 321 -326.
NORDSTROM, K., MOLIN, S. and LIGHT, J., 1984. Control of replication of bacterial plasmids: genet- ics, moecular biology and physiology of the plasmid R1 system. Plasmid, 12, 71 -90.
OBUCHOWSKI, M. and WQGRZYN, G., 1996. Synthesis of the bacteriophage I P protein in amino acid- starved Escherichia coli cells. Biochem. Biophys. Res. Commun., 222,612-618.
OGURA, T., NIKI, H., W o , Y., IMAMOTO, F. and HIRAGA, S., 1990. Maintenance of plasmids in HU and IHF mutants of Escherichia coli. Mol. Gen. Genet., 220, 197-203.
PAL, S. K., MASON, R. J. and CHAITORAJ, D. K., 1986. P1 plasmid replication: role of initiator titra- tion in copy number control. J. Mol. Biol., 192, 275-285.
REDDY, P. S., RAGHAVAN, A. and CHATTERJI, D., 1995. Evidence for ppGpp binding site on E. coli RNA polymerase: proximity relationship with the rifampicin binding domain. Mol. Microbiol., 15, 255 -265.
RIETHDORF, S., SCHROETER, A. and HECKER, M., 1989. RelA mutation and pBR322 plasmid amplifi- cation in amino acid-starved cells of Escherichia coli. Genet. Res., 54, 167-171.
SAMBROOK, J., FRITSCH, E. F. and MANIATIS, T., 1989. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Coled Spring Harbor, NY.
SCHREIBER, G., RON, E. Z. and GLASER, G., 1995. ppGpp-mediated regulation of DNA replication and cell division in Escherichia coli. Cum. Microbiol., 30, 27-32.
SCHROETER, A., RIETHDORF, S. and HECKER, M., 1988. Amplification of different ColEl plasmids in an Escherichia coli relA strain. J. Basic Microbiol., 28,553-555.
SEROR, S. J., VANNIER, F., LEVINE, A. and HENCKES, G., 1986. Stringent control of initiation of chro- mosomal replication in Bacillus subrilis. Nature, 321, 709-7 10.
SZALEWSKA-PALASZ, A. and WQGRZYN, G., 1994. An additional role of transcriptional activation of ori I in the regulation of I plasmid replication in Escherichia coli. Biochem. Biophys. Res. Com- mun., 205,802-806.
SZALEWSKA-PALASZ, A. and WFGRZYN, G., 1995. Inhibition of transcription starting from bacterio- phage I pR promoter during the stringent response in Escherichia coli: implications for I DNA replication. Acta Biochim. Polon., 42,233 -240.
SZALEWSKA-PALASZ, A., WEGRZYN, A., HERMAN, A. and WQGRZYN, G., 1994. The mechanism of the stringent control of I plasmid DNA replication. EMBO J., 13, 5779-5785.
WEGFUYN, G., 1995. Amplification of I plasmids in Escherichia coEi relA mutants. J. Biotechnol., 43, 139-143.
WQGRZYN, G. and TAYLOR, K., 1992. Inheritance of the replication complex by one of two daughter copies during I plasmid replication in Escherichia coli. J. Mol. Biol., 226,681-688.
WGGRZYN, A. and WQGRZYN, G., 1995. Transcriptional activation of on' I regulates I plasmid replica- tion in amino acid-starved Eschen'chiu coli cells. Biochem. Biophys. Res. Commun., 214,978-984.
W~$%~YN, G., KWASNIK, E. and TAYLOR, K., 1991a. Replication of I. plasmid in amino acid-starved strains of Escherichia coli. Acta Biochim. Polon., 38, 181 -1 86.
WQGRZYN, G., NEUBAUER, P., KRUEGER, S., HECKER, M. and TAYLOR, K., 1991 b. Stringent control of replication of plasmids derived from coliphage I . Mol. Gen. Genet., 225,94-98.
WEGRZYN, A., WEGRZYN, G. and TAYLOR, K., 1995. Plasmid and host functions required for I plas- mid replication carried out by the inherited replication complex. Mol. Gen. Genet., 247, 501 -508.
WQGRZYN, A., TAYLOR, K. and WEGRZYN, G., 1996a. The cbpA chaperone gene function compen- sates for dnaJ in 1 plasmid replication during amino acid starvation of Escherichia coli. J. Bacte- r i~ l . , 178, 5847-5849.

Stringent control of plasmid DNA replication 463
W~GRZYN, A., WQGRZYN, G., HERMAN, A. and TAYLOR, K., 1996. Protein inheritance: 1 plasmid re- plication perpetuated by the heritable replication complex. Genes Cells, 1,953 -963.
WROBEL, B. and WQGRZYN, G., 1997. Replication and amplification of 1 plasmids in Escherichiu coli during amino acid starvation and limitation. FEMS Microbiol. Lett., 153, 151 -157.
WU, F., GOLDBERG, I. and RLUTOWICZ, M., 1992. Roles of a 106-bp origin enhancer and Escherichiu coli DnaA protein in replication of plasmid R6K. Nucleic Acids Res., 20, 81 1-817.
XIAO, H., KALMAN, M., IKEHARA, K., ZEMEL, S., GLASER, G. and CASHEL, M., 1991. Residual gua- nosine 3',5'-bispyrophosphate synthetic activity of re& null mutants can be eliminated by spoT null mutations. J. Biol. Chem., 266, 5980-5990.
YAVACHEV, L. and IVANOV, I., 1988. What does the homology between E. coli tRNAs and RNAs controling ColEl plasmid replication mean? J. Theor. Biol., 131,235-241.
Mailing address: Dr. GRZEGORZ WQGRZYN, Department of Molecular Biology, University of Gdahsk, Kladki 24, 80-822 Gdansk, Poland e-mail: [email protected]




















