Embed Size (px)
Citation preview


ISSN 0325-8718ISSN 1669-2314
Institución Editora: Instituto Nacional de Tecnología Agropecuaria (INTA)
Cuatrimestral
Vol. 46 N.º 2Agosto 2020, Buenos Aires, Argentina
Propiedad IntelectualPropiedad Intelectual N.º 5231354
SEDE EDITORIAL
Chile 460 2.º piso Ciudad Autónoma de Buenos Aires, Argentina
CP (C1098AAJ) +54 (011) 4339-0600http://ria.inta.gob.ar
La Revista de Investigaciones Agropecuarias (RIA) es una publicación de información
científica editada por el INTA. Los artículos presentados son admitidos
por un Comité Editorial compuesto por prestigiosas figuras del ámbito académico
y científico agropecuario. Abarca diferentes temas preparados
por profesionales del INTA y de otras instituciones, y está destinada
a la comunidad científica nacional e internacional.
Lo expresado por autores, corresponsales o columnistas no necesariamente reflejan
el pensamiento del Comité Editorial, de la revista o de su institución editora.
Indexada en: Scopus
Latindex (Folio 3346)CABI Publishing
DOAJ (Directory of Open Access Journals)Redalyc.org
Núcleo BásicoThomson Reuters
DialnetScielo
CiteFactor
Esta publicación es propiedad del Instituto Nacional de Tecnología Agropecuaria. Rivadavia 1439. CABA. Buenos Aires, Argentina.
MIEMBROS DEL CONSEJO DIRECTIVO DEL INTA
Presidente:
Dr. Carlos Di Bella. Universidad de Buenos Aires (UBA) y Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Argentina.
Vocales:Ing. Agr. Jorge Mariotti. Universidad Nacional de Salta (UNSa). Argentina.
Dr. Tomás Miguel Schlichter. Instituto Nacional de Tecnología Agropecuaria (INTA). Argentina.
Dra. Ana María Romero. Universidad de Buenos Aires (UBA). Argentina.
Ing. Agr. M. Sc. María Eugenia Beget. Instituto Nacional de Tecnología Agropecuaria (INTA). Argentina.
Dr. Miguel Ángel Taboada. Instituto Nacional de Tecnolo-gía Agropecuaria (INTA) y Consejo Nacional de Investi-gaciones Científicas y Técnicas (CONICET). Argentina.
Dr. Martín Durante. Instituto Nacional de Tecnología Agropecuaria (INTA). Argentina.
Dr. Frédéric Goulet. Instituto Nacional de Tecnología Agropecuaria (INTA) y Centre de Coopération Internationale en Recherche Agronomique pour le Développement (CIRAD). Francia.
Dr. Ariel Pereda. Instituto Nacional de Tecnología Agro-pecuaria (INTA) y Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Argentina.
Dr. Laureano S. Frizzo. Universidad Nacional del Litoral (UNL).
Dr. Claudio Machado. Universidad Nacional del Centro de la Provincia de Buenos Aires (UNCPBA). Argentina.
Dr. Juan Wahren. Instituto de Investigaciones Gino Ger-mani (IIGG). Facultad de Ciencias Sociales. Universidad de Buenos Aires. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Argentina.
Dra. Karina Trono. Instituto Nacional de Tecnología Agropecuaria (INTA) y Consejo Nacional de Investigacio-nes Científicas y Técnicas (CONICET). Argentina.
Dra. Gabriela Posse. Instituto Nacionas de Tecnología Agropecuaria (INTA). Argentina.
COMITÉ EDITORIAL
Presidenta:Ing. Agr. Susana Mirassou
Vicepresidente:Ing. Agr. Tomás Schlichter
Consejeros Nacionales: Dr. Manuel Pedreira. MinAgroIng. Agr. Liliana Monterroso. Facultad de Cs. AgronómicasMéd. Vet Humberto Luis José Occhi. Facultades de Ciencias Veterinarias Ing. Agr. Alejandro Conci. AACREA Ing. Agr. Mario Nicolás Carlino. CONINAGRO Ing. Agr. Sergio Melgarejo. CRADr. Pablo Paillole. FAA Sr. Santos Zuberbhuler. SRA
Director Nacional:Ing. Agr. Carlos Parera
Coordinador Nacional Técnico:Ing. Agr. José Luis Sponton
Directores Nacionales Asistentes:DN Asistente de Investigación, Desarrollo y Planificación Ing. Agr. Martín Irurueta
DN Asistente de Transferencia y Extensión Ing. Agr. María Rosa Scala
DG de Sistemas de Información, Comunicación y Procesos Ing. Fernando Ricitelli
DG de Capital Humano Ing. Prod. Agr. Eduardo Alberto Quargnolo
DG de Administración CPN Lic. Rosendo Tarsetti
CN de Vinculación Tecnológica y Relaciones InstitucionalesIng. Zootecnista Juan Sebastián Llorens
Ing. Agr. M. Sc. Marina Di Filippo. Instituto Nacional de Tecnología Agropecuaria (INTA). Argentina.
Ing. Agr. M. Sc. Emiliano Quiroga. Instituto Nacional de Tecnología Agropecuaria (INTA). Argentina.
Dr. Darío Fernández. Instituto Nacional de Tecnología Agropecuaria (INTA). Argentina
Ing. Agr. M. Sc. Alejandro Rago. Instituto Nacional de Tecnología Agropecuaria (INTA). Argentina.
M. Sc. María Isabel Tort. Instituto Nacional de Tecnología Agropecuaria (INTA). Argentina.
Ing. Agr. M. Sc. Damián Vega. Universidad de Buenos Aires (UBA). Argentina.
Ing. Agr. Sergio E. Feingold. Instituto Nacional de Tecnología Agropecuaria (INTA). Argentina.
COMITÉ ASESOR
Dirección:Dra. Norma Pensel Lic. Máximo Bontempo
Coordinación editorial:Per. Giselle MiquetMg. Mario Migliorati
Asistente editorial:Lic. Gustavo Ciuffo Lic. Cecilie Esperbent
Producción Editorial:Asesores técnicos y de contenido Ing. Agr. Cristian Zuchini Dr. Martín IruruetaDra. Mariana Insani
Diseño y Edición:Comunicación Visual, Gerencia de Comunicación e Imagen Institucional
Fotografía:Audiovisual, Gerencia de Comunicación e Imagen Institucional
STAFF

Editorial
El consumo de carnes ha aumentado sustancialmente en las últimas décadas como consecuencia del crecimiento po-blacional y del incremento en los ingresos de los países emer-gentes y en vías de desarrollo, y según la Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO) se espera que esta tendencia continúe. La Organización de las Naciones Unidas (ONU) estima que hacia el 2050 la po-blación mundial se encontrará entre los 9.4 y 10.1 billones de habitantes. Para garantizar la seguridad alimentaria de esta población en aumento será necesario un incremento en la producción de proteínas para que todos tengan la posibilidad de una dieta equilibrada. Como consecuencia, los sistemas alimentarios a nivel global enfrentan un gran desafío, ya que los recursos se agotan conforme crece la demanda.
En paralelo, el mundo afronta la urgente necesidad de adop-tar medidas que ayuden a mitigar el impacto del calentamiento global. Esto, sumado a un incremento de la conciencia social por el medio ambiente, el bienestar animal y la salud son las principales motivaciones de una tendencia creciente hacia el cambio de dietas ricas en carne a otras con un mayor aporte de vegetales, con el consenso de los organismos mundiales sobre sus beneficios asociados. En este sentido, en agosto de 2019, el Panel Intergubernamental de Cambio Climático de las Naciones Unidas, publicó un informe que remarca como punto clave la necesidad de la adopción de una dieta con un mayor aporte de vegetales para combatir el calentamiento global.
Dicha tendencia sostenible combinada con la necesidad cre-ciente de proteínas de una población en expansión, ha promo-vido la obtención de nuevas fuentes proteicas alternativas a las de origen animal, entre las cuales se destacan las derivadas de semillas de cereales y legumbres.
Además de la relevancia que poseen estas nuevas fuentes proteicas para la alimentación, su procesamiento ha permiti-do obtener productos similares a la carne, también conocidos como análogos o imitadores, que pretenden ser una réplica de esta en todas sus propiedades (flavor, aroma, textura, mordida, humedad, sensación en boca, apariencia y color) con avances muy significativos. La sustitución parcial de carne a través de la mezcla o el blending con proteínas vegetales constituye en la actualidad otra estrategia emergente utilizada para lograr pro-ductos con una menor proporción de carne pero que respeten su esencia.
En relación a ello, se vislumbra un posicionamiento de estas nuevas categorías de productos siendo sus principales inver-sores y promotores las mayores empresas cárnicas del mundo. Recientemente INTA ha iniciado líneas de trabajo vinculadas al desarrollo de análogos de carne y blends preparados a partir de proteínas de legumbres.
El trabajo del INTA en una línea de selección de genotipos, para su adaptación a las variadas condiciones que presenta el territorio argentino, involucra a cultivos alternativos como la quinua y amaranto en ambientes con limitantes de sales o pre-cipitaciones.
No debe perderse de vista que, en busca del éxito, las al-ternativas cárnicas deben lograr reunir los mismos impulsores que promueven el consumo de carne: sabor y textura, precio y conveniencia, y aspectos nutricionales.
Este escenario plantea una oportunidad de posicionamiento para el mercado argentino de legumbres y proteínas obtenidas a partir de las mismas, con innegable capacidad de responder a estas nuevas tendencias.
Fuentes proteicas alternativas

RIA / Vol. 43 N.º 3 Diciembre 2017, Argentina.
Contenido
Actualidad en I+DEditorial
Fuentes proteicas alternativas134
Experimentan una tendencia creciente donde se destacan las derivadas de semillas de cereales y legumbres.
Notas
Alimentos del futuro: crece la demanda de proteínas alternativas a la carne
136
Especialistas evalúan cómo satisfacer estas necesidades y abastecer un mercado en alza en Argentina.
Camas biológicas: una herramienta versátil y proacti-va para el uso adecuado de fitosanitarios
140
La preservación del ecosistema demanda articulación públi-co-privada para dar soluciones a problemas del productor.
Construyen una biblioteca de nanoanticuerpos de llamas contra la COVID-19
145
Aporta información útil sobre anticuerpos que producen estos camélidos contra el antígeno de interés.
RIA / Vol. 46 N.º 2 Agosto 2020, Argentina.
Entrevista: “Hay muy pocas instituciones en el mundo como el INTA”
148
Carlos Parera repasa su gestión como director nacional y la situación del INTA frente a la COVID-19.
Caracterización de accesiones de garbanzo (Cicer arieti-num L.) mediante descriptores morfo agronómicos cuan-titativos en una colección del Banco de Germoplasma de la Universidad Nacional de Córdoba
156
Castillo, V.; Collavino, N.; Farfan, E.; Carreras, J.; Fekete, A.; Pocovi, M.
RevisiónTrabajos
Técnica de sexado de pupas de Anthonomus grandis (Coleoptera: Curculionidae): una forma rápida y fácil de sexar para estudios biológicos
167
Blettler, D.C.; Gúndez, G.A.; Chemez, D.M.
Comunicación corta
Artículos
Niveles plasmáticos de calcio, magnesio y fósforo en rodeos lecheros del Valle de Lerma, Salta, Argentina
181
Martínez, G.M.; Micheloud, J.F.; Suárez, V.H.; Rosa, D.E.; Ventura, M.V.; Ortega, A.; Sánchez, C.; Mattioli, G.A.
Prevalencia y factores de riesgo asociados a las enfermedades de transmisión sexual en bovinos de la provincia de La Pampa, Argentina
187
Miranda, A.; Fort, M.; Fuchs, L.; Baldone, V.; Giménez, H.; Carné, L.; Sago, A.; Otermin, H.; Dubié, D.; Goyeneche, P.; Campero, C.
Respuesta a la frecuencia de defoliación de plantas de festuca alta de una población naturalizada infectada con endófito silvestre y un cultivar comercial libre
195
Petigrosso, L.R.; Navarro, D.; Assuero, S.G.; Vignolio, O.R.; Castaño, J.A.; Colabelli, M.N.
Propiedades mecánicas de la madera de álamos de cortinas forestales de Río Negro, Patagonia, Argentina
203
Medina, A.A.; Manzione, P., Baucis, A.G.; Catalán, M.A.; Laffit-te, L.; Andía, I.R.
Seed priming induces biochemical changes in melon plants and increase salt tolerance
208
Castañares, J.L.; Bouzo, C.A.
Caracterización socioproductiva de los productores hortícolas en el partido de Junín (Buenos Aires)
218
Muzi, M.E.; Prividera, G.; Melilli, P.; Beribe, M.J.; Monsutti, M.V.; Telleria, M.G.; Fernandez, L.; Meccia, J.
¿Cómo ha cambiado la precipitación en la provincia de Santa Fe?
226
Lovino, M.A.; Müller, G.V.; Sgroi, L.C.
Evaluación del expeller de soja en dietas de cerdos en recría y engorde
240
Murcia, V.N.; Pordomingo, A.J.; Pordomingo, A.B.; Franco, R.
Transformación eficiente de Solanum tuberosum cv. Spunta mediada por Agrobacterium tumefaciens utili-zando higromicina como agente selectivo
248
Décima Oneto, C.; Coronel, J.; Storani, L.; González, M.N.; Feingold, S.; Massa, G.A.
Promotion of Lotus tenuis and calf early weaning as a good management practice for breeding herds in marginal soils of the Flooding Pampa (Argentina)
267
Bailleres, M.; Campestre, M.P.; Antonelli, C.J.; Melani, G.; Menéndez, A.; Ruiz, O.A.
Entrevista: “El desafío es generar herramientas que lleguen al productor”
153
Yésica Chazarreta recorre su inicio en la investigación orien-tada en el control genético y ambiental del maíz.
Restricción del crecimiento intrauterino y peso del cerebro en relación con sus órganos vitales en lechones neonatos: consideraciones para la producción porcina
171
Romanelli, A.; Decundo, J.M.; Martínez, G.; Fernandez Pag-gi, M.B.; Pérez Gaudio, D.S.; Dieguez, S.N.; Soraci, A.L.
First report of Bacterial Spot with gum production in almond fruits of Argentina
177
Marangi, M.J.; Temperini, C.V.; Fernández, D.; Pardo, A.G.; Pose, G.N.
Evaluación de la calidad de subproductos de la soja258
Seijas Noya, I.; Jaurena, G.; Wawrzkiewicz, M.; Gaggiotti, M.; Romero, L.A.; Méndez, M.

136 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2136 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2
Alimentos del futuro: crece la demanda de proteínas alternativas a la carne
Cada vez son más los consumidores que eligen dietas especiales basados en sus valores. En su mayoría, los motiva la conciencia ambiental, el bienestar ani-mal y la salud. Este marcado interés en saber qué contienen y cómo fueron ob-tenidos los productos que los argentinos ponen en su mesa impulsa una tenden-cia en auge.
Más informados y exigentes, los con-sumidores tienden a evitar alimentos con ingredientes o aditivos sintéticos en exceso y demandan aquellos sa-ludables, nutritivos y clean label o con
etiquetado limpio. Esta tendencia, que comenzó de la mano de los vegetaria-nos y se extendió con los veganos, hoy alcanza a los flexitarianos.
Esta nueva categoría de consumidores agrupa a quienes, por diversos motivos, decidieron reducir al mínimo el consu-mo de carne animal, denominados ve-getarianos flexibles. Según una reciente investigación de mercado realizada por Innova Market Insights (empresa líder mundial de conocimiento en la industria de alimentos y bebidas), Alemania lide-ra este movimiento con un 69 % de su
población que reconoce consumir carne una vez a la semana, seguido por el 53 % en el Reino Unido y el 38 % en Esta-dos Unidos.
Por su parte, un informe de enero de 2020 de Markets and Markets estima que el mercado mundial de sustitutos de la carne para 2026 alcanzará los 3,5 mil millones de dólares, lo que repre-senta una tasa compuesta anual del 12 % durante el período de pronóstico. El mercado está impulsado principalmen-te por la creciente demanda entre los millennials.
Alimentos del futuro: crece la demanda de proteínas alternativas a la carne
TENDENCIA CRECIENTE
POR LAURA PÉREZ CASAR
Motivados por la conciencia ambiental, el bienestar animal y la salud, cada vez más consumidores prefieren alimentos de origen vegetal y análogos de la carne. En este contexto, las legumbres ocupan un lugar preferencial y se presentan como una oportunidad única para la Argentina, que cuenta con potencial para abastecer un mercado en auge.

137Agosto 2020, Argentina 137Agosto 2020, Argentina
Alimentos del futuro: crece la demanda de proteínas alternativas a la carne
Ahora bien, ¿qué es un producto aná-logo de la carne?, ¿existen alimentos capaces de reemplazar a la proteína ani-mal?, ¿qué lugar ocupan las legumbres en este escenario?, ¿es una oportunidad para la Argentina?, ¿hay potencial para satisfacer este mercado en auge?
Para Trinidad Soteras, investigadora del Instituto de Tecnología de Alimentos del INTA Castelar, “a escala global, exis-te una demanda creciente de consumi-dores que buscan productos alternativos a la carne motivados, principalmente, por aspectos ambientales, nutricionales y relacionados con la conciencia animal. Aunque, también, exigen variedad y buen sabor”.
Así, surgen los análogos de carne que componen una clase de produc-tos alimenticios sin proteínas cárnicas que intentan imitar sus características desde el punto de vista organoléptico y nutricional, simulando las clásicas ham-burguesas, salchichas y nuggets, entre otros.
Un estudio de la Dirección Nacional de Alimentos y Bebidas del ex Ministerio de Agroindustria de la Nación –actual Ministerio de Agricultura, Ganadería y Pesca- confirmó que se consolida una tendencia hacia la búsqueda de produc-
tos sustitutivos de la carne. Entre los alimentos emergentes se destacan las legumbres, semillas, granos y cereales ancestrales o cultivos andinos, y los fru-tos secos. En esta línea, aseguran que se trata de “una oportunidad para em-prender con éxito”.
Gabriel Prieto –referente del INTA en cultivos de invierno– dio un paso más y reconoció: “Desde mediados de 2019 vemos una tendencia creciente y sos-tenida en nuestro país”, destacó el es-pecialista de Arroyo Seco –Santa Fe–, quien confirmó que “la demanda aumen-tó significativamente”.
Es que, a la tendencia global de ali-mentación saludable, se suman factores tales como un menor precio de las le-gumbres frente a los productos cárnicos, el mayor rendimiento culinario y su fácil conservación.
Como si fuera poca la evidencia, una reciente encuesta sobre los hábitos de los consumidores realizada por el Insti-tuto de Promoción de la Carne Vacuna Argentina (IPCVA) mostró que un tercio de los consultados declaró haber redu-cido la proporción de carnes en la ali-mentación, lo que lo amplía el margen de habitantes dentro de la categoría de flexitarianos.
Para Soteras, “la necesidad de dispo-ner de alimentos de calidad para una po-blación en expansión sumada a la impe-rante necesidad mundial de lograr que las cadenas de valor sean más sosteni-bles, hace que las proteínas provenien-tes de fuentes distintas de los animales entren en escena con fuerza, solas o en combinación con la carne”.
Según la investigadora, “es indiscutible que los paradigmas de la alimentación se encuentran en plena transformación y los esfuerzos de la industria de aná-logos están enfocados en reducir al mí-nimo la brecha entre el gusto, la textura y la experiencia de comer carne con aquella experimentada al consumir sus análogos libres de carne”.
Y agregó: “Si bien la adopción de los productos alternativos a la carne se en-cuentra en pleno desarrollo, se vislum-bra el auge de estos por parte del sec-tor alimentario, al tiempo que se espera que el número de flexitarianos continúe creciendo”.
Legumbres: protagonistas en la transformación
Arvejas, lentejas y garbanzos. Las tres especies de legumbres se presentan
Desde mediados de 2019, la demanda
de legumbres en la Argentina crece de
manera significativa.

138 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2138 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2
Alimentos del futuro: crece la demanda de proteínas alternativas a la carne
como alternativas promisorias. En el li-bro “Legumbres, semillas nutritivas para un futuro sostenible” de la Organización de las Naciones Unidas para la Alimen-tación y la Agricultura (FAO) se destacan las propiedades de este alimento y se lo considera como uno de los más nu-tritivos del planeta y extraordinariamente beneficioso para la salud.
Además, se las describe como plantas muy eficientes: su cultivo permite intensi-ficar las rotaciones, con beneficios en los cultivos sucesores con el nitrógeno fijado naturalmente y una mayor disponibilidad de agua. A su vez, tienen un costo bajo para los agricultores, pueden ser cultiva-das en tierras secas con un tiempo de conservación prolongado.
En este mismo sentido, Prieto subrayó que “una de las principales ventajas que tiene la incorporación de legumbres en los esquemas productivos, a diferencia de los cereales, es la capacidad de fijar nitrógeno del aire, mediante la simbiosis con bacterias específicas, y mejora la es-tructura del suelo a partir de rotaciones más intensas en siembra directa”.
“Cuando se incorporan legumbres, como antecesor de los cultivos de vera-no, los beneficios se multiplican, no solo porque tienen un consumo de agua me-nor que el trigo o porque dejan un mejor balance de nitrógeno en el suelo, sino
que, además, desocupan el lote 20 días antes que el trigo lo que permite asegu-rar así más recursos para el cultivo suce-sor”, destacó Prieto.
Con respecto al garbanzo, Ana Fekete, investigadora del INTA Cerrillos –Salta– señaló que “tiene perspectivas positi-vas en cuanto a su participación en el comercio exterior” y destacó la extensa trayectoria del INTA Salta en el desarro-llo de cultivares mejorados en un con-texto nacional con poca disponibilidad de variedades.
“Más allá de ser una leguminosa para grano seco importante por la cantidad y calidad nutritiva de sus componentes, hoy representa una oportunidad para la Argentina, por su calidad para exporta-ción a países Mediterráneos de Europa, así como del Mercosur y andinos de América”, expresó Fekete.
Un consumidor que demanda, un sector que ¿responde?
Las últimas cifras de la Cámara de Legumbres de la República Argentina (CLERA) coinciden con las publicadas por la Bolsa de Comercio de Rosario (BCR) y aseguran que la producción de legumbres en la campaña 2017/18 alcanzó las 686.500 toneladas en la Ar-gentina, mientras que las exportaciones
alcanzaron el 75 % de la producción con 512.000 toneladas.
Incluso, hay estimaciones para el sec-tor que calculan una proyección de cre-cimiento de hasta el 20 % en los próxi-mos años en la demanda de alimentos de origen vegetal y análogos de la carne que ubican a las leguminosas en un rol protagónico. Ahora bien, ¿cuáles son los desafíos a los que se enfrenta el sec-tor?, ¿la Argentina puede abastecer este mercado en auge?, ¿es realmente una oportunidad?
Para Prieto, “la Argentina es el país con mejores chances del mundo de sa-tisfacer estas necesidades y abastecer este mercado en alza”. De acuerdo con el especialista, nuestro país presenta ventajas competitivas clave tales como la bondad de sus inviernos y la fertilidad
Por la bondad del clima y la fertilidad de los suelos, la Argentina se destaca en el mundo por su potencial para abastecer lacreciente demanda.
Una encuesta delIPCVA mostró que un tercio de consumidoresdeclaró haber reducido la proporción decarnes en su alimentación.

139Agosto 2020, Argentina 139Agosto 2020, Argentina
Pocos países del mundo “pueden ha-cer doble cultivo como la Argentina por-que los inviernos son tan crudos que no tienen tierra disponible para trabajar”, indicó Prieto.
La promoción, un desafíoDe acuerdo con el referente en legum-
bres del INTA, esta creciente demanda plantea un desafío para los productores y toda la cadena de legumbres, princi-palmente, referida a la promoción para la inclusión de estos cultivos.
“Es importante que desde el gobierno y desde los organismos técnicos difun-damos las bondades de este cultivo, fo-
Alimentos del futuro: crece la demanda de proteínas alternativas a la carne
Los cambios de hábito en la alimentación
representan un desafío y una oportunidad
para el sector productivo nacional.
de sus suelos, con menores costos que implica la fertilización o riego.
Es que, según detalló, las legumbres de invierno –arveja, lenteja y garbanzo– se siembran sin resignar a los cultivos de verano como soja y maíz. Además, no compiten con otros cultivos de invierno como la cebada o el trigo.
“Durante el invierno, se ocupa entre el 25 y el 35 % de la tierra disponible en la zona núcleo”, aseguró, al tiempo que recalcó: “Hay un 60 % de suelo barbe-chado durante los meses invernales con capacidades oseosas, a merced de las malezas tolerantes y la pérdida de agua por el consumo de las malezas o por la evaporación directa”.
mentemos su incorporación a las dietas forrajeras y hasta impulsemos su incor-poración como ingrediente a la industria alimenticia”, recalcó Prieto.
Es que, para el referente del INTA, cor-tar la harina de trigo con harina de legum-bres en la industria panificadora implica potenciar el aporte adicional de proteínas y de otros nutrientes tales como hierro y zinc a los alimentos. También puede ser incluida en otras recetas y sumarlo desde las golosinas hasta los fideos.
Más información: Trinidad Soteras [email protected]; Gabriel Prieto [email protected]; Ana Fekete [email protected]

140 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2140 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2
Camas biológicas: una herramienta versátil y proactiva para el uso adecuado de fitosanitarios
“Las BPA tienen 7 requisitos o criterios para poder certificar. Uno de los requisitos habla de la gestión sostenible y responsable de los fitosanitarios” (M. C. Rivas).
La presión por el aumento de la pro-ductividad conlleva una exigencia mayor en los recursos naturales y en los recur-sos humanos que están afectados a esa producción. El mundo demanda cada vez más productos que cuenten con cer-tificación de su proceso de producción, y el valor agregado ambiental resulta fun-damental en un contexto de consumido-res cada vez más exigentes y atentos al cuidado de los recursos naturales.
Nuestro país se encuentra en el cuarto lugar en cuanto al consumo de fitosani-tarios, luego de China, Estados Unidos y Brasil. Por ese motivo resulta fundamen-tal desarrollar tecnologías innovadoras y accesibles que respondan de una mane-ra sustentable al problema de la contami-nación por lo cual las camas biológicas o biobeds se convirtieron en una solución
BUENAS PRÁCTICAS
POR VALERIA GUERRAPOR VALERIA GUERRAFOTOS GENTILEZA CASAFEFOTOS GENTILEZA CASAFE
viable y práctica para proteger los recur-sos naturales y la salud humana.
De acuerdo con la Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO), las Buenas Prác-ticas Agrícolas (BPA) consisten en “la aplicación del conocimiento disponible a la utilización sostenible de los recursos naturales básicos para la producción, en forma benévola, de productos agrícolas alimentarios y no alimentarios inocuos y saludables, a la vez que se procura la viabilidad económica y la estabilidad social”.
“Las BPA tienen 7 requisitos o criterios para poder certificar. Uno de los requi-sitos habla de la gestión sostenible y responsable de los fitosanitarios”, indi-có María del Carmen Rivas, investiga-dora del Instituto de Suelos del Centro
Camas biológicas: una herramienta versátil y proactiva para el uso adecuado de fitosanitariosEsta tecnología simple, económica y de fácil mantenimiento permite la preservación del ecosistema, el cuidado de la salud humana y el cumplimiento de las nuevas exigencias quedemanda el mercado nacional e internacional bajo el principio de bioprofilaxis.
de Investigación de Recursos Naturales (CIRN) del INTA.
En las reglas de juego que define el mercado actual, una herramienta de dife-renciación con un rol cada vez más pro-tagónico es el valor agregado ambiental

141Agosto 2020, Argentina 141Agosto 2020, Argentina
Camas biológicas: una herramienta versátil y proactiva para el uso adecuado de fitosanitarios
“Es importante buscar la mejora continua
y apostar por el cuidado de los
recursos naturales” (M. C. Rivas).
para los productos agropecuarios. “Este valor se obtiene cuando se pone en evi-dencia del consumidor que todo el pro-ceso que involucró obtener ese alimento fue inocuo con respecto al ambiente, lo cual satisface su preocupación con respecto al cuidado de los recursos na-turales y cómo se están gestionando”, expresó Rivas.
El origen de las camas data de Suecia en la década de 1990, a través de la in-vestigadora María del Pilar Castillo del Instituto Sueco de Ingeniería Agrícola y Ambiental, como una respuesta a la ne-cesidad de encontrar un sistema sencillo y efectivo para minimizar la contamina-ción por plaguicidas. Es una iniciativa proactiva para abordar y minimizar el riesgo ambiental de la manipulación de fitosanitarios con métodos y protocolos de fácil adopción para el productor.
En ese contexto, desde el INTA se ha comenzado a promocionar el uso de las camas biológicas a raíz del vínculo ge-nerado con Castillo, quién fue invitada en el año 2013 al Primer Taller Interna-cional de Biorremediación (PRITIBIO), organizado por el Instituto de Suelos del
INTA y la Facultad de Agronomía de la UBA (FAUBA), del cual resultó la poste-rior publicación del libro Biorremediación de los recursos naturales, que incluye un capítulo dedicado a esta tecnología, es-crito por Castillo y Leticia Pizzul.
¿En qué consisten las camas biológicas?
Son una construcción sencilla y ver-sátil diseñada para retener derrames y degradar los fitosanitarios. Se imple-mentan varios diseños, según diferentes situaciones, tipo de producción agrícola, condiciones climáticas y disponibilidad de recursos. Consisten en una excava-ción en el suelo que varía de los 60 cm a 1 metro de profundidad, rellenada en el modelo sueco de abajo hacia arriba por una capa de arcilla, una biomezcla de paja, suelo y turba y una capa de cés-ped en la superficie. El propósito es la retención de los líquidos, en caso de que ocurran derrames accidentales durante el llenado del equipo, y la degradación de los compuestos químicos a través de la acción de los microorganismos que se desarrollaron en la biomezcla.
Hay dos momentos críticos en el ma-nejo seguro de los fitosanitarios: el pre-parado del caldo y llenado del tanque-mochila aplicadora; el lavado (interno y externo) del equipo aplicador y de los envases junto con su correcta disposi-ción final. En esos momentos críticos la recomendación clásica tradicional es contar con equipos o elementos de contención antiderrames que actúan de una forma reactiva cuando el “daño” ya sucedió.
En ese sentido, y de acuerdo con Ri-vas, lo que caracteriza a las camas bio-lógicas, además de su versatilidad es su proactividad. “Queremos recuperar el concepto de bioprofilaxis, que es de origen griego, integrada por pro que sig-nifica “antes”, y por philax que designa a un “guardián o protector”, subrayó.
Además, agregó: “Es importante bus-car la mejora continua y apostar por el cuidado de los recursos naturales. Al productor esto además le tiene que ce-rrar económicamente. Se puede hacer tan simple o tan sofisticado como la si-tuación de escala de producción lo de-mande y esa es la versatilidad que tiene”.
Cama biológica inditrecta.

142 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2142 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2
En 2018 se propuso en el IRAM ha-cer una normativa, y durante el mes de agosto salió a consulta pública la Norma IRAM “Guía para la construcción y ma-nejo de camas biológicas”, la cual, una vez publicada, resultará fundamental para el desarrollo masivo de esta he-rramienta. En la comisión de Calidad de Suelos trabajaron no solamente desde el INTA, sino también desde otros orga-nismos como, por ejemplo: CONICET, el Instituto de Desarrollo Tecnológico para la Industria Química (INTEC) de Santa Fe y la Cámara de Sanidad Agropecua-ria y Fertilizantes (CASAFE).
El especialista en implementación de Buenas Prácticas en el uso de Fitosani-tarios de CASAFE, Federico Elorza, ase-guró que vienen trabajando en la herra-mienta de camas biológicas desde 2013, cuando entró en contacto con el Instituto de Sanidad y Calidad Agroalimentaria de Mendoza (ISCAMEN). “Ellos son la autoridad de aplicación con el tema de agroquímicos de la provincia, y no sa-bían cómo hacer una disposición fi nal,
Cama biológica, aspectos a tener en cuenta
Se debe hacer un plan básico del sitio donde se va a instalar. Defi nir el tipo de cama más conveniente. Es fundamental tener en cuenta la cubierta vegetal, el sistema de impermeabilización, el sistema de recir-culación, el suelo, los sustratos orgánicos humifi cables y los materiales lignocelulósicos que intervendrán en la biomezclaLa cama tiene que estar ubicada en áreas con pendientes menores que el 10 % y separada 150 metros de cualquier curso o cuerpo de agua. Y a 150 metros de áreas anegables. En cuanto a la temperatura, si bien la ideal para la degradación es de 20°, las camas funcionan tanto en Suecia como en climas tropicales. Es necesario mantener un 60 % de humedad para que la degradación sea la correcta. En el caso de zonas con abundancia de precipitaciones es importante resguardarla poniéndole un techo o un plástico separado 20 cm del suelo para que corra aire y permita la evaporación del agua de la cama biológica para que el producto se degrade.
Camas biológicas: una herramienta versátil y proactiva para el uso adecuado de fi tosanitarios
por ello comenzamos a probar la técnica a ver si era efi ciente y dio unos resulta-dos muy buenos”.
CASAFE forma parte de lo que es Cro-pLife Latinoamérica, quienes en el año 2015 hicieron su Congreso en Argen-tina. “Trajimos a Pizzul de Suecia para difundir el uso de las camas biológicas y a partir de ese momento estuvimos trabajando muy fuerte en llevar adelante distintos ensayos e iniciativas en las que están nuestras compañías socias”, afi r-mó Elorza. En ese sentido, explicó que en 2019 lograron instalar las primeras 5 camas a nivel de productores extensivos y hoy hay en construcción 6 más en las provincias de Buenos Aires, Santa Fe y Córdoba. “La primera cama fue realizada en una empresa socia con un contratista de Pergamino”.
Actualmente, CASAFE está llevando adelante una serie de ensayos en Cha-co y Corrientes con el Instituto Fuentes Godo para evaluar la combinación que mejor funciona con distintos materiales. Además, están con un proyecto que
arrancó a principios de año, pero se tuvo que parar por diferentes inconvenientes, que es la instalación de camas bioló-gicas en distintas universidades, para que puedan hacer una disposición fi nal adecuada de los fi tosanitarios que usan para sus ensayos. “Creemos que es muy importante brindar esta tecnología, brin-dar información y ayudar a los jóvenes a hacer tesis de investigación al respecto”, aseguró el especialista.
Tipos de camas biológicasExisten las directas cerradas y abier-
tas, y las indirectas. En las directas se deposita la pulverizadora arriba de la cama biológica y el producto cae direc-tamente sobre la biomezcla. “Dentro de las directas, existen las directas cerradas donde la salida del líquido es mediante la evaporación, y las directas abiertas que tienen debajo de esta excavación un caño que recoge los líquidos. Luego cuando la pulverizadora no está estacio-nada arriba de la cama biológica, se as-
“Hay una necesidad muy fuerte deincorporar este tipo de herramientas. Es muy importante la colaboración del INTA…” (F. Elorza).
Biobeds abierta y cerrada.

143Agosto 2020, Argentina 143Agosto 2020, Argentina
“Desde el INTA generamos información
y tecnologías para el sector agropecuario
y, asimismo, evaluamos las tecnologías desde
todo punto de vista” (J. Montoya).
Camas biológicas: una herramienta versátil y proactiva para el uso adecuado de fitosanitarios
perja el líquido sobre esta, de manera de terminar de degradar todo el producto”, explicó el especialista de CASAFE.
El tercer tipo son las camas biológicas indirectas: “La pulverizadora se estacio-na sobre una plataforma de material que este impermeabilizada para recoger to-dos los líquidos en un tanque y asperjar en recipientes apartados de la pulveriza-dora. Ese tipo de cama biológica es una de las más económicas y es la que más se está implementando hoy en Argenti-na”, aseguró.
Tampoco deja de lado la posibilidad de hacer una cama biológica sencilla, en un tacho de 200 litros donde se coloca la biomezcla y permite apoyar una mochila pulverizadora. “Eso sirve para los peque-ños productores y el costo es muy bajo”.
Desde CASAFE han notado la alta de-manda que hay en capacitación de ope-rarios. “Hay una necesidad muy fuerte de incorporar este tipo de herramientas. Es muy importante la colaboración del INTA, porque en la articulación público-privada está la mejor combinación para solucionar este tipo de problemas del productor. Ahora necesitamos legisla-ción al respecto, porque esta tecnología es muy sencilla y permite bajar el costo al productor y hacer una disposición ade-cuada y ambientalmente amigable”.
Las biomezclas, el corazón de la cama
Está compuesta por un 50 % de mate-rial vegetal con un alto contenido de lig-nina, un 25 % de suelo del lugar, porque los microorganismos están adaptados a
El cuidado ambiental en la cartera de proyectos
“Desde el INTA generamos información y tecnologías para el sector agropecuario y, asimismo, evaluamos las tecnologías desde todo punto de vista, incluyendo la identificación y efectos colaterales que pudieran estar afectando negativamente al ambiente. Se trabaja también desde el punto de vista de mitigar los potenciales problemas que puedan ocasio-nar en el ambiente”, afirmó Jorgelina Montoya, coordinadora del Proyec-to Gestión Sostenible de Fitosanitarios del INTA.Como es sabido, el manejo inadecuado de fitosanitarios puede afectar negativamente a la calidad del suelo, al aire, al agua, a la biodiversidad y a la salud de la población. Por una parte, trabajos realizados por la institución han identificado situaciones de contaminación de agua subte-rránea asociada a eventos de derrames o manejo inadecuado del caldo de la pulverizadora. Por otra parte, el INTA recibe inquietudes e interro-gantes por parte de los mismos usuarios y aplicadores, interesados en conocer cómo manejar y qué hacer con los remanentes y residuos que se generan durante el uso de fitosanitarios. Todo esto ha motivado la ne-cesidad de profundizar desde la institución cómo manejar esos residuos, y la posibilidad del uso de las camas biológicas resulta una excelente opción. De acuerdo con Montoya, “lo importante de mostrar a la comunidad en lo que se está trabajando, es que a partir de ello se generan intercam-bios entre el sector público y privado que hacen que la temática se re-troalimente”. Y reconoció: “INTA genera el conocimiento científico técnico para llevar adelante las cosas, pero hace falta bajarlo al terreno, conocer en el territorio cuál es la demanda en concreto y su adaptación al medio, y ahí es donde el sector privado juega un rol fundamental”. En tal sentido, “desde hace aproximadamente 20 años se trabaja en el INTA en torno al tema de la contaminación agroambiental que incluye el estudio del impacto de los fitosanitarios en el ambiente. Esto ha ido creciendo en las carteras de proyectos y con una mirada cada vez más integral, con mayores interrelaciones entre los distintos proyectos en marcha”.Y concluyó: “Desde el proyecto se estudia el impacto que tienen los fitosanitarios en el ambiente y se trabaja generando tecnologías e información que mejoren la gestión de los fitosanitarios en el ámbito rural. Resulta indispensable la optimización del uso de los fitosanitarios, profundizando en el conocimiento y ajustando las buenas prácticas de manejo y capacitando en tal sentido”.
Cama biológica directa cerrada.

144 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2144 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2
Camas biológicas: una herramienta versátil y proactiva para el uso adecuado de fitosanitarios
esos productos fitosanitarios, y un 25 % de material humificado, que puede ser turba o reemplazarlo por compost, dado que la turba es un recurso finito y no re-novable. El compost tiene un alto conte-nido de materia orgánica y sirve para re-tener la humedad y al igual que la turba le da una buena estructura y ayuda en las propiedades físicas de la biomezcla.
Respecto a la turba, “en Suecia tienen suficiente, pero para el resto de los paí-ses no están al alcance de forma sen-cilla”, indicó Lucrecia Brutti, del Instituto de Suelos del INTA. En ese sentido, “la normativa del IRAM tiene una tabla de las distintas composiciones de carbono y nitrógeno de los residuos orgánicos, como paja de trigo, de cebada, de maíz, de girasol”.
Existen diferentes investigaciones del INTA y CONICET sobre biomezclas de
acuerdo a la zona donde esté ubicada la cama, los productos que se utilicen y la cantidad. Por ejemplo, en Argentina se probó a escala de laboratorio que la degradación de glifosato con una mezcla de residuos de alfalfa, rastrojo de trigo y resaca de río, mezclado con un porcen-taje de suelo, alcanzó un 99 % de la de-gradación en 63 días.
A la hora de explicar el proceso de de-gradación de los fitosanitarios Brutti ex-plicó que “crecen los microorganismos y usan para su crecimiento el carbono y el nitrógeno que hay en esa biomez-cla. Cuando hay degradación actúan por orden, primero los hongos. Cuando se acaba su principal insumo la población decae porque al no tener comida mue-re, y esa misma población es comida por otra tanda, son las bacterias que nece-sitan nitrógeno. De esa manera sigue el proceso. Van consumiendo carbono y eliminando oxígeno y anhídrido carbóni-co que va al aire”. El proceso es aeróbi-co. La paja es la que alberga el hongo blanco que degrada la lignina y tiene las enzimas que son capaces de degradar los fitosanitarios.
Economía circular En todo lo que respecta a las camas
biológicas entra en juego también el con-
“Las camas biológicas no son eternas,
podrían durar alrededor de 3 años
en nuestro país” (L. Brutti).
cepto de economía circular, que: recicla los residuos, minimiza el ingreso energé-tico a los sistemas, protege el ambiente y sus recursos naturales
“Las camas biológicas no son eternas, podrían durar alrededor de 3 años en nuestro país. Una vez que se colmató la cama, tengo que decidir qué hacer con ese residuo”, afirmó Brutti.
Es claro que el desdoblamiento de fi-tosanitarios no quiere decir que los pro-ductos resultantes sean inocuos. Ante ello, de acuerdo con la investigadora, pueden hacerse, en un laboratorio, prue-bas de ecotoxicidad usando organismos sensibles y manipulables como lombri-ces o semillas.
“Hay pruebas rápidas que hacen en 5 días. Ese material si no tiene problemas se puede utilizar en el campo desparra-mándolo en pequeñas cantidades, caso contrario se puede hacer una biopila ta-pada con plástico y colocada en lugar seguro con piso impermeabilizado y ca-nales de recolección de los posibles lixi-viados. En el lapso de 9 meses a 1 año se degradará lo que no se degradó en la cama. Incluso, ese compostaje se pue-de utilizar a futuro en una nueva cama o como acondicionador orgánico.
Siguiendo a Brutti, desde el Instituto de Suelos la propuesta que están haciendo es que ese residuo podría formar parte de un sustrato para hacer plantines flo-rales. “Desde Castelar le vimos la veta referida al AMBA y extendida al periurba-no: el fitosanitario en conflicto con el ha-bitante”, indicó y agregó: “Pretendemos trabajar para producciones intensivas, huertas, flores, que nos permitan inter-venir en esa problemática”.
En otras unidades del INTA, como Hi-lario Ascasubi, se está trabajando en la construcción de una cama biológica de la mano de Guillermo Tucat, de la Uni-versidad Nacional del Sur, CERZOS-CONICET. Por su parte, el INTA Cerrillos implementó una cama biológica para una demostración durante el Primer Congreso Nacional de Fitosanitarios, organizado en 2017 por CASAFE, INTA y el ex Ministerio de Agroindustria de la Nación -actual Ministerio de Agricultura, Ganadería y Pesca-.
Más información: Lucrecia [email protected]; Jorgelina Montoya [email protected]; María del Carmen [email protected]; Federico Elorza [email protected]
Cama biológica indirecta.

145Agosto 2020, Argentina 145Agosto 2020, Argentina
Construyen una biblioteca de nanoanticuerpos de llamas contra la COVID-19
BIOTECNOLOGÍA
POR CECILIE ESPERBENTFOTOS: INTA CASTELAR
Construyen una biblioteca de nanoanticuerpos de llamas contra la COVID-19Se trata del primer reservorio elaborado en la Argentina y que contiene información sobre los anticuerpos que producen estos camélidos frente al SARS-CoV-2. Con la información obtenida de la biblioteca se podrán obtener nanoanticuerpos con la capacidad de inhibir la infección viral provocada por el coronavirus-2. Fue lograda gracias al trabajo de investigadores del INTA y del Instituto Milstein junto con consorcios nacionales e internacionales.
Identificado como una de las maquina-rias biológicas más refinadas que exis-ten, el sistema inmunológico del ser hu-mano es la principal barrera de defensa frente al ataque de un microorganismo –bacteria, parásito o virus–. Sin embar-go, este sofisticado escudo natural no siempre responde a tiempo. De hecho, frente al SARS-CoV-2 está en desventa-ja debido a que mientras el virus puede hacer hasta 100.000 copias de sí mismo en 24 horas, el sistema inmune tarda en-tre 15 y 21 días en proteger al cuerpo de la enfermedad.
Para ayudarlo, un equipo de investiga-dores –integrado por especialistas del INTA y del Instituto de Ciencia y Tecnolo-
gía “Dr. César Milstein”, en colaboración con el Instituto Nacional para la Salud (NIH), de los Estados Unidos y el Conse-jo Nacional de Investigación de Canadá (NRCC)– construyó la primera biblioteca de nanoanticuerpos de llamas o VHH, un reservorio genético que permitirá selec-cionar los anticuerpos que generan es-tos camélidos cuando son expuestos al SARS-CoV-2 y que poseen la capacidad de neutralizar la infección viral.
“Gracias a un gran equipo, integrado por Itatí Ibañez, Florencia Pavan e inves-tigadores de INCUINTA, pudimos cons-truir la biblioteca de los VHH en solo 10 días”, afirmó eufórica Viviana Parreño, coordinadora científica de INCUINTA del
INTA, y agregó: “Para nosotros esto es una hazaña, que nos llena de orgullo y demuestra lo importante que es el traba-jo interinstitucional”.
El tratamiento con nanoanticuerpos de llamas es una de las alternativas más prometedoras para enfrentar la pande-mia. Para producir los nanoanticuerpos, los investigadores inmunizaron a Spike –nombre de la llama– con la proteína que forma la corona del SARS-CoV-2; luego de cuatro dosis de vacunas, extrajeron una muestra de sangre y, de allí, los linfo-citos circulantes. A partir de esas células se purificó el ARN –ácido ribonucleico– mensajero, que contiene información de los anticuerpos que elabora el camélido.

146 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2146 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2
Construyen una biblioteca de nanoanticuerpos de llamas contra la COVID-19
“Buscamos los VHHque puedan neutralizarla infección causada por el SARS-CoV-2” (M. Bok).
“Toda esa información codificada sirve para armar un gran archivo de genes que se usará para seleccionar los nanoan-ticuerpos que están dirigidos contra el antígeno de interés”, indicó Itatí Ibañez –viróloga molecular del Instituto Milstein-CONICET y especialista en la construc-ción de este tipo de bibliotecas–.
Con la información codificada en los genes VHH se pueden obtener anticuer-pos monoclonales que servirán para inhibir una infección viral o como reac-tivos dentro de un método diagnóstico para detectar el virus. En general, estos nanoanticuerpos se utilizan para el diag-nóstico de una enfermedad, entre otras aplicaciones.
“Buscamos los VHH que puedan neutralizar la infección causada por el SARS-CoV-2”, expresó Marina Bok –in-vestigadora de INCUINTA y una de las especialistas que trabaja en el armado de la biblioteca– y agregó: “Esos nano-bodies servirán como fármaco preventi-vo o terapéutico. Nuestro objetivo es que
sirvan para una terapia de inmunidad pasiva complementaria a la vacunación”.
Una aguja en un pajarEncontrar e identificar un gen en un
genoma es una tarea compleja y, mu-chas veces, se asocia con un objetivo muy difícil de cumplir. Sin embargo, las librerías génicas están diseñadas para guardar y ordenar grandes volúmenes de información sobre el material gené-tico de interés.
De acuerdo con Bok, los nanoanticuer-pos son moléculas muy pequeñas y para poder producirlos es necesario obtener todo el repertorio de genes que codifican para los anticuerpos que la llama posee, a ese repertorio se lo conoce como bi-blioteca de genes. “De ese gran ‘archivo’ se seleccionan los nanobodies específi-cos que neutralizan al virus”, señaló.
En este sentido, Ibañez agregó que “la biblioteca de genes debe representar lo mejor posible el repertorio de anticuer-
pos que posee una llama, inmunizada con el antígeno de interés, para mejorar las chances de encontrar aquellos con las propiedades deseadas”.
Por esto, es importante que la bibliote-ca tenga un buen tamaño y una alta va-riabilidad. De este modo, “nos asegura-mos que la información que contiene sea de buena calidad”, afirmó Ibañez quien advirtió que el procedimiento para la ge-neración de una biblioteca implica varios pasos y puede llevar varias semanas (el tiempo depende de los reactivos dispo-nibles y de los problemas técnicos que puedan surgir).
Cómo construir una librería de genes
Para construir la biblioteca de nanoanticuerpos de llamas, el equipo de investigadores utilizó bacterias (E. coli) que se infectan con un virus (fago helper). Durante este proceso, los fagos se enfrentan al antígeno (en este caso proteínas del virus SARS-CoV-2) y se seleccionan los que lo recono-cen. Luego, se infectan nuevas bacterias con los fagos que recono-cieron las proteínas del virus. Este procedimiento se repite tres veces seguidas para aumentar las chances de encontrar nanoanticuerpos específicos. Finalmente, se seleccionan las bacterias que contienen el gen del nanoanticuerpo que neutraliza la proteína del SARS-CoV-2, se las mul-tiplica en placas con medio de crecimiento y se seleccionan las colonias de bacterias que poseen los distintos genes que codifican para distintos nanoanticuerpos.
“Pudimos construir la biblioteca de los VHH en solo 10 días” (V. Parreño).

147Agosto 2020, Argentina 147Agosto 2020, Argentina
Construyen una biblioteca de nanoanticuerpos de llamas contra la COVID-19
La implementación de un tratamiento con
nanoanticuerpos de llamas (VHH) es una
alternativa prometedora para enfrentar
la pandemia.
Cómo ayudarán los VHH al sistema inmunológico
Ahora mismo, científicos de todo el mundo trabajan en el laboratorio día y noche en busca de estrategias que ayuden a luchar y combatir al coronavirus-2. En términos generales, los virus no son fáciles de combatir. Esto se debe, sobre todo, a su naturaleza: están constituidos por genes que contienen ácidos nucleicos que forman moléculas largas de ADN o ARN, rodeadas de proteínas. El SARS-CoV-2 está integrado por una única molécula de ARN (ácido ribonucleico), es de cadena simple y de polari-dad positiva.Básicamente, cada molécula de virus posee las instrucciones necesarias para replicarse a sí misma. De este modo, al infectar una célula, la usa para multiplicarse y propagarse rápidamente. Durante este proceso, las células se deterioran y eso provoca la enfermedad. A par, el sistema inmunológico se activa y se pone en funcionamiento para identificar al intruso y neutralizarlo. Cada virus posee una “huella” característica, llamada antígeno, que per-mite distinguirlo de otros. Así, le sirve al sistema inmunológico que bus-ca, mediante los anticuerpos, cualquier cosa que contenga ese antígeno para destruir al virus o neutralizarlo. Todo este proceso se desarrolla en dos semanas aproximadamente, mientras que el virus puede hacer hasta 100.000 copias de sí mismo en solo 24 horas.Por esto, la implementación de un tratamiento con nanoanticuerpos de llamas (VHH) es una alternativa prometedora para enfrentar la pande-mia. Si bien no evitan la enfermedad, pueden minimizar los efectos.Por su pequeñez y capacidad de escabullirse, los VHH reconocen la parte interna del virus y pueden neutralizarlo. De este modo, se detiene la infección.
“La biblioteca de genes debe representar lo
mejor posible el repertorio de anticuerpos
que posee una llama” (I. Ibañez).
ronavirus-2, el equipo de investigadoras avanza con el análisis de la secuencia que está realizando Andrea Puebla, del servicio de secuenciación del INTA, y pa-ralelamente comienza la expresión y pu-rificación de los potenciales candidatos para luego realizar las pruebas in vitro que servirán para caracterizar funcional-mente a los nanoanticuerpos.
Asimismo, Parreño afirmó que “junto con Elsa Baumeister –del Servicio de Vi-rosis Respiratorias del Instituto Nacional de Enfermedades Infecciosas (INEI) de la ANLIS-Malbrán– esperamos realizar pronto los ensayos de neutralización in vitro del virus”.
“Anhelamos contar con uno o varios anticuerpos monoclonales, de fácil pro-ducción y purificación, que serían una herramienta clave para el tratamiento de pacientes en estado avanzado y/o con enfermedades de base”, insistió Parreño.
La sinergia lograda entre las institu-ciones y los investigadores hace que se pueda tener una respuesta excepcional-mente rápida frente a esta situación de emergencia. “Este no es el trabajo de un solo investigador, sino que es el resulta-do de la articulación, el compromiso y la labor en equipo de Itatí Ibáñez, Marina Bok, Florencia Pavan, Juan Pablo Malito, Gisela Marcoppido, Diego Franco y Laura López –apoyo fundamental en limpieza y en esterilizado–; Andrés Wigdorovitz, Celina Vega y Laura Crispino –apoyo en compras y en logística–”, resaltó.
Más información: Viviana Parreño [email protected]; Marina Bok [email protected]; Itatí Ibañez [email protected]
Sin embargo, más allá de la inmuniza-ción del animal, para obtener los frag-mentos de VHH es necesario realizar un biopaneo. Con esta técnica, “se pueden identificar los nanoanticuerpos deseados que servirán para desarrollar un trata-miento preventivo”, sumó Ibañez.
Con la biblioteca armada y con varios clones seleccionados capaces de reco-nocer y unirse a la proteína Spike del co-

148 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2148 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2
“Hay muy pocas instituciones en el mundo como el INTA”
INSTITUCIONAL
POR MARIO MIGLIORATI FOTOS: INTA
“Hay muy pocas instituciones en el mundo como el INTA”La afirmación es de Carlos Parera, director nacional del INTA, quien repasó los primeros meses de su gestión en un INTA atravesado por las reglas que impone la COVID-19. Destacó la labor desarrollada por la institución para adaptarse a una nueva realidad y “dar respuesta a los problemas que tenemos como país”.
“Fue y es un desafío enorme”, afirmó Carlos Parera para referirse al trabajo realizado por el INTA. Después que la Organización Mundial de la Salud (OMS) evaluara –el pasado 11 de marzo– que la COVID-19 podía ser caracterizada como una pandemia y que el Poder Ejecutivo Nacional decretara el aislamiento social, preventivo y obligatorio (ASPO) para todo el territorio nacional, “se presentó un nuevo marco para dar respuestas como INTA en todo el país”, apuntó.
En los 9 meses transcurridos desde que asumió como director nacional y con la función de coordinar, planificar y ges-
tionar las actividades del instituto duran-te los próximos cuatro años, reconoció la importancia de dar continuidad a la agen-da del organismo donde fueron clave los acuerdos con municipios y organizacio-nes de productores para responder a las demandas en el territorio.
El INTA se organizó con el objeto de mantener el nivel operativo y colaborar con distintas instituciones del país y del extranjero para enfrentar la grave crisis sanitaria con aportes a la salud pública, el sector productivo y la educación. “Y se pudo cumplir con los objetivos pauta-dos”, explicó Parera, ingeniero agrónomo
egresado de la Universidad Nacional de Cuyo y con un doctorado de la Universi-dad de Florida –Estados Unidos–.
Parera destacó la reacción para ayudar a “los pequeños y medianos productores a organizarse” y ponderó “el rol de los consejos locales y regionales”, cuya par-ticipación en el funcionamiento institucio-nal la define como: “una de las grandes fortalezas de INTA”.
Mendocino y vinculado a la investiga-ción en temas de fisiología vegetal y el uso de los recursos naturales, ingresó al INTA como becario en la Agencia de Ex-tensión Rural La Consulta –Mendoza–,

149Agosto 2020, Argentina 149Agosto 2020, Argentina
“Hay muy pocas instituciones en el mundo como el INTA”
en 1980. Se desempeñó como director de la Estación Experimental Agropecua-ria San Juan, desde 1992 hasta 2003, y luego como director del Centro Regional Mendoza – San Juan, desde 2003 has-ta 2014. En 2019, tras tomar el cargo de director nacional, expresó que se trata-ba de “una oportunidad para contribuir y aportar al desarrollo nacional”.
La situación generada por la pan-demia del COVID-19 llevó a los orga-nismos del Estado a reorganizarse en muy poco tiempo para seguir cum-pliendo sus funciones ¿Cómo fue el trabajo desarrollado por el INTA?
En el marco de la pandemia por CO-VID-19 tuvimos que reconfigurar la insti-tución. Esto nos llevó a trabajar en dife-rentes planos. Uno de ellos respondió al funcionamiento interno relacionado con las rutinas de trabajo y de funcionamien-to que se daban en el INTA. Tuvimos que cambiar rutinas rápidamente para adap-tarnos a las formas que indicaba el ais-lamiento social, preventivo y obligatorio (ASPO), así como también al modo en que fue evolucionando la situación. Es decir, nos encontró en un momento ins-titucional donde contamos con procesos informatizados, lo cual nos ayudó, y con personal capacitado. Este hecho no es menor y resultó, si nos observamos fren-te al desempeño de otras instituciones, en una ventaja.
Además, nos permitió responder rá-pidamente sin que se detuviera ningún
proceso. Por el contrario, en el segundo trimestre de este año tuvimos un nivel de ejecución presupuestario histórico en un mes, lo que no se puede alcanzar si no se tienen bien coordinados los mecanis-mos y si no se cuenta con las capacida-des. Son aspectos que resalto y destaco de manera significativa.
Sin duda esta capacidad demostra-da es el resultado de la planificación y gestión en los últimos años desde la Dirección Nacional…
Sí, por supuesto. Y es también una vi-sión institucional para anticiparse o ir en línea en cómo avanza la disponibilidad tecnológica. En este sentido, el INTA armó sus programas a partir de generar el software propio, adaptarlo o adquirirlo, permitiéndonos contar en este momento con las áreas administrativas informati-zadas. Otros aportes surgieron desde el gobierno nacional, al informatizar proce-sos que no resultaron menores en este contexto para el funcionamiento de la administración pública.
¿Qué sucedió con la presencia del INTA en el territorio?
Con respecto al todo el trabajo externo realizado por la institución, se presen-taron distintas realidades. La pandemia nos encontró en un momento que po-dríamos señalar como crítico en cuanto a la operatoria institucional, sobre todo en el tema de extensión y en lo que ha-
cía a la distribución de semillas de la campaña otoño-invierno del programa ProHuerta en el territorio. Normalmente la distribución de semillas se realiza de persona a persona y a grupos con mu-cho contacto, ya que es la dinámica de la extensión. Este nuevo contexto obligó a repensar las estrategias para la distri-bución de las semillas y llevó a tener que asociarnos con municipios y organiza-ciones de productores. Es decir, generar distintas instancias que, más allá de esta situación, permitieron que la totalidad de las semillas fueran distribuidas.
Es una tarea importante en cuanto a la situación de pobreza que se está vi-viendo y que se va a vivir. Y esta es una herramienta que estamos ofreciendo a nivel institucional junto con el Ministerio de Desarrollo Social de la Nación como aporte a la lucha contra la pobreza y con-tra el hambre en Argentina. Desde ese punto de vista hemos podido cumplir con este programa.
También se brindó apoyo en estrate-gias de comercialización a pequeños productores…
Si bien ya se trabajaba y contábamos con experiencias en todo el país, hubo una importante reacción para ayudar a los pequeños y medianos productores a organizarse en la comercialización de la producción de alimentos destinados a los consumidores. El INTA participó de inicia-tivas para que pudieran ofrecer bolsones de verduras a precios accesibles, esto en
“Tenemos la responsabilidad
institucional de apoyar a la producción
de alimentos que presenta diferentes
aristas”.
“Hubo una importante reacción para ayudar
a los pequeños y medianos productores
a organizarse en la comercialización”.

150 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2150 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2
cuanto a la necesidad de avanzar en cir-cuitos cortos de comercialización.
Al asumir el cargo de director usted remarcaba: “El INTA tiene que adap-tarse a esta realidad para dar respues-ta con tecnologías e innovación a fin de resolver los problemas reales de la sociedad” ¿Cómo se trabajó en este tiempo para guiar la gestión y brindar respuestas frente a estas demandas? En un escenario actual que era impo-sible prever en ese momento.
Esta era una demanda de la sociedad y le dimos respuesta rápidamente, ¡lo remarco! A nivel interno hicimos ajustes para dar respuesta y, entre otras accio-nes, se conformó un comité de crisis. Se trabajó en la reorganización institucional que no estaba prevista antes del mes de marzo de este año.
También comenzamos a observar la necesidad de trabajar en temas estruc-turales del sector y para el país. En este sentido, trabajamos en muchos frentes en términos del tema sustentabilidad.
Esto en cuanto a iniciativas de leyes como la de protección de humedales y fitosanitarios junto con otras que se en-cuentran en la agenda legislativa y que son centrales para el país. Como no nos quedamos quietos, trabajamos con gru-pos técnicos desde el rol institucional de ofrecer información científica probada y evaluada a partir de sumar la labor de los mejores profesionales que integran el INTA. Es una tarea importante acompa-ñar al Estado nacional y al poder legis-lativo con información técnica y científica probada.
En el INTA comienzan a generarse ámbitos desde la Dirección Nacional para expresar opiniones y miradas desde la investigación, desarrollo e innovación que permiten reconocer que conviven distintas visiones so-bre, por ejemplo, los modelos pro-ductivos ¿Estos espacios apuntan a saldar debates o buscan hallar pun-tos de encuentro y formas de poder convivir?
El INTA es un organismo donde la cien-cia y la tecnología constituyen uno de los ejes importantes del funcionamiento institucional. En este sentido, no estoy de acuerdo en que haya posiciones ho-mogéneas porque resulta muy difícil ya que somos unas 7000 personas que con-formamos la institución. Pero considero que el desafío es acordar en los temas centrales, es decir: acordar, pero no fijar posición. Esto implica considerar 4 o 5 ejes en común a los que el INTA no re-
nuncie, cualquiera sea la visión o la idea que represente a algún grupo, ya sea por cuestiones ideológicas o porque lo ha comprobado.
Está el caso de la producción agroeco-lógica con cero intervenciones y la pro-ducción industrial que representan los dos extremos. Recientemente hicimos un seminario donde tomamos el tema y lo pusimos a discusión y lo que vimos es que vamos acercando posiciones. Los equilibrios son importantes. Tenemos la responsabilidad institucional de apoyar a la producción de alimentos que presenta diferentes aristas y cualquier interven-ción para producir va a generar un dis-turbio en ese ambiente. Lo importante en esto es lograr el equilibrio, al asegurar la producción, la disponibilidad de alimen-tos y materias primas para la agroindus-tria con el menor impacto posible sobre el ambiente. Este es el gran desafío técnico que tiene el INTA en este momento.
¿Cómo es el diálogo con las organi-zaciones del campo? Con las que tie-nen representación en el Consejo Di-rectivo del INTA y aquellas vinculadas a la agricultura familiar.
Con las organizaciones tradicionales del sector productivo que forman parte del Consejo Directivo del INTA no solo nos reunimos en ese espacio, sino que mantenemos reuniones sectoriales. Con-formamos agenda sobre temas en co-mún, pero también estamos empezando a establecer lazos con otras organizacio-nes que no son parte formal del Consejo
“Estamos restableciendo estas relaciones y ampliando la base de participación que, a mi entender, es la clave del INTA”.
“Hay muy pocas instituciones en el mundo como el INTA”
De izq. a der.: Susana Mirassou, presidenta del INTA, Diana Guillén, jefa de gabinete del Mi-nisterio de Agricultura, Ganadería y Pesca de la Nación, y Carlos Parera, director nacional.

151Agosto 2020, Argentina 151Agosto 2020, Argentina
Directivo pero que, en muchos casos, representan en las regiones a un alto nú-mero de productores o cuentan con un número importante de productores aso-ciados. Con estas estamos recreando la relación como, por ejemplo, la Asociación Argentina de Productores Algodoneros que no está sentada en el consejo, pero cuenta con una importante representati-vidad en el NEA –nordeste– del país. Y también con las organizaciones sociales como la UTT –Unión de Trabajadores de la Tierra– y otras con presencia de pe-queños productores. Y si bien estaban un poco dormidas, estamos restableciendo estas relaciones y ampliando la base de participación que, a mi entender, es la clave del INTA. Ampliar la base de par-ticipación implica no quedarnos con un solo grupo o una visión única.
El ProHuerta acaba de cumplir 30 años, ¿qué desafíos tiene por delante el INTA junto con el Ministerio de De-sarrollo Social en temas de seguridad alimentaria?
Me interesa hacer una aclaración, ya que el INTA trabajaba en ProHuerta antes de que existiera como tal. Era un programa muy similar que no llevaba ese nombre, pero que formaba parte de una tradición y se lo conocía como Hogar Ru-ral, un área que era parte de las activida-des de extensión.
En este sentido, el ProHuerta se con-solidó en el INTA y en el país con alti-bajos, valles y picos, con momentos muy cambiantes dado que siempre tuvo un
importante componente político. En este momento, con el Ministerio de Desarro-llo Social de la Nación nos encontramos ajustando una propuesta de ProHuerta 2021. Consideramos con las autorida-des del ministerio que es necesario ha-cer una revisión y realizar ajustes, de ahí que en este último trimestre hemos mantenido reuniones donde participó la presidenta del INTA, Susana Mirassou, y la responsable de la Dirección Nacional Asistente de Transferencia y Extensión, María Rosa Scala, que tiene a cargo la ejecución del programa. El objetivo es plantear un ProHuerta en función de las experiencias que reconocemos para que las cosas que están bien, se continúen, las que hay que mejorar, se ordenen y, en cuanto a las ideas nuevas, ver qué traen.
Los aportes del INTA en investi-gación y desarrollo son relevantes y reconocidos. En estos meses de pandemia se demostró la capacidad para dar respuestas en investigación, por ejemplo, en cuanto a trabajar con productos biológicos contra el SARS-CoV-2 o sumarse a la red de diagnós-tico de Covid-19, y, al mismo tiempo, no dejar de estar con el productor en el territorio ¿Qué es lo que viene en investigación desde el trabajo de coo-peración y bajo el concepto de Una Salud?
Esta idea de Una Salud es el desafío que tiene el país y el INTA como un ac-tor central en este tema. La respuesta y
“Esta idea de Una Salud es el desafío que tiene el país y el INTA como
un actor central en este tema”.
capacidad demostrada por la institución con aportes al sistema de salud fue in-mediata. Y no se trató de una respuesta “caprichosa”. Son años de inversión y formación y, en consecuencia, cuando se habla de ciencia y tecnología no se debe pensar en un corto plazo, sino a largo plazo. En su momento se pensó así y por eso contamos con las capacidades del personal y de la infraestructura. Por-que en algún momento se invirtió y vio que esto era un tema que a futuro iba a tener algún impacto. Esto nos permitió sumarnos rápidamente con aportes en el diagnóstico y que no es posible si no se cuenta con las instalaciones, la tecnolo-gía y las capacidades del personal. Rea-lizando determinaciones para el diag-nóstico de COVID-19 para la provincia de Buenos Aires desde el INTA Castelar, para Córdoba, desde la Estación Experi-mental Agropecuaria de Marcos Juárez, y recientemente se integró Entre Ríos, donde la estación experimental de Con-cepción del Uruguay empezará a anali-zar muestras.
Lo importante de destinar y sostener la inversión en ciencia y tecnología para el país…
Es consecuencia de inversión y de haber planificado la formación de inves-tigadores. Con respecto a la repuesta brindada, hemos logrado financiamiento para proyectos de la Agencia de Promo-ción de la Investigación, el Desarrollo y la Innovación en la convocatoria “Ideas Proyecto COVID-19”. Entre los proyectos financiados se encuentra el que lidera la investigadora Viviana Parreño y que está orientado a la obtención de nanoan-ticuerpos monoclonales recombinantes derivados de llamas y anticuerpos poli-clonales, derivados de la yema de huevo, para el diagnóstico y el tratamiento pre-ventivo y terapéutico de COVID-19. Una investigación que se venía realizando y que se adaptó al tema de COVID-19, por lo que fue posible trabajar en brindar una respuesta. Estos proyectos se suman a otros 6 proyectos que desde distintas unidades del país lograron financiamien-
“Hay muy pocas instituciones en el mundo como el INTA”

152 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2152 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2
to de la agencia en la convocatoria: “Pro-grama de articulación y fortalecimiento federal de las capacidades en ciencia y tecnología COVID-19”. También hay otras iniciativas con instituciones y em-presas interesadas en desarrollos en esta línea.
Como ha sucedido en el último tiem-po, a esta altura ya se habla del pre-supuesto del próximo año, ¿qué previ-sión y expectativas tiene el INTA para el período 2021?
Como INTA hemos elevado una pro-puesta para el armado del presupuesto. Contamos con la aprobación del Ministe-rio de Agricultura y estamos haciendo los presupuestos basados en demandas. En estas se incorporó el relevamiento de in-fraestructura, donde tenemos algún défi-cit, e hicimos el pedido de las partidas 2 y 3, las más operativas, en función de los proyectos que actualmente tenemos en ejecución. En esta partida se contempla el crecimiento de la cartera con los pro-yectos locales que abordan problemas y oportunidades de localización geográfica acotada no contemplados en el proceso de priorización a nivel nacional, así como también algunos proyectos eventuales que puedan llegar a salir.
Lo solicitado está dentro de las pre-visiones que requerimos para funcio-namiento y necesidades. Quizás sea el inciso 4 donde se tendría que haber discutido más, ya que queríamos darles un impulso fuerte a algunas inversiones, por ejemplo: vehículos, equipamiento de laboratorio y construcciones. Estoy espe-ranzado en que vayamos a tener un pre-supuesto acorde con lo que necesitamos.
¿Cómo viene construyendo el vín-culo con el Ministerio de Agricultura, Ganadería y Pesca de la Nación?
Tenemos una muy buena relación en cuanto a equipos de trabajo. Estamos coordinando con la Secretaría de Agricul-tura Familiar, Campesina e Indígena en diversas actividades. Nos integramos en proyectos con UNICEF, en un proyecto de microcréditos. Con el ministerio avan-zamos en diversos frentes desarrollando acciones de manera conjunta y con mu-cha expectativa de continuar colaboran-do. Lo que observo es una vocación de pensar al INTA desde sus capacidades en potenciales políticas públicas que pueda generar el ministerio.
En este tiempo transcurrido en el INTA, ¿cuál es la valoración que hace del organismo?
Lo que quiero señalar acerca del INTA trasciende la manera en la que me he criado y formado profesionalmente en esta institución. Hay muy pocas institu-ciones en el mundo como el INTA. He tenido la suerte de visitar e interactuar en muchas organizaciones similares, pero son pocas las que se le parecen. Por-que cuenta con componentes que son centrales en el ADN institucional que la hacen única.
Le otorga un lugar destacado a la participación en los consejos locales y regionales.
Lo primero es la participación del sec-tor agropecuario. Si bien habría que hacer algunos ajustes, es fundamental que el sector participe. Contamos apro-
ximadamente con 3000 personas que forman parte de los consejos locales y regionales en todo el país de manera ad honorem y realizan aportes relevan-tes a la institución. Los considero muy importantes porque no permiten que nos vayamos de contexto, sino que nos mantienen atados a la realidad. Por esto es fundamental y prioritario el fortaleci-miento de esta herramienta de control social.
Lo segundo es la capacidad que hemos logrado para readaptarnos a situacio-nes y nuevos desafíos. En esto somos un ejemplo. No somos una institución anquilosada, sino que somos capaces de armar una cartera de proyectos con una nueva visión basada en problemas y oportunidades. Generamos una estructu-ra programática que no estaba y que son las plataformas de innovación territorial. Resultan una importante herramienta para identificar y priorizar problemas y oportunidades que aportan a la innova-ción regional a corto, mediano y largo plazo. Y si en el lapso de un tiempo, sea porque cambió el contexto, no funciona más, podemos armar otra porque conta-mos con la capacidad para hacerlo.
El tercer aspecto es el personal. El INTA cuenta con un nivel de formación y capacitación que hace a la diferencia. Es lo que no queremos perder y vamos a trabajar en la nueva visión institucional, es decir, continuar con el fortalecimiento de la capacitación del personal del INTA.
Más información: Carlos Parera [email protected]
“Estoy esperanzado en que vayamos a tener un presupuesto acorde con lo que necesitamos”.
“Hay muy pocas instituciones en el mundo como el INTA”

153Agosto 2020, Argentina 153Agosto 2020, Argentina
CIENCIA JOVEN
POR MARIO MIGLIORATIFOTOS: GENTILEZA INVESTIGADORA
“El desafío es generar herramientas que lleguen al productor”Así describe el objetivo que persigue en investigación Yésica Chazarreta, genetista e integrante del equipo de Ecofisiología de Cultivos del INTA Pergamino –Buenos Aires-. Estudia el impacto de la fecha de siembra del maíz sobre el llenado, el secado y la calidad de los granos, lo que ya le valió ser reconocida en Latinoamérica como investigadora joven en innovación del cultivo.
Del campo a la ciudad es una manera de describir un día de esta profesional que realiza diversos ensayos con híbri-dos de maíz en Pergamino, centro de la zona núcleo de la producción agrícola bonaerense. Al referirse con pasión so-bre sus experimentos de campo, Yésica Chazarreta (25) explica que su deseo de estudiar Licenciatura en Genética, en la Universidad Nacional del Noroeste de la Provincia de Buenos Aires (UNNOBA), surge después de terminar la escuela agrotécnica -en esa misma localidad- donde se despierta su interés. Esta disci-plina dentro de la biología le abrió un am-plio espectro de aplicación para abordar diversas temáticas de la investigación científica, aunque eligió especializarse en la rama de la genética vegetal.
Durante la carrera de grado realizó pa-santías en empresas de la región y co-misión de estudios en el Grupo Mejora-miento de Maíz del INTA Pergamino. En el año 2017, en esa estación experimen-tal inició los primeros muestreos para la tesis de grado en el cultivo de maíz. Se integró al trabajo de otros becarios de la unidad enfocada en el llenado y secado de los granos de este cereal. Esta línea de investigación le permitió estudiar la evolución del peso seco y del contenido de humedad, comparando los maíces de fecha de siembra temprana y tardía. No solo dando lugar a sus tesis de grado, sino que le permitió participar junto con María Elena Otegui, investigadora del INTA - CONICET y FAUBA, y Juan Igna-cio Amas, becario doctoral del CONICET en el INTA Pergamino, en un convenio público-privado para desarrollar esta lí-nea de investigación en otros materiales.
Actualmente, como becaria del CO-NICET y con un doctorado en curso en Ciencias Agropecuarias en la Facultad de Agronomía de la UBA, investiga acerca del control genético y ambiental de los determinantes fisiológicos del llenado, se-cado y calidad de granos de maíz en ge-notipos destinados para grano o ensilaje.
¿Cuándo surge la decisión de ser investigadora?
Mi interés sobre los cultivos surge du-rante mis estudios en la escuela agrotéc-
nica, un ámbito en el que nos habíamos vinculado y, una vez finalizados, me per-mitieron anotarme en la Licenciatura en Genética. Pero la realidad es que des-conocía qué hace un especialista en el tema. Uno se imagina que desarrollan aspectos más vinculados con prácticas de laboratorio. También desconocía cómo era el funcionamiento del sistema científico en el país. En el transcurso de la carrera me fui interiorizando, sobre todo en cuanto a cómo hacer para in-gresar al sistema. Vale señalar que me enteré bastante tarde sobre su funcio-
“El desafío es generar herramientas que lleguen al productor”

154 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2154 ACTUALIDAD EN I+D RIA / Vol. 46 / N.º 2
namiento, en lo que hace a las becas, escribir un proyecto, o, cómo encontrar un grupo de trabajo.
Instancias de la profesionalización y de la investigación de las que poco se habla…
Sí, porque cuando uno se presenta a una beca tiene que contar con un grupo de trabajo, es decir, un lugar de asiento físico definido, un director y un proyecto. El desconocimiento de estos aspectos demanda un saber del que, hasta ese momento, donde se empieza a definir el futuro profesional, no se cuenta con información.
Fuiste premiada internacionalmente en tu área de estudio por la innovación en maíz, ¿qué significa este logro?
Se trató del premio de Innovación Ju-venil en Maíz 2019 - América Latina, a la que postulamos con el grupo de investi-gación de Ecofisiología de Cultivos. Los premios tienen como objetivo identificar a mujeres y hombres jóvenes innova-dores que pueden servir para inspirar a otros a involucrarse en los sistemas agroalimentarios basados en el maíz. Así como también crear una plataforma que permita a jóvenes innovadores de todo el mundo establecer contactos y
compartir sus experiencias. Mi tema de presentación se relacionó con los resul-tados obtenidos en mi tesis de grado y en los experimentos que actualmente lle-vamos a cabo para mi doctorado.
¿Qué te implica este reconocimiento como investigadora?
Fue un premio inesperado, ya que no vengo de una formación agronómica, pero a su vez es muy valioso en lo perso-nal para poder mostrar la investigación que desarrollamos, poder contarlo y que este conocimiento genere un intercam-bio positivo de otras voces que también pueden aportar ideas valiosas al tema. Se trata centralmente, más allá del re-conocimiento, de un trabajo en conjunto
con otros investigadores del INTA Perga-mino, entre los que se encuentran María Elena Otegui (directora de tesis), Alfredo Cirilo (director de la beca FONCyT que tuve al inicio de mi doctorado), y Juan Ig-nacio Amas, con quien he compartido los ensayos a campo.
¿Cuál es tu percepción de la inves-tigación en esta línea de desarrollo?, teniendo en cuenta lo que te permitió este intercambio con otros jóvenes de América Latina
En mi opinión, los investigadores ar-gentinos son valorados en el continente, algo que se pudo ver en la XXIII Reunión Latinoamericana del Maíz de la que par-ticipé en Colombia, durante la premia-
Un reconocimiento a la innovación
En el año 2019, Yésica Chazarreta recibió el premio de Innovación Juvenil en Maíz 2019 de América Latina. Esta distinción lo impulsó el Programa de Investigación de Maíz del CGIAR, una asociación de investigación global para un futuro con seguridad alimentaria, a fin de promover la participación de mujeres y hombres jóvenes en los siste-mas agroalimentarios basados en el maíz. Un reconocimiento a las contribuciones de jóvenes menores de 35 años que están implementando innovaciones en los sistemas agroalimenta-rios basados en el maíz de América Latina, que incluyen investigación para el desarrollo, sistemas de semillas, agroindustria e intensificación sustentable.La premiación fue en el marco de la XXIII Reunión Latinoamericana del Maíz y el IV Congreso de Semillas realizado en Colombia, organizada por la Asociación Colombiana de Semillas y Biotecnología (Acosemillas) junto con la Corporación Colombiana de Investigación Agropecuaria (Agrosavia), el Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT) y la Federación Nacional de Cultivadores de Cereales y Leguminosas (Fenalce).
“En mi opinión, los investigadoresargentinos son valorados en el continente”.
“El desafío es generar herramientas que lleguen al productor”

155Agosto 2020, Argentina 155Agosto 2020, Argentina
ción. Lo que se destaca es la forma de producción de este cereal en la Argenti-na, ya sea en la forma y escala como en los usos respecto de otros países de la región. Se valora contar con profesiona-les argentinos en los grupos de trabajo. De hecho, en el Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT, según su sigla en inglés) cuentan con pasantes y becarios argentinos que rea-lizan allí sus experiencias. Este interés por los investigadores nacionales es algo que se percibe en América Latina como en el resto del mundo.
Y respecto del lugar de la mujer en la ciencia…
Desde hace algunos años se ha puesto en discusión el lugar de la mujer en la sociedad y esto muy importante. Es muy positivo que estos debates ocurran. En muchos lugares del mundo, las mujeres aún siguen ocupando minoritariamente puestos jerárquicos y relacionados con la ciencia. Afortunadamente, en Argenti-na se vive un clima de gran paridad de género en el ámbito científico. Es impor-tante que los estereotipos de género vin-culados con el imaginario común de un profesional de la agronomía o un científi-co sean derribados.
¿Qué tan importante fue contar con una universidad en tu región?
Lo primero, decir que soy de las pri-meras profesionales en mi familia. En segundo lugar, destacar que la UNNO-BA cuenta con una beca destinada al primer estudiante universitario de una
familia que sea egresado de una escue-la agrotécnica. Una iniciativa de la cual participé en contarles a otros jóvenes la posibilidad de esta beca. Que la univer-sidad esté mucho más cerca de la casa de uno, hace a que gran parte de los estudiantes sean primera generación de estudiantes universitarios en una familia. Es un impacto muy positivo para toda la sociedad, no solo para quien asiste sino para la comunidad que lo rodea. Este pa-saje por la universidad, más allá de las experiencias, nos cambia en la forma de percibir el mundo que nos rodea.
Respecto a hacer una proyección de tu futuro, ¿cómo y dónde te imaginas?
Estoy cursando el doctorado y tengo por delante el desarrollo de varios expe-rimentos que culminarán con la escritura de la tesis, pero me veo capacitándome para los desafíos que vienen por delan-te. El conocimiento adquirido debe ser-virnos para que las investigaciones lle-guen a la sociedad. Al fin y al cabo, sus aportes son los que financian mis estu-dios. El desafío es generar herramientas que lleguen al productor.
En un cultivo como el maíz, donde se han logrado tantos avances, ¿en qué más se puede mejorar?
La realidad es que el maíz ha tenido muchos cambios a lo largo la historia, resta mucho por investigar y por hacer. Durante los últimos años en el país hubo un hecho que cambió la forma de produ-cir y que responde a sembrar más tarde, en lo que conocemos como maíz tardío.
En este sentido, y en la línea de inves-tigación en la que participo, pude obser-var que la evolución de la pérdida de humedad en los granos del maíz tardío, si bien seguía el mismo patrón que los maíces de fecha de siembra temprana, presentaban como aspecto de interés que la pérdida de humedad de los gra-nos se producía de una forma más lenta.
“Afortunadamente, en Argentina se vive un
clima de gran paridad de género en el ámbito
científico”.
“El conocimiento adquirido debe servirnos
para que las investigaciones lleguen
a la sociedad”.
“El desafío es generar herramientas que lleguen al productor”
Donde la humedad de comercialización del maíz en el país es del 14,5 por cien-to y esta humedad el cultivo la alcanza-ba en los meses de junio y julio. Esto le produce muchos problemas al productor porque tiene que mantener el cultivo en pie en el campo en una época donde las temperaturas son menores, hay menos radiación solar, más humedad con el consecuente aumento de la incidencia de enfermedades de espiga. Esto baja la calidad y genera la presencia de mico-toxinas, que pueden tener efectos noci-vos para la salud si se consume, como así también perdidas de rendimiento por quebrado y vuelco de plantas.
En cuanto a maíces tardíos, ¿cuáles son los aportes buscados en la tesis de doctorado?
Uno de los objetivos de la tesis es po-der generar, a partir de los experimentos realizados y los que llevamos a cabo ac-tualmente, modelos de predicción de la evolución de la humedad de los granos para poder contar con herramientas de predicción durante el momento de cose-cha; donde los productores puedan con-tar con esas herramientas para la toma de decisiones acerca de cuándo cose-char su maíz tardío.
Más información: Yésica Chazarreta [email protected]
“Este pasaje por la universidad, más allá
de las experiencias, nos cambia en la forma de
percibir el mundo que nos rodea”.

156
Caracterización de accesiones de garbanzo (Cicer arietinum L.) mediante descriptores morfo agronómicos cuantitativos en una colección (...)
RIA / Vol. 46 / N.º 2REVISIONES156156
Caracterización de accesiones de garbanzo (Cicer arietinum L.) mediante descriptores morfo agronómicos cuantitativos en una colección del Banco de Germoplasma de la Universidad Nacional de Córdoba
RESUMENEl garbanzo (Cicer arietinum L) es la segunda leguminosa más cultivada en el mundo. La producción na-
cional está basada en pocos cultivares (Norteño, Chañaritos S-156, Kiara UNC-INTA, Felipe UNC-INTA, TUC 403, TUC 464 y TUC G 470). Debido a que el área cultivada está creciendo resulta importante disponer de nuevos materiales genéticos mejor adaptados a diferentes condiciones agroecológicas. Los objetivos del presente trabajo fueron caracterizar diferentes accesiones de garbanzo mediante descriptores morfo agronó-micos cuantitativos, evaluar la variabilidad fenotípica de los materiales genéticos, determinar los descriptores que efectivamente contribuyen a la discriminación de las accesiones estudiadas y obtener una función canó-nica que permita clasificar nuevas accesiones según su origen.
Se estudiaron 93 accesiones de garbanzo de diferentes orígenes utilizando diferentes aproximaciones es-tadísticas uni y multivariadas. La caracterización de las accesiones se realizó sobre cinco plantas, evaluando 17 descriptores morfo agronómicos. Los resultados de los estadísticos simples revelaron que las variables número de vainas por planta y peso de 100 semillas presentaron valores altos de CV. Los componentes de peso y volumen de semillas resultaron asociarse positivamente entre sí, pero negativamente con el número de vainas por planta. En el gráfico del perfil multivariado los caracteres que permitieron la mayor diferencia-ción de las accesiones fueron número de semillas por planta, número de vainas por planta y rendimiento por planta. En el análisis de conglomerados se dio la conformación de tres grupos diferenciados por las variables pesos de 100 semillas, altura de planta, número de vainas y semillas por planta. En el análisis de componen-tes principales se encontró que las tres primeras componentes principales explicaron el 66% de la variación total. En el C1 las variables que mayor contribución realizaron a la separación de los tipos de garbanzo fueron: número de vainas y semillas por planta, tamaño de semillas, peso de 100 semillas y altura de planta. Mediante el análisis discriminante se lograron detectar las variables morfo agronómicas que mejor discriminan (número de vainas y semillas por planta) las accesiones. Este estudio permitió concluir que existe una considerable variabilidad en todos los caracteres estudiados. De este modo podría ser utilizada la información para introdu-cir germoplasma con mayor divergencia genética, seleccionando los progenitores con los mayores contrastes genéticos para los programas de mejora y ampliar la base genética para el garbanzo en Argentina.
Palabras clave: garbanzo, variabilidad, accesiones, descriptores, análisis multivariado.
1Universidad Nacional de Salta (UNSa), Facultad de Ciencias Naturales, Sede Central y Sede Metán-Rosario de la Frontera, Cátedra de Mejoramiento Genético Vegetal. Av. Bolivia, 5150 (4400), Salta, Argentina. Correo electrónico: [email protected] Nacional de Salta (UNSa), Facultad de Ciencias Naturales, Cátedra de Mejoramiento Genético Vegetal. 3Universidad Nacional de Córdoba (UNCo), Facultad de Ciencias Agrarias, Cátedra de Mejoramiento Genético Vegetal. 4Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Cerrillos, Salta. 5Universidad Nacional de Salta (UNSa), Facultad de Ciencias NaturalesCátedra de Genética.
Recibido 20 de noviembre de 2019 // Aceptado 13 de mayo de 2020 // Publicado online 21 de octubre de 2020
CASTILLO, V.1; COLLAVINO, N.2; FARFAN, E.2; CARRERAS, J.3; FEKETE, A.4; POCOVI, M.5

157
CASTILLO, V.; COLLAVINO, N.; FARFAN, E.; CARRERAS, J.; FEKETE, A.; POCOVI, M.
Agosto 2020, Argentina 157157
INTRODUCCIÓN
El garbanzo (Cicer arietinum L) es la segunda legumino-sa más cultivada en el mundo, desplazando a la arveja en 2016 (FAO, 2016). Se estima que en Argentina en el año 2018 se sembraron 110.000 hectáreas distribuidas principal-mente entre Córdoba (45.000 ha), Tucumán (20.000 ha) y Salta (15.000 ha), mientras que las provincias de Catamar-ca, Santa Fe y Buenos Aires sumaron 30.000 ha (Panorama EEOC, 2018). La producción nacional está basada en pocos cultivares de garbanzo (Norteño, Chañaritos S-156, Kiara UNC-INTA, Felipe UNC-INTA, TUC 403, TUC 464 y TUC G 470) con adaptabilidad amplia. Las variedades Chañaritos S-156 y Norteño ocupan la mayor parte de la superficie cul-tivada, aunque comenzaron a difundirse Kiara y Felipe, muy relacionadas genéticamente con Norteño.
Debido a que el área cultivada con garbanzo está creciendo de manera sostenida, sería importante dispo-ner de nuevos materiales mejor adaptados a diferentes condiciones agroecológicas tanto en áreas tradicionales como de expansión. La Universidad Nacional de Córdoba cuenta con una colección de germoplasma que incluye tanto materiales de centros internacionales (ICARDA; ICRISAT; IFAP-UCO) como variedades locales, líneas
y cultivares obtenidos por el propio programa de mejora del garbanzo desarrollado por la Universidad Nacional de Córdoba, INTA y Universidad Nacional de Salta. Conocer la variabilidad fenotípica de estos materiales es impor-tante para el diseño de futuros cruzamientos con fines específicos de mejora. La caracterización genética de las colecciones de germoplasma puede estimarse utilizando diferentes métodos que van desde los convencionales como el uso de listas de descriptores morfológicos, méto-dos bioquímicos y moleculares (Carvalho, 2004). En gar-banzo los descriptores morfológicos son muy utilizados para la descripción de los materiales genéticos, incluso la UPOV (Unión Internacional para la Protección de Obten-ciones Vegetales) acepta un total de 19 caracteres mor-fológicos para la descripción de variedades de garbanzo. Los objetivos del presente trabajo fueron caracterizar 93 accesiones de garbanzo con descriptores morfo agro-nómicos cuantitativos, evaluar la variabilidad fenotípica de los materiales genéticos, determinar los descriptores morfo agronómicos que efectivamente contribuyen a la discriminación de las accesiones estudiadas y obtener una función canónica que permita clasificar nuevas acce-siones según su origen.
ABSTRACT
Chickpea (Cicer arietinum L.) is the second most important cultivated legume in the world. The production of Argentina is based on few cultivars (Norteño, Chañaritos S-156, Kiara UNC-INTA, Felipe UNC-INTA, TUC 403, TUC 464 and TUC G 470). As the cultivated area is increasing, it would be important to select new genetic materials better adapted to different agroecological conditions. The objectives of this research were to charac-terize different accessions of chickpea using quantitative morpho agronomic descriptors, to evaluate the phe-notypic variability of genetic materials, determine the descriptors that effectively contribute to the discrimination of genetic materials of the accessions and obtain a canonical function that allows classifying new accessions according to its origin. The characterization of the accessions was carried out on five plants, evaluating 17 agronomic morpho descriptors in the field and laboratory. The results of descriptive statistics revealed that the pod numbers per plant and 100 seed weight variables presented high CV values. Seed weight and volume components were positively associated among them, but negatively correlated to pod numbers per plant. In the Multivariate Profile Analisys, the characters that allowed the greatest differentiation of accessions were seed numbers per plant, pod numbers per plant and yield per plant. In the cluster analysis, there were three groups differentiated by 100 seed weight, height of plant, pod numbers per plant and seed numbers per plant variables. In the principal component analysis, it was found that the first three main components accounted for 66% of the total variation. In C1 the variables that made the greatest contribution to the separation of chickpea types were: pod numbers per plant, seed numbers per plant, seed size, 100 seed weight and height of plant. Discriminant Analysis detected that the most discriminating morpho-agronomic variables were pod numbers per plant and seed numbers per plant. This study allows us to conclude that there is significant variation for all the characters studied. This variability can be used for safe introducing different germplasm and for selecting parents with the highest genetic contrasts for chickpea genetic improvement programs, and broadening the genetic bases of this crop in Argentina.
Keywords: chickpea, variability, accessions, descriptors, multivariate analysis.
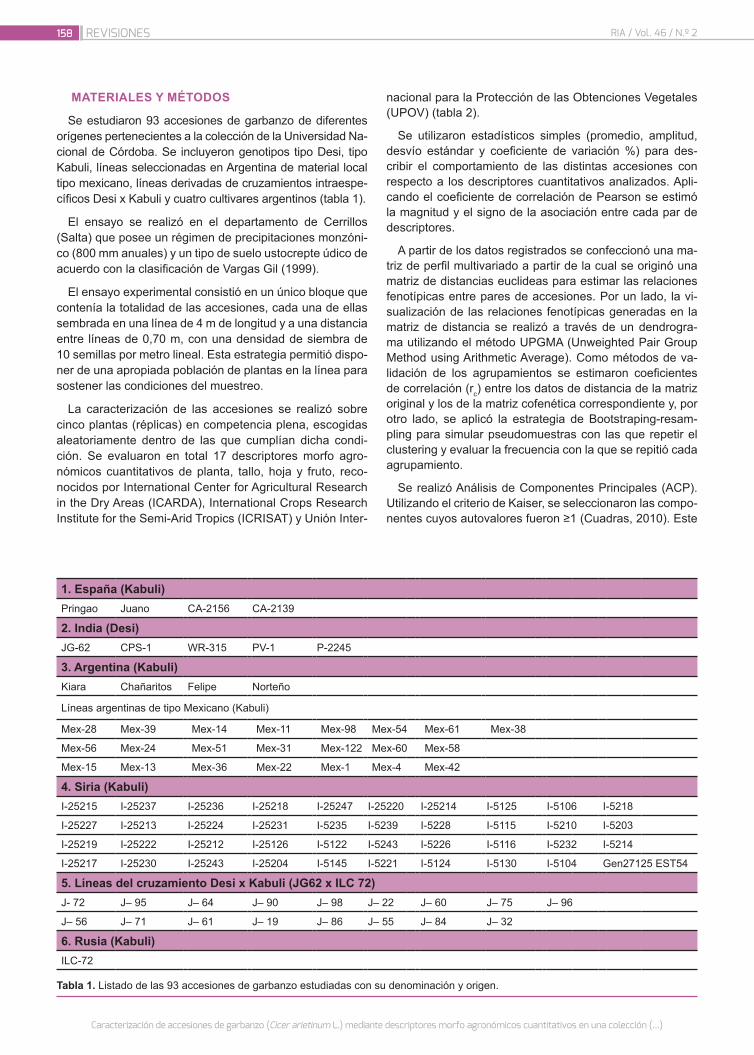
158
Caracterización de accesiones de garbanzo (Cicer arietinum L.) mediante descriptores morfo agronómicos cuantitativos en una colección (...)
RIA / Vol. 46 / N.º 2REVISIONES158158
MATERIALES Y MÉTODOS
Se estudiaron 93 accesiones de garbanzo de diferentes orígenes pertenecientes a la colección de la Universidad Na-cional de Córdoba. Se incluyeron genotipos tipo Desi, tipo Kabuli, líneas seleccionadas en Argentina de material local tipo mexicano, líneas derivadas de cruzamientos intraespe-cíficos Desi x Kabuli y cuatro cultivares argentinos (tabla 1).
El ensayo se realizó en el departamento de Cerrillos (Salta) que posee un régimen de precipitaciones monzóni-co (800 mm anuales) y un tipo de suelo ustocrepte údico de acuerdo con la clasificación de Vargas Gil (1999).
El ensayo experimental consistió en un único bloque que contenía la totalidad de las accesiones, cada una de ellas sembrada en una línea de 4 m de longitud y a una distancia entre líneas de 0,70 m, con una densidad de siembra de 10 semillas por metro lineal. Esta estrategia permitió dispo-ner de una apropiada población de plantas en la línea para sostener las condiciones del muestreo.
La caracterización de las accesiones se realizó sobre cinco plantas (réplicas) en competencia plena, escogidas aleatoriamente dentro de las que cumplían dicha condi-ción. Se evaluaron en total 17 descriptores morfo agro-nómicos cuantitativos de planta, tallo, hoja y fruto, reco-nocidos por International Center for Agricultural Research in the Dry Areas (ICARDA), International Crops Research Institute for the Semi-Arid Tropics (ICRISAT) y Unión Inter-
Tabla 1. Listado de las 93 accesiones de garbanzo estudiadas con su denominación y origen.
nacional para la Protección de las Obtenciones Vegetales (UPOV) (tabla 2).
Se utilizaron estadísticos simples (promedio, amplitud, desvío estándar y coeficiente de variación %) para des-cribir el comportamiento de las distintas accesiones con respecto a los descriptores cuantitativos analizados. Apli-cando el coeficiente de correlación de Pearson se estimó la magnitud y el signo de la asociación entre cada par de descriptores.
A partir de los datos registrados se confeccionó una ma-triz de perfil multivariado a partir de la cual se originó una matriz de distancias euclideas para estimar las relaciones fenotípicas entre pares de accesiones. Por un lado, la vi-sualización de las relaciones fenotípicas generadas en la matriz de distancia se realizó a través de un dendrogra-ma utilizando el método UPGMA (Unweighted Pair Group Method using Arithmetic Average). Como métodos de va-lidación de los agrupamientos se estimaron coeficientes de correlación (rc) entre los datos de distancia de la matriz original y los de la matriz cofenética correspondiente y, por otro lado, se aplicó la estrategia de Bootstraping-resam-pling para simular pseudomuestras con las que repetir el clustering y evaluar la frecuencia con la que se repitió cada agrupamiento.
Se realizó Análisis de Componentes Principales (ACP). Utilizando el criterio de Kaiser, se seleccionaron las compo-nentes cuyos autovalores fueron ≥1 (Cuadras, 2010). Este
1. España (Kabuli)Pringao Juano CA-2156 CA-2139
2. India (Desi)JG-62 CPS-1 WR-315 PV-1 P-2245
3. Argentina (Kabuli)Kiara Chañaritos Felipe Norteño
Líneas argentinas de tipo Mexicano (Kabuli)
Mex-28 Mex-39 Mex-14 Mex-11 Mex-98 Mex-54 Mex-61 Mex-38
Mex-56 Mex-24 Mex-51 Mex-31 Mex-122 Mex-60 Mex-58
Mex-15 Mex-13 Mex-36 Mex-22 Mex-1 Mex-4 Mex-42
4. Siria (Kabuli)I-25215 I-25237 I-25236 I-25218 I-25247 I-25220 I-25214 I-5125 I-5106 I-5218
I-25227 I-25213 I-25224 I-25231 I-5235 I-5239 I-5228 I-5115 I-5210 I-5203
I-25219 I-25222 I-25212 I-25126 I-5122 I-5243 I-5226 I-5116 I-5232 I-5214
I-25217 I-25230 I-25243 I-25204 I-5145 I-5221 I-5124 I-5130 I-5104 Gen27125 EST54
5. Líneas del cruzamiento Desi x Kabuli (JG62 x ILC 72)J- 72 J– 95 J– 64 J– 90 J– 98 J– 22 J– 60 J– 75 J– 96
J– 56 J– 71 J– 61 J– 19 J– 86 J– 55 J– 84 J– 32
6. Rusia (Kabuli)ILC-72
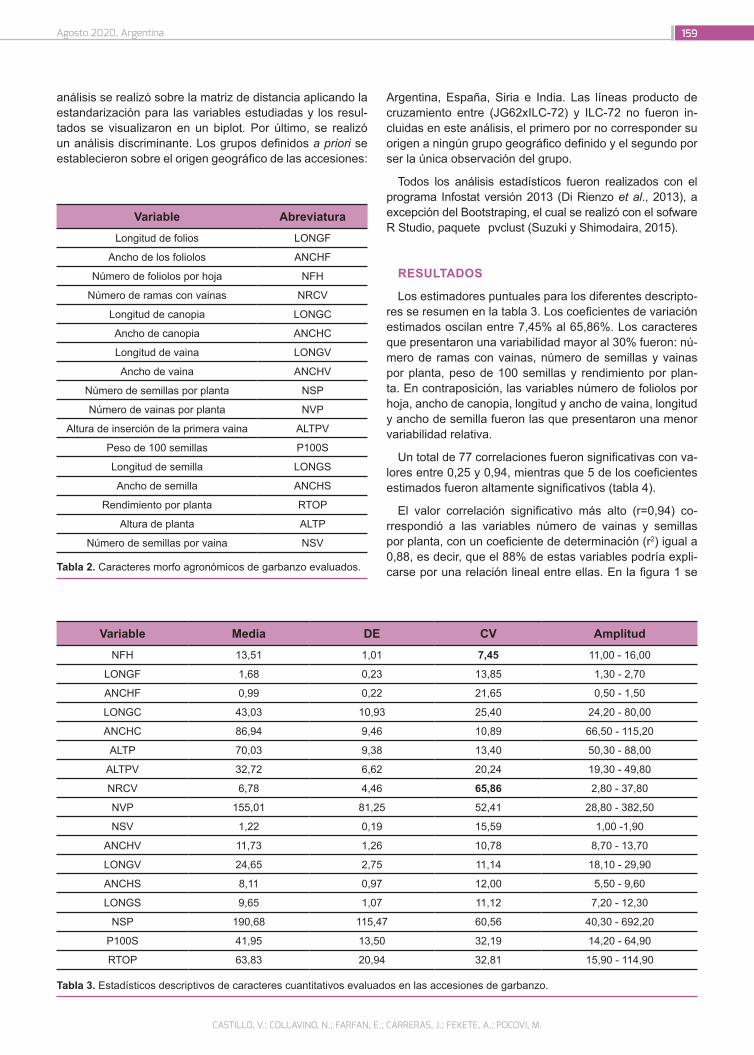
159
CASTILLO, V.; COLLAVINO, N.; FARFAN, E.; CARRERAS, J.; FEKETE, A.; POCOVI, M.
Agosto 2020, Argentina 159159
Tabla 3. Estadísticos descriptivos de caracteres cuantitativos evaluados en las accesiones de garbanzo.
análisis se realizó sobre la matriz de distancia aplicando la estandarización para las variables estudiadas y los resul-tados se visualizaron en un biplot. Por último, se realizó un análisis discriminante. Los grupos definidos a priori se establecieron sobre el origen geográfico de las accesiones:
Variable AbreviaturaLongitud de folios LONGF
Ancho de los foliolos ANCHF
Número de foliolos por hoja NFH
Número de ramas con vainas NRCV
Longitud de canopia LONGC
Ancho de canopia ANCHC
Longitud de vaina LONGV
Ancho de vaina ANCHV
Número de semillas por planta NSP
Número de vainas por planta NVP
Altura de inserción de la primera vaina ALTPV
Peso de 100 semillas P100S
Longitud de semilla LONGS
Ancho de semilla ANCHS
Rendimiento por planta RTOP
Altura de planta ALTP
Número de semillas por vaina NSV
Tabla 2. Caracteres morfo agronómicos de garbanzo evaluados.
Variable Media DE CV AmplitudNFH 13,51 1,01 7,45 11,00 - 16,00
LONGF 1,68 0,23 13,85 1,30 - 2,70
ANCHF 0,99 0,22 21,65 0,50 - 1,50
LONGC 43,03 10,93 25,40 24,20 - 80,00
ANCHC 86,94 9,46 10,89 66,50 - 115,20
ALTP 70,03 9,38 13,40 50,30 - 88,00
ALTPV 32,72 6,62 20,24 19,30 - 49,80
NRCV 6,78 4,46 65,86 2,80 - 37,80
NVP 155,01 81,25 52,41 28,80 - 382,50
NSV 1,22 0,19 15,59 1,00 -1,90
ANCHV 11,73 1,26 10,78 8,70 - 13,70
LONGV 24,65 2,75 11,14 18,10 - 29,90
ANCHS 8,11 0,97 12,00 5,50 - 9,60
LONGS 9,65 1,07 11,12 7,20 - 12,30
NSP 190,68 115,47 60,56 40,30 - 692,20
P100S 41,95 13,50 32,19 14,20 - 64,90
RTOP 63,83 20,94 32,81 15,90 - 114,90
Argentina, España, Siria e India. Las líneas producto de cruzamiento entre (JG62xILC-72) y ILC-72 no fueron in-cluidas en este análisis, el primero por no corresponder su origen a ningún grupo geográfico definido y el segundo por ser la única observación del grupo.
Todos los análisis estadísticos fueron realizados con el programa Infostat versión 2013 (Di Rienzo et al., 2013), a excepción del Bootstraping, el cual se realizó con el sofware R Studio, paquete pvclust (Suzuki y Shimodaira, 2015).
RESULTADOS
Los estimadores puntuales para los diferentes descripto-res se resumen en la tabla 3. Los coeficientes de variación estimados oscilan entre 7,45% al 65,86%. Los caracteres que presentaron una variabilidad mayor al 30% fueron: nú-mero de ramas con vainas, número de semillas y vainas por planta, peso de 100 semillas y rendimiento por plan-ta. En contraposición, las variables número de foliolos por hoja, ancho de canopia, longitud y ancho de vaina, longitud y ancho de semilla fueron las que presentaron una menor variabilidad relativa.
Un total de 77 correlaciones fueron significativas con va-lores entre 0,25 y 0,94, mientras que 5 de los coeficientes estimados fueron altamente significativos (tabla 4).
El valor correlación significativo más alto (r=0,94) co-rrespondió a las variables número de vainas y semillas por planta, con un coeficiente de determinación (r2) igual a 0,88, es decir, que el 88% de estas variables podría expli-carse por una relación lineal entre ellas. En la figura 1 se
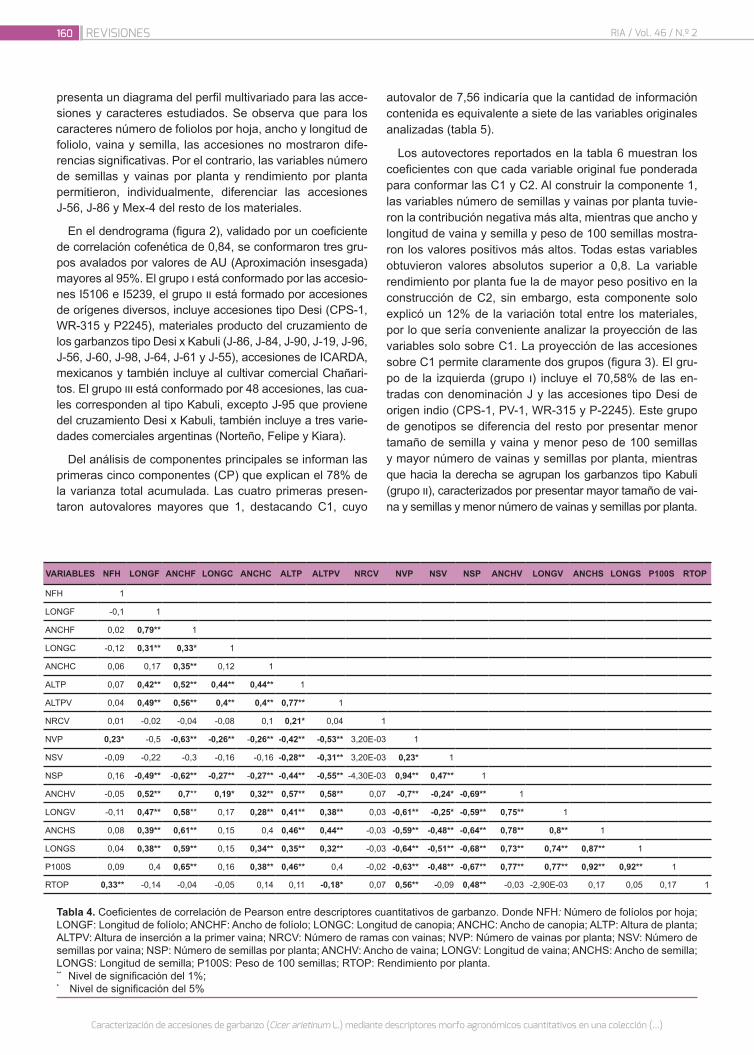
160
Caracterización de accesiones de garbanzo (Cicer arietinum L.) mediante descriptores morfo agronómicos cuantitativos en una colección (...)
RIA / Vol. 46 / N.º 2REVISIONES160160
presenta un diagrama del perfil multivariado para las acce-siones y caracteres estudiados. Se observa que para los caracteres número de foliolos por hoja, ancho y longitud de foliolo, vaina y semilla, las accesiones no mostraron dife-rencias significativas. Por el contrario, las variables número de semillas y vainas por planta y rendimiento por planta permitieron, individualmente, diferenciar las accesiones J-56, J-86 y Mex-4 del resto de los materiales.
En el dendrograma (figura 2), validado por un coeficiente de correlación cofenética de 0,84, se conformaron tres gru-pos avalados por valores de AU (Aproximación insesgada) mayores al 95%. El grupo i está conformado por las accesio-nes I5106 e I5239, el grupo ii está formado por accesiones de orígenes diversos, incluye accesiones tipo Desi (CPS-1, WR-315 y P2245), materiales producto del cruzamiento de los garbanzos tipo Desi x Kabuli (J-86, J-84, J-90, J-19, J-96, J-56, J-60, J-98, J-64, J-61 y J-55), accesiones de ICARDA, mexicanos y también incluye al cultivar comercial Chañari-tos. El grupo iii está conformado por 48 accesiones, las cua-les corresponden al tipo Kabuli, excepto J-95 que proviene del cruzamiento Desi x Kabuli, también incluye a tres varie-dades comerciales argentinas (Norteño, Felipe y Kiara).
Del análisis de componentes principales se informan las primeras cinco componentes (CP) que explican el 78% de la varianza total acumulada. Las cuatro primeras presen-taron autovalores mayores que 1, destacando C1, cuyo
autovalor de 7,56 indicaría que la cantidad de información contenida es equivalente a siete de las variables originales analizadas (tabla 5).
Los autovectores reportados en la tabla 6 muestran los coeficientes con que cada variable original fue ponderada para conformar las C1 y C2. Al construir la componente 1, las variables número de semillas y vainas por planta tuvie-ron la contribución negativa más alta, mientras que ancho y longitud de vaina y semilla y peso de 100 semillas mostra-ron los valores positivos más altos. Todas estas variables obtuvieron valores absolutos superior a 0,8. La variable rendimiento por planta fue la de mayor peso positivo en la construcción de C2, sin embargo, esta componente solo explicó un 12% de la variación total entre los materiales, por lo que sería conveniente analizar la proyección de las variables solo sobre C1. La proyección de las accesiones sobre C1 permite claramente dos grupos (figura 3). El gru-po de la izquierda (grupo i) incluye el 70,58% de las en-tradas con denominación J y las accesiones tipo Desi de origen indio (CPS-1, PV-1, WR-315 y P-2245). Este grupo de genotipos se diferencia del resto por presentar menor tamaño de semilla y vaina y menor peso de 100 semillas y mayor número de vainas y semillas por planta, mientras que hacia la derecha se agrupan los garbanzos tipo Kabuli (grupo ii), caracterizados por presentar mayor tamaño de vai-na y semillas y menor número de vainas y semillas por planta.
Tabla 4. Coeficientes de correlación de Pearson entre descriptores cuantitativos de garbanzo. Donde NFH: Número de folíolos por hoja; LONGF: Longitud de folíolo; ANCHF: Ancho de folíolo; LONGC: Longitud de canopia; ANCHC: Ancho de canopia; ALTP: Altura de planta; ALTPV: Altura de inserción a la primer vaina; NRCV: Número de ramas con vainas; NVP: Número de vainas por planta; NSV: Número de semillas por vaina; NSP: Número de semillas por planta; ANCHV: Ancho de vaina; LONGV: Longitud de vaina; ANCHS: Ancho de semilla; LONGS: Longitud de semilla; P100S: Peso de 100 semillas; RTOP: Rendimiento por planta.** Nivel de significación del 1%;* Nivel de significación del 5%
VARIABLES NFH LONGF ANCHF LONGC ANCHC ALTP ALTPV NRCV NVP NSV NSP ANCHV LONGV ANCHS LONGS P100S RTOP
NFH 1
LONGF -0,1 1
ANCHF 0,02 0,79** 1
LONGC -0,12 0,31** 0,33* 1
ANCHC 0,06 0,17 0,35** 0,12 1
ALTP 0,07 0,42** 0,52** 0,44** 0,44** 1
ALTPV 0,04 0,49** 0,56** 0,4** 0,4** 0,77** 1
NRCV 0,01 -0,02 -0,04 -0,08 0,1 0,21* 0,04 1
NVP 0,23* -0,5 -0,63** -0,26** -0,26** -0,42** -0,53** 3,20E-03 1
NSV -0,09 -0,22 -0,3 -0,16 -0,16 -0,28** -0,31** 3,20E-03 0,23* 1
NSP 0,16 -0,49** -0,62** -0,27** -0,27** -0,44** -0,55** -4,30E-03 0,94** 0,47** 1
ANCHV -0,05 0,52** 0,7** 0,19* 0,32** 0,57** 0,58** 0,07 -0,7** -0,24* -0,69** 1
LONGV -0,11 0,47** 0,58** 0,17 0,28** 0,41** 0,38** 0,03 -0,61** -0,25* -0,59** 0,75** 1
ANCHS 0,08 0,39** 0,61** 0,15 0,4 0,46** 0,44** -0,03 -0,59** -0,48** -0,64** 0,78** 0,8** 1
LONGS 0,04 0,38** 0,59** 0,15 0,34** 0,35** 0,32** -0,03 -0,64** -0,51** -0,68** 0,73** 0,74** 0,87** 1
P100S 0,09 0,4 0,65** 0,16 0,38** 0,46** 0,4 -0,02 -0,63** -0,48** -0,67** 0,77** 0,77** 0,92** 0,92** 1
RTOP 0,33** -0,14 -0,04 -0,05 0,14 0,11 -0,18* 0,07 0,56** -0,09 0,48** -0,03 -2,90E-03 0,17 0,05 0,17 1
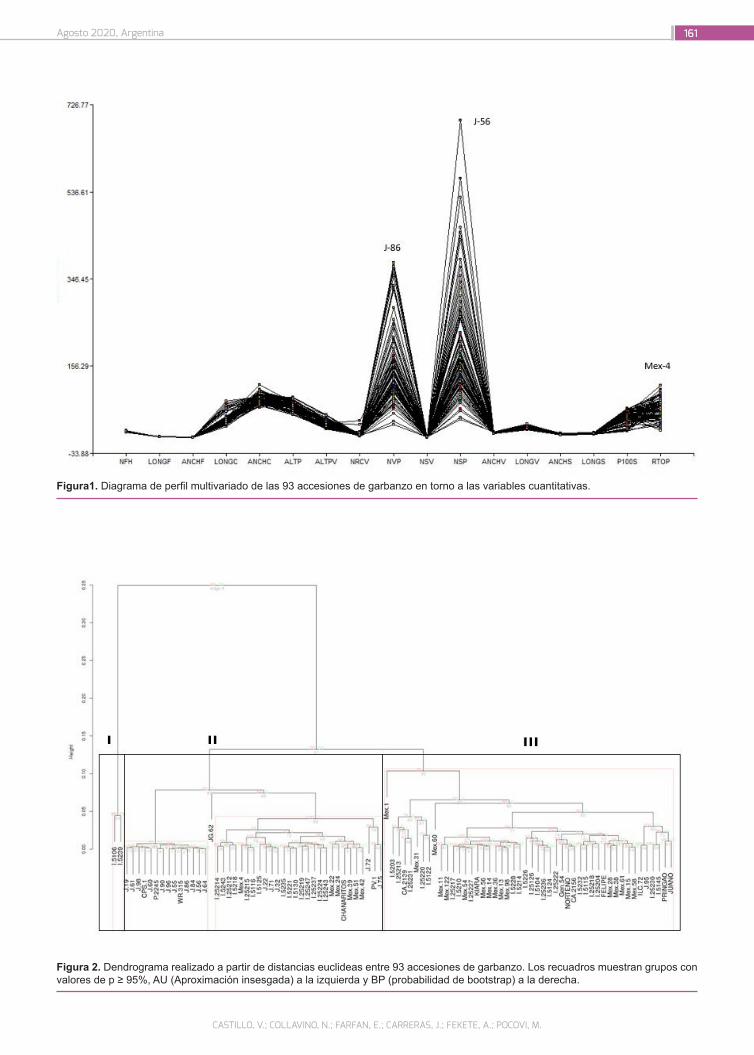
161
CASTILLO, V.; COLLAVINO, N.; FARFAN, E.; CARRERAS, J.; FEKETE, A.; POCOVI, M.
Agosto 2020, Argentina 161161
Figura1. Diagrama de perfil multivariado de las 93 accesiones de garbanzo en torno a las variables cuantitativas.
Figura 2. Dendrograma realizado a partir de distancias euclideas entre 93 accesiones de garbanzo. Los recuadros muestran grupos con valores de p ≥ 95%, AU (Aproximación insesgada) a la izquierda y BP (probabilidad de bootstrap) a la derecha.
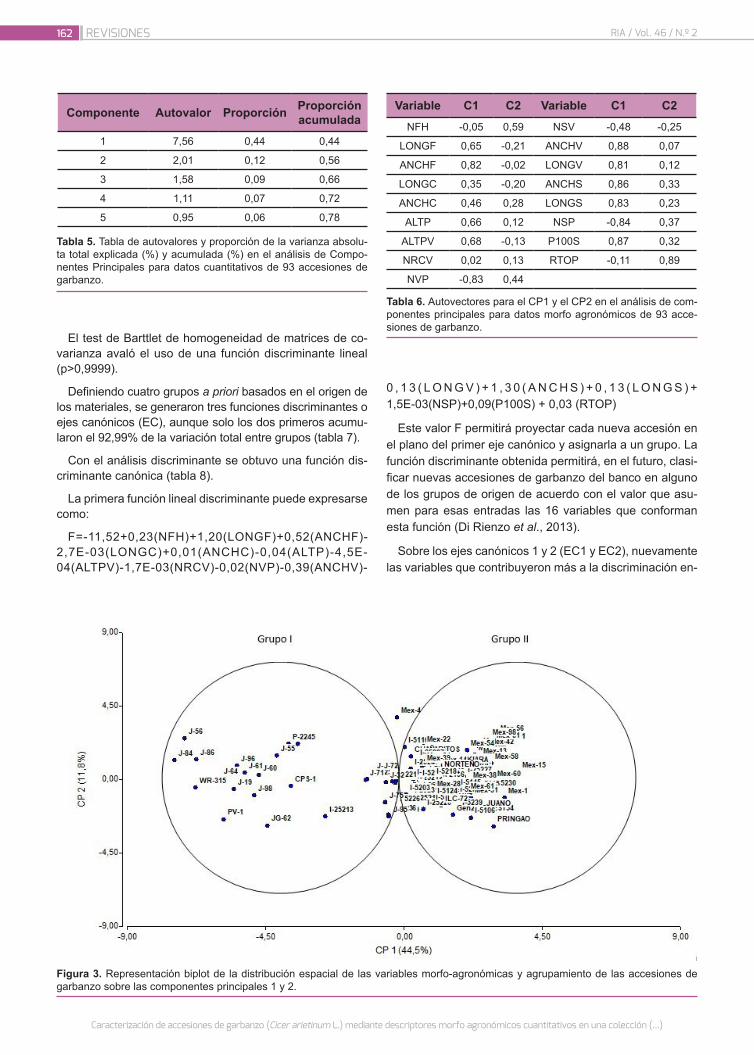
162
Caracterización de accesiones de garbanzo (Cicer arietinum L.) mediante descriptores morfo agronómicos cuantitativos en una colección (...)
RIA / Vol. 46 / N.º 2REVISIONES162162
Componente Autovalor Proporción Proporción acumulada
1 7,56 0,44 0,44
2 2,01 0,12 0,56
3 1,58 0,09 0,66
4 1,11 0,07 0,72
5 0,95 0,06 0,78
Tabla 5. Tabla de autovalores y proporción de la varianza absolu-ta total explicada (%) y acumulada (%) en el análisis de Compo-nentes Principales para datos cuantitativos de 93 accesiones de garbanzo.
Tabla 6. Autovectores para el CP1 y el CP2 en el análisis de com-ponentes principales para datos morfo agronómicos de 93 acce-siones de garbanzo.
Variable C1 C2 Variable C1 C2NFH -0,05 0,59 NSV -0,48 -0,25
LONGF 0,65 -0,21 ANCHV 0,88 0,07
ANCHF 0,82 -0,02 LONGV 0,81 0,12
LONGC 0,35 -0,20 ANCHS 0,86 0,33
ANCHC 0,46 0,28 LONGS 0,83 0,23
ALTP 0,66 0,12 NSP -0,84 0,37
ALTPV 0,68 -0,13 P100S 0,87 0,32
NRCV 0,02 0,13 RTOP -0,11 0,89
NVP -0,83 0,44
Figura 3. Representación biplot de la distribución espacial de las variables morfo-agronómicas y agrupamiento de las accesiones de garbanzo sobre las componentes principales 1 y 2.
El test de Barttlet de homogeneidad de matrices de co-varianza avaló el uso de una función discriminante lineal (p>0,9999).
Definiendo cuatro grupos a priori basados en el origen de los materiales, se generaron tres funciones discriminantes o ejes canónicos (EC), aunque solo los dos primeros acumu-laron el 92,99% de la variación total entre grupos (tabla 7).
Con el análisis discriminante se obtuvo una función dis-criminante canónica (tabla 8).
La primera función lineal discriminante puede expresarse como:
F=-11,52+0,23(NFH)+1,20(LONGF)+0,52(ANCHF)-2,7E-03(LONGC)+0,01(ANCHC)-0,04(ALTP)-4,5E-04(ALTPV)-1,7E-03(NRCV)-0,02(NVP)-0,39(ANCHV)-
0 , 1 3 ( L O N G V ) + 1 , 3 0 ( A N C H S ) + 0 , 1 3 ( L O N G S ) + 1,5E-03(NSP)+0,09(P100S) + 0,03 (RTOP)
Este valor F permitirá proyectar cada nueva accesión en el plano del primer eje canónico y asignarla a un grupo. La función discriminante obtenida permitirá, en el futuro, clasi-ficar nuevas accesiones de garbanzo del banco en alguno de los grupos de origen de acuerdo con el valor que asu-men para esas entradas las 16 variables que conforman esta función (Di Rienzo et al., 2013).
Sobre los ejes canónicos 1 y 2 (EC1 y EC2), nuevamente las variables que contribuyeron más a la discriminación en-
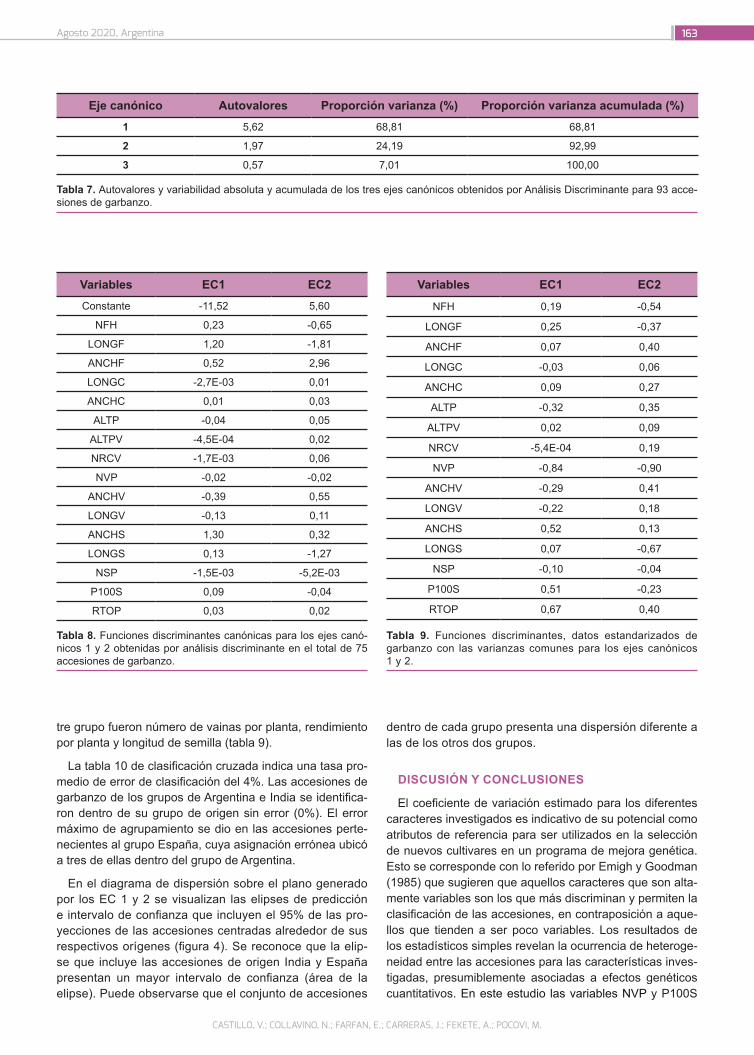
163
CASTILLO, V.; COLLAVINO, N.; FARFAN, E.; CARRERAS, J.; FEKETE, A.; POCOVI, M.
Agosto 2020, Argentina 163163
Tabla 7. Autovalores y variabilidad absoluta y acumulada de los tres ejes canónicos obtenidos por Análisis Discriminante para 93 acce-siones de garbanzo.
Tabla 8. Funciones discriminantes canónicas para los ejes canó-nicos 1 y 2 obtenidas por análisis discriminante en el total de 75 accesiones de garbanzo.
Tabla 9. Funciones discriminantes, datos estandarizados de garbanzo con las varianzas comunes para los ejes canónicos 1 y 2.
Eje canónico Autovalores Proporción varianza (%) Proporción varianza acumulada (%)1 5,62 68,81 68,81
2 1,97 24,19 92,99
3 0,57 7,01 100,00
Variables EC1 EC2Constante -11,52 5,60
NFH 0,23 -0,65
LONGF 1,20 -1,81
ANCHF 0,52 2,96
LONGC -2,7E-03 0,01
ANCHC 0,01 0,03
ALTP -0,04 0,05
ALTPV -4,5E-04 0,02
NRCV -1,7E-03 0,06
NVP -0,02 -0,02
ANCHV -0,39 0,55
LONGV -0,13 0,11
ANCHS 1,30 0,32
LONGS 0,13 -1,27
NSP -1,5E-03 -5,2E-03
P100S 0,09 -0,04
RTOP 0,03 0,02
Variables EC1 EC2
NFH 0,19 -0,54
LONGF 0,25 -0,37
ANCHF 0,07 0,40
LONGC -0,03 0,06
ANCHC 0,09 0,27
ALTP -0,32 0,35
ALTPV 0,02 0,09
NRCV -5,4E-04 0,19
NVP -0,84 -0,90
ANCHV -0,29 0,41
LONGV -0,22 0,18
ANCHS 0,52 0,13
LONGS 0,07 -0,67
NSP -0,10 -0,04
P100S 0,51 -0,23
RTOP 0,67 0,40
tre grupo fueron número de vainas por planta, rendimiento por planta y longitud de semilla (tabla 9).
La tabla 10 de clasificación cruzada indica una tasa pro-medio de error de clasificación del 4%. Las accesiones de garbanzo de los grupos de Argentina e India se identifica-ron dentro de su grupo de origen sin error (0%). El error máximo de agrupamiento se dio en las accesiones perte-necientes al grupo España, cuya asignación errónea ubicó a tres de ellas dentro del grupo de Argentina.
En el diagrama de dispersión sobre el plano generado por los EC 1 y 2 se visualizan las elipses de predicción e intervalo de confianza que incluyen el 95% de las pro-yecciones de las accesiones centradas alrededor de sus respectivos orígenes (figura 4). Se reconoce que la elip-se que incluye las accesiones de origen India y España presentan un mayor intervalo de confianza (área de la elipse). Puede observarse que el conjunto de accesiones
dentro de cada grupo presenta una dispersión diferente a las de los otros dos grupos.
DISCUSIÓN Y CONCLUSIONES
El coeficiente de variación estimado para los diferentes caracteres investigados es indicativo de su potencial como atributos de referencia para ser utilizados en la selección de nuevos cultivares en un programa de mejora genética. Esto se corresponde con lo referido por Emigh y Goodman (1985) que sugieren que aquellos caracteres que son alta-mente variables son los que más discriminan y permiten la clasificación de las accesiones, en contraposición a aque-llos que tienden a ser poco variables. Los resultados de los estadísticos simples revelan la ocurrencia de heteroge-neidad entre las accesiones para las características inves-tigadas, presumiblemente asociadas a efectos genéticos cuantitativos. En este estudio las variables NVP y P100S
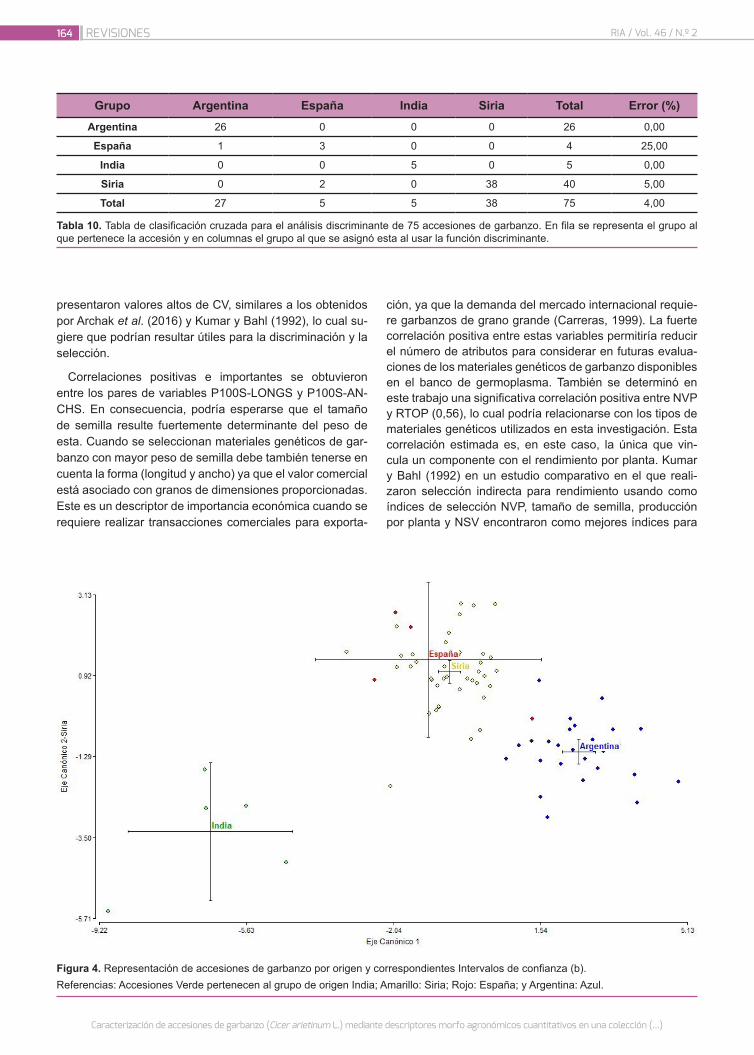
164
Caracterización de accesiones de garbanzo (Cicer arietinum L.) mediante descriptores morfo agronómicos cuantitativos en una colección (...)
RIA / Vol. 46 / N.º 2REVISIONES164164
presentaron valores altos de CV, similares a los obtenidos por Archak et al. (2016) y Kumar y Bahl (1992), lo cual su-giere que podrían resultar útiles para la discriminación y la selección.
Correlaciones positivas e importantes se obtuvieron entre los pares de variables P100S-LONGS y P100S-AN-CHS. En consecuencia, podría esperarse que el tamaño de semilla resulte fuertemente determinante del peso de esta. Cuando se seleccionan materiales genéticos de gar-banzo con mayor peso de semilla debe también tenerse en cuenta la forma (longitud y ancho) ya que el valor comercial está asociado con granos de dimensiones proporcionadas. Este es un descriptor de importancia económica cuando se requiere realizar transacciones comerciales para exporta-
Figura 4. Representación de accesiones de garbanzo por origen y correspondientes Intervalos de confianza (b). Referencias: Accesiones Verde pertenecen al grupo de origen India; Amarillo: Siria; Rojo: España; y Argentina: Azul.
ción, ya que la demanda del mercado internacional requie-re garbanzos de grano grande (Carreras, 1999). La fuerte correlación positiva entre estas variables permitiría reducir el número de atributos para considerar en futuras evalua-ciones de los materiales genéticos de garbanzo disponibles en el banco de germoplasma. También se determinó en este trabajo una significativa correlación positiva entre NVP y RTOP (0,56), lo cual podría relacionarse con los tipos de materiales genéticos utilizados en esta investigación. Esta correlación estimada es, en este caso, la única que vin-cula un componente con el rendimiento por planta. Kumar y Bahl (1992) en un estudio comparativo en el que reali-zaron selección indirecta para rendimiento usando como índices de selección NVP, tamaño de semilla, producción por planta y NSV encontraron como mejores índices para
Tabla 10. Tabla de clasificación cruzada para el análisis discriminante de 75 accesiones de garbanzo. En fila se representa el grupo al que pertenece la accesión y en columnas el grupo al que se asignó esta al usar la función discriminante.
Grupo Argentina España India Siria Total Error (%)Argentina 26 0 0 0 26 0,00
España 1 3 0 0 4 25,00
India 0 0 5 0 5 0,00
Siria 0 2 0 38 40 5,00
Total 27 5 5 38 75 4,00

165
CASTILLO, V.; COLLAVINO, N.; FARFAN, E.; CARRERAS, J.; FEKETE, A.; POCOVI, M.
Agosto 2020, Argentina 165165
rendimiento el NVP y el tamaño de semilla. En este trabajo, por una parte, los componentes de peso y volumen de se-millas resultan asociados positivamente entre sí, pero ne-gativamente correlacionados con el NVP. Debido a ello se pone en evidencia que se tratan de efectos competitivos en la determinación del RTOP, ya que, si se incrementara el NVP, redundaría en una significativa reducción de volumen y peso de la semilla. Este efecto de compensación explica la relativamente baja determinación del número de vainas sobre el rendimiento por planta (de apenas el 31,3%), lo que sugiere que los componentes de tamaño y peso de semilla contribuyen indirectamente al RTOP. Por otra parte, y en coincidencia con Padmavathi et al. (1995) la detec-ción de elevadas correlaciones entre descriptores puede ser valiosa al momento de ser utilizada como herramienta tanto en selección directa como indirecta para incremen-tar productividad. La correlación negativa obtenida en este estudio entre las variables P100S y NVP coincide con el trabajo realizado de correlación por Pundir (1988) en una población de 12.010 individuos de garbanzo.
En el análisis del perfil multivariado los caracteres que per-mitieron la mayor diferenciación de las accesiones fueron NSP, NVP y RTOP. Este resultado es similar al obtenido por Cantarutti et al. (2016) a pesar de que trabajó con solo 22 materiales resultantes del cruzamiento entre JG-62 y ILC-72.
Producto de la caracterización morfológica en las 93 ac-cesiones de garbanzo, se determinó que existe variabilidad fenotípica, ya que por medio del análisis de conglomerados se dio la conformación de cuatro grupos diferenciados, lo que fue una de las principales diferencias entre los grupos P100S, ALTP, NVP y NSP.
En el ACP se encontró que las tres primeras compo-nentes principales explicaron el 66% de la variabilidad total, donde el primer componente aportó 44% de ese to-tal. Resultado similar al obtenido por Shagarodsky et al. (2001) al trabajar con 19 entradas de distintos orígenes, en el cual las tres primeras componentes explicaron el 64,3% de la variabilidad total. En el CP1 las variables de mayor peso fueron: NVP, NSP, tamaño de vaina y semilla, P100S, LONGF, ALTP y ALTPV. Este resultado coincide parcialmente con Archak et al. (2016), en que las varia-bles NVP, P100S y ALTP fueron determinantes en el CP1. En general se aprecia una gran variabilidad en el conjunto de accesiones de garbanzo estudiadas con los 17 des-criptores morfo agronómicos evaluados. Los caracteres seleccionados mediante el análisis de componentes prin-cipales han contribuido a la descripción de los dos tipos de garbanzo (Desi y Kabuli). Las variables que realizaron mayor contribución a la separación de los tipos fueron: NVP, NSP, tamaño de semillas, P100S y ALTP. Todos estos caracteres a excepción de ALTP son citados como importantes componentes del rendimiento en garbanzo (Kharrat et al., 1991; Kumar y Bahl 1992; Maynez et al., 1993; Rubio et al., 1998). Los resultados obtenidos ade-más prueban que la metodología utilizada fue la apropia-da para diferenciar claramente los distintos tipos de mate-riales incluidos en el estudio.
Mediante el análisis discriminante se lograron detectar las variables morfo agronómicas que permitieron discrimi-nar genotipos en la colección de garbanzos. Si bien hubo coincidencias con los resultados de ACP y con el gráfico de perfil multivariado en relación con los descriptores más discriminantes (NVP y NSP), este nuevo estudio permitió asignar probabilísticamente las accesiones a los diferentes grupos propuestos con buen grado de aceptabilidad ya que los niveles de confusiones son bajos. Este resultado coinci-de con el estudio realizado por Sosa et al. (2018) en el que trabajaron con 42 accesiones de orígenes diferentes y con-cluyeron que una de las variables más discriminantes fue el descriptor NVP. Un importante aporte a los programas de mejora genética de garbanzo fue haber obtenido una función canónica que permitirá en un futuro clasificar, con un valor de probabilidad asociado, nuevas accesiones del banco de germoplasma en alguno de los grupos de origen establecidos en este trabajo.
La información presentada permite concluir que existe variación fenotípica para todos los caracteres estudiados en las 93 accesiones evaluadas. Esta variabilidad puede ser utilizada para introducir con mayor seguridad diferente germoplasma y seleccionar los progenitores con los mayo-res contrastes genéticos en los programas de mejora de garbanzo y ampliar la base genética de este cultivo en Ar-gentina, ya que actualmente se encuentra limitada a unos pocos cultivares emparentados entre sí.
AGRADECIMIENTOS
Al Proyecto INTA-AUDEAS-CONADEV, CIAC-940138 por la colaboración y apoyo durante el transcurso de este trabajo.
BIBLIOGRAFÍA ARCHAK, S.; RISHI, K.T.; HARER, P.N.; MAHASE, L.B.;
SINGH, N., DAHIYA OM, P.; ABDUL, N.M., SINGH, M.; TILEKAR, V.; KUMAR, V.; DUTTA, M.; SINGH, N.P., BANSAL, K.C. 2016. Characterization of chickpea germplasm conserved in the Indian National Genebank and development of a core set using qualita-tive and quantitative trait data. The Crop Journal 4. 417-424 pp.
CANTARUTTI, N.; LUCARELLI, L.; SOLONYENZY, K. 2016. Caracterización multivariada de genotipos de garbanzo según ca-racteres morfológicos y de rendimiento. Universidad Nacional de Córdoba.
CARRERAS, J. 1999. Caracterización de líneas selectas de garbanzo (Cicer arietinum L.). Tesis Maestría Tecnología. Bibliote-ca Facultad de Ciencias Agropecuarias. Universidad Nacional de Córdoba. Biblioteca 11Ca:10446. 144 p.
CARRERAS, J. 2013. Establecimiento de bases genéticas para la mejora del garbanzo (Cicer arietinum L.) en Argentina. Tesis doctoral. UCO-España.
CARRERAS, J.; MAZZUFERI, V.; KARLIN, M. 2016. El Cultivo de Garbanzo (Cicer arietinum L.) en Argentina. 1.a ed. Universidad Nacional de Córdoba. Córdoba.
CUADRAS, C.M. 2010. Nuevos Métodos de Análisis Multiva-riantes. CMC Editions. Barcelona. 285 p.
Di RIENZO, J.A.; CASANOVES, F.; BALZARINI, M.G.; GONZA-LEZ, L.; TABLADA, M.; ROBLEDO, C.W. 2013. InfoStat, versión 2013,

166
Caracterización de accesiones de garbanzo (Cicer arietinum L.) mediante descriptores morfo agronómicos cuantitativos en una colección (...)
RIA / Vol. 46 / N.º 2REVISIONES166166
Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. (Disponible: http://www.InfoStat.com.ar verificado: mayo de 2020).
EEOC (ESTACIÓN EXPERIMENTAL OBISPO COLOMBRES). 2018. PANORAMA Campaña de poroto 2018. Siembra de garban-zo. Tucuman. (Disponible: http://www.eeaoc.org.ar/tematica/44/ Legumbres-secas.html verificado: mayo de 2020).
EMIGH, T.H.; GOODMAN, M.M. 1985. Multivariate analysis in nematede taxonomy. En: BARKER, K.R.; CARTER, C.E.; SASSER, J.N. (eds.). An advanced treatise on Meloidogyne. Vol JI methodology. Nort Carolina State University Graphics. Raleigh, N.C. 197-204 pp.
FAO. 2016. El estado mundial de la agricultura y la alimentac-ión. Roma.
FELSENSTEIN, J. 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39:783-791.
ICARDA. 1993. Food legume improvement Program. AnnualRe-port for 1993. ICARDA, Aleppo, Syria, 284 p.
ICRISAT. 1984. The seventh International Pearl Millet Adapta-tion Trial (IPMA T 7), 1981. Patancheru, A.P. 502 324, India:. Inter-national Crops Research Institute for the Semi-Arid Tropics (Lim-ited distribution.)
JENSEN, E.S.; HAUGGAARD NIELSEN, H. 2003. How can increased use of biological N2 fixation in agriculture benefit the environment? Plant and Soil 252:177-186.
KHARRAT, M.; BEN SALAH, H.; HALILA, H.M. 1991. Faba bean status and prospects in Tunisia. Options Méditerranéennes. Série Séminaires, 10: 169-172.
KUMAR, J.; BAHL, P.N. 1992. Direct and indirect selection for yield in chickpea. Euphytica 60:197-199.
MAYNEZ, M.; MOREMO, M.T.; GIL, J. 1993 Desi/Kabuli intro-gression for yield improvement in chickpea (Cicer arietinum L.). J. Genet Breed 47:61-64.
MORENO, M.T.; CUBERO, J.I. 1978. Variation in (Cicer Arieti-num L.). Euphytica 27:465-485.
PADMAVATHI, P.V.; SREEMANNARAYANA MURTHY, S.; SATYANARAYANA RAO, V.; LAL AHAMED, M. 2013. Correlation
and Path Coefficient Analysis in Kabuli Chickpea (Cicer arietinum L.). International Journal of Applied Biology and Pharmaceutical Technology. Volumen 4, Issue-3.
PUNDIR, R.P.S.; REDDY, K.N.; MELAK, H. 1988. ICRISAT Chickpea germplasm catalog: evaluation and análisis. Catalog: Evaluation and Analysis. Volumen 1 de ICRISAT Chickpea Germ-plasm Catalog, Melak H. Mengesha. International Crops Research Institute for the Semi-Arid Tropics. Cornell University. 94 p.
PUSHPAMMA, P.; GEERVANI, P. 1987. Utilisation of chickpea. En: SAXENA, M.C.; SINGH, K.B. (eds.). The Chickpea. CAB Inter-national, Wallingford, Reino Unido. 357-368 pp.
RUBIO, J.; MORENO, M.T.; CUBERO, J.I.; GIL, J. 1998. Effect of the gene for double pod in chickpea on yield, yield components and stability of yield. Plant Breeding 117:585-587.
SHAGARODSKY, T.; CHIANG, M.L.; LÓPEZ, Y. 2001. Eval-uación de cultivares de garbanzo (Cicer arietinum L.) en Cuba. Agronomía Mesoamericana, vol. 12, núm. 95-98 pp.
SOSA, M.; CASTILLO, V.; FARFÁN, E.; DELGADO, R.; PE-REYRA, C.; CARRERAS, J.; COLLAVINO, G.; FÉKETE, A.; POCOVÍ, M. 2018. Análisis discriminante en la clasificación morfo agronómica de accesiones de garbanzo y comparación con grupos definidos a priori mediante microsatélites. Libro de Resúmenes del Congreso Argentino de Horticultura.
SUZUKI, R.; SHIMODAIRA, H. 2015. Hierarchical Clustering with P-Values via Multiscale Bootstrap Resampling. Package: pv-clust. Versión: 2.0-0.
TANNO, K.; WILLCOX, G. 2006. The origins of cultivation of Cicer arietinum L. and Vicia faba L.: early finds from Tell el-Kerkh, north-west Syria, late 10th millennium BP. Vegetation History and Archaeobotany 15 (3): 197-204.
UPOV. 2005. International Union for Protection of Vegetal Ob-taining.
VARGAS GIL, J.R. 1999. Carta de suelos de la República Argentina, provincia de Salta- Valle de Lerma. EEA INTA Cer-rillos. Salta.

ALMADA, M.S.; NUSSENBAUM, A.L.
Agosto 2020, Argentina 167167
Técnica de sexado de pupas de Anthonomus grandis (Coleoptera: Curculionidae): una forma rápida y fácil de sexar para estudios biológicos
RESUMEN El picudo del algodonero, Anthonomus grandis Boheman (Coleoptera: Curculionidae), es considerado una
de las plagas más perjudiciales del cultivo de algodón en América. Muchos estudios requieren la determina-ción del sexo para lo que se utilizan diferentes técnicas, pero estas tienen limitaciones o requieren cierto en-trenamiento previo. En el presente trabajo, contribuimos con mejores ilustraciones y fotografías actualizadas de las diferencias entre sexos de pupas de A. grandis para proporcionar una técnica más fácil y rápida para la diferenciación de sexos.
Palabras clave: algodón, determinación del sexo, picudo del algodonero, plaga.
ABSTRACT
Boll weevil, Anthonomus grandis Boheman (Coleoptera: Curculionidae), is considered the most destructive cotton pest in America. Many studies require sex determination for which different techniques are used, but they have limitations or require previous training. In the present work, we contribute with better illustrations and updated photographs of the differences between sexes of A. grandis pupae to provide an easy and quick technique for sex differentiation.
Keywords: cotton, sex determination, boll weevil, pest.
1Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Reconquista, Ruta 11, km 773 (3560). Reconquista, Santa Fe, Argentina; 2Universidad Católica de Santa Fe, Facultad de Ciencias Agropecuarias, Reconquista, Santa Fe, Argentina. Correo electrónico: [email protected] Nacional de Tecnología Agropecuaria (INTA), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Instituto de Genética “Ewald A. Favret” (IGEAF), G.V.A. Instituto de Agrobiotecnología y Biología Molecurlar (IABIMO), Laboratorio de Insectos de Importancia Agronómica, Hurlingham, Buenos Aires, Argentina. Correo electrónico: [email protected]
Recibido 29 de octubre de 2019 // Aceptado 06 de abril de 2020 // Publicado online 16 de septiembre de 2020
ALMADA, M.S.1,2; NUSSENBAUM, A.L.3

Técnica de sexado de pupas de Anthonomus grandis (Coleoptera: Curculionidae): una forma rápida y fácil de sexar para estudios biológicos
COMUNICACIÓN CORTA RIA / Vol. 46 / N.º 2168168
INTRODUCCIÓN
El picudo del algodonero, Anthonomus grandis Boheman (Coleoptera: Curculionidae), es una de las plagas más im-portantes del algodón Gossypium hirsutum L. (Malvales: Malvaceae) en el continente americano. En la República Argentina se capturó por primera vez en 1993 en áreas no cultivadas de la provincia de Misiones, y en 1994 cerca de cultivos de algodón en el límite con Paraguay (Marengo Lozada y Whitcomb, 1993; Stadler y Buteler, 2007). Luego de los primeros informes en Argentina, la plaga se ha dis-persado a nuevas áreas y, finalmente, a pesar de la fuerte acción del gobierno nacional para disminuir su propaga-ción, se obtuvieron registros de la plaga en las principales zonas algodoneras de las provincias de Formosa (1994), Corrientes (1997), Chaco (2002), Santa Fe (2004) y San-tiago del Estero (2015) (Lanteri et al., 2003; Sosa et al., 2009), donde se encuentra actualmente establecida.
El picudo del algodonero daña las estructuras reproduc-tivas de las plantas de algodón debido a su estrategia de alimentación y oviposición, produciendo pérdida de calidad de fibra y abscisión de pimpollos (Showler, 2008). Las hem-bras oviponen en el interior del pimpollo o cápsulas peque-ñas y permanecen hasta completar su desarrollo larval en tres estados. Las larvas del segundo o tercer estadio ge-neran la abscisión de la cápsula, lo que permite culminar el desarrollo dentro de este en el suelo, hasta que los adultos emergen (Coakley et al., 1969; Showler, 2005; Showler y Cantú, 2005). La rápida colonización de la plaga está rela-cionada con características del ciclo de vida de esta espe-cie, como por ejemplo, alta capacidad reproductiva, ciclo de vida multivoltino, desarrollo endofítico de las etapas in-maduras, junto con las condiciones ambientales, donde la depredación y el parasitismo proporcionan un control defi-ciente, aumentando rápidamente los niveles de infestación (Grilli et al., 2012).
Muchos estudios, especialmente estudios ecológicos de esta plaga, requieren diferenciar los sexos tanto de adultos como de pupas. Sin embargo, las diferentes técnicas utili-zadas para el sexado tienen limitaciones o requieren entre-namiento previo. Entre las diferentes técnicas, se pueden mencionar la disección de adultos y la observación de la morfología interna, técnica posible únicamente cuando el estudio no requiere del insecto vivo. Otros métodos que permite el sexado es oprimir suavemente el abdomen del adulto hasta extraer parcialmente los genitales, o levantar los élitros hasta observar la cantidad de terguitos que es diferencial entre los sexos, aunque estos métodos gene-ralmente pueden producir daños en el adulto (Sappington y Spurgeon, 2000).
La técnica más utilizada para sexar al picudo del algodo-nero fue descripta por Agee (1964) y mejorada por Sapping-ton y Spurgeon (2000). El método para sexar adultos implica la observación del octavo (último) terguito abdominal, que en el macho es fijo y presenta una muesca en el centro, mientras que en la hembra es móvil y se proyecta junto con el ovipositor. Esta técnica fue ampliamente utilizada en dife-rentes estudios porque es fácil y muy precisa (Firmino et al.,
2013; Greenberg et al., 2004a; Lewis et al., 2002; Spurgeon et al., 2003). Sin embargo, se requiere experiencia y habili-dad para manipular adultos vivos sin dañarlos o que sufran estrés, fundamental en insectos que luego serán evaluados en ensayos de comportamiento. Otra limitación de esta téc-nica es que, cuando se necesitan obtener adultos vírgenes es necesario separar adultos recién emergidos para garan-tizar que los insectos no copulen, teniendo en cuenta que el apareamiento podría ocurrir luego de 12 horas de la emer-gencia (Mayer y Brazzel, 1963).
Burke (1968) y Anderson (1968) realizaron una descrip-ción de las pupas de la tribu Anthonomini, principalmente de las especies A. grandis grandis y A. grandis thurberiae. Se reconocieron pupas hembras por la presencia de un par de lóbulos en una vista posteroventral; están ausentes en pupas machos. Esta diferencia de sexo es fácil de distinguir bajo el microscopio binocular. Las ventajas de esta técnica comprenden una manipulación factible de las pupas y permi-ten el sexado antes de la aparición de adultos. Por lo tanto, es fácil obtener insectos recién emergidos y vírgenes.
Nuestro objetivo es mostrar fotografías y dibujos inter-pretativos para ilustrar las diferencias entre las pupas de Anthonomus grandis machos y hembras, y mejorar la téc-nica de sexado en esta especie.
MATERIALES Y MÉTODOS
Los insectos fueron recolectados en el campo y obteni-dos de una cría en laboratorio con dieta artificial. Se evaluó un total de 120 pupas (recolección de campo y de cría). La recolección de pupas en el campo consistió en 30 pimpollos de algodón presentes en el suelo, durante el ciclo repro-ductivo del cultivo de algodón en la Estación Experimental Agropecuaria (EEA-INTA) Reconquista, Santa Fe, Argenti-na (29° 11´ S, 59° 52´O) de enero a abril de 2017. Respec-to a las pupas criadas en laboratorio, se obtuvieron pupas de la colonia mantenida en el Instituto de Microbiología y Zoología Agrícola (IMYZA), INTA Castelar, iniciada con in-dividuos recolectados en áreas rurales de la provincia de Santa Fe, Argentina en 2007 y de la colonia mantenida en EEA INTA Reconquista. Ambas colonias fueron vigorizadas con gorgojos silvestres una vez al año. Las condiciones de cría fueron 27 ± 1 °C, 50 ± 10% HR y 12:12 fotoperiodo. Las larvas y los adultos fueron alimentados con una dieta artificial, de acuerdo con las modificaciones realizadas en la metodología propuesta por Monnerat et al. (2000).
Para sexar las pupas se siguieron las diferencias des-critas por Anderson (1968). Estas diferencias se ilustraron utilizando la técnica de dibujo de grafito del último esternón abdominal visible, y las pupas se fotografiaron con un mi-croscopio binocular. Además, observamos diariamente las pupas para evaluar cuándo se identifican estas caracterís-ticas desde el principio hasta el final del estadio.
Finalmente, para evaluar las diferencias entre sexos en el tamaño corporal, se midieron la longitud y el peso de las pupas tanto para los insectos recolectados en el campo como para los insectos de la cría obtenida de laboratorio.
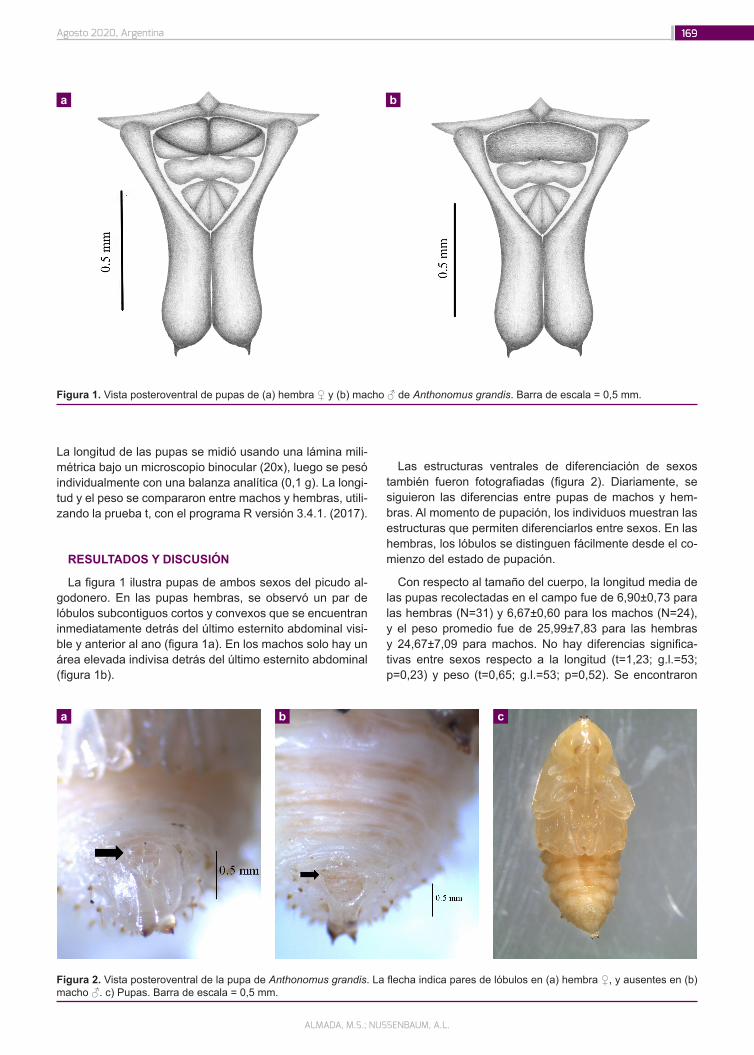
ALMADA, M.S.; NUSSENBAUM, A.L.
Agosto 2020, Argentina 169169
Figura 1. Vista posteroventral de pupas de (a) hembra ♀ y (b) macho ♂ de Anthonomus grandis. Barra de escala = 0,5 mm.
Figura 2. Vista posteroventral de la pupa de Anthonomus grandis. La flecha indica pares de lóbulos en (a) hembra ♀, y ausentes en (b) macho ♂. c) Pupas. Barra de escala = 0,5 mm.
La longitud de las pupas se midió usando una lámina mili-métrica bajo un microscopio binocular (20x), luego se pesó individualmente con una balanza analítica (0,1 g). La longi-tud y el peso se compararon entre machos y hembras, utili-zando la prueba t, con el programa R versión 3.4.1. (2017).
RESULTADOS Y DISCUSIÓN
La figura 1 ilustra pupas de ambos sexos del picudo al-godonero. En las pupas hembras, se observó un par de lóbulos subcontiguos cortos y convexos que se encuentran inmediatamente detrás del último esternito abdominal visi-ble y anterior al ano (figura 1a). En los machos solo hay un área elevada indivisa detrás del último esternito abdominal (figura 1b).
Las estructuras ventrales de diferenciación de sexos también fueron fotografiadas (figura 2). Diariamente, se siguieron las diferencias entre pupas de machos y hem-bras. Al momento de pupación, los individuos muestran las estructuras que permiten diferenciarlos entre sexos. En las hembras, los lóbulos se distinguen fácilmente desde el co-mienzo del estado de pupación.
Con respecto al tamaño del cuerpo, la longitud media de las pupas recolectadas en el campo fue de 6,90±0,73 para las hembras (N=31) y 6,67±0,60 para los machos (N=24), y el peso promedio fue de 25,99±7,83 para las hembras y 24,67±7,09 para machos. No hay diferencias significa-tivas entre sexos respecto a la longitud (t=1,23; g.l.=53; p=0,23) y peso (t=0,65; g.l.=53; p=0,52). Se encontraron
a b
a b c

Técnica de sexado de pupas de Anthonomus grandis (Coleoptera: Curculionidae): una forma rápida y fácil de sexar para estudios biológicos
COMUNICACIÓN CORTA RIA / Vol. 46 / N.º 2170170
los mismos resultados para las pupas de cría en labora-torio. La longitud promedio fue de 7,10 ± 0,77 para las hembras (N=28) y 6,82 ± 0,74 para los machos (N=30), y el peso promedio fue de 24,49±7,34 para las hembras y 21,99±7,30 para los machos. Tampoco se encontraron diferencias significativas entre sexos respecto a la longitud (t=1,41; g.l.=56; p=0,16) y peso (t=1,30; g.l.=56; p=0,20). Estos resultados son similares a los de estudios previos en los que no existen diferencias entre los sexos para la longitud corporal y el peso (Sappington y Spurgeon, 2000; Greenberg et al., 2004b). La variación en el acceso al ali-mento (calidad y disponibilidad) entre las larvas podría dar como resultado una amplia gama de tamaños de adultos (Greenberg et al., 2005) independientemente de su sexo.
En este trabajo, las diferencias entre sexos se identifica-ron rápidamente en todas las pupas observadas. Nuestros resultados actualizan información ilustrativa permitiendo la separación por sexo de pupas de A. grandis, así como as-pectos sobre su biología. Esta técnica es fácil y rápida para los rasgos de sexado y es una herramienta útil al momento de realizar acciones necesarias para el Manejo Integrado de Plagas del picudo algodonero en Argentina.
AGRADECIMIENTOS
Se agradece la colaboración del personal técnico y auxi-liar de la Estación Experimental Agropecuaria Reconquista de INTA, al Consejo Nacional de Investigaciones Científi-cas y Tecnológicas (CONICET), a la Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT), y al Minis-terio de Ciencias de Santa Fe ASaCTeI (2010-160-16) por su financiamiento.
BIBLIOGRAFÍAAGEE, H.R. 1964. Characters for determination of sex of the boll
weevil. Journal of Economic Entomology, 57, 500-501.ANDERSON, D.M. 1968. Observations on the Pupae of Antho-
nomus grandis grandis Boheman and A. grandis thurberiae Pierce (Coleoptera: Curculionidae), Annals of the Entomological Society of America, 61(1), 125-129.
BURKE, H.R. 1968. Pupae of the weevil tribe Anthonomini (Co-leoptera: Curculi- onidae). Texas Agri. Exp. Sta. Tech. Monog. 5, 92 p.
COAKLEY, J.M.; MAXWELL, F.G.; JENKINS, J.N. 1969. Influen-ce of feeding, oviposition, and egg and larval development of the boll weevil on abscission of cotton squares. Journal of Economic Entomology, 62, 244-245.
FIRMINO, A.A.P; FONSECA, F.C.A.; DE MACEDO, L.L.P.; COELHO, R.R.; ANTONINO DE SOUZA, JR.J.D.; TOGAWA, R.C.; SILVA-JUNIOR B.O.; PAPPAS, JR.G.J.; DA SILVA, M.C.M.; ENGLER, G.; DE SA, M.F.G. 2013. Transcriptome analysis in co-tton boll weevil (Anthonomus grandis) and RNA interference in insect pests. PLoS ONE 8(12), e85079. https://doi. org/10.1371/journal.pone.0085079
GREENBERG, S.M.; SAPPINGTON, T.W.; COLEMAN, R.J. 2004b. Population dynamics of overwintering boll weevils, Antho-nomus grandis grandis (Boheman) in the Lower Rio Grande Valley of Texas, Entomology Presentations, Posters and Proceedings. (Disponible: 4.https://lib.dr.iastate.edu/ent_conf/4 verificado: 21 de julio de 2019).
GREENBERG, S.M.; SAPPINGTON, T.W.; COLEMAN, R.J. 2004b. Population dynamics of overwintering boll weevils, Antho-nomus grandis grandis (Boheman) in the Lower Rio Grande Valley of Texas, Entomology Presentations, Posters and Proceedings. (Disponible: 4.https://lib.dr.iastate.edu/ent_conf/4).
GREENBERG, S.M.; SPURGEON, D.W.; SAPPINGTON, T.W.; SETAMOU, M. 2005. Size-dependent feeding and reproduction by boll weevil (Coleoptera: Curculionidae). Journal of Economic Ento-mology, 98(3), 749-756.
GRILLI, M.P.; BRUNO, M.A.; PEDEMONTE, M.L.; SHOWLER, A.T. 2012. Boll weevil invasion process in Argentina. Journal of Pest Science, 85, 47-54.
LANTERI, A.A.A.; CONFALONIERI, V.A.; SCATAGLINI, M.A. 2003. El picudo del algodonero en la Argentina: Principales resul-tados e implicancias de los estudios moleculares. Revista de la Sociedad Entomológica Argentina, 62, 1-15.
LEWIS, D.K.; SPURGEON, D.; SAPPINGTON, T.W.; KEELEY, L.L. 2002. A hexamerin protein, AgSP-1, is associated with diapause in the boll weevil. Journal of Insect Physiology, 48(9), 887-901.
MARENGO LOZADA, R.M.; WHITCOMB, W.H. 1993. Hospederas alternantes del picudo mejicano del algodonero (Anthonomus grandis Boh.). Ministerio Agricultura Ganadería. Asunción, Paraguay, 40 p.
MAYER, M.S.; BRAZZEL, J.F. 1963. The mating behavior of the boll weevil, Anthonomus grandis. Journal of Economic Entomolo-gy, 56(5), 605-609.
MONNERAT, R.G.; DIAS, S.C.; OLIVEIRA NETO, O.B. de; NO-BRE, S. D.; SILVA-WERNECK, J. O.; SÁ, M.F.G. de. 2000. Criação massal do bicudo do algodoeiro Anthonomus grandis em labora-tório. Brasília: Embrapa Recursos Genéticos e Biotecnologia, Co-municado Técnico, 4p.
R CORE TEAM. 2017. R: A language and environment for sta-tistical computing. R Foundation for Statistical Computing, Vienna, Austria. (Disponible: https://www.R-roject.org/ verificado: 31 de agosto de 2019).
SAPPINGTON, T.W.; SPURGEON, D.W. 2000. Preferred tech-nique for adult sex determination of the boll weevil, Anthonomus grandis Boheman (Coleoptera: Curculionidae). Annals of the Ento-mological Society of America, 93(3), 610-615.
SHOWLER, A.T. 2005. Relationships of different cotton square sizes to boll weevil (Coleoptera: Curculionidae) feeding and ovipo-sition in field conditions. Journal of Economic Entomology, 98(5), 1572-1579.
SHOWLER, A.T. 2008. Longevity and egg development of adult female boll weevils fed exclusively on different parts and stages of cotton fruiting bodies. Entomologia Experimentalis et Applicata, 127, 125-132.
SHOWLER, A.T.; CANTÚ, R.V. 2005. Intervals between boll weevil (Coleoptera: Curculionidae) oviposition and square abscis-sion, and development to adulthood in Lower Rio Grande Valley, Texas, field conditions. Southwestern Entomology, 30, 161-164.
SOSA, M.A.; VITTI, D.E.; ALMADA, M.S. 2009. Avance del pi-cudo del algodonero (Anthonomus grandis Boheman) (Coleoptera: Curculionidae) en el departamento General Obligado (Santa Fe). xiii Jornadas Fitosanitarias Argentinas. Fitosanidad responsable, base de la calidad de vida. Termas de Río Hondo. Santiago del Estero.
SPURGEON, D.W.; SAPPINGTON, T.W.; SUH, C.P.C. 2003. A system for characterizing reproductive and diapause morphology in the boll weevil (Coleoptera: Curculionidae), Annals of the Ento-mological Society of America, 96(1), 1-11.
STADLER, T.; BUTELER, M. 2007. Migration and dispersal of Anthonomus grandis (Coleoptera: Curculionidae) in South Amer-ica, Revista de la Sociedad Entomológica Argentina, 66, 205-217.

ROMANELLI, A.; DECUNDO, J.M.; MARTÍNEZ, G.; FERNANDEZ PAGGI, M.B.; PÉREZ GAUDIO, D.S.; DIEGUEZ, S.N.; SORACI, A. L.
Agosto 2020, Argentina 171171
Restricción del crecimiento intrauterino y peso del cerebro en relación con sus órganos vitales en lechones neonatos: consideraciones para la producción porcina
RESUMENLa restricción del crecimiento intrauterino (IUGR) se refiere al desarrollo insuficiente del embrión/feto y sus
órganos, particularmente sistema nervioso, generando lechones con bajo peso al nacimiento. La insuficiencia placentaria es su principal causa, adaptando la circulación fetal a favor del cerebro (brain sparing) para pre-servar el suministro de oxígeno y nutrientes. El objetivo de este trabajo fue evaluar la relación entre el peso de los lechones y el ratio peso cerebro/órganos vitales como indicador de insuficiencia placentaria. Se utilizaron ochenta (n=80) lechones de 35 cerdas (n=35) clínicamente sanas de primer parto. Se realizó la necropsia de todo neonato muerto durante el periparto hasta las 24 h de vida. Se registró sexo, peso del lechón y de sus órganos (hígado, corazón, pulmón, cerebro, riñones, intestino, bazo, estómago lleno y vacío). Se los clasificó como lechones normo peso (>1 kg), bajo peso (<1 kg) y restringidos (<1 kg y con cabezas tamaño relativo mayor que su cuerpo con forma semejante a un foco, exoftalmia u ojos saltones y hocico arrugado con plie-gues perpendiculares a la boca). Se calcularon las relaciones (ratios) peso del cerebro/órganos vitales y se determinaron las causas de muerte: nacido muerto, aplastado, inanición, y otras causas. Se analizaron las correlaciones entre el peso de los lechones y cada ratio a través del coeficiente de correlación de Spearman y se evaluó para cada ratio el efecto del grupo (normo peso, bajo peso y lechones IUGR) mediante ANOVA y test de Tukey. El peso promedio de los lechones bajo peso e IUGR fue 0,817±0,132 kg y 0,559±0,116 kg, respectivamente vs. los normo peso con 1,307±0,181 kg. Los ratios peso del cerebro/órganos vitales fueron significativamente mayores para lechones IUGR que para bajo peso y normo peso. Se observaron correla-ciones estadísticamente significativas (p<0,05) entre el peso de los lechones y todos los ratios con excepción del cerebro/bazo. Los nutrientes en fetos IUGR se destinan preferentemente al cerebro con un crecimiento asimétrico de los órganos vitales. La identificación de lechones IUGR asociado a insuficiencia placentaria, debería ser utilizada como herramienta para aplicar estrategias de manejo y selección genética que mejoren la productividad en granjas porcinas comerciales.
Palabras clave: crecimiento intrauterino retardado, crías porcinas, nutrición placentaria.
1Universidad Nacional del Centro de la Provincia de Buenos Aires, Facultad de Ciencias Veterinarias, Departamento de Fisiopatología (UNCPBA - FCV), Pinto 399, Tandil B7000, Argentina. Correo electrónico: [email protected] Nacional del Centro de la Provincia de Buenos Aires, Facultad de Ciencias Veterinarias, Departamento. de Producción Animal, Área Producción Porcina (UNCPBA - FCV), Pinto 399, Tandil B7000, Argentina.3Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Buenos Aires, Argentina.4Centro de Investigación Veterinaria de Tandil (CIVETAN), UNCPBA-CONICET-CICPBA, Facultad de Ciencias Veterinarias, Campus Universitario, (7000) Tandil, Buenos Aires, Argentina.5Comisión de Investigaciones Científicas de la Provincia de Buenos Aires (CIC-PBA), La Plata B1900, Buenos aires, Argentina.
Recibido 22 de octubre de 2019 // Aceptado 23 de abril de 2020 // Publicado online 07 de octubre de 2020
ROMANELLI, A.1,3,4; DECUNDO, J.M.1,3,4; MARTÍNEZ, G.1,3,4; FERNANDEZ PAGGI, M.B.2,3,4; PÉREZ GAUDIO, D.S.1,3,4; DIEGUEZ, S.N.1,5; SORACI, A. L.1,3,4

Restricción del crecimiento intrauterino y peso del cerebro en relación con sus órganos vitales en lechones neonatos: consideraciones (...)
COMUNICACIÓN CORTA RIA / Vol. 46 / N.º 2172172
INTRODUCCIÓN
La restricción del crecimiento intrauterino (IUGR) o creci-miento intrauterino retardado (CIUR) se refiere al desarrollo insuficiente del embrión/feto de los mamíferos o sus órga-nos durante la gestación (Wu et al., 2006). La insuficiencia placentaria es su principal causa (Pardi et al., 2002; Cohen et al., 2015), lo que conduce a una adaptación de la circu-lación fetal a favor de los órganos vitales, como el cerebro, conocido como brain sparing, una estrategia fetal para pre-servar el suministro de oxígeno y nutrientes para su desa-rrollo ante un menor flujo sanguíneo. Es una afección de importancia tanto en medicina humana como en medicina veterinaria, incluyendo la producción porcina (Dong et al., 2014; Ladinig et al., 2014). Alrededor del 5-10% de los neo-natos humanos y del 15-20% de los lechones de cerdas hiperprolíficas padecen de IUGR (Wang et al., 2018). Entre otras etiologías menos frecuentes de IUGR se pueden ci-tar anomalías cromosómicas fetales, infecciones fetales y anomalías estructurales (Krishna y Bhalerao, 2011).
En la especie porcina, especie politoca, los IUGR se diag-nostican clínicamente como los neonatos con un peso al nacimiento menor a la distribución normal de peso al naci-miento dentro de las camadas (Royston et al., 1982) y según criterios morfológicos de su cabeza (ovalada, tamaño rela-tivo mayor que su cuerpo, ojos saltones) y criterios de com-portamiento (aislamiento de sus compañeros y/o decúbito lateral) (Sacy et al., 2010). Estos animales presentan baja eficiencia de utilización y absorción de nutrientes provenien-tes de la placenta, especialmente de aminoácidos funciona-
les para el desarrollo fetoplacentario (Wang et al., 2018) con mayor susceptibilidad a padecer enfermedades y por con-siguiente mayor mortalidad. El bajo peso y heterogeneidad al nacimiento genera pérdidas antes y después del destete hasta el momento del sacrificio (Matheson et al., 2018) con menor crecimiento muscular y menores rendimientos de la canal (Wu et al., 2006), requiriendo 23 días más para llegar al peso de sacrificio de 110 kg (Town et al., 2005).
El peso al nacimiento puede ser usado como un criterio práctico para la detección de IUGR, ya que es un paráme-tro de fácil medición en las granjas de cría porcina. Sin em-bargo, por sí solo no alcanza para poder identificar estos animales ya que, si bien todos los IUGR son lechones de bajo peso, no todos los animales pequeños para la edad gestacional son IUGR. Por este motivo además del peso del lechón se deben considerar la morfología corporal, el índice de masa corporal y los ratios o relaciones del peso del cerebro/órganos vitales (Almeida, 2009; Ladinig et al., 2014; Sacy et al., 2010). Por lo tanto, el presente trabajo tuvo como objetivo evaluar la relación entre el peso de los lechones y el ratio peso cerebro/órganos vitales como indi-cador de insuficiencia placentaria.
MATERIALES Y MÉTODOS
El estudio se realizó siguiendo las normas éticas apro-badas por el Comité de Bienestar Animal de la Facultad de Ciencias Veterinarias, Universidad Nacional del Centro de la Provincia de Buenos Aires, Argentina. El trabajo experi-
ABSTRACT
Intrauterine growth restriction (IUGR) refers to the insufficient development of the embryo/fetus and its or-gans, resulting in low birth weights piglets. It is most commonly caused by placental insufficiency, in response to which the fetus adapts its circulation to preserve oxygen and nutrient supply to the brain (‘brain-sparing’). The aim of this study was to evaluate the relationship between piglets’ weight and brain/vital organ weight ratio, as an indicator of placental insufficiency. Eighty (n:80) piglets from 35 (n:35) healthy sows of first partum were used. All piglets which died during the peripartum and up to the first 24 h after birth were necropsied. Piglets gender, animal and organs weights (liver, heart, lungs, brain, kidneys, intestine, spleen, empty and full stomach) were noted. They were classified as normo-weight (> 1 kg), low-weight (< 1kg) and restricted piglets (< 1kg and with light bulb-like forehead, bulging eyes, wrinkles perpendicular to the mouth). Brain/organ weight ratios and causes of death: stillborn, crushed, dead due starvation and other causes were recorded. Spearman’s rank correlations were used to identify relationships between variables and ANOVA and Tukey Test was used to evaluate group effect (normo-weight, low-weight and IUGR) for each ratio. The mean weight of the low-weight and IUGR was 0,817±0,132 kg and 0,559±0,116 kg, respectively vs normo-weight with 1,307±0,181 kg. IUGR piglets showed significantly higher brain/vital organs ratios than low and normo-weight piglets. Statistically significant correlations (p<0,05) between piglets’ weights and ratios, except brain/spleen, were found. Nutrients in IUGR fetuses are preferable destined for the brain with asymmetric growth of vital organs. Identification of IUGR piglets associated with placental insufficiency, should be used as a tool to apply strategies of management and genetic selection to improve productivity in commercial pig farms.
Keywords: intrauterine growth restriction, piglets, placental nutrition.
ROMANELLI, A.; DECUNDO, J.M.; MARTÍNEZ, G.; FERNANDEZ PAGGI, M.B.; PÉREZ GAUDIO, D.S.; DIEGUEZ, S.N.; SORACI, A. L.
Agosto 2020, Argentina 173173
Tabla 1. Promedio ± DE del peso del lechón (kg) y de cada ratio cerebro/órganos vitales según el grupo (normo peso, bajo peso y restringi-dos o IUGR). Correlaciones (coeficiente de Spearman) halladas entre el peso del lechón (kg) y los distintos ratios cerebro/órganos vitales. *Todas las correlaciones mostraron ser estadísticamente significativas (p<0,05) a excepción del peso del lechón con el cerebro/bazo.
Figura 1. Ilustración de un lechón normal (izquierda) y un lechón restringido (IUGR) (derecha). Nótese las diferencias morfológicas de la cabeza con semejanza a un foco. (Adaptado de Hales et al., 2013).
mental se desarrolló en una granja comercial ubicada en el partido de Tandil, provincia de Buenos Aires. La explotación está organizada de manera intensiva, en confinamiento to-tal, con ciclo completo en un solo sitio con 400 hembras en producción. Se utilizaron ochenta (n=80) lechones de 35 (n=35) cerdas clínicamente sanas, de una misma línea genética y de primer parto las cuales fueron inducidas a los 114 días de gestación. Su alimentación consistió en una dieta comercial gestación (3,0 McalEM/kg-14% proteína), antes del parto y una dieta comercial lactancia (3,2 Mca-lEM/kg-16% proteína) durante la lactación.
Se realizó la necropsia de todo neonato muerto durante el periparto hasta las 24 h de vida, provenientes de dichas cerdas. Se registró sexo, peso del lechón y de sus órga-nos (hígado, corazón, pulmón, cerebro, riñones, intestino, bazo, estómago lleno y vacío). Se los clasificó como le-chones normo peso n=38 (>1 kg), bajo peso n=30 (<1 kg) y restringidos n=12 (<1 kg y con cabezas tamaño relati-vo mayor que su cuerpo con forma semejante a un foco,
exoftalmia u ojos saltones y hocico arrugado con pliegues perpendiculares a la boca) (Sacy et al., 2010) (figura 1).
Se calcularon los ratios peso del cerebro/peso de órga-nos vitales y se determinaron las causas de muerte según las categorías: “nacido muerto”, lechones cuyos pulmones no demostraron flotación en el agua por falta de oxigena-ción del tejido pulmonar; “aplastado”, lechones con signos de trauma y edema subcutáneo, en alguna parte del cuer-po; “inanición”, lechones sin evidencia de aplastamiento, pero con emaciación marcada, sin o poco contenido en el estómago/intestino; y “otras causas”, lechones que mu-rieron por afecciones que no pudieron determinarse con exactitud mediante el examen post mórtem.
La normalidad de la distribución fue evaluada con el test de Shapiro-Wilk. Para analizar la asociación entre variables se utilizó la prueba de correlación de Pearson o Spearman según corresponda. Se llevó a cabo un análisis de varian-za ANOVA para evaluar el efecto grupo (normo peso, bajo peso y restringido) sobre cada ratio considerado, posterior-
Grupo Peso (kg)
RATIOS
Cerebro/Hígado
Cerebro/Corazón
Cerebro/Pulmón
Cerebro/Intestino
Cerebro/Riñones
Cerebro/Estómago
lleno
Cerebro/Estómago
vacío
Cerebro/Bazo
Normo peso 1,307±0,181 0,789±0,365 2,599±0,740 1,045±0,350 0,423±0,143 1,989±0,746 1,905±1,564 3,339±1,153 23,217±8,249
Bajo peso 0,817±0,132 1,190±0,399 3,455±1,010 1,419±0,463 0,731±0,235 2,901±0,750 3,415±2,269 5,192±2,471 26,500±6,615
Restringidos 0,559±0,116 2,002±0,742 5,047±1,180 2,456±0,686 1,028±0,178 3,868±1,460 5,895±1,353 6,800±2,032 26,667±5,211
Correlación Peso (kg) -0,7276* -0,682* -0,6996* -0,8245* -0,7037* -0,5717* -0,6494* -0,1631
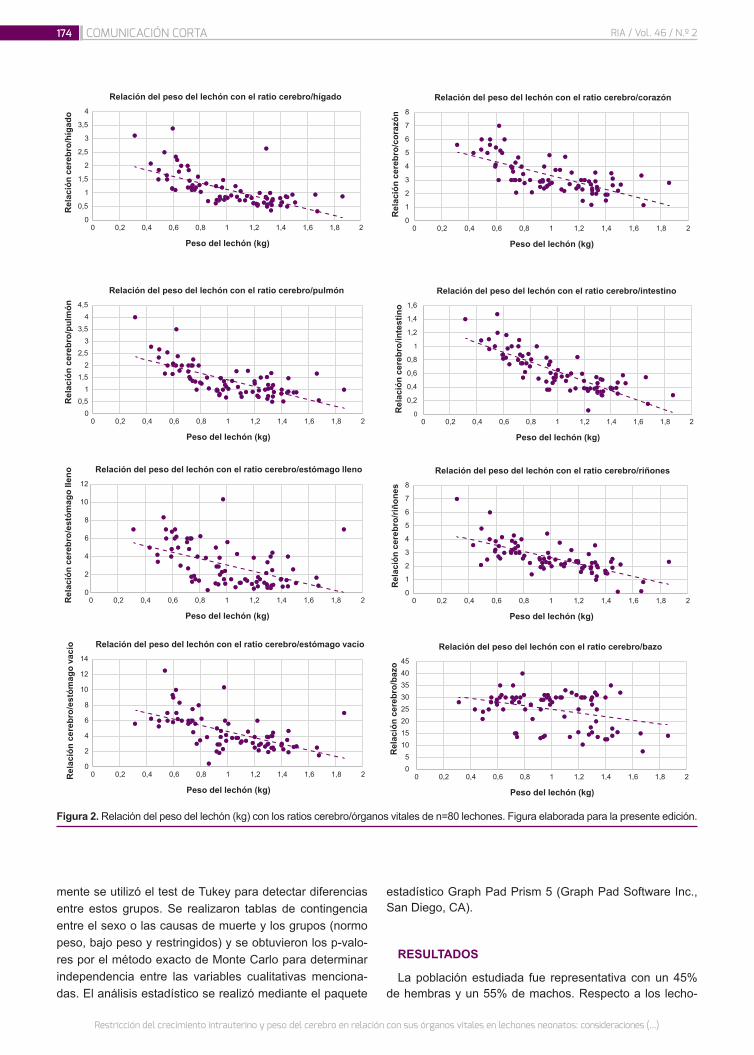
Restricción del crecimiento intrauterino y peso del cerebro en relación con sus órganos vitales en lechones neonatos: consideraciones (...)
COMUNICACIÓN CORTA RIA / Vol. 46 / N.º 2174174
Figura 2. Relación del peso del lechón (kg) con los ratios cerebro/órganos vitales de n=80 lechones. Figura elaborada para la presente edición.
0
0,5
1
1,5
2
2,5
3
3,5
4
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 2
Rel
ació
n ce
rebr
o/hí
gado
Peso del lechón (kg)
Relación del peso del lechón con el ratio cerebro/hígado
0
1
2
3
4
5
6
7
8
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 2
Rel
ació
n ce
rebr
o/co
razó
n
Peso del lechón (kg)
Relación del peso del lechón con el ratio cerebro/corazón
00,5
11,5
22,5
33,5
44,5
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 2
Rel
ació
n ce
rebr
o/pu
lmón
Peso del lechón (kg)
Relación del peso del lechón con el ratio cerebro/pulmón
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 2
Rel
ació
n ce
rebr
o/in
test
ino
Peso del lechón (kg)
Relación del peso del lechón con el ratio cerebro/intestino
0
1
2
3
4
5
6
7
8
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 2
Rel
ació
n ce
rebr
o/riñ
ones
Peso del lechón (kg)
Relación del peso del lechón con el ratio cerebro/riñones
0
2
4
6
8
10
12
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 2Rel
ació
n ce
rebr
o/es
tóm
ago
lleno
Peso del lechón (kg)
Relación del peso del lechón con el ratio cerebro/estómago lleno
0
2
4
6
8
10
12
14
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 2Rel
ació
n ce
rebr
o/es
tóm
ago
vací
o
Peso del lechón (kg)
Relación del peso del lechón con el ratio cerebro/estómago vacío
05
1015202530354045
0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 2
Rel
ació
n ce
rebr
o/ba
zo
Peso del lechón (kg)
Relación del peso del lechón con el ratio cerebro/bazo
mente se utilizó el test de Tukey para detectar diferencias entre estos grupos. Se realizaron tablas de contingencia entre el sexo o las causas de muerte y los grupos (normo peso, bajo peso y restringidos) y se obtuvieron los p-valo-res por el método exacto de Monte Carlo para determinar independencia entre las variables cualitativas menciona-das. El análisis estadístico se realizó mediante el paquete
estadístico Graph Pad Prism 5 (Graph Pad Software Inc., San Diego, CA).
RESULTADOS
La población estudiada fue representativa con un 45% de hembras y un 55% de machos. Respecto a los lecho-
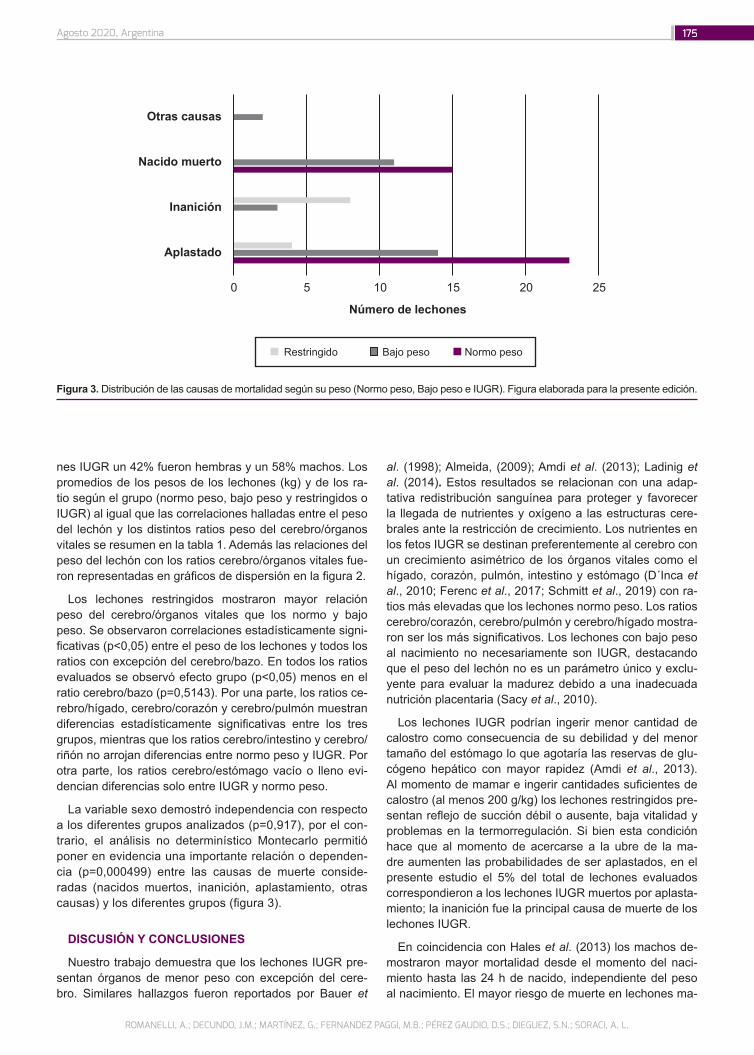
ROMANELLI, A.; DECUNDO, J.M.; MARTÍNEZ, G.; FERNANDEZ PAGGI, M.B.; PÉREZ GAUDIO, D.S.; DIEGUEZ, S.N.; SORACI, A. L.
Agosto 2020, Argentina 175175
nes IUGR un 42% fueron hembras y un 58% machos. Los promedios de los pesos de los lechones (kg) y de los ra-tio según el grupo (normo peso, bajo peso y restringidos o IUGR) al igual que las correlaciones halladas entre el peso del lechón y los distintos ratios peso del cerebro/órganos vitales se resumen en la tabla 1. Además las relaciones del peso del lechón con los ratios cerebro/órganos vitales fue-ron representadas en gráficos de dispersión en la figura 2.
Los lechones restringidos mostraron mayor relación peso del cerebro/órganos vitales que los normo y bajo peso. Se observaron correlaciones estadísticamente signi-ficativas (p<0,05) entre el peso de los lechones y todos los ratios con excepción del cerebro/bazo. En todos los ratios evaluados se observó efecto grupo (p<0,05) menos en el ratio cerebro/bazo (p=0,5143). Por una parte, los ratios ce-rebro/hígado, cerebro/corazón y cerebro/pulmón muestran diferencias estadísticamente significativas entre los tres grupos, mientras que los ratios cerebro/intestino y cerebro/riñón no arrojan diferencias entre normo peso y IUGR. Por otra parte, los ratios cerebro/estómago vacío o lleno evi-dencian diferencias solo entre IUGR y normo peso.
La variable sexo demostró independencia con respecto a los diferentes grupos analizados (p=0,917), por el con-trario, el análisis no determinístico Montecarlo permitió poner en evidencia una importante relación o dependen-cia (p=0,000499) entre las causas de muerte conside-radas (nacidos muertos, inanición, aplastamiento, otras causas) y los diferentes grupos (figura 3).
DISCUSIÓN Y CONCLUSIONES
Nuestro trabajo demuestra que los lechones IUGR pre-sentan órganos de menor peso con excepción del cere-bro. Similares hallazgos fueron reportados por Bauer et
al. (1998); Almeida, (2009); Amdi et al. (2013); Ladinig et al. (2014). Estos resultados se relacionan con una adap-tativa redistribución sanguínea para proteger y favorecer la llegada de nutrientes y oxígeno a las estructuras cere-brales ante la restricción de crecimiento. Los nutrientes en los fetos IUGR se destinan preferentemente al cerebro con un crecimiento asimétrico de los órganos vitales como el hígado, corazón, pulmón, intestino y estómago (D´Inca et al., 2010; Ferenc et al., 2017; Schmitt et al., 2019) con ra-tios más elevadas que los lechones normo peso. Los ratios cerebro/corazón, cerebro/pulmón y cerebro/hígado mostra-ron ser los más significativos. Los lechones con bajo peso al nacimiento no necesariamente son IUGR, destacando que el peso del lechón no es un parámetro único y exclu-yente para evaluar la madurez debido a una inadecuada nutrición placentaria (Sacy et al., 2010).
Los lechones IUGR podrían ingerir menor cantidad de calostro como consecuencia de su debilidad y del menor tamaño del estómago lo que agotaría las reservas de glu-cógeno hepático con mayor rapidez (Amdi et al., 2013). Al momento de mamar e ingerir cantidades suficientes de calostro (al menos 200 g/kg) los lechones restringidos pre-sentan reflejo de succión débil o ausente, baja vitalidad y problemas en la termorregulación. Si bien esta condición hace que al momento de acercarse a la ubre de la ma-dre aumenten las probabilidades de ser aplastados, en el presente estudio el 5% del total de lechones evaluados correspondieron a los lechones IUGR muertos por aplasta-miento; la inanición fue la principal causa de muerte de los lechones IUGR.
En coincidencia con Hales et al. (2013) los machos de-mostraron mayor mortalidad desde el momento del naci-miento hasta las 24 h de nacido, independiente del peso al nacimiento. El mayor riesgo de muerte en lechones ma-
Figura 3. Distribución de las causas de mortalidad según su peso (Normo peso, Bajo peso e IUGR). Figura elaborada para la presente edición.
0 5 10 15 20 25
Aplastado
Inanición
Nacido muerto
Otras causas
Número de lechones
Restringido Bajo peso Normo peso

Restricción del crecimiento intrauterino y peso del cerebro en relación con sus órganos vitales en lechones neonatos: consideraciones (...)
COMUNICACIÓN CORTA RIA / Vol. 46 / N.º 2176176
chos podría ser causa de los niveles de testosterona aso-ciado con rápidos cambios metabólicos. El aumento de tes-tosterona en los lechones machos incrementa la energía para los procesos de degradación y producción proteica, con una disponibilidad menor de energía para regular la temperatura corporal e ingerir calostro. (Bate et al., 1985; Hales et al., 2013).
Finalmente, los resultados de este trabajo sugieren un punto de partida para mejorar la productividad de los sis-temas de cría porcina dado que los lechones IUGR oca-sionan costos adicionales asociados a mortalidad, manejo, retraso en el flujo de la cadena de producción. La eva-luación regular de las camadas para identificar lechones IUGR asociado a insuficiencia placentaria debería ser una herramienta para aplicar estrategias de manejo y selección genética que mejoren la productividad en granjas porcinas comerciales.
BIBLIOGRAFÍAALMEIDA, F.R.C.L. 2009. Influência da nutrição da fêmea so-
bre a qualidade do leitão ao nascer. Acta Scientiae Veterinariae, 37(Supl 1), 31-34.
AMDI, C.; KROGH, U.; FLUMMER, C.; OKSBJERG, N.; HAN-SEN, C.F.; THEI, P.K. 2013. Intrauterine growth restricted piglets defined by their head shape ingest insufficient amounts of colos-trum. Journal of Animal Science, 91(12), 5605-5613. https://doi.org/10.2527/jas2013-6824
BATE, L.A.; HACKER, R.R.; KREUKNIET, M.B. 1985. The rela-tionship between serum testosterone levels, sex and teat-seeking ability of newborn piglets. Canadian Journal of Animal Science, 65(3), 627-630.
BAUER, R.; WALTER, B.; HOPPE, A.; GASER, E.; LAMPE, V.; KAUF, E.; ZWIENER, U. 1998. Body weight distribution and organ size in newborn swine (sus scrofa domestica) – a study describing an animal model for asymmetrical intrauterine growth retardation. Experimental and Toxicologic Pathology, 50(1), 59-65. https://doi.org/10.1016/S0940-2993(98)80071-7
COHEN, E.; BAERTS, W.; VAN BEL, F. 2015. Brain-Sparing in In-trauterine Growth Restriction: Considerations for the Neonatologist. Neonatology, 108(4), 269-276. https://doi.org/10.1159/000438451
CHE, L.; YANG, Z.; XU, M.; XU, S.; CHE, L.; LIN, Y.; FANG, Z.; FENG, B.; LI, J.; CHEN, D.; WU, D. 2017. Maternal nutrition modu-lates fetal development by inducing placental efficiency changes in gilts. BMC Genomics, 18(1), 213. https://doi.org/10.1186/s12864-017-3601-1
D’INCA, R.; GRAS-LE GUEN, C.; CHE, L.; SANGILD, P.T.; LE HUËROU-LURON, I. 2010. Intrauterine growth restriction delays feeding-induced gut adaptation in term newborn pigs. Neonatolo-gy, 99(3), 208-216. https://doi.org/10.1159/000314919
DONG, L.; ZHONG, X.; AHMAD, H.; LI, W.; WANG, Y.; ZHANG, L.; WANG, T. 2014. Intrauterine Growth Restriction Impairs Small Intestinal Mucosal Immunity in Neonatal Piglets. Journal of Histochemistry and Cytochemistry, 62(7), 510-518. https://doi.org/10.1369/0022155414532655
FERENC, K.; PILZYS, T.; SKRZYPEK, T.; GARBICZ, D.; MARCINKOWSKI, M.; DYLEWSKA, M.; ZABIELSKI, R. 2017. Structure and Function of Enterocyte in Intrauterine
Growth Retarded Pig Neonates. Disease Marker. https://doi.org/10.1155/2017/5238134
FERENC, K.; PIETRZAK, P.; WIERZBICKA, M.; MATYBA, P.; GRZESIUK, E.; GAJEWSKI, Z.; ZABIELSKI, R. 2018. Alterations in the liver of intrauterine growth retarded piglets may predispose to development of insulin resistance and obesity in later life. Jour-nal of Physiology and Pharmacology, 69(2), 211-218. https://doi.org/10.26402/jpp.2018.2.06
HALES, J.; MOUSTSEN, V.A.; NIELSEN, M.B.F.; HANSEN, C.F. 2013. Individual physical characteristics of neonatal piglets affect preweaning survival of piglets born in a noncrated system. Jour-nal of Animal Science, 91(10), 4991-5003. https://doi.org/10.2527/jas2012-5740
KRISHNA, U.; BHALERAO, S. 2011. Placental insufficiency and fetal growth restriction. The Journal of Obstetrics and Gynecology of India, 61(5), 505-511.
LADINIG, A.; FOXCROFT, G.; ASHLEY, C.; LUNNEY, J.K.; PLASTOW, G.; HARDING, J.C.S. 2014. Birth weight, intrauterine growth retardation and fetal susceptibility to porcine reproductive and respiratory syndrome virus. PLoS ONE, 9(10). https://doi.org/10.1371/journal.pone.0109541
LIN, G.; WANG, X.; WU, G.; FENG, C.; ZHOU, H.; LI, D.; WANG, J. 2014. Improving amino acid nutrition to prevent intrauterine growth restriction in mammals. Amino Acids, 46(7), 1605-1623. https://doi.org/10.1007/s00726-014-1725-z
MATHESON, S.M.; WALLING, G.A.; EDWARDS, S.A. 2018. Genetic selection against intrauterine growth retardation in piglets: a problem at the piglet level with a solution at the sow level. Ge-netics Selection Evolution, 50, 46. https://doi.org/10.1186/s12711-018-0417-7
PARDI, G.; MARCONI, A.M.; CETIN, I. 2002. Placental-fetal in-terrelationship in IUGR fetuses - A review. Placenta, 23(SUPPL. 1), 136-141. https://doi.org/10.1053/plac.2002.0802
ROYSTON, J.P.; FLECKNELL, P.A.; WOOTTON, R. 1982. New evidence that the intrauterine growth-retarded piglet is a member of a discrete subpopulation. Biology of the Neonate, 42(1-2), 100-104.
SACY, A.; TREUT, Y.L.E.; SCHMIDELY, P.; CHEVAUX, E. 2010. Caractérisation de l’ immaturité des porcelets à la naissance. Journees Recherche Porcine, 259-260.
SCHMITT, O.; O’DRISCOLL, K.; BAXTER, E.M. 2019. Explor-atory study of the effects of intrauterine growth retardation and neonatal energy supplementation of low birth-weight piglets on their post-weaning cognitive abilities. Animal Cognition, 0(0), 0. https://doi.org/10.1007/s10071-019-01251-8
TOWN, S.C.; PATTERSON, J.L.; PEREIRA, C.Z.; GOUR-LEY, G.; FOXCROFT, G.R. 2005. Embryonic and fetal develop-ment in a commercial dam-line genotype. Animal Reproduction Science, 85(3-4), 301-316. https://doi.org/10.1016/j.anirepros-ci.2004.05.019
WANG, J.; CHEN, L.; LI, D.; YIN, Y.; WANG, X.; LI, P.; DAN-GOTT, L.J.; HU, W.; WU, G. 2018. Intrauterine Growth Restriction Affects the Proteomes of the Small Intestine, Liver, and Skeletal Muscle in Newborn Pigs. The Journal of Nutrition, 138(1), 60-66. https://doi.org/10.1093/jn/138.1.60
WU, G.; BAZER, F.W.; WALLACE, J.M.; SPENCER, T.E. 2006. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. Journal of Animal Science, 84(9), 2316-2337. https://doi.org/10.2527/jas.2006-156

MARANGI, M.J.; TEMPERINI, C.V.; FERNÁNDEZ, D.; PARDO, A.G.; POSE, G.N.
Agosto 2020, Argentina 177177
First report of Bacterial Spot with gum production in almond fruits of Argentina
ABSTRACTBacterial Spot of Almond is a pathology caused by Xanthomonas arboricola pv. pruni responsible for sig-
nificant yield losses. During 2017, young symptomatic almond fruits were observed in orchards of Northern Patagonia, Argentina. Symptoms consisted of small watery circular lesions, from which an amber gummy substance emerged. The objective of this work was to determine the etiology of this pathology considering that the symptoms were coincident with those previously described for the bacterial spot with gum production caused by Xanthomonas arboricola pv. pruni on almond fruits in other parts of the world. The presence of Xanthomonas arboricola in damaged fruits was determined confirming their identity by molecular analysis. Symptomatology and gum production were demonstrated by performing pathogenicity tests. As a result, Bac-terial Spot with gum production is reported for the first time on almond fruits in Argentina.
Keywords: Bacterial Spot, amber gum, almond, Xanthomonas arboricola pv. pruni.
RESUMEN
La mancha bacteriana de la almendra es una patología causada por Xanthomonas arborícola pv. pruni y es responsable de importantes pérdidas en el rendimiento productivo. Durante 2017, en plantaciones del norte de la Patagonia Argentina, se observaron frutos jóvenes de almendro con pequeñas lesiones circulares, acuosas, de las cuales emergía una sustancia ámbar gomosa. El objetivo de este trabajo fue determinar la causa de esta patología, considerando que los síntomas eran coincidentes con los previamente descriptos para la mancha bacteriana con producción de goma, causada por Xanthomonas arborícola pv. pruni en frutos de almendro en otras partes del mundo. Se determinó la presencia de Xanthomonas arborícola en los frutos lesionados confirmándose la identidad del patógeno por análisis molecular. A través de pruebas de patoge-nicidad se demostró la sintomatología y la producción de goma en frutos sanos inoculados con la bacteria. A partir del estudio realizado se reporta por primera vez la mancha bacteriana con producción de goma en frutos de almendro en Argentina.
Palabras clave: mancha bacteriana, goma ámbar, almendra, Xanthomonas arborícola pv. pruni.
1Universidad Nacional de Río Negro, Río Negro, Argentina. Mitre 331, (8336) Villa Regina, provincia de Río Negro, Argentina. Correo electrónico: [email protected] 2Universidad Nacional de Quilmes, Buenos Aires, Argentina. Roque Sáenz Peña 352, (B1876) Bernal, provincia de Buenos Aires, Argentina.3Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Alto Valle, Agencia de Extensión Rural (AER) Valle Medio. Villa Galense 575, (8361) Luis Beltrán, Río Negro.4Consejo Nacional de Investigaciones Científicas y Tecnológicas (CONICET), Argentina.aInstitución actual: Universidad Nacional de Quilmes, Laboratorio de Micología y Cultivo de Hongos Comestibles - INTECH (CONICET).
Received September 10th 2019// Accepted May 27th 2020 // Published online November 04th 2020
MARANGI, M.J.1,4; TEMPERINI, C.V.1,4; FERNÁNDEZ, D.3; PARDO, A.G.2,4; POSE, G.N.1,4,a

First report of Bacterial Spot with gum production in almond fruits of Argentina
COMUNICACIÓN CORTA RIA / Vol. 46 / N.º 2178178
INTRODUCTION
Bacterial Spot of Almond is a pathology caused by Xan-thomonas arboricola pv. pruni. This disease occurs on leaves, branches and fruits of almost all Prunus species. On almond trees, it can be observed on leaves and buds, but the most evident symptoms are observed on fruits (Adaskaveg et al., 2015). Regarding symptoms on immature fruits, lesions begin in the shell as small watery brown spots that slowly increase in diameter (2 to 4 mm, generally smaller than 5 mm). These le-sions produce a light to dark amber gummy substance, whose color is important because it helps to distinguish bacterial spot from other types of damage (Adaskaveg et al., 2015).
Bacterial Spot caused by Xanthomonas arboricola pv. pruni was described for the first time in the United States on Jap-anese plum (Smith, 1903) and, lately, it is being reported in almost every continent where stone fruits are grown (EPPO, 2006). It has been determined on almond trees in Japan (Ishi-yama, 1923), Sicily (Italy) (Ciccarone, 1959), New Zealand (Young, 1977), Spain (Palacio-Bielsa et al., 2010), Australia and California (United States) (Adaskaveg et al., 2015). Previ-ous studies indicate that this pathogen led to reduction in the quality and marketability of fruit, reduction in orchard produc-tivity and increases in production costs, resulting in significant economic losses (Lamichhane, 2014).
The leading almond producer in the world is the Unit-ed States, followed by Spain. Also, Australia emerges as a strong growing producer. In Argentina, in recent years, the sector has been recovered and modern plantations with late bloom varieties were incorporated (Iannamico, 2015). Currently, there are about 4,200 cultivated hectares, mainly distributed in the provinces of Mendoza, San Juan, La Rio-ja, Salta, Río Negro and Neuquén. In Río Negro (Northern Patagonia), growing areas are mainly located in the High Valley productive region, while smaller orchards are located
in the Lower Valley and Middle Valley productive regions (Iannamico, 2015). Although the almond trees integrate a group of fruit trees that are not traditionally grown in North-ern Patagonia, it has enormous potential due to its agro-nomic behavior and its good profitability and growth possi-bilities (Iannamico, 2015).
During October 2017 in cultivars of Luis Beltrán, a pro-ductive region of the Middle Valley of Río Negro, young al-mond fruits with external lesions were observed. Symptoms consisted of small watery circular lesions, from which an amber gummy substance emerged (fig. 1). The objective of this work was to determine the etiology of this pathology, considering that the symptoms were coincident with those previously described for diseases caused by Xanthomonas arboricola pv. pruni and the fact that Xanthomonas arbori-cola pv. juglandis had previously been reported in the re-gion as a causal agent of Walnut Blight and Brown Apical Necrosis (Temperini et al., 2017).
MATERIALS AND METHODS
A total of 5 symptomatic immature almond fruits were chosen for analysis. The fruits were washed with tap wa-ter, superficially disinfected by soaking them in a solution of 1% Sodium Hypochlorite (NaClO) for 3 minutes, and rinsed twice with sterile distilled water. Then, they were placed in-dividually in closed plastic jars, containing a piece of ster-ilized cotton soaked in water to create a wet chamber to allow microbial development. They were incubated at 27 °C for 14 days. After the incubation period, internal pieces of injured tissue were aseptically removed and placed in Ep-pendorf tubes containing sterile physiological solution. After homogenization, 0.1 ml of the suspension was inoculated in Petri dishes containing Luria Bertani (LB) medium. They were incubated at 27 °C for 4 days. Colonies that showed morphological characteristics equal or similar to those de-scribed for the genus Xanthomonas on LB medium were cultured on a differential medium for this genus, Xan-D (Yung-Ann et al., 2009). They were incubated at 27 °C for 7 days. The colonies that were positive on the Xan-D medium were confirmed by molecular techniques.
DNA extractions were performed using the “DNeasy blood and tissue mini kit” with a pre-treatment protocol for gram-negative bacteria as for the manufacturer’s instruc-tions (Qiagen, Intl.). The extracted genomic DNA was quan-tified with a fluorimeter Qubit 2.0 (Life Technologies, Intl.). PCR amplifications were performed using Xanthomonas species-specific primers XarbQF (GCGAGATCAATGC-GACCTCGTC) and XarbQR (GGTGACCACATCGAACCG-CGCA) for qumA gene, a quinate dehydrogenase (Pothier et al., 2011), according to Temperini et al. (2017). After mo-lecular identification, Xanthomonas isolates were stored at -80 °C in broth Yeast Extract Peptone Dextrose.
Pathogenicity tests were conducted on 8 healthy im-mature almonds fruits, 40 days after the setting, and the results were verified according to Koch’s postulates. The disinfected fruits were injured at a depth of 0.1 mm in the
Figure 1. External symptoms of the disease on an immature al-mond fruit.
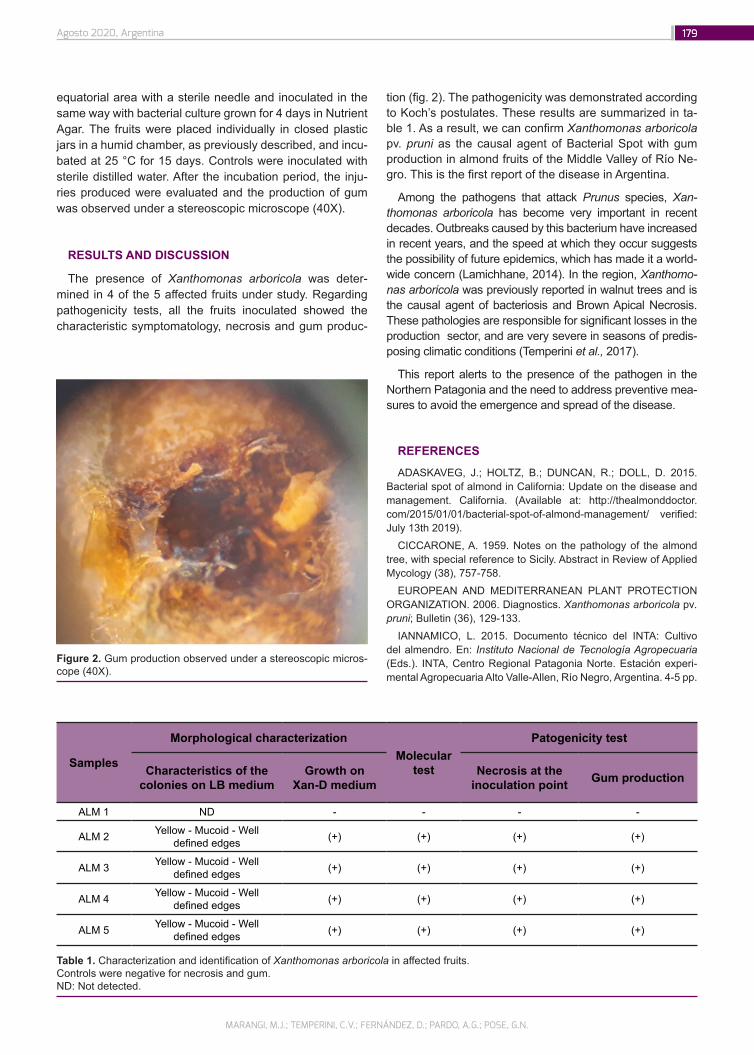
MARANGI, M.J.; TEMPERINI, C.V.; FERNÁNDEZ, D.; PARDO, A.G.; POSE, G.N.
Agosto 2020, Argentina 179179
equatorial area with a sterile needle and inoculated in the same way with bacterial culture grown for 4 days in Nutrient Agar. The fruits were placed individually in closed plastic jars in a humid chamber, as previously described, and incu-bated at 25 °C for 15 days. Controls were inoculated with sterile distilled water. After the incubation period, the inju-ries produced were evaluated and the production of gum was observed under a stereoscopic microscope (40X).
RESULTS AND DISCUSSION
The presence of Xanthomonas arboricola was deter-mined in 4 of the 5 affected fruits under study. Regarding pathogenicity tests, all the fruits inoculated showed the characteristic symptomatology, necrosis and gum produc-
Figure 2. Gum production observed under a stereoscopic micros-cope (40X).
Table 1. Characterization and identification of Xanthomonas arboricola in affected fruits. Controls were negative for necrosis and gum.ND: Not detected.
Samples
Morphological characterizationMolecular
test
Patogenicity test
Characteristics of the colonies on LB medium
Growth on Xan-D medium
Necrosis at the inoculation point Gum production
ALM 1 ND - - - -
ALM 2 Yellow - Mucoid - Well defined edges (+) (+) (+) (+)
ALM 3 Yellow - Mucoid - Well defined edges (+) (+) (+) (+)
ALM 4 Yellow - Mucoid - Well defined edges (+) (+) (+) (+)
ALM 5 Yellow - Mucoid - Well defined edges (+) (+) (+) (+)
tion (fig. 2). The pathogenicity was demonstrated according to Koch’s postulates. These results are summarized in ta-ble 1. As a result, we can confirm Xanthomonas arboricola pv. pruni as the causal agent of Bacterial Spot with gum production in almond fruits of the Middle Valley of Río Ne-gro. This is the first report of the disease in Argentina.
Among the pathogens that attack Prunus species, Xan-thomonas arboricola has become very important in recent decades. Outbreaks caused by this bacterium have increased in recent years, and the speed at which they occur suggests the possibility of future epidemics, which has made it a world-wide concern (Lamichhane, 2014). In the region, Xanthomo-nas arboricola was previously reported in walnut trees and is the causal agent of bacteriosis and Brown Apical Necrosis. These pathologies are responsible for significant losses in the production sector, and are very severe in seasons of predis-posing climatic conditions (Temperini et al., 2017).
This report alerts to the presence of the pathogen in the Northern Patagonia and the need to address preventive mea-sures to avoid the emergence and spread of the disease.
REFERENCESADASKAVEG, J.; HOLTZ, B.; DUNCAN, R.; DOLL, D. 2015.
Bacterial spot of almond in California: Update on the disease and management. California. (Available at: http://thealmonddoctor. com/2015/01/01/bacterial-spot-of-almond-management/ verified: July 13th 2019).
CICCARONE, A. 1959. Notes on the pathology of the almond tree, with special reference to Sicily. Abstract in Review of Applied Mycology (38), 757-758.
EUROPEAN AND MEDITERRANEAN PLANT PROTECTION ORGANIZATION. 2006. Diagnostics. Xanthomonas arboricola pv. pruni; Bulletin (36), 129-133.
IANNAMICO, L. 2015. Documento técnico del INTA: Cultivo del almendro. En: Instituto Nacional de Tecnología Agropecuaria (Eds.). INTA, Centro Regional Patagonia Norte. Estación experi-mental Agropecuaria Alto Valle-Allen, Río Negro, Argentina. 4-5 pp.

First report of Bacterial Spot with gum production in almond fruits of Argentina
COMUNICACIÓN CORTA RIA / Vol. 46 / N.º 2180180
ISHIYAMA, S. 1923. Studies of blackspot disease of plum. Japa-nese Journal of Botany (1), 21.
LAMICHHANE, J.R. 2014. Xanthomonas arboricola disease of stone fruit, almond, and walnut tres: Progress toward understand-ing and management. Plant Disease (98), 1600-1610.
PALACIO-BIELSA, A.; ROSELLÓ, M.; CAMBRA, M.A.; LÓPEZ, M.M. 2010. First report on almond in Europe of bacterial spot dis-ease of stone fruits caused by Xanthomonas arboricola pv. pruni. Plant Disease (94), 786.
POTHIER, J.F.; PAGANI, M.C.; PELLUDAT, C.; RITCHIE, D.F.; DUFFY, B. 2011. A duplex - PCR method for species - and pa-thovar - level identification and detection of the quarantine plant pathogen Xanthomonas arboricola pv. pruni. Journal of microbiolo-gical methods (86), 16-24.
SMITH, E.F. 1903. Observations on a hither to unreported bac-terial disease, the cause of which enters the plant through ordinary stomata. Science (17), 456-457.
TEMPERINI, C.V.; PARDO, A.G.; POSE, G.N. 2017. First re-port of apical necrosis in walnut cultivars from Northern Argen-tinean Patagonia. Journal of Plant Pathology and Microbiology (8), 414-419.
YOUNG, J.M. 1977. Xanthomonas pruni in almond in New Zea-land. New Zealand Journal of Agricultural Research (20), 105-107.
YUNG-ANN, L.; AI-NING, S.; TZU-FEN, L.; YUNG-SHAN, L. 2009. Combination of chromogenic differential médium and es-tA-specific PCR for isolation and detection of Phytopathogenic Xanthomonas spp. Applied and Environmental Microbiology (75) 6831-6838.

MARTÍNEZ, G.M.; MICHELOUD, J.F.; SUÁREZ, V.H.; ROSA, D.E.; VENTURA, M.V.; ORTEGA, A.; SÁNCHEZ, C.; MATTIOLI, G.A.
Agosto 2020, Argentina 181181
Niveles plasmáticos de calcio, magnesio y fósforo en rodeos lecheros del Valle de Lerma, Salta, Argentina
RESUMENLa cuenca lechera del Valle de Lerma representa una de las economías regionales más relevantes para la
provincia de Salta. Si bien es conocida la importancia que tienen macrominerales como calcio (Ca), fósforo (P) y magnesio (Mg) sobre la producción y la salud de los rodeos lecheros, se desconoce la prevalencia de sus desequilibrios. Con el propósito de conocer estos datos se muestrearon 584 vacas de 4 tambos del Valle de Lerma, en 4 oportunidades y a intervalos de 3 a 4 meses entre sí. En cada visita se sangraron por punción yugular entre 9 y 10 animales de cada categoría: preparto, vaca fresca, 2.º tercio de lactancia y 3.º tercio de lactancia. Se determinaron los valores de Ca y Mg sérico por espectrofotometría de absorción atómica y P por espectrofotometría UV-visible. Para los análisis estadísticos los valores de referencia para considerar hipocal-cemia clínica y subclínica fueron <5,5 y de 5,5 a <8,5 mg/dl respectivamente. Para el caso del magnesio se consideró hipomagnesémicos los valores inferiores a 1,8 mg/dl. Los valores de referencia para hipofosfate-mia, e hiperfosfatemia fueron de <4 y >6,18 mg/dl respectivamente. Las asociaciones entre variables fueron analizadas mediante correlación lineal (Pearson) y regresión lineal simple, además de analizar la probabilidad de ocurrencia de hipocalcemia mediante odds ratio. Las diferencias entre variables y periodos de lactancia se examinaron mediante los análisis de varianza y test de Tukey y de chi cuadrado usando el test de Fisher. La media del Ca sérico fue de 8,85 mg/dl, sin hallarse diferencias entre periodos de lactancia. La prevalencia media de hipocalcemia fue del 46,2% en todos los periodos productivos. La media del Mg sérico fue de 2,26%, con una prevalencia de hipomagnesemia del 10,6% y una mayor proporción de vacas hipomagnesémicas al preparto. Los animales con hipomagnesemia presentaron una probabilidad 3,5 veces mayores de sufrir hipo-calcemia respecto a aquellos con valores normales. La prevalencia de animales con valores anormales de P fue elevada (48,3%), y si bien no se asociaron con los niveles de hipocalcemia, demostraron que el excesivo aporte dietario representa un riesgo sanitario y también medioambiental. En virtud a los resultados obtenidos en el presente trabajo se advierte de los riesgos potenciales de desequilibrios de Ca, Mg y P en los rodeos lecheros del Valle de Lerma.
Palabras clave: desbalance, minerales, vacas lecheras.
ABSTRACT
The dairy production of the Lerma Valley is one of the most relevant regional economies of the province of Salta. In order to known imbalances prevalence of calcium (Ca), phosphorus (P) and magnesium (Mg), 584 dairy cows of 4 farms of basin Lerma Valley were sampled in 4 opportunities. At each visit were bled from the
1Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Salta, RN 68 km 172 (4403) Cerrillos, Salta. Correo electrónico: [email protected] Nacional de Tecnología Agropecuaria (INTA), Instituto de investigación animal para el Chaco Semiárido, Área de Salud Animal.3Universidad Nacional de la Plata, Facultad de Veterinaria.4Universidad Nacional de Salta, Facultad de Ciencias Naturales.
MARTÍNEZ, G.M.1; MICHELOUD, J.F.2; SUÁREZ, V.H.2; ROSA, D.E.3; VENTURA, M.V.3; ORTEGA, A.1; SÁNCHEZ, C.4; MATTIOLI, G.A.3
Recibido 09 de agosto de 2017 // Aceptado 25 de septiembre de 2018 // Publicado online 12 de agosto de 2020

Niveles plasmáticos de calcio, magnesio y fósforo en rodeos lecheros del Valle de Lerma, Salta, Argentina
ARTÍCULOS RIA / Vol. 46 / N.º 2182182
jugular vein of 9/10 animals in each category: prepartum, fresh cow, 2nd and 3rd third of lactation. Values Ca and Mg serum were obtained by atomic absorption spectrophotometry and P values by UV-visible spectro-photometry. Associations between variables were analyzed using linear correlation and linear regression, in addition to analyzing the probability of occurrence of hypocalcemia by odds ratio. Differences between varia-bles and moment of lactation were examined by analysis of variance and Tukey’s test and Chi square using Fisher’s test. The mean serum Ca was 8.85 mg / dl, with no differences found all over the periods. The average prevalence of hypocalcemia was 46.2% in all production periods. The mean serum Mg was 2.26%, with a pre-valence of 10.6% hypomagnesemia. Animals with hypomagnesemic had a 3.5 times greater chance of having hypocalcemia compared to those with normal values. The prevalence of animals with abnormal values of P was high (48.3%), without association with hypocalcemia levels. Pursuant to the results obtained in this study are warned of the potential risks of imbalances Ca, Mg and P in the dairy herds Lerma Valley.
Keywords: imbalance, minerals, dairy cows.
INTRODUCCIÓN
La producción lechera en la cuenca del Valle de Lerma representa una de las economías regionales más relevan-tes para la provincia de Salta. La producción media diaria es de 21,5±5,3 l/vaca, con producción promedio por tambo de 4900 l/día; y con rodeos promedio de 208±105 anima-les en ordeñe (Suárez y Martínez, 2015a). El sistema de producción predominante es de tipo pastoril, basado en pasturas templadas con altas tasas de suplementación y animales de elevado valor genético, similares a otras cuen-cas lecheras de Argentina.
Se conoce la importancia que tienen los macrominera-les como calcio (Ca), fósforo (P) y magnesio (Mg) sobre la producción y la salud de los rodeos lecheros (Goff y Horst, 2003). Los desbalances minerales constituyen un tema de preocupación asociado al bienestar animal y a la rentabili-dad de la industria lechera. Sin embargo, su impacto en los sistemas productivos por lo general pasa desapercibido, ya que en la mayoría de los casos tienen una presentación subclínica, afectando la salud y los niveles productivos, pu-diendo confundirse con otras alteraciones (Contreras et al., 1990). Los ejes metabólico-nutricionales que controlan los niveles de Ca, P y Mg parecen ser los más comprometidos durante las primeras etapas posparto, y cada vez existen más evidencias sobre la estrecha relación entre los siste-mas endócrinos que controlan el metabolismo y aquellos que regulan el sistema inmunológico (Elsasser, 1992).
El diagnóstico de los desbalances de macrominerales constituye una de las bases de la medicina preventiva en los rodeos lecheros (Wittwer, 2007). De este modo la medi-ción de los niveles de Ca, P, y Mg sérico resulta ser una he-rramienta útil para monitorear estos desequilibrios (Herdt et al., 2000). A nivel nacional algunos trabajos ponen de manifiesto la importancia de estos desbalances minerales para el sector lechero (CLAVES, 2009). El informe CLAVES identifica dentro de las causas de mortandad más impor-tante en los primeros noventa días posparto la asociación entre hipocalcemia e hipomagnesemia. A su vez, además de resultar las enfermedades diagnosticadas con mayor
frecuencia explican alrededor del 10,5% de las muertes durante este periodo. Por una parte dicho informe destaca que aquellos animales adultos que sufren un episodio de desbalance mineral, sea de Ca o Mg durante los primeros 3 meses de lactancia, presentan una disminución a lo largo de este periodo de 166 litros de leche. Por otra parte ade-más de las pérdidas de producción lácteas es importan-te poner de manifiesto lo oneroso de los tratamientos, es decir, insumos y horas hombre que conllevan este tipo de patologías (CLAVES, 2009).
Si bien los trabajos de relevamiento sérico resultan esen-ciales en el diagnóstico de los desbalances minerales en el sistema lechero, no existen antecedentes para la Cuenca del Valle del Lerma (Salta) en este sentido. El objetivo del presen-te trabajo fue realizar un análisis descriptivo y comparativo de los niveles séricos de Ca, P y Mg, evaluando la prevalencia de sus desbalances y su asociación entre sí y con las etapas productivas en rodeos lecheros de la cuenca salteña.
MATERIALES Y MÉTODOS
Se seleccionaron 4 tambos comerciales de la cuenca le-chera del Valle de Lerma en Salta.En todos los estableci-mientos la alimentación fue de tipo pastoril, siendo la base forrajera pasturas templadas (praderas polífíticas) con altas tasas de suplementación tanto energética como proteica, lo que es representativo de la región (Suárez y Martínez, 2015a). Las prácticas de manejo tanto nutricionales como sanitarias, como así también el biotipo lechero empleado fueron similares en los cuatro establecimientos y la produc-ción promedio fue de 7979 l de leche/vaca/lactancia con extremos de 7176 y 8472 litros. Cada establecimiento fue muestreado 4 veces a intervalos de 3 a 4 meses entre sí (primavera 2013, verano de 2014, otoño 2014 y primavera 2014). En cada muestreo se seleccionaron al azar 36 a 40 vacas de 4 periodos productivos (9 a 10 por periodo) que fueron: Preparto, aproximadamente 21 días antes de la fe-cha probable de parto; Lactancia temprana o Vaca Fresca, durante los primeros 30 días de lactancia; Lactancia media,

MARTÍNEZ, G.M.; MICHELOUD, J.F.; SUÁREZ, V.H.; ROSA, D.E.; VENTURA, M.V.; ORTEGA, A.; SÁNCHEZ, C.; MATTIOLI, G.A.
Agosto 2020, Argentina 183183
En este estudio los valores promedio de Ca se ubicaron por dentro de los rangos normales de referencia y no mos-traron diferencias significativas entre periodos productivos, aunque en todos ellos se halló un porcentaje elevado de vacas con hipocalcemia (tabla 1). Se cita que la hipocalce-mia es más frecuente en el periparto, es decir, 4 semanas antes y 4 después del parto (Shank et al., 1981; Curtis et al., 1983; Stevenson y Lean, 1998). En el presente traba-jo esta diferencia no llegó a ser significativa, pero el valor promedio más bajo y el mayor porcentaje de valores con hipocalcemia (53%) se observaron durante el preparto. En el posparto el 46% presentó hipocalcemia, lo cual es gra-ve considerando que valores inferiores a 8,4-8,6 mg/dl du-rante los primeros días posparto se asocian con mayores concentraciones plasmáticas de ácidos grasos no esteri-ficados y de β-hidroxibutirato, con aumentos de metritis y del intervalo parto concepción, y con menor producción de leche (Chapinal et al., 2012; Martínez et al., 2012).
Se identificaron 13 animales con hipocalcemia severa (2,22%), inferior al 5% reportada por otros autores para rodeos con alimentación de base pastoril (Roche, 2003; Goff, 2006). Sin embargo, los presentes resultados coinci-den con declaraciones de los tamberos del Valle de Lerma, quienes afirman padecer un 2,8±1,4% de vacas caídas pre-sumiblemente debido a hipocalcemia (Suárez et al., 2015). Sánchez y Saborío Montero (2014), en un relevamiento en vacas Holstein en sistemas pastoriles, obtuvieron una pre-valencia general de hipocalcemia de 57% (n=86), donde el 6,6% fueron casos clínicos y el 50% subclínicos; preva-lencia similar al 46% de hipocalcemias observadas en el presente trabajo.
La importancia de la prevención de la hipocalcemia al parto se basa en sus consecuencias. Trabajos de Curtis et al. (1985) y Gröhn et al. (1990) indican que cada vaca que padece un caso de hipocalcemia al parto presenta 7,2 veces más posibilidades de tener un parto distócico, 5,7 veces más posibilidades de padecer retención de placenta y 5,4 veces más posibilidades de manifestar casos clíni-cos de mastitis durante los primeros 90 días en leche, a la vez que aumenta la posibilidad de episodios de metritis, desplazamiento del abomaso y cetosis. Una causa de hipo-calcemias al parto es la elevada concentración de potasio en el alimento, lo cual genera alcalosis metabólica (Rérat
Tabla 1. Valores de calcio sérico por periodo de lactancia.D.E.: Desvío Estándar.Medias con una letra común no son significativamente diferentes (p>0,05).
entre 90 y 120 días en leche y Lactancia tardía, entre 200-305 días. La totalidad de las muestras correspondieron a vacas de más de 2 partos.
Determinación de Ca, Mg y P en suero
Las muestras de sangre se obtuvieron por punción yu-gular. Se extrajo suero libre de hemólisis por centrifugación para la determinación de Ca, P y Mg. Las concentraciones de Ca y Mg se midieron por espectrofotometría de absor-ción atómica de llama (Perkin Elmer AAnalyst 200), previa dilución en LaCl (0,1 N), y las determinaciones de P se realizaron mediante espectrofotometría UV-visible (Perkin Elmer-Lanbda25), según indicaciones del fabricante.
Análisis estadísticos
Para los análisis estadísticos los valores de referencia para considerar hipocalcemia clínica y subclínica fueron <5,5 y de 5,5 a <8,5 mg/dl respectivamente (McDowell, 2002; Goff, 2006; Chapinal et al., 2012). Para el caso del Mg se consideró hipomagnesemia a los valores inferiores a 1,8 mg/dl (McDowell, 2002). Los valores de referencia para hipofosfatemia e hiperfosfatemia fueron <4 y >6,18 mg/dl respectivamente (Goff, 2006).
Las relaciones entre variables de los valores séricos fueron analizadas usando correlación lineal (Pearson) y regresión lineal simple, además de analizar la probabilidad de ocurrencia de hipocalcemia mediante odds ratio. Las diferencias entre variables y periodos de lactancia se exa-minaron mediante los análisis de varianza y test de Tukey y de chi cuadrado usando el test de Fisher. Para los aná-lisis indicados se utilizó el paquete estadístico InfoStat (Di Rienzo et al., 2008).
RESULTADOS Y DISCUSIÓN
Concentraciones séricas de Ca los diferentes periodos productivos
La hipocalcemia clínica se asocia a valores de hipocal-cemia severa, inferiores a 5,5 mg/dl, mientras que la forma subclínica se asocia a valores de hipocalcemia moderada, de entre 5,5 a 8,0 mg/dl (McDowell, 2002; Goff, 2006).
Periodo n Media (mg/dl) D.E. Hipocalcémicas (%)Preparto 135 8,55 a 1,94 53
Vaca Fresca(hasta 30 días posparto)
155 8,82 a 1,94 46
2.o tercio lactancia 140 8,96 a 1,94 46
3.o tercio lactancia 154 9,06 a 1,89 41
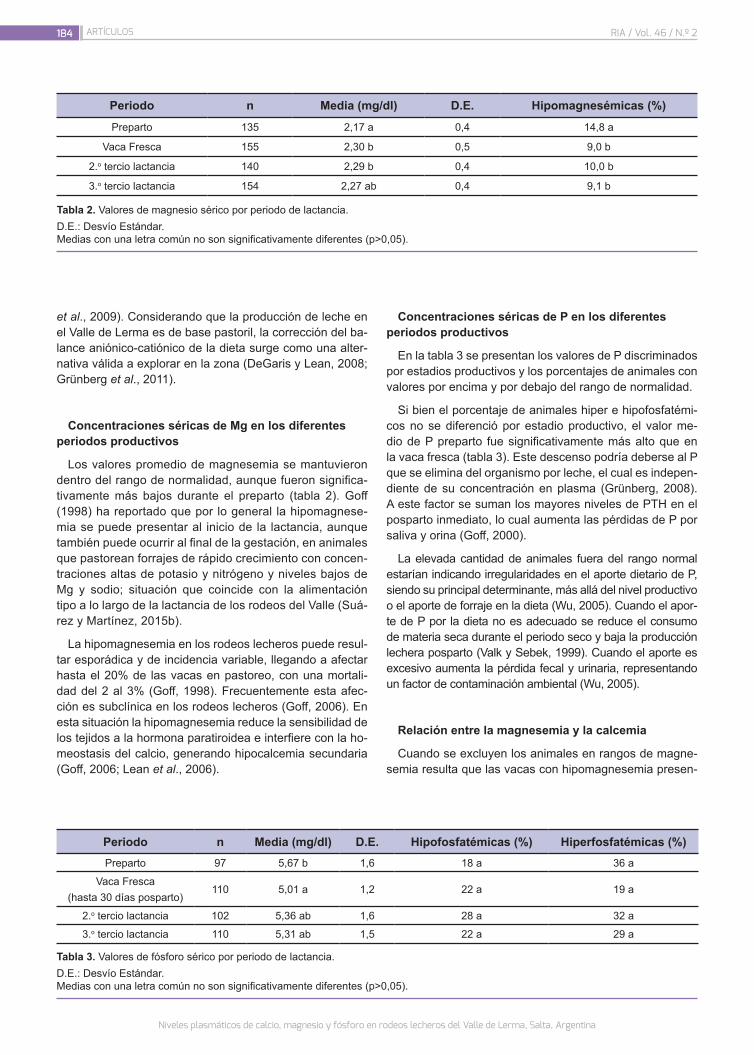
Niveles plasmáticos de calcio, magnesio y fósforo en rodeos lecheros del Valle de Lerma, Salta, Argentina
ARTÍCULOS RIA / Vol. 46 / N.º 2184184
Tabla 3. Valores de fósforo sérico por periodo de lactancia.D.E.: Desvío Estándar.Medias con una letra común no son significativamente diferentes (p>0,05).
Tabla 2. Valores de magnesio sérico por periodo de lactancia.D.E.: Desvío Estándar.Medias con una letra común no son significativamente diferentes (p>0,05).
et al., 2009). Considerando que la producción de leche en el Valle de Lerma es de base pastoril, la corrección del ba-lance aniónico-catiónico de la dieta surge como una alter-nativa válida a explorar en la zona (DeGaris y Lean, 2008; Grünberg et al., 2011).
Concentraciones séricas de Mg en los diferentes periodos productivos
Los valores promedio de magnesemia se mantuvieron dentro del rango de normalidad, aunque fueron significa-tivamente más bajos durante el preparto (tabla 2). Goff (1998) ha reportado que por lo general la hipomagnese-mia se puede presentar al inicio de la lactancia, aunque también puede ocurrir al final de la gestación, en animales que pastorean forrajes de rápido crecimiento con concen-traciones altas de potasio y nitrógeno y niveles bajos de Mg y sodio; situación que coincide con la alimentación tipo a lo largo de la lactancia de los rodeos del Valle (Suá-rez y Martínez, 2015b).
La hipomagnesemia en los rodeos lecheros puede resul-tar esporádica y de incidencia variable, llegando a afectar hasta el 20% de las vacas en pastoreo, con una mortali-dad del 2 al 3% (Goff, 1998). Frecuentemente esta afec-ción es subclínica en los rodeos lecheros (Goff, 2006). En esta situación la hipomagnesemia reduce la sensibilidad de los tejidos a la hormona paratiroidea e interfiere con la ho-meostasis del calcio, generando hipocalcemia secundaria (Goff, 2006; Lean et al., 2006).
Concentraciones séricas de P en los diferentes periodos productivos
En la tabla 3 se presentan los valores de P discriminados por estadios productivos y los porcentajes de animales con valores por encima y por debajo del rango de normalidad.
Si bien el porcentaje de animales hiper e hipofosfatémi-cos no se diferenció por estadio productivo, el valor me-dio de P preparto fue significativamente más alto que en la vaca fresca (tabla 3). Este descenso podría deberse al P que se elimina del organismo por leche, el cual es indepen-diente de su concentración en plasma (Grünberg, 2008). A este factor se suman los mayores niveles de PTH en el posparto inmediato, lo cual aumenta las pérdidas de P por saliva y orina (Goff, 2000).
La elevada cantidad de animales fuera del rango normal estarían indicando irregularidades en el aporte dietario de P, siendo su principal determinante, más allá del nivel productivo o el aporte de forraje en la dieta (Wu, 2005). Cuando el apor-te de P por la dieta no es adecuado se reduce el consumo de materia seca durante el periodo seco y baja la producción lechera posparto (Valk y Sebek, 1999). Cuando el aporte es excesivo aumenta la pérdida fecal y urinaria, representando un factor de contaminación ambiental (Wu, 2005).
Relación entre la magnesemia y la calcemia
Cuando se excluyen los animales en rangos de magne-semia resulta que las vacas con hipomagnesemia presen-
Periodo n Media (mg/dl) D.E. Hipomagnesémicas (%)Preparto 135 2,17 a 0,4 14,8 a
Vaca Fresca 155 2,30 b 0,5 9,0 b
2.o tercio lactancia 140 2,29 b 0,4 10,0 b
3.o tercio lactancia 154 2,27 ab 0,4 9,1 b
Periodo n Media (mg/dl) D.E. Hipofosfatémicas (%) Hiperfosfatémicas (%)Preparto 97 5,67 b 1,6 18 a 36 a
Vaca Fresca(hasta 30 días posparto)
110 5,01 a 1,2 22 a 19 a
2.o tercio lactancia 102 5,36 ab 1,6 28 a 32 a
3.o tercio lactancia 110 5,31 ab 1,5 22 a 29 a

MARTÍNEZ, G.M.; MICHELOUD, J.F.; SUÁREZ, V.H.; ROSA, D.E.; VENTURA, M.V.; ORTEGA, A.; SÁNCHEZ, C.; MATTIOLI, G.A.
Agosto 2020, Argentina 185185
Tabla 4. Concentración media de calcio sérico en relación con los rangos de magnesemia.D.E.: Desvío Estándar.
Tabla 5. Tabla de contingencia que ubica el número de vacas con o sin hipocalcemia en función de padecer o no hipomagnesemia (Chi 19,72; p<0,0001).
tan a su vez valores inferiores de calcemia (p<0,0001; tabla 4). Como se explicara anteriormente, la reducción de los nive-les de magnesio en la sangre comprometen la homeostasis del calcio haciendo que los tejidos blanco pierdan sensibilidad para la PTH y la vitamina D activa (Reinhardt et al., 1988).
Al evaluar la asociación correspondiente entre el nivel de magnesio y calcio séricos se obtuvo un coeficiente de corre-lación de 0,3 y la determinación del Ca sérico a partir del Mg sérico como variable regresiva resultó significativa y de tipo lineal (Ca mg/dl=5,42+1,6x [Mg mg/dl]; r2: 0,09; p<0,0001).
Trabajos más recientes proponen elevar el rango inferior de calcemia a 8,4 mg/dl en vacas de alto mérito genético y productividad (Chapinal et al., 2012; Martínez et al., 2012). Aplicando este valor a la ecuación anterior, el límite inferior de normomagnesemia debería elevarse a 1,87 mg/dl, al menos en sistemas lecheros de este tipo.
Si bien la deficiencia de Mg no resultaría ser la patología más relevante, aquellos animales con hipomagnesemia presentaron una probabilidad 3,5 mayor de sufrir hipocalce-mia respecto a aquellos con valores normales (odds ratio: 3,53 LI 95% 1,98 LS 95% 6,29). Por un lado, al ordenar los datos en una tabla de contingencia, se observa que solo el 7,7% de los animales presentaron una deficiencia conjunta de hipomagnesemia con hipocalcemia (tabla 5). Esto lleva a pensar que los casos de hipocalcemia observados se de-berían fundamentalmente a aspectos biológicos propios de la homeostasis del Ca y no secundarios a la carencia del Mg, coincidiendo además con informes previos (Sánchez y et al., 2014). Por otro lado, esta distribución podría interpre-tarse en función de que la hipomagnesemia sería respon-sable de la hipocalcemia, por lo cual el 73% de las vacas con hipomagnesemia presentan hipocalcemia secundaria a la carencia de Mg, mientras que de los animales que pre-sentan hipocalcemia, posiblemente como consecuencia primaria de su estado fisiológico, solo el 5% presentaba hipomagnesemia. La relación causal sigue siendo que la hipomagnesemia genera una hipocalcemia secundaria, pero no a la inversa, en coincidencia con trabajos anterio-res (Goff, 2006; Lean et al., 2006).
Relación entre los valores de fósforo sérico y la calcemia
No se hallaron relaciones significativas entre aquellos animales que presentaron hiperfosfatemia (Chi 0,03; p<0,85) o hipofosfatemia (Chi 0,9; p<0,35) y a su vez hi-
pocalcemia. Sin embargo, la hipocalcemia puede estar asociada a ambas. La mayoría de los casos de hipocal-cemia clínica (paresia puerperal) se encuentran asocia-dos a hipofosfatemias, de modo que la corrección con Ca parenteral genera la recuperación espontánea del P, mientras que cuando esto no ocurre, la hipofosfatemia persistente en una causa frecuente del síndrome de vaca caída (Grünberg, 2008). Las hiperfosfatemias debidas al exceso de P en el preparto conducen a hipocalcemias se-cundarias en el posparto. Con aportes 0,21; 0,31 y 0,44% P en el preparto de vacas lecheras en ningún caso se alteró el metabolismo óseo ni los niveles de PTH o vit. D (Peterson et al., 2005). Sin embargo, en este trabajo se observó que los animales con mayor aporte de P presen-taron las menores calcemias durante el posparto, sugi-riendo un efecto de interferencia. Este ocurriría cuando la dieta aporta exceso de P (>50 g/día) en el preparto de vacas lecheras, inhibiendo la activación renal de vit. D y predisponiendo a la paresia puerperal (Schonewille et al., 1999). NRC (2001) sugiere una relación no inferior 1:1 y no superior 7:1 siempre y cuando el consumo de fósforo se ajuste a los requerimientos de los animales. Valores por encima o por debajo de estos límites se asocian a una mayor incidencia de hipocalcemia.
CONCLUSIONES
Al estudiar las prevalencias de hipocalcemia, hipomag-nesemia y de los desbalances del P en los rodeos del Valle de Lerma, se destaca la elevada prevalencia de hipocalce-mia, de 46,2% promedio en todos los periodos de lactan-cia. En cuanto a la prevalencia de hipomagnesemia, que resultó del 10,6% fue más elevada durante el preparto que en el resto de las categorías. También se halló una alta pro-porción de vacas (48,3%) con desbalances en la concen-tración de P, siendo más frecuentes las hiperfosfatemias que las hipofosfatemias.
Es importante destacar que la deficiencia de magnesio parece tener un rol muy importante en la aparición de hipo-calcemias posparto, ya el 73% de las vacas con hipomagne-semia presentan hipocalcemia, indicando que estas tuvieron una probabilidad 3,5 veces mayor de sufrir hipocalcemia respecto a aquellas con valores normales de Mg. Aunque se halló un porcentaje elevado de animales con niveles de P fuera del rango normal, no se observaron asociaciones entre este mineral con los de incidencia de hipocalcemia.
Rango Media (mg/dl) n D. E.Hipomagnesemia
(<1,80 mg/dl) 7,98 a 62 2,36
Normomagnesemia (≥1,80 mg/dl) 9,17 b 522 2,28
Hipo Mg Normo Mg Total
HipoCa 45 225 270
Normo Ca 17 297 314
Total 62 522 584

Niveles plasmáticos de calcio, magnesio y fósforo en rodeos lecheros del Valle de Lerma, Salta, Argentina
ARTÍCULOS RIA / Vol. 46 / N.º 2186186
En virtud a los resultados obtenidos en el presente traba-jo se sugiere corregir el aporte de minerales en las dietas de los rodeos lecheros del Valle de Lerma con un énfasis particular en evitar las hipocalcemias, especialmente se-cundarias a bajos aportes de Mg, así como los excesos de P en la dieta, a fin de minimizar los riesgos sanitarios y ambientales que conllevan.
BIBLIOGRAFÍACHAPINAL, N.; LEBLANC, M.; CARSON, K.; LESLIE,
S.; GODDEN, M.; CAPEL, J.; SANTOS, M.; OVERTON, N.; DUFFIELD, T. 2012. Herd-level association of serum metabolites in the transition period with disease, milk production, and early lac-tation reproductive performance. J. Dairy Sci. 95,5676-5682.
CLAVES. 2009. Organización y Análisis de un Sistema de Re-gistros de Enfermedades del Periparto en Vacas Lecheras: su In-cidencia e Impacto Económico sobre las Empresas. (Disponible: http://www.infortambo.com/admin/upload/arch/Claves%20I%20reporte%20final.pdf verificado: 02 de agosto de 2017).
CONTRERAS, P.A.; WITTWER, F.; BÖHMWALD, H. 1990. Con-centraciones de calcio, fósforo y magnesio en suero sanguíneo de bovinos de leche en 40 predios lecheros de la x Región, Chile. Arch. Med. Vet. 22, 185-189.
CURTIS, C.R.; ERB, H.N.; SNIFFEN, G.J.; SMITH, R.D.; POWERS P.A.; SMITH, M.C.; WHITE, M.E.; HILLMAN, R.B.; PEAR-SON, E.J. 1983. Association of parturient hypocalcemia with peri-parturient disorders in Hols. J. Am. Vet. Med. Assoc. 183(5), 559-61.
CURTIS, C.R.; ERB, H.N.; SNIFFEN, C.J.; SMITH, R.D.; KRON-FELD, D.S. 1985. Path analysis of dry period nutrition, postpar-tum metabolic and reproductive disorders, and mastitis in Holstein cows. J Dairy Sci. 68(9), 2347-60.
DEGARIS, P.J.; LEAN, I.J. 2008. Milk fever in dairy cows: a review of pathophysiology and control principles. Vet. J. 176(1), 58-69.
DI RIENZO, J.A.; CASANOVES, F.; BALZARINI, M.G.; GONZALEZ, L.; TABLADA, M.; ROBLEDO, C.W. 2008. InfoStat, versión 2008, Gru-po InfoStat, FCA, Universidad Nacional de Córdoba, Argentina.
ELSASSER, T.H. 1992. The impact of production disease stress on the character of growth and productivity of food producing ani-mal. Proc. 8th. Int. Conf. on Prod. Diseases in Farm Animals, Uni-versity of Berne, Suiza. 1-15 pp.
GOFF, J.P; HORST, R.L. 2003. Milk fever control in the United States. Acta Vet Scand Suppl. 97, 145-147.
GOFF, J.P. 1998. Ruminant hypomagnesemic tetanies. Current Veterinary Therap: Food Animal Practice. 4th Ed. Edited by Jimmy L. Howard. W.B. Saunders. Philadelphia, Pa. EUA. 1-9 pp.
GOFF, J.P. 2000. Pathopysiology of calcium and phosphorus di-sorders. Vet. Clin. North Am. Food Anim. Pract. 13, 619-639.
GOFF, J.P. 2006. Macromineral physiology and application to the feeding of the dairy cow for prevention of milkfever and other periparturient mineral disorders. Animal Feed Science and Tech-nology 126, 237-257.
GRÖHN, Y.T.; ERB, H.N.; MCCULLOCH, C.E.; SALONIEMI, H.S. 1990. Epidemiology of reproductive disorders in dairy cattle: Associations among host characteristics, disease and production. Prev. Vet. Med. 8, 25-39.
GRÜNBERG, W.; DONKIN, S.S.; CONSTABLE, P.D. 2011. Pe-riparturient effects of feeding a low dietary cation-anion difference diet on acid-base, calcium, and phosphorus homeostasis and on intravenous glucose tolerance test in high-producing dairy cows. J Dairy Sci. 94(2), 727-745.
GRÜNBERG, W. 2008. Phosphorus Homeostasis in Dairy Catt-le: Some Answers, More Questions. Tri-State Dairy Nutrition Con-
ference, 29-35.HERDT, T.H.; RUMBEIHA, W., BRASELTON, W.E. 2000. The
use of blood analyses to evaluate mineral status in livestock. Vet Clin North Am Food Anim Pract. 16(3), 423-444.
LEAN, I.J.; DEGARIS, P.J.; MCNEIL, D.M.; BLOCK, E. 2006. Hypocalcemia in dairy cows: meta-analysis and dietary cation anion difference theory revisited. J Dairy Sci. 89(2), 669-684.
MCDOWELL, L.R. 2002. Recent advances in mineral and vita-mins on nutrition of lactating cows. Pakistan Journal of Nutrition 1(1), 8-19.
MARTINEZ, N.; RISCO, C.; LIMA, F.; BISINOTTO, R.; GRECO, L.; RIBEIRO, E.; GALVAO, K.; SANTOS, J. 2012. Evaluation of peripartal calcium status, energetic profile, and neutrophil function in dairy cows at low or high risk of developing uterine disease. J. Dairy Sci. 95, 7158-7172.
NRC (NATIONAL RESEARCH COUNCIL). 2001. Nutrient Re-quirements of Dairy Cattle: Seventh Revised Edition, 2001. Natio-nal Academy Press. Washington, D. C., EUA.
OETZEL, G.R. 2002. Milk Fever. Encyclopedia Dairy Sciense. Vol 2. Editorial Academic Press. Biblioteca de la División Académi-ca de Ciencias Agropecuarias de la Universidad Juarez Autónoma de Tabasco. México. 824-830 pp.
REINHARDT, T.A.; HORST, R.L.; GOFF, J.P. 1988. Calcium, phosphorus, and magnesium homeostasis in ruminants. Vet. Clin. North Am. Food Anim. Pract. 4, 331.
RÉRAT, M.; PHILIPP, A.; HESS, H.D.; LIESEGANG, A. 2009. Effect of different potassium levels in hay on acid-base status and mineral balance in periparturient dairy cows. J Dairy Sci. 92(12), 6123-6133.
ROCHE, J.R. 2003. The incidence and control of hipocalcemia in pasture-based systems. Acta Vet. Scand. 97,141-144.
SÁNCHEZ, J.M.; SABORÍO-MONTERO, A. 2014. Hipocalcemia e Hipomagnesemia en un hato de vacas Holstein, Jersey y Guern-sey en pastoreo. Agronomía Costarricense 38(2), 55-65.
SCHONEWILLE, J.T.; VAN, A.T.; KLOOSTER, T.; WOURTE-SE, H.; BEYNEN, A.C. 1999. Hypocalcemia induced by intrave-nous administration of disodium ethylenedaminotetracetate and its effects on excretion of calcium in urine of cows fed a high chloride diet. J Dairy Sci 82, 1317-1324.
SHANK, R.D.; FREEMAN, A.E.; DICKINSON, F.N. 1981. Pos-tpartum distribution of costs and disorders of health. J Dairy Sci. 64, 683-688.
STEVENSON, M.A.; LEAN, I.J. 1998. Descriptive epidemiolo-gical study on culling and deaths in eight dairy herds. Australian Veterinary Journal 76, 482-488.
SUÁREZ, V.H.; MARTÍNEZ, G.M. 2015a. Caracterización de los sistemas productivos de leche del Valle de Lerma, Salta. Revista Argentina de Producción Animal 35(1), 109.
SUÁREZ, V.H.; MICHELOUD, J.F., MARTINEZ, G.M., BERTO-NI, E.A., NEUMANN, R.D. 2015. Presencia de enfermedades y problemas sanitarios en los tambos del Valle de Lerma, Salta. Re-vista Argentina de Producción Animal 35(1), 75.
SUÁREZ, V.; MARTÍNEZ, G.M. 2015b. Características y Proble-máticas Productivas - Sanitarias de la Lechería del Valle de Lerma (Salta). Ed INTA. 66 p.
VALK, H.; SEBEK, L.B. 1999. Influence of long-term feeding of limited amounts of phosphorus on dry matter intake, milk production, and body weight of dairy cows. J Dairy Sci. 82(10), 2157-2163.
WITTWER, F. 2007. Diagnóstico y control de carencias minera-les en bovinos. Seminario Internacional de Nutrición y Alimenta-ción Animal UNISARC, Risaralda, Colombia.
WU, Z. 2005. Utilization of phosphorus in lactating cows fed varying amounts of phosphorus and sources of fiber. J Dairy Sci. 88(8), 2850-2859.

MIRANDA, A.; FORT, M.; FUCHS, L.; BALDONE, V.; GIMÉNEZ, H.; CARNÉ, L.; SAGO, A.; OTERMIN, H.; DUBIÉ, D.; GOYENECHE, P.; CAMPERO, C.
Agosto 2020, Argentina 187187
Prevalencia y factores de riesgo asociados a las enfermedades de transmisión sexual en bovinos de la provincia de La Pampa, Argentina
RESUMENLa tricomonosis bovina (TB) y campylobacteriosis genital bovina (CGB) son enfermedades de transmisión
sexual (ETS) que causan muerte embrionaria temprana, infertilidad transitoria y abortos, impactando en el rendimiento reproductivo de los rodeos. El objetivo del presente estudio fue estimar la prevalencia general y por zonas fitogeográficas (Z1: Este pampeana, Z2: Monte central Z3: Monte occidental), así como los factores de riesgo asociados a ambas enfermedades en la provincia de La Pampa, Argentina. Se realizó un estudio transversal entre enero y diciembre del 2006 que abarcó a 4018 propietarios de rodeos con un stock de 684116 vacas y 33969 toros (54% y 51,5% del stock provincial, respectivamente). Se detectaron 622 (15,5%) rodeos con al menos 1 toro positivo a TB y/o CGB. Las prevalencias halladas fueron de 2,15% (Z1:1,62%; Z2:1,90%; Z3:4,61%) y 2,36% (Z1:2,08%; Z2: 2,28%; Z3: 3,51%) para TB y CGB, respectivamente. Los fac-tores de riesgo asociados a la presencia de ETS fueron: traspaso de animales vecinos al establecimiento (Odds Ratio – OR = 1,8; Intervalo de Confianza-IC: 1,5-2,2) realización de servicio continuo (OR=1,3; IC: 1,1-1,6), productores que no realizan el tacto rectal (OR=1,2; IC: 1,0-1,5) y alta relación vaca/toro (χ2 =35,1, p=<0,0001). Asimismo, las ETS se relacionaron con presencia de abortos (OR=1,3; IC: 1,0-1,7) y bajo por-centaje de preñez (OR=1,8; IC: 1,3-2,6). Este estudio provee información de interés para ser utilizada en los planes de control y erradicación en áreas en donde las ETS son de carácter endémico.
Palabras clave: enfermedades transmisión sexual, bovinos, tricomonosis, campylobacteriosis, prevalencia, factores de riesgo.
ABSTRACT
Bovine Tricomonosis (BT) and Bovine Genital Campylobacteriosis (BGC) are sexual transmission diseases (STD) that produce early embryonic death, transitory infertility and abortion, impacting on livestock reproduc-tive performance. The objective of the study was to estimate the global and specific prevalence by phytogeo-graphy area (Z1: East Pampeana, Z2: Central Forest, Z3. West Forest) as well as the risk factor associated to both diseases in La Pampa province, Argentina. A cross sectional study was carried out between January and December 2006 which included 4018 farmers having a stock of 684116 cows and 33969 bulls (54% and 51.5% of the provincial stock, respectively). Six hundred and twenty two (15.5%) cattle herds were detected with at
1Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Anguil, Grupo Sanidad Animal, Ruta Nacional N.o 5 km 580 (6326), Anguil, La Pampa, Argentina. Correo electrónico: [email protected] 2Servicio Nacional de Sanidad y Calidad Agroalimentaria (SENASA), Centro Regional La Pampa, San Luis, Argentina.3Colegio Veterinario de la provincia de La Pampa, La Pampa, Argentina.4Subsecretaría Agricultura del Gobierno de la Provincia de La Pampa, La Pampa, Argentina.5Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Balcarce, Grupo de Sanidad Animal, Balcarce, Argentina.
MIRANDA, A.1; FORT, M.1; FUCHS, L.1; BALDONE, V.1; GIMÉNEZ, H.1; CARNÉ, L.2; SAGO, A.2; OTERMIN, H.3; DUBIÉ, D.3; GOYENECHE, P.4; CAMPERO, C.5
Recibido 06 de agosto de 2018 // Aceptado 29 de octubre de 2018 // Publicado online 28 de octubre de 2020

Prevalencia y factores de riesgo asociados a las enfermedades de transmisión sexual en bovinos de la provincia de La Pampa, Argentina
ARTÍCULOS RIA / Vol. 46 / N.º 2188188
least one bull positive for TB and / or CGB. Prevalence was 2.15% (Z1: 1.62%, Z2: 1.90%, Z3: 4.61%) and 2.36% (Z1: 2.08%; Z2: 2.28%; Z3: 3.51%) for BT and BGC, respectively. Risk factors associated with the pre-sence of STD were: Passage of bulls from in-contact herds (Odds ratio OR=1,8; Confidence Intervals-IC: 1,5-2,2), no defined breeding season (OR=1,3; IC: 1,1-1,6), farmers who do not detect pregnancy by rectal exami-nation (OR=1,2; IC: 1,0-1,5) and high cows/bulls ratio (χ2 =35,1, p=<0,0001). Moreover, STD were related to abortion (OR=1.3; IC: 1.0-1.7) and low pregnancy rate (OR=1.8; IC: 1.3-2.6). This study provided information to use by control programs and erradication in a region where these diseases are considered to be endemic.
Keywords: Sexual transmisión deseases, bovine tricomonosis, bovine genital campylobacteriosis, preva-lence, risk factor.
INTRODUCCIÓN
Las enfermedades de transmisión sexual (ETS) afectan la capacidad reproductiva de los rodeos para cría bovina en los sistemas ganaderos extensivos, ocasionando severas pérdi-das económicas en todo el mundo (BonDurant, 2007). Den-tro de las ETS, la tricomonosis (TB), causada por el protozoo Tritrichomonas foetus, y la campylobacteriosis genital bovina (CGB), ocasionada por Campylobacter fetus subsp. fetus y venerealis) y Campylobacter fetus subsp. venerealis biovar intermedius, producen infertilidad y abortos en bovinos. Es-tos microorganismos colonizan el tracto genital femenino causando inflamación, infertilidad, pérdidas embrionarias y abortos, mientras que el toro actúa como hospedador inapa-rente (carrier) (Parsonson, 1974; BonDurant, 2007; Agnew, 2008). Las pérdidas indirectas se asocian a la venta de ani-males por descarte y a la reposición o reemplazo de anima-les infectados (BonDurant, 2007).
Las ETS han sido detectadas en todos los países que realizan producción bovina para carne en forma extensiva o semiextensiva, en lugares en donde el servicio natural es una práctica común (Yule et al., 1989; Anderson, 2007). Se han notificado diferentes prevalencias para TB en EUA con datos muy dispares que variaron del 0,1% a 30,4% según zonas como Florida, (Rae et al., 1999; Rae, 2004), Nebraska (Grotelueschen et al., 1994), Nevada (Kvasnic-ka et al., 1989), California (BonDurant et al.,1990) y Texas (Szonyi et al., 2012). En el centro oeste de Canadá, Riley et al. (1995) reportaron prevalencias del 6% en tanto que en Costa Rica (Pérez et al., 1992) y en el noroeste de España (Martín-Gómez et al., 1998) mencionaron prevalencias del 3,9% y 2,9%, respectivamente. Similar comportamiento en cuanto a la distribución y prevalencias es reportado para CGB (Stoessel, 1982). En ciertos países como Australia (Hum et al., 2009) es considerada como la principal ETS. En Uruguay, Repiso et al. (2005) reportaron prevalencias en toros del 28,1% en tanto que a nivel predial del 37%. En tanto que en otras regiones ganaderas del país, Campero y Martínez (2010) informaron prevalencias en rodeos infec-tados a TB del 5% al 25% mientras que para CGB de 2% al 15%. Así mismo, en regiones ganaderas de la provincia de San Luis, con características agroecológicas similares a las reportadas en el presente estudio, Rossanigo et al. (2004) reportan prevalencias de 10,8% y 9,5% a nivel de
rodeo y 1,1% y 0,72% a nivel de toros para TB y CGB, res-pectivamente. En la provincia de La Pampa, Argentina, se han reportado prevalencias del 2,3% y 20% a TB y del 0,8 y 10% a CGB en toros y rodeos infectados, respectivamente (Fort et al., 2004a, b).
Las ETS han sido erradicadas en ciertos países del mun-do y pueden ser controladas con el uso de la inseminación artificial, el control sanitario del semen, la venta de anima-les infectados y el apropiado conocimiento de los factores que predisponen a su presentación.
Trabajos realizados en regiones ganaderas reportan asociación entre factores de riesgo (bajo porcentaje de preñez, traspaso de toros de rodeos vecinos, ETS repor-tada previamente en el establecimiento) y la presencia de ETS, con efectos desfavorables sobre la rentabilidad de los rodeos afectados (Mardones et al., 2008; Jiménez et al., 2011; Ondrak, 2016). Sin embargo, dadas las carac-terísticas agroecológicas dispares en relación con el área bajo estudio del presente trabajo, el conocimiento de los factores de riesgo asociados constituye una herramienta relevante para tratar de controlar ambas enfermedades y disminuir el impacto de las pérdidas productivas.
La provincia de La Pampa cuenta con un total de 3781109 cabezas bovinas distribuidas en 143440 km2 representan-do el 6,5% de la existencia bovina del país. Desde el año 2007 y por resoluciones oficiales provinciales y nacionales (Servicio Nacional de Sanidad y calidad Agroalimentaria (SENASA, 2008)) se estableció la obligatoriedad del mues-tro para ETS en los toros de los rodeos provinciales, lo cual permitió contar con información para determinar la preva-lencia de dichas enfermedades en los rodeos de cría de la provincia. En el presente trabajo se plantea como objetivo informar sobre la prevalencia de las ETS y determinar los factores de riesgo asociados a su presentación, con espe-cial énfasis sobre características fitogeográfica específicas de cada zona de producción de la provincia de La Pampa.
MATERIALES Y MÉTODOS
El presente trabajo se llevó a cabo en conjunto con téc-nicos de la Estación Experimental del INTA Anguil, vete-rinarios y laboratorios de la actividad privada, el Colegio

MIRANDA, A.; FORT, M.; FUCHS, L.; BALDONE, V.; GIMÉNEZ, H.; CARNÉ, L.; SAGO, A.; OTERMIN, H.; DUBIÉ, D.; GOYENECHE, P.; CAMPERO, C.
Agosto 2020, Argentina 189189
Veterinario de la provincia de La Pampa y el SENASA. La información fue recopilada utilizando para ello una encues-ta modalidad cara a cara entre el veterinario privado, en-cargado de realizar la toma de muestras para el diagnós-tico de las ETS en los toros, y el productor agropecuario.
Área de estudio
El estudio transversal abarcó establecimientos distribui-dos a lo largo de todo el territorio provincial. En la tabla 1 se muestra el área de cobertura en km2, el número estimado de bovinos extraídos del registro de productores agrope-cuarios de la provincia de La Pampa (Repagro, 2008) y las precipitaciones anuales promedio (isohietas) para cada región en particular (Bellini Saibene, 2012).
Las explotaciones agropecuarias se agruparon de acuer-
do a características fitogeográficas de la provincia confor-mándose 3 regiones a saber: Zona 1 (Z1): Estepa Pam-peana, ubicada en el centro y noreste de la provincia con características productivas favorables para el desarrollo de agricultura de cosecha y ganadería de cría, recría y en-gorde. Zona 2 (Z2): Productores ubicados en la región del monte central con predominio de monte de caldén, gana-dería de cría sobre campo natural. Zona 3 (Z3): Producto-res ubicados en la región de “Monte Occidental” de arbus-tos bajos con limitada oferta forrajera, región en la cual se desarrolla la cría de bovinos exclusivamente (figura 1). Las cargas animales de las distintas regiones ascienden de 0,1; 0,5 y 2 vacas/ha para las Z3, Z2 y Z1, respectivamente.
Cada uno de los productores encuestados fue georrefe-renciado; la información fue procesada según el Sistema In-tegrado de Gestión de Sanidad Animal (SIGSA) provisto por
Zona fitogeográfica
Superficie en km2
N.º de animalesestimados
Precipitaciones en milímetros
Z1 28295 2009165 800
Z2 53905 1358948 600
Z3 61240 412996 400
Total 143440 3781109
Tabla 1. Superficie en km2, número de animales y precipitación anual por zona fitogeográfica analizada.
Figura 1. Mapa de la Argentina y de la provincia de La Pampa delimitando las zonas fitogeográficas estudiadas.
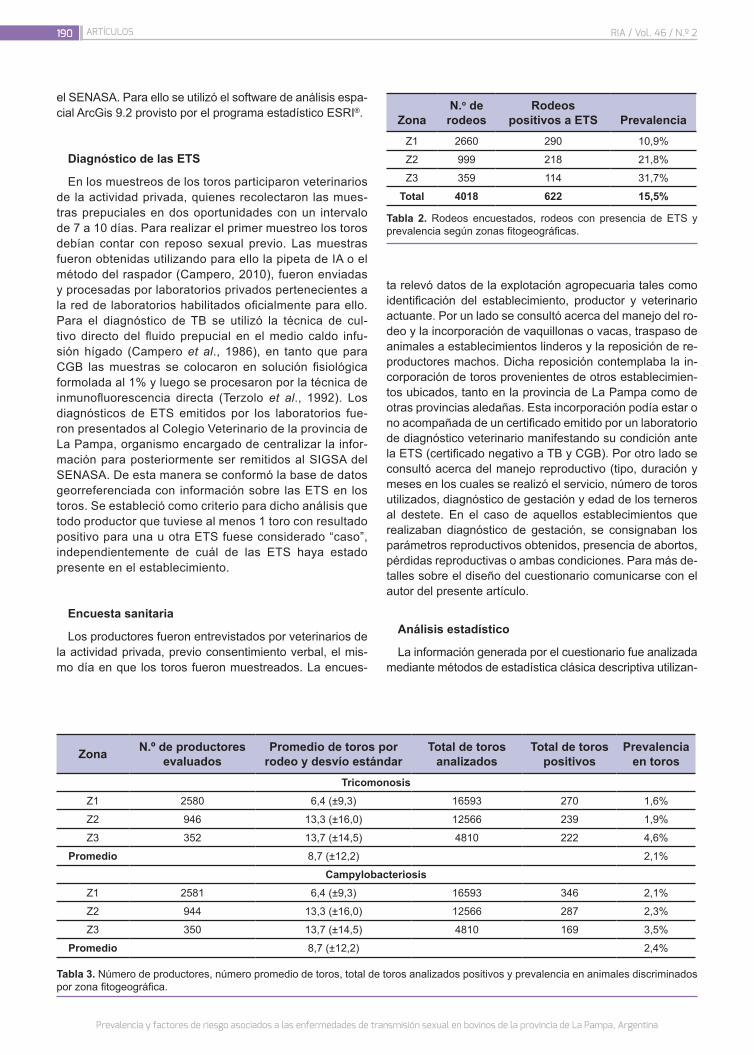
Prevalencia y factores de riesgo asociados a las enfermedades de transmisión sexual en bovinos de la provincia de La Pampa, Argentina
ARTÍCULOS RIA / Vol. 46 / N.º 2190190
Tabla 2. Rodeos encuestados, rodeos con presencia de ETS y prevalencia según zonas fitogeográficas.
Tabla 3. Número de productores, número promedio de toros, total de toros analizados positivos y prevalencia en animales discriminados por zona fitogeográfica.
el SENASA. Para ello se utilizó el software de análisis espa-cial ArcGis 9.2 provisto por el programa estadístico ESRI®.
Diagnóstico de las ETS
En los muestreos de los toros participaron veterinarios de la actividad privada, quienes recolectaron las mues-tras prepuciales en dos oportunidades con un intervalo de 7 a 10 días. Para realizar el primer muestreo los toros debían contar con reposo sexual previo. Las muestras fueron obtenidas utilizando para ello la pipeta de IA o el método del raspador (Campero, 2010), fueron enviadas y procesadas por laboratorios privados pertenecientes a la red de laboratorios habilitados oficialmente para ello. Para el diagnóstico de TB se utilizó la técnica de cul-tivo directo del fluido prepucial en el medio caldo infu-sión hígado (Campero et al., 1986), en tanto que para CGB las muestras se colocaron en solución fisiológica formolada al 1% y luego se procesaron por la técnica de inmunofluorescencia directa (Terzolo et al., 1992). Los diagnósticos de ETS emitidos por los laboratorios fue-ron presentados al Colegio Veterinario de la provincia de La Pampa, organismo encargado de centralizar la infor-mación para posteriormente ser remitidos al SIGSA del SENASA. De esta manera se conformó la base de datos georreferenciada con información sobre las ETS en los toros. Se estableció como criterio para dicho análisis que todo productor que tuviese al menos 1 toro con resultado positivo para una u otra ETS fuese considerado “caso”, independientemente de cuál de las ETS haya estado presente en el establecimiento.
Encuesta sanitaria
Los productores fueron entrevistados por veterinarios de la actividad privada, previo consentimiento verbal, el mis-mo día en que los toros fueron muestreados. La encues-
ta relevó datos de la explotación agropecuaria tales como identificación del establecimiento, productor y veterinario actuante. Por un lado se consultó acerca del manejo del ro-deo y la incorporación de vaquillonas o vacas, traspaso de animales a establecimientos linderos y la reposición de re-productores machos. Dicha reposición contemplaba la in-corporación de toros provenientes de otros establecimien-tos ubicados, tanto en la provincia de La Pampa como de otras provincias aledañas. Esta incorporación podía estar o no acompañada de un certificado emitido por un laboratorio de diagnóstico veterinario manifestando su condición ante la ETS (certificado negativo a TB y CGB). Por otro lado se consultó acerca del manejo reproductivo (tipo, duración y meses en los cuales se realizó el servicio, número de toros utilizados, diagnóstico de gestación y edad de los terneros al destete. En el caso de aquellos establecimientos que realizaban diagnóstico de gestación, se consignaban los parámetros reproductivos obtenidos, presencia de abortos, pérdidas reproductivas o ambas condiciones. Para más de-talles sobre el diseño del cuestionario comunicarse con el autor del presente artículo.
Análisis estadístico
La información generada por el cuestionario fue analizada mediante métodos de estadística clásica descriptiva utilizan-
ZonaN.o de rodeos
Rodeospositivos a ETS Prevalencia
Z1 2660 290 10,9%
Z2 999 218 21,8%
Z3 359 114 31,7%
Total 4018 622 15,5%
Zona N.º de productores evaluados
Promedio de toros por rodeo y desvío estándar
Total de toros analizados
Total de toros positivos
Prevalencia en toros
TricomonosisZ1 2580 6,4 (±9,3) 16593 270 1,6%
Z2 946 13,3 (±16,0) 12566 239 1,9%
Z3 352 13,7 (±14,5) 4810 222 4,6%
Promedio 8,7 (±12,2) 2,1%
CampylobacteriosisZ1 2581 6,4 (±9,3) 16593 346 2,1%
Z2 944 13,3 (±16,0) 12566 287 2,3%
Z3 350 13,7 (±14,5) 4810 169 3,5%
Promedio 8,7 (±12,2) 2,4%
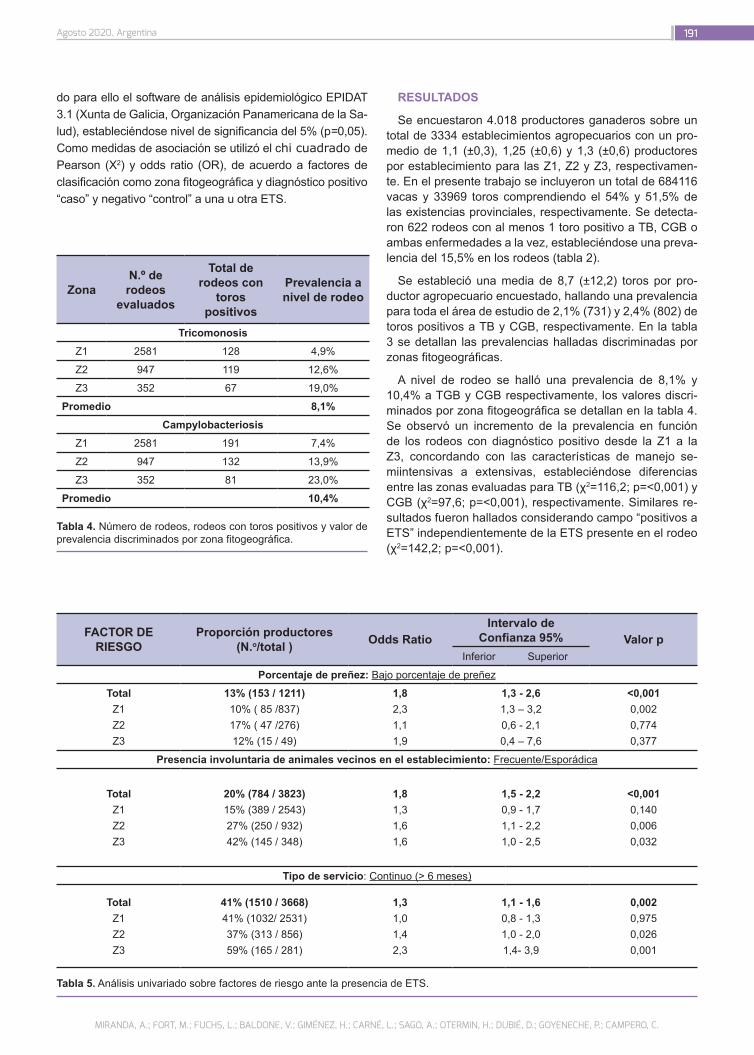
MIRANDA, A.; FORT, M.; FUCHS, L.; BALDONE, V.; GIMÉNEZ, H.; CARNÉ, L.; SAGO, A.; OTERMIN, H.; DUBIÉ, D.; GOYENECHE, P.; CAMPERO, C.
Agosto 2020, Argentina 191191
do para ello el software de análisis epidemiológico EPIDAT 3.1 (Xunta de Galicia, Organización Panamericana de la Sa-lud), estableciéndose nivel de significancia del 5% (p=0,05). Como medidas de asociación se utilizó el chi cuadrado de Pearson (X2) y odds ratio (OR), de acuerdo a factores de clasificación como zona fitogeográfica y diagnóstico positivo “caso” y negativo “control” a una u otra ETS.
ZonaN.º de rodeos
evaluados
Total de rodeos con
toros positivos
Prevalencia a nivel de rodeo
TricomonosisZ1 2581 128 4,9%
Z2 947 119 12,6%
Z3 352 67 19,0%
Promedio 8,1%Campylobacteriosis
Z1 2581 191 7,4%
Z2 947 132 13,9%
Z3 352 81 23,0%
Promedio 10,4%
Tabla 4. Número de rodeos, rodeos con toros positivos y valor de prevalencia discriminados por zona fitogeográfica.
Tabla 5. Análisis univariado sobre factores de riesgo ante la presencia de ETS.
RESULTADOS
Se encuestaron 4.018 productores ganaderos sobre un total de 3334 establecimientos agropecuarios con un pro-medio de 1,1 (±0,3), 1,25 (±0,6) y 1,3 (±0,6) productores por establecimiento para las Z1, Z2 y Z3, respectivamen-te. En el presente trabajo se incluyeron un total de 684116 vacas y 33969 toros comprendiendo el 54% y 51,5% de las existencias provinciales, respectivamente. Se detecta-ron 622 rodeos con al menos 1 toro positivo a TB, CGB o ambas enfermedades a la vez, estableciéndose una preva-lencia del 15,5% en los rodeos (tabla 2).
Se estableció una media de 8,7 (±12,2) toros por pro-ductor agropecuario encuestado, hallando una prevalencia para toda el área de estudio de 2,1% (731) y 2,4% (802) de toros positivos a TB y CGB, respectivamente. En la tabla 3 se detallan las prevalencias halladas discriminadas por zonas fitogeográficas.
A nivel de rodeo se halló una prevalencia de 8,1% y 10,4% a TGB y CGB respectivamente, los valores discri-minados por zona fitogeográfica se detallan en la tabla 4. Se observó un incremento de la prevalencia en función de los rodeos con diagnóstico positivo desde la Z1 a la Z3, concordando con las características de manejo se-miintensivas a extensivas, estableciéndose diferencias entre las zonas evaluadas para TB (χ2=116,2; p=<0,001) y CGB (χ2=97,6; p=<0,001), respectivamente. Similares re-sultados fueron hallados considerando campo “positivos a ETS” independientemente de la ETS presente en el rodeo (χ2=142,2; p=<0,001).
FACTOR DE RIESGO
Proporción productores (N.o/total ) Odds Ratio
Intervalo de Confianza 95% Valor p
Inferior Superior
Porcentaje de preñez: Bajo porcentaje de preñez
TotalZ1Z2Z3
13% (153 / 1211)10% ( 85 /837)17% ( 47 /276)12% (15 / 49)
1,82,31,11,9
1,3 - 2,61,3 – 3,20,6 - 2,10,4 – 7,6
<0,0010,0020,7740,377
Presencia involuntaria de animales vecinos en el establecimiento: Frecuente/Esporádica
TotalZ1Z2Z3
20% (784 / 3823)15% (389 / 2543)27% (250 / 932)42% (145 / 348)
1,81,31,61,6
1,5 - 2,20,9 - 1,71,1 - 2,21,0 - 2,5
<0,0010,1400,0060,032
Tipo de servicio: Continuo (> 6 meses)
TotalZ1Z2Z3
41% (1510 / 3668)41% (1032/ 2531) 37% (313 / 856)59% (165 / 281)
1,31,01,42,3
1,1 - 1,60,8 - 1,31,0 - 2,01,4- 3,9
0,0020,9750,0260,001

Prevalencia y factores de riesgo asociados a las enfermedades de transmisión sexual en bovinos de la provincia de La Pampa, Argentina
ARTÍCULOS RIA / Vol. 46 / N.º 2192192
En la tabla 5 se muestran los resultados más relevantes en cuanto a variables de riesgo analizadas. Se determi-nó un porcentaje de preñez promedio general del 83,6% (±12,3) evaluado sobre un total de 1255 productores, esta-bleciéndose 85,1% (±11,9), 80,5% (±13,4) y 76,5% (±13,3) para las Z1, Z2 y Z3, respectivamente. Por un lado la pre-sencia de ETS se asoció estadísticamente al bajo porcen-taje de preñez (P<0,001) utilizando como punto de corte el promedio de preñez general (83,6%) (tabla 5). Por otro lado las ETS estuvieron asociadas a un mayor número de vientres (χ2=12,4; p<0,001) en el rodeo.
De un total de 1716 productores que respondieron la en-cuesta (Z1: 10247, Z2: 535 y Z3: 154) se pudieron consignar datos de interés referentes al número de toros empleados en el servicio extensivo. Para el presente análisis se excluyeron aquellos productores que contaron, al momento de la en-cuesta, con un número inferior a 100 vientres. Se determinó un promedio de 4,1% (±1,4) de toros por cada 100 vientres en servicio encontrándose porcentajes de toros variables según zonas y por ende, ambientes y diferentes manejos, entre los establecimientos, siendo de 3,7% (±1,4), 4,5 (±1,7) y 5,1 (±1,7) para las Zonas 1, 2 y 3 respectivamente.
La presencia de ETS se asoció a un mayor número de toros por vientre (χ2=35,1; p=<0,0001) evaluado a través de la conformación de dos categorías utilizando para ello un punto de corte del 4%. Así mismo se hallaron diferen-cias estadísticas (χ2=128,1; p<0,0001) entre las zonas fi-togeográficas analizadas observándose un mayor porcen-taje de productores con alto porcentaje de toros, en la Z3 (67%), seguido por la Z2 (57%) y por último la Z1 (32%), respectivamente. Un total de 3765 productores desteta los terneros a los 6,1 (±1,6) meses. El 25% (666), 26% (254) y 30% (106) de los productores expresaron que poseían regular o mal estado de los alambrados para las Z1, Z2 y Z3, respectivamente, no hallándose diferencias para esta variable. La reposición de toros, de acuerdo a la informa-ción brindada por 3968 productores, el 16% (649), 39% (1559) y 44% (1760) utilizaron toros Propios, de Compra y Ambas opciones a la vez, respectivamente, no hallando diferencias para esta variable de interés. El 65% (1637), 72% (663) y el 67% (224) de los productores de las Z1, Z2 y Z3, respectivamente, solicitaron certificado negativo a ETS al momento de la compra de los toros. Si bien, un alto porcentaje de productores no lo hace o lo hace esporádi-camente, no se pudo asociar esta variable a la presencia de las ETS en los rodeos.
En relación con el contacto de animales del rodeo con animales de rodeos vecinos o viceversa y para una mejor interpretación de los resultados, la variable respuesta “fre-cuente” fue agrupada con la variable “esporádica” consi-deradas ambas variables como presencia efectiva de con-tacto entre animales. En tanto que la variable respuesta “raro” fue considerada como ausencia de contacto. Se ha-lló una prevalencia en rodeos del 22,4% (176/784) y 13,7% (417/3039) a ETS de productores que declararon la pre-sencia de animales de establecimientos linderos o vicever-sa, en forma frecuente/esporádica y raro, respectivamente, hallándose diferencias estadísticas significativas (χ2=36,2;
p=<0,0001). El análisis de riesgo univariado por zona nos permite determinar que estas diferencias se observaron en las Z2 (χ2=7,5; p=<0,006) y Z3 (χ2=4,6; p=<0,032) pero no así en la Z1 (tabla 5).
El 41% (1510) y el 59% (2158) de los productores realiza el servicio continuo (>6meses) y estacionado (<6 meses), respectivamente. En tanto que estacionan el servicio el 59%, 63% y 41% de los productores considerando las Z1, Z2 y Z3 respectivamente. La permanencia de los toros a lo largo del año en servicio como variable de riesgo estuvo asociado a la presencia de ETS (χ2=9,3; p=0,002). Estos resultados tu-vieron un similar comportamiento en la Z2 (χ2=4,9; p=0,027) y Z3 (χ2=10,0; p=0,001), respectivamente, no hallándose di-ferencias en el análisis para la Z1 (tabla 5).
El servicio de los vientres se realiza durante los me-ses de octubre (46,3%) y noviembre (37,0%) Z1, octubre (40,0%) y noviembre (46,2%) Z2 y octubre (24,6%), no-viembre (58,5%) y diciembre (12,7%) para la Z3; con una duración de 3,1 (±1,2), 3,5 (±1,3) y 3,8 (±1,2) meses para cada zona, respectivamente.
Las prevalencias a ETS en aquellos productores que no realizan tacto (64,8%, 2423) en relación con aquellos productores que sí realizan tacto (35,2%, 1316) fueron de 16,4% y 13,9%, respectivamente, estableciéndose diferen-cias significativas (χ2=4,0; p=0,045). La escasa adopción de dicha tecnología fue hallada como factor de riesgo en el presente trabajo (OR=1,2; IC: 1,0-1,5).
El 14,2% (495) de 3484 productores describe haber te-nido problemas de abortos, de vacas abortadas o de am-bas condiciones en su rodeo. Esta descripción estuvo aso-ciada a la presencia de ETS (OR=1,3; IC: 1,0-1,7) (χ2=6,3; p=0,012). Un similar comportamiento se pudo observar en la Z1 (χ2=16,0; p=0,001) no así para los rodeos de las Z2 y Z3.
DISCUSIÓN
El contacto de animales del rodeo con animales de esta-blecimientos vecinos (toros/vacas), el servicio continuo, la descripción por parte del productor de presencia de abor-tos y un bajo porcentaje de preñez en el rodeo fueron los factores de riesgo que se asociaron significativamente a la presencia de ETS en el presente estudio. Asimismo, se observó un alto porcentaje de toros por vientre en servicio en rodeos infectados. Esta práctica de manejo que se ob-serva frecuentemente tiene su explicación en el aumento de las copulas realizadas por los toros debidas a las fallas en la concepción, ocasionando un desgaste excesivo e in-cremento de toros en los rodeos.
Trabajos realizados en La Pampa Húmeda en la región de la Cuenca del Salado por Quiroz García et al. (2002) mostraron resultados superiores a TB, pero significativa-mente inferiores en cuanto a CGB. En tanto que en regiones semiáridas con características agroecológicas análogas a la región en estudio se reportan resultados de prevalencias a nivel de rodeo y toros similares a los hallados en el pre-sente estudio (Rossanigo et al., 2004). Los resultados a

MIRANDA, A.; FORT, M.; FUCHS, L.; BALDONE, V.; GIMÉNEZ, H.; CARNÉ, L.; SAGO, A.; OTERMIN, H.; DUBIÉ, D.; GOYENECHE, P.; CAMPERO, C.
Agosto 2020, Argentina 193193
por Mardones et al. (2008) y Jiménez et al (2011), situa-ción fitogeográfica similar a la Z1 del presente estudio, los cuales presentaron 2,3 y 3,1 más riesgo de tener la en-fermedad ante la presencia de abortos, respectivamente. Por un lado, el estacionamiento del servicio permite orde-nar los rodeos, programar la época de parición con la de mayor oferta forrajera, concentrar la parición mejorando el manejo del rodeo y predecir posibles fallas reproductivas en los vientres, situación que se observó en el presente trabajo al asociar estadísticamente la mayor presencia de enfermedades en servicios continuos (OR=1,3, p=0,002). Por otro lado, la compra de reproductores negativos a TGB y CGB actuó como factor de protección concordando con los resultados reportados por Jiménez et al. (2011) en la provincia de Buenos Aires.
Una correcta identificación de los factores de riesgo aso-ciados a la presencia de ETS constituye una herramienta de gran valor para la toma de decisiones. Esto permitirá implementar medidas de manejo en el rodeo que mejora-rán los índices reproductivos, sobre todo en un contexto en donde la prevención mediante vacunas, fundamentalmente a TB, todavía es discutible (Baltzell, 2013).
CONCLUSIÓNES
Las prevalencias halladas para TB (2,1%) y CGB (2,4%) permiten avanzar sobre el conocimiento epidemiológico de dichas enfermedades así como también poder realizar es-timaciones sobre pérdidas económicas consecuentes ge-neradas en los rodeos de la provincia de La Pampa. Iden-tificar factores de riesgo inherentes a la presencia de las ETS como servicio continuo, traspaso de animales (toros/vacas) de vecinos y la no realización de tacto rectal per-miten avanzar sobre el control de TB y CGB, evitando la perpetuidad de dichas enfermedades en los rodeos.
AGRADECIMIENTOS
Se agradece la desinteresada colaboración de los pro-ductores agropecuarios y veterinarios privados por la in-formación suministrada, al Ministerio de la Producción del Gobierno de la provincia de La Pampa, al Colegio Médico Veterinario de la provincia de La Pampa, al SENASA y a todo el personal del grupo de sanidad animal de la EEA INTA Anguil, por el compromiso generado con la sanidad de los rodeos de nuestra provincia.
BIBLIOGRAFÍAAGNEW, D.W.; MUNSON, L.; COBO, E.R.; OLESEN,
D.;CORBEIL, L.B.; BONDURANT, R.H. 2008. Comparative histo-pathology and antibody responses of non-Tritrichomonas foetus trichomonad and Tritrichomonas foetus genital infections in virgin heifers. Veterinary Parasitology. 151, 170-180.
ANDERSON, M.L. 2007. Infectious causes of bovine abortion during mid-to late-gestation. Theriogenology. 68, 474-486.
BALTZELL, P.; NEWTON, H.; O’CONNOR, A.M. 2013. A Criti-cal Review and Meta-Analysis of the Efficacy of Whole-Cell Killed
TB, obtenidos por Fort et al. (2004a) en la provincia de La Pampa son similares a los hallados en el presente trabajo, en tanto que a nivel de rodeo se describen prevalencias algo inferiores. En cuanto a CGB las prevalencias son le-vemente superiores a las descriptas por el mismo autor, diferencias que no se observaron a nivel de rodeo (Fort et al. 2004b). Considerando que los resultados hallados en el presente trabajo y los reportados previamente por Fort et al. (2004 a, b) corresponden a la misma región bajo estu-dio con valores similares de prevalencias, se puede inducir que la información generada sistemáticamente por labora-torios de la actividad privada constituye una herramienta epidemiológica de gran utilidad, sobre todo en áreas en donde se carece de información previa basada en estudios epidemiológicos exploratorios (Rojas, 2011).
Por un lado, los valores de prevalencias más altos a ETS hallados en la Z3 y Z2 se explicarían por diferentes razo-nes a saber: en primer lugar, se observa un mayor número de rodeos por establecimiento, esta práctica común entre ganaderos conlleva a que animales de distinta procedencia compartan pasturas, instalaciones y muchas veces machos reproductores, favoreciendo el contacto sexual de animales y perpetuando, de esta manera, las enfermedades en los rodeos. Por otro lado y en el mismo sentido, el contacto de animales vecinos constituyó el factor de riesgo que mostró mayores diferencias estadísticas, entre aquellos producto-res con diagnóstico positivo a TGB y CGB, aumentando el riesgo de contraer las enfermedades 1,8 veces. De igual manera, estudios previos (Mardones et al. 2008; Jiménez et al. 2011) realizados en rodeos de la provincia de Buenos Aires reportaron esta variable como un importante factor de riesgo. La infertilidad de los vientres, observada a través de un bajo porcentaje de preñez al tacto rectal, así como un porcentaje mayor de preñeces correspondientes al final del servicio, constituye una de las primeras manifestaciones clínicas sobre la presencia de las enfermedades. En el pre-sente estudio se pudo determinar que el riesgo de presen-tar una u otra enfermedad o ambas a la vez aumentaba 1,8 veces para aquellos establecimientos con hallazgo positivo y bajo número de vientres preñados. Similares resultados fueron hallados por Mardones et al. (2008) y Jiménez et al. (2011), quienes asociaron estadísticamente esta variable de riesgo a la presencia de ETS estableciendo un OR de 4,1 y 2,0 para cada uno respectivamente.
Otro de los factores evaluados y consultados al produc-tor agropecuario fue la presencia de abortos y vacas abor-tadas en el rodeo. Las ETS causan muerte embrionaria o aborto espontáneo en el primer trimestre de la gestación, aunque ocasionalmente puede ocurrir entre el 4 y 7 meses (BonDurant, 2007; Anderson, 2007). Se halló asociación entre la presencia de ETS y los reportes de abortos para la Z1 (OR: 1,8). Estos resultados posiblemente se deban a que esta zona se caracteriza por manejos más intensivos con superficies significativamente inferiores al resto de las regiones, con un mayor número de potreros y campos lim-pios (sin bosque natural), condición que facilitaría la iden-tificación de vientres o del feto abortado. Estos resultados son similares a los hallados en la provincia de Buenos Aires

Prevalencia y factores de riesgo asociados a las enfermedades de transmisión sexual en bovinos de la provincia de La Pampa, Argentina
ARTÍCULOS RIA / Vol. 46 / N.º 2194194
Tritrichomonas foetus Vaccines in Beef Cattle. J Vet Intern Med.; 27: 760-770.
BELLINI SAIBENE, Y.; RAMOS, L.; CASAGRANDE, G.; VER-GARA, G. 2011. El uso de AgroTics para generar las cartas agro-climáticas de la provincia de La Pampa. (Disponible: www.inta.gob.ar verificado: 11 de abril de 2008.
BONDURANT, R.H.; ANDERSON, M.L.; BLANCHARD, P.; HIRD, D.; DANAYE-ELMI, C.; PALMER, C. SISCHO, W.M.; SUTHER, D. UTTERBACK, W.; WEIGLER, B.L. 1990. Prevalen-ce of Trichomoniasis among California beef herds. J Am Vet Med Assoc. 196, 1590-3.
BONDURANT, R.H. 2007. Selected diseases and conditions associated with bovine conceptus loss in the first trimester. Therio-genology 68, 461-473.
CAMPERO, C.M.; CATENA, M.C., MEDINA, D. 1986. Caldo infusión hígado para el cultivo de Tritrichomona foetus. Vet. Arg. 3:80-81.
CAMPERO, C.M.; MARTINEZ, A. 2010. Enfermedades de transmisión sexual en los bovinos. Actualización en Enfermedades de los Bovinos. Azul. Bs. As. 9 p.
FORT, M.C.; ROJAS, M.C.; ESAÍN, F.H; PÉREZ, L.R. 2004a. El control de la Campylobacteriosis genital bovina en 5 departamen-tos de la provincia de La Pampa durante el período 2000-2003. Publicación técnica N.º 58. EEA INTA Anguil, La Pampa. 5 p.
FORT, M.C.; ROJAS, M.C.; PÉREZ, L.R.; ESAÍN, F.H. 2004b. El control de la Trichomoniasis genital bovina en siete departamentos de la provincia de La Pampa durante el período 2000-2003. Publi-cación técnica N.º 58. EEA INTA Anguil, La Pampa. 9 p.
GROTELUESCHEN, D.M.; CHENEY, J.; HUDSON, D.B.; SCHWEITZER, D.J.; KIMBERLING, C.V.; TATON-ALLEN, G.F.; NIELSEN, K.A.; MARASH, D.J. 1994. Bovine Trichomoniasis: Re-sults of a salughter survey in Colorado and Nebraska. Theriogeno-logy. 42 (1), 165-171.
HUM, S.; HORNITZKY, M.; BERG, T. 2009. Bovine Genital Cam-pylobacteriosis. Australia and New Zealand Standard Diagnostic Procedures. (Disponible: http://www.scahls.org.au verificado: 01 de marzo de 2016).
JIMÉNEZ, D.F.; PEREZ, A.M.; CARPENTER, T.E.; MARTINEZ, A. 2011. Factor associated with infection by Campylobacter fetus in beef herds in the Province of Buenos Aires, Argentina. Preventive Veterinary Medicine. 101, 157-162.
KVASNICKA, W.G.; TAYLO, R.E.L.; HUAG; HANKS, D.; TRONSTAD, R.J.; BOSOMWORTH, A.; HALL, M.R. 1989. Investi-gations of the incidence of bovine trichomoniasis in Nevada and of the efficacy of immunizing cattle with vaccines containing Tritricho-monas foetus. Theriogenology 31 (5), 963-971.
MARTÍN-GÓMEZ, S.; GONZÁLES-PANIELLO, R.; PEREIRA-BUENO, J.; ORTEGA-MORA, L.M. 1998. Prevalence of Tritricho-monas foetus infection in beef bulls in northwestern Spain. Veteri-nary Parasitology 75, 265-268.
MARDONES, F.O.; PEREZ, A.M.; MARTÍNEZ, A.; CARPEN-TER, T.E. 2008. Risk factors associated with Tritrichomonas foetus infection in beef herds in the Province of Buenos Aires, Argentina. Veterinary Parasitology. 153, 231-237.
ONDRAK, J. 2016. Tritrichomonas foetus Prevention and Con-trol in Cattle. Vet Clin Food Anim 32: 411-423.
PARSONSON, I.M.; CLARK, B.; DUFTY, J. 1974. The patho-genesis of Tritrichomonas foetus infection in the bull. Aust. Vet. J. 50: 421-423.
PÉREZ, E.; CONRAD, P.A.; HIRD, D.; ORTUNO, A.; CHACON, J.; BONDURANT, R.; NOORDHUIZEN, J. 1992. Prevalence and
risk factors for Trichomonas foetus infection in cattle in northeas-tern Costa Rica. Prev. Vet. Med. 14, 155-165.
QUIROZ GARCÍA, J.L.; MARESCA, S., VERDIER, M., 2002, Diagnóstico de enfermedades venéreas en la cuenca del Salado, provincia de Bs As. Años 2001.E 22. xiv Reunión Científico Técni-ca, Asoc. Arg. Vet. Lab. Diag. Gral Belgrano, Córdoba.
RAE, D.O.; CHENOWETH, P.J.; GENHO, P.C.; MCINTOSH, A.D. CROSBY, C.E.; MOORE, S.A. 1999. Prevalence of Tritricho-monas fetus in a bull population and effect on production in a large cow-calf enterprise. J. Am. Vet. Med. Assoc. 214, 1051-1055.
RAE, D.O.; CREWS, J.E.; GREINER, E.C.; DONOVAN, G.A. 2004. Epidemiology of Tritrichomonas foetus in beef bull popula-tions in Florida. Theriogenology. 61, 605-618.
REPAGRO (REGISTRO PROVINCIAL AGROPECUARIO). 2008. (Disponible: http://www.estadistica.lapampa.gov.ar verifica-do: 01 de marzo de 2016).
REPISO, M.V.; GIL, A.; BAÑALES, P.; D´ANATRO, N.; FER-NÁNDEZ, L.; GUARINO, H.; HERRERA, B.; NÚÑEZ, A.; OLIVE-RA, M.; TAKESHI, O.; SILVA, M. 2005. Prevalencia de las princi-pales enfermedades infecciosas que afectan el comportamiento reproductivo en la ganadería de carne y caracterización de los establecimientos de cría del Uruguay. INIA. Serie FPTA N.º 13.
RILEY, D.E.; WAGNER, B.; POLLEY, L.; KRIEGER, J. 1995. PCR-based study of conserved and variable DNA sequences of Tritrichomonas foetus isolates from Saskatchewan, Canada. J. Clin. Microbiol. 33 (5), 1308-13.
ROJAS, M.; VÁZQUEZ, P.; VERDIER, M.; CAMPERO, C.M. 2011. Evolución y distribución de las enfermedades de transmisión sexual en bovinos del partido de Rauch, prov. Buenos Aires, años 2001-2009. Veterinaria Argentina. 18: 273. (Disponible: http://www. veterinariar-gentina.com./revista/2011/01 verificado: 04 de octubre de 2018).
ROSSANIGO, C.E.; ÁVILA, J.D.; LÓPEZ ROCA, A.; INSUA, C.;
PIVIDAL, J. 2004. Las enfermedades venéreas en los rodeos de cría bovina de la región semiárida subhúmeda central. Prevalen-cia, diagnóstico y control. (Disponible: http://www.produccion-ani-mal.com.ar verificado: 04 de octubre de 2018).
SENASA (SERVICIO NACIONAL DE SANIDAD Y CALIDAD AGROALIMENTARIA). 2008. Resolución Nº 358/08, decreto Nº 374/07. Gobierno de la provincia de La Pampa. (Disponible: http://www.senasa.gov.ar verificado: 11 de abril de 2013).
STOESSEL, F. 1982. Las enfermedades venéreas de los bovi-nos: Trichomoniasis y vibriosis genital. 1.a ed. Acribia. Zaragoza, España. 6-16 pp.
TERZOLO, H.R.; ARGENTO, E.; CATENA, M.C.; CIPOLLA, A.L.; MARTINEZ, A.H.; TEJADA, G.; VILLA, C.; BENTANCOR, L.; CAMPERO, C.M.; CORDEVIOLA, J.M.; PASINI, M.I. 1992. Procedimientos de laboratorio para el Diagnóstico de la Campylo-bacteriosis y Tricomonosis Genital Bovina. Documento de la Co-misión Científica Permanente de Enfermedades Venéreas de los Bovinos. Asociación Argentina de Veterinarios de Laboratorios de Diagnóstico. Centro Regional Buenos Aires Sur (CERBAS) INTA Balcarce. 1-33 pp.
YULE, A.; SKIRROW, S.Z.; BONDURANT, R.H. 1989. Bovine trichomoniasis. Parasitology today 12, 373-377.
SZONYI, B.; SRINATH, I.; SCHWARTZ, A.; CLAVIJO, A.; IVA-NEK, R. 2012. Spatio-temporal epidemiology of Tritrichomonas foetus infection in Texas bulls based on state-wide diagnostic labo-ratory data. Veterinary Parasitology, 186, 450-455.

PETIGROSSO, L.R.; NAVARRO, D.; ASSUERO, S.G.; VIGNOLIO, O.R.; CASTAÑO, A.; COLABELLI, M.N.
Agosto 2020, Argentina 195195
Respuesta a la frecuencia de defoliación de plantas de festuca alta de una población naturalizada infectada con endófito silvestre y un cultivar comercial libre
RESUMENEl objetivo del presente trabajo fue evaluar el efecto de dos frecuencias de defoliación sobre el crecimiento
vegetativo y reproductivo de plantas de festuca alta provenientes de semilla cosechada en una población na-turalizada e infectada con Epichloë coenophiala (FNE+) y de un cultivar comercial libre de endófito (FCE-, cv. Continental Taita). Además, en las plantas de FNE+ se analizó el efecto de la frecuencia de defoliación sobre la transmisión vertical del endófito a las semillas. La siembra se realizó el 02/05/2016 y la densidad de plantas para todos los tratamientos fue de 200 plantas m-2. Los factores experimentales fueron: tipo de micropastura de festuca alta (FNE+ y FCE-) y frecuencia de defoliación (AF: alta y BF: baja, defoliaciones cada 450±100°Cd y cada 850±100°Cd, respectivamente, en el período comprendido entre el 20/10/2016 y el 16/08/2017). Se realizaron 8 cortes para el tratamiento AF y 4 cortes para el BF. Se empleó un diseño experimental completa-mente aleatorizado con arreglo factorial y 5 repeticiones. Se halló que con AF las plantas de FNE+ presenta-ron mayor acumulación de biomasa total que las FCE- (168,19±8,84 y 149,12±3,74 g MS m-2 respectivamen-te, p=0,009), mientras que con BF la situación fue inversa (FNE+: 104,13±7,07 y FCE-: 120,13±5,79 g MS m-2, p=0,009). El número de macollos en las plantas de ambos tipos de micropastura fue mayor con AF que con BF (p<0,0001); el promedio fue 2346±302 y 1940±250 macollos m-2 respectivamente. El número de panojas por planta fue mayor en FNE+ que en FCE- (2,85±0,20 y 1,55±0,34, respectivamente, p=0,002) y no fue afectado por la frecuencia de defoliación (p=0,065). Las plantas de FNE+ tendieron a mostrar mayor esfuerzo repro-ductivo que las FCE-, aunque la diferencia solo fue significativa en BF. La producción de semillas fue mayor en las plantas de FNE+ que en las de FCE- (316,44±35,03 y 183,53±18,19 semillas/planta, respectivamente, p=0,0033) y no fue afectada por la frecuencia de defoliación (p=0,2581), aunque el aumento de la frecuencia de defoliación tendió a disminuir la producción de semillas en las plantas FNE+. Todas las semillas analizadas producidas por las plantas FNE+ resultaron positivas al diagnóstico de endófito, por lo que no hubo efecto de la frecuencia de defoliación sobre la transmisión de este. En conclusión, a pesar de que la frecuencia de defoliación afectó en forma diferencial la producción de biomasa total por planta en ambos tipos de micropas-turas, las plantas de FNE+ siempre mostraron una mejor performance reproductiva. Esta última respuesta y la perfecta transmisión del endófito a las semillas observadas en ambas frecuencias de defoliación contribuirían a explicar el avance de los porcentajes de plantas infectadas comúnmente observado a campo.
Palabras clave: Festuca arundinacea, Epichloë coenophiala, producción de biomasa, número de macollos por planta, esfuerzo reproductivo, transmisión vertical del endófito.
1Universidad Nacional de Mar del Plata, Facultad de Ciencias Agrarias, Ruta 226 km 73,5, (7620), Balcarce, Argentina. Correo electrónico: [email protected]; [email protected]; [email protected]; [email protected]; [email protected] Nacional de Tecnología Agropecuaria (INTA), Centro Regional Buenos Aires Sur, Estación Experimental Agropecuaria (EEA), Bal-carce, Ruta 226 km 73,5, (7620), Balcarce, Argentina. Correo electrónico: [email protected]
PETIGROSSO, L.R.1; NAVARRO, D.1; ASSUERO, S.G.1; VIGNOLIO, O.R.1; CASTAÑO, J.A.1,2; COLABELLI, M.N.1
Recibido 05 de julio de 2018 // Aceptado 15 de marzo de 2019 // Publicado online 09 de septiembre de 2020

Respuesta a la frecuencia de defoliación de plantas de festuca alta de una población naturalizada infectada con endófito silvestre y un cultivar comercial libre
ARTÍCULOS RIA / Vol. 46 / N.º 2196196
ABSTRACT
The objective of the present work was to evaluate the effect of two defoliation frequencies on the vegetative and reproductive growth of tall fescue plants originated from seed harvested in a naturalized population infec-ted with Epichloë coenophiala (FNE+) and from a commercial endophyte-free cultivar (FCE-, cv Continental Taita). In addition, the effect of the defoliation frequencies applied on the vertical transmission of the endophyte to the seeds produced by FNE+ plants was analyzed. On 2 May 2016 seeds were sown in micropastures at a density of 200 plants m-2. The experimental factors were: type of tall fescue micropasture (FNE+ and FCE-) and frequency of defoliation (AF: high and BF: low, defoliated every 450±100°Cd and every 850±100°Cd, res-pectively, during the period between 20 October 2016 and 16 August 2017). Eight cuts were carried out in AF and 4 cuts in BF. A completely randomized experimental design with a factorial arrangement and five replicates was used. Under AF, FNE+ plants presented higher accumulation of total biomass than FCE- (168.19±8.84 and 149.12±3.74 g DM m-2 respectively, p=0.009), while the opposite occurred under BF (FNE+: 104.13±7.07 and FCE-: 120.13±5.79 g DM m-2, p=0.009). Independently of micropasture type, the number of tillers per plant was higher under AF than under BF (2346±302 and 1940±250 tillers m-2 respectively, p<0.0001). The number of panicles per plant was higher in FNE+ than in FCE- (2.85±0.20 and 1.55±0.34, respectively, p=0.002) and was not affected by the frequency of defoliation (p=0.065). The FNE+ plants tended to show a greater repro-ductive effort than FCE-, although the difference was only significant under BF. Seed production was higher in FNE+ than in FCE- plants (316.44±35.03 and 183.53±18.19 seeds/plant, respectively, p=0.0033) and it was not affected by the defoliation frequency (p=0.2581), although the highest frequency of defoliation tended to decrease the seed production of FNE+ plants. All the analyzed seeds produced by FNE+ plants were positive to the endophyte diagnosis, so endophyte transmission was not affected by the frequency of defoliation. In conclusion, despite the fact that the defoliation frequency affected differentially total plant biomass of both types of micropastures, FNE+ plants always showed a better reproductive performance than FCE- plants. The last finding together with the fact that the transmission of the endophyte to the seeds was perfect and not affected by the defoliation frequency, would help to explain the progress of the percentages of infected plants commonly observed in the field.
Keywords: Festuca arundinacea, Epichloë coenophiala, biomass production, number of tillers per plant, reproductive effort, endophyte vertical transmission.
INTRODUCCIÓN
Festuca alta, Schedonorus arundinaceus (Schreb.) Du-mort [sinónimos: Festuca arundinacea Schreb., Lolium arundinaceum (Schreb.) S.J. Darbyshire, Schedonorus phoenix (Scop.) Holub], es una gramínea C3 perenne muy valorada en los sistemas de producción ganadera por su adaptación a un amplio rango de condiciones edafoclimáti-cas y su alto potencial de producción de forraje (Hannaway et al., 1999; Fribourg y Hannaway, 2007; Scheneiter et al., 2015). Al igual que otras gramíneas C3 puede establecer una relación generalmente simbiótica con hongos endófitos asexuales de transmisión vertical (Saikkonen et al., 1998). El endófito que infecta festuca alta, Epichloë coenophiala (Leuchtmann et al., 2014) ex Neotyphodium coenophialum Glenn, Bacon & Hanlin (= Acremonium Link Sección Albo-lanosa Morgan-Jones; Gams) (Glenn et al., 1996), desarro-lla su ciclo de vida dentro de la planta hospedante, es asin-tomático y su dispersión tiene lugar únicamente a través de semillas infectadas, lo que se conoce como “transmisión vertical” (White et al., 1993; Clay y Schardl, 2002). Por un
lado, la importancia agronómica de esta asociación está dada por los efectos positivos inducidos por la simbiosis pasto-hongo sobre el crecimiento y la tolerancia a estreses bióticos y abióticos (Malinowski y Belesky, 2000; White y Torres, 2009; Omacini et al., 2013) y por causar “festuco-sis”. Esta última hace referencia a distintas patologías que los alcaloides tóxicos producidos por el hongo provocan al ganado que consume plantas infectadas (Bacon et al., 1977; Hoveland, 1993; De Battista et al., 1995).
Por otro lado, se ha observado que pasturas de festuca alta sembradas con semilla libre de endófito son suscep-tibles a la invasión por plantas silvestres infectadas (ej.: resiembra natural); los mecanismos de contaminación son con semillas infectadas presentes en el banco o inmigra-ción desde los bordes o campos aledaños, un factor clave en el avance de la infección endofítica (Shelby et al., 1989; Shelby y Schmidt, 1991; Shelby y Dalrymple, 1993; Bar-ker et al., 2005). Estos mecanismos dependerán del tipo de pastura y del manejo de esta. Por ejemplo, se pueden mencionar el uso de cultivares de festuca libre de endófito

PETIGROSSO, L.R.; NAVARRO, D.; ASSUERO, S.G.; VIGNOLIO, O.R.; CASTAÑO, A.; COLABELLI, M.N.
Agosto 2020, Argentina 197197
proveniente del horizonte A de un Argiudol típico (53 ppm de P Bray I, 4,3% MO y 30,1 ppm N-NO3
-) en contenedores de 0,6 m de largo x 0,4 m de ancho x 0,2 m de profundidad ubicados al aire libre.
Se empleó semilla de festuca alta infectada con endófito silvestre (100% de infección, poder germinativo 82%) pro-veniente de un pastizal del partido de Mar Chiquita (37° 32’ S; 57° 55’ O) y semilla de festuca alta cultivar Continental TAITA, libre de infección (0% de infección, poder germina-tivo 87%), suministrada por GENTOS S.A. Los niveles de infección se corroboraron previo a la siembra empleando la técnica de Saha et al. (1988). Para ello, se realizó un pretratamiento que consiste en sumergir las semillas en hi-dróxido de sodio NaOH al 5% a temperatura ambiente por 12 h. Al cabo de este tiempo las semillas se enjuagaron con agua y se colocaron individualmente sobre un portaobjeto para separar las glumelas y proceder a su tinción (colora-ción directa) con una gota del colorante rosa de Bengala durante 2-3 minutos. Posteriormente se colocó un cubre-objeto y se observó al microscopio óptico Olympus CHK (400x) la presencia de hifas del hongo entre las células aleuroníferas del endosperma. Así, las semillas que conte-nían el endófito fueron diagnosticadas como positivas (E+) y las no infectadas como negativas (E-).
La siembra se realizó el 02/05/2016 ubicando 13 semi-llas a 5 mm de profundidad en cada uno de cuatro surcos distanciados 12 cm entre sí (52 plantas por contenedor, ej. 200 pl m-2). Las fallas en la emergencia se subsanaron dentro de los 30 días después de la siembra, mediante el trasplante de plántulas extra, de tamaño similar, cultivadas paralelamente en almácigos. La fecha de emergencia pro-medio de las plántulas en las unidades experimentales fue el 25/05/2016, equivalente a un tiempo térmico desde la siembra de 144°Cd (temperatura base 4 °C; Colabelli et al., 1998; Agnusdei y Mazzanti, 2001).
El 20/10/16 se llevó a cabo un corte de limpieza para uniformar las micropasturas. A partir de esa fecha, se rea-lizaron 8 cortes para los tratamientos con alta frecuencia y
Tabla 1. Fechas de corte y tiempo térmico acumulado desde el corte de limpieza efectuado el 20 de octubre de 2016 (°Cd, tem-peratura base 4 °C) para dos frecuencias de defoliación: alta (AF, cada 450±100°Cd) y baja (BF, cada 850±100°Cd).
o infectados con endófito amigable (Bouton et al., 2002; Gundel et al., 2013; Lugtenberg et al., 2016), el éxito re-productivo diferencial de plantas infectadas respecto a las libres de endófito (Gundel et al., 2008; 2011), la eficiencia de transmisión del endófito (Gundel et al., 2008; 2011), la aplicación de herbicidas, Defelice y Henning, 1990; Hill et al., 2010; Trevor et al., 2016), la intersiembra con legumino-sas o gramíneas libre de endófito (De Battista et al., 1997; Tekeli y Ates, 2005; Petigrosso et al., 2018), el momento e intensidad de pastoreo (Evans et al., 2012) y el movimiento de los animales entre parcelas (Odriozola et al., 2002).
La transmisión vertical del endófito puede ser imperfecta, es decir, que las plantas infectadas pueden producir una proporción variable de semillas no infectadas, dependien-do de la especie o población en estudio y de las condicio-nes ambientales de crecimiento (Afkhami y Rudgers, 2008; Gundel et al., 2009, 2011; García Parisi et al., 2012). En este sentido, se sabe que la sequía y la aplicación de her-bicidas disminuyen la transmisión vertical del endófito (Da-vitt et al., 2011; Gundel et al., 2011), pero poco se conoce sobre el efecto del pastoreo sobre esta. Por ejemplo, Gar-cía Parisi et al. (2012) analizando el impacto del pastoreo (simulado mediante cortes y pisoteo) sobre la dinámica de transmisión del endófito E. occultans en Lolium multiflorum Lam. (raigrás anual) en microcosmos bajo diferentes con-textos agroecológicos encontraron que la defoliación con pisoteo en dos momentos durante el ciclo de crecimiento (macollaje y encañazón) redujeron la producción de semilla y provocaron fallas en la transmisión del endófito respecto a los tratamientos no defoliados. Sin embargo, es escasa la información acerca de cómo operan los distintos facto-res ecológicos y de manejo agronómico en la regulación del reemplazo de plantas libres por plantas infectadas con endófito de tipo silvestre, que es común observar en las pasturas de la Cuenca del Salado (Colabelli et al., 2006; Petigrosso et al., 2013).
A partir de lo mencionado, y con la finalidad de desarro-llar prácticas de manejo que permitan prevenir o disminuir la invasión de festuca infectada en pasturas libres de en-dófito, el presente trabajo tuvo como objetivo avanzar en el entendimiento de los factores que regulan dicha inva-sión. Para ello, se evaluó el efecto de dos frecuencias de defoliación sobre el crecimiento vegetativo y reproductivo de plantas de festuca alta de una población naturalizada e infectada con Epichloë coenophiala y de un cultivar comer-cial libre de endófito. Además, en las plantas infectadas se analizó el efecto de la frecuencia de defoliación sobre la transmisión vertical del endófito a las semillas.
MATERIALES Y MÉTODOS
Sitio y diseño experimental
El experimento se realizó en la Unidad Integrada Balcar-ce (Facultad de Ciencias Agrarias UNMdP–Estación Expe-rimental Agropecuaria INTA Balcarce; 37° 45’ S, 58° 17’ O, 130 m s. n. m.) en micropasturas (unidades experimenta-les) establecidas sobre suelo tamizado y homogeneizado
FechaFrecuencia de
defoliaciónAF (°Cd) BF (°Cd)
23/11/2016 AF 393 -
19/12/2016 AF y BF 833 833
03/01/2017 AF 1119 -
30/01/2017 AF y BF 1645 1645
24/02/2017 AF 2098 -
27/03/2017 AF y BF 2561 2561
10/05/2017 AF 3092 -
16/08/2017 AF y BF 3701 3701

Respuesta a la frecuencia de defoliación de plantas de festuca alta de una población naturalizada infectada con endófito silvestre y un cultivar comercial libre
ARTÍCULOS RIA / Vol. 46 / N.º 2198198
4 cortes para los de baja frecuencia (tabla 1). La frecuencia de defoliación se estableció considerando el tiempo térmi-co acumulado (frecuencia alta cada 450±100°Cd y baja cada 850±100°Cd respectivamente). Los cortes se realiza-ron a 70 mm de altura desde el nivel del suelo con tijera de mano. El último corte se efectuó el 16/08/17 con el objetivo de permitir que las plantas pasen a estado reproductivo y evitar el corte de las varas florales.
Se empleó un diseño experimental completamente alea-torizado con arreglo factorial. Los factores experimentales controlados fueron: tipo de pastura de festuca alta (FNE+: naturalizada e infectada con endófito silvestre, y FCE-: comercial libre de endófito) y nivel de frecuencia de defo-liación (AF: alta, y BF: baja). En total se establecieron 20 contenedores experimentales: 2 niveles de defoliación x 2 niveles de infección endofítica x 5 repeticiones.
Determinaciones
Producción de biomasa aérea acumulada y número de macollos
Los cortes para alta y baja frecuencia de defoliación se realizaron en promedio a 470°Cd y 950°Cd, respectiva-mente, durante el período comprendido entre el 20/10/16 y el 16/08/17. La biomasa fresca se secó en estufa de aire forzado a 60 °C hasta peso constante y se pesó. En cada micropastura se identificaron 4 plantas de festuca alta en las que se contó el número total de macollos previo a la realización de cada corte.
Número de panojas, esfuerzo reproductivo, peso y número de semillas
En cada una de las 4 plantas marcadas se registró el número de macollos totales (reproductivos y vegetativos) para calcular el esfuerzo reproductivo (ej. panojas/macollos totales). Las panojas se cosecharon entre el 27/11/2017 y el 5/12/2017, cuando el raquis de estas presentaba un co-lor amarillento. Se determinó el peso total de las semillas producidas en cada planta y, a partir de tres submuestras de 100 semillas se estimó el peso promedio de esas 100 semillas y así, con el dato de peso total, obtener el número de semillas por planta.
Determinación de la transmisión vertical del endófito
Se realizó el diagnóstico de la presencia del endófito en 20 semillas de cada una de las cuatro plantas identificada en cada micropastura empleando la técnica de Saha et al. (1988) previamente descripta. Con los resultados obtenidos se cal-culó el porcentaje de semillas infectadas en cada planta.
Datos meteorológicos y mantenimiento de los experimentos
El tiempo térmico acumulado se calculó utilizando los registros de temperatura del aire en casilla meteorológi-ca a 1,50 m de altura de la Estación de Agrometereología de la EEA Balcarce del INTA, ubicada aproximadamente a
300 m del sitio experimental. Los contenedores se mantu-vieron libres de malezas y con adecuada disponibilidad de agua durante el período experimental. Se aplicó nitrógeno (150 kg N ha-1 año-1 en forma dividida: 50 kg N ha-1 a la siembra, 50 kg N ha-1 luego del corte limpieza y 50 kg N ha-1 después del último corte realizado previo a la encañazón). No se observaron ataques de plagas ni patógenos durante el desarrollo del experimento.
Análisis estadístico
Debido a la diferencia en el número de cortes entre ambos tratamientos de frecuencia de defoliación (ej. 8 y 4 valores para alta y baja frecuencia de defoliación, res-pectivamente), el cálculo de biomasa aérea acumulada con alta frecuencia de defoliación se efectuó sumando las cantidades cosechadas en cortes sucesivos de manera de obtener el valor de biomasa acumulada correspondiente al mismo período de acumulación de los tratamientos con baja frecuencia (ej. 4 valores en total para cada frecuencia de defoliación). Los efectos de los factores experimentales y su posible interacción se analizaron mediante ANOVA el cual, para las variables biomasa aérea acumulada y nú-mero de macollos, se efectuó con medidas repetidas en el tiempo. Se realizaron comparaciones de medias (LSD p<0,05) cuando se encontraron diferencias significativas (α=0,05). Debido a que de la variable respuesta número de macollo no cumplió con los supuestos de normalidad ni homogeneidad de varianzas debió transformarse (ej. ln del número de macollos). Todos los análisis se realizaron empleando el software estadístico R (R Development Core Team, 2016). Los modelos se ajustaron con la función lme del paquete nlme (Pinheiro et al., 2012). En la sección re-sultados se muestran como media ± error estándar (EE).
RESULTADOS
Producción de biomasa aérea acumulada y número de macollos
Al analizar la biomasa aérea acumulada de las micropas-turas (figura 1) no se detectó interacción significativa entre el tipo de pastura, la frecuencia de defoliación y la fecha de corte (p=0,117), pero sí se encontró interacción significativa entre el tipo de pastura y la frecuencia de defoliación (p=0,009). En este sentido, se halló que con AF las plantas de FNE+ presentaron mayor acumulación de biomasa total que las de FCE- (168,19±8,84 y 149,12±3,74 g MS m-2 respectivamen-te), mientras que con baja frecuencia la situación fue inversa (FNE+: 104,13±7,07 y FCE-: 120,13±5,79 g MS m-2).
El número de macollos en las plantas de ambos tipos de pastura fue mayor con AF que con BF (p<0,0001) siendo en promedio 2346±302 y 1940±250 macollos m-2 respectivamen-te. El efecto tipo de pastura no fue significativo (p=0,061).
Número de panojas, esfuerzo reproductivo y número de semillas por planta
En la tabla 2 se muestran los resultados de las variables reproductivas. El número de panojas por planta fue ma-
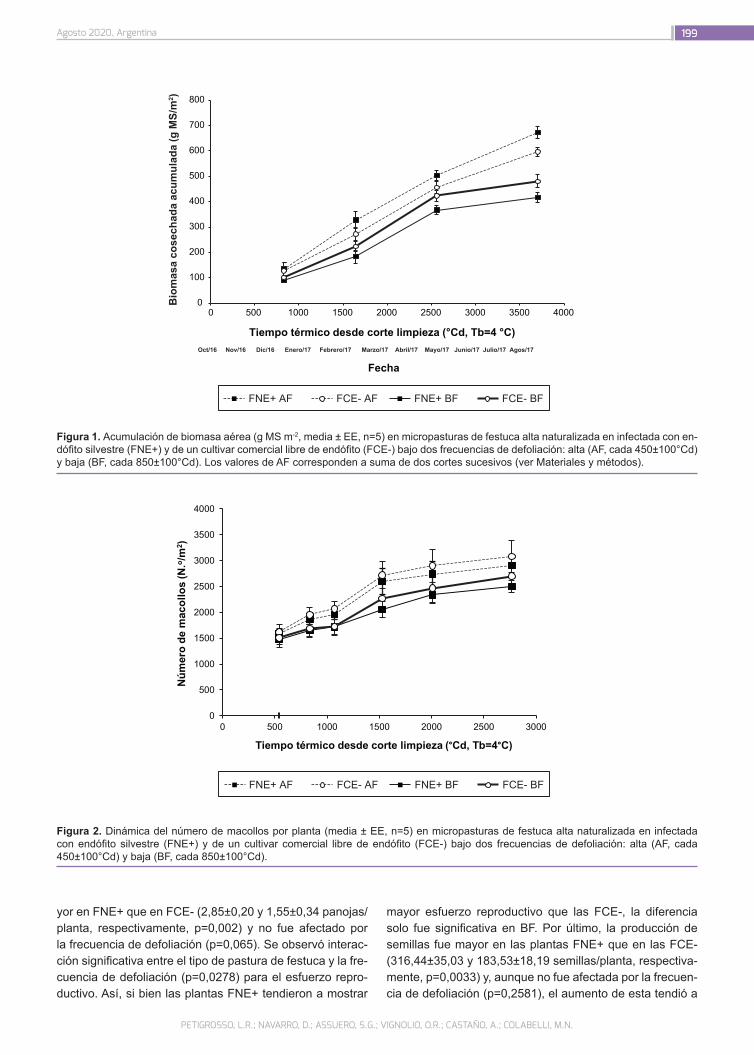
PETIGROSSO, L.R.; NAVARRO, D.; ASSUERO, S.G.; VIGNOLIO, O.R.; CASTAÑO, A.; COLABELLI, M.N.
Agosto 2020, Argentina 199199
Figura 1. Acumulación de biomasa aérea (g MS m-2, media ± EE, n=5) en micropasturas de festuca alta naturalizada en infectada con en-dófito silvestre (FNE+) y de un cultivar comercial libre de endófito (FCE-) bajo dos frecuencias de defoliación: alta (AF, cada 450±100°Cd) y baja (BF, cada 850±100°Cd). Los valores de AF corresponden a suma de dos cortes sucesivos (ver Materiales y métodos).
Figura 2. Dinámica del número de macollos por planta (media ± EE, n=5) en micropasturas de festuca alta naturalizada en infectada con endófito silvestre (FNE+) y de un cultivar comercial libre de endófito (FCE-) bajo dos frecuencias de defoliación: alta (AF, cada 450±100°Cd) y baja (BF, cada 850±100°Cd).
0
100
200
300
400
500
600
700
800
0 500 1000 1500 2000 2500 3000 3500 4000
Bio
mas
a co
sech
ada
acum
ulad
a (g
MS/
m2 )
Tiempo térmico desde corte limpieza (°Cd, Tb=4 °C)
Fecha
Oct/16 Nov/16 Dic/16 Enero/17 Febrero/17 Marzo/17 Abril/17 Mayo/17 Junio/17 Julio/17 Agos/17
FNE+ AF FCE- AF FNE+ BF FCE- BF
0
500
1000
1500
2000
2500
3000
3500
4000
0 500 1000 1500 2000 2500 3000
Núm
ero
de m
acol
los
(N.o /m
2 )
Tiempo térmico desde corte limpieza (°Cd, Tb=4°C)
FNE+ AF FCE- AF FNE+ BF FCE- BF
yor en FNE+ que en FCE- (2,85±0,20 y 1,55±0,34 panojas/planta, respectivamente, p=0,002) y no fue afectado por la frecuencia de defoliación (p=0,065). Se observó interac-ción significativa entre el tipo de pastura de festuca y la fre-cuencia de defoliación (p=0,0278) para el esfuerzo repro-ductivo. Así, si bien las plantas FNE+ tendieron a mostrar
mayor esfuerzo reproductivo que las FCE-, la diferencia solo fue significativa en BF. Por último, la producción de semillas fue mayor en las plantas FNE+ que en las FCE- (316,44±35,03 y 183,53±18,19 semillas/planta, respectiva-mente, p=0,0033) y, aunque no fue afectada por la frecuen-cia de defoliación (p=0,2581), el aumento de esta tendió a
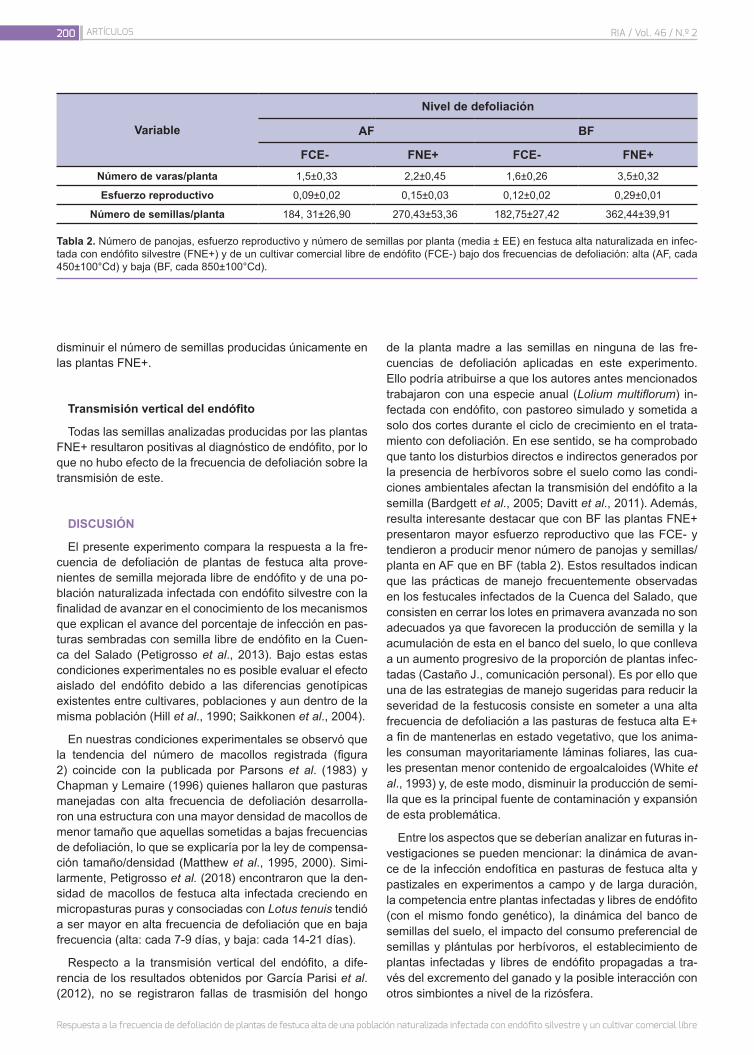
Respuesta a la frecuencia de defoliación de plantas de festuca alta de una población naturalizada infectada con endófito silvestre y un cultivar comercial libre
ARTÍCULOS RIA / Vol. 46 / N.º 2200200
disminuir el número de semillas producidas únicamente en las plantas FNE+.
Transmisión vertical del endófito
Todas las semillas analizadas producidas por las plantas FNE+ resultaron positivas al diagnóstico de endófito, por lo que no hubo efecto de la frecuencia de defoliación sobre la transmisión de este.
DISCUSIÓN
El presente experimento compara la respuesta a la fre-cuencia de defoliación de plantas de festuca alta prove-nientes de semilla mejorada libre de endófito y de una po-blación naturalizada infectada con endófito silvestre con la finalidad de avanzar en el conocimiento de los mecanismos que explican el avance del porcentaje de infección en pas-turas sembradas con semilla libre de endófito en la Cuen-ca del Salado (Petigrosso et al., 2013). Bajo estas estas condiciones experimentales no es posible evaluar el efecto aislado del endófito debido a las diferencias genotípicas existentes entre cultivares, poblaciones y aun dentro de la misma población (Hill et al., 1990; Saikkonen et al., 2004).
En nuestras condiciones experimentales se observó que la tendencia del número de macollos registrada (figura 2) coincide con la publicada por Parsons et al. (1983) y Chapman y Lemaire (1996) quienes hallaron que pasturas manejadas con alta frecuencia de defoliación desarrolla-ron una estructura con una mayor densidad de macollos de menor tamaño que aquellas sometidas a bajas frecuencias de defoliación, lo que se explicaría por la ley de compensa-ción tamaño/densidad (Matthew et al., 1995, 2000). Simi-larmente, Petigrosso et al. (2018) encontraron que la den-sidad de macollos de festuca alta infectada creciendo en micropasturas puras y consociadas con Lotus tenuis tendió a ser mayor en alta frecuencia de defoliación que en baja frecuencia (alta: cada 7-9 días, y baja: cada 14-21 días).
Respecto a la transmisión vertical del endófito, a dife-rencia de los resultados obtenidos por García Parisi et al. (2012), no se registraron fallas de trasmisión del hongo
de la planta madre a las semillas en ninguna de las fre-cuencias de defoliación aplicadas en este experimento. Ello podría atribuirse a que los autores antes mencionados trabajaron con una especie anual (Lolium multiflorum) in-fectada con endófito, con pastoreo simulado y sometida a solo dos cortes durante el ciclo de crecimiento en el trata-miento con defoliación. En ese sentido, se ha comprobado que tanto los disturbios directos e indirectos generados por la presencia de herbívoros sobre el suelo como las condi-ciones ambientales afectan la transmisión del endófito a la semilla (Bardgett et al., 2005; Davitt et al., 2011). Además, resulta interesante destacar que con BF las plantas FNE+ presentaron mayor esfuerzo reproductivo que las FCE- y tendieron a producir menor número de panojas y semillas/planta en AF que en BF (tabla 2). Estos resultados indican que las prácticas de manejo frecuentemente observadas en los festucales infectados de la Cuenca del Salado, que consisten en cerrar los lotes en primavera avanzada no son adecuados ya que favorecen la producción de semilla y la acumulación de esta en el banco del suelo, lo que conlleva a un aumento progresivo de la proporción de plantas infec-tadas (Castaño J., comunicación personal). Es por ello que una de las estrategias de manejo sugeridas para reducir la severidad de la festucosis consiste en someter a una alta frecuencia de defoliación a las pasturas de festuca alta E+ a fin de mantenerlas en estado vegetativo, que los anima-les consuman mayoritariamente láminas foliares, las cua-les presentan menor contenido de ergoalcaloides (White et al., 1993) y, de este modo, disminuir la producción de semi-lla que es la principal fuente de contaminación y expansión de esta problemática.
Entre los aspectos que se deberían analizar en futuras in-vestigaciones se pueden mencionar: la dinámica de avan-ce de la infección endofítica en pasturas de festuca alta y pastizales en experimentos a campo y de larga duración, la competencia entre plantas infectadas y libres de endófito (con el mismo fondo genético), la dinámica del banco de semillas del suelo, el impacto del consumo preferencial de semillas y plántulas por herbívoros, el establecimiento de plantas infectadas y libres de endófito propagadas a tra-vés del excremento del ganado y la posible interacción con otros simbiontes a nivel de la rizósfera.
Tabla 2. Número de panojas, esfuerzo reproductivo y número de semillas por planta (media ± EE) en festuca alta naturalizada en infec-tada con endófito silvestre (FNE+) y de un cultivar comercial libre de endófito (FCE-) bajo dos frecuencias de defoliación: alta (AF, cada 450±100°Cd) y baja (BF, cada 850±100°Cd).
Variable
Nivel de defoliación
AF BF
FCE- FNE+ FCE- FNE+Número de varas/planta 1,5±0,33 2,2±0,45 1,6±0,26 3,5±0,32
Esfuerzo reproductivo 0,09±0,02 0,15±0,03 0,12±0,02 0,29±0,01
Número de semillas/planta 184, 31±26,90 270,43±53,36 182,75±27,42 362,44±39,91

PETIGROSSO, L.R.; NAVARRO, D.; ASSUERO, S.G.; VIGNOLIO, O.R.; CASTAÑO, A.; COLABELLI, M.N.
Agosto 2020, Argentina 201201
CONCLUSIONES
En función de los resultados obtenidos podemos concluir que, bajo nuestras condiciones experimentales, la frecuen-cia de defoliación alta favoreció la producción de biomasa aérea de las plantas de festuca de la población naturaliza-da e infectada con respecto al cultivar libre de endófito, sin embargo ocurrió lo contrario en las plantas sometidas a la frecuencia de defoliación baja. Independientemente de la frecuencia de defoliación, las plantas de la población natu-ralizada e infectada tendieron a producir mayor número de panojas y semillas que las del cultivar libre de endófito, aun-que la producción de estas últimas tendió a disminuir con el aumento de la frecuencia de defoliación. No se registraron fallas en la transmisión vertical del endófito a la semilla pro-ducida por las primeras. Con base en estos resultados, sería recomendable aplicar una frecuencia de defoliación que, sin afectar la supervivencia de las plantas, sea lo suficientemen-te alta para permitir mantener la pastura en estado vegeta-tivo y disminuir la producción de semilla (principal fuente de contaminación y expansión de festuca E+).
AGRADECIMIENTOS
El presente estudio se desarrolló en el marco del Pro-yecto 15/A542, AGR540/17 de la Facultad de Ciencias Agrarias de la Universidad Nacional de Mar del Plata. Los autores agradecen a la empresa GENTOS S.A. por el su-ministro de la semilla empleada.
BIBLIOGRAFÍAAFKHAMI, M.E.; RUDGERS, J.A. 2008. Symbiosis lost: imper-
fect vertical transmission of fungal endophytes in grasses. Am. Nat. 172: 405-416.
AGNUSDEI, M.G.; MAZZANTI, A. 2001. Frequency of defolia-tion of native and naturalized species of the Flooding Pampas (Ar-gentina). Grass Forage Sci. 56:344-351.
BACON, C.W.; PORTER, J.K.; ROBBINS, J.D.; LUTTRELL, E.S. 1977. Epichloe typhina from toxic tall fescue grasses. App. Environ. Microbiol. 34:576-581.
BARDGETT, R.D.; BOWMAN, W.D.; KAUFMANN, R.; SCH-MIDT, S.K. 2005. A temporal approach to linking aboveground and belowground ecology. Trends Ecol. Evol. 20: 634-641.
BARKER, D.J.; SULC, R.M.; BULTEMEIER, T.L.; MC CORMICK, J.S.; LITTLE, R.; PENROSE, C.D.; SAMPLES, D. 2005. Contras-ting toxic-endophyte contamination between endophyte-free and nontoxic-endophyte tall fescue pastures. Crop Sci. 45: 616-625.
BOUTON, J.R.; GATES, R.; HILL, N.; HOVELAND, C. 2002. Agronomic traits with MaxQ tall fescue. Proc. Tall Fescue Toxico-sis Workshop, SERAIEG-8, Wildersvillle, TN. Missouri Forage and Grassland Council. 40-41 pp.
CHAPMAN, D.F.; LEMAIRE, G. 1996. Morphogenetic and struc-tural determinants of plant regrowth after defoliation. N.Z.J. Agric. Res. 26: 159-168.
CLAY, K.; SCHARDL, C. 2002. Evolutionary origins and ecolog-ical consequences of endophyte symbiosis with grasses. Am. Nat. 160: 99-127.
CLAY, K.; HOLAH, J.; RUDGERS, J.R. 2005. Herbivores cause a rapid increase in hereditary symbiosis and alter plant community composition. PNAS 102 (35): 12465-12470.
COLABELLI, M.; AGNUSDEI, M.; MAZZANTI, A.; LABREVEUX, M. 1998. El proceso de crecimiento y desarrollo de gramíneas for-rajeras como base para el manejo de la defoliación. Boletín Técni-co N.° 148. INTA EEA Balcarce. Buenos Aires, Argentina. 1-14 pp.
COLABELLI, M.N.; SALOMONE, L.; FERNÁNDEZ, F.; SAN MARTINO, S. 2006. Niveles de infección de Neotyphodium coeno-phialum en poblaciones de festuca naturalizadas en el sudeste bo-naerense. 29.º Congreso AAPA. Mar del Plata. 239-240 pp.
DAVITT, A.J.; CHEN, C.; RUDGERS, J.A. 2011. Understanding context-dependency in plant-microbe symbiosis: The influence of abiotic and biotic contexts on host fitness and the rate of symbiont transmission. Environ. Exp. Bot. 71: 137-45.
DE BATTISTA, J.; PERETTI, A.; CARLETTI, S.; RAMIREZ, A.; COSTA, M.; SCHULTZ, L. 1995. Evolución de la incidencia de la infección de Acremonium coenophialum en la oferta de semilla de festuca alta en Argentina. Período 1987-1994. Rev. Arg. Prod. Anim. 15: 300-302.
DE BATTISTA, J.; ALTIER, N.; GALDAMES, D.R.; DALL’AG-NOL, M. 1997. Significance of endophyte toxicosis and current practices in dealing with the problema in South America. En: BA-CON, C.W.; HILL, N.S. (eds.). Neotyphodium/Grass Interactions. Plenum Press: Nueva York. 383-388 pp.
DEFELICE, M.S.; HENNING, J.C. 1990. Renovation of endo-phyte (Acremonium coenophialum) infected tall fescue (Festuca arundinacea) pastures with herbicides. Weed Sci. 38: 628-633.
FRIBOURG, H.A.; HANNAWAY, D.B. 2007. Tall fescue. on-line monograph, forage information system. (Disponible: http://forages.oregonstate. edu/is/tfis/ monograph.html verificado: 08 de marzo de 2013).
GARCÍA PARISI, P.A.; CASAS, C.; GUNDEL, P.E.; OMACINI, M. 2012. Consequences of grazing on the vertical transmission of a fungal Neotyphodium symbiont in an annual grass population. Austral Ecol. 37:620-628.
GLENN, A.E.; BACON, C.W.; PRICE, R.; HANDIL, R.T. 1996. Molecular phylogeny of Acremonium and its taxonomic implica-tions. Mycología 88: 369-383.
GUNDEL, P.E.; PÉREZ, L.I.; HELANDER, M.; SAIKKONEN, K. 2013. Symbiotically modified organisms: nontoxic fungal endo-phytes in grasses. Trends Plant Sci. 18: 420-427.
GUNDEL, P.E.; RUDGERS, J.A.; GHERSA, C.M. 2011. Incor-porating the process of vertical transmission into understanding of host symbiont dynamics. Oikos. 120: 1121-1128.
GUNDEL, P.E.; GARIBALDI, L.A.; TOGNETTI, P.M.; ARAGÓN, R.; GHERSA, C.M.; OMACINI, M. 2009. Imperfect vertical trans-mission of the endophyte Neotyphodium in exotic grasses in grass-lands of the Flooding Pampa. Microbial. Ecol. 57: 740-748.
GUNDEL, P.E.; BATISTA, W.H.; TEXEIRA M.; MARTINEZ-GHERSA, M.A.; OMACINI, M.; GHERSA, C.M. 2008. Neotypho-dium endophyte infection frequency in annual grass populations: relative importance of mutualism and transmission efficiency. Pro-ceedings of the Royal Society of London. 275: 897-905.
HANNAWAY, D.; FRANSEN, S.; CROPPER, J.; TEEL, M.; CHANEY. M.; GRIGGS, T.; HALSE, R.; HART, J.; CHEEKE, P.; HANSEN, D.; KLINGER, R.; LANE, W. 1999. Tall fescue (Festu-ca arundinacea Schreb.). Oregon State University: Oregon. EUA. PNW 504: 1-20.
HILL, N.S.; STRINGER, W.C.; ROTTINGHAUS, G.E.; BE-LESKY, D.P.; PARTOT, W.A.; POPE, D.D. 1990. Growth, morpho-logical and chemical component responses of tall fescue to Acre-monium coenophialum. Crop Sci. 30: 156-161.
HILL, N.S.; ANDRAE, J.G.; DURHAM, R.G; HANCOCK, D.W. 2010. Herbicide Treatments to Renovate Toxic Endophyte Infec-

Respuesta a la frecuencia de defoliación de plantas de festuca alta de una población naturalizada infectada con endófito silvestre y un cultivar comercial libre
ARTÍCULOS RIA / Vol. 46 / N.º 2202202
ted Tall Fescue Pastures with ‘Jesup’ MaxQ. Crop Sci. 50 (3): 1086-1089.
HOVELAND, C.S. 1993. Economic importance of Acremonium endophytes. Agr. Ecosyst. Environ. 44: 3-12.
LEUCHTMANN, A.; BACON, C.W.; SCHARDL, C.L.; WHITE, J.F.; TADYCH, M. 2014. Nomenclatural realignment of Neotypho-dium species with genus Epichloë. Mycol. 106: 202-215.
LUGTENBERG, B.J.J.; CARADUS, J.R.; JOHNSON, L.J. 2016. Fungal endophytes for sustainable crop production. Microbiol. Ecol. 92 (12):1-17.
MALINOWSKI, D.; BELESKY, D. 2000. Adaptations of endo-phyte-infected cool-season grasses to environmental stresses: mechanisms of drought and mineral stress tolerance. Crop Sci. 40: 923-940.
MATTHEW, C.; ASSUERO, S.G.; BLACK, C.K.; SACKVILLE HAMILTON, N.R. 2000. Tiller dynamics of grazed swards. En: LEMAIRE, G.; HODGSON, J.; DE MORAES, A.; NABINGER, C.; CARVALHO, P.C. DE F. (eds.). Grassland ecofisiology and grazing ecology. CABI Publishing, University Press, Cambridge, Reino Unido. 127-151 pp.
MATTHEW, C.; LEMAIRE, G.; SACKVILLE HAMILTON, R.N. 1995. A modified self-thinning equation to describe size density re-lationships for defoliation swards. Ann. Bot. 76: 579-587.
ODRIOZOLA, E.; IRAGUEN PAGATE, I.; LLOBERAS, M.M.; COSENTINO, I; PORTEY, R.; OROMÍ, J. 2002. Festuca tóxica. Su efecto en diferentes razas bovinas. Rev. Vet. Arg. 19:12-21.
PARSONS, A.J.; LEAFE, E.L.; COLLET, B.; STILES, W. 1983. The physiology of grass production under grazing. i. Characte-ristics of lead and canopy photosynthesis of continuously-grazed swards. J. Appl. Ecol. 20: 117-126.
PETIGROSSO, L.R.; COLABELLI, M.N.; FERNÁNDEZ, O.N.; ISPIZÚA, V., CENDOYA, M.G. 2013. Incidence of the endophy-te fungus Neotyphodium coenophialum in pastures of tall fescue differing in age and soil characteristics. Afr. J. Agric. Res. 8 (22): 2655-2662.
PETIGROSSO, L.R.; ASSUERO, S.G.; VIGNOLIO, O.R.; RO-MANO, Y.; COLABELLI, M.N., SALVAT, A.; CRISTOS, D. 2018. Interacción entre festuca alta infectada con endófito y Lotus tenuis bajo dos frecuencias de defoliación. RIA. 44(1):41-48.
PINHEIRO, J.; BATES, D.; DEBROY, S.; SARKAR, D. 2012. R Development Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-103.
R DEVELOPMENT CORE TEAM. 2016. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. (Disponible: http://www.R-project.org/ verificado: 15 de junio de 2016).
SAHA, C.D.; JACKSON, M.A.; JOHNSON-CICALESE, J.M. 1988. A rapid staining method for detection of endophytic fungi in turf and forage grass. Phytopathol. 78: 237-239.
SAIKKONEN, K.; FAETH, S.H.; HELANDER, M.; SULLIVAN, T.J. 1998. Fungal Endophytes: A Continuum of Interactions with Host Plants. Annu. Rev. Ecol. Syst. 29:319-343.
SAIKKONEN, K.; WALI, P.; HELANDER, M.; FAETH, S.H. 2004. Evolution of endophyte-plant symbioses. Trends Plant Sci. 9:275-280.
SAIKKONEN, K.; LEHTONEN, P.; HELANDER, M.; KORICHE-VA, J.; FAETH, S.H. 2006. Model systems in ecology: Dissecting the endophyte-grass literature. Trends Plant Sci. 11: 428-433.
SCHENEITER, J.O.; KAUFMANN, I.I.; FERREYRA, A.R.; LLO-RENTE, R.T. 2015. The herbage productivity of tall fescue in the Pampas region of Argentina is correlated to its ecological niche. Grass Forage Sci. 71: 403-412.
SHELBY, R.A.; SCHMIDT, S.P.; RUSSELL, R.W.; GREGORY, W.H. 1989. Spread of tall fescue endophyte by cattle. Leaflet 104, Alabama Agric, Exp. Stn.
SHELBY, R.A.; SCHMIDT, S.P. 1991. Survival of the tall fescue endophyte in the digestive tract of cattle and horses. Plant Dis. 75: 776-778.
SHELBY, R.A.; DALRYMPLE, L.W. 1993. Long-term changes of endophyte infection in tall fescue stands. Grass Forage Sci. 48: 356-361.
TEKELI, A.S.; ATES, E. 2005. Yield potential and mineral com-position of white clover (Trifolium repens L.) – tall fescue (Festuca arundinacea Schreb.) mixtures. J. Cent. Eur. Agr. 6: 27-34.
TREVOR, D.I.; BATES, G.E.; MUELLER,C.T.; WALLER, J.C.; RHODES, G.N.JR. 2016. Effects of Aminocyclopyrachlor Plus Met-sulfuron on Tall Fescue Yield, Forage Quality, and Ergot Alkaloid Concentration. Weed Technol. 30(1):171-180.
WHITE, J.F.JR.; MORGAN-JONES, G.; MORROW, A.C. 1993. Taxonomy, life cicle, reproduction and detection of Acremonium endophytes. Agr. Ecosys. Environ. 44: 13-37.
WHITE, J.F.JR.; TORRES, M. 2009. Defensive mutualism in mi-crobial symbiosis. CRC Press: Boca Raton, FL. 333-334 pp.

MEDINA, A.A.; MANZIONE, P.; BAUCIS, A.G.; CATALÁN, M.A.; LAFFITTE, L.; ANDÍA, I.R.
Agosto 2020, Argentina 203203
Propiedades mecánicas de la madera de álamos de cortinas forestales de Río Negro, Patagonia, Argentina
RESUMENEn la provincia de Río Negro, Argentina, existen en la actualidad 6000 km de longitud lineal de cortinas
cortavientos de álamo. Su función principal es la protección de cultivos frutihortícolas, pero son además un recurso forestal real y potencial. El objetivo de este trabajo es conocer algunas propiedades mecánicas de la madera estos álamos, información no existente y esencial para optimizar su aprovechamiento y maximizar su valor en diferentes usos. Se apearon 15 ejemplares de los álamos más representados en cortinas forestales (álamo chileno, conti 12 y boleana) de chacras de la localidad de Allen. De cada árbol se extrajo una troza de 1,5 m de largo desde la cual se obtuvieron probetas para ensayos de propiedades mecánicas de resistencia estática de flexión, compresión (Norma ASTM D-143), resistencia al aplastamiento (Norma ASTM D-5764-97) y dureza (Norma IRAM-9570). Por un lado, los resultados obtenidos fueron del orden de los reportados para madera de otros álamos cultivados en diferentes zonas de nuestro país, algunas de ellas habilitadas para uso estructural. Por otro lado, los valores obtenidos fueron del rango de los reportados para pino ponderosa, madera ampliamente utilizada en Patagonia en la construcción de cabreadas, estructuras portantes (tirantes, vigas, columnas, etc.) y bastidores de entramados en paneles estructurales. La madera de boleana y conti 12 presentó mayores valores de resistencia a la flexión, a la compresión y al desgaste que la del álamo chi-leno (MOR flexión: 66,7 Mpa, MOE flexión: 8562,15 Mpa, MOR compresión: 31,22 Mpa, MOE compresión: 8270,52 Mpa, dureza radial: 1,63 kN/cm2). La resistencia al aplastamiento no presentó variaciones significa-tivas entre ellos (conti 12: 29,42 Mpa, boleana: 30,08 Mpa y chileno: 29,65 Mpa). Estos resultados resaltan el potencial que presenta la madera de los álamos de cortinas forestales de los valles irrigados de Río Negro para ampliar sus posibilidades de uso.
Palabras clave: tecnología de la madera, cortinas protectoras, populicultura.
ABSTRACT
In Río Negro province, Argentina, there are currently 6000 km of linear length of poplar windbreaks. Its main function is the protection of fruit and vegetable crops but they are also a real and potential forest resource. The objective of this work is to know the mechanical properties of the wood these poplars, nonexistent and essential information to optimize their use and maximize their value in different uses. Fifteen specimens of the most represented poplars in forest curtains (chileno, conti 12 and boleana) of farms in Allen were cutted. From
1Universidad Nacional del Comahue, Asentamiento Universitario San Martín de los Andes, Pasaje de la Paz 235, (8370), San Martín de los Andes, Neuquén, Argentina. Correo electrónico: [email protected] Nacional del Comahue, Facultad de Ingeniería, Departamento de Mecánica Aplicada, Grupo de Estudio de Polímeros Sintéti-cos y Naturales (GEPSyN), Buenos Aires 1400 (8300) Neuquén.
MEDINA, A.A.1; MANZIONE, P.2; BAUCIS, A.G.1; CATALÁN, M.A.1; LAFFITTE, L.1; ANDÍA, I.R.1
Recibido 21 junio de 2018 // Aceptado 25 de marzo de 2019 // Publicado online 05 de agosto de 2020

Propiedades mecánicas de la madera de álamos de cortinas forestales de Río Negro, Patagonia, Argentina
ARTÍCULOS RIA / Vol. 46 / N.º 2204204
each tree a 1.5 m long log was extracted to obtain wood for mechanical properties tests of static resistance of bending, compression (ASTM D-143), dowel bearing strenght (ASTM D-5764-97) and hardness (IRAM-9570). The results obtained were of the order of those reported for wood from other poplars grown in different areas of our country, some of them accepted for structural use. On the other hand, the values obtained were from the range of those reported for ponderosa pine, wood widely used in Patagonia in the construction, load-bearing structures (beams, columns, etc.) and frames of trusses in structural panels. The wood of boleana and conti 12 presented higher values of resistance to flexion, compression and hardness than chilean poplar (MOR flexion: 66.7 Mpa, MOE flexion: 8562.15 Mpa, MOR compression: 31.22 Mpa, MOE compression: 8270.52 MPa, radial hardness: 1.63 kN / cm2). The dowel bearing strenght did not show significant variations among them (conti 12: 29.42 MPa, boleana: 30.08 MPa and chilean: 29.65 MPa). These conclusions highlight the potential of poplar wood from Río Negro forest windbreaks to expand their possibilities of use.
Keywords: wood technology, forest curtains, poplar cultivation.
INTRODUCCIÓN
La plantación de álamos en los valles irrigados del norte de la Patagonia comenzó a principios del siglo pasado con el objetivo de establecer cortinas protectoras de cultivos frutihortícolas. Tempranamente se implantaron Populus ni-gra L. cv. italica “álamo criollo” y P. nigra cv. thaisiana “ála-mo chileno”, este último caracterizado por tener un periodo fenológico extendido, brindando protección prolongada a los cultivos. Posteriormente se fueron plantando otros cul-tivares de P. nigra y del híbrido Euroamericano Populus x canadensis (P. deltoides x P. nigra); de este último el clon ´conti 12´ tuvo preferencia por su forma disciplinada y gran rusticidad. Otra especie ampliamente implantada fue Po-pulus alba L. var. bolleana (Lauche) Otto, conocida en la región como “boleana” (García, 2011; Serventi, 2011).
En la provincia de Río Negro existen en la actualidad 6000 km de longitud lineal de cortinas y unas 900 ha de plantaciones en macizo de álamo. El destino de la madera de estas cortinas ha sido tradicionalmente la fabricación de envases y embalajes frutihortícolas, pero también se utiliza con otros fines. En el año 2011 se consumieron 68565 m3 de madera de álamo en los aserraderos del Alto Valle de Río Negro, alcanzando una producción de 36177 m3, prin-cipalmente de cajones, madera aserrada, pallets y bins, tanto con destino local como regional e internacional (CFI, 2012). El empleo de una tecnología inapropiada ha gene-rado la idea de que la madera de álamo es de baja calidad, sin embargo algunos ejemplos de fabricación de muebles, aberturas y construcción de viviendas prefabricadas con un manejo adecuado de esta muestran excelentes resultados (García, 2011).
El conocimiento de las propiedades mecánicas de estas maderas es esencial para optimizar su aprovechamiento y maximizar su valor en diferentes usos ya que estas definen su comportamiento frente a procesos de transformación. Si bien existe información de propiedades mecánicas de la madera de álamos cultivados en Argentina (Cortizo et al.,
2014; Calderón et al., 2006; Membrivez et al., 2007; Mana-vella et al., 2014; Cortizo y Monteverde, 2017; Filippetti et al., 2017), esta es nula de los álamos de cortinas forestales de la región Norpatagónica.
Como lo indican numerosas referencias bibliográficas (Beaudoin et al., 1992; Zhang et al., 2003; Monteoliva y Senisterra, 2008; Yu et al., 2008; Huda et al., 2014) la ma-dera de los álamos puede variar significativamente entre sitios y entre especies, variedades y clones, por lo que es necesario conocer sus propiedades para cada especie o entidad genética en cada zona de cultivo.
El objetivo de este trabajo es conocer las propiedades mecánicas de la madera de tres de los álamos más cultiva-dos en valles irrigados de Río Negro (álamo chileno, conti 12 y boleana).
Estos resultados serán de utilidad para analizar la posi-ble ampliación del abanico de uso de estas maderas. En futuros escenarios de desarrollo forestal de la región tam-bién podrán ser importantes en la elección de especies y el desarrollo de programas de mejoramiento forestal.
MATERIALES Y MÉTODOS
Se apearon 5 ejemplares de cada especie (tabla1) de buen estado sanitario de cortinas protectoras de chacras de la localidad de Allen, provincia de Río Negro (39° 0’ 17.06” S, 67° 50’ 41.14” O; 240 m s. n. m.). Se eligieron estas especies por su representatividad en las cortinas de la zona (SSDFI, 2017).
El clima de la región es árido frío, con temperatura media anual 15,5 °C y precipitación media anual 243,7 mm. Los suelos son xerofluventes típicos, con más de 100 cm de profundidad, de muy escasa evolución y con un horizonte A bajo en materia orgánica. En profundidad se superponen distintas capas de acumulación fluvial de antiguos planos aluviales (Bran, 2014).
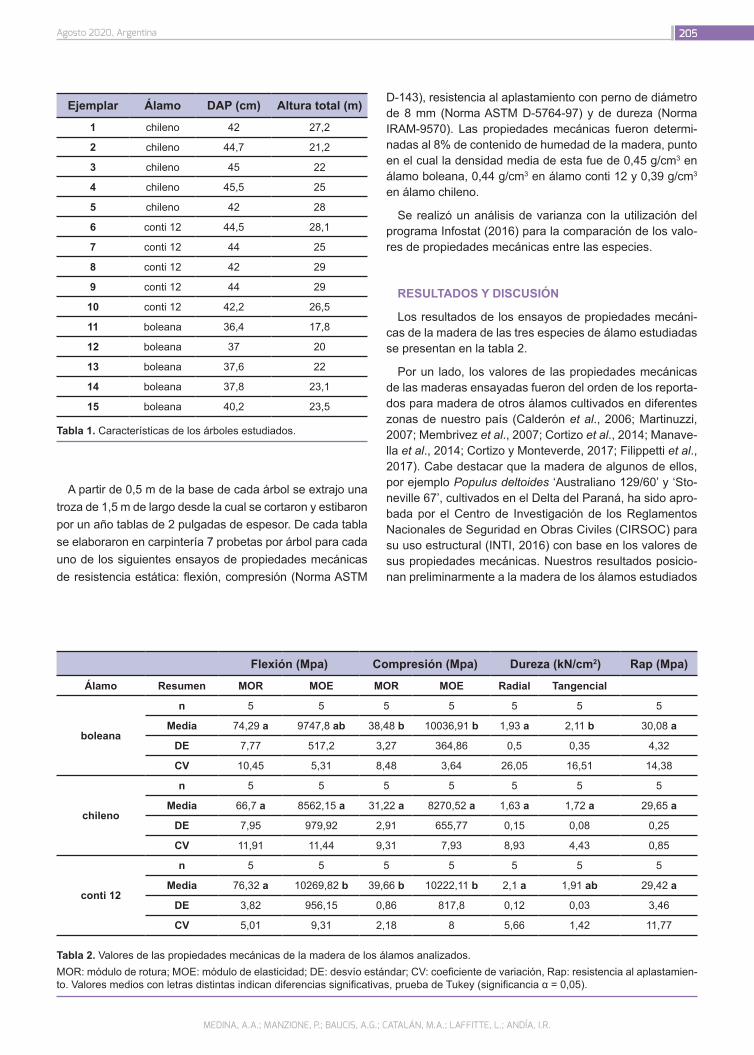
MEDINA, A.A.; MANZIONE, P.; BAUCIS, A.G.; CATALÁN, M.A.; LAFFITTE, L.; ANDÍA, I.R.
Agosto 2020, Argentina 205205
D-143), resistencia al aplastamiento con perno de diámetro de 8 mm (Norma ASTM D-5764-97) y de dureza (Norma IRAM-9570). Las propiedades mecánicas fueron determi-nadas al 8% de contenido de humedad de la madera, punto en el cual la densidad media de esta fue de 0,45 g/cm3 en álamo boleana, 0,44 g/cm3 en álamo conti 12 y 0,39 g/cm3 en álamo chileno.
Se realizó un análisis de varianza con la utilización del programa Infostat (2016) para la comparación de los valo-res de propiedades mecánicas entre las especies.
RESULTADOS Y DISCUSIÓN
Los resultados de los ensayos de propiedades mecáni-cas de la madera de las tres especies de álamo estudiadas se presentan en la tabla 2.
Por un lado, los valores de las propiedades mecánicas de las maderas ensayadas fueron del orden de los reporta-dos para madera de otros álamos cultivados en diferentes zonas de nuestro país (Calderón et al., 2006; Martinuzzi, 2007; Membrivez et al., 2007; Cortizo et al., 2014; Manave-lla et al., 2014; Cortizo y Monteverde, 2017; Filippetti et al., 2017). Cabe destacar que la madera de algunos de ellos, por ejemplo Populus deltoides ‘Australiano 129/60’ y ‘Sto-neville 67’, cultivados en el Delta del Paraná, ha sido apro-bada por el Centro de Investigación de los Reglamentos Nacionales de Seguridad en Obras Civiles (CIRSOC) para su uso estructural (INTI, 2016) con base en los valores de sus propiedades mecánicas. Nuestros resultados posicio-nan preliminarmente a la madera de los álamos estudiados
Tabla 1. Características de los árboles estudiados.
Tabla 2. Valores de las propiedades mecánicas de la madera de los álamos analizados.MOR: módulo de rotura; MOE: módulo de elasticidad; DE: desvío estándar; CV: coeficiente de variación, Rap: resistencia al aplastamien-to. Valores medios con letras distintas indican diferencias significativas, prueba de Tukey (significancia α = 0,05).
Ejemplar Álamo DAP (cm) Altura total (m)1 chileno 42 27,2
2 chileno 44,7 21,2
3 chileno 45 22
4 chileno 45,5 25
5 chileno 42 28
6 conti 12 44,5 28,1
7 conti 12 44 25
8 conti 12 42 29
9 conti 12 44 29
10 conti 12 42,2 26,5
11 boleana 36,4 17,8
12 boleana 37 20
13 boleana 37,6 22
14 boleana 37,8 23,1
15 boleana 40,2 23,5
Flexión (Mpa) Compresión (Mpa) Dureza (kN/cm2) Rap (Mpa)Álamo Resumen MOR MOE MOR MOE Radial Tangencial
boleana
n 5 5 5 5 5 5 5
Media 74,29 a 9747,8 ab 38,48 b 10036,91 b 1,93 a 2,11 b 30,08 a
DE 7,77 517,2 3,27 364,86 0,5 0,35 4,32
CV 10,45 5,31 8,48 3,64 26,05 16,51 14,38
chileno
n 5 5 5 5 5 5 5
Media 66,7 a 8562,15 a 31,22 a 8270,52 a 1,63 a 1,72 a 29,65 a
DE 7,95 979,92 2,91 655,77 0,15 0,08 0,25
CV 11,91 11,44 9,31 7,93 8,93 4,43 0,85
conti 12
n 5 5 5 5 5 5 5
Media 76,32 a 10269,82 b 39,66 b 10222,11 b 2,1 a 1,91 ab 29,42 a
DE 3,82 956,15 0,86 817,8 0,12 0,03 3,46
CV 5,01 9,31 2,18 8 5,66 1,42 11,77
A partir de 0,5 m de la base de cada árbol se extrajo una troza de 1,5 m de largo desde la cual se cortaron y estibaron por un año tablas de 2 pulgadas de espesor. De cada tabla se elaboraron en carpintería 7 probetas por árbol para cada uno de los siguientes ensayos de propiedades mecánicas de resistencia estática: flexión, compresión (Norma ASTM

Propiedades mecánicas de la madera de álamos de cortinas forestales de Río Negro, Patagonia, Argentina
ARTÍCULOS RIA / Vol. 46 / N.º 2206206
en posibilidad de competencia con las de otros álamos cul-tivados en diferentes regiones de Argentina.
Por otro lado, los valores de las propiedades estudiadas fueron del rango de los reportados para Pinus ponderosa cultivado en Patagonia (Martinuzzi, 2007; Letourneau et al., 2014), madera ampliamente utilizada en la región con fines estructurales en la construcción de cabreadas, estructuras portantes (tirantes, vigas, columnas, etc.) y bastidores de entramados en paneles estructurales.
La comparación estadística entre los diferentes álamos mostró que la madera de los álamos boleana y conti 12 pre-sentó mayores valores de resistencia a la flexión, a la com-presión y al desgaste (dureza) que el álamo chileno. La resis-tencia al aplastamiento no presentó variaciones significativas entre ellos para el diámetro de bulón ensayado (tabla 2).
CONCLUSIONES
Los resultados obtenidos en el presente trabajo indican la posibilidad de ampliar el abanico de usos de la madera de álamos de cortinas forestales implantadas en los valles irri-gados de Río Negro, utilizada tradicionalmente para la fabri-cación de envases y embalajes frutihortícolas. En este sen-tido, alientan a la realización de nuevos estudios orientados a consolidar su uso en estructuras portantes de distinto tipo.
A su vez, la incorporación de las construcciones con ma-dera al sistema tradicional (INTI, 2016) señala a estos ár-boles como un recurso forestal potencial y a la populicultu-ra como una interesante actividad para tener en cuenta en proyectos de diversificación de la matriz productiva regional.
Por un lado, sería de gran utilidad proseguir con estos estudios en mayor número de ejemplares, en otras espe-cies de álamo y en distintos sitios de cultivo con el fin de conocer características tan importantes respecto a calidad de la madera como la variabilidad de sus propiedades.
Por otro lado, y debido a que el CIRSOC (INTI, 2016) se basa en la actualidad en la técnica de determinación de pro-piedades mecánicas en tablas de madera a escala de utili-zación para su aprobación para usos estructurales, vemos la apremiante necesidad de avanzar en dichos ensayos.
AGRADECIMIENTOS
Agradecemos el apoyo del técnico forestal Nicolás Man-tilaro en la elaboración de probetas para ensayos de pro-piedades mecánicas. La colaboración del técnico regional de la Subsecretaría de Desarrollo Foresto-Industrial del Ministerio de Agroindustria de la Nación Julio García, al consultor privado e ingeniero agrónomo Norberto Serventi y al presidente de la Cámara de Forestadores y Empresa-rios Madereros y Afines de Norpatagonia Nazareno Olivet-ti, en la elección de las cortinas de álamo y los contactos para la colección del material y su procesado inicial. A la estudiante Evelyn Riquelme por su colaboración en ensa-yos de laboratorio.
BIBLIOGRAFÍAASTM D 143-94. 2000. Standard Methods of Testing Small
Clear Specimens of Timber. Annual Book of ASTM Standards. Section 4, Construction. Volume 04.09 Wood. Philadelphia, EUA.
ASTM D 5764-97a. 2002. Standard Test Methods for Evaluating Dowel Bearing-Strength of Wood and Wood-Based Products.
BEAUDOIN, M.; HERNÁNDEZ, R.; KOUBAA, A.; POLIQUIN, J. 1992. Interclonal, intraclonal and within-tree variation in wood density of poplar hybrid clones. Wood Fiber Science 24 (2): 147-153.
BRAN, D.E. 2014. Inventario Integrado de los Recursos Natu-rales de la Provincia de Río Negro. INTA, Provincia de Río Negro. Ediciones INTA.
CALDERÓN, A.D.; ROIG, F.A.; ZANETTI, R.P.; AGUADO, E.G.; FURLANI, A.M.; ARTINEZ, F.G. 2006. Características físico mecá-nicas de maderas de clones de álamo ensayadas en la provincia de Mendoza, República Argentina–Parte ii. iii Congreso Internacio-nal de Salicáceas en Argentina, Buenos Aires.
CFI (CONSEJO FEDERAL DE INVERSIONES). 2012. Censo de Industrias de la Madera de la Provincia del Neuquén y zonas de influencia del Alto Valle del Río Negro. Neuquén. 69 p.
CORTIZO, S.; MONTEVERDE, S.; FERNÁNDEZ, E.; REFORT, M.; TARABORELLI, C.; KEIL, G.; ABBIATI, N. 2014. Características técnicas de un nuevo genotipo de álamo para su inscripción en el Registro Nacional de Variedades. iv Congreso Internacional de Sali-cáceas en la Argentina. La Plata, Buenos Aires, Argentina.
CORTIZO, S.; MONTEVERDE, M.S. 2017. Seis nuevos clo-nes de Álamo, recientemente incorporados al Registro Nacional de Cultivares de Argentina, mejoran la oferta clonal de la cadena foresto industrial de las Salicáceas. Jornadas de Salicáceas 2017. v Congreso Internacional de Salicáceas en Talca, Chile, 9 p.
DI RIENZO, J.A.; CASANOVES, F.; BALZARINI, M.G.; GONZA-LEZ, L.; TABLADA, M.; ROBLEDO, C.W. 2016. INFOSTAT, Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. (Disponi-ble: http://www.infostat.com.ar verificado: 02 de junio de 2018).
FILIPPETTI, M.C.; GUILLAUMET, A.A.; TORTORIELLO, M.; MOSSRE, C.A. 2017. Potencial uso estructural de la madera de Populus deltoides cultivado en el noroeste de la provincia de bue-nos Aires. i Congreso Latinoamericano de Estructuras de Madera. ii Congreso Ibero-Latinoamericano de la Madera en la Construc-ción. Junín, Argentina.
GARCÍA, J. 2011. El cultivo de las Salicáceas en los valles irri-gados de la Patagonia, pasado, presente y futuro. 3.er Congreso Internacional de Salicáceas en Argentina. Neuquén, Argentina.
HUDA, A.; KOUBAA, A.; CLOUTIER, A.; HERNÁNDEZ, R.; FORTIN, Y. 2014. Variation of physical and mechanical properties of hybrid poplar clones. Bioresources 9 (1): 1456-1471.
INTI (INSTITUTO NACIONAL DE TECNOLOGÍA INDUS-TRIAL). 2016. Centro de Investigación de los Reglamentos Na-cionales de Seguridad en Obras Civiles (CIRSOC). Reglamento Argentino de Estructuras de Madera (601). Buenos Aires.
IRAM (INSTITUTO ARGENTINO DE NORMALIZACIÓN Y CER-TIFICACIÓN) 9570. 1971. Método de ensayo de la dureza Janka. Instituto Argentino de Racionalización de Materiales. 6 p.
LETOURNEAU, F.J.; MEDINA, A.A.; ANDIA, I.R.; ANDEN-MATTEN, E.; DE AGOSTINI, N.; MANTILARO, N. 2014. Carac-terización xilo-tecnológica de la madera de una plantación adulta de Pinus ponderosa de la Patagonia Argentina. Revista de investi-gaciones agropecuarias, 40 (2), 196-201.
MANAVELLA, R.D.; GUILLAUMET, A.A.; FILIPPETTI, M. C.; TORTORIELLO, M. 2014. Determinación de la Resistencia al Aplastamiento en Madera de Álamo para Uniones Tipo Clavija.

MEDINA, A.A.; MANZIONE, P.; BAUCIS, A.G.; CATALÁN, M.A.; LAFFITTE, L.; ANDÍA, I.R.
Agosto 2020, Argentina 207207
xxxvi Jornadas Sudamericanas de Ingeniería Estructural. Mon-tevideo. Uruguay.
MARTINUZZI, F. 2007. Fichas Técnicas de Maderas. Buenos Aires, Argentina: Instituto Nacional de Tecnología Industrial, Madera y Muebles. (Disponible: http://www.inti.gob.ar/maderaymuebles ve-rificado: 20 de junio de 2018).
MEMBRIVEZ, F.; GENOVESE, F.; REVIGLIO, H.; ARCIDÍA-CONO, R.; BONINO, G.; DI MENZA, A; LUCERO, C.; MARTÍ-NEZ ARAUJO, N.; NAVARRO, N.; NIETO, A.; OBERTI, N. 2007. Caracterización físico mecánica de la madera de álamo del sur de Mendoza. Iberomadera, Buenos Aires, Argentina. 21 p.
MONTEOLIVA, S.; SENISTERRA, G. 2008. Efecto del sitio, el origen y el clon sobre el crecimiento y propiedades de la madera de Populus. Investigación Agraria: Sistemas y Recursos Foresta-les, 17 (3): 261-270.
SERVENTI, N. 2011. Las cortinas forestales en los valles irriga-dos de Norpatagonia. 3.er Congreso Internacional de Salicáceas en Argentina. Neuquén, Argentina.
SSDFI (SUBSECRETARÍA DE DESARROLLO FORESTO INDUSTRIAL). 2017. Inventario Nacional de Plantaciones Fores-tales bajo riego. Región Patagónica. Ministerio de Agroindustrias, Presidencia de la Nación.
YU, Q.; ZHANG, S.Y.; PLIURA, A.; MACKAY, J.; BOUS-QUET, J.; PERINET, P. 2008. Variation in Mechanical Properties of Selected Young Poplar Hybrid Crosses. Forest Science 54 (3) 255-259.
ZHANG, S.Y.; YU, Q.; CHAURET, G.; KOUBAA, A. 2003. Se-lection for Both Growth and Wood Properties in Hybrid Poplar Clo-nes. Forest Science, 49 (6): 901-908

Seed priming induces biochemical changes in melon plants and increase salt tolerance
ARTÍCULOS RIA / Vol. 46 / N.º 2208208
Seed priming induces biochemical changes in melon plants and increase salt tolerance
ABSTRACTAn experiment was conducted to evaluate the effect of priming on germination and initial growth of melon
plants under salinity conditions. Two osmotic agents (NaCl and CaCl2) and two treatment durations (2 and 4 days) were compared. Germination percentage and growth of seedlings were evaluated at electrical conducti-vity 8.0 dS m-1. Total chlorophyll content, relative water content, root viability, proline content, relative electroly-te leakage, malondialdehyde concentration, peroxidase and catalase activity, Na+/K+ ratio in green parts, and K+ leakage from roots were measured. The best germination was obtained when seeds were soaked either with NaCl or CaCl2 for two days. Priming improved growth parameters and plants presented a better response in all the evaluated biochemical parameters. Results suggest that priming could be used to improve the perfor-mance of seeds and seedlings in situations of high salinity.
Keywords: Cucumis melo, osmo priming, germination, salt stress, oxidative stress.
RESUMEN
Se realizó un experimento para evaluar el efecto del ´priming´ de semillas sobre la germinación y creci-miento inicial de plantas de melón en salinidad. Se compararon dos agentes osmóticos (NaCl y CaCl2) y dos duraciones de tratamiento (2 y 4 días). Se evaluó el porcentaje de germinación y crecimiento de las plántulas a una conductividad eléctrica de 8.0 dS m-1. Se midió el contenido total de clorofila, contenido relativo de agua, viabilidad de raíces, contenido de prolina, pérdida relativa de electrolitos, concentración de malondialdehído, actividad peroxidasa y la catalasa, proporción de Na+/K+ en partes verdes y pérdida de K+ por raíces. La mejor germinación se obtuvo cuando las semillas se trataron con NaCl o CaCl2 durante dos días. El ́ priming´ mejoró los parámetros de crecimiento y las plantas mostraron una mejor respuesta en todos los parámetros bioquí-micos evaluados. Los resultados sugieren que el ´priming´ podría usarse para mejorar el rendimiento de las semillas y las plántulas en situaciones de alta salinidad.
Palabras clave: Cucumis melo, osmo priming, germinación, estrés salino, estrés oxidativo.
1Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Área Metropolitana Buenos Aires (AMBA). Udaondo 1695, Ituzaingó, Buenos Aires, Argentina. Correo electrónico: [email protected] Nacional de Luján (UNLu), Departamento de Ciencias Básicas, Laboratorio de Fisiología Vegetal. Ruta 5 y Avenida Constitución, Luján, Buenos Aires, Argentina. 3Universidad Nacional del Litoral, Facultad de Ciencias Agrarias, Laboratorio de Biología Molecular y Fisiología Vegetal. Kreder 2805, Esperan-za, Santa Fe, Argentina.
CASTAÑARES, J.L.1,2; BOUZO, C.A.3
Received July 04th 2019 // Accepted December 13th 2019// Published online September 30th 2020

CASTAÑARES, J.L.; BOUZO, C.A.
Agosto 2020, Argentina 209209
INTRODUCTION
Soil salinity is one of the most important stresses limi-ting crop production. This phenomenon affects 20% of cul-tivated lands of the world and causes important yield los-ses in crops (Qadir et al., 2014). Irrigation with poor quality water is the main cause of increased soil salinity in culti-vated land (Yadav et al., 2011). Although irrigated crops represent only 15% of total cultivated land, this represents one third of the world’s food production given that yields double what is produced in non-irrigated crops (Munns, 2005). For this reason, it is unlikely that crop irrigation will be abandoned as a normal practice and, instead, an in-crease in its application could be expected (Shabala and Munns, 2012).
Melon (Cucumis melo L.) is an important horticultural crop with a worldwide production of 27.3 million tons. Chi-na, Iran, Turkey, Egypt, and the United States represent 68% of the world´s production (Iqbal et al., 2016). It is culti-vated in all regions of the world with warm and slightly rainy weather, where salinity is becoming a problem (Yasar et al., 2006). This crop is considered as moderately sensitive to salt (salinity threshold in the range of 0.75 - 6.8 dS m-1) (Tedeschi et al., 2011). It has been reported that the hig-hest sensibility occurs in early vegetative stages, while its tolerance increases between fruit development and harvest (Franco et al., 1993; Nukaya et al., 1984). High salinity can delay or inhibit germination (Carvajal et al., 1998; Tanveer et al., 2012), inhibit growth (Bokochoy-Nbok and Вokochoy, 1991; Botía et al., 1998) and reduce yield (Huang et al., 2012; Sivritepe et al., 2005).
Seed priming is a technique of controlled hydration that allows activation of numerous metabolic processes (pre-germinative metabolism) which occur in early stages of imbibition, before radicle protrusion (Paparella et al., 2015). There are different priming methods, which vary based on the way the entry of water is controlled: osmo-priming (seeds are soaked in an osmotic solution such as polyethylene glycol, mannitol or salts), hydropriming (seeds are soaked in a determined amount of water, or limited imbibition periods), or matrix priming (organic or inorganic solids are mixed with water to reduce osmotic potential) (Wojtyla et al., 2016). Priming can improve seed germination under favorable and unfavorable conditions, although its effects are more evident under adverse con-ditions (Jisha et al., 2013). The advantages of priming are not exclusive to the seed stage but it can also improve growth and yield of crops under different abiotic stressors (Castañares and Bouzo, 2017; Zheng et al., 2016; Far-houdi and Sharifzadeh, 2006).
The objective of this work was to evaluate the effect of priming on germination, initial growth, and biochemical res-ponses of melon plants grown in saline conditions.
MATERIALS AND METHODS
Seeds of F1 hybrid ´Charentais´ cantaloupe ´Planter´s Jumbo´ (Bonanza Seeds, California, USA) were used. The
different reagents and salts (analytical grade) used were obtained from Sigma-Aldrich (Buenos Aires, Argentina).
Germination tests
Seeds were surface-disinfected using a 10% sodium hypochlorite solution (NaOCl in water, v/v) for 10 min. Then, the seeds were rinsed with deionized water, pla-ced between two filter papers (Whatman No 1) in 8.5 cm diameter Petri dishes, and soaked with either one of two different osmotic solutions: NaCl or CaCl2, according to a previous study (Castañares and Bouzo, 2018), in a ratio of 1:5 (wt/v) (Farooq et al., 2013). The concentration of these salts was calculated to achieve an osmotic potential (Ψo) of -1.5 MPa according to the van’t Hoff equation (Shabala and Munns, 2012). Two priming durations were studied: two and four days. Ten seeds per dish were used, equidis-tant from each other, with five replicates per treatment. The assay was performed in a germination chamber (Semedic, model I-500 PF, Argentina) at 25 °C (± 0.5 °C) in darkness (Nascimento, 2003). After priming, seeds were removed, rinsed three times in distilled water, and air-dried at 25 °C for 24 h to reduce the moisture content to < 10%. Later, the seeds were germinated between two filter papers and imbibed with an 80 mM NaCl solution (EC = 8.0 dS m-1). Control treatment (Control S1) consisted of seeds without priming, treated with distilled water. Germination was re-corded daily during 8 days (ISTA, 2013) and seeds were labeled as germinated when the seed coat was broken and the radicle was visible (>2 mm). Germination percentage (GP) was calculated as the relation between germinated seeds and seeds put to germinate.
Growth of seedlings
The two treatments with the highest GP from the pre-vious experiment were chosen to evaluate the growth response of seedlings. After priming, seeds were rinsed three times in distilled water and sown in 1000 cm3 pots previously filled with perlite, and watered with Hoagland´s nutrient solution (Hoagland and Arnon, 1950), with 2.0 dS m-1 electrical conductivity (EC). After first leaf emergence, plants started to be watered with the nutrient solution with the addition of approximately 60 mM NaCl, to achieve 8.0 dS m-1 EC. EC was monitored daily with a portable con-ductivity meter (Milwaukee model WP MC66 0-10MS/C, USA). Two control groups were established: Control S0, plants derived from non-primed seeds and watered with a 2.0 dS m-1 EC solution; and Control S1, plants derived from non-primed seeds and watered with an 8.0 dS m-1 EC solution. The experiment was performed in a phytotron, with a 16 h light/8 h darkness cycle and temperatures of 25/18 °C (± 1.0 °C), optimal for melon vegetative growth (Reche Mármol, 2008).
The experiment had a completely randomized design. The position of the pots was randomly changed daily to minimize positional effects in the phytotron. All values re-ported are the means of five replicates. Data obtained were

Seed priming induces biochemical changes in melon plants and increase salt tolerance
ARTÍCULOS RIA / Vol. 46 / N.º 2210210
leaf material was homogenized in 10 mL of 3% aqueous 5-sulfosalicylic acid solution, and the homogenate was filte-red using Whatman No 1 paper. An aliquot of 2 mL of filtrate was reacted with 2 mL of acid ninhydrin (1,2,3-triketohydrin-dene hydrate) and 2 mL of glacial acetic acid (C2H4O2) for 1 h at 100 °C. The reaction was terminated on ice for 15 min. The reaction mixture was then extracted with 4 mL of toluene (C7H8) and vortexed for 20 s. The absorption of the upper phase was measured at 520 nm using a spectropho-tometer and proline concentration was calculated using a standard curve and expressed as µmol g-1 FW.
Relative electrolyte leakage (EL) was estimated according to Dionisio-Sese and Tobita (1998). A total of 100 mg of fresh leaves were cut into 5 mm segments and placed in test tubes with 10 mL of deionized water. The tubes were incubated 2 h at 30 °C and the initial EC of the medium was measu-red (EC1) using a conductivity meter (Altronix model CTX-II, USA). Then the test tubes were incubated at 100 °C for 15 min and the EC was measured again (EC2). EL was estima-ted according to the formula: EL (%) = EC1/EC2 × 100.
To evaluate the level of malondialdehyde (MDA), a sample of 300 mg of fresh leaves was macerated with 3 mL of 0.1% trichloroacetic acid (Cl3C2O2H) (TCA). The homogenate was centrifuged at 3,000 x g for 10 min. Then, 1 mL of the su-pernatant was placed in a test tube, and 1 mL of the reagent TCA-BHT-TBA (TCA 20%, thiobarbituric acid (TBA) 0.37% and butyl hydroxyl toluene (BHT) 0.01 g) was added. The sample was incubated in a water bath for 25 min at 95 °C. The reaction was terminated on ice. Then the sample was centri-fuged at 10,000 x g for 10 min. The supernatant was used to measure the absorbance at 532 and 600 nm. The content of MDA was expressed as μM g-1 FW (Heath and Packer, 1968).
Peroxidase and catalase activity
After 20 days of growth, fresh leaf samples (60 mg) were ground in a mortar, re-suspended with 1.5 mL of 10 mM phosphate buffer (pH 6), and centrifuged at 12,000 x g for 20 min. The supernatant was used for the following assays.
To determine the peroxidase activity (POX), 100 μL of 10 mM sodium phosphate buffer (pH 6), 20 μL of guaiacol ((CH3O) C6H4OH) 0.25% (v/v) and 50 μL of 0.88 M H2O2 were added to 100 μL of the extract. A reduction in absorbance at 470 nm was measured at 0.5, 1.0, 1.5, and 2.0 min. POX ac-tivity was expressed as Abs min-1 mg-1 FW (George, 1953).
Catalase activity (CAT) was measured according to Aebi (1984). An aliquot of 50 μL was extracted, and 2000 μL of 50 mM sodium potassium phosphate buffer (pH 7) and 20 μL of 0.88 M H2O2 were added. The activity was determined by measuring the decrease in absorbance due to H2O2 de-composition for 1.0 min at 240 nm and expressed as μmol of H2O2 reduced g-1 FW.
Determination Na+/K+ Ratio and K+ leakage from roots
The following determinations were also made in seed-lings after having grown for 20 days in salinity.
analyzed using Tukey’s test at 0.05 confidence level using Infostat statistical software (Di Rienzo et al., 2011).
Growth evaluation
Five plants were extracted from each treatment after 40 days, and the following parameters were determined: leaf number, main stem height, leaf area, and total dry weight. Leaf area was determined using digitized images of leaves with Image J software (Schneider et al., 2012). For stem height, the distance from the substrate surface up to the in-sertion of the petiole of the uppermost leaf was considered.
Xylematic potential measurement
Xylematic potential (Ψx) of plants was measured using a Scholander pressure chamber (Biocontrol model 0-6 MPA, Argentina). Measurements were made on developed lea-ves, located in the middle third of the stem (Scholander et al., 1964), after 20 days of growth.
Determination of chlorophyll content, relative water content, root viability, proline and malondialdehyde concentration, and relative electrolyte leakage
After 20 days of growth in salinity, five plants per treatment were harvested and used for biochemical determinations.
Chlorophyll content (Ch) was determined according to Lichtenthaler and Wellburn (1983). A sample of 100 mg of fresh leaves (PF) was homogenized with 20 mL (V) of 80% (v/v) acetone in a mortar. Then the mixture was centrifuged at 8,000 × g for 10 min. The absorbance (A) of supernatant was measured using a spectrophotometer (Shimadzu mo-del UV-1800, Japan) at 646.8 and 663.2 nm, and chlorophyll content was determined according to the following formula: Ch (mg g-1 FW) = (7.15 A 663.2 nm + 18.71 A 646.8 nm) x (V/PF).
Relative water content (RWC) was calculated as descri-bed by Cao et al. (2015). The second leaves were extracted and fresh weight (FW) was determined. Then the leaves were placed in a beaker with distilled water for 5 h to deter-mine saturated weight (SW). Finally, the leaves were dried at 80 °C until constant weight and dry weight (DW) was determined. RWC was calculated according the equation: RWC (%) = (FW–DW)/(SW–DW) × 100.
Root viability was estimated by measuring the activity of the dehydrogenase enzyme by using the 2,3,5- triphenyl te-trazolium chloride (TTC) reduction technique (Clemensson-Lindell, 1994). A sample of 500 mg of fresh roots was cut into small pieces, put into test tubes with 5 mL of TTC 0.4 % and 5 mL of phosphate buffer (pH 7.0), and incubated for 3 h at 37 °C. The samples were then extracted in ethyl aceta-te for 15 min. The absorbance of samples was measured at 485 nm. Results were expressed as absorbance in relation to root dry weight, determined after drying in an 80 °C oven until constant weight (A485 nm g-1 DW).
Proline content was measured using the method develo-ped by Bates et al. (1973). Approximately 300 mg of fresh
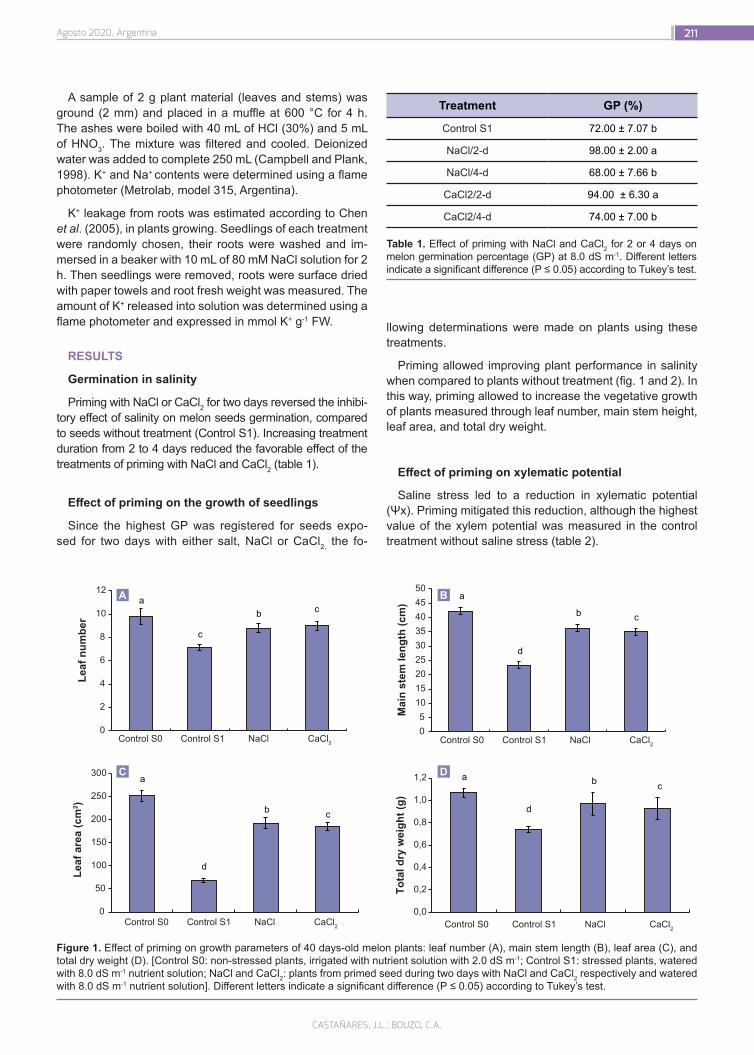
CASTAÑARES, J.L.; BOUZO, C.A.
Agosto 2020, Argentina 211211
A sample of 2 g plant material (leaves and stems) was ground (2 mm) and placed in a muffle at 600 °C for 4 h. The ashes were boiled with 40 mL of HCl (30%) and 5 mL of HNO3. The mixture was filtered and cooled. Deionized water was added to complete 250 mL (Campbell and Plank, 1998). K+ and Na+ contents were determined using a flame photometer (Metrolab, model 315, Argentina).
K+ leakage from roots was estimated according to Chen et al. (2005), in plants growing. Seedlings of each treatment were randomly chosen, their roots were washed and im-mersed in a beaker with 10 mL of 80 mM NaCl solution for 2 h. Then seedlings were removed, roots were surface dried with paper towels and root fresh weight was measured. The amount of K+ released into solution was determined using a flame photometer and expressed in mmol K+ g-1 FW.
RESULTS
Germination in salinity
Priming with NaCl or CaCl2 for two days reversed the inhibi-tory effect of salinity on melon seeds germination, compared to seeds without treatment (Control S1). Increasing treatment duration from 2 to 4 days reduced the favorable effect of the treatments of priming with NaCl and CaCl2 (table 1).
Effect of priming on the growth of seedlings
Since the highest GP was registered for seeds expo-sed for two days with either salt, NaCl or CaCl2, the fo-
llowing determinations were made on plants using these treatments.
Priming allowed improving plant performance in salinity when compared to plants without treatment (fig. 1 and 2). In this way, priming allowed to increase the vegetative growth of plants measured through leaf number, main stem height, leaf area, and total dry weight.
Effect of priming on xylematic potential
Saline stress led to a reduction in xylematic potential (Ψx). Priming mitigated this reduction, although the highest value of the xylem potential was measured in the control treatment without saline stress (table 2).
Table 1. Effect of priming with NaCl and CaCl2 for 2 or 4 days on melon germination percentage (GP) at 8.0 dS m-1. Different letters indicate a significant difference (P ≤ 0.05) according to Tukey’s test.
Treatment GP (%)
Control S1 72.00 ± 7.07 b
NaCl/2-d 98.00 ± 2.00 a
NaCl/4-d 68.00 ± 7.66 b
CaCl2/2-d 94.00 ± 6.30 a
CaCl2/4-d 74.00 ± 7.00 b
Figure 1. Effect of priming on growth parameters of 40 days-old melon plants: leaf number (A), main stem length (B), leaf area (C), and total dry weight (D). [Control S0: non-stressed plants, irrigated with nutrient solution with 2.0 dS m-1; Control S1: stressed plants, watered with 8.0 dS m-1 nutrient solution; NaCl and CaCl2: plants from primed seed during two days with NaCl and CaCl2 respectively and watered with 8.0 dS m-1 nutrient solution]. Different letters indicate a significant difference (P ≤ 0.05) according to Tukey’s test.
a
c
b c
0
2
4
6
8
10
12
Leaf
num
ber
A B
C D
Control S0 Control S1 NaCl CaCl2
a
d
b c
05
101520253035404550
Mai
n st
em le
ngth
(cm
)
Control S0 Control S1 NaCl CaCl2
a
d
b c
0
50
100
150
200
250
300
Leaf
are
a (c
m2 )
Control S0 Control S1 NaCl CaCl2
a
d
b c
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Tota
l dry
wei
ght (
g)
Control S0 Control S1 NaCl CaCl2
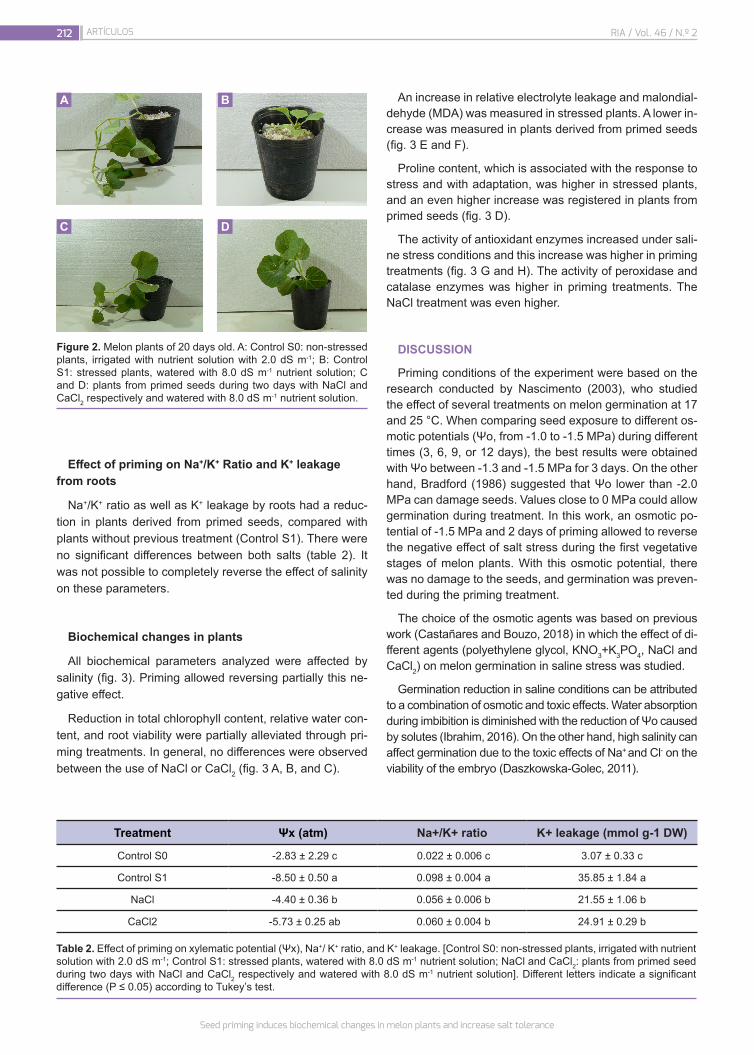
Seed priming induces biochemical changes in melon plants and increase salt tolerance
ARTÍCULOS RIA / Vol. 46 / N.º 2212212
Figure 2. Melon plants of 20 days old. A: Control S0: non-stressed plants, irrigated with nutrient solution with 2.0 dS m-1; B: Control S1: stressed plants, watered with 8.0 dS m-1 nutrient solution; C and D: plants from primed seeds during two days with NaCl and CaCl2 respectively and watered with 8.0 dS m-1 nutrient solution.
A
C
B
D
Effect of priming on Na+/K+ Ratio and K+ leakage from roots
Na+/K+ ratio as well as K+ leakage by roots had a reduc-tion in plants derived from primed seeds, compared with plants without previous treatment (Control S1). There were no significant differences between both salts (table 2). It was not possible to completely reverse the effect of salinity on these parameters.
Biochemical changes in plants
All biochemical parameters analyzed were affected by salinity (fig. 3). Priming allowed reversing partially this ne-gative effect.
Reduction in total chlorophyll content, relative water con-tent, and root viability were partially alleviated through pri-ming treatments. In general, no differences were observed between the use of NaCl or CaCl2 (fig. 3 A, B, and C).
Table 2. Effect of priming on xylematic potential (Ψx), Na+/ K+ ratio, and K+ leakage. [Control S0: non-stressed plants, irrigated with nutrient solution with 2.0 dS m-1; Control S1: stressed plants, watered with 8.0 dS m-1 nutrient solution; NaCl and CaCl2: plants from primed seed during two days with NaCl and CaCl2 respectively and watered with 8.0 dS m-1 nutrient solution]. Different letters indicate a significant difference (P ≤ 0.05) according to Tukey’s test.
Treatment Ψx (atm) Na+/K+ ratio K+ leakage (mmol g-1 DW)
Control S0 -2.83 ± 2.29 c 0.022 ± 0.006 c 3.07 ± 0.33 c
Control S1 -8.50 ± 0.50 a 0.098 ± 0.004 a 35.85 ± 1.84 a
NaCl -4.40 ± 0.36 b 0.056 ± 0.006 b 21.55 ± 1.06 b
CaCl2 -5.73 ± 0.25 ab 0.060 ± 0.004 b 24.91 ± 0.29 b
An increase in relative electrolyte leakage and malondial-dehyde (MDA) was measured in stressed plants. A lower in-crease was measured in plants derived from primed seeds (fig. 3 E and F).
Proline content, which is associated with the response to stress and with adaptation, was higher in stressed plants, and an even higher increase was registered in plants from primed seeds (fig. 3 D).
The activity of antioxidant enzymes increased under sali-ne stress conditions and this increase was higher in priming treatments (fig. 3 G and H). The activity of peroxidase and catalase enzymes was higher in priming treatments. The NaCl treatment was even higher.
DISCUSSION
Priming conditions of the experiment were based on the research conducted by Nascimento (2003), who studied the effect of several treatments on melon germination at 17 and 25 °C. When comparing seed exposure to different os-motic potentials (Ψo, from -1.0 to -1.5 MPa) during different times (3, 6, 9, or 12 days), the best results were obtained with Ψo between -1.3 and -1.5 MPa for 3 days. On the other hand, Bradford (1986) suggested that Ψo lower than -2.0 MPa can damage seeds. Values close to 0 MPa could allow germination during treatment. In this work, an osmotic po-tential of -1.5 MPa and 2 days of priming allowed to reverse the negative effect of salt stress during the first vegetative stages of melon plants. With this osmotic potential, there was no damage to the seeds, and germination was preven-ted during the priming treatment.
The choice of the osmotic agents was based on previous work (Castañares and Bouzo, 2018) in which the effect of di-fferent agents (polyethylene glycol, KNO3+K3PO4, NaCl and CaCl2) on melon germination in saline stress was studied.
Germination reduction in saline conditions can be attributed to a combination of osmotic and toxic effects. Water absorption during imbibition is diminished with the reduction of Ψo caused by solutes (Ibrahim, 2016). On the other hand, high salinity can affect germination due to the toxic effects of Na+ and Cl- on the viability of the embryo (Daszkowska-Golec, 2011).
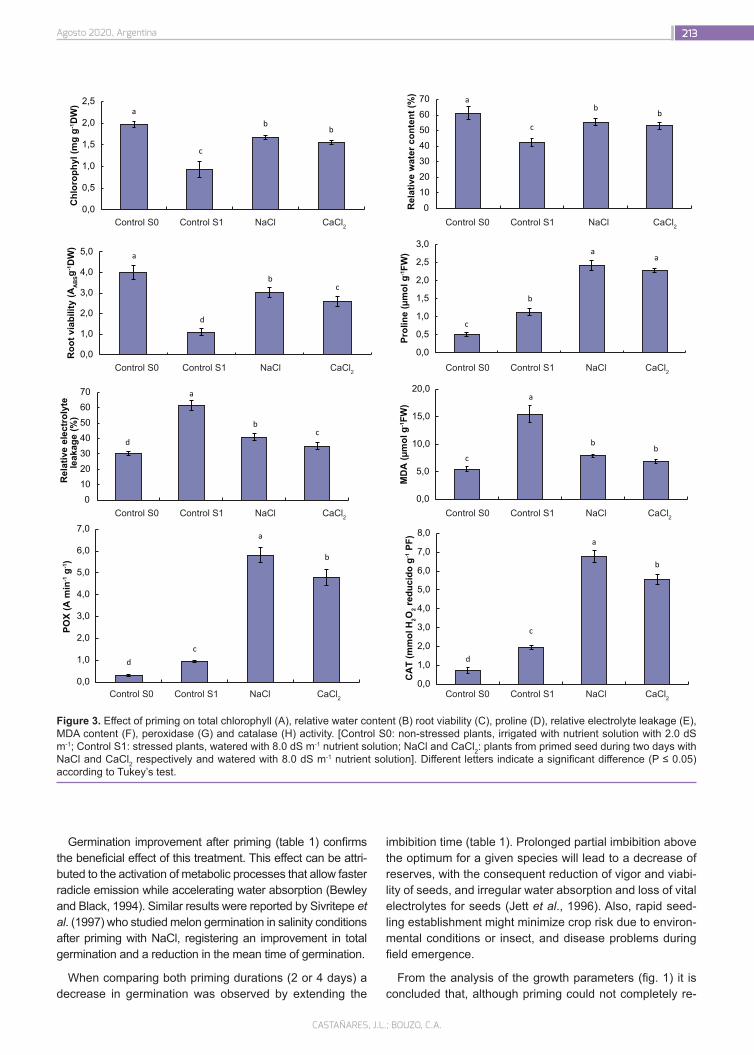
CASTAÑARES, J.L.; BOUZO, C.A.
Agosto 2020, Argentina 213213
Figure 3. Effect of priming on total chlorophyll (A), relative water content (B) root viability (C), proline (D), relative electrolyte leakage (E), MDA content (F), peroxidase (G) and catalase (H) activity. [Control S0: non-stressed plants, irrigated with nutrient solution with 2.0 dS m-1; Control S1: stressed plants, watered with 8.0 dS m-1 nutrient solution; NaCl and CaCl2: plants from primed seed during two days with NaCl and CaCl2 respectively and watered with 8.0 dS m-1 nutrient solution]. Different letters indicate a significant difference (P ≤ 0.05) according to Tukey’s test.
Germination improvement after priming (table 1) confirms the beneficial effect of this treatment. This effect can be attri-buted to the activation of metabolic processes that allow faster radicle emission while accelerating water absorption (Bewley and Black, 1994). Similar results were reported by Sivritepe et al. (1997) who studied melon germination in salinity conditions after priming with NaCl, registering an improvement in total germination and a reduction in the mean time of germination.
When comparing both priming durations (2 or 4 days) a decrease in germination was observed by extending the
imbibition time (table 1). Prolonged partial imbibition above the optimum for a given species will lead to a decrease of reserves, with the consequent reduction of vigor and viabi-lity of seeds, and irregular water absorption and loss of vital electrolytes for seeds (Jett et al., 1996). Also, rapid seed-ling establishment might minimize crop risk due to environ-mental conditions or insect, and disease problems during field emergence.
From the analysis of the growth parameters (fig. 1) it is concluded that, although priming could not completely re-
a
c
bb
0,0
0,5
1,0
1,5
2,0
2,5
Chl
orop
hyl (
mg
g-1D
W) a
c
bb
010203040506070
Rel
ativ
e w
ater
con
tent
(%)
a
d
bc
0,0
1,0
2,0
3,0
4,0
5,0
Roo
t via
bilit
y (A
AB
Sg-1D
W)
c
b
aa
0,0
0,5
1,0
1,5
2,0
2,5
3,0
Prol
ine
(µm
ol g
-1FW
)
d
a
cb
010203040506070
Rel
ativ
e el
ectr
olyt
e le
akag
e (%
)
c
a
bb
0,0
5,0
10,0
15,0
20,0M
DA
(µm
ol g
-1FW
)
dc
a
b
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
POX
(A m
in-1 g
-1)
d
c
a
b
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
8,0
CA
T (m
mol
H2O
2 red
ucid
o g-1
PF)
Control S0 Control S1 NaCl CaCl2 Control S0 Control S1 NaCl CaCl2
Control S0 Control S1 NaCl CaCl2 Control S0 Control S1 NaCl CaCl2
Control S0 Control S1 NaCl CaCl2 Control S0 Control S1 NaCl CaCl2
Control S0 Control S1 NaCl CaCl2 Control S0 Control S1 NaCl CaCl2

Seed priming induces biochemical changes in melon plants and increase salt tolerance
ARTÍCULOS RIA / Vol. 46 / N.º 2214214
verse the effect of salt stress, an increase in tolerance was recorded. Sivritepe et al. (2003) observed that with EC va-lues of up to 9.0 dS m-1, there was a difference in the dry weight of melon plants from primed seeds. At higher levels of salinity, there was no difference between treated and un-treated plants.
Xylematic potential (Ψx) indicates water tension in plants’ vessels which is related to the degree of stress (Selles and Ferreyra, 2005). The reduction of Ψx registered in plants growing under salinity conditions (table 2) confirms its effect on the hydric state of plants, due to the reduction of water availability (Shabala and Munns, 2012). The de-crease in Ψx was lower in NaCl treatment, which suggests the activation of osmotic adjustment mechanisms, such as accumulation of compatible osmolytes, mainly proline (fig. 3 D), glycine betaine and sucrose, which reduce water po-tential of cells, protect against membrane damage and sta-bilize proteins and enzymes (Singh et al., 2015). Proline ac-cumulation in plants partially reduces the negative effects of stress given the ability of this molecule to participate in processes of osmotic adjustment and neutralization of ROS (Rejeb et al., 2014). As observed in fig. 3 D, stressed plants increased proline content and an even higher content of this amino acid was measured in priming treatments. The increase in proline content in salt stressed plants was wi-dely documented. In primed seeds, for example, was regis-tered in pepper (Capsicum annuum L.) (Aloui et al., 2014), rice (Oryza sativa L.) (Li and Zhang, 2012) and corn (Zea mays L.) (Bakht et al., 2011).
Reduction of chlorophyll content in plants under saline stress can be related to a reduction of plant biomass and an increase in lipid peroxidation of the chloroplast´s mem-brane due to oxidative stress, and the accumulation to to-xic levels of ions that affect synthesis (Ashraf and Harris, 2013). The increase or reduction of chlorophyll content in saline stress has been proposed in melon as an indicator of tolerance or sensitivity to salinity, respectively (Romero et al., 1997). There was a lower reduction of this pigment’s content in primed treatments (fig. 3 A). When working with wheat (Triticum aestivum L.), Farooq et al. (2013), found a high correlation between the reduction of chlorophyll con-tent in plants under salt stress and the degree of lipid pe-roxidation and membrane damage. Our results confirm this relationship (fig. 3 A, E, and F).
Reduction of relative water content (RWC) in stressed plants (fig. 3 B) can be related to lower water absorption due to damage of the root system or the low Ψ of the subs-trate (Kukreja et al., 2005). In this way, the increase of RWC in priming treatments can be explained, on the one hand, by the lower root damage, evidenced with the highest va-lues of reaction to TTC (fig. 3 C), as an indicator of root viability (Ruf and Brunner, 2003).
Stress in plants triggers numerous processes that lead to the generation of ROS. These ROS do not necessarily have a negative effect on plants, but they do have an im-portant function in intracellular communication that allows the best acclimatization to environmental stress (Rejeb et
al., 2014). However, excessive accumulation causes the initiation of a significant number of auto oxidative chain re-actions, mainly lipid peroxidation, DNA damage and protein degradation (Mittler, 2002), since these are molecules with a highly unstable configuration. Therefore, radicals quickly react with other molecules, generating more free radicals (Ashraf, 2009). Hydrogen peroxide (H2O2), which was not measured here, is the most harmful ROS and has been wi-dely studied because of its relative stability (Mhamdi et al., 2010). Antioxidant enzymes, particularly peroxidases (POX) and catalases (CAT) are very efficient at degrading H2O2 (Li et al., 2014). Therefore, the high values measured here of these enzymes (fig. 3 G and H) indicate effective mitiga-tion of the possible damage of H2O2 on melon plants. Saline stress caused an increase in the activity of POX (fig. 3 G) and CAT (fig. 3 H) enzymes. Priming led to an even greater increase, primarily for NaCl followed by CaCl2. These re-sults agree with was reported by Islam et al. (2015) who studied the response of priming with CaCl2 and KCl in two wheat cultivars (Triticum aestivum L.) grown in saline soil, and reported a higher antioxidant activity. Salah et al. (2015) cultivated rice plants (Oryza sativa L.) under toxic levels of Zn, and observed a higher POX and CAT activity, and lower levels of MDA, in plants from seeds primed with PEG.
Under high salinity conditions, Na+ has the ability to com-pete with K+. Many physiological studies have demonstrated that Na+ toxicity is caused by the ability of Na+ to compete for K+ binding sites to disrupt K+ homeostasis (Hasegawa, 2013). One of the main characteristics of salt tolerant plants is the ability to maintain a low Na+/ K+ ratio (Tester and Da-venport, 2003). In our experiments, plant growing in saline conditions increased Na+/K+ (table 2). This can be explai-ned by a greater Na+ accumulation in tissues and an increa-se in K+ leakage by cells (Poustini and Siosemardeh, 2004). The reduction of Na+/K+ ratio in plants from primed seeds contributes to a better adaptation to salinity. In wheat, Ab-basdokht and Edalatpisheh (2012) observed that plants from primed seeds had low Na+/K+ ratio, while the untreated plants exhibited higher ratios. Working with tomato (Sola-num lycopersicum L.), Cayuela et al. (1996) reported that priming with NaCl allowed reducing Na+ content in plants that best grow in salinity. Plants growing in salt stress can experience an irreversible loss of K+, which is particularly important in roots. This is related to the loss of integrity of membranes, reducing the ability to retain K+ (Demidchik et al., 2014). Low K+ leakage from roots is accepted as an in-dicator of salt tolerance (Chen et al., 2005). The lower K+ release measured in priming treatments (table 2) confirms the greater stability of membranes, also estimated from re-lative electrolyte leakage (fig. 3 E) and MDA (fig. 3 F). K+ is recognized as an important factor limiting yield and quality of crops, and functions include maintenance of cell turgor, growth, activation of numerous enzymes, stabilization of protein synthesis, the formation of membrane potential, and maintenance of cytosolic pH (Cakmak, 2005). Relative electrolyte leakage is considered an estimator of membra-ne integrity (Demidchik et al., 2014). A greater release of electrolytes, estimated from EC, indicates membrane da-mage. This was measured in stressed plants (Control S1)

CASTAÑARES, J.L.; BOUZO, C.A.
Agosto 2020, Argentina 215215
while priming allowed reducing this damage (fig. 3 E). The final product of polyunsaturated fatty acids oxidation is ma-londialdehyde (MDA). This molecule reacts with thiobarbi-turic acid (TBA) and generated a pink compound detectable by spectrophotometry (Hodges et al., 1999). The increase of the MDA level in the conditions of the experiment (fig. 3 F) confirms the membrane damage, which corroborates the results discussed above by the electrolyte leakage measu-rement (fig. 3 E). Thus, priming allowed improving the inte-grity of membranes under the conditions of the experiment. This response is in agreement with what was observed by Randhir and Shetty (2005) in corn (Zea mays L.) and Amooaghaie (2011) in alfalfa (Medicago sativa L.).
CONCLUSIONS
The results of this study show that priming induces bio-chemical changes in melon seedlings increasing the adap-tive response to salinity. Melon is cultivated intensively and with complementary irrigation, which promotes the accu-mulation of salts in the soil. Given that the studied salts have a relatively low cost and are easy to manipulate, the information obtained could be of interest to producers.
REFERENCESABBASDOKHT, H.; EDALATPISHEH, M.R. 2012. The effect of
priming and salinity on physiological and chemical characteristics of wheat (Triticum aestivum L.). Desert 17, 183–192.
AEBI, H. 1984. Catalase in vitro, in: PACKER, L. (Ed.). Methods in Enzymology. Academic Press, New York. 121–126 pp.
ALOUI, H.; SOUGUIR, M.; LATIQUE, S.; HANNACHI, C. 2014. Germination and growth in control and primed seeds of pepper as affected by salt stress. Cercet. Agron. Mold. 47, 83–95.
AMOOAGHAIE, R. 2011. The effect of hydro and osmopriming on alfalfa seed germination and antioxidant defenses under salt stress. African J. Biotechnol. 10, 6269–6275.
ASHRAF, M. 2009. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv. 27, 84–93. https://doi.org/10.1016/j.biotechadv.2008.09.003
ASHRAF, M.; HARRIS, P.J.C. 2013. Photosynthesis under stressful environments: an overview. Photosynthetica 51, 163–190.
BAKHT, J.; SHAFI, M.; JAMAL, Y.; SHER, H. 2011. Response of maize (Zea mays L.) to seed priming with NaCl and salinity stress. Spanish J. Agric. Res. 9, 252–261.
BATES, L.S.; WALDREN, R.P.; TEARE, I.D. 1973. Rapid de-termination of free proline for water-stress studies. Plant Soil 39, 205–207. https://doi.org/10.1007/BF00018060
BEWLEY, J.D.; BLACK, M. 1994. Seeds, in: Seeds. Springer, New York. 1–33 pp.
BOKOCHOY-NBOK, H.; ВOKOCHOY, А. 1991. Response of melon plants to salt: 1. Growth, morphology and root membrane properties. J. Plant Physiol 139, 100–105.
BOTÍA, P.; CARVAJAL, M.; CERDÁ, A.; MARTÍNEZ, V. 1998. Response of eight Cucumis melo cultivars to salinity during ger-mination and early vegetative growth. Agronomie 18, 503–513. https://doi.org/10.1051/agro:19980801
BRADFORD, K.J. 1986. Manipulation of seed water relations via osmotic priming to improve germination under stress conditions. HortScience (USA) 21, 1105–1113.
CAKMAK, I. 2005. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 168, 521–530.
CAMPBELL, C.R.; PLANK, C.O. 1998. Preparation of plant tis-sue for laboratory analysis, in: KALRA, Y.P. (Ed.), Handbook of Methods for Plant Analysis. Taylor & Francis Group, New York. 51–63 pp.
CAO, Y.Y.; YANG, M.T.; CHEN, S.Y.; ZHOU, Z.Q.; LI, X.; WANG, X.J.; BAI, J.G. 2015. Exogenous sucrose influences antioxidant enzyme activities and reduces lipid peroxidation in water-stressed cucumber leaves. Biol. Plant. 59, 147–153.
CARVAJAL, M.; DEL AMOR, F.M.; FERNANDEZ-BALLESTER, G.; MARTÍNEZ, V.; CERDÁ, A. 1998. Time course of solute accu-mulation and water relations in muskmelon plants exposed to salt during different growth stages. Plant Sci. 138, 103–112.
CASTAÑARES, J.L.; BOUZO, C.A. 2018. Effect of different pri-ming treatments and priming durations on melon germination be-havior under suboptimal conditions. Open Agric. 3, 386–392.
CASTAÑARES, J.L.; BOUZO, C.A. 2017. Effect of priming on germination and initial growth of melon plants under salt stress. FAVE Sección Ciencias Agrar. 16, 7–16.
CAYUELA, E.; PÉREZ-ALFOCEA, F.; CARO, M.; BOLARÍN, M.C. 1996. Priming of seeds with NaCl induces physiological chan-ges in tomato plants grown under salt stress. Physiol. Plant. 96, 231–236. https://doi.org/10.1111/j.1399-3054.1996.tb00207.x
CHEN, Z.; NEWMAN, I.; ZHOU, M.; MENDHAM, N.; ZHANG, G.; SHABALA, S. 2005. Screening plants for salt tolerance by measuring K+ flux: a case study for barley. Plant. Cell Environ. 28, 1230–1246.
CLEMENSSON-LINDELL, A. 1994. Triphenyltetrazolium chlo-ride as an indicator of fine-root vitality and environmental stress in coniferous forest stands: applications and limitations. Plant Soil 159, 297–300.
DASZKOWSKA-GOLEC, A. 2011. Arabidopsis seed germina-tion under abiotic stress as a concert of action of phytohormones. Omi. a J. Integr. Biol. 15, 763–774.
DEMIDCHIK, V.; STRALTSOVA, D.; MEDVEDEV, S.S.; PO-ZHVANOV, G.A.; SOKOLIK, A.; YURIN, V. 2014. Stress-induced electrolyte leakage: the role of K+-permeable channels and invol-vement in programmed cell death and metabolic adjustment. J. Exp. Bot. 65, 1259–1270.
DI RIENZO, J.A.; CASANOVES, F.; BALZARINI, M.G.; GONZÁ-LEZ, L.; TABLADA, M.; ROBLEDO, Y.C.W. 2011. InfoStat versión 2011. Grup. InfoStat, FCA, Univ. Nac. Córdoba, Argentina. (Available at: http//www. infostat. com.ar verified: December 2019) 195–199.
DIONISIO-SESE, M.L.; TOBITA, S. 1998. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 135, 1–9.
FARHOUDI, R.; SHARIFZADEH, F. 2006. The effects of NaCl priming on salt tolerance in canola (Brassica napus L.) seedlings grown under saline conditions. Indian J Crop Sci 1, 74–78.
FAROOQ, M.; IRFAN, M.; AZIZ, T.; AHMAD, I.; CHEEMA, S.A. 2013. Seed priming with ascorbic acid improves drought resistan-ce of wheat. J. Agron. Crop Sci. 66, 12–22.
FRANCO, J.A.; ESTEBAN, C.; RODRIGUEZ, C. 1993. Effects of salinity on various growth stages of muskmelon cv. Revigal. J. Hortic. Sci. 68, 899–904.
GEORGE, P. 1953. The chemical nature of the second hydrogen peroxide compound formed by cytochrome c peroxidase and hor-seradish peroxidase. 1. Titration with reducing agents. Biochem. J. 54, 267–276.
HASEGAWA, P.M. 2013. Sodium (Na+) homeostasis and salt tolerance of plants. Environ. Exp. Bot. 92, 19–31.

Seed priming induces biochemical changes in melon plants and increase salt tolerance
ARTÍCULOS RIA / Vol. 46 / N.º 2216216
HEATH, R.L.; PACKER, L. 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxida-tion. Arch. Biochem. Biophys. 125, 189–198.
HOAGLAND, D.R.; ARNON, D.I. 1950. The Water-Culture Method for Growing Plants without Soil. Calif. Agric. Exp. Stn. 51, 914–916.
HODGES, D.M.; DELONG, J.M.; FORNEY, C.F.; PRANGE, R.K. 1999. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing antho-cyanin and other interfering compounds. Planta 207, 604–611.
HUANG, C.H., ZONG, L.; BUONANNO, M.; XUE, X.; WANG, T.; TEDESCHI, A. 2012. Impact of saline water irrigation on yield and quality of melon (Cucumis melo cv. Huanghemi) in northwest China. Eur. J. Agron. 43, 68–76.
IBRAHIM, E.A. 2016. Seed priming to alleviate salinity stress in germinating seeds. J. Plant Physiol. 192, 38–46. https://doi.org/10.1016/j.jplph.2015.12.011
IQBAL, M.; UL HAQ, Z.; JAMIL, Y.; NISAR, J. 2016. Pre-sowing seed magnetic field treatment influence on germination, seedling growth and enzymatic activities of melon (Cucumis melo L.). Bio-catal. Agric. Biotechnol. 6, 176–183.
ISLAM, F.; YASMEEN, T.; ALI, S.; ALI, B.; FAROOQ, M.A.; GILL, R.A. 2015. Priming-induced antioxidative responses in two wheat cultivars under saline stress. Acta Physiol. Plant. 37, 153–165.
ISTA. 2013. International rules for seed testing. Seed Sci. Tech-nol. 27, 25–30.
JETT, L.W.; WELBAUM, G.E.; MORSE, R.D. 1996. Effects of matric and osmotic priming treatments on broccoli seed germina-tion. J. Am. Soc. Hortic. Sci. 121, 423–429.
JISHA, K.C.; VIJAYAKUMARI, K.; PUTHUR, J.T. 2013. Seed pri-ming for abiotic stress tolerance: an overview. Acta Physiol. Plant. 35, 1381–1396.
KUKREJA, S.; NANDWAL, A.S.; KUMAR, N.; SHARMA, S.K.; UNVI, V.; SHARMA, P.K. 2005. Plant water status, H2 O2 scaven-ging enzymes, ethylene evolution and membrane integrity of Cicer arietinum roots as affected by salinity. Biol. Plant. 49, 305–308.
LI, C.; TAN, D.X.; LIANG, D.; CHANG, C.; JIA, D.; MA, F. 2014. Melatonin mediates the regulation of ABA metabolism, free-radical scavenging, and stomatal behaviour in two Malus species under drought stress. J. Exp. Bot. 66, 669–680.
LI, X.; ZHANG, L. 2012. SA and PEG-induced priming for water stress tolerance in rice seedling, in: Information Technology and Agricultural Engineering. Springer, 881–887 pp.
LICHTENTHALER, H.K.; WELLBURN, A.R. 1983. Determina-tions of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 11, 591–592.
MHAMDI, A.; QUEVAL, G.; CHAOUCH, S.; VANDERAUWERA, S.; VAN BREUSEGEM, F.; NOCTOR, G. 2010. Catalase function in plants: a focus on Arabidopsis mutants as stress-mimic models. J. Exp. Bot. 61, 4197–4220.
MITTLER, R. 2002. Oxidative stress, antioxidants and stress to-lerance. Trends Plant Sci. 7, 405–410.
MUNNS, R. 2005. Genes and salt tolerance: bringing them to-gether. New Phytol. 167, 645–663.
NASCIMENTO, W.M. 2003. Muskmelon seed germination and seed-ling development in response to seed priming. Sci. Agric. 60, 71–75.
NUKAYA, A.; MASUI, M.; ISHIDA, A. 1984. Salt tolerance of muskmelons as affected by diluted sea water applied at different growth stages in nutrient solution culture. J. Japanese Soc. Hortic. Sci. 53, 168–175.
PAPARELLA, S.; ARAÚJO, S.S.; ROSSI, G.; WIJAYASINGHE, M.; CARBONERA, D.; BALESTRAZZI, A. 2015. Seed priming: sta-
te of the art and new perspectives. Plant Cell Rep. 34, 1281–1293. https://doi.org/10.1007/s00299-015-1784-y
PASTOR, V.; BALMER, A.; GAMIR, J.; FLORS, V.; MAUCH-MANI, B. 2014. Preparing to fight back: generation and storage of priming compounds. Front. Plant Sci. 5, 1–12.
POUSTINI, K.; SIOSEMARDEH, A. 2004. Ion distribution in wheat cultivars in response to salinity stress. F. rops Res 85, 125–133.
QADIR, M.; QUILLÉROU, E.; NANGIA, V.; MURTAZA, G.; SINGH, M.; THOMAS, R.J.; DRECHSEL, P.; NOBLE, A.D. 2014. Economics of salt induced land degradation and restoration, in: Natural Resources Forum. Wiley Online Library. 282–295 pp.
RANDHIR, R.; SHETTY, K. 2005. Developmental stimulation of total phenolics and related antioxidant activity in light-and dark-ger-minated corn by natural elicitors. Process Biochem. 40, 1721–1732.
RECHE MÁRMOL, J. 2008. Cultivo del melón en invernadero, Generalitat Valenciana Conselleria d’Agricultura i Pesca. Valencia, España. Secretaría General Técnica. Servicio de Publicaciones y Divulgación, Andalucía.
REJEB, K.B.; ABDELLY, C.; SAVOURÉ, A. 2014. How reacti-ve oxygen species and proline face stress together. Plant Physiol. Biochem. 80, 278–284.
ROMERO, L.; BELAKBIR, A.; RAGALA, L.; RUIZ, J.M. 1997. Response of plant yield and leaf pigments to saline conditions: Effectiveness of different rootstocks in melon plants (Cucumis melo L.). Soil Sci. Plant Nutr. 43, 855–862. https://doi.org/10.1080/00380768.1997.10414652
RUF, M.; BRUNNER, I. 2003. Vitality of tree fine roots: reevalua-tion of the tetrazolium test. Tree Physiol. 23, 257–263.
SALAH, S.M.; YAJING, G.; DONGDONG, C.; JIE, L.; AAMIR, N.; QIJUAN, H.; WEIMIN, H.; MINGYU, N.; JIN, H. 2015. Seed priming with polyethylene glycol regulating the physiological and molecular mechanism in rice (Oryza sativa L.) under nano-ZnO stress. Sci. Rep. 5, 1–14.
SCHNEIDER, C.A.; RASBAND, W.S.; ELICEIRI, K.W. 2012. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 9, 671–675.
SCHOLANDER, P.F.; HAMMEL, H.T.; HEMMINGSEN, E.A.; BRADSTREET, E.D. 1964. Hydrostatic pressure and osmotic po-tential in leaves of mangroves and some other plants. Proc. Natl. Acad. Sci. 52, 119–125.
SELLES, G.; FERREYRA, R. 2005. Criterios para controlar el riego en uva de mesa. INIA (ed). Manejo Riego y Suelo en Vides para Vino y Mesa. INIA, Santiago, Chile 15–53.
SHABALA, S.; MUNNS, R. 2012. Salinity stress: physiological constraints and adaptative mechanism, in: SHABALA, S. (Ed.). Plant Stress Physiology. CAB International, London. 59–93 pp.
SINGH, M.; KUMAR, J.; SINGH, S.; SINGH, V.P.; PRASAD, S.M. 2015. Roles of osmoprotectants in improving salinity and drought tolerance in plants: a review. Rev. Environ. Sci. Bio/Tech-nology 14, 407–426.
SIVRITEPE, H.Ö.; SIVRITEPE, N.; ERIŞ, A.; TURHAN, E. 2005. The effects of NaCl pre-treatments on salt tolerance of melons grown under long-term salinity. Sci. Hortic. (Amsterdam). 106, 568–581.
SIVRITEPE, N.; SIVRITEPE, H.Ö.; ERIS, A. 2003. The effects of NaCl priming on salt tolerance in melon seedlings grown under saline conditions. Sci. Hortic. (Amsterdam). 97, 229–237.
SIVRITEPE, Ö.H.; ERIS, A.; SIVRITEPE, N. 1997. The effects of priming treatments on salt tolerance in melon seeds, in: I Interna-tional Symposium on Cucurbits 492. 287–296 pp.
TANVEER, A.; SALMAN ARSHAD, M.; AYUB, M.; MANSOOR JAVAID, M.; YASEEN, M. 2012. Effect of temperature, light, sali-

CASTAÑARES, J.L.; BOUZO, C.A.
Agosto 2020, Argentina 217217
nity, drought stress and seeding depth on germination of Cucumis melo var. agrestis. Pakistan J. Weed Sci. Res. 18, 445–459.
TEDESCHI, A.; LAVINI, A.; RICCARDI, M.; PULVENTO, C.; D’ANDRIA, R. 2011. Melon crops (Cucumis melo L., cv. Tendral) grown in a mediterranean environment under saline-sodic condi-tions: Part I. Yield and quality. Agric. water Manag. 98, 1329–1338.
TESTER, M.; DAVENPORT, R. 2003. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 91, 503–527.
WOJTYLA, Ł.; LECHOWSKA, K.; KUBALA, S.; GARNCZARS-KA, M. 2016. Molecular processes induced in primed seeds—in-creasing the potential to stabilize crop yields under drought condi-tions. J. Plant Physiol. 203, 116–126.
YADAV, S.; IRFAN, M.D.; AHMAD, A.; HAYAT, S. 2011. Causes of salinity and plant manifestations to salt stress: A review. J. Envi-ron. Biol. 32, 667–685.
YASAR, F.; KUSVURAN, S.; ELLIALTIOGLU, S. 2006. Determi-nation of anti-oxidant activities in some melon (Cucumis melo L.) varieties and cultivars under salt stress. J. Hortic. Sci. Biotechnol. 81, 627–630.
ZHENG, M.; TAO, Y.; HUSSAIN, S.; JIANG, Q.; PENG, S.; HUANG, J.; CUI, K.; NIE, L. 2016. Seed priming in dry direct-seed-ed rice: consequences for emergence, seedling growth and asso-ciated metabolic events under drought stress. Plant Growth Regul. 78, 167–178.

Caracterización socioproductiva de los productores hortícolas en el partido de Junín (Buenos Aires)
ARTÍCULOS RIA / Vol. 46 / N.º 2218218
Caracterización socioproductiva de los productores hortícolas en el partido de Junín (Buenos Aires)
RESUMENEl partido de Junín se ubica al noroeste de la provincia de Buenos Aires, abarca 226337 hectáreas y cuenta
con 90305 habitantes. La ciudad de Junín se configuró históricamente como el principal centro administrativo, turístico, educativo, industrial, de salud y comercial de la región. Debido a la aptitud de los suelos, se ubica en la zona núcleo de producción agropecuaria. Desde los 90 nuevos perfiles socioproductivos surgieron en gran cantidad de agrociudades intermedias, no solamente en el sector de campo extensivo, sino también en las zonas periurbanas. Los productores hortícolas son unos de los más característicos. El objetivo de este trabajo consiste en realizar una caracterización socioproductiva de los productores que integran el sector hortícola del partido de Junín. Para ello se elaboró un dispositivo metodológico basado en la realización de una encuesta en la que se relevan variables sociales y productivas. La metodología consiste en un barrido total de los productores hortícolas del partido de Junín. Se llega a la totalidad a partir de la conformación de un listado de productores en el que participaron las diferentes instituciones. A los fines de confirmar la llegada a la totalidad de los productores para corroborar el barrido se utiliza la fotointerpretación de Google Earth con imágenes del año 2016. Entre los principales resultados se encuentra que el 60% de los productores son de origen argentino, generalmente juninenses, y el 40% de origen boliviano, principalmente del sur. Los produc-tores juninenses fueron quienes, en continuidad con la actividad productiva de los padres, tuvieron acceso a la propiedad de la tierra, mientras que los productores que llegan de otras zonas son arrendatarios. Sin embar-go, los productores bolivianos desarrollaron la estrategia productiva e innovadora de la producción de horta-lizas bajo cubierta. Otra estrategia desarrollada por estos productores corresponde a la organización familiar exclusiva del trabajo productivo. Del total de los productores, el 56% realiza venta directa en quinta, el 28% vende en ferias y el 76% en verdulerías locales. Sin embargo, la principal problemática percibida correspon-de a los bajos precios del mercado y a la competencia. En este sentido, se considera que el sector hortícola del partido de Junín tiene potencial para el desarrollo de políticas públicas vinculadas a la comercialización y abastecimiento local de productos hortícolas frescos.
Palabras clave: productores hortícolas, periurbano, perfiles socioproductivos, estrategias.
1Instituto Nacional de Tecnología Agropecuaria (INTA), Instituto de Investigación y DesarrolloTegnológico para la Agrigultura Familiar Región Pampeana (IPAF Región Pampeana), Calle 403 s/n, Villa Elisa, Buenos Aires, Argentina. Correo electrónico: [email protected] Nacional de Tecnología Agropecuaria (INTA), Instituto de Investigación y Desarrollo Tecnológico para la Agricultura Familiar Región Pampeana (IPAF Región Pampeana), Calle 403 s/n, Villa Elisa, Buenos Aires, Argentina. Correo electrónico: [email protected] Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Pergamino, Agencia de Extensión Rural (AER), Junín, Brasil N° 149, Junín, Buenos Aires, Argentina. Correo electrónico: [email protected] Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Pergamino, Av. Frondizi, Ruta 32, km 4,5, Pergamino, Buenos Aires. Correo electrónico: [email protected] Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Pergamino, Av. Frondizi Ruta 32 km 4,5, Pergamino, Buenos Aires. Correo electrónico: [email protected] Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Pergamino, Agencia de Extensión Rural (AER) Junín, Brasil N° 149, Junín, Buenos Aires. Argentina.7Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA), Pergamino, Agencia de Extensión Rural (AER) Junín, Brasil N° 149, Junín, Buenos Aires, Argentina. Correo electrónico: [email protected] de Junín (Economía y Producción), España 37, Buenos Aires, Argentina. Correo electrónico: producció[email protected]
MUZI, M.E.1; PRIVIDERA, G.2; MELILLI, P.3; BERIBE, M.J.4; MONSUTTI, M.V.5; TELLERIA, M.G.6; FERNANDEZ, L.7; MECCIA, J.8
Recibido 17 de abril de 2018 // Aceptado 05 de febrero de 2020 // Publicado online 14 de octubre de 2020

MUZI, M.E.; PRIVIDERA, G.; MELILLI, P.; BERIBE, M.J.; MONSUTTI, M.V.; TELLERIA, M.G.; FERNANDEZ, L.; MECCIA, J.
Agosto 2020, Argentina 219219
ABSTRACT
The district of Junín is located in the northwest of Buenos Aires Province, covers 226337 hectares and has 90305 inhabitants. Junín city was historically configured as the main administrative, tourist, educational, in-dustrial, health and commercial center of the region. Due to the aptitude of the soils, it is located in the core of agricultural production zone. Since 1990, new socio-productive profiles have emerged in a large number of intermediate agro-cities, not only in the extensive field sector, but also in peri-urban areas. Horticultural producers are some of the most characteristic, but their features and potential are still unknown. The aim of this work is to carry out a socio-productive characterization of horticultural producers in Junín. In order to do this, a methodological device was elaborated based on the realization of a survey with social and productive variables. The methodology consists in a total scan of horticultural producers in Junín. This totality is reached from the creation of a list of producers made with different institutions. In order to confirm the total cover of producers, to corroborate the scan, a photointerpretation of google earth was used. The main results show that 60% of the producers are argentine, generally from Junín; and 40% are bolivian, mainly from the south. Producers born in Junín are those who, in continuity with the productive activity of the parents, had access to land ownership, while the immigrant producers are tenants. However, Bolivian producers developed producti-ve and innovative strategies of the production of vegetables under cover. Another strategy developed by these producers corresponds to the exclusive family organization of social-productive work. 56% of all producers make direct sales in the horticultural farm, 28% sell in fairs and 76% in local greengrocers. However, the main problem perceived corresponds to low market prices and competition. In this sense, it is considered that horti-cultural sector of Junín has potential for the development of public policies linked to the commercialization and local supply of fresh horticultural products.
Keywords: horticultural producers, peri-urban, socio-productive profiles, strategies.
INTRODUCCIÓN
El partido de Junín se ubica en el noroeste de la provin-cia de Buenos Aires, a 260 km de Capital Federal y abarca una superficie aproximada de 226.337 ha (Tauber, 1996). Cuenta con una población de 90305 habitantes, de los cua-les el 97% es población urbana (CNPHyV, 2010). Junín es la ciudad cabecera del partido y se constituyó como un nú-cleo regional con autonomía y dinámica propia y una gran oferta de servicios públicos y privados. Su rol se consolida por poseer una universidad nacional, nuclear la adminis-tración pública de los organismos de gobierno y judiciales, ser centro de comercialización y consumo, de provisión de insumos y servicios de la región. Tiene una estratégica ac-cesibilidad y conectividad hacia el oeste, norte y sur, hacia las grandes ciudades y puertos del país. En este sentido, el partido se ubica en un lugar privilegiado y altamente com-petitivo para la producción agropecuaria.
Las transformaciones en el uso del suelo vinculadas al proceso de agriculturización comienzan con la Revolución Verde y se profundizan en la década de 1970. Durante la década de 1990, el aumento de los precios internacionales de los cereales y oleaginosas se combinó con la creciente demanda internacional de soja y la liberación al mercado nacional de la semilla Roundup Ready. Durante este pro-ceso se generaliza dicho cultivo, disminuyendo la diversi-ficación agrícola y la actividad ganadera, desdibujando las particularidades locales que imprimían cierta dinámica par-ticular al espacio rural juninense.
Al mismo tiempo se profundiza un éxodo del campo como espacio de residencia permanente. A pesar de ha-berse incrementado la población total del partido desde el CNPHyV 1991 al 20101, este proceso se produce particu-larmente en las zonas urbanas, mientras que en las zonas rurales el proceso es inverso. En este sentido, la población urbana en Junín se fue incrementando hacia la periferia.
De este modo, nuevos perfiles socioproductivos surgie-ron en gran cantidad de agrociudades intermedias, como es el caso de Junín. El proceso no se percibe solamente en el sector de campo extensivo, sino también en las zo-nas periurbanas, donde se localizan productores hortíco-las. Según el informe agropecuario realizado por el Área de Asuntos Agropecuarios del Municipio de Junín en el año 2010, el partido concentra una incipiente actividad hortíco-la en paulatino crecimiento. La última información con la que se cuenta corresponde al Censo Hortiflorícola 2005 de la Provincia de Buenos Aires, en el que resulta que en el partido existen 26 establecimientos que corresponden a la actividad hortícola. Las quintas2 fueron localizadas en dos cuencas en la periferia capitalina: una en las inmediaciones del barrio San Antonio, al norte de la ciudad, y otra cercana al barrio de Villa del Parque, al sudeste de la ciudad. En
1 La variación poblacional consiste en un crecimiento del 1,85%. 2 En este trabajo se utilizará la denominación de “quintas” y “esta-blecimientos hortícolas” de manera indistinta. Se corresponde con el modo de denominación local.

Caracterización socioproductiva de los productores hortícolas en el partido de Junín (Buenos Aires)
ARTÍCULOS RIA / Vol. 46 / N.º 2220220
una localización más dispersa, le sigue el corredor de la ruta 7. Se estima que el sector hortícola juninense tiene un gran potencial socioproductivo vinculado al abastecimiento local y a la generación de espacios de comercialización, pero la falta de información actualizada implica dificulta-des para realizar cualquier diagnóstico. En lo que respecta particularmente al partido de Junín, tomando en cuenta los trabajos de investigación vinculados a las migraciones de productores hortícolas hacia y en el país (Benencia, 2012) se considera que, posterior a la fecha censada, se produce el asentamiento de productores nuevos en el partido.
Se inicia entonces un trabajo articulado entre el IPAF Re-gión Pampeana (CIPAF-INTA), la AER Junín, la EEA Per-gamino (CERBAN-INTA) y el municipio de Junín. A los fi-nes de lograr un conocimiento integral del sector hortícola, se conformó un equipo de investigación interdisciplinario en el que intervienen las ciencias agronómicas, sociales y estadísticas. El objetivo del trabajo consiste en realizar una caracterización socioproductiva del sector hortícola del partido de Junín, con la finalidad de conocer su potenciali-dad socioproductiva.
MATERIALES Y MÉTODOS
Para lograr los objetivos planteados se diseñó una en-cuesta en la que se relevan aspectos sociales y produc-tivos. La metodología consiste en un barrido total de los productores hortícolas del partido de Junín, con la finalidad de conocer sus características socioproductivas y las po-
Figura 1. Localización de los establecimientos hortícolas en el partido de Junín.Fuente: Elaboración propia en Google Earth.
tencialidades presentes en el territorio.
En este sentido, a partir de la conformación de un lista-do total de productores, mediante informantes clave, en el que participaron las diferentes instituciones, se realizó un operativo de campo visitando por zonas a cada uno de los productores para realizar la encuesta. Para corroborar el barrido total de los productores, se utilizó la fotointerpreta-ción de Google Earth, dado que la producción hortícola a campo presenta un patrón distinguible.
El relevamiento se realizó durante el mes de mayo, junio y julio de 2017. Se han encuestado 25 productores, loca-lizándose 25 quintas, una menos de las relevadas en el Censo Horti-florícola del 20053. El procesamiento de datos se realizó en una matriz, permitiendo un análisis cuantitati-vo descriptivo del sector.
RESULTADOS
Territorio, residencia y tradición
A partir de la georreferenciación de los establecimientos de los productores hortícolas, se identifica que hay dos nú-cleos de localización de las quintas alrededor de la ciudad de Junín.
3 Nuestro relevamiento no toma la base de datos del Censo Horti-florícola 2005. Se trata de una base propia. Este trabajo no tiene por objetivo dar cuenta de la continuidad ni desaparición de las quintas de aquel censo.
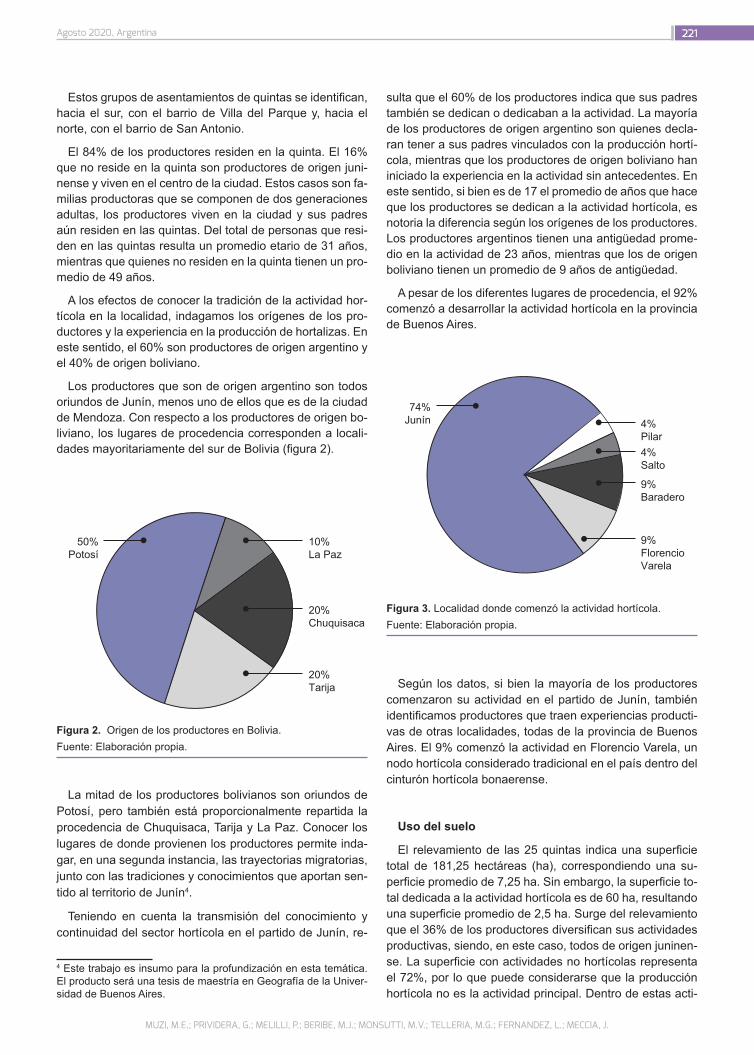
MUZI, M.E.; PRIVIDERA, G.; MELILLI, P.; BERIBE, M.J.; MONSUTTI, M.V.; TELLERIA, M.G.; FERNANDEZ, L.; MECCIA, J.
Agosto 2020, Argentina 221221
Figura 2. Origen de los productores en Bolivia.Fuente: Elaboración propia.
Figura 3. Localidad donde comenzó la actividad hortícola.Fuente: Elaboración propia.
Estos grupos de asentamientos de quintas se identifican, hacia el sur, con el barrio de Villa del Parque y, hacia el norte, con el barrio de San Antonio.
El 84% de los productores residen en la quinta. El 16% que no reside en la quinta son productores de origen juni-nense y viven en el centro de la ciudad. Estos casos son fa-milias productoras que se componen de dos generaciones adultas, los productores viven en la ciudad y sus padres aún residen en las quintas. Del total de personas que resi-den en las quintas resulta un promedio etario de 31 años, mientras que quienes no residen en la quinta tienen un pro-medio de 49 años.
A los efectos de conocer la tradición de la actividad hor-tícola en la localidad, indagamos los orígenes de los pro-ductores y la experiencia en la producción de hortalizas. En este sentido, el 60% son productores de origen argentino y el 40% de origen boliviano.
Los productores que son de origen argentino son todos oriundos de Junín, menos uno de ellos que es de la ciudad de Mendoza. Con respecto a los productores de origen bo-liviano, los lugares de procedencia corresponden a locali-dades mayoritariamente del sur de Bolivia (figura 2).
sulta que el 60% de los productores indica que sus padres también se dedican o dedicaban a la actividad. La mayoría de los productores de origen argentino son quienes decla-ran tener a sus padres vinculados con la producción hortí-cola, mientras que los productores de origen boliviano han iniciado la experiencia en la actividad sin antecedentes. En este sentido, si bien es de 17 el promedio de años que hace que los productores se dedican a la actividad hortícola, es notoria la diferencia según los orígenes de los productores. Los productores argentinos tienen una antigüedad prome-dio en la actividad de 23 años, mientras que los de origen boliviano tienen un promedio de 9 años de antigüedad.
A pesar de los diferentes lugares de procedencia, el 92% comenzó a desarrollar la actividad hortícola en la provincia de Buenos Aires.
10%La Paz
50%Potosí
20%Chuquisaca
20%Tarija
4%Pilar4%Salto
74%Junín
9%Baradero
9%FlorencioVarela
La mitad de los productores bolivianos son oriundos de Potosí, pero también está proporcionalmente repartida la procedencia de Chuquisaca, Tarija y La Paz. Conocer los lugares de donde provienen los productores permite inda-gar, en una segunda instancia, las trayectorias migratorias, junto con las tradiciones y conocimientos que aportan sen-tido al territorio de Junín4.
Teniendo en cuenta la transmisión del conocimiento y continuidad del sector hortícola en el partido de Junín, re-
4 Este trabajo es insumo para la profundización en esta temática. El producto será una tesis de maestría en Geografía de la Univer-sidad de Buenos Aires.
Según los datos, si bien la mayoría de los productores comenzaron su actividad en el partido de Junín, también identificamos productores que traen experiencias producti-vas de otras localidades, todas de la provincia de Buenos Aires. El 9% comenzó la actividad en Florencio Varela, un nodo hortícola considerado tradicional en el país dentro del cinturón hortícola bonaerense.
Uso del suelo
El relevamiento de las 25 quintas indica una superficie total de 181,25 hectáreas (ha), correspondiendo una su-perficie promedio de 7,25 ha. Sin embargo, la superficie to-tal dedicada a la actividad hortícola es de 60 ha, resultando una superficie promedio de 2,5 ha. Surge del relevamiento que el 36% de los productores diversifican sus actividades productivas, siendo, en este caso, todos de origen juninen-se. La superficie con actividades no hortícolas representa el 72%, por lo que puede considerarse que la producción hortícola no es la actividad principal. Dentro de estas acti-

Caracterización socioproductiva de los productores hortícolas en el partido de Junín (Buenos Aires)
ARTÍCULOS RIA / Vol. 46 / N.º 2222222
vidades, la actividad agrícola compromete el 67% de la su-perficie, el 5% restante está representado por un productor que desarrolla la actividad porcina y otro que no especifica la actividad. Ninguno de estos productores que combinan acti-vidades productivas tiene superficie bajo cubierta para hor-ticultura, es decir, que quienes tienen invernáculo son aque-llos productores que solo se dedican a la actividad hortícola.
Para abordar la producción hortícola, consideramos la producción a campo y bajo cubierta. En este sentido, en-contramos que el 100% de los productores tienen produc-ción de hortalizas a campo y que un 32%, además, realiza producción bajo cubierta.
Desde la década de 1990 se desarrollan trabajos de in-vestigación situados en diferentes espacios geográficos a lo largo del país que dan cuenta de la implementación de la modalidad del cultivo bajo cubierta a partir del creciente uso de invernáculos por parte de los productores de origen boliviano (Benencia, (1994, 1997, 2005 y 2012); Benencia y Geymonat, (2005); García, (2010); Lorda, (2015) y Rivas y Natera Rivas, (2007)).
En este sentido, al analizar las características de los pro-ductores en Junín, notamos que la práctica hortícola bajo cubierta está generalizada solamente entre los productores de origen boliviano. De la totalidad de los productores hor-tícolas de origen boliviano en Junín, un 70% realiza pro-ducción en invernáculos comprometiendo una superficie cubierta total de 43000 m2.
La superficie media destinada a la producción hortícola a campo en Junín es de 2,2 ha por productor.
Respecto de las especies producidas en el partido, los productores cultivan 10 tipos en promedio. Más del 55% de los productores siembran: acelga, achicoria, berenjena, ce-
Figura 4. Tenencia de la tierra.Fuente: Elaboración propia.
Figura 5. Canales de venta.Fuente: Elaboración propia.
bolla de verdeo, espinaca, lechuga, rúcula, tomate redondo y zapallito de tronco.
Tenencia de la tierra
Si se analiza el régimen de tenencia, encontramos bási-camente quintas en propiedad y en arrendamiento.
Si tenemos en cuenta el régimen de propiedad, resulta que el 52% de los productores son propietarios (todos de origen juninense) y ocupan el 89% de la superficie total. El tamaño promedio de los establecimientos es de 12,40 ha, con una superficie mínima de 3 ha y máxima de 100 ha.
Si consideramos la superficie bajo arrendamiento, se encuentra menos proporción de tierra (10,8%) en casi la misma cantidad de productores (44%). Así, el tamaño pro-medio es de 1,8 ha, presentándose establecimientos hor-tícolas con una superficie mínima de 1,5 ha y máxima de 3 ha. En este caso, quienes trabajan en tierras arrendadas son los productores provenientes de otras zonas.
En todos los casos se trata del tipo jurídico “Persona física”, no encontramos cooperativas, sociedades u otras figuras jurídicas colectivas.
Comercialización y mercados
Respecto de la comercialización, aproximadamente la mitad de los productores maneja solo un canal de venta, mayoritariamente verdulerías locales, mientras que la otra mitad de productores desarrolla cierta estrategia con dos o tres canales de venta.
Del total de los productores, el 56% realiza venta directa en quinta, el 28% vende en ferias y el 76% en verdule-rías locales. Estos porcentajes no son excluyentes, puede tratarse de canales de venta combinados. Es interesante notar que el 100% de los productores de origen boliviano utiliza como canal de venta las verdulerías locales, en algu-nos casos combinan con la venta directa y las ferias. Esto
Superficie (ha) Cant. quintas
Propiedad Arrendamiento Préstamo
89,1%
52%44%
4%0,14%
10,76%
16%3 canales de venta
48%1 canal
de venta
36%2 canales de venta

MUZI, M.E.; PRIVIDERA, G.; MELILLI, P.; BERIBE, M.J.; MONSUTTI, M.V.; TELLERIA, M.G.; FERNANDEZ, L.; MECCIA, J.
Agosto 2020, Argentina 223223
se da de manera inversa en el caso de los productores de origen juninense, quienes utilizan la venta directa y las ferias como canales de venta principales y solo algunos de ellos también venden en verdulerías.
Organización social del trabajo
Teniendo en cuenta el factor de producción “Trabajo”, hay trabajadores familiares en todos los establecimientos rele-vados, con un promedio de 2 trabajadores familiares. Del relevamiento resulta que existe muy poca contratación de mano de obra. Ningún productor boliviano contrata mano de obra para las labores. Solo el 8% contrata mano de obra permanente y el 16% trabajadores transitorios. En el 60% de los casos donde existe contratación de mano de obra asa-lariada se trata de establecimientos que también se dedican a la actividad agrícola. Solo uno de los casos es particular, dado que se trata de una quinta que se dedica exclusiva-mente a la actividad hortícola, con 20000 m2 de superficie bajo cubierta. Este caso además es el único que combina la contratación de mano de obra permanente y transitoria.
El 28% de los establecimientos terceriza servicios de gestión/administración.
Composición social de los hogares
La población total en los hogares correspondientes a los establecimientos es de 87 personas y el promedio por hogar es de 3,5 personas (similar al promedio nacional de hogares urbanos y rurales según el CNPHyV 2010). Sin embargo, si analizamos los hogares por procedencia, encontramos que los hogares de origen boliviano son más numerosos, resul-tando un promedio de 5 miembros por hogar con un prome-dio etario de 20 años. Mientras que los hogares de origen argentino tienen un promedio de 2,5 personas (la mitad) con un promedio etario de 52 años (más del doble).
Del total de los miembros de los hogares, el 55% realiza trabajos en la quinta.
Figura 6. Tipo de trabajo.Fuente: Elaboración propia.
Figura 7. Género y tipo de trabajo.Fuente: Elaboración propia.
De las personas que trabajan en quinta, encontramos que existe mayor participación en las tareas físicas y ad-ministrativas exclusivas. Las tareas se vuelven administra-tivas a medida que aumenta el promedio de edad. En este sentido, resulta un promedio de 39 años en las tareas pu-ramente físicas, de 44 en las mixtas y 68 en las puramente administrativas. El total de las tareas las realizan el 67% personas de sexo masculino y el 33% de sexo femenino.
En general, las tareas administrativas corren por cuenta de personas de sexo masculino, son los “hombres mayo-res del hogar los que llevan las cuentas”, mientras que las mujeres se dedican a trabajar en las labores de la quinta.
Respecto de la composición de ingresos de los hogares, el 92% considera que la fuente principal de ingresos es el establecimiento. A pesar de ello, encontramos que el 56% de los hogares tiene asignaciones estatales: en el caso de los hogares de origen boliviano se destaca la Asignación Universal por Hijo, mientras que en los hogares de origen argentino prevalecen las jubilaciones.
Intereses y problemáticas
Respecto de los intereses que expresan los productores y considerando que no son excluyentes, de la totalidad de los productores el 56% menciona el interés en asesorar-se comercialmente, el 84% en recibir capacitación técnica/productiva y el 84% considera importante capacitarse en sanidad y manejo de plagas.
El 56% de los productores mencionó tener algún tipo de problemática vinculada a la producción y comercialización.
4%Tareas físicasy administrativas
56%Tareas
administrativas
40%Tareas físicas
Femenino Masculino
Tareasfísicas
Tareasadministrativas
Tareasfísicas y
administrativas
96%32%
68%
4%
100%

Caracterización socioproductiva de los productores hortícolas en el partido de Junín (Buenos Aires)
ARTÍCULOS RIA / Vol. 46 / N.º 2224224
Respecto del tipo de problemática percibida por los pro-ductores, la mitad menciona los bajos precios del mercado y la competencia como problemática principal.
DISCUSIÓN Y CONCLUSIONES
Considerando el Censo Hortiflorícola (2005) y teniendo en cuenta este relevamiento, se observa que en 12 años la cantidad de productores se mantuvo, aunque aún se des-conoce si efectivamente son los mismos o hubo transfor-maciones o migraciones en el sector, objetivo planteado para una segunda etapa del trabajo de investigación.
En términos generales pueden percibirse diferentes con-dicionamientos estructurales y prácticas culturales vincu-ladas a los orígenes de los productores. Los productores oriundos de Junín conllevan una tradición local, tienen a sus padres vinculados a la actividad hortícola y agrícola en general, los cuales viven, en su mayoría, en la ciudad de Junín, mientras que los productores bolivianos se han iniciado más recientemente en la actividad y se especiali-zan en horticultura. Esto podría estar vinculado al proceso migratorio desde el país de origen. La trayectoria y la per-tenencia espacial local, pareciera estar relacionada con la propiedad de la tierra de los productores juninenses. Sin embargo, a pesar que los productores oriundos de otros si-tios son quienes arriendan las tierras que trabajan, la prác-tica del cultivo bajo cubierta es una innovación llegada a Junín como parte de la experiencia que migra junto con los productores de origen boliviano. Por un lado, esta práctica tiene una consecuencia económica vinculada a una mejora en sus ingresos. Pero también, a pesar de no ser propieta-rios de la tierra que trabajan, la construcción de los inver-náculos para la producción se relaciona con cierto arraigo territorial vinculado particularmente al lugar.
Por otro lado, se perciben ciertas diferencias vinculadas a las estrategias de la organización social del trabajo en la unidad productiva. Los hogares de productores de origen boliviano tienen más cantidad de miembros y, a la vez, son
más jóvenes. Al mismo tiempo, ningún productor boliviano contrata mano de obra asalariada para las labores, estas pueden ser realizadas por los miembros de la familia.
Dado que las familias que integran los establecimientos de productores juninenses son mayores en edad y que las tareas se vuelven administrativas a medida que aumenta el promedio etario, las tareas físicas necesitan ser realizadas por mano de obra extrafamiliar contratada.
Resulta del trabajo de análisis que para la mayoría de los entrevistados la producción es su principal fuente de ingre-so y que perciben como una importante problemática los bajos precios del mercado y la competencia, generar es-pacios de comercialización donde no se encuentren com-petencias, donde se respete el precio de los productores sin intermediarios, puede ser una política pertinente para el sector hortícola local.
Teniendo en cuenta las diferentes características de los productores, nos preguntamos, ¿pueden percibirse diferen-tes perfiles sociales y productivos?, ¿cómo pueden com-prenderse en el territorio en relación con sus trayectorias? En este sentido, considerando los resultados y reflexiones de este análisis de corte sincrónico, interesa continuar una investigación con cierta profundidad temporal.
AGRADECIMIENTOS
Los autores agradecen la colaboración de Marina Poggi, de los productores hortícolas de Junín y al financiamiento del PN-SEPT 1129022 (INTA) y del PRET BANOR 1271103 (INTA).
BIBLIOGRAFÍABENENCIA, R. 1994. La horticultura bonaerense: lógicas pro-
ductivas y cambios en el mercado de trabajo. Desarrollo Econó-mico, 53-73.
BENENCIA, R. 1997. De peones a patrones quinteros. Movi-lidad social de familias bolivianas en la periferia bonaerense, en Estudios Migratorios Latinoamericanos V.12, n.o 35, CEMLA, Bue-nos Aires, 63-102.
BENENCIA, R. 2005. Producción, trabajo y nacionalidad: confi-guraciones territoriales de la producción hortícola del cinturón ver-de bonaerense. Tesis doctoral. Facultad de Ciencias Económicas. Universidad de Buenos Aires. Bueno Aires.
BENENCIA, R. 2012. Participación de los inmigrantes bolivia-nos en espacios específicos de la producción hortícola en la Ar-gentina/Migrations, Work and Agricultural Global Chains. Política y Sociedad, 49(1), 163.
BENENCIA, R.; GEYMONAT, M. 2005. Migración transnacional y redes sociales en la creación de territorios productivos en la Ar-gentina. Río Cuarto, Córdoba. Cuadernos de desarrollo rural, (55).
CENSO HORTIFLORÍCOLA DE BUENOS AIRES (CHFBA’05). 2005. Ministerio de Asuntos Agrarios y Ministerio de Economía de la Provincia de Buenos Aires (Argentina) 2006.
GARCÍA, M. 2010. Inicios, consolidación y diferenciación de la horticultura platense. Agricultura periurbana en Argentina y globa-lización. Escenarios, recorridos y problemas, 69-85.
INFORME AGROPECUARIO. 2010. Gobierno Local, Produc-ción, Asuntos Agropecuarios. Junín. Buenos Aires.
Figura 8. Problemáticas.Fuente: Elaboración propia.
14%Manejo productivo de vecinos
15%Sanidad /manejo de plagas
7%Inseguridad
50%Precios
14%Consecuenciaspor clima

MUZI, M.E.; PRIVIDERA, G.; MELILLI, P.; BERIBE, M.J.; MONSUTTI, M.V.; TELLERIA, M.G.; FERNANDEZ, L.; MECCIA, J.
Agosto 2020, Argentina 225225
INSTITUTO NACIONAL DE ESTADÍSTICAS Y CENSOS (IN-DEC). 1991, 2001 y 2010. Censo Nacional de Población y Vivien-da. República Argentina.
LORDA, M.A. 2015. La comprensión del territorio a partir del modelo de formación socio espacial desde la práctica de la horti-cultura en el periurbano de Bahía Blanca, Argentina. InterEspaço: Revista de Geografia e Interdisciplinaridade, 1(3), 32-55.
RIVAS, A.I.; NATERA RIVAS, J.J. 2007. Inserción de la inmi-gración boliviana en la actividad hortícola del Departamento Lules (Tucumán, Argentina) a mediados de la década de los noventa. Cuadernos Geográficos, (41).
TAUBER, F. 1996. Reflexiones y datos para una estrategia de desarrollo. Junín. Municipalidad. Buenos Aires.

¿Cómo ha cambiado la precipitación en la provincia de Santa Fe?
ARTÍCULOS RIA / Vol. 46 / N.º 2226226
¿Cómo ha cambiado la precipitaciónen la provincia de Santa Fe?
RESUMENLa provincia de Santa Fe concentra un gran porcentaje de la producción agroindustrial de Argentina. En los
últimos años, los eventos extremos relacionados con excesos y déficits de precipitación han provocado ane-gamientos prolongados y severas sequías impactando gravemente la estructura agroindustrial en la provincia. Estos eventos pueden estar influenciados por cambios en la precipitación que se produjeron a partir de me-diados del siglo xx. Este trabajo investiga la variabilidad y los cambios en la precipitación y sus extremos en las distintas regiones de la provincia de Santa Fe. Para ello se utilizaron datos observados con registros pro-longados y calidad controlada. Se aplicó un Análisis Espectral Singular para examinar la variabilidad temporal de la precipitación anual y los eventos extremos. Los eventos extremos se analizaron a través de los valores máximos anuales de precipitación diaria y de precipitación acumulada en 5 días consecutivos, el índice simple de intensidad diaria y los días secos consecutivos. Los resultados muestran un incremento de la precipitación anual a partir de la década de 1960 y un aumento considerable de la frecuencia y magnitud de los eventos de lluvias intensas en toda la provincia. Además, la precipitación está influenciada por el fenómeno El Niño-Oscilación del Sur (ENOS) con frecuencias de entre 2 y 6 años. En las estaciones estudiadas de las regiones norte y centro, la precipitación anual aumentó un 15% desde la década de 1960 en comparación con la pri-mera mitad del siglo xx, aunque las tendencias crecientes parecen haberse estabilizado a partir de 1990. En contraposición, los días secos consecutivos han aumentado de manera significativa en las últimas décadas. En las estaciones analizadas de las regiones centro-sur y sur se dio un incremento de la precipitación anual y las lluvias intensas con tendencias crecientes hasta la actualidad. Estas tendencias podrían provocar en los próximos años un incremento aún mayor de las lluvias intensas de gran magnitud. Estos resultados sugieren que habrá menor cantidad de días con lluvias de mayor intensidad y mayor cantidad de agua, dando lugar a excesos hídricos frecuentes en toda la provincia. Además, se producirán déficits estacionales con mayor frecuencia principalmente en el norte de la provincia.
Palabras clave: variabilidad climática, cambio climático, lluvias intensas, sequías.
ABSTRACT
The province of Santa Fe concentrates a large percentage of Argentina’s agroindustrial production. In recent years, extreme events related to precipitation excesses and deficits have caused extended waterlogging and large droughts, severely affecting the agroindustry in the province. These extreme events may be influenced by changes in precipitation that occurred since mid-twentieth century. This study investigates the variability
1Centro de Estudios de Variabilidad y Cambio Climático (CEVARCAM), Facultad de Ingeniería y Ciencias Hídricas (FICH), Universidad Na-cional del Litoral (UNL), Santa Fe, Argentina. Correo electrónico: [email protected] Nacional de Investigaciones Científicas y Técnicas (CONICET).
LOVINO, M.A.1; MÜLLER, G.V.1,2; SGROI, L.C.1
Recibido 26 de marzo de 2019 // Aceptado 02 de marzo de 2020 // Publicado online 26 de agosto de 2020

LOVINO, M.A.; MÜLLER, G.V.; SGROI, L.C.
Agosto 2020, Argentina 227227
and changes in precipitation and its extremes in different regions of the province of Santa Fe. To fulfill this aim, we used observed data with long records and controlled quality. A Singular Spectral Analysis was applied to examine the temporal variability of annual precipitation and extreme events. The extreme events were cha-racterized by the maximum annual daily precipitation, the maximum amount of precipitation accumulated in 5 consecutive days per year, the simple daily intensity index and the consecutive dry days. The results show an increase in annual precipitation since the 1960s and a considerable rise in the frequency and magnitude of heavy rainfall throughout the province. In addition, the El Niño Southern Oscillation influence precipitation with frequencies between 2 and 6 years. In the studied stations of the northern and central regions, the annual rainfall registered an average increase of 15% since 1960 compared with the first half of the twentieth century, although upward trends seem to have stabilized after 1990. In contrast, consecutive dry days have raised sig-nificantly in recent decades. In the stations of the central-southern and southern regions, there was an increa-se in annual precipitation and heavy rainfall with upward trends to date. If these trends continue rising, they will generate an even greater increase in heavy rains of great magnitude in the coming years. These findings suggest that precipitation will be concentrated in fewer days with rain of greater intensity and quantity of water for each event, leading to water excesses in the whole province. Furthermore, seasonal deficits will occur more frequently, mainly in the north of the province.
Keywords: climate variability, climate change, heavy rainfalls, droughts.
INTRODUCCIÓN
La Pampa húmeda tiene una gran importancia econó-mica y demográfica en Sudamérica y es clave para la se-guridad alimentaria de la región y del mundo. La provincia de Santa Fe, como centro neurálgico de la región núcleo agrícola ganadera, desempeña un rol estratégico a nivel nacional ya que concentra un gran porcentaje de la produc-ción agroindustrial del país. En este contexto, el clima tiene un protagonismo destacado, dado que impacta en dicha producción en forma directa.
El cambio climático es un cambio en el estado medio del clima o en su variabilidad, que se manifiesta por una ten-dencia en las variables climáticas durante un prolongado período (en general, se adopta un período de 30 años como período normal de referencia, definición de IPCC, 2013). En todo el planeta, los extremos de temperatura han cam-biado hacia condiciones más cálidas durante los últimos 60 años y las lluvias intensas aumentaron en una fracción mayor de la superficie terrestre que en la que decrecieron, aunque ampliamente influenciados por variaciones regio-nales y subregionales y variabilidad en escalas temporales de interanual a decadal (Seneviratne et al., 2012; Donat et al., 2013). En el sudeste de Sudamérica, consistentemen-te con los cambios globales, los eventos hidroclimáticos tienden hacia condiciones más cálidas y húmedas ya que aumentó la magnitud y la frecuencia de extremos de tem-peratura y de precipitaciones intensas (Rusticucci, 2012; Cavalcanti et al., 2015; Carril et al., 2016). En esta región, los incrementos de la precipitación (especialmente lluvias intensas) y de los caudales contribuyeron, junto con fac-tores antrópicos, a aumentar la frecuencia de crecidas e
inundaciones que afectaron la productividad agrícola y ga-nadera e impactaron los asentamientos urbanos y rurales (Magrin et al., 2014).
En el nordeste argentino se produjo un importante cambio climático a partir de mediados del siglo xx con un considera-ble aumento de la precipitación en toda la región (Lovino et al., 2014; Lovino et al., 2018). Este cambio podría haber es-tado influenciado por condiciones antrópicas y por variabili-dad decadal natural. Entre los forzantes naturales, se puede citar el enfriamiento del océano Atlántico tropical en las últi-mas décadas y el calentamiento del océano Pacífico tropical que podrían haber favorecido las condiciones más húme-das (Seager et al., 2010; Jacques-Coper y Garreaud, 2015). También, dentro de los forzantes antrópicos, es posible que los cambios de uso de suelo hayan reducido la evapotrans-piración y favorecido el escurrimiento superficial, induciendo un aumento de los caudales (Doyle y Barros, 2011).
Los impactos de los extremos climáticos y el potencial de desastres se acrecientan en las regiones con alta exposición y vulnerabilidad, pudiendo exacerbarse aún más ante un contexto complejo de variabilidad y cambio climático (IPCC, 2014). La provincia de Santa Fe es una de estas regiones ya que sus actividades agroindustriales están influenciadas por un complejo sistema de recursos hídricos superficiales en una gran llanura con escasas pendientes, concentra im-portantes centros urbanos y está afectada por variabilidad y cambios en la precipitación que conllevan a frecuentes y severas sequías e inundaciones (Lovino et al., 2018).
Dado el contexto enunciado, este trabajo se propone investigar la variabilidad y los cambios observados en la precipitación y sus extremos en las distintas regiones de
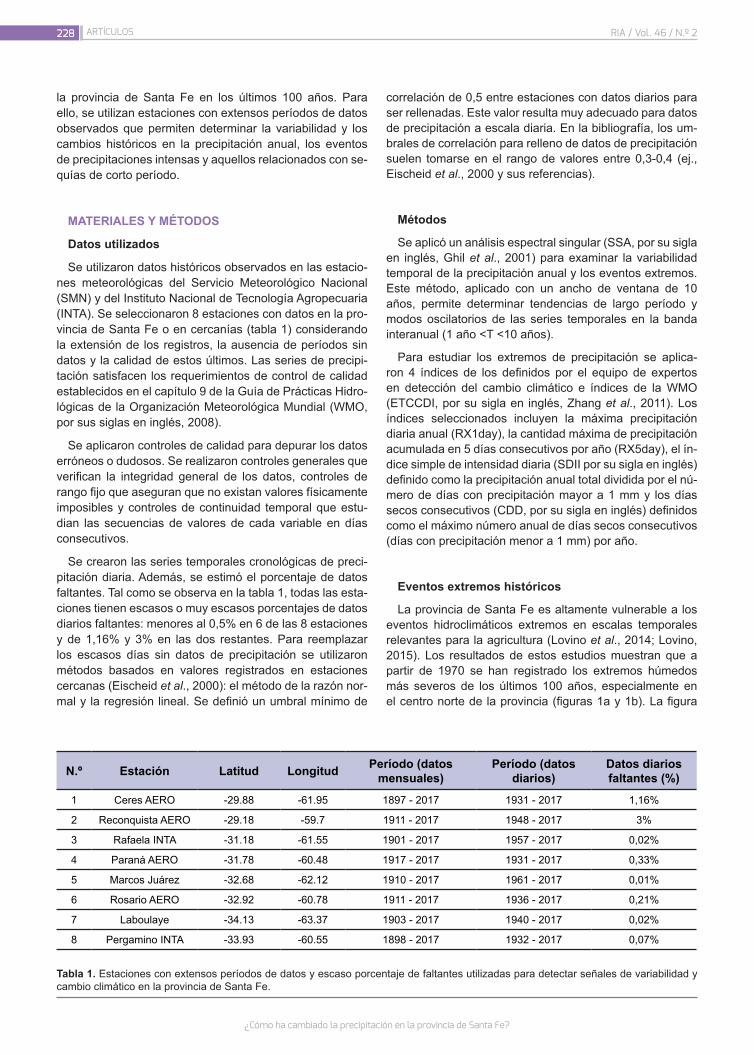
¿Cómo ha cambiado la precipitación en la provincia de Santa Fe?
ARTÍCULOS RIA / Vol. 46 / N.º 2228228
correlación de 0,5 entre estaciones con datos diarios para ser rellenadas. Este valor resulta muy adecuado para datos de precipitación a escala diaria. En la bibliografía, los um-brales de correlación para relleno de datos de precipitación suelen tomarse en el rango de valores entre 0,3-0,4 (ej., Eischeid et al., 2000 y sus referencias).
Métodos
Se aplicó un análisis espectral singular (SSA, por su sigla en inglés, Ghil et al., 2001) para examinar la variabilidad temporal de la precipitación anual y los eventos extremos. Este método, aplicado con un ancho de ventana de 10 años, permite determinar tendencias de largo período y modos oscilatorios de las series temporales en la banda interanual (1 año <T <10 años).
Para estudiar los extremos de precipitación se aplica-ron 4 índices de los definidos por el equipo de expertos en detección del cambio climático e índices de la WMO (ETCCDI, por su sigla en inglés, Zhang et al., 2011). Los índices seleccionados incluyen la máxima precipitación diaria anual (RX1day), la cantidad máxima de precipitación acumulada en 5 días consecutivos por año (RX5day), el ín-dice simple de intensidad diaria (SDII por su sigla en inglés) definido como la precipitación anual total dividida por el nú-mero de días con precipitación mayor a 1 mm y los días secos consecutivos (CDD, por su sigla en inglés) definidos como el máximo número anual de días secos consecutivos (días con precipitación menor a 1 mm) por año.
Eventos extremos históricos
La provincia de Santa Fe es altamente vulnerable a los eventos hidroclimáticos extremos en escalas temporales relevantes para la agricultura (Lovino et al., 2014; Lovino, 2015). Los resultados de estos estudios muestran que a partir de 1970 se han registrado los extremos húmedos más severos de los últimos 100 años, especialmente en el centro norte de la provincia (figuras 1a y 1b). La figura
Tabla 1. Estaciones con extensos períodos de datos y escaso porcentaje de faltantes utilizadas para detectar señales de variabilidad y cambio climático en la provincia de Santa Fe.
la provincia de Santa Fe en los últimos 100 años. Para ello, se utilizan estaciones con extensos períodos de datos observados que permiten determinar la variabilidad y los cambios históricos en la precipitación anual, los eventos de precipitaciones intensas y aquellos relacionados con se-quías de corto período.
MATERIALES Y MÉTODOS
Datos utilizados
Se utilizaron datos históricos observados en las estacio-nes meteorológicas del Servicio Meteorológico Nacional (SMN) y del Instituto Nacional de Tecnología Agropecuaria (INTA). Se seleccionaron 8 estaciones con datos en la pro-vincia de Santa Fe o en cercanías (tabla 1) considerando la extensión de los registros, la ausencia de períodos sin datos y la calidad de estos últimos. Las series de precipi-tación satisfacen los requerimientos de control de calidad establecidos en el capítulo 9 de la Guía de Prácticas Hidro-lógicas de la Organización Meteorológica Mundial (WMO, por sus siglas en inglés, 2008).
Se aplicaron controles de calidad para depurar los datos erróneos o dudosos. Se realizaron controles generales que verifican la integridad general de los datos, controles de rango fijo que aseguran que no existan valores físicamente imposibles y controles de continuidad temporal que estu-dian las secuencias de valores de cada variable en días consecutivos.
Se crearon las series temporales cronológicas de preci-pitación diaria. Además, se estimó el porcentaje de datos faltantes. Tal como se observa en la tabla 1, todas las esta-ciones tienen escasos o muy escasos porcentajes de datos diarios faltantes: menores al 0,5% en 6 de las 8 estaciones y de 1,16% y 3% en las dos restantes. Para reemplazar los escasos días sin datos de precipitación se utilizaron métodos basados en valores registrados en estaciones cercanas (Eischeid et al., 2000): el método de la razón nor-mal y la regresión lineal. Se definió un umbral mínimo de
N.º Estación Latitud Longitud Período (datos mensuales)
Período (datos diarios)
Datos diarios faltantes (%)
1 Ceres AERO -29.88 -61.95 1897 - 2017 1931 - 2017 1,16%
2 Reconquista AERO -29.18 -59.7 1911 - 2017 1948 - 2017 3%
3 Rafaela INTA -31.18 -61.55 1901 - 2017 1957 - 2017 0,02%
4 Paraná AERO -31.78 -60.48 1917 - 2017 1931 - 2017 0,33%
5 Marcos Juárez -32.68 -62.12 1910 - 2017 1961 - 2017 0,01%
6 Rosario AERO -32.92 -60.78 1911 - 2017 1936 - 2017 0,21%
7 Laboulaye -34.13 -63.37 1903 - 2017 1940 - 2017 0,02%
8 Pergamino INTA -33.93 -60.55 1898 - 2017 1932 - 2017 0,07%
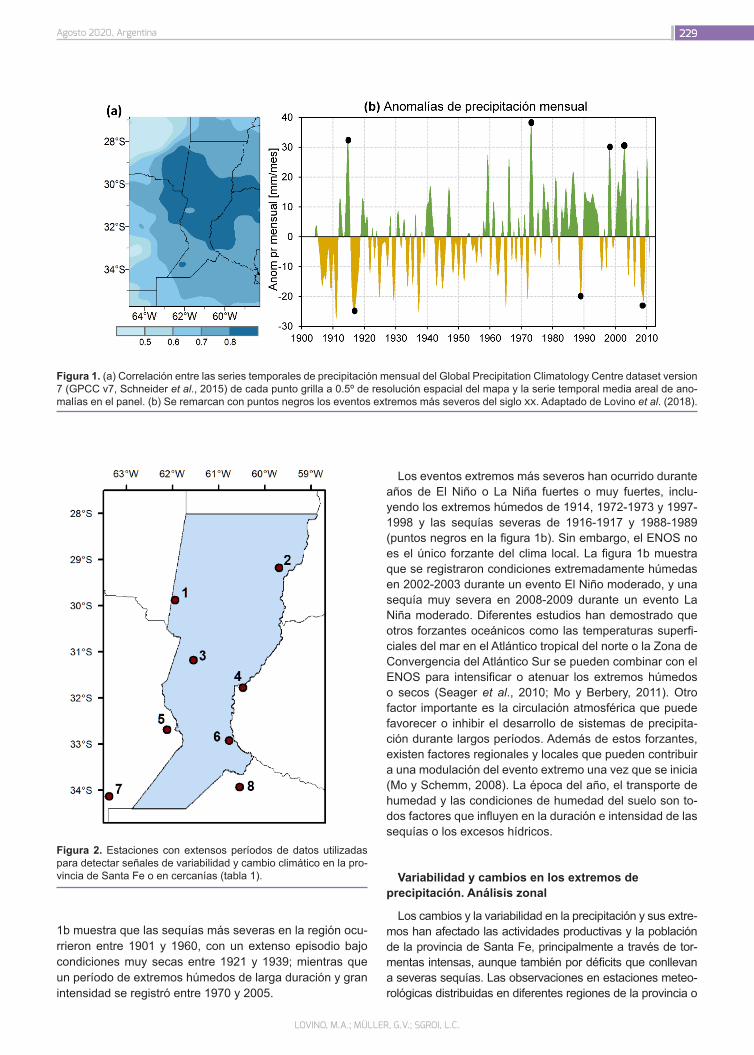
LOVINO, M.A.; MÜLLER, G.V.; SGROI, L.C.
Agosto 2020, Argentina 229229
Figura 1. (a) Correlación entre las series temporales de precipitación mensual del Global Precipitation Climatology Centre dataset version 7 (GPCC v7, Schneider et al., 2015) de cada punto grilla a 0.5º de resolución espacial del mapa y la serie temporal media areal de ano-malías en el panel. (b) Se remarcan con puntos negros los eventos extremos más severos del siglo xx. Adaptado de Lovino et al. (2018).
Figura 2. Estaciones con extensos períodos de datos utilizadas para detectar señales de variabilidad y cambio climático en la pro-vincia de Santa Fe o en cercanías (tabla 1).
1b muestra que las sequías más severas en la región ocu-rrieron entre 1901 y 1960, con un extenso episodio bajo condiciones muy secas entre 1921 y 1939; mientras que un período de extremos húmedos de larga duración y gran intensidad se registró entre 1970 y 2005.
Los eventos extremos más severos han ocurrido durante años de El Niño o La Niña fuertes o muy fuertes, inclu-yendo los extremos húmedos de 1914, 1972-1973 y 1997-1998 y las sequías severas de 1916-1917 y 1988-1989 (puntos negros en la figura 1b). Sin embargo, el ENOS no es el único forzante del clima local. La figura 1b muestra que se registraron condiciones extremadamente húmedas en 2002-2003 durante un evento El Niño moderado, y una sequía muy severa en 2008-2009 durante un evento La Niña moderado. Diferentes estudios han demostrado que otros forzantes oceánicos como las temperaturas superfi-ciales del mar en el Atlántico tropical del norte o la Zona de Convergencia del Atlántico Sur se pueden combinar con el ENOS para intensificar o atenuar los extremos húmedos o secos (Seager et al., 2010; Mo y Berbery, 2011). Otro factor importante es la circulación atmosférica que puede favorecer o inhibir el desarrollo de sistemas de precipita-ción durante largos períodos. Además de estos forzantes, existen factores regionales y locales que pueden contribuir a una modulación del evento extremo una vez que se inicia (Mo y Schemm, 2008). La época del año, el transporte de humedad y las condiciones de humedad del suelo son to-dos factores que influyen en la duración e intensidad de las sequías o los excesos hídricos.
Variabilidad y cambios en los extremos de precipitación. Análisis zonal
Los cambios y la variabilidad en la precipitación y sus extre-mos han afectado las actividades productivas y la población de la provincia de Santa Fe, principalmente a través de tor-mentas intensas, aunque también por déficits que conllevan a severas sequías. Las observaciones en estaciones meteo-rológicas distribuidas en diferentes regiones de la provincia o
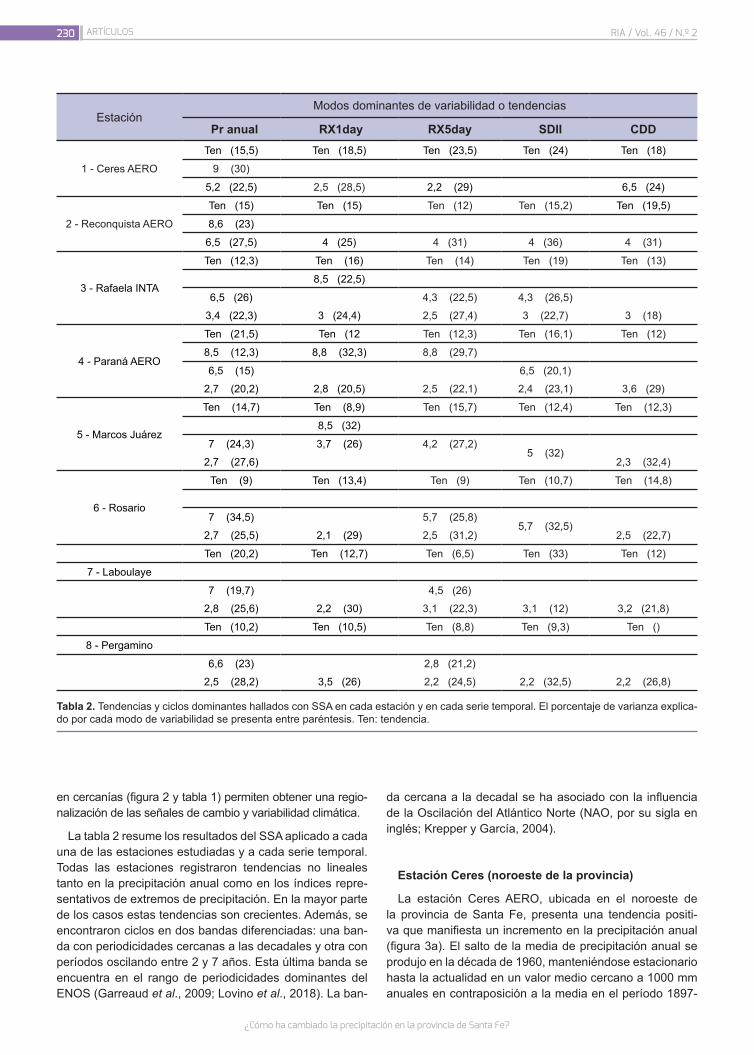
¿Cómo ha cambiado la precipitación en la provincia de Santa Fe?
ARTÍCULOS RIA / Vol. 46 / N.º 2230230
en cercanías (figura 2 y tabla 1) permiten obtener una regio-nalización de las señales de cambio y variabilidad climática.
La tabla 2 resume los resultados del SSA aplicado a cada una de las estaciones estudiadas y a cada serie temporal. Todas las estaciones registraron tendencias no lineales tanto en la precipitación anual como en los índices repre-sentativos de extremos de precipitación. En la mayor parte de los casos estas tendencias son crecientes. Además, se encontraron ciclos en dos bandas diferenciadas: una ban-da con periodicidades cercanas a las decadales y otra con períodos oscilando entre 2 y 7 años. Esta última banda se encuentra en el rango de periodicidades dominantes del ENOS (Garreaud et al., 2009; Lovino et al., 2018). La ban-
da cercana a la decadal se ha asociado con la influencia de la Oscilación del Atlántico Norte (NAO, por su sigla en inglés; Krepper y García, 2004).
Estación Ceres (noroeste de la provincia)
La estación Ceres AERO, ubicada en el noroeste de la provincia de Santa Fe, presenta una tendencia positi-va que manifiesta un incremento en la precipitación anual (figura 3a). El salto de la media de precipitación anual se produjo en la década de 1960, manteniéndose estacionario hasta la actualidad en un valor medio cercano a 1000 mm anuales en contraposición a la media en el período 1897-
Tabla 2. Tendencias y ciclos dominantes hallados con SSA en cada estación y en cada serie temporal. El porcentaje de varianza explica-do por cada modo de variabilidad se presenta entre paréntesis. Ten: tendencia.
EstaciónModos dominantes de variabilidad o tendencias
Pr anual RX1day RX5day SDII CDD
1 - Ceres AERO
Ten (15,5) Ten (18,5) Ten (23,5) Ten (24) Ten (18)
9 (30)
5,2 (22,5) 2,5 (28,5) 2,2 (29) 6,5 (24)
2 - Reconquista AERO
Ten (15) Ten (15) Ten (12) Ten (15,2) Ten (19,5)
8,6 (23)
6,5 (27,5) 4 (25) 4 (31) 4 (36) 4 (31)
3 - Rafaela INTA
Ten (12,3) Ten (16) Ten (14) Ten (19) Ten (13)
8,5 (22,5)
6,5 (26)3,4 (22,3) 3 (24,4)
4,3 (22,5)2,5 (27,4)
4,3 (26,5)3 (22,7) 3 (18)
4 - Paraná AERO
Ten (21,5) Ten (12 Ten (12,3) Ten (16,1) Ten (12)
8,5 (12,3) 8,8 (32,3) 8,8 (29,7)
6,5 (15)2,7 (20,2) 2,8 (20,5) 2,5 (22,1)
6,5 (20,1)2,4 (23,1) 3,6 (29)
5 - Marcos Juárez
Ten (14,7) Ten (8,9) Ten (15,7) Ten (12,4) Ten (12,3)
8,5 (32)
7 (24,3)2,7 (27,6)
3,7 (26) 4,2 (27,2)5 (32)
2,3 (32,4)
6 - Rosario
Ten (9) Ten (13,4) Ten (9) Ten (10,7) Ten (14,8)
7 (34,5)2,7 (25,5) 2,1 (29)
5,7 (25,8)2,5 (31,2)
5,7 (32,5)2,5 (22,7)
Ten (20,2) Ten (12,7) Ten (6,5) Ten (33) Ten (12)
7 - Laboulaye
7 (19,7)2,8 (25,6) 2,2 (30)
4,5 (26)3,1 (22,3) 3,1 (12) 3,2 (21,8)
Ten (10,2) Ten (10,5) Ten (8,8) Ten (9,3) Ten ()
8 - Pergamino
6,6 (23)2,5 (28,2) 3,5 (26)
2,8 (21,2)2,2 (24,5) 2,2 (32,5) 2,2 (26,8)
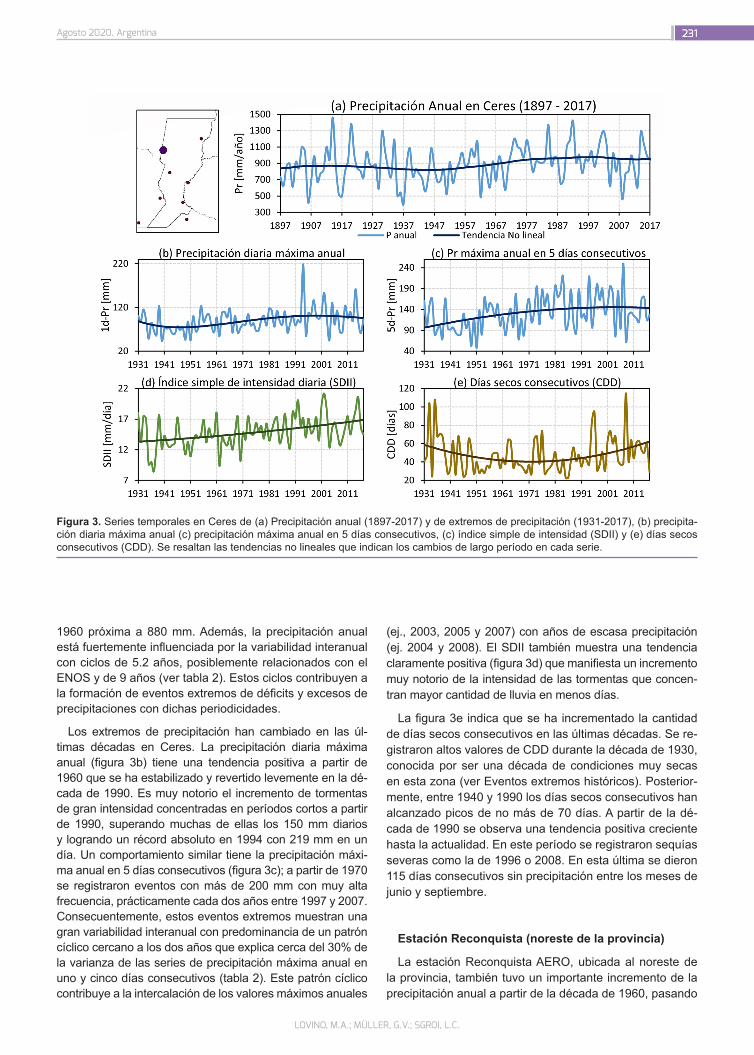
LOVINO, M.A.; MÜLLER, G.V.; SGROI, L.C.
Agosto 2020, Argentina 231231
1960 próxima a 880 mm. Además, la precipitación anual está fuertemente influenciada por la variabilidad interanual con ciclos de 5.2 años, posiblemente relacionados con el ENOS y de 9 años (ver tabla 2). Estos ciclos contribuyen a la formación de eventos extremos de déficits y excesos de precipitaciones con dichas periodicidades.
Los extremos de precipitación han cambiado en las úl-timas décadas en Ceres. La precipitación diaria máxima anual (figura 3b) tiene una tendencia positiva a partir de 1960 que se ha estabilizado y revertido levemente en la dé-cada de 1990. Es muy notorio el incremento de tormentas de gran intensidad concentradas en períodos cortos a partir de 1990, superando muchas de ellas los 150 mm diarios y logrando un récord absoluto en 1994 con 219 mm en un día. Un comportamiento similar tiene la precipitación máxi-ma anual en 5 días consecutivos (figura 3c); a partir de 1970 se registraron eventos con más de 200 mm con muy alta frecuencia, prácticamente cada dos años entre 1997 y 2007. Consecuentemente, estos eventos extremos muestran una gran variabilidad interanual con predominancia de un patrón cíclico cercano a los dos años que explica cerca del 30% de la varianza de las series de precipitación máxima anual en uno y cinco días consecutivos (tabla 2). Este patrón cíclico contribuye a la intercalación de los valores máximos anuales
(ej., 2003, 2005 y 2007) con años de escasa precipitación (ej. 2004 y 2008). El SDII también muestra una tendencia claramente positiva (figura 3d) que manifiesta un incremento muy notorio de la intensidad de las tormentas que concen-tran mayor cantidad de lluvia en menos días.
La figura 3e indica que se ha incrementado la cantidad de días secos consecutivos en las últimas décadas. Se re-gistraron altos valores de CDD durante la década de 1930, conocida por ser una década de condiciones muy secas en esta zona (ver Eventos extremos históricos). Posterior-mente, entre 1940 y 1990 los días secos consecutivos han alcanzado picos de no más de 70 días. A partir de la dé-cada de 1990 se observa una tendencia positiva creciente hasta la actualidad. En este período se registraron sequías severas como la de 1996 o 2008. En esta última se dieron 115 días consecutivos sin precipitación entre los meses de junio y septiembre.
Estación Reconquista (noreste de la provincia)
La estación Reconquista AERO, ubicada al noreste de la provincia, también tuvo un importante incremento de la precipitación anual a partir de la década de 1960, pasando
Figura 3. Series temporales en Ceres de (a) Precipitación anual (1897-2017) y de extremos de precipitación (1931-2017), (b) precipita-ción diaria máxima anual (c) precipitación máxima anual en 5 días consecutivos, (c) índice simple de intensidad (SDII) y (e) días secos consecutivos (CDD). Se resaltan las tendencias no lineales que indican los cambios de largo período en cada serie.
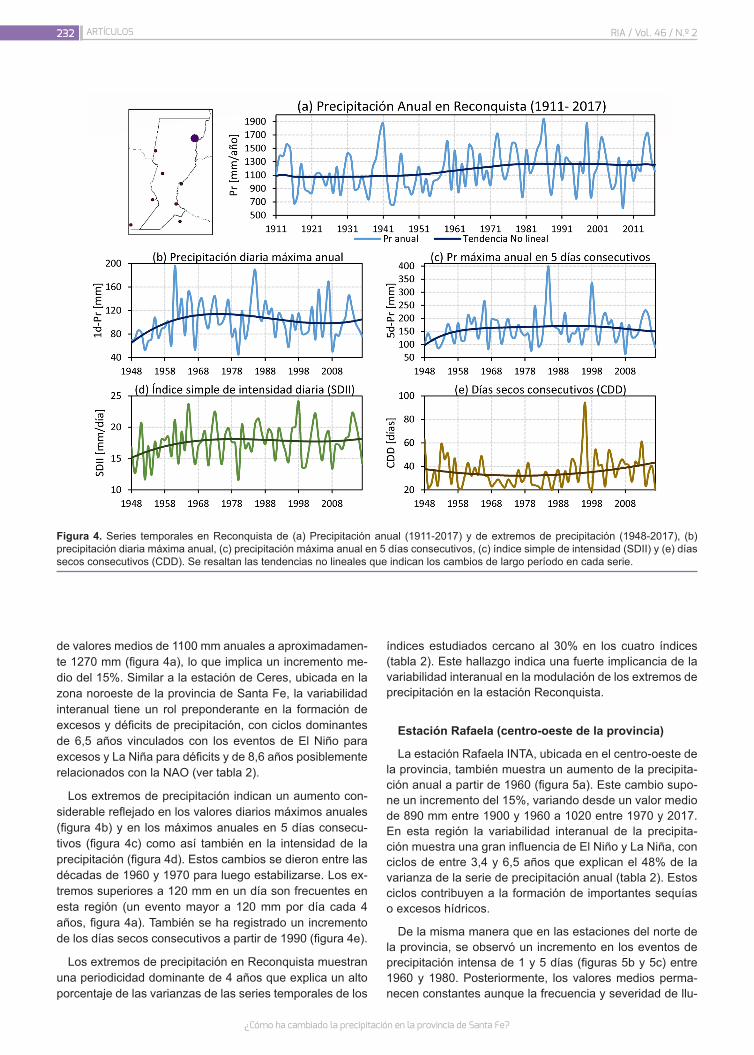
¿Cómo ha cambiado la precipitación en la provincia de Santa Fe?
ARTÍCULOS RIA / Vol. 46 / N.º 2232232
de valores medios de 1100 mm anuales a aproximadamen-te 1270 mm (figura 4a), lo que implica un incremento me-dio del 15%. Similar a la estación de Ceres, ubicada en la zona noroeste de la provincia de Santa Fe, la variabilidad interanual tiene un rol preponderante en la formación de excesos y déficits de precipitación, con ciclos dominantes de 6,5 años vinculados con los eventos de El Niño para excesos y La Niña para déficits y de 8,6 años posiblemente relacionados con la NAO (ver tabla 2).
Los extremos de precipitación indican un aumento con-siderable reflejado en los valores diarios máximos anuales (figura 4b) y en los máximos anuales en 5 días consecu-tivos (figura 4c) como así también en la intensidad de la precipitación (figura 4d). Estos cambios se dieron entre las décadas de 1960 y 1970 para luego estabilizarse. Los ex-tremos superiores a 120 mm en un día son frecuentes en esta región (un evento mayor a 120 mm por día cada 4 años, figura 4a). También se ha registrado un incremento de los días secos consecutivos a partir de 1990 (figura 4e).
Los extremos de precipitación en Reconquista muestran una periodicidad dominante de 4 años que explica un alto porcentaje de las varianzas de las series temporales de los
índices estudiados cercano al 30% en los cuatro índices (tabla 2). Este hallazgo indica una fuerte implicancia de la variabilidad interanual en la modulación de los extremos de precipitación en la estación Reconquista.
Estación Rafaela (centro-oeste de la provincia)
La estación Rafaela INTA, ubicada en el centro-oeste de la provincia, también muestra un aumento de la precipita-ción anual a partir de 1960 (figura 5a). Este cambio supo-ne un incremento del 15%, variando desde un valor medio de 890 mm entre 1900 y 1960 a 1020 entre 1970 y 2017. En esta región la variabilidad interanual de la precipita-ción muestra una gran influencia de El Niño y La Niña, con ciclos de entre 3,4 y 6,5 años que explican el 48% de la varianza de la serie de precipitación anual (tabla 2). Estos ciclos contribuyen a la formación de importantes sequías o excesos hídricos.
De la misma manera que en las estaciones del norte de la provincia, se observó un incremento en los eventos de precipitación intensa de 1 y 5 días (figuras 5b y 5c) entre 1960 y 1980. Posteriormente, los valores medios perma-necen constantes aunque la frecuencia y severidad de llu-
Figura 4. Series temporales en Reconquista de (a) Precipitación anual (1911-2017) y de extremos de precipitación (1948-2017), (b) precipitación diaria máxima anual, (c) precipitación máxima anual en 5 días consecutivos, (c) índice simple de intensidad (SDII) y (e) días secos consecutivos (CDD). Se resaltan las tendencias no lineales que indican los cambios de largo período en cada serie.
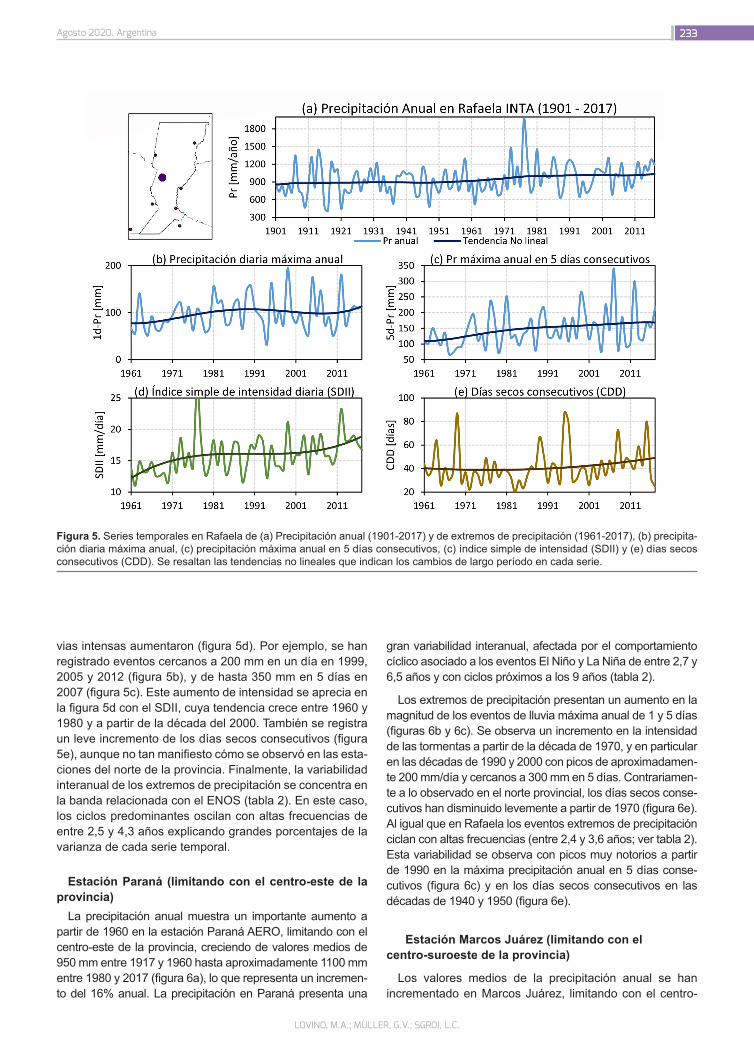
LOVINO, M.A.; MÜLLER, G.V.; SGROI, L.C.
Agosto 2020, Argentina 233233
vias intensas aumentaron (figura 5d). Por ejemplo, se han registrado eventos cercanos a 200 mm en un día en 1999, 2005 y 2012 (figura 5b), y de hasta 350 mm en 5 días en 2007 (figura 5c). Este aumento de intensidad se aprecia en la figura 5d con el SDII, cuya tendencia crece entre 1960 y 1980 y a partir de la década del 2000. También se registra un leve incremento de los días secos consecutivos (figura 5e), aunque no tan manifiesto cómo se observó en las esta-ciones del norte de la provincia. Finalmente, la variabilidad interanual de los extremos de precipitación se concentra en la banda relacionada con el ENOS (tabla 2). En este caso, los ciclos predominantes oscilan con altas frecuencias de entre 2,5 y 4,3 años explicando grandes porcentajes de la varianza de cada serie temporal.
Estación Paraná (limitando con el centro-este de la provincia)
La precipitación anual muestra un importante aumento a partir de 1960 en la estación Paraná AERO, limitando con el centro-este de la provincia, creciendo de valores medios de 950 mm entre 1917 y 1960 hasta aproximadamente 1100 mm entre 1980 y 2017 (figura 6a), lo que representa un incremen-to del 16% anual. La precipitación en Paraná presenta una
gran variabilidad interanual, afectada por el comportamiento cíclico asociado a los eventos El Niño y La Niña de entre 2,7 y 6,5 años y con ciclos próximos a los 9 años (tabla 2).
Los extremos de precipitación presentan un aumento en la magnitud de los eventos de lluvia máxima anual de 1 y 5 días (figuras 6b y 6c). Se observa un incremento en la intensidad de las tormentas a partir de la década de 1970, y en particular en las décadas de 1990 y 2000 con picos de aproximadamen-te 200 mm/día y cercanos a 300 mm en 5 días. Contrariamen-te a lo observado en el norte provincial, los días secos conse-cutivos han disminuido levemente a partir de 1970 (figura 6e). Al igual que en Rafaela los eventos extremos de precipitación ciclan con altas frecuencias (entre 2,4 y 3,6 años; ver tabla 2). Esta variabilidad se observa con picos muy notorios a partir de 1990 en la máxima precipitación anual en 5 días conse-cutivos (figura 6c) y en los días secos consecutivos en las décadas de 1940 y 1950 (figura 6e).
Estación Marcos Juárez (limitando con el centro-suroeste de la provincia)
Los valores medios de la precipitación anual se han incrementado en Marcos Juárez, limitando con el centro-
Figura 5. Series temporales en Rafaela de (a) Precipitación anual (1901-2017) y de extremos de precipitación (1961-2017), (b) precipita-ción diaria máxima anual, (c) precipitación máxima anual en 5 días consecutivos, (c) índice simple de intensidad (SDII) y (e) días secos consecutivos (CDD). Se resaltan las tendencias no lineales que indican los cambios de largo período en cada serie.
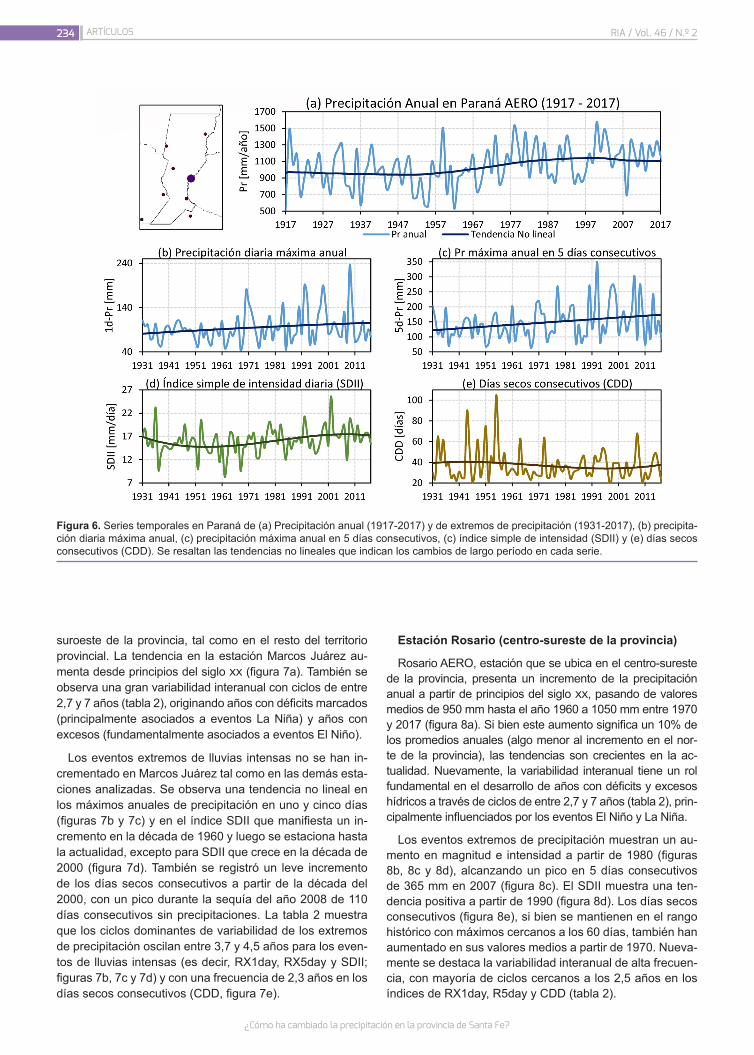
¿Cómo ha cambiado la precipitación en la provincia de Santa Fe?
ARTÍCULOS RIA / Vol. 46 / N.º 2234234
suroeste de la provincia, tal como en el resto del territorio provincial. La tendencia en la estación Marcos Juárez au-menta desde principios del siglo xx (figura 7a). También se observa una gran variabilidad interanual con ciclos de entre 2,7 y 7 años (tabla 2), originando años con déficits marcados (principalmente asociados a eventos La Niña) y años con excesos (fundamentalmente asociados a eventos El Niño).
Los eventos extremos de lluvias intensas no se han in-crementado en Marcos Juárez tal como en las demás esta-ciones analizadas. Se observa una tendencia no lineal en los máximos anuales de precipitación en uno y cinco días (figuras 7b y 7c) y en el índice SDII que manifiesta un in-cremento en la década de 1960 y luego se estaciona hasta la actualidad, excepto para SDII que crece en la década de 2000 (figura 7d). También se registró un leve incremento de los días secos consecutivos a partir de la década del 2000, con un pico durante la sequía del año 2008 de 110 días consecutivos sin precipitaciones. La tabla 2 muestra que los ciclos dominantes de variabilidad de los extremos de precipitación oscilan entre 3,7 y 4,5 años para los even-tos de lluvias intensas (es decir, RX1day, RX5day y SDII; figuras 7b, 7c y 7d) y con una frecuencia de 2,3 años en los días secos consecutivos (CDD, figura 7e).
Estación Rosario (centro-sureste de la provincia)
Rosario AERO, estación que se ubica en el centro-sureste de la provincia, presenta un incremento de la precipitación anual a partir de principios del siglo xx, pasando de valores medios de 950 mm hasta el año 1960 a 1050 mm entre 1970 y 2017 (figura 8a). Si bien este aumento significa un 10% de los promedios anuales (algo menor al incremento en el nor-te de la provincia), las tendencias son crecientes en la ac-tualidad. Nuevamente, la variabilidad interanual tiene un rol fundamental en el desarrollo de años con déficits y excesos hídricos a través de ciclos de entre 2,7 y 7 años (tabla 2), prin-cipalmente influenciados por los eventos El Niño y La Niña.
Los eventos extremos de precipitación muestran un au-mento en magnitud e intensidad a partir de 1980 (figuras 8b, 8c y 8d), alcanzando un pico en 5 días consecutivos de 365 mm en 2007 (figura 8c). El SDII muestra una ten-dencia positiva a partir de 1990 (figura 8d). Los días secos consecutivos (figura 8e), si bien se mantienen en el rango histórico con máximos cercanos a los 60 días, también han aumentado en sus valores medios a partir de 1970. Nueva-mente se destaca la variabilidad interanual de alta frecuen-cia, con mayoría de ciclos cercanos a los 2,5 años en los índices de RX1day, R5day y CDD (tabla 2).
Figura 6. Series temporales en Paraná de (a) Precipitación anual (1917-2017) y de extremos de precipitación (1931-2017), (b) precipita-ción diaria máxima anual, (c) precipitación máxima anual en 5 días consecutivos, (c) índice simple de intensidad (SDII) y (e) días secos consecutivos (CDD). Se resaltan las tendencias no lineales que indican los cambios de largo período en cada serie.
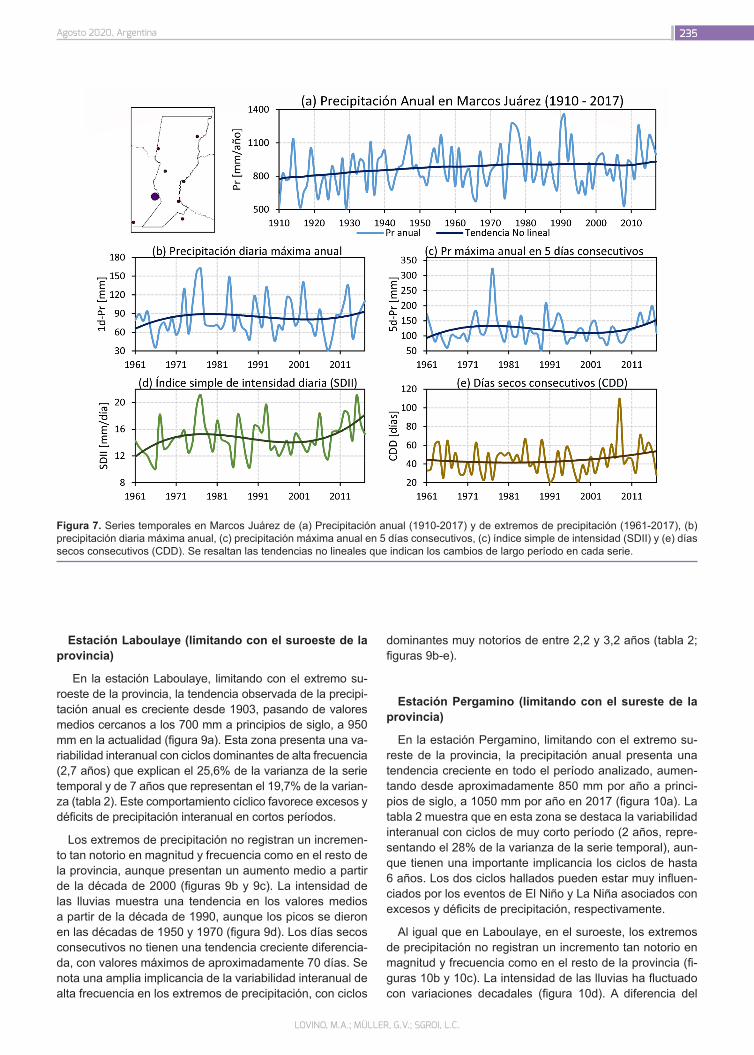
LOVINO, M.A.; MÜLLER, G.V.; SGROI, L.C.
Agosto 2020, Argentina 235235
Estación Laboulaye (limitando con el suroeste de la provincia)
En la estación Laboulaye, limitando con el extremo su-roeste de la provincia, la tendencia observada de la precipi-tación anual es creciente desde 1903, pasando de valores medios cercanos a los 700 mm a principios de siglo, a 950 mm en la actualidad (figura 9a). Esta zona presenta una va-riabilidad interanual con ciclos dominantes de alta frecuencia (2,7 años) que explican el 25,6% de la varianza de la serie temporal y de 7 años que representan el 19,7% de la varian-za (tabla 2). Este comportamiento cíclico favorece excesos y déficits de precipitación interanual en cortos períodos.
Los extremos de precipitación no registran un incremen-to tan notorio en magnitud y frecuencia como en el resto de la provincia, aunque presentan un aumento medio a partir de la década de 2000 (figuras 9b y 9c). La intensidad de las lluvias muestra una tendencia en los valores medios a partir de la década de 1990, aunque los picos se dieron en las décadas de 1950 y 1970 (figura 9d). Los días secos consecutivos no tienen una tendencia creciente diferencia-da, con valores máximos de aproximadamente 70 días. Se nota una amplia implicancia de la variabilidad interanual de alta frecuencia en los extremos de precipitación, con ciclos
dominantes muy notorios de entre 2,2 y 3,2 años (tabla 2; figuras 9b-e).
Estación Pergamino (limitando con el sureste de la provincia)
En la estación Pergamino, limitando con el extremo su-reste de la provincia, la precipitación anual presenta una tendencia creciente en todo el período analizado, aumen-tando desde aproximadamente 850 mm por año a princi-pios de siglo, a 1050 mm por año en 2017 (figura 10a). La tabla 2 muestra que en esta zona se destaca la variabilidad interanual con ciclos de muy corto período (2 años, repre-sentando el 28% de la varianza de la serie temporal), aun-que tienen una importante implicancia los ciclos de hasta 6 años. Los dos ciclos hallados pueden estar muy influen-ciados por los eventos de El Niño y La Niña asociados con excesos y déficits de precipitación, respectivamente.
Al igual que en Laboulaye, en el suroeste, los extremos de precipitación no registran un incremento tan notorio en magnitud y frecuencia como en el resto de la provincia (fi-guras 10b y 10c). La intensidad de las lluvias ha fluctuado con variaciones decadales (figura 10d). A diferencia del
Figura 7. Series temporales en Marcos Juárez de (a) Precipitación anual (1910-2017) y de extremos de precipitación (1961-2017), (b) precipitación diaria máxima anual, (c) precipitación máxima anual en 5 días consecutivos, (c) índice simple de intensidad (SDII) y (e) días secos consecutivos (CDD). Se resaltan las tendencias no lineales que indican los cambios de largo período en cada serie.
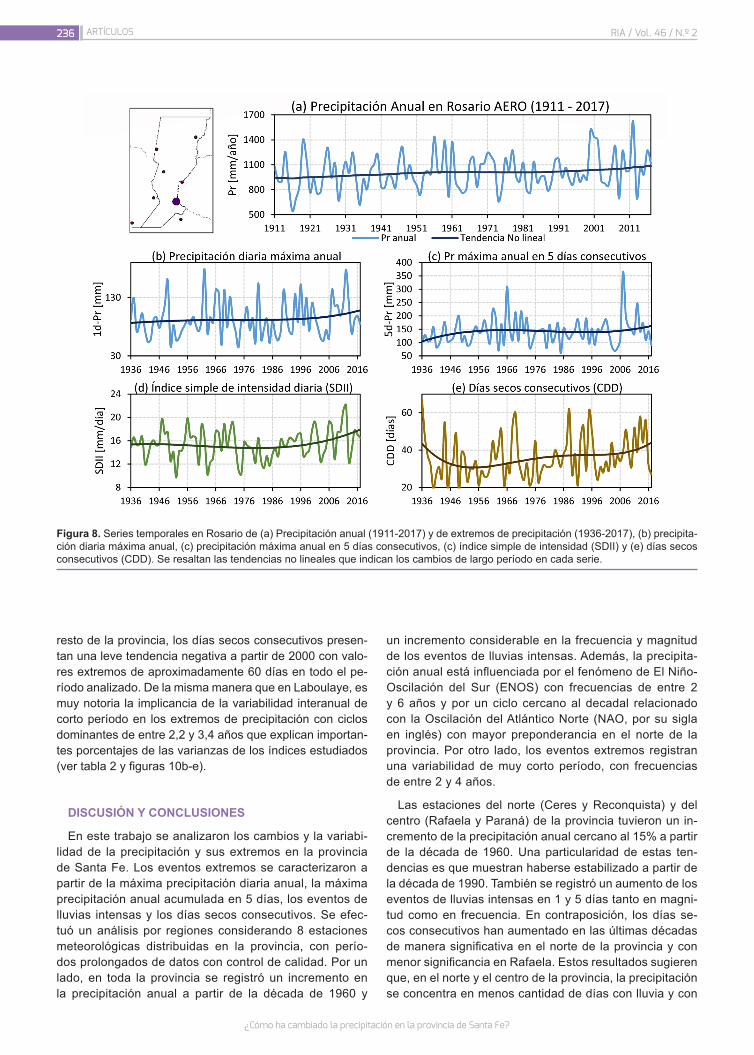
¿Cómo ha cambiado la precipitación en la provincia de Santa Fe?
ARTÍCULOS RIA / Vol. 46 / N.º 2236236
resto de la provincia, los días secos consecutivos presen-tan una leve tendencia negativa a partir de 2000 con valo-res extremos de aproximadamente 60 días en todo el pe-ríodo analizado. De la misma manera que en Laboulaye, es muy notoria la implicancia de la variabilidad interanual de corto período en los extremos de precipitación con ciclos dominantes de entre 2,2 y 3,4 años que explican importan-tes porcentajes de las varianzas de los índices estudiados (ver tabla 2 y figuras 10b-e).
DISCUSIÓN Y CONCLUSIONES
En este trabajo se analizaron los cambios y la variabi-lidad de la precipitación y sus extremos en la provincia de Santa Fe. Los eventos extremos se caracterizaron a partir de la máxima precipitación diaria anual, la máxima precipitación anual acumulada en 5 días, los eventos de lluvias intensas y los días secos consecutivos. Se efec-tuó un análisis por regiones considerando 8 estaciones meteorológicas distribuidas en la provincia, con perío-dos prolongados de datos con control de calidad. Por un lado, en toda la provincia se registró un incremento en la precipitación anual a partir de la década de 1960 y
un incremento considerable en la frecuencia y magnitud de los eventos de lluvias intensas. Además, la precipita-ción anual está influenciada por el fenómeno de El Niño-Oscilación del Sur (ENOS) con frecuencias de entre 2 y 6 años y por un ciclo cercano al decadal relacionado con la Oscilación del Atlántico Norte (NAO, por su sigla en inglés) con mayor preponderancia en el norte de la provincia. Por otro lado, los eventos extremos registran una variabilidad de muy corto período, con frecuencias de entre 2 y 4 años.
Las estaciones del norte (Ceres y Reconquista) y del centro (Rafaela y Paraná) de la provincia tuvieron un in-cremento de la precipitación anual cercano al 15% a partir de la década de 1960. Una particularidad de estas ten-dencias es que muestran haberse estabilizado a partir de la década de 1990. También se registró un aumento de los eventos de lluvias intensas en 1 y 5 días tanto en magni-tud como en frecuencia. En contraposición, los días se-cos consecutivos han aumentado en las últimas décadas de manera significativa en el norte de la provincia y con menor significancia en Rafaela. Estos resultados sugieren que, en el norte y el centro de la provincia, la precipitación se concentra en menos cantidad de días con lluvia y con
Figura 8. Series temporales en Rosario de (a) Precipitación anual (1911-2017) y de extremos de precipitación (1936-2017), (b) precipita-ción diaria máxima anual, (c) precipitación máxima anual en 5 días consecutivos, (c) índice simple de intensidad (SDII) y (e) días secos consecutivos (CDD). Se resaltan las tendencias no lineales que indican los cambios de largo período en cada serie.
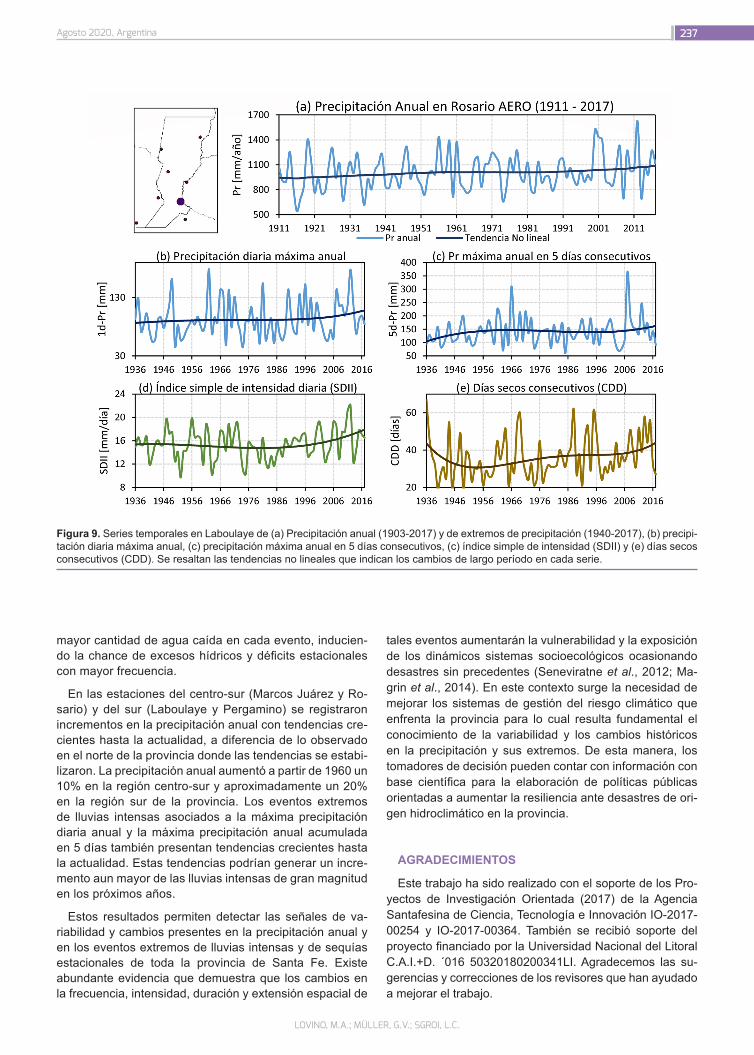
LOVINO, M.A.; MÜLLER, G.V.; SGROI, L.C.
Agosto 2020, Argentina 237237
Figura 9. Series temporales en Laboulaye de (a) Precipitación anual (1903-2017) y de extremos de precipitación (1940-2017), (b) precipi-tación diaria máxima anual, (c) precipitación máxima anual en 5 días consecutivos, (c) índice simple de intensidad (SDII) y (e) días secos consecutivos (CDD). Se resaltan las tendencias no lineales que indican los cambios de largo período en cada serie.
mayor cantidad de agua caída en cada evento, inducien-do la chance de excesos hídricos y déficits estacionales con mayor frecuencia.
En las estaciones del centro-sur (Marcos Juárez y Ro-sario) y del sur (Laboulaye y Pergamino) se registraron incrementos en la precipitación anual con tendencias cre-cientes hasta la actualidad, a diferencia de lo observado en el norte de la provincia donde las tendencias se estabi-lizaron. La precipitación anual aumentó a partir de 1960 un 10% en la región centro-sur y aproximadamente un 20% en la región sur de la provincia. Los eventos extremos de lluvias intensas asociados a la máxima precipitación diaria anual y la máxima precipitación anual acumulada en 5 días también presentan tendencias crecientes hasta la actualidad. Estas tendencias podrían generar un incre-mento aun mayor de las lluvias intensas de gran magnitud en los próximos años.
Estos resultados permiten detectar las señales de va-riabilidad y cambios presentes en la precipitación anual y en los eventos extremos de lluvias intensas y de sequías estacionales de toda la provincia de Santa Fe. Existe abundante evidencia que demuestra que los cambios en la frecuencia, intensidad, duración y extensión espacial de
tales eventos aumentarán la vulnerabilidad y la exposición de los dinámicos sistemas socioecológicos ocasionando desastres sin precedentes (Seneviratne et al., 2012; Ma-grin et al., 2014). En este contexto surge la necesidad de mejorar los sistemas de gestión del riesgo climático que enfrenta la provincia para lo cual resulta fundamental el conocimiento de la variabilidad y los cambios históricos en la precipitación y sus extremos. De esta manera, los tomadores de decisión pueden contar con información con base científica para la elaboración de políticas públicas orientadas a aumentar la resiliencia ante desastres de ori-gen hidroclimático en la provincia.
AGRADECIMIENTOS
Este trabajo ha sido realizado con el soporte de los Pro-yectos de Investigación Orientada (2017) de la Agencia Santafesina de Ciencia, Tecnología e Innovación IO-2017-00254 y IO-2017-00364. También se recibió soporte del proyecto financiado por la Universidad Nacional del Litoral C.A.I.+D. ´016 50320180200341LI. Agradecemos las su-gerencias y correcciones de los revisores que han ayudado a mejorar el trabajo.
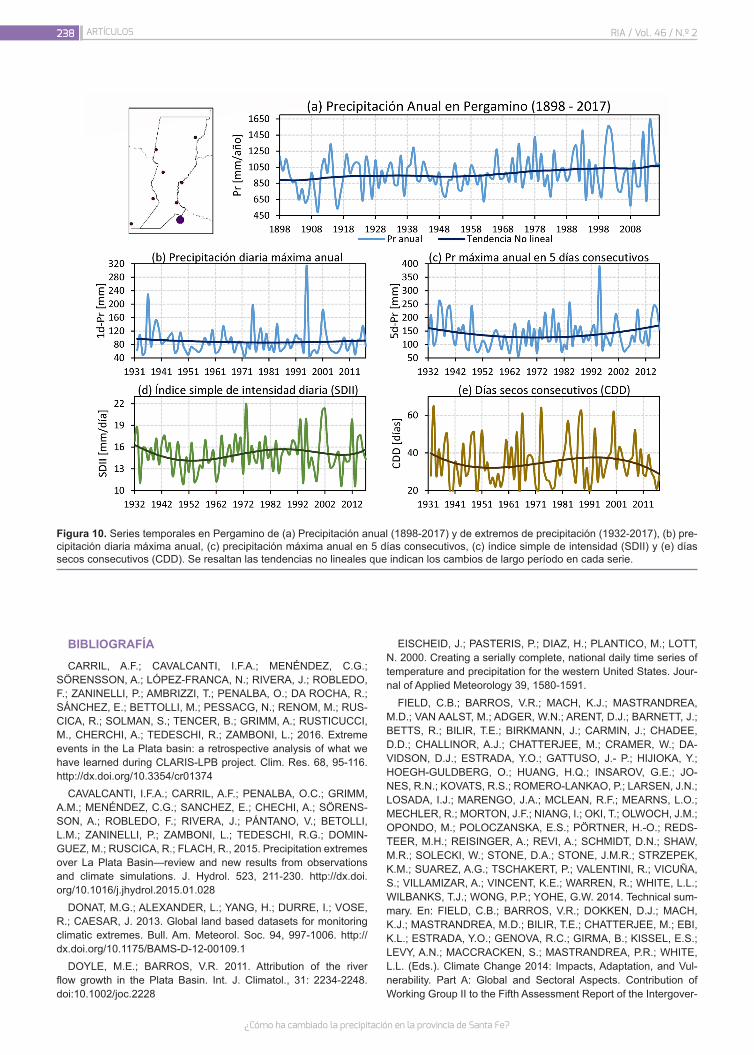
¿Cómo ha cambiado la precipitación en la provincia de Santa Fe?
ARTÍCULOS RIA / Vol. 46 / N.º 2238238
Figura 10. Series temporales en Pergamino de (a) Precipitación anual (1898-2017) y de extremos de precipitación (1932-2017), (b) pre-cipitación diaria máxima anual, (c) precipitación máxima anual en 5 días consecutivos, (c) índice simple de intensidad (SDII) y (e) días secos consecutivos (CDD). Se resaltan las tendencias no lineales que indican los cambios de largo período en cada serie.
BIBLIOGRAFÍACARRIL, A.F.; CAVALCANTI, I.F.A.; MENÉNDEZ, C.G.;
SÖRENSSON, A.; LÓPEZ-FRANCA, N.; RIVERA, J.; ROBLEDO, F.; ZANINELLI, P.; AMBRIZZI, T.; PENALBA, O.; DA ROCHA, R.; SÁNCHEZ, E.; BETTOLLI, M.; PESSACG, N.; RENOM, M.; RUS-CICA, R.; SOLMAN, S.; TENCER, B.; GRIMM, A.; RUSTICUCCI, M., CHERCHI, A.; TEDESCHI, R.; ZAMBONI, L.; 2016. Extreme events in the La Plata basin: a retrospective analysis of what we have learned during CLARIS-LPB project. Clim. Res. 68, 95-116. http://dx.doi.org/10.3354/cr01374
CAVALCANTI, I.F.A.; CARRIL, A.F.; PENALBA, O.C.; GRIMM, A.M.; MENÉNDEZ, C.G.; SANCHEZ, E.; CHECHI, A.; SÖRENS-SON, A.; ROBLEDO, F.; RIVERA, J.; PÁNTANO, V.; BETOLLI, L.M.; ZANINELLI, P.; ZAMBONI, L.; TEDESCHI, R.G.; DOMIN-GUEZ, M.; RUSCICA, R.; FLACH, R., 2015. Precipitation extremes over La Plata Basin—review and new results from observations and climate simulations. J. Hydrol. 523, 211-230. http://dx.doi.org/10.1016/j.jhydrol.2015.01.028
DONAT, M.G.; ALEXANDER, L.; YANG, H.; DURRE, I.; VOSE, R.; CAESAR, J. 2013. Global land based datasets for monitoring climatic extremes. Bull. Am. Meteorol. Soc. 94, 997-1006. http://dx.doi.org/10.1175/BAMS-D-12-00109.1
DOYLE, M.E.; BARROS, V.R. 2011. Attribution of the river flow growth in the Plata Basin. Int. J. Climatol., 31: 2234-2248. doi:10.1002/joc.2228
EISCHEID, J.; PASTERIS, P.; DIAZ, H.; PLANTICO, M.; LOTT, N. 2000. Creating a serially complete, national daily time series of temperature and precipitation for the western United States. Jour-nal of Applied Meteorology 39, 1580-1591.
FIELD, C.B.; BARROS, V.R.; MACH, K.J.; MASTRANDREA, M.D.; VAN AALST, M.; ADGER, W.N.; ARENT, D.J.; BARNETT, J.; BETTS, R.; BILIR, T.E.; BIRKMANN, J.; CARMIN, J.; CHADEE, D.D.; CHALLINOR, A.J.; CHATTERJEE, M.; CRAMER, W.; DA-VIDSON, D.J.; ESTRADA, Y.O.; GATTUSO, J.- P.; HIJIOKA, Y.; HOEGH-GULDBERG, O.; HUANG, H.Q.; INSAROV, G.E.; JO-NES, R.N.; KOVATS, R.S.; ROMERO-LANKAO, P.; LARSEN, J.N.; LOSADA, I.J.; MARENGO, J.A.; MCLEAN, R.F.; MEARNS, L.O.; MECHLER, R.; MORTON, J.F.; NIANG, I.; OKI, T.; OLWOCH, J.M.; OPONDO, M.; POLOCZANSKA, E.S.; PÖRTNER, H.-O.; REDS-TEER, M.H.; REISINGER, A.; REVI, A.; SCHMIDT, D.N.; SHAW, M.R.; SOLECKI, W.; STONE, D.A.; STONE, J.M.R.; STRZEPEK, K.M.; SUAREZ, A.G.; TSCHAKERT, P.; VALENTINI, R.; VICUÑA, S.; VILLAMIZAR, A.; VINCENT, K.E.; WARREN, R.; WHITE, L.L.; WILBANKS, T.J.; WONG, P.P.; YOHE, G.W. 2014. Technical sum-mary. En: FIELD, C.B.; BARROS, V.R.; DOKKEN, D.J.; MACH, K.J.; MASTRANDREA, M.D.; BILIR, T.E.; CHATTERJEE, M.; EBI, K.L.; ESTRADA, Y.O.; GENOVA, R.C.; GIRMA, B.; KISSEL, E.S.; LEVY, A.N.; MACCRACKEN, S.; MASTRANDREA, P.R.; WHITE, L.L. (Eds.). Climate Change 2014: Impacts, Adaptation, and Vul-nerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergover-

LOVINO, M.A.; MÜLLER, G.V.; SGROI, L.C.
Agosto 2020, Argentina 239239
nmental Panel on Climate Change. Cambridge University Press, Cambridge, Reino Unido y NuevaYork, NY, EUA, 35-94 pp.
FIELD, C.B.; BARROS, V.; STOCKER, T.F.; QIN, D.; DOKKEN, D.J.; EBI, K.L.; MASTRANDREA, M.D.; MACH, K.J.; PLATTNER, G.K.; ALLEN, S.K.; TIGNOR, M.; MIDGLEY, P.M. 2014. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the AR5 of the IPCC. Cambridge University Press, Cambridge, Reino Unido y Nueva York, EUA.
GARREAUD, R.D.; VUILLE, M.; COMPAGNUCCI, R.; MAREN-GO, J. 2009. Present-day South American climate, Paleogeogr. Pa-leocl., 281, 180-195. https://doi.org/10.1016/j.palaeo.2007.10.032.
GHIL, M.; ALLEN, M.; DETTINGER, M.D.; IDE, K.; KON-DRASHOV, D.; MANN, M.; ROBERTSON, A.; SAUNDERS, A.; TIAN, Y.; VARADI, F.; YIOU, P. 2001. Advanced spectral method for climatic time series. Rev. Geophys. 40, 1-41. http://dx.doi.org/10.1029/2001RG000092.
IPCC. 2013. Planton, S. (ed). Annex III: Glossary. En: STOC-KER, T.F.; QIN, D.; PLATTNER, G.K.; TIGNOR, M.; ALLEN, s.k.; BOSCHUNG, J.; NAUELS, A.; XIA, Y.;V. BEX, V.; MIDGLEY, P.M. (eds.). Climate Change 2013: The Physical Science Basis. Con-tribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. IPCC. Cambridge University Press, Cambridge, Reino Unido y Nueva York, EUA.
JACQUES-COPER, M.; GARREAUD, R.D. 2015. Characteriza-tion of the 1970s climate shift in South America, Int. J. Climatol., 35, 2164-2179, https://doi.org/10.1002/joc.4120
KREPPER, C.M.; GARCÍA, N.O. 2004. Spatial and temporal structure of trends and interannual variability of precipitation over La Plata Basin, Quartern. Int., 114, 11-21, https://doi.org/10.1016/S1040-6182(03)00038-7
LOVINO, M.A.; GARCÍA, N., BAETHGEN, W. 2014. Spatiotem-poral analysis of extreme precipitation events in the Northeast re-gion of Argentina (NEA). J. Hydrol.: Reg. Stud. 2, 140-158. http://dx.doi.org/10.1016/j.ejrh.2014.09.001
LOVINO, M.A. 2015. Impacto ambiental de la variabilidad climá-tica y los eventos extremos en la provincia de Santa Fe, en el con-texto del cambio climático. Tesis Doctoral, UNL, Argentina. (Dis-ponible: http://bibliotecavirtual.unl.edu.ar:8080/tesis/handle/11185/ 707 verificado: 20 de marzo de 2019). 238 p.
LOVINO, M.A.; MÜLLER, O.V.; MÜLLER, G.V.; SGROI, L.C.; BAETHGEN, W.E. 2018. Interannual-to-multidecadal Hydroclima-te Variability and its Sectoral Impacts in northeastern Argentina, Hydrol. Earth Syst. Sci., 22, 3155-3174, https://doi.org/10.5194/hess-22-3155-2018
MAGRIN, G.O.; MARENGO, J.A.; BOULANGER, J.-P.; BUCKE-RIDGE, M.S.; CASTELLANOS, E.; POVEDA, G.; SCARANO, F.R.; VICUÑA, S. 2014. Central and South America. En: BARROS, V.R.; FIELD, C.B.; DOKKEN, D.J.; MASTRANDREA, M.D.; MACH, K.J.;
BILIR, T.E.; CHATTERJEE, M.; EBI, K.L.; ESTRADA, Y.O.; GENO-VA, R.C.; GIRMA, B.; KISSEL, E.S.; LEVY, A.N.; MACCRACKEN, S.; MASTRANDREA, P.R.; WHITE, L.L. (Eds.). Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional As-pects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cam-bridge University Press, Cambridge, Reino Unido y NuevaYork, EUA. 1499-1566 pp.
MO, K.C.; SCHEMM, J.E. 2008. Relationships between ENSO and drought over the southeastern United States, Geophysical Re-search Letters, 35 (15).
MO, K.C.; BERBERY, E.H. 2011. Drought and Persistent Wet Spells over South America Based on Observations and the U.S. CLIVAR Drought Experiments, J. Climate, 24, 1801-1820. https://doi.org/10.1175/2010JCLI3874.1
RUSTICUCCI, M. 2012. Observed and simulated variability of extreme temperature events over South America. Atmospheric Re-search, 106, 1-17.
SEAGER, R.; NAIK, N.; BAETHGEN, W.; ROBERTSON, A.; KUSHNIR, Y.; NAKAMURA, J.; JURBURG, S. 2010. Tropical Oceanic Causes of Interannual to Multidecadal Precipitation Varia-bility in Southeast South America over the Past Century, J. Climate, 23, 5517-5539. https://doi.org/10.1175/2010JCLI3578.1
SCHNEIDER, U.; BECKER, A.; FINGER, P.; MEYER-CHRIS-TOFFER, A., RUDOLF, B.; ZIESE, M. 2015. GPCC Full Data Rea-nalysis Version 7.0 at 0.5: Monthly Land-Surface Precipitation from Rain Gauges built on GTS-based and Historic Data. https://doi.org/10.5676/DWD_GPCC/FD_M_V7_050
SENEVIRATNE, S.I.; NICHOLLS, N.; EASTERLING, D.; GOO-DESS, C.M.; KANAE, S.; KOSSIN, J.; LUO, Y.; MARENGO, J.; MCINNES, K.; RAHIMI, M.; REICHSTEIN, M.; SORTEBERG, A.; VERA, C.; ZHANG, X. 2012. Changes in climate extremes and their impacts on the natural physical environment. Managing the Risks of Extreme Events and Disasters to Advance Climate Chan-ge Adaptation. En: FIELD, C.B.; BARROS, V.; STOCKER, T.F.; QIN, D.; DOKKEN, D.J.; EBI, K.L.; MASTRANDREA, M.D.; MACH, K.J.; PLATTNER, G.K.; ALLEN, S.K.; TIGNOR, M.; MIDGLEY, P.M. (Eds.), A Special Report of Working Groups I and II of the IPCC. Cambridge University Press, Cambridge, Reino Unido y Nueva York, EUA. 109-230 pp.
WORLD METEOROLOGICAL ORGANIZATION (WMO), 2009. Manual for Estimation of Probable Maximum Precipitation, Publi-cation N.o 1045, World Meteorological Organization, Ginebra.
ZHANG, X.; ALEXANDER, L.V.; HEGERL, G.C.; KLEIN-TANK, A.; PETERSON, T.C.; TREWIN, B.; ZWIERS, F.W. 2011. Indices for monitoring changes in extremes based on daily temperature and precipitation data. Wiley Interdiscip. Rev. Clim. Chang. 2, 851-870. http://dx.doi.org/10.1002/wcc.147.

Evaluación del expeller de soja en dietas de cerdos en recría y engorde
ARTÍCULOS RIA / Vol. 46 / N.º 2240240
Evaluación del expeller de soja en dietas de cerdos en recría y engorde
RESUMENEl objetivo de este estudio fue evaluar la respuesta animal, calidad de la canal y de la carne de cerdos re-
criados y engordados con dietas con diferentes niveles de reemplazo de harina de soja por expeller de soja. Treinta y seis animales de 40±2 kg de peso vivo (PV) inicial fueron alimentados con 4 dietas isoproteicas (19 y 16% para recría y engorde, respectivamente) compuestas por maíz, expeller y harina de soja y el núcleo vitamínico mineral, determinadas por el porcentaje de inclusión del expeller de soja como oferente proteico en reemplazo de harina de soja. Los tratamientos fueron identificados como: D0: 100% harina de soja (dieta control); D33: 33,3% expeller de soja, 66,7% harina de soja; D66: 66,7% expeller de soja, 33,3% harina de soja y la D100: 100% expeller de soja. Superados los 100 kg de PV se realizó la faena. No se encontraron diferencias ni contrastes significativos (P>0,05) en los parámetros productivos evaluados. La ganancia me-dia diaria fue de 895,5±0,04 g, el índice de conversión promedio de 2,4±0,33 y el 53±0,86% de magro. Los tratamientos establecidos no tuvieron efectos significativos (P>0,05) sobre las características físicas de la carcasa. Los valores promedios de pH a los 45 min y 24 h post mórtem fueron de 5,95±0,22 y 5,74±0,21, respectivamente. El color se puntúo con 1,96±0,43 y el marmoleado del longissimus dorsi (LD) se valoró con 2,04±0,54 según las cartillas estándar de National Pork Producers Council (NPPC, 2000), respectivamente. Los perfiles de lípidos del LD y de la grasa subcutánea no fueron afectados por las dietas (P>0,05). Se obtu-vieron en promedio un 40% de AGS, 46% de AGMI y 14% AGPI y un 42% AGS, 37,21% AGMI y un 19,79% AGPI para la grasa intramuscular del LD y la grasa subcutánea, respectivamente. El análisis económico de las dietas utilizadas determinó que, en el contexto económico actual, la utilización del expeller de soja disminuye los costos económicos de alimentación sin afectar rendimientos productivos, características de la canal ni calidad de la carne.
Palabras clave: ácidos grasos, calidad de la canal, rendimiento productivo.
ABSTRACT
This study evaluated the effect of replacing soybean meal for press extruded soy beans (ExpS) in the diet on growth performance, meat quality parameters and fatty acid composition of the longissimus dorsi muscle of growing - fattening pigs. The experiment used 32 pigs. Eight animals were determined for each of following treatments: D1 (control diet) diet based on soybean meal with no ExpS, D2 which included 33.3% of ExpS and 67.3% of soybean meal, D3 which included 67.3% of ExpS and 33.3% of soybean meal, and D4 which inclu-ded 100% ExpS. No treatment effects or significant contrasts were detected (P>0.05) for the parameters eva-luated. Average daily weight gain was 895.5±0.04 g, the overall all average of gain: feed ratio was 2.44±0.33 and the overall lean yield was 53±0.86% across treatments. Diet treatments did not affect (P>0.05) pH at 45 min and ultimate pH (5.95±0.22 and 5.74±0.21, respectively across treatments). Color and marbling scores of
1Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Anguil, Ruta Nacional N.o 5, km 580 (6326), La Pampa. Universidad Nacional de la Pampa. Correo electrónico: [email protected] Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Anguil, Ruta Nacional N.o 5, km 580 (6326), La Pampa. Correo electrónico: [email protected]
MURCIA, V.N.1; PORDOMINGO, A.J.1; PORDOMINGO, A.B.1; FRANCO, R.2
Recibido 04 de diciembre de 2019 // Aceptado 23 de abril de 2020 // Publicado online 02 de septiembre de 2020

MURCIA, V.N.; PORDOMINGO, A.J.; PORDOMINGO, A.B.; FRANCO, R.
Agosto 2020, Argentina 241241
the LD muscle were 1.96±0.43; 2.04±0.54, respectively. Likewise, no differences were detected (P>0.05) for the lipid groups and main single lipids of LD intramuscular fat or subcutaneous fat longissimus dorsi intramus-cular lipid proportions of total lipids were comprised of 40% SFA, 46%, MUFA, 14% PUFA. Subcutaneous fat proportions were 42% SFA, 37.21% MUFA and 19.79% PUFA. In conclusion, this study suggests that press-extruded soybeans fed in the diet would have a similar response compared with soybean meal on performance and meat quality attributes in growing –fattening pig programs. The substitution would be cost effective.
Keywords: fatty acids, carcass quality, grown performance.
INTRODUCCIÓN
El 90-93% de la producción de poroto de soja de Ar-gentina es procesado por empresas aceiteras, generando harina y expeller de soja como subproductos del proceso (Juan et al., 2015). Ambos son alimentos de alto valor nu-tritivo porque representan la fuente principal de proteínas y de aminoácidos esenciales de monogástricos y rumiantes (Gallardo, 2005). Sin embargo, según el sistema de extrac-ción de aceite utilizado, existen diferencias nutricionales en cuanto al contenido de aceites remanentes, 9% y <2% en expeller y harina de soja, respectivamente (Lawrence et al., 2003; Méndez et al., 2010; Juan et al., 2015).
La industria porcina ha ido cambiando según las pre-ferencias de los consumidores, destinando la producción al consumo de carnes seguras, saludables y sabrosas (Cheng et al., 2017), seleccionando genotipos magros y ali-mentándolos con dietas que contienen grasas insaturadas (Viljoen y Ras, 1991).
Si bien, el incremento de ácido linolénico (ALA C18:3 n-3) y otros ácidos grasos poliinsaturados (AGPI) resultan beneficiosos para la salud, son indeseables en el procesa-miento de la carne porcina porque generan grasa blanda que dificultan la manipulación e incrementan la rancidez (Averette Gatlin et al., 2002a). Teniendo en cuenta que en Argentina existe escasa o nula información sobre los efec-tos de la inclusión del expeller de soja en la dieta de cerdos recriados y engordados es que cobra importancia conocer el rendimiento productivo y características de la carne de los cerdos alimentados con expeller de soja como suple-mento proteico.
El objetivo del trabajo fue determinar la respuesta pro-ductiva y económica, y la calidad de la canal y de la carne de cerdos recriados y engordados con diferentes niveles de inclusión del expeller de soja en la dieta en reemplazo de harina de soja para lo cual, se plantean los siguientes objetivos particulares: (1) evaluar el efecto de la inclusión de expeller de soja en reemplazo de harina de soja en la dieta sobre el rendimiento, las características de la carcasa y respuesta animal; (2) analizar el perfil de ácidos grasos del longissimus dorsi (LD) y grasa subcutánea de los ani-males alimentados con diferentes porcentajes de inclusión del expeller de soja en la dieta.
MATERIALES Y MÉTODOS
El ensayo de campo se llevó a cabo en la Estación Ex-perimental Agropecuaria INTA Marcos Juárez, Córdoba. Tuvo una duración de 70 días, comprendiendo los meses noviembre de 2017 hasta febrero de 2018. La faena de los animales utilizados se realizó en las instalaciones del frigorífico Mattievich de la localidad de Carcarañá, San-ta Fe. Las determinaciones de calidad de carne se rea-lizaron en las instalaciones del laboratorio de calidad de producto de la Estación Experimental Agropecuaria INTA Anguil, La Pampa.
Se utilizaron 36 cerdos, procedentes de la progenie de cruzamientos de madres híbridas con padrillos terminales de empresas genéticas, con un peso vivo inicial promedio del individuo de 40±2 kg hasta un peso vivo final superior a los 100 kg. Estos se encontraban en buen estado sanita-rio, inmunizados contra enfermedades infecto-contagiosas y desparasitados. Los animales fueron alojados al azar en los corrales de alimentación de a 3 por corral, hasta el momento de la faena y alimentados ad libitum con acceso libre de agua. Previamente al inicio del ensayo tuvieron un periodo de adaptación de siete días. Se utilizaron tres co-rrales por tratamiento, los cuales estaban equipados con comederos tolva, bebederos chupetes y piso ranurado.
Se trabajó con cuatro dietas isoproteicas (19 y 16% para recría y engorde, respectivamente) compuestas por maíz, expeller y harina de soja y el núcleo vitamínico mineral. La composición química de los ingredientes utilizados se muestra en la tabla 1. Las raciones se formularon cubrien-do los requerimientos nutricionales de acuerdo a NRC (2012) para cerdos en recría (40 a 70 kg PV) y engorde (70 a 105 kg PV). Las dietas se definieron por el nivel de sustitución de harina de soja por expeller de soja (tabla 2). Las dietas fueron identificadas como: D0 = 100% harina de soja (dieta control); D33 = 33,3% expeller de soja, 66,7% harina de soja; D66 = 66,7% expeller de soja, 33,3% harina de soja y la D100 = 100% expeller de soja.
Los animales fueron pesados individualmente con balan-za electrónica al iniciar el ensayo y luego cada 15 días has-ta alcanzar el peso de faena predeterminado. A partir de los valores de PV se calculó la ganancia media diaria (GMD, kg/día) por diferencia de peso entre pesadas.
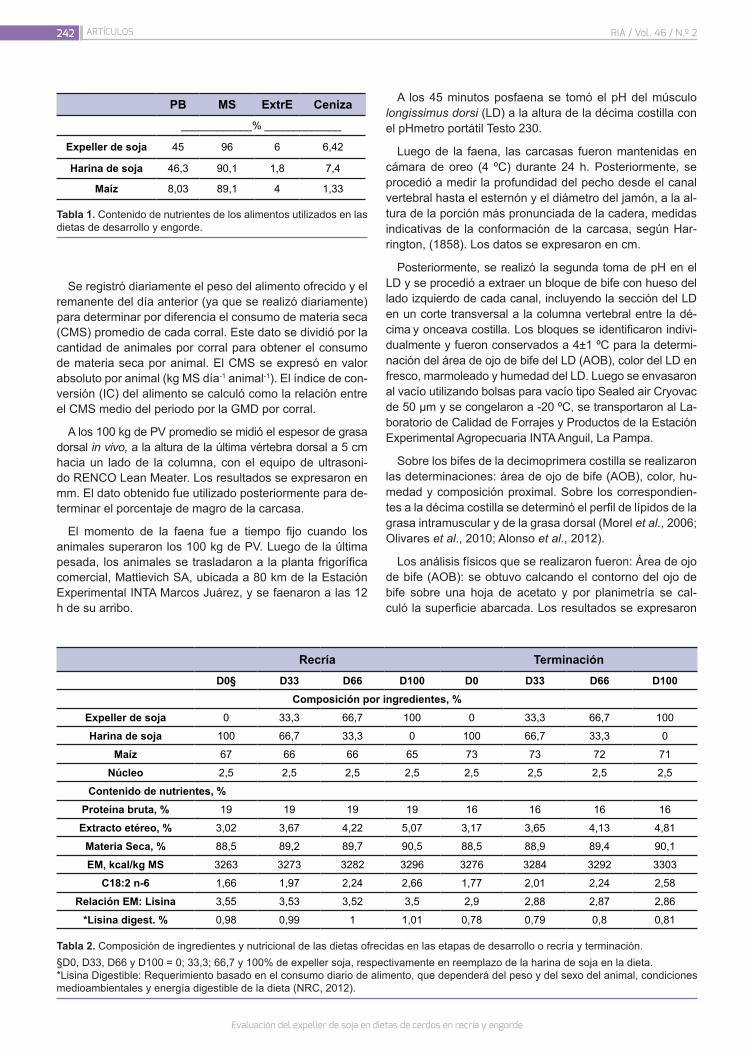
Evaluación del expeller de soja en dietas de cerdos en recría y engorde
ARTÍCULOS RIA / Vol. 46 / N.º 2242242
Se registró diariamente el peso del alimento ofrecido y el remanente del día anterior (ya que se realizó diariamente) para determinar por diferencia el consumo de materia seca (CMS) promedio de cada corral. Este dato se dividió por la cantidad de animales por corral para obtener el consumo de materia seca por animal. El CMS se expresó en valor absoluto por animal (kg MS día-1 animal-1). El índice de con-versión (IC) del alimento se calculó como la relación entre el CMS medio del periodo por la GMD por corral.
A los 100 kg de PV promedio se midió el espesor de grasa dorsal in vivo, a la altura de la última vértebra dorsal a 5 cm hacia un lado de la columna, con el equipo de ultrasoni-do RENCO Lean Meater. Los resultados se expresaron en mm. El dato obtenido fue utilizado posteriormente para de-terminar el porcentaje de magro de la carcasa.
El momento de la faena fue a tiempo fijo cuando los animales superaron los 100 kg de PV. Luego de la última pesada, los animales se trasladaron a la planta frigorífica comercial, Mattievich SA, ubicada a 80 km de la Estación Experimental INTA Marcos Juárez, y se faenaron a las 12 h de su arribo.
Tabla 1. Contenido de nutrientes de los alimentos utilizados en las dietas de desarrollo y engorde.
A los 45 minutos posfaena se tomó el pH del músculo longissimus dorsi (LD) a la altura de la décima costilla con el pHmetro portátil Testo 230.
Luego de la faena, las carcasas fueron mantenidas en cámara de oreo (4 ºC) durante 24 h. Posteriormente, se procedió a medir la profundidad del pecho desde el canal vertebral hasta el esternón y el diámetro del jamón, a la al-tura de la porción más pronunciada de la cadera, medidas indicativas de la conformación de la carcasa, según Har-rington, (1858). Los datos se expresaron en cm.
Posteriormente, se realizó la segunda toma de pH en el LD y se procedió a extraer un bloque de bife con hueso del lado izquierdo de cada canal, incluyendo la sección del LD en un corte transversal a la columna vertebral entre la dé-cima y onceava costilla. Los bloques se identificaron indivi-dualmente y fueron conservados a 4±1 ºC para la determi-nación del área de ojo de bife del LD (AOB), color del LD en fresco, marmoleado y humedad del LD. Luego se envasaron al vacío utilizando bolsas para vacío tipo Sealed air Cryovac de 50 µm y se congelaron a -20 ºC, se transportaron al La-boratorio de Calidad de Forrajes y Productos de la Estación Experimental Agropecuaria INTA Anguil, La Pampa.
Sobre los bifes de la decimoprimera costilla se realizaron las determinaciones: área de ojo de bife (AOB), color, hu-medad y composición proximal. Sobre los correspondien-tes a la décima costilla se determinó el perfil de lípidos de la grasa intramuscular y de la grasa dorsal (Morel et al., 2006; Olivares et al., 2010; Alonso et al., 2012).
Los análisis físicos que se realizaron fueron: Área de ojo de bife (AOB): se obtuvo calcando el contorno del ojo de bife sobre una hoja de acetato y por planimetría se cal-culó la superficie abarcada. Los resultados se expresaron
PB MS ExtrE Ceniza____________% _____________
Expeller de soja 45 96 6 6,42
Harina de soja 46,3 90,1 1,8 7,4
Maíz 8,03 89,1 4 1,33
Recría Terminación D0§ D33 D66 D100 D0 D33 D66 D100
Composición por ingredientes, %Expeller de soja 0 33,3 66,7 100 0 33,3 66,7 100
Harina de soja 100 66,7 33,3 0 100 66,7 33,3 0
Maíz 67 66 66 65 73 73 72 71
Núcleo 2,5 2,5 2,5 2,5 2,5 2,5 2,5 2,5
Contenido de nutrientes, %Proteína bruta, % 19 19 19 19 16 16 16 16
Extracto etéreo, % 3,02 3,67 4,22 5,07 3,17 3,65 4,13 4,81
Materia Seca, % 88,5 89,2 89,7 90,5 88,5 88,9 89,4 90,1
EM, kcal/kg MS 3263 3273 3282 3296 3276 3284 3292 3303
C18:2 n-6 1,66 1,97 2,24 2,66 1,77 2,01 2,24 2,58
Relación EM: Lisina 3,55 3,53 3,52 3,5 2,9 2,88 2,87 2,86
*Lisina digest. % 0,98 0,99 1 1,01 0,78 0,79 0,8 0,81
Tabla 2. Composición de ingredientes y nutricional de las dietas ofrecidas en las etapas de desarrollo o recría y terminación.§D0, D33, D66 y D100 = 0; 33,3; 66,7 y 100% de expeller soja, respectivamente en reemplazo de la harina de soja en la dieta.*Lisina Digestible: Requerimiento basado en el consumo diario de alimento, que dependerá del peso y del sexo del animal, condiciones medioambientales y energía digestible de la dieta (NRC, 2012).

MURCIA, V.N.; PORDOMINGO, A.J.; PORDOMINGO, A.B.; FRANCO, R.
Agosto 2020, Argentina 243243
en cm2. Color del LD: se determinó mediante score visual utilizando las cartillas estándar de puntuación de National Pork Producers Council (NPPC, 2000), posterior al tiempo de desarrollo del blooming. La escala de las cartillas (rango 1-6) comienza en las carnes más pálidas hasta las más oscuras pasando por los colores intermedios. Marmoleado del LD: se evaluó la cantidad de grasa dispuestas entre las fibras musculares del LD tomando como referencia el rango de 1 a 10 de las cartillas estándar de puntuación de National Pork Producers Council (NPPC, 2000).
Para realizar los análisis bioquímicos previstos los bifes fueron descongelados a 4±1 oC durante 24 horas (Almada et al., 2011). Se apartó la grasa externa y el tejido conec-tivo de los bifes seleccionados para las determinaciones de composición proximal y perfil de lípidos. Posteriormente las muestras de LD se molieron utilizando una procesadora para carne. Una alícuota se utilizó para la determinación de humedad obtenida por diferencia de peso después de 24 h en una estufa a 100 ºC. El resto fue liofilizado en el liofilizador Labconco (EE. UU., Kansas).
Los contenidos de humedad, proteína, ceniza y grasa se determinaron acorde a los métodos descriptos por la AOAC (2000). Los valores de PB se determinaron por el método Kjeldahl (contenido de nitrógeno por el factor 6,25). El con-tenido de ceniza se obtuvo mediante incineración en una mufla durante 8 h a 600 ºC. Los lípidos totales se extraje-ron con hexano en ebullición en un equipo Tecator según el método Soxhlet (AOAC, 2005). Todos los resultados se expresaron en porcentaje.
Sobre una alícuota de las muestras liofilizadas se rea-lizó la transmetilación de los lípidos según el método de Park y Goins (1994). Los ácidos grasos metilados se ana-lizaron en un cromatógrafo gaseoso Perkin Elmer CLA-RUS 600, equipado con un detector de ionización a llama (FID). La separación de las muestras se realizó con una columna Agilent Technologies CP-Sil88 (100 m x 0,25 mm id) y nitrógeno como gas portador. La temperatura inicial del horno fue 150 oC/min, luego se programaron 3 rampas de temperatura. La rampa 1 incrementó de 2 ºC/min hasta 174 durante 0 min; la rampa 2: incrementó de 0,2 °C /min hasta 178 °C por 0 min y la rampa 3: incrementó de 2 °C /min hasta 225 durante 7,5 min acorde con Duckett et al. (2013). Las muestras se corrieron por duplicado, con una relación de split de 100:1 para ácidos grasos de cadena larga y trans C18:1 y una relación de split 10:1 para los isómeros CLA y ácidos grasos de cadena larga omega 3. Los ácidos grasos individuales se identificaron por compa-ración de los tiempos de retención con estándares Supel-co Mix 37 (47885-U Supelco –Aldrich) y se cuantificaron incorporando un estándar interno (ácido metiltricosanoic, C23:0) dentro de cada muestra durante la metilación. Los ácidos grasos individuales se agruparon en: AGS [mi-rístico (C14:0) + palmítico (C16:0) + esteárico (C18:0)]; AGMI [palmitoleico (C16:1) + oleico (C18:1 cis 9) + C18:1 cis 11]; AGPI n-3 [linolénico (C18:3) EPA (C20:5) + DPA (C22:5) + DHA (C22:6)]; AGPI n-6 [α-linoleico (C18:2) + Araquidónico (C20:4)] y AGPI [AGPI n-3 + AGPI n-6]. Los resultados se expresaron como porcentaje en peso del
total de ácidos grasos y se determinaron las relaciones AGPI: AGS y n-6: n-3.
Los animales se distribuyeron al azar en 12 corrales, de a 3 por corral. Los corrales fueron las unidades experimen-tales sobre las cuales se aleatorizaron los cuatro tratamien-tos, se generaron 3 repeticiones por tratamiento. El análisis estadístico para todas las variables se realizó utilizando el procedimiento PROC GLM (SAS, 1999). Se aplicaron con-trastes ortogonales (Bonferroni, 1936).
RESULTADOS Y DISCUSIÓN
En la tabla 3 se sintetizaron los resultados de respues-ta productiva y de calidad de la carcasa. No se encontra-ron diferencias significativas en los parámetros evaluados (P>0,05) entre ninguno de los tratamientos. En promedio de todos los tratamientos los animales ingresaron con un peso inicial de 40,9±1,2 kg y alcanzaron 103,6±3,4 kg de peso final. Tuvieron una GMD de 895,5±0,04 g, un IC promedio de 2,4±0,3 y el 53±0,8% de magro. Resultados similares de crecimiento y engorde fueron obtenidos por Olivares et al. (2010) quienes utilizaron dietas formuladas a base de maíz y trigo con el agregado de ácidos grasos sa-turados (aceite de palma) o con ácidos grasos insaturados (soja extruida rica en C18:2 n 6).
Sin embargo, Averette Gatlin et al. (2002b) trabajaron con dietas establecidas por el porcentaje de inclusión del acei-te de soja en un 0; 33,3; 66,7; 100%. Las variaciones en las concentraciones de AL (C18:2 n 6) fueron desde 4,11 al 1,56% del total de la dieta, reportaron disminución del ren-dimiento productivo; mientras que Cheng et al. (2017), Teye et al. (2006) y Wiseman y Agunbiade (1998) no encontra-ron diferencias con la adición de ingredientes ricos en AL (C18:2 n 6) como el aceite de orégano, palma, poroto de soja sobre el rendimiento productivo. De la misma mane-ra, Landero et al. (2012) y Lawrence et al. (2003) evaluaron nutricionalmente la harina de soja obtenida por solvente y por prensado sin encontrar diferencias en el rendimiento en animales de recría y engorde. Por un lado, la dieta control no contenía harina de soja, mientras que las otras estuvieron determinadas por la inclusión del 20 y 40% de harina de soja o expeller de soja, en reemplazo de la harina de pescado y de sangre. Por otro lado, el rendimiento de la carcasa, el es-pesor de grasa dorsal y el pH muscular no fueron afectados por el remplazo de la harina de soja por el expeller de soja. Del mismo modo ocurrió en el trabajo reportado por Teye et al. (2006) quienes utilizaron aceites de diferentes materias primas ricas en el contenido de AL (C18:2 n 6).
En la tabla 4 se representan las variables evaluadas para las características físicas y químicas del LD. No presenta-ron diferencias (P>0,05) entre los tratamientos utilizados: D0= 100% harina de soja (dieta control); D33 = 33,3% ex-peller de soja, 66,7% harina de soja; D66 = 66,7% expeller de soja, 33,3% harina de soja y la D100 = 100% expeller de soja. Los valores promedios de pH a los 45 min y 24 h post mórtem fueron de 5,9 ± 0,22 y 5,7 ± 0,21, respectivamente. Estos valores encontrados de pH a las 24 h coinciden con
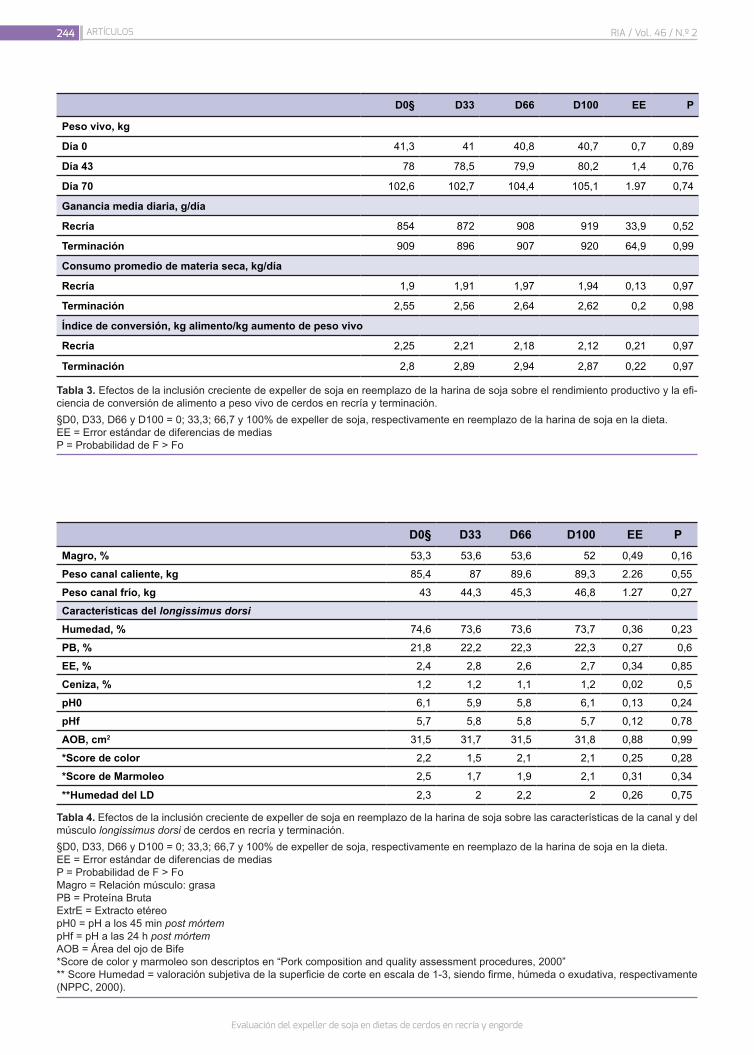
Evaluación del expeller de soja en dietas de cerdos en recría y engorde
ARTÍCULOS RIA / Vol. 46 / N.º 2244244
Tabla 3. Efectos de la inclusión creciente de expeller de soja en reemplazo de la harina de soja sobre el rendimiento productivo y la efi-ciencia de conversión de alimento a peso vivo de cerdos en recría y terminación.§D0, D33, D66 y D100 = 0; 33,3; 66,7 y 100% de expeller de soja, respectivamente en reemplazo de la harina de soja en la dieta.EE = Error estándar de diferencias de mediasP = Probabilidad de F > Fo
Tabla 4. Efectos de la inclusión creciente de expeller de soja en reemplazo de la harina de soja sobre las características de la canal y del músculo longissimus dorsi de cerdos en recría y terminación.§D0, D33, D66 y D100 = 0; 33,3; 66,7 y 100% de expeller de soja, respectivamente en reemplazo de la harina de soja en la dieta.EE = Error estándar de diferencias de mediasP = Probabilidad de F > FoMagro = Relación músculo: grasaPB = Proteína BrutaExtrE = Extracto etéreopH0 = pH a los 45 min post mórtempHf = pH a las 24 h post mórtemAOB = Área del ojo de Bife*Score de color y marmoleo son descriptos en “Pork composition and quality assessment procedures, 2000”** Score Humedad = valoración subjetiva de la superficie de corte en escala de 1-3, siendo firme, húmeda o exudativa, respectivamente (NPPC, 2000).
D0§ D33 D66 D100 EE P
Peso vivo, kg
Día 0 41,3 41 40,8 40,7 0,7 0,89
Día 43 78 78,5 79,9 80,2 1,4 0,76
Día 70 102,6 102,7 104,4 105,1 1.97 0,74
Ganancia media diaria, g/día
Recría 854 872 908 919 33,9 0,52
Terminación 909 896 907 920 64,9 0,99
Consumo promedio de materia seca, kg/día
Recría 1,9 1,91 1,97 1,94 0,13 0,97
Terminación 2,55 2,56 2,64 2,62 0,2 0,98
Índice de conversión, kg alimento/kg aumento de peso vivo
Recría 2,25 2,21 2,18 2,12 0,21 0,97
Terminación 2,8 2,89 2,94 2,87 0,22 0,97
D0§ D33 D66 D100 EE P Magro, % 53,3 53,6 53,6 52 0,49 0,16
Peso canal caliente, kg 85,4 87 89,6 89,3 2.26 0,55
Peso canal frío, kg 43 44,3 45,3 46,8 1.27 0,27
Características del longissimus dorsiHumedad, % 74,6 73,6 73,6 73,7 0,36 0,23
PB, % 21,8 22,2 22,3 22,3 0,27 0,6
EE, % 2,4 2,8 2,6 2,7 0,34 0,85
Ceniza, % 1,2 1,2 1,1 1,2 0,02 0,5
pH0 6,1 5,9 5,8 6,1 0,13 0,24
pHf 5,7 5,8 5,8 5,7 0,12 0,78
AOB, cm2 31,5 31,7 31,5 31,8 0,88 0,99
*Score de color 2,2 1,5 2,1 2,1 0,25 0,28
*Score de Marmoleo 2,5 1,7 1,9 2,1 0,31 0,34
**Humedad del LD 2,3 2 2,2 2 0,26 0,75
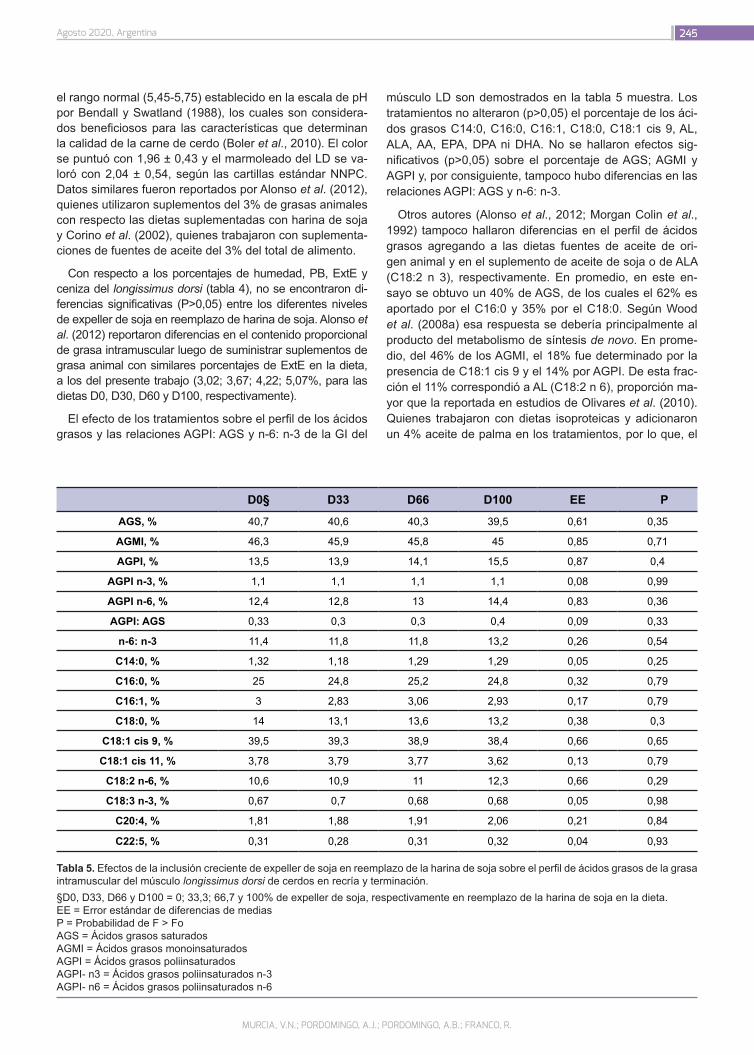
MURCIA, V.N.; PORDOMINGO, A.J.; PORDOMINGO, A.B.; FRANCO, R.
Agosto 2020, Argentina 245245
el rango normal (5,45-5,75) establecido en la escala de pH por Bendall y Swatland (1988), los cuales son considera-dos beneficiosos para las características que determinan la calidad de la carne de cerdo (Boler et al., 2010). El color se puntuó con 1,96 ± 0,43 y el marmoleado del LD se va-loró con 2,04 ± 0,54, según las cartillas estándar NNPC. Datos similares fueron reportados por Alonso et al. (2012), quienes utilizaron suplementos del 3% de grasas animales con respecto las dietas suplementadas con harina de soja y Corino et al. (2002), quienes trabajaron con suplementa-ciones de fuentes de aceite del 3% del total de alimento.
Con respecto a los porcentajes de humedad, PB, ExtE y ceniza del longissimus dorsi (tabla 4), no se encontraron di-ferencias significativas (P>0,05) entre los diferentes niveles de expeller de soja en reemplazo de harina de soja. Alonso et al. (2012) reportaron diferencias en el contenido proporcional de grasa intramuscular luego de suministrar suplementos de grasa animal con similares porcentajes de ExtE en la dieta, a los del presente trabajo (3,02; 3,67; 4,22; 5,07%, para las dietas D0, D30, D60 y D100, respectivamente).
El efecto de los tratamientos sobre el perfil de los ácidos grasos y las relaciones AGPI: AGS y n-6: n-3 de la GI del
músculo LD son demostrados en la tabla 5 muestra. Los tratamientos no alteraron (p>0,05) el porcentaje de los áci-dos grasos C14:0, C16:0, C16:1, C18:0, C18:1 cis 9, AL, ALA, AA, EPA, DPA ni DHA. No se hallaron efectos sig-nificativos (p>0,05) sobre el porcentaje de AGS; AGMI y AGPI y, por consiguiente, tampoco hubo diferencias en las relaciones AGPI: AGS y n-6: n-3.
Otros autores (Alonso et al., 2012; Morgan Colin et al., 1992) tampoco hallaron diferencias en el perfil de ácidos grasos agregando a las dietas fuentes de aceite de ori-gen animal y en el suplemento de aceite de soja o de ALA (C18:2 n 3), respectivamente. En promedio, en este en-sayo se obtuvo un 40% de AGS, de los cuales el 62% es aportado por el C16:0 y 35% por el C18:0. Según Wood et al. (2008a) esa respuesta se debería principalmente al producto del metabolismo de síntesis de novo. En prome-dio, del 46% de los AGMI, el 18% fue determinado por la presencia de C18:1 cis 9 y el 14% por AGPI. De esta frac-ción el 11% correspondió a AL (C18:2 n 6), proporción ma-yor que la reportada en estudios de Olivares et al. (2010). Quienes trabajaron con dietas isoproteicas y adicionaron un 4% aceite de palma en los tratamientos, por lo que, el
Tabla 5. Efectos de la inclusión creciente de expeller de soja en reemplazo de la harina de soja sobre el perfil de ácidos grasos de la grasa intramuscular del músculo longissimus dorsi de cerdos en recría y terminación.§D0, D33, D66 y D100 = 0; 33,3; 66,7 y 100% de expeller de soja, respectivamente en reemplazo de la harina de soja en la dieta.EE = Error estándar de diferencias de mediasP = Probabilidad de F > FoAGS = Ácidos grasos saturadosAGMI = Ácidos grasos monoinsaturadosAGPI = Ácidos grasos poliinsaturadosAGPI- n3 = Ácidos grasos poliinsaturados n-3AGPI- n6 = Ácidos grasos poliinsaturados n-6
D0§ D33 D66 D100 EE P AGS, % 40,7 40,6 40,3 39,5 0,61 0,35
AGMI, % 46,3 45,9 45,8 45 0,85 0,71
AGPI, % 13,5 13,9 14,1 15,5 0,87 0,4
AGPI n-3, % 1,1 1,1 1,1 1,1 0,08 0,99
AGPI n-6, % 12,4 12,8 13 14,4 0,83 0,36
AGPI: AGS 0,33 0,3 0,3 0,4 0,09 0,33
n-6: n-3 11,4 11,8 11,8 13,2 0,26 0,54
C14:0, % 1,32 1,18 1,29 1,29 0,05 0,25
C16:0, % 25 24,8 25,2 24,8 0,32 0,79
C16:1, % 3 2,83 3,06 2,93 0,17 0,79
C18:0, % 14 13,1 13,6 13,2 0,38 0,3
C18:1 cis 9, % 39,5 39,3 38,9 38,4 0,66 0,65
C18:1 cis 11, % 3,78 3,79 3,77 3,62 0,13 0,79
C18:2 n-6, % 10,6 10,9 11 12,3 0,66 0,29
C18:3 n-3, % 0,67 0,7 0,68 0,68 0,05 0,98
C20:4, % 1,81 1,88 1,91 2,06 0,21 0,84
C22:5, % 0,31 0,28 0,31 0,32 0,04 0,93
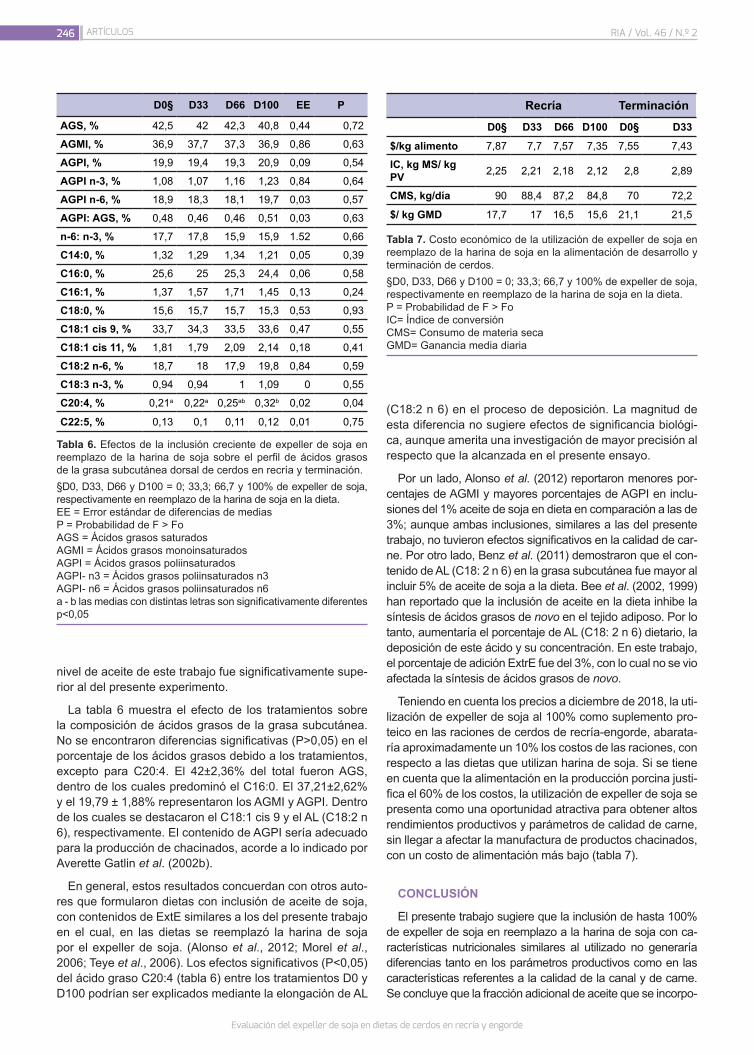
Evaluación del expeller de soja en dietas de cerdos en recría y engorde
ARTÍCULOS RIA / Vol. 46 / N.º 2246246
(C18:2 n 6) en el proceso de deposición. La magnitud de esta diferencia no sugiere efectos de significancia biológi-ca, aunque amerita una investigación de mayor precisión al respecto que la alcanzada en el presente ensayo.
Por un lado, Alonso et al. (2012) reportaron menores por-centajes de AGMI y mayores porcentajes de AGPI en inclu-siones del 1% aceite de soja en dieta en comparación a las de 3%; aunque ambas inclusiones, similares a las del presente trabajo, no tuvieron efectos significativos en la calidad de car-ne. Por otro lado, Benz et al. (2011) demostraron que el con-tenido de AL (C18: 2 n 6) en la grasa subcutánea fue mayor al incluir 5% de aceite de soja a la dieta. Bee et al. (2002, 1999) han reportado que la inclusión de aceite en la dieta inhibe la síntesis de ácidos grasos de novo en el tejido adiposo. Por lo tanto, aumentaría el porcentaje de AL (C18: 2 n 6) dietario, la deposición de este ácido y su concentración. En este trabajo, el porcentaje de adición ExtrE fue del 3%, con lo cual no se vio afectada la síntesis de ácidos grasos de novo.
Teniendo en cuenta los precios a diciembre de 2018, la uti-lización de expeller de soja al 100% como suplemento pro-teico en las raciones de cerdos de recría-engorde, abarata-ría aproximadamente un 10% los costos de las raciones, con respecto a las dietas que utilizan harina de soja. Si se tiene en cuenta que la alimentación en la producción porcina justi-fica el 60% de los costos, la utilización de expeller de soja se presenta como una oportunidad atractiva para obtener altos rendimientos productivos y parámetros de calidad de carne, sin llegar a afectar la manufactura de productos chacinados, con un costo de alimentación más bajo (tabla 7).
CONCLUSIÓN
El presente trabajo sugiere que la inclusión de hasta 100% de expeller de soja en reemplazo a la harina de soja con ca-racterísticas nutricionales similares al utilizado no generaría diferencias tanto en los parámetros productivos como en las características referentes a la calidad de la canal y de carne. Se concluye que la fracción adicional de aceite que se incorpo-
D0§ D33 D66 D100 EE P
AGS, % 42,5 42 42,3 40,8 0,44 0,72
AGMI, % 36,9 37,7 37,3 36,9 0,86 0,63
AGPI, % 19,9 19,4 19,3 20,9 0,09 0,54
AGPI n-3, % 1,08 1,07 1,16 1,23 0,84 0,64
AGPI n-6, % 18,9 18,3 18,1 19,7 0,03 0,57
AGPI: AGS, % 0,48 0,46 0,46 0,51 0,03 0,63
n-6: n-3, % 17,7 17,8 15,9 15,9 1.52 0,66
C14:0, % 1,32 1,29 1,34 1,21 0,05 0,39
C16:0, % 25,6 25 25,3 24,4 0,06 0,58
C16:1, % 1,37 1,57 1,71 1,45 0,13 0,24
C18:0, % 15,6 15,7 15,7 15,3 0,53 0,93
C18:1 cis 9, % 33,7 34,3 33,5 33,6 0,47 0,55
C18:1 cis 11, % 1,81 1,79 2,09 2,14 0,18 0,41
C18:2 n-6, % 18,7 18 17,9 19,8 0,84 0,59
C18:3 n-3, % 0,94 0,94 1 1,09 0 0,55
C20:4, % 0,21a 0,22a 0,25ab 0,32b 0,02 0,04
C22:5, % 0,13 0,1 0,11 0,12 0,01 0,75
Recría Terminación D0§ D33 D66 D100 D0§ D33$/kg alimento 7,87 7,7 7,57 7,35 7,55 7,43
IC, kg MS/ kg PV 2,25 2,21 2,18 2,12 2,8 2,89
CMS, kg/día 90 88,4 87,2 84,8 70 72,2
$/ kg GMD 17,7 17 16,5 15,6 21,1 21,5
Tabla 6. Efectos de la inclusión creciente de expeller de soja en reemplazo de la harina de soja sobre el perfil de ácidos grasos de la grasa subcutánea dorsal de cerdos en recría y terminación.§D0, D33, D66 y D100 = 0; 33,3; 66,7 y 100% de expeller de soja, respectivamente en reemplazo de la harina de soja en la dieta.EE = Error estándar de diferencias de mediasP = Probabilidad de F > FoAGS = Ácidos grasos saturadosAGMI = Ácidos grasos monoinsaturadosAGPI = Ácidos grasos poliinsaturadosAGPI- n3 = Ácidos grasos poliinsaturados n3AGPI- n6 = Ácidos grasos poliinsaturados n6a - b las medias con distintas letras son significativamente diferentes p<0,05
Tabla 7. Costo económico de la utilización de expeller de soja en reemplazo de la harina de soja en la alimentación de desarrollo y terminación de cerdos.§D0, D33, D66 y D100 = 0; 33,3; 66,7 y 100% de expeller de soja, respectivamente en reemplazo de la harina de soja en la dieta.P = Probabilidad de F > FoIC= Índice de conversiónCMS= Consumo de materia secaGMD= Ganancia media diaria
nivel de aceite de este trabajo fue significativamente supe-rior al del presente experimento.
La tabla 6 muestra el efecto de los tratamientos sobre la composición de ácidos grasos de la grasa subcutánea. No se encontraron diferencias significativas (P>0,05) en el porcentaje de los ácidos grasos debido a los tratamientos, excepto para C20:4. El 42±2,36% del total fueron AGS, dentro de los cuales predominó el C16:0. El 37,21±2,62% y el 19,79 ± 1,88% representaron los AGMI y AGPI. Dentro de los cuales se destacaron el C18:1 cis 9 y el AL (C18:2 n 6), respectivamente. El contenido de AGPI sería adecuado para la producción de chacinados, acorde a lo indicado por Averette Gatlin et al. (2002b).
En general, estos resultados concuerdan con otros auto-res que formularon dietas con inclusión de aceite de soja, con contenidos de ExtE similares a los del presente trabajo en el cual, en las dietas se reemplazó la harina de soja por el expeller de soja. (Alonso et al., 2012; Morel et al., 2006; Teye et al., 2006). Los efectos significativos (P<0,05) del ácido graso C20:4 (tabla 6) entre los tratamientos D0 y D100 podrían ser explicados mediante la elongación de AL

MURCIA, V.N.; PORDOMINGO, A.J.; PORDOMINGO, A.B.; FRANCO, R.
Agosto 2020, Argentina 247247
ra con el aporte del expeller no tendría efectos de significancia sobre las características físicas de la grasa o de la composi-ción de los lípidos musculares o subcutáneos. En el contexto de las dietas ofrecidas, la inclusión de expeller de soja respec-to de la harina permitiría una reducción del 10% de los costos de alimentación por lo que su uso sería recomendable.
BIBLIOGRAFÍAALMADA, C.A.; BONATO, P.I.; CARDUZA, F.; COSSU, M.E.;
GRIGIONE, G.M.; IRURUETA, M.; PERLO, F.; PICALLO, A.B.; TEIRA, G.A. 2011. Manual de procedimientos. Determinación de los parámetros de calidad física y sensorial de carne bovina. INTA-UBA-UN Lujan-UNNER. Ed. INTA.
ALONSO, V.; NAJES, L.M.; PROVINCIAL, L.; GUILLÉN, E.; GIL, M.;. RONCALÉS, P.; BELTRÁN, J.A. 2012. Influence of dietary fat on pork eating quality. Meat Science 92(4): 366-73.
AOAC. 2000. Official methods of analysis. 17th ed. Assoc. of Off. Anal. Chem., Washington, D. C.
AOAC. 2005. Official methods of analysis. 18th ed. 4th supple-ment, Arlington, VA, EUA.
AVERETTE GATLIN, L.; SEE, M.T.; LARICK, D.K.; LIN, X.; ODLE, J. 2002a. Conjugated linoleic acid in combination with sup-plemental dietary fat alters pork fat quality. American Society for Nutritional Sciences.132 (10): 3105-12.
AVERETTE GATLIN, L.; SEE, M.T.; HANSEN, J.A.; SUTTON, D.; ODLE, J. 2002b. The effects of dietary fat sources, levels, and feeding intervals on pork fatty acid composition. Journal of Animal Science 80(6): 1606-15.
BEE, G.; MESSIKOMMER, R.; GEBERT, S. 1999. Dietary fats and energy levels differently affect lipogenic enzyme activity in fin-ishing pigs. Felt/Lipid 101:336-342.
BEE, G.; GEBERT, S.; MESSIKOMER, R. 2002. Effect of dietary energy supply and fat source on the fatty acid pattern of adipose and lean tissues and lipogenesis in the pig. Journal Animal Sci-ence. 80:1564-1574.
BENDALL, J.R.; SWATLAND, H.J. 1988. A review of the rela-tionships of pH with physical aspects of pork quality. Meat science 24(2), 85-126.
BENZ, J.M.; TOKACH, M.D.; DRITZ, S.S.; NELSSEN, J.L.; DER-OUCHEY, J.M.; SULABO, R.C.; GOODBAND, R.D. 2011. Effects of choice white grease and soybean oil on growth performance, carcass characteristics, and carcass fat quality of growing-finishing pigs. Journal of Animal Science 89(2): 404-13.
BOLER, D.D.; DILGER, A.C.; BIDNER, B.S.; CARR, S.N.; EGGERT,J.M.; DAY, J.W.; ELLIS, M.; MCKEITH,F.K.; KILLEFER, J. 2010. Ultimate pH explains variation in pork quality traits. Jour-nal of Muscle foods 21(217): 119-30.
BONFERRONI, C. 1936. Teoria statistica delle classi e calcolo delle probabilita. Pubblicazioni del R Istituto Superiore di Scienze Economiche e Commericiali di Firenze, 8, 3-62.
CHENG, C.; LIU, Z.; ZHOU, Y.; WEI, H.; ZHANG, X.; XIA, M. 2017. Effect of oregano essential oil supplementation to a re-duced-protein, amino acid-supplemented diet on meat quality, fatty acid composition, and oxidative stability of longissimus thoracic muscle in growing-finishing pigs. Meat Science 133: 103-9.
CORINO, C.; MAGNI, S.; PAGLIARINI, E.; ROSSI, R.; PASTOREL-LI, G.; CHIESA, L.M. 2002. Effects of dietary fats on meat quality and sensory characteristics of heavy pig loins. Meat Science 60(1): 1-8.
DUCKETT, S. K.; NEEL, J.P.S.; LEWIS, J.P.; FONTENOT, J.P.; CLAPHAM, W.P. 2013. Effects of winter stocker growth rate and
finishing system on: III. Tissue proximate, fatty acid, vitamin, and cholesterol content. Journal Animal Science. 91: 1454-1467.
GALLARDO, G. M. 2005.Soja: harinas de extracción para la ali-mentación del ganado. Un análisis de las cualidades nutricionales de los diferentes tipos, de acuerdo al método de extracción utilizado. Ins-tituto Nacional de Tecnología Agropecuario-INTA. Rafaela, Argentina.
HARRINGTON, G. 1958. Pig carcass evaluation. Common-wealth Agricultural Bureaus: England 1-119.
JUAN, N.A.; MASSIGOGUE, J.I.; ERRASQUIN, L.; MÉNDEZ, J.M.; OCHANDIO, D.C.; SAAVEDRA, M.C.; PAOLILLI, M.C.; AL-LADIO, R.M.; ACCORONI, C.; BEHR, E.F. 2015. Calidad de la soja procesada y de expeller producido por la industria de extrusado-prensado en Argentina. Ediciones INTA: 31.
LANDERO, J.L.; BELTRANENA, E.; CERVANTES, M.; ARAIZA, A.B.; ZIJLSTRA, R. 2012. The effect of feeding expeller-pressed canola meal on growth performance and diet nutrient digestibility in weaned pigs. Animal Feed Science and Technology 171(2-4): 240-45.
LAWRENCE, K.R.; GOODBAND, R.D.; TOKACH, S.S.; DRITZ NELSSEN, J.L.; DEROUCHEY, J.M.; HASTAD, C.W.; GROES-BECK, C.N. 2003. Effects of extruded-expelled soybean meal and solvent extracted soybean meal level of growth performance of weanling pigs 1, 2. Swine Day: 90-96.
MENDEZ, J.M.; COVACEVICH, M.; CAPURRO, J. 2010. Proce-samiento del grano de soja en la provincia de Santa Fe mediante extrusado y prensado. Para mejorar la Producción 45 47: 137-40.
MOREL, P.C.H.; MCINTOSH, J.C.; JANZ, J.A.M. 2006. Al-teration of the fatty acid profile of pork by dietary manipulation. Asian-Australasian Journal of Animal Sciences 19(3): 431-37.
MORGAN COLIN, A.; RAYMOND, N.; MASSIMO, C.; MCCA-RTNEY, R. 1992. Manipulation of the fatty acid composition of pig meat lipids by dietary means. Journal of the Science of Food and Agriculture 58(3): 357-68.
NATIONAL RESEARCH COUNCIL. 2012. Nutrient requirements of swine. National Academies Press.
NPPC. NATIONAL PORK PRODUCERS COUNCIL 2000. Pork composition and quality assessment procedures. In Natl. Des Moines, IA: Pork Board.
OLIVARES, A.; DAZA, A.; REY, A.I.; LÓPEZ-BOTE, C.J. 2010. Effect of diet saturation on growth performance, carcass character-istics and fat quality of heavy pigs. Food Science and Technology International 16(4): 321-27.
PARK, P.W.; GOINS, R.E. 1994. In situ preparation of fatty acid methyl esters for analysis of fatty acid composition in foods. Jour-nal of Food Science 59 (6): 1262-1266.
SAS. STATISCAL ANALYSIS SYSTEM. 1999. SAS User’s Guide: Statistics (Version 8). SAS Institute Inc., Cary, NC, EUA. 3884 p.
TEYE, G.A.; SHEARD, P.R.; WHITTINGTON, F.M.; NUTE, G.R.; STEWART, A.; WOOD, J.D. 2006. Influence of dietary oils and pro-tein level on pork quality. 1. Effects on muscle fatty acid composi-tion, carcass, meat and eating quality. Meat Science 73(1): 157-65.
VILJOEN, J.; RAS, M.N. 1991. The effect of different dietary fat sourc-es, withdrawal times, and castration on the fatty acid composition of back-fat in bacon pigs. South Africa. Journal Animal Science. 21: 76-79.
WISEMAN, J.; AGUNBIADE, J.A. 1998. The influence of chang-es in dietary fat and oils on fatty acid profiles of carcass fat in finish-ing pigs. Livestock Production Science 54(3): 217-27.
WOOD, J.D.; RICHARDSON, R.I.; NUTE, G.R.; FISHER, A.V.; CAMPO, M.M.; KASAPIDOU, E.; SHEARD, P.R.; ENSER, M. 2003. Effects of fatty acids on meat quality: a review. Meat Science 66(1): 21-32.

Transformación eficiente de Solanum tuberosum cv. Spunta mediada por Agrobacterium tumefaciens utilizandohigromicina como agente selectivo
ARTÍCULOS RIA / Vol. 46 / N.º 2248248
Transformación eficiente de Solanum tuberosum cv. Spunta mediada por Agrobacterium tumefaciens utilizando higromicina como agente selectivo
RESUMENLa transformación genética de plantas es una herramienta biotecnológica de alto impacto en el mejoramien-
to vegetal que brinda la posibilidad de obtener nuevos cultivos que se adapten a las demandas de producción y de consumo. Actualmente la estrategia de transformación más utilizada en papa se basa en la incorporación de genes completos vía Agrobacterium tumefaciens. El objetivo del presente trabajo fue desarrollar el primer protocolo de transformación para Solanum tuberosum cv. Spunta mediado por Agrobacterium tumefaciens utilizando el antibiótico higromicina como agente de selección. Para ello se transformaron explantes de papa cv. Spunta utilizando A. tumefaciens con un vector binario que contenía el gen hpt que confiere resistencia al antibiótico higromicina. Se realizaron determinaciones moleculares y fenotípicas para validar tanto la efi-ciencia del protocolo de transformación como la acción del antibiótico higromicina como agente de selección. Se obtuvieron 62 ápices regenerados de los explantes inoculados con A. tumefaciens (62% de regeneración) de las cuales 30 plantas fueron positivas para el gen hpt. Considerando como eficiencia de transformación el número de plantas transgénicas/el número de explantes inoculados, obtuvimos un 30% de eficiencia. El presente trabajo permitió desarrollar un protocolo sencillo y eficiente de transformación genética de papa cv. Spunta mediante A. tumefaciens utilizando higromicina como agente selectivo.
Palabras claves: papa, transformación vegetal, selección, mejoramiento genético.
ABSTRACT
Plant genetic transformation is a biotechnological tool with a high impact on plant breeding since it offers the possibility of introducing new traits in crops to fulfill the demands of production and consumption. The most widely used transformation strategy in potato is based on integration of genes via A. tumefaciens. The objec-tive of the present work was to develop a transformation protocol for Solanum tuberosum cv. Spunta mediated by A. tumefaciens using hygromycin as a selective agent. For this, explants of potato cv. Spunta were trans-formed using A. tumefaciens with a binary vector containing the hygromycin conferring resistance transgene (hpt) under the control of NOS promoter. Molecular and phenotypic determinations were performed to validate both the efficiency of the transformation protocol and the performance of hygromycin as a selection agent. Sixty-two regenerated shoots were obtained from explants inoculated with A. tumefaciens (62% regeneration) of which 30 plants were positive for the hpt gene. Thirty percent transformation efficiency -calculated as the
1Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Balcarce, Laboratorio de Agrobiotecnolo-gía, Balcarce CC 276 (7620), Balcarce, Argentina.2Universidad Nacional de Mar del Plata, Facultad de Ciencias Agrarias, Balcarce, Argentina.3Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina. Correo electrónico: [email protected]
DÉCIMA ONETO, C.1,2 ; CORONEL, J.1; STORANI, L.1,3; GONZÁLEZ, M.N.1,3; FEINGOLD, S.1 ; MASSA, G. A.1,2,3
Recibido 10 de octubre de 2017 // Aceptado 25 de abril de 2020 // Publicado online 23 de septiembre de 2020

DÉCIMA ONETO, C.; CORONEL, J.; STORANI, L.; GONZÁLEZ, M.N.; FEINGOLD, S. ; MASSA, G. A.
Agosto 2020, Argentina 249249
number of transgenic plants / the number of explants inoculated- was obtained using this protocol. The present work developed presents a simple and efficient protocol for genetic transformation of potato cv. Spunta by A. tumefaciens using hygromycin as a selective agent.
Keywords: potato, vegetal transformation, selection, genetic breeding.
INTRODUCCIÓN
El cultivo de papa (Solanum tuberosum L.) es el tercer cul-tivo alimenticio más importante del mundo, con un total de 22 millones de hectáreas y una producción aproximada de 458 millones de toneladas. En Argentina, durante la cam-paña 2015/2016 se reportó una producción de 2.430.000 t (FAOSTAT, 2018). El cultivar Spunta es el más utilizado en Argentina debido a que posee un alto rendimiento, se adap-ta a regiones con doble cosecha y posee muy buena capaci-dad de conservación (Huarte y Capezio, 2013).
Uno de los objetivos de los programas de mejoramien-to es el aumento de la producción con el fin de cumplir con la demanda del sector. La transformación genética de plantas, entendida como la introducción controlada de áci-dos nucleicos en un genoma receptor, es una herramienta biotecnológica de alto impacto que brinda la posibilidad de obtener con mayor rapidez cultivos mejorados a partir de la introducción de una característica deseada, por ejemplo: aumento en la producción, calidad nutricional y tolerancia a estreses tantos bióticos como abióticos (Waterer et al., 2010; Imbo et al., 2016). Los métodos de transformación más utilizados actualmente se basan en la obtención de células transgénicas mediante Agrobacterium tumefaciens o biobalística, y posterior regeneración de plantas comple-tas y fértiles a partir de cultivo y selección in vitro. Durante el proceso de transformación son pocas las células blanco que sobreviven e incorporan establemente el ADN foráneo (Birch, 1997). Es esencial, por lo tanto, detectar las células transformadas dentro del gran número de las no transfor-madas y establecer condiciones adecuadas de cultivos de tejidos, de transferencia de genes y de selección que per-mitan desarrollar plantas enteras a partir de esta estrate-gia. Para ello se utilizan genes “marcadores” que permiten crecer a los explantes transformados en medios selectivos, lo que permite diferenciarlos de los no transformados.
En la actualidad, se encuentran reportados múltiples pro-tocolos de transformación de papa vía A. tumefaciens uti-lizando diversos explantes, en distintos cultivares y con di-ferentes agentes de selección (Millam, 2007; Halterman et al., 2016; Craze et al., 2018; Bruce y Rupp, 2019; Bakhsh et al., 2020). La relevancia de este tema se debe principal-mente a que la papa cultivada, Solanum tuberosum Grupo Tuberosum L. (2n = 4x = 48), es un cultivo tetraploide he-terocigoto que se propaga clonalmente y debido a la fal-
ta de reproducción sexual y su segregación concomitante de alelos, la ingeniería genética es una forma eficiente de introducir rasgos de mejoramiento de cultivos de manera más rápida que el mejoramiento convencional (Nadakuduti et al., 2019).
Cabe destacar que a la fecha no se encuentra publicado un protocolo de transformación de papa para el cultivar Spunta.
A su vez, en diversos trabajos se indica que la transfor-mación genética vegetal mediada por A. tumefaciens es dependiente del genotipo (Heeres et al., 2002; Cingel et al., 2010; Barpete et al., 2015; Chakraborty et al., 2020) por lo que resulta de interés desarrollar un protocolo de trans-formación genética para cultivares comerciales de papa, como Spunta, lo que permitirá en un futuro ser aplicado en planes de mejoramiento.
MATERIALES Y MÉTODOS
1. Material vegetal
Ápices de S. tuberosum cv. Spunta se cultivaron in vitro en medio de cultivo Murashige Skoog (MS; medio basal Murashige Skoog 1X suplementado con vitaminas (Phyto-technology Laboratories, Cat. N.o M519); 20 g.L-1 sacarosa (Biopack, Cat. N.o 9789.08); 8 g.L-1 de agar (Phytotechno-logy Laboratories, Cat. N.o A296) durante 3 a 4 semanas en tubos de ensayos para la determinación de la curva de sensibilidad y en cajas magenta de plástico para los en-sayos de transformación (Phytotechnology Laboratories, Cat. N.o C2100). El medio MS se ajustó a pH 5,8 con 0,1 N KOH o HCl antes de esterilizar en autoclave (121 ºC, 20 min). Para los experimentos de transformación se coloca-ron cinco ápices por recipiente de cultivo, en un gabinete de flujo laminar, y se cultivaron en cuarto de cultivo con un fotoperiodo de 16/8 h (luz/oscuridad) (80/90 µE/m2/s luz blanca fría) a 21 ºC, hasta el momento de realizar la trans-formación vegetal.
2. Protocolo de selección: curva de sensibilidad al agente selectivo
Se evaluaron los efectos de distintas concentraciones del agente selectivo higromicina en plántulas de papa cv. Spunta sin transformar (WT) durante 30 días de cultivo in

Transformación eficiente de Solanum tuberosum cv. Spunta mediada por Agrobacterium tumefaciens utilizandohigromicina como agente selectivo
ARTÍCULOS RIA / Vol. 46 / N.º 2250250
logy Laboratories, Cat. N.o Z899) y 8 g.L-1 de agar (Phyto-technology Laboratories, Cat. N.o A296).
Luego del periodo de cocultivo, se subcultivaron un total de 10 explantes por placa en medio inductor de callo (CIM) durante 12 días a 21 °C con fotoperiodo 16/8 (luz/oscuridad; 80-110 µE/m2/s). El medio CIM está compuesto por medio MS suplementado con 0,2 mg.L-1 de ANA, 0,02 mg.L-1 de GA3, 2,5 mg.L-1 de ZR, 2,5 mg.L-1 de cefotaxime (Phytote-chnology Laboratories, Cat. N.o A296), 100 mg.L-1 de ampi-cilina (Sigma, Cat. N.o 7177-48-2) y 8 g.L-1 de agar.
Posteriormente, los explantes se subcultivaron cada 15 días en medio inductor de ápices (SIM) a 21 °C con foto-periodo 16/8 (luz/oscuridad; 80-110 µE/m2/s). El medio SIM está compuesto por medio MS suplementado con 0,2 mg.L-1
de ANA, 0,2 mg.L-1 de GA3, 2,5 mg.L-1 de ZR, 400 mg.L-1 de cefotaxima, 100 mg.L 1 de ampicilina, 8 mg.L-1 de higromi-cina y 8 g.L -1 de agar.
A medida que se formaron los ápices (5-10 mm de tama-ño), se cortaron cuidadosamente de los callos y se pasaron a tubos que contenían medio MS y se mantuvieron a 21 °C bajo fotoperiodo 16/8 (luz/oscuridad; 80-110 µE/m2/s).
Se continuó transfiriendo los explantes a intervalos de 15 días a medio SIM. La disección de los nuevos ápices se realizó de manera de asegurar que los nuevos brotes provinieran de callos independientes.
5. Detección del gen hpt mediante la reacción en cadena de la polimerasa (PCR)
La extracción de ADN de las plántulas provenientes del ensayo de transformación se realizó según lo descripto por Haymes y colaboradores (1996). Se determinó la concen-tración de ADN por absorbancia a 260 nm con un espectro-fotómetro digital (SmartSpectTM3000 BIORAD, EE. UU.). Se determinó la calidad del ADN mediante la relación A260/A280 y por electroforesis en geles de agarosa al 1% teñi-dos con GelRed (Genbiotech, Argentina).
El ADN extraído de las muestras se utilizó como molde para amplificar el gen hpt mediante PCR. Para tal fin se utilizaron los iniciadores Hig Fw e Hig Rv (5´gaagatgttggc-gacctcgta 3´ y 5´aagttcgacagcgtctccga 3´, respectivamen-te). Como control negativo se utilizó ADN de plantas de papa cv. Spunta WT y como control positivo se utilizó ADN plasmídico del vector pHPT. Las condiciones de amplifica-ción fueron: 94 °C durante 3 min, 35 ciclos: 94 °C por 30 seg, 52 °C por 15 seg, 72 °C por 1 min y por último, 1 ciclo a 72 °C durante 5 min. Los productos obtenidos fueron con-firmados por electroforesis en gel de agarosa al 1% teñidos con GelRed (Genbiotech, Argentina) en presencia de mar-cadores de peso molecular (Axygen, EE. UU.) y revelados con luz ultravioleta.
6. Análisis estadístico
Para el análisis estadístico de los datos fenotípicos de la curva de sensibilidad se realizó un ANOVA de medidas
vitro en medio BM. Para realizar la curva de sensibilidad se utilizaron 5 concentraciones diferentes de higromicina (2 mg.L-1, 4 mg.L-1, 6 mg.L-1, 8 mg.L-1 o 10 mg.L-1). Como control se utilizó medio BM sin higromicina. Para cada cur-va se analizaron 10 plántulas durante cuatro semanas. En este periodo se realizaron registros tres veces por sema-na de los parámetros: altura de las plántulas, presencia de raíz, número y coloración de hojas.
3. Vector de expresión vegetal
Para la transformación genética se usó el vector bina-rio pHPT, (13277 pb). Este plásmido contiene el gen hpt que codifica para la enzima higromicin-fosfotransferasa, la cual otorga resistencia al antibiótico higromicina bajo con-trol transcripcional del promotor NOS (del inglés: nopaline synthase; An et al., 1990) y el terminador NOS entre los bordes del ADN-T. El vector pHPT se introdujo en la cepa EHA105 de A. tumefaciens por electroporación. Las cé-lulas transformadas se seleccionaron para resistencia en medio Luria Bertani (LB) suplementado con 25 mg.L-1 de rifampicina y 50 mg.L-1 de kanamicina.
3.1 Preparación del inóculo de Agrobacterium tumefaciens para transformación
Se realizó un cultivo inicial de 5 ml de medio LB, suple-mentado con 25 mg.L-1 de rifampicina y 50 mg.L-1 de kana-micina, partiendo de un cultivo en placa fresca. El cultivo se incubó durante 16 h a 28 °C en un agitador a 200 rpm. Luego de la incubación de 16 h se transfirieron los 5 ml de cultivo a un erlenmeyer de 250 ml que contenía 50 ml de medio LB suplementado con 25 mg.L-1 de rifampicina y 50 mg.L-1 de kanamicina. El cultivo se incubó durante 16 h a 28 °C en un agitador a 200 rpm. En el momento de la transformación se midió la densidad óptica (DO) a 600 nm.
4. Protocolo de transformación
Se utilizaron como explantes hojas de plántulas de 3 o 4 semanas cultivadas in vitro. Las hojas fueron cortadas eliminando el pecíolo. Se colocaron 20 explantes por caja de Petri (OneLab Ø90mm) y 20 ml de medio de infección líquido compuesto por sacarosa 20 g.L-1 (Biopack, Cat. N.o 9789.08) y 4,3 g.L-1 sales con vitaminas Murashige y Skoog (PhytoTechnology Laboratories, Cat. N.o M519). A cada pla-ca se le adicionó la cantidad necesaria de inóculo de A. tumefaciens para lograr una DO final de 0,2/0,3.
Inmediatamente, las placas se incubaron a 21 °C, en os-curidad con una agitación a 50 rpm durante 45 minutos. Luego, los explantes se secaron en papel estéril y se sub-cultivaron en medio de cocultivo durante 48 h a 21 °C con un fotoperiodo 16/8 (luz/oscuridad; 20 µE/m2/s). El medio de cocultivo está compuesto por medio MS suplementado con 0,2 mg.L-1 de ácido naftalén acético (ANA; Sigma, Cat. N.o NN0640), 0,02 mg.L-1 de ácido giberélico (GA3; Sigma, Cat. N.o x), 2,5 mg.L-1 de zeatin ribósido (ZR; PhytoTechno-
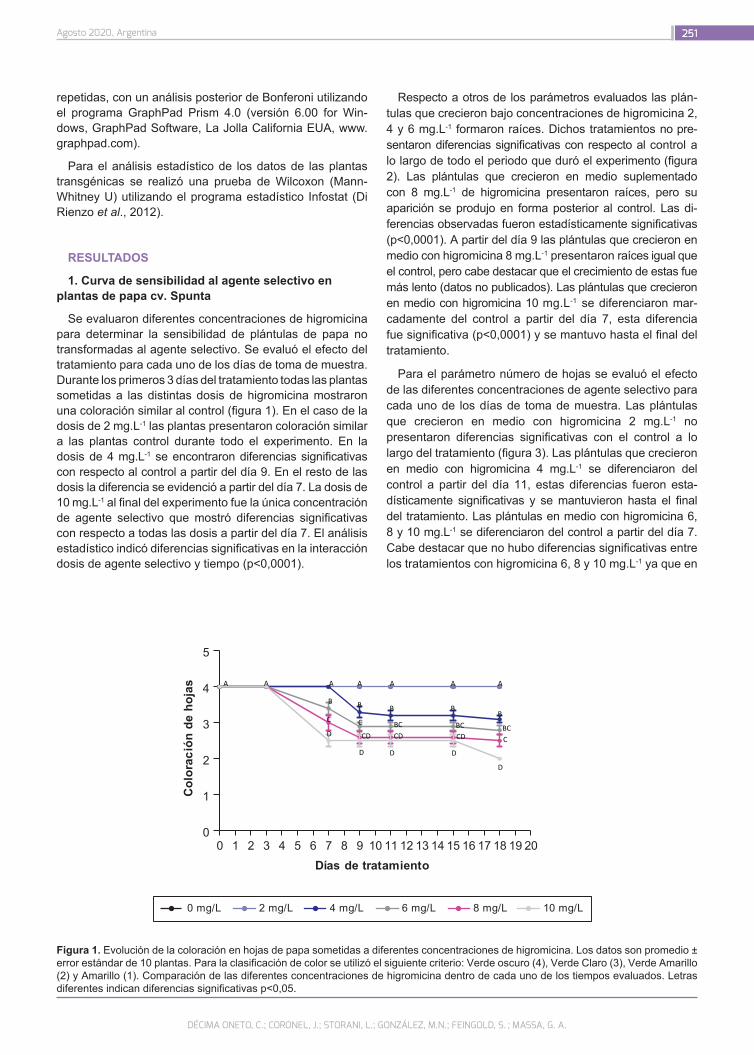
DÉCIMA ONETO, C.; CORONEL, J.; STORANI, L.; GONZÁLEZ, M.N.; FEINGOLD, S. ; MASSA, G. A.
Agosto 2020, Argentina 251251
Figura 1. Evolución de la coloración en hojas de papa sometidas a diferentes concentraciones de higromicina. Los datos son promedio ± error estándar de 10 plantas. Para la clasificación de color se utilizó el siguiente criterio: Verde oscuro (4), Verde Claro (3), Verde Amarillo (2) y Amarillo (1). Comparación de las diferentes concentraciones de higromicina dentro de cada uno de los tiempos evaluados. Letras diferentes indican diferencias significativas p<0,05.
repetidas, con un análisis posterior de Bonferoni utilizando el programa GraphPad Prism 4.0 (versión 6.00 for Win-dows, GraphPad Software, La Jolla California EUA, www.graphpad.com).
Para el análisis estadístico de los datos de las plantas transgénicas se realizó una prueba de Wilcoxon (Mann-Whitney U) utilizando el programa estadístico Infostat (Di Rienzo et al., 2012).
RESULTADOS
1. Curva de sensibilidad al agente selectivo en plantas de papa cv. Spunta
Se evaluaron diferentes concentraciones de higromicina para determinar la sensibilidad de plántulas de papa no transformadas al agente selectivo. Se evaluó el efecto del tratamiento para cada uno de los días de toma de muestra. Durante los primeros 3 días del tratamiento todas las plantas sometidas a las distintas dosis de higromicina mostraron una coloración similar al control (figura 1). En el caso de la dosis de 2 mg.L-1 las plantas presentaron coloración similar a las plantas control durante todo el experimento. En la dosis de 4 mg.L-1 se encontraron diferencias significativas con respecto al control a partir del día 9. En el resto de las dosis la diferencia se evidenció a partir del día 7. La dosis de 10 mg.L-1 al final del experimento fue la única concentración de agente selectivo que mostró diferencias significativas con respecto a todas las dosis a partir del día 7. El análisis estadístico indicó diferencias significativas en la interacción dosis de agente selectivo y tiempo (p<0,0001).
Respecto a otros de los parámetros evaluados las plán-tulas que crecieron bajo concentraciones de higromi cina 2, 4 y 6 mg.L-1 formaron raíces. Dichos tratamientos no pre-sentaron diferencias significativas con respecto al control a lo largo de todo el periodo que duró el experimento (figura 2). Las plántulas que crecieron en medio suplementado con 8 mg.L-1 de higromicina presentaron raíces, pero su a parición se produjo en forma posterior al control. Las di-ferencias observadas fueron estadísticamente signifi ca tivas (p<0,0001). A partir del día 9 las plántulas que cre cieron en medio con higromicina 8 mg.L-1 presentaron raí ces igual que el control, pero cabe destacar que el creci miento de estas fue más lento (datos no publicados). Las plántulas que crecieron en medio con higromicina 10 mg.L-1 se diferenciaron mar-cadamente del control a partir del día 7, esta diferencia fue significativa (p<0,0001) y se mantuvo hasta el final del tratamiento.
Para el parámetro número de hojas se evaluó el efecto de las diferentes concentraciones de agente selectivo para cada uno de los días de toma de muestra. Las plántulas que crecieron en medio con higromicina 2 mg.L-1 no presentaron diferencias significativas con el control a lo largo del tratamiento (figura 3). Las plántulas que crecieron en medio con higromicina 4 mg.L-1 se diferenciaron del control a partir del día 11, estas diferencias fueron esta-dísticamente significativas y se mantuvieron hasta el final del tratamiento. Las plántulas en medio con higromicina 6, 8 y 10 mg.L-1 se diferenciaron del control a partir del día 7. Cabe destacar que no hubo diferencias significativas entre los tratamientos con higromicina 6, 8 y 10 mg.L-1 ya que en
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 200
1
2
3
4
5
0 mg/L 2 mg/L 4 mg/L 6 mg/L 8 mg/L 10 mg/L
Días de tratamiento
Col
orac
ión
de h
ojas A A A A AA A
B B BB
B
C BC BC BCC
CCDCDCDD
DD
D
D
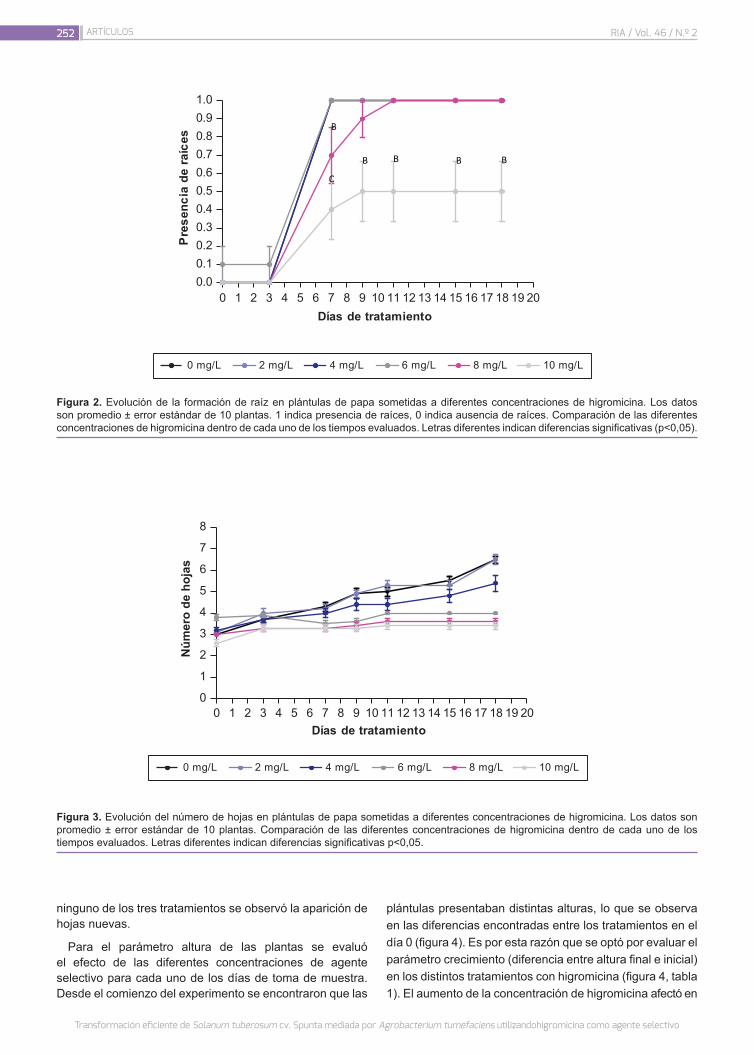
Transformación eficiente de Solanum tuberosum cv. Spunta mediada por Agrobacterium tumefaciens utilizandohigromicina como agente selectivo
ARTÍCULOS RIA / Vol. 46 / N.º 2252252
ninguno de los tres tratamientos se observó la aparición de hojas nuevas.
Para el parámetro altura de las plantas se evaluó el efecto de las diferentes concentraciones de agente selectivo para cada uno de los días de toma de muestra. Desde el comienzo del experimento se encontraron que las
plántulas presentaban distintas alturas, lo que se observa en las diferencias encontradas entre los tratamientos en el día 0 (figura 4). Es por esta razón que se optó por evaluar el parámetro crecimiento (diferencia entre altura final e inicial) en los distintos tratamientos con higromicina (figura 4, tabla 1). El aumento de la concentración de higromicina afectó en
Figura 2. Evolución de la formación de raíz en plántulas de papa sometidas a diferentes concentraciones de higromicina. Los datos son promedio ± error estándar de 10 plantas. 1 indica presencia de raíces, 0 indica ausencia de raíces. Comparación de las diferentes concentraciones de higromicina dentro de cada uno de los tiempos evaluados. Letras diferentes indican diferencias significativas (p<0,05).
Figura 3. Evolución del número de hojas en plántulas de papa sometidas a diferentes concentraciones de higromicina. Los datos son promedio ± error estándar de 10 plantas. Comparación de las diferentes concentraciones de higromicina dentro de cada uno de los tiempos evaluados. Letras diferentes indican diferencias significativas p<0,05.
0 mg/L 2 mg/L 4 mg/L 6 mg/L 8 mg/L 10 mg/L
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 200.00.10.20.30.40.50.60.70.80.91.0
Días de tratamiento
Pre
senc
ia d
e ra
íces
B
C
B B B B
0 mg/L 2 mg/L 4 mg/L 6 mg/L 8 mg/L 10 mg/L
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 200
1
2
3
4
5
6
7
8
Días de tratamiento
Núm
ero
de h
ojas
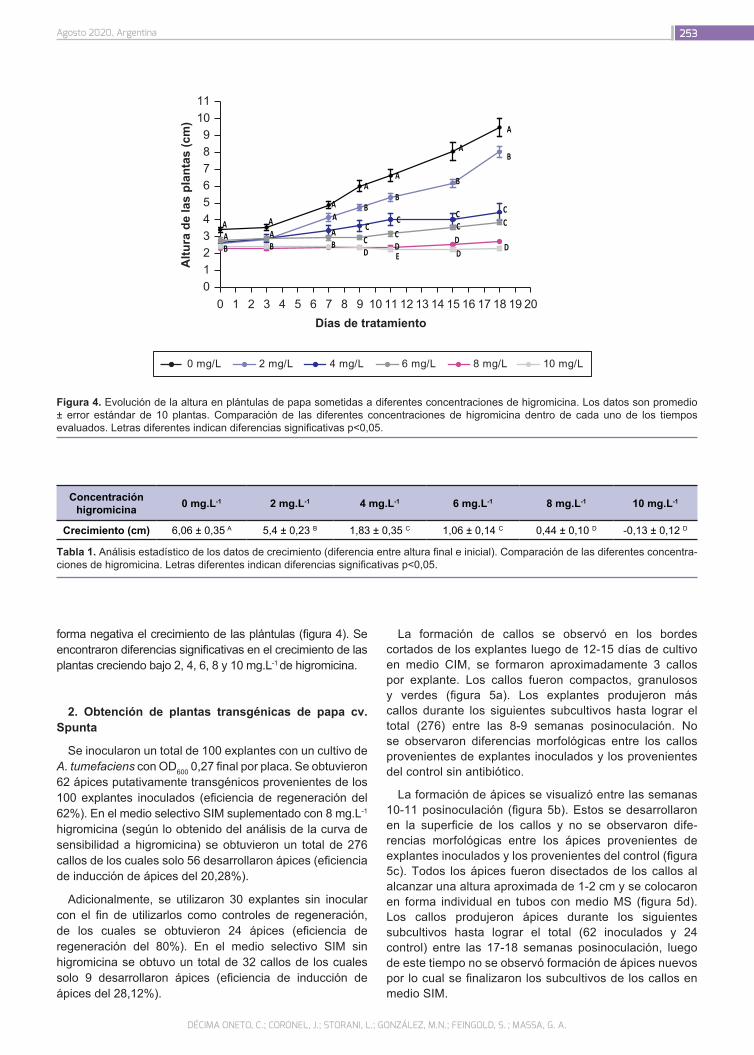
DÉCIMA ONETO, C.; CORONEL, J.; STORANI, L.; GONZÁLEZ, M.N.; FEINGOLD, S. ; MASSA, G. A.
Agosto 2020, Argentina 253253
forma negativa el crecimiento de las plántulas (figura 4). Se encontraron diferencias significativas en el crecimiento de las plantas creciendo bajo 2, 4, 6, 8 y 10 mg.L-1 de higromicina.
2. Obtención de plantas transgénicas de papa cv. Spunta
Se inocularon un total de 100 explantes con un cultivo de A. tumefaciens con OD600 0,27 final por placa. Se obtuvieron 62 ápices putativamente transgénicos provenientes de los 100 explantes inoculados (eficiencia de regeneración del 62%). En el medio selectivo SIM suplementado con 8 mg.L-1 higromicina (según lo obtenido del análisis de la curva de sensibilidad a higromicina) se obtuvieron un total de 276 callos de los cuales solo 56 desarrollaron ápices (eficiencia de inducción de ápices del 20,28%).
Adicionalmente, se utilizaron 30 explantes sin inocular con el fin de utilizarlos como controles de regeneración, de los cuales se obtuvieron 24 ápices (eficiencia de regeneración del 80%). En el medio selectivo SIM sin higromicina se obtuvo un total de 32 callos de los cuales solo 9 desarrollaron ápices (eficiencia de inducción de ápices del 28,12%).
La formación de callos se observó en los bordes cortados de los explantes luego de 12-15 días de cultivo en medio CIM, se formaron aproximadamente 3 callos por explante. Los callos fueron compactos, granulosos y verdes (figura 5a). Los explantes produjeron más callos durante los siguientes subcultivos hasta lograr el total (276) entre las 8-9 semanas posinoculación. No se observaron diferencias morfológicas entre los callos provenientes de explantes inoculados y los provenientes del control sin antibiótico.
La formación de ápices se visualizó entre las semanas 10-11 posinoculación (figura 5b). Estos se desarrollaron en la superficie de los callos y no se observaron dife-ren cias morfológicas entre los ápices provenientes de explantes inoculados y los provenientes del control (figura 5c). Todos los ápices fueron disectados de los callos al alcanzar una altura aproximada de 1-2 cm y se colocaron en forma individual en tubos con medio MS (figura 5d). Los callos produjeron ápices durante los siguientes subcultivos hasta lograr el total (62 inoculados y 24 control) entre las 17-18 semanas posinoculación, luego de este tiempo no se observó formación de ápices nuevos por lo cual se finalizaron los subcultivos de los callos en medio SIM.
Figura 4. Evolución de la altura en plántulas de papa sometidas a diferentes concentraciones de higromicina. Los datos son promedio ± error estándar de 10 plantas. Comparación de las diferentes concentraciones de higromicina dentro de cada uno de los tiempos evaluados. Letras diferentes indican diferencias significativas p<0,05.
Tabla 1. Análisis estadístico de los datos de crecimiento (di fe rencia entre altura final e inicial). Comparación de las diferentes co ncen tra-ciones de higromicina. Letras diferentes indican diferencias significativas p<0,05.
0 mg/L 2 mg/L 4 mg/L 6 mg/L 8 mg/L 10 mg/L
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 200123456789
1011
Días de tratamiento
Altu
ra d
e la
s pl
anta
s (c
m)
AAB
AAB
AAAB
A
B
CCD
A
B
CCDE
A
B
CC
DD
A
B
CC
D
Concentración higromicina 0 mg.L-1 2 mg.L-1 4 mg.L-1 6 mg.L-1 8 mg.L-1 10 mg.L-1
Crecimiento (cm) 6,06 ± 0,35 A 5,4 ± 0,23 B 1,83 ± 0,35 C 1,06 ± 0,14 C 0,44 ± 0,10 D -0,13 ± 0,12 D

Transformación eficiente de Solanum tuberosum cv. Spunta mediada por Agrobacterium tumefaciens utilizandohigromicina como agente selectivo
ARTÍCULOS RIA / Vol. 46 / N.º 2254254
2.1 Evaluación de plantas transgénicas cultivadas en medio selectivo
A fin de corroborar los resultados de la curva de sensibi-lidad, se seleccionó la concentración de 8 mg.L-1 de higro-micina para evaluar su eficacia como agente de selección de las plantas transgénicas del cv. Spunta. Se seleccionó la concentración de 8 mg.l-1 de higromicina debido a que fue la menor concentración en la que se observaron dife-rencias significativas en todos los parámetros estudiados comparando con los controles.
Se tomaron los 62 ápices de papa cv. Spunta provenien-tes de los ensayos de transformación genética mediada por Agrobacterium tumefaciens, se subcultivaron en tubos que contenían medio MS suplementado con 8 mg.L-1 de hi-gromicina y fueron evaluados fenotípica y molecularmente a fin de confirmar la presencia del transgen hpt.
2.2 Evaluación molecular
Luego de tres semanas se procedió a la extracción de ADN genómico de las 62 plántulas subcultivadas en me-dio MS. El ADN extraído fue de buena calidad ya que se observó una banda íntegra de alto peso molecular y la re-lación de A260/A280 se encontró en el rango de 1,6 a 2,3. Se
procedió a realizar una PCR a fin de poder determinar la presencia del gen hpt. De 62 plantas analizadas, 30 resul-taron positivas para la presencia del gen hpt (hpt+, 648 pb) y se clasificaron como plantas transgénicas. Las 32 plantas restantes no presentaron el gen hpt por lo que clasificaron como plantas no transformadas y fueron consideradas es-capes. Estos resultados indican que la eficiencia de trans-formación obtenida fue del 30%, calculada como número de plantas transgénicas obtenidas/número de explantes inoculados.
2.3 Evaluación fenotípica de las plantas hpt+ y plantas no transformadas
Se separaron las plantas en dos grupos, aquellas que fueron positivas para hpt (hpt+) y las negativas para hpt (hpt-) (escapes), se colocaron en tubos que contenían BM suplementado con 8 mg.L-1 de higromicina y se evaluaron los parámetros medidos anteriormente durante la curva de sensibilidad. En plantas hpt+ se observó aparición de hojas nuevas, efecto no observado en plantas hpt- (p=0,0001; 1,25±0,14 y 0±0 ca). La coloración de todas las plantas hpt+ fue de verde intenso, mientras que en plantas hpt- varió desde amarillo a verde amarillento (p<0,0001; 4±0
Figura 5. Transformación genética de papa cv. Spunta mediada por Agrobacterium tumefaciens. A, B y C: callos obtenidos en medio de cultivo CIM con selección; D: ápices provenientes de explantes inoculados en medio SIM; E y F: ápices provenientes de explantes inoculados en medio BM (en tubo de ensayo), G: plántulas provenientes de explantes inoculados, cortados del callo en medio BM.
A
D
B
E
C
F

DÉCIMA ONETO, C.; CORONEL, J.; STORANI, L.; GONZÁLEZ, M.N.; FEINGOLD, S. ; MASSA, G. A.
Agosto 2020, Argentina 255255
y 1,38±0,16 ca). En el caso del crecimiento se observó que fue mayor en plantas hpt+, mientas que en plantas hpt- el crecimiento fue casi nulo (p=0,0002; 3,34±0,28 cm; 0,09±0,05 cm ca). Por último se evaluó la presencia de raí-ces. Si bien no todas las plantas desarrollaron raíces, la aparición de estas fue mayor en plantas hpt+ que en plan-tas hpt- (p=0,0201; 0,67±0,15 y 0,13±0,11 ca).
DISCUSIÓN
El desarrollo de un protocolo de regeneración confiable es un paso crucial para la transformación genética en papa (Ahmed et al., 2018) debido a que el genotipo es un factor importante que afecta tanto la eficiencia de regeneración como la eficiencia de transformación genética (Borna et al., 2010; Ahloowalia et al., 1982; Romano et al., 2003). La se-lección de explantes sanos de la planta wild type de papa es crítica para generar una alta frecuencia de callos, brotes y regeneración de raíces (Khatun et al., 2012).
En el presente trabajo se ajustó por primera vez un pro-tocolo reproducible de transformación genética de papa cv. Spunta mediante la utilización de Agrobacterium tumefa-ciens utilizando como agente selectivo el antibiótico higro-micina. Mediante la utilización de este protocolo se logró una eficiencia de transformación del 30%. En diversos tra-bajos se reportan eficiencias de transformación superiores, en un rango entre el 60% y el 90% (Beaujean et al., 1998; Douches et al., 1998; Khatun et al., 2012; Borna et al., 2010; Ahmed et al., 2018; Bakhsh et al., 2020) para cultiva-res como Désirée, Lemhi Russet, Kaptah Vandel, Cardinal, Heera, Diamant, Granola, Lady Olympia, Agria e Innovator. En ninguno de estos trabajos está incluido el cv. Spunta, por lo que no es posible lograr una adecuada comparación entre las eficiencias de transformación reportadas en los trabajos mencionados y el nuestro.
La menor eficiencia de transformación obtenida en nues-tro ensayo puede ser explicada por factores como el geno-tipo, el agente de selección, la construcción utilizada para la transformación o la cepa de Agrobacterium utilizada (Roman et al., 2015). Por un lado, se obtuvo un 50% de ápices re-generantes en medio SIM que resultaron escapes (hpt-). Este hecho también fue reportado en la transformación del cv. Desiree (Roman et al., 2015). El alto número de escapes se puede deber a la gran capacidad de regeneración in vitro que caracteriza a ciertos cultivares de papa (Cañedo y Cisne-ros, 2004), permitiendo la proliferación y desarrollo de ciertos regenerantes no transformados en presencia del agente se-lector. Como resultado se obtiene una planta con resistencia o tolerancia natural (López y Chaparro, 2007). También es posible que varios de estos regenerantes no estuvieran en contacto permanente en el medio que contiene el agente se-lector permitiendo su desarrollo (Monserrat et al., 2001). Esto puede deberse a la forma globular que presentan los callos donde no todo el callo se encuentra en contacto con el medio que contiene el agente selectivo. Es por esta razón que los ápices obtenidos en medio SIM se deben cultivar en forma individual en tubos que contengan medio BM suplementado con 8 mg.L-1 de higromicina, para asegurar el contacto del
ápice con el agente de selección y de esta forma reducir el número de plantas en los análisis moleculares.
Por otro lado, el antibiótico higromicina se utilizó por pri-mera vez como agente selectivo en la transformación ge-nética de Nicotiana tabacum por el grupo de Van den El-zen (1985) y en 1989 fue utilizado por primera vez en papa (Solanum tuberosum x S. phureja y Solanum tuberosum x S. chacoense) por Masson (1989). Posteriormente, fue uti-lizado en numerosos trabajos como agente selectivo en la transformación de diversos cultivares de papa y batata (Su-rov et al., 1998; Okada et al., 2001; Cavatorta et al., 2011; Orbegozo et al., 2016), no describiéndose su acción en el cv. Spunta. A través de los ensayos de sensibilidad al antibióti-co higromicina se determinó que la dosis letal para los ex-plantes no transformados fue de 8 mg L-1 de higromicina. De acuerdo con los resultados obtenidos y luego de comparar las diferentes curvas de sensibilidad al antibiótico, se pudo concluir que el aumento de la concentración del agente se-lectivo en el medio de cultivo provocó una disminución en el crecimiento, decoloración del tejido, disminución del número de hojas y perjudicó la formación de raíces. Las plantas que presentaron mejores parámetros morfofisiológicos en medio selectivo portaban el gen hpt. El método de selección pro-puesto para el cv. Spunta (8 mg L-1 de higromicina) presentó un porcentaje de identificación del 48,38% entre las plantas transgénicas de las plántulas no transformadas.
En relación con el uso de higromicina durante el proceso de obtención de transformantes se concluye que la eficien-cia fue baja (48,38%) en el presente trabajo; esto se de-terminó al comparar los resultados obtenidos con ensayos reportados previamente donde se describe el uso del mis-mo antibiótico, pero en diferentes variedades (M’Hamdi et al., 2003; Kashani et al., 2012). Estas diferencias pueden deberse a múltiples factores asociados a las condiciones de transformación o al tipo de explante usado (Lagunes, 2009). No obstante la principal causa puede deberse a que los ensayos de regeneración y transformación son genoti-po dependiente del tipo de cultivo que se use en los expe-rimentos para realizar (Orbegozo et al., 2013). En futuros trabajos de transformación del cv. Spunta utilizando como agente de selección higromicina, se deberá ajustar este parámetro y utilizar concentraciones más altas.
Este protocolo es una primera aproximación a la transfor-mación genética (de un cultivar de papa de amplio interés económico y agronómico en Argentina como lo es el culti-var Spunta) mediada por Agrobacterium tumefaciens utili-zando como agente de selección el antibiótico higromicina.
CONCLUSIONES
Se logró la obtención de plantas transgénicas de papa cv. Spunta resistentes al antibiótico higromicina. Se logró es-tablecer un protocolo de transformación reproducible que puede ser aplicado, como primer acercamiento, en otros cultivares de interés económico y agronómico en Argentina, sobre los cuales no existen reportes de protocolos de trans-formación. Para cada caso se deberá estudiar la respuesta de cada cultivar al método desarrollado para Spunta.

Transformación eficiente de Solanum tuberosum cv. Spunta mediada por Agrobacterium tumefaciens utilizandohigromicina como agente selectivo
ARTÍCULOS RIA / Vol. 46 / N.º 2256256
Finalmente, luego de realizar la curva de dosis respues-ta con el agente de selección higromicina en plántulas de papa cv. Spunta se concluyó que las variables analizadas fueron adecuadas para determinar la sensibilidad al anti-biótico y que la concentración de 8 mg L-1 de higromici-na, si bien permitió diferenciar plantas de papa cv. Spunta transgénicas de las plantas WT, no fue del todo suficiente y es necesario aumentar la dosis de este en los medios de cultivo. Las variables evaluadas “aparición de hojas nue-vas”, “emergencia de raíces” y “crecimiento absoluto” son las más adecuadas para tener en cuenta al momento de seleccionar plantas transgénicas que utilicen el antibiótico higromicina como agente de selección.
AGRADECIMIENTOS
Los autores agradecen a Silvina Divito y a Cristina Cinalli por su asistencia técnica. Este trabajo fue financiado por el proyecto INTA PNBIO1131024 “Desarrollo de sistemas alternativos de generación y utilización de variabilidad ge-nética y su aplicación al mejoramiento de los cultivos” y por el proyecto joven PICT 2011-0411 “Mutagénesis dirigida en papa mediante la tecnología de nucleasas zinc-finger”. MNG posee una beca doctoral de CONICET y LS una beca posdoctoral de CONICET.
BIBLIOGRAFÍA
AHLOOWALIA, B.S. 1982. Plant regeneration from callus cul-ture in potato. Euphytica. 31, 755-759.
AHMED, H.A.; BARPETE, S.; AKDOGAN, G.; AYDIN, G.; SANKAT, C.; OZCAN, S. 2018. Effient regeneration and mediated genetic transformation of potato (L.). Feb-fresenius environmental bulletin, 3020.
AN, G.; COSTA, M.; HA, S. 1990. Nopaline Synthase Promoter 1s Wound lnducible and Auxin lnducible. American Society of Plant Physiologists. The plant cell vol. 2, 225-233.
BAKHSH, A. 2020. Development of efficient, reproducible and stable Agrobacterium-mediated genetic transformation of five po-tato cultivars. Food Technology and Biotechnology, 58(1), 57-63.
BARPETE, S.; OGUZ, M.C.; ÖZCAN, S.F.; ANAYOL, E.; AHMED, H.A. 2015. Effect of temperature on germination, seed vigor index and seedling growth of five Turkish cotton (Gossypium hirsutum L.) cultivars. Fresen. Environ. Bull. 24, 2561-2566.
BETTANY, A.; DALTON, S.; TIMMS, E.; DHANOA, M.; MORRIS, P. 2002. Effect of selectable gene to reporter gene ratio on the frequency of co-transformation and co-expression of uidA and hpt transgenes in protoplast-derived plants of tall fescue. Plant cell, tissue and organ culture, 68(2), 177-186.
BEAUJEAN, A.; SANGWAN, R.S.; LECARDONNEL, A.; SANG-WAN-NORREEL, B.S. 1998. Agrobacterium-mediated transforma-tion of three economically important potato cultivars using sliced internodal explants: an efficient protocol of transformation. Journal of experimental Botany, 49(326), 1589-1595.
BIRCH, R. 1997. PLANT TRANSFORMATION: Problems and Strategies for Practical. Application. Annu. Rev. Plant Physiol. Plant Mol. Biol. 48:297-326.
BIRCHLER, J. 2011. Plant Chromosome Engineering: Meth-ods and Protocols, Methods in Molecular Biology, vol. 701, DOI 10.1007/978-1-61737-957-4_1.
BORNA, R.S.; HOQUE, M.I.; SARKER, R.H. 2010. Agrobacte-rium mediated genetic transformation for local cultivars of potato (Solanum tuberosum L.) using marker genes. Plant Tiss Cult. Bio-techno. 20, 145-155.
BRUCE, M.A.; SHOUP RUPP, J.L. 2019. Agrobacterium-Me-diated Transformation of Solanum tuberosum L., Potato. En: KUMAR, S.; BARONE, P.; SMITH, M. (eds.). Transgenic Plants. Methods in Molecular Biology, vol 1864. Humana Press, New York, Nueva York.
CAÑEDO, V.; CISNEROS, F. 2004. Clones de papa transforma-das con la toxina de Bacillus thurigensis (Berliner) contra la polilla de la papa, Phthorimea operculella (Zeller). i. Transformación de clones de papa y verificación de la presencia del gen cry1A (b). Revista Peruana de Entomología 44:89-93.
CAVATORTA, J.; PEREZ, K.; GRAY, S.; VAN ECK, J., YEAM, I.; JAHN, M. 2011. Engineering virus resistance using a modified potato gene. Plant biotechnology journal, 9(9), 1014-1021.
CHAKRABORTY, N.; CHAKRABORTY, P.; Sen, M.; BANDOPAD-HYAY, R. 2020. Choice of explant for plant genetic transformation. Biolistic DNA Delivery in Plants. Humana, Nueva York. 107-123 pp.
CINGEL, A.; VINTERHALTER, B.; VINTERHALTER, D.; DRA-GOSAVAC, D.A.; SMIGOCKI, A.; NINKOVI, S. 2010. Agrobac-terium-mediated transformation of two Serbian potato cultivars (Solanum tuberosum L. cv. Draga_evka and cv. Jelica). Afr. J. Bio-techno. 9, 4644-4650.
CRAZE, M.; BATES, R.; BOWDEN, S.; WALLINGTON, E.J. 2018. Highly Efficient Agrobacterium Mediated Transformation of Potato (Solanum tuberosum) and Production of Transgenic Micro-tubers. Current protocols in plant biology, 3(1), 33-41.
DI RIENZO, J.; CASANOVES, F.; BALZARINI, M.; GONZÁLEZ, L.; TABLADA, M.; ROBLEDO, C. 2012. Programa Estadístico In-foStat. version 2012, para Windows, Grupo InfoStat. Universidad Nacional de Córdoba, Argentina.
DOUCHES, D.S.; WESTEDT, A.L.; ZARKA, K.; SCHROETER, B.; GRAFIUS, E.J. 1998. Breeding, Cultivars, Rootstocks, & Germ-plasm Resources-Potato Transformation to Combine Natural and Engineered Resistance for Controlling Tuber Moth. Hortscience, 33(6), 1053-1056.
FAO. Estadísticas. (Disponible: http://www.fao.org/statistics/es/ verificado: 05 de septiembre de 2017).
HALTERMAN, D.; GUENTHNER, J.; COLLINGE, S.; BUTLER, N.; DOUCHES, D. 2016. Biotech potatoes in the 21st century: 20 years since the first biotech potato. American journal of potato re-search, 93(1), 1-20.
HAYMES, K. 1996. Mini-prep method suitable for plant breeding programs. Plant Mol. Biol. Rep. 14: 280-284.
HEERES, P.; SCHIPPERS, R.M.; JACOBSEN, E.; VISSER, R.G.F. 2002. Transformation of a large number of potato varieties: genotypedependent variation in efficiency and somaclonal variabil-ity. Euphytica. 124, 13-22.
HUARTE, M.; CAPEZIO, S. 2013. Cultivo de papa. (Disponi-ble: http://inta.gob.ar/sites/default/files/script-tmp-inta-_huarte_ca-pezio_papa2013.pdf verificado: agosto de 2017).
IMBO, M.C.; BUDAMBULA, N.; MWEU, C.M.; MULI, J.K.; ANA-MI, S.E. 2016. Genetic transformation of sweet potato for improved tolerance to stress: a review. Advances in Life Science and Tech-nology Vol. 49
KASHANI, K.; JAVARAN, M.J.; MOHEBODINI, M.; MOIENI, A.; SHEIKI, M.; ABAD, D. 2012. Regeneration and Agrobacterium-me-diated transformation of three potato cultivars (Solanum tubero-sum cv. Désirée Agria and Marfona) by human proinsulin gene. AJCS 6(7):1212-1220.

DÉCIMA ONETO, C.; CORONEL, J.; STORANI, L.; GONZÁLEZ, M.N.; FEINGOLD, S. ; MASSA, G. A.
Agosto 2020, Argentina 257257
KHATUN, A.; HASAN, M.M.; BACHCHU, M.A.A.; MONIRUZ-ZAMAN, M.; NASIRUDDIN, K.M. 2012. Agrobacterium-mediated genetic transformation of potato (Solanum tuberosum L.) var Car-dinal and Heera. The Agricult. Scientific J. Krishi Found. 10, 81-86.
LAGUNES, F.E. 2009. Transformación genética de ajo (Allium sativum L.) mediante Agrobacterium tumefaciens. Transformación genética de ajo (Allium sativum L.) mediante Agrobacterium tume-faciens. Tesis, Magister en Recursos Genéticos y Productividad, mención en Ciencias. Colegio de Postgraduados Campus Monte-cillo, Texcoco, Edo. de México. 95 p.
LÓPEZ, A.; CHAPARRO, A. 2007. A system for transformation potato plants (Solanum tuberosum sp. andigena var. Pastusa su-prema) mediated through Agrobacterium tumefaciens. Agronomía Colombiana 25(1):16-25.
MASSON, J.; LANCELIN, D.; BELLINI, C.; LECERF, M.; GUER-CHE, P.; PELLETIER, G. 1989. Selection of somatic hybrids between diploid clones of potato (Solanum tuberosum L.) trans-formed by direct gene transfer. Theoretical and applied genetics, 78(2), 153-159.
M’HAMDI, M.; ROUVIERE, C.; ROJAS-BELTRAN, J.; DU JAR-DIN, P. 2003. Optimisation de la transformation génétique de la ponme de terre par Agrobacterium tumefaciens. Utilisation de la résistance à l’hygromycine comme marqueur sélectif. Biotechnol. Agron. Soc. Environ. 7 (3-4):183-188.
MILLAM, S. 2007. Potato (Solanum tuberosum L.). Agrobacteri-um Protocols Volume 2, 25-35.
MONSERRAT, E.; MARFA, V.; MELÉ, E.; MESSENGUER, J. 2001. Study of different antibiotic combinations for use in the elim-ination of Agrobacterium with kanamycin selection in cRNAation. Plant Cell Tissue and Organ Culture 65: 211-220. DOI: http://dx.do i.org/10.1023/A:1010630726444.
MURASHIGE, T.; SKOOG, F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 15, 473-497.
NADAKUDUTI, S.S.; STARKER, C.G.; VOYTAS, D.F.; BUELL, C.R.; DOUCHES, D.S. 2019. Genome editing in potato with CRIS-PR/Cas9. In Plant Genome Editing with CRISPR Systems . Huma-na Press, Nueva York. 183-201 pp.
OKADA, Y.; SAITO, A.; NISHIGUCHI, M.; KIMURA, T.; MORI, M.; HANADA, K.; MURATA, T. 2001. Virus resistance in transgenic sweetpotato [Ipomoea batatas L.(Lam)] expressing the coat pro-tein gene of sweet potato feathery mottle virus. Theoretical and Applied Genetics, 103(5), 743-751.
ORBEGOZO, J.; ROMAN, M.; RIVERA, C. TOVAR, J.C., PÉREZ, W.; GAMBOA, S.; GHISLAIN, M. 2013. Agrotransforma-ción y evaluación de la resistencia a Phytophthora infestans en Solanum tuberosum L. variedad Désirée. Revista peruana de bio-logía, 20(3), 205-210.
ORBEGOZO, J.; ROMAN, M.; RIVERA, C.; GAMBOA, S.; TO-VAR, J.; FORBES, G.; GHISLAIN, M. 2016. Rpi-blb2 gene from Solanum bulbocastanum confers extreme resistance to late blight disease in potato. Plant Cell, Tissue and Organ Culture (PCTOC), 125(2), 269-281.
PANDEY, S.; MISHRA, A.; PATEL, M.K.; JHA, B. 2013. An effi-cient method for Agrobacterium-mediated genetic transformation and plant regeneration in cumin (Cuminum cyminum L.). Applied biochemistry and biotechnology, 171(1), 1-9.
PLACKETT, A.; HUANG, L.; SANDERS, H.; LANGDALE, J. 2014. High-efficiency stable transformation of the model fern spe-cies Ceratopteris richardii via microparticle bombardment. Plant physiology, 165(1), 3-14.
ROMÁN, M.L.; RIVERA, C.; ORBEGOZO, J.; SERNA, F.; GAM-BOA, S.; PEREZ, W., GHISLAN, M. 2015. Resistencia a Phyto-phthora infestans linaje clonal EC-1 en Solanum tuberosum me-diante la introducción del gen RB. Revista peruana de biología, 22(1), 63-70.
ROMANO, A.; RAEMAKERS, K.; BERNARDI, J.; VISSER, R.; MOOIBROEK, H. 2003. Transgene organisation in potato after particle bombardment-mediated (co-) transformation using plas-mids and gene cassettes. Transgen. Res. 12, 461-473.
SUROV, T.; AVIV, D.; ALY, R.; JOEL, D.M., GOLDMAN-GUEZ, T.; GRESSEL, J. 1998. Generation of transgenic asulam-resistant potatoes to facilitate eradication of parasitic broomrapes (Oro-banche spp.), with the sul gene as the selectable marker. Theoret-ical and Applied Genetics, 96(1), 132-137.
VAN DEN ELZEN, P.; TOWNSEND, J.; LEE, K.; BEDBROOK, J. 1985. A chimaeric hygromycin resistance gene as a selectable marker in plant cells.Plant Molecular Biology, 5(5), 299-302.
WATERER, D.; BENNING, N.T.; WU, G.; LUO, X.; LIU, X.; GUS-TA, M.; GUSTA, L.V. 2010. Evaluation of abiotic stress tolerance of genetically modified potatoes (Solanum tuberosum cv. Desiree). Molecular breeding, 25(3), 527-540.
WENDT, T.; HOLM, P.; STARKER, C.; CHRISTIAN, M.; VOY-TAS, D.; BRINCH-PEDERSEN, H.; HOLME, I. 2013. TAL effec-tor nucleases induce mutations at a pre-selected location in the genome of primary barley transformants. Plant molecular biology, 83(3), 279-285.

Evaluación de la calidad de subproductos de la soja
ARTÍCULOS RIA / Vol. 46 / N.º 2258258
Evaluación de la calidad de subproductos de la soja
RESUMENLos subproductos provenientes de la agroindustria suelen presentar una alta variabilidad en su calidad nu-
tricional. Esto resulta problemático al momento de utilizarlos en dietas destinadas a producción animal. En el presente trabajo se analizó mediante ensayos químicos, in sacco y encuestas el perfil nutricional y la variabi-lidad de expellers y harinas de soja provenientes de la región de Rafaela, Santa Fe. A partir de las muestras se observó que la variabilidad en los indicadores de la calidad nutricional de estos subproductos, tales como contenido de proteína bruta (PB), extracto etéreo (EE) y degradabilidad, no fue diferente a la reportada por la bibliografía nacional. Además, dicha variabilidad pudo ser explicada por factores tales como los contenidos de EE y fibra cruda (FC), y estuvo asociada con la solubilidad de la proteína en hidróxido de potasio (PB-KOH). En correspondencia con esto, los expellers presentaron menor degradabilidad que las harinas. Entre los factores del procesamiento industrial que modificaron la variabilidad del expeller se encontró que la limpieza del grano previo a la extracción del aceite aumentó el contenido de EE del poroto y disminuyó el del expeller, incrementando un 22% la eficiencia de extracción. El tratamiento térmico modificó las fracciones de degrada-bilidad a través de la disminución de la fracción a (inmediatamente soluble) y el aumento de la fracción b (po-tencialmente degradable). Además de limpiar el grano, este trabajo indica que para disminuir la variabilidad de estos subproductos deberían implementarse controles de calidad sencillos y confiables como PB-KOH. Esto permitiría generar planes de protocolización y reducir la variabilidad del EE en estos subproductos.
Palabras clave: expeller de soja, variabilidad nutricional, protocolos de proceso, evaluación de alimentos.
ABSTRACT
By-products from agribusiness tend to have high variability in nutritional quality. This is problematic when used in diets intended for animal production. In the present work, the nutritional profile and variability of ex-pellers and soy flour from the Rafaela, Santa Fe region were analyzed through chemical tests, in sacco and surveys. From the samples it was observed that the variability in the indicators of nutritional quality of these by-products, such as crude protein content (PB), ethereal extract (EE) and degradability, was not different from that reported by the national literature. In addition, this variability could be explained by factors such as the contents of EE and crude fiber (FC), and was associated with the solubility of the protein in potassium hydro-xide (PB-KOH). In correspondence with this, the expellers presented less degradability than the flours. Among the factors of industrial processing that modified the variability of the expeller, it was found that cleaning the grain prior to oil extraction increased the EE content of the bean and decreased that of the expeller, increasing
1Universidad de Buenos Aires (UBA), Facultad de Agronomía, Cátedra de Nutrición Animal. San Martín 4544, CABA. 2Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Rafaela, Ruta 34, km 227 (2300), Rafae-la, Santa Fe, Argentina. 3Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria (EEA) Oliveros, Agencia de Extensión Rural (AER) Totoras, Av. Maipú 1138 (2144). Correo electrónico: [email protected]
SEIJAS NOYA, I.1; JAURENA, G.1; WAWRZKIEWICZ, M.1; GAGGIOTTI, M.2; ROMERO, L.A.2; MÉNDEZ, M.3
Recibido 21 de abril de 2018 // Aceptado 05 de mayo de 2020 // Publicado online 19 de agosto de 2020

SEIJAS NOYA, I.; JAURENA, G.; WAWRZKIEWICZ, M.; GAGGIOTTI, M.; ROMERO, L.A.; MÉNDEZ, M.
Agosto 2020, Argentina 259259
the extraction efficiency by 22%. The heat treatment modified the degradability fractions through the decrease in fraction a (immediately soluble) and the increase in fraction b (potentially degradable). In addition to cleaning the grain, this work indicates that to reduce the variability of these by-products, simple and reliable quality con-trols such as PB-KOH should be implemented. This would allow generating protocolization plans and reducing EE variability in these by-products.
Keywords: soybean expeller, nutritional variability, process protocols, feed evaluation.
INTRODUCCIÓN
El principal objetivo de la industrialización de la soja es la obtención de aceite, sin embargo, los subproductos de dicho proceso (expellers y harinas) son de crucial impor-tancia económica y comercial por su masiva utilización para la alimentación animal. Estos resultan valiosos es-pecialmente por su alto contenido y calidad de proteína. Básicamente existen dos principios para la extracción del aceite: mediante extrusión y prensado, y mediante el uso de solventes orgánicos (también existen diversos sistemas que trabajan con ambos principios). El subproducto obteni-do por extrusión y prensado es conocido con el nombre de expeller, mientras que aquel excedente de la extracción por solventes se denomina harina.
Naturalmente, la variabilidad de calidad de materias pri-mas y de los diversos procesos de extracción que existen determinan que se generen subproductos de distinta cali-dad (INTA, 2011).
El poroto de soja se caracteriza por la presencia de com-puestos antinutricionales de naturaleza proteica, tales como inhibidores de tripsina y quimiotripsina, ureasa, fitohema-glutininas, y factores alergénicos como la glicinia y la beta-conglicinina, los cuales disminuyen la digestibilidad (Cinque, 2011) y valor nutricional del alimento. Sin embargo, dado que la mayoría de las proteínas de la soja son globulinas ter-mosensibles (Cheftel et al., 1989) los tratamientos térmicos desactivan dichos factores antinutricionales. Una incorrecta desactivación (por exceso o por defecto) puede traer apare-jado una disminución de la calidad de la proteína útil.
Existen diferentes determinaciones que pretenden eva-luar el valor nutricional de la soja y sus subproductos para las distintas especies animales, ya sea a través de la carac-terización del aporte y calidad de la proteína (ej. proteína bruta (PB), nitrógeno insoluble en detergente ácido (NIDA), solubilidad de proteína en hidróxido de potasio (PB-KOH)), o por el grado de actividad de los factores antinutricionales (ej. actividad ureásica, AU).
En el caso de los expellers, evaluaciones realizadas en la Estación Experimental Agrícola (EEA) INTA Rafaela mostraron valores de PB entre 30% y 42% (base seca), con degradabilidades de dicha PB que variaron desde 21% hasta 42% para vacas lecheras, y digestibilidades de la proteína no degradable que variaron entre 0% y 93% de
acuerdo a los cálculos del NRC (2001). Como se puede apreciar, la variación es muy significativa y hasta la actuali-dad hay escasos estudios que caractericen acabadamente estos ingredientes en nuestro país.
Las razones de esta variabilidad son diversas y respon-den a causas asociadas con las características propias de los cultivos (ej. genética, ambiente de cada ciclo) así como de los procesos de industrialización y conservación. La im-plementación de protocolos para los procesos industriales, la utilización de normativas y la fijación de estándares de calidad podría disminuir la variabilidad antes menciona-da y aumentar la calidad nutricional y comercial de estos subproductos, aumentando el valor agregado a la cadena agroindustrial argentina (Latimori et al., 2013).
El objetivo del trabajo fue contribuir a la caracterización químico-nutricional de los expellers de soja provenientes de la zona centro-norte de la provincia de Santa Fe, así como también describir la influencia de los procesos indus-triales utilizados sobre la calidad de estos.
MATERIALES Y MÉTODOS
Análisis de laboratorio y ensayos
Caracterización químico-nutricional
Se utilizaron tres expellers (identificados como Exp08, Exp10 y Exp17), obtenidos de la zona de influencia del INTA EEA Rafaela (Santa Fe). Adicionalmente, con el pro-pósito de contar con un patrón de comparación, se eva-luó una harina de soja (extraída con solvente) en su forma “cruda” (HS00), o tratada térmicamente a 116 ºC por 30 minutos (HS30) o por 60 minutos (HS60).
Las muestras fueron analizadas de acuerdo a los proto-colos sugeridos por el PROMEFA (Programa de Mejora-miento de Forrajes y Alimentos; Wawrzkiewicz y Jaurena, 2013). En breve, la materia seca (MS) total se obtuvo so-metiendo las muestras a 105 °C en estufa durante 4 h. Las cenizas (Ce) se obtuvieron por ignición a 550 °C durante 12 h (AOAC, 1990). El contenido de proteína bruta (PB) se es-timó multiplicando el dato de nitrógeno total (NT) por 6,25 (Kjeldahl, AOAC, 1995), y el extracto etéreo (EE) mediante lavados con éter de petróleo (AOAC, 1990). La FC se obtu-vo como el residuo seco luego de la digestión de las mues-tras a un medio ácido (ácido sulfúrico, 1,25% H2SO4) y otro alcalino (hidróxido de sodio, 1,25% NaOH) (AOAC, 1990).
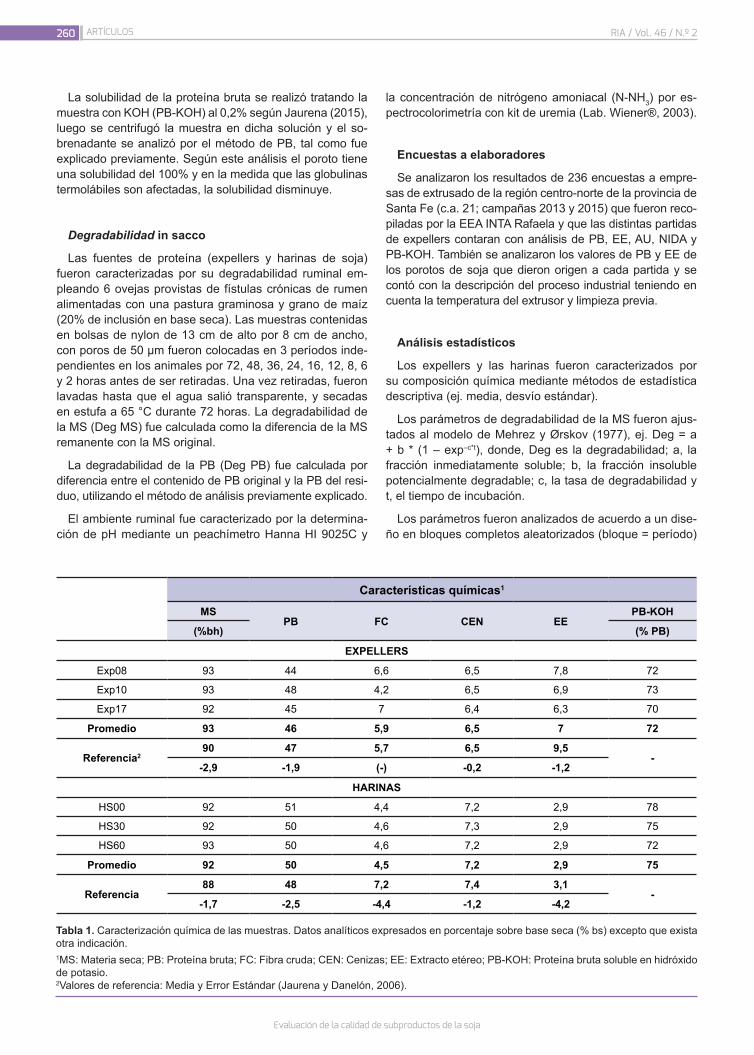
Evaluación de la calidad de subproductos de la soja
ARTÍCULOS RIA / Vol. 46 / N.º 2260260
La solubilidad de la proteína bruta se realizó tratando la muestra con KOH (PB-KOH) al 0,2% según Jaurena (2015), luego se centrifugó la muestra en dicha solución y el so-brenadante se analizó por el método de PB, tal como fue explicado previamente. Según este análisis el poroto tiene una solubilidad del 100% y en la medida que las globulinas termolábiles son afectadas, la solubilidad disminuye.
Degradabilidad in sacco
Las fuentes de proteína (expellers y harinas de soja) fueron caracterizadas por su degradabilidad ruminal em-pleando 6 ovejas provistas de fístulas crónicas de rumen alimentadas con una pastura graminosa y grano de maíz (20% de inclusión en base seca). Las muestras contenidas en bolsas de nylon de 13 cm de alto por 8 cm de ancho, con poros de 50 µm fueron colocadas en 3 períodos inde-pendientes en los animales por 72, 48, 36, 24, 16, 12, 8, 6 y 2 horas antes de ser retiradas. Una vez retiradas, fueron lavadas hasta que el agua salió transparente, y secadas en estufa a 65 °C durante 72 horas. La degradabilidad de la MS (Deg MS) fue calculada como la diferencia de la MS remanente con la MS original.
La degradabilidad de la PB (Deg PB) fue calculada por diferencia entre el contenido de PB original y la PB del resi-duo, utilizando el método de análisis previamente explicado.
El ambiente ruminal fue caracterizado por la determina-ción de pH mediante un peachímetro Hanna HI 9025C y
Tabla 1. Caracterización química de las muestras. Datos analíticos expresados en porcentaje sobre base seca (% bs) excepto que exista otra indicación.1MS: Materia seca; PB: Proteína bruta; FC: Fibra cruda; CEN: Cenizas; EE: Extracto etéreo; PB-KOH: Proteína bruta soluble en hidróxido de potasio.2Valores de referencia: Media y Error Estándar (Jaurena y Danelón, 2006).
la concentración de nitrógeno amoniacal (N-NH3) por es-pectrocolorimetría con kit de uremia (Lab. Wiener®, 2003).
Encuestas a elaboradores
Se analizaron los resultados de 236 encuestas a empre-sas de extrusado de la región centro-norte de la provincia de Santa Fe (c.a. 21; campañas 2013 y 2015) que fueron reco-piladas por la EEA INTA Rafaela y que las distintas partidas de expellers contaran con análisis de PB, EE, AU, NIDA y PB-KOH. También se analizaron los valores de PB y EE de los porotos de soja que dieron origen a cada partida y se contó con la descripción del proceso industrial teniendo en cuenta la temperatura del extrusor y limpieza previa.
Análisis estadísticos
Los expellers y las harinas fueron caracterizados por su composición química mediante métodos de estadística descriptiva (ej. media, desvío estándar).
Los parámetros de degradabilidad de la MS fueron ajus-tados al modelo de Mehrez y Ørskov (1977), ej. Deg = a + b * (1 – exp–c*t), donde, Deg es la degradabilidad; a, la fracción inmediatamente soluble; b, la fracción insoluble potencialmente degradable; c, la tasa de degradabilidad y t, el tiempo de incubación.
Los parámetros fueron analizados de acuerdo a un dise-ño en bloques completos aleatorizados (bloque = período)
Características químicas1
MSPB FC CEN EE
PB-KOH
(%bh) (% PB)
EXPELLERS
Exp08 93 44 6,6 6,5 7,8 72
Exp10 93 48 4,2 6,5 6,9 73
Exp17 92 45 7 6,4 6,3 70
Promedio 93 46 5,9 6,5 7 72
Referencia290 47 5,7 6,5 9,5
--2,9 -1,9 (-) -0,2 -1,2
HARINAS
HS00 92 51 4,4 7,2 2,9 78
HS30 92 50 4,6 7,3 2,9 75
HS60 93 50 4,6 7,2 2,9 72
Promedio 92 50 4,5 7,2 2,9 75
Referencia88 48 7,2 7,4 3,1
--1,7 -2,5 -4,4 -1,2 -4,2

SEIJAS NOYA, I.; JAURENA, G.; WAWRZKIEWICZ, M.; GAGGIOTTI, M.; ROMERO, L.A.; MÉNDEZ, M.
Agosto 2020, Argentina 261261
y se comprobaron los resultados por normalidad y homoce-dasticidad. También se evaluó linealidad entre temperatura aplicada a las HS y degradabilidad, y se comparó por con-trastes ortogonales. Los análisis se realizaron utilizando el software InfoStat (2009).
Los resultados de las encuestas se resumieron por es-tadística descriptiva, y se analizaron por ANVA de acuerdo a un modelo anidado, identificando como fuentes de va-riación las plantas de origen, el año y la temperatura de extrusión. Las medias fueron comparadas por contrastes ortogonales. Se analizó la PB y el EE del expeller en un modelo que tuvo como covariables la PB y el EE del poroto correspondiente para corroborar si la variabilidad de la ma-teria prima aporta de manera significativa a la variabilidad del subproducto final. En todos los casos las diferencias se declararon significativas cuando p<0,05.
RESULTADOS Y DISCUSIÓN
Análisis de laboratorio y ensayos
Caracterización químico-nutricional
Los resultados de los ingredientes estudiados (tabla 1) se encontraron dentro de los rangos esperados de acuerdo a lo reportado anteriormente (Jaurena et al., 2006).
En comparación con las harinas, los expellers presen-taron mayor concentración de EE (7,0 y 2,9% respectiva-mente), y menor contenido de PB (50 y 46%), diferencia que se explica por el proceso de extracción lipídica más eficiente en las harinas (Coscia, 1982), lo cual genera una mayor acumulación de compuestos remanentes.
Los resultados de PB-KOH para los expellers (70% - 73%) sugirieron que el tratamiento térmico fue superior a lo esperado (75%-85%, Gallardo, 2008). En cuanto a las harinas, se reportaron valores adecuados para la muestra testigo (HS00) y para el tratamiento térmico HS30. Por el contrario, HS60 presentó un valor de PB-KOH (72%) que sugirió una disminución en su degradabilidad potencial por exceso de calor. La variabilidad de los subproductos ana-lizados fue escasa, inferior a 10 unidades de porcentaje (rango de valores entre 70% y 78%), como probable con-secuencia del número limitado de materiales analizados.
Según Dale (1988), la tasa de crecimiento de pollos para carne fue menor cuando las harinas fueron insuficiente-mente procesadas (87% de PB-KOH) y también cuando sufrieron un daño excesivo (≤72% de PB-KOH). Esto se debería a que un tratamiento térmico insuficiente no desac-tiva de manera completa los factores antinutricionales, y un tratamiento excesivo produce la desnaturalización de ami-noácidos de alto valor biológico como la lisina, en conjunto con las condiciones predisponentes para las reacciones de Maillard. Estas reacciones ocurren cuando la soja es sobrecalentada, y el grupo amino epsilon de principalmen-te lisina (y otros aminoácidos) se une irreversiblemente al grupo carbonil proporcionado por los azúcares reducibles (Dale, 1988).
La solubilidad de la PB en KOH para subproductos de soja estaría asociada con la digestibilidad verdadera de los ami-noácidos (Martínez et al., 1996). Según estos lineamientos, las fuentes proteicas corresponderían a expellers y harinas de calidad media, con una consecuente reducción de la dis-ponibilidad de proteína digestible para monogástricos y ru-miantes jóvenes debido a un tratamiento térmico excesivo.
Degradabilidad in sacco
No se hallaron diferencias significativas entre ambientes ruminales de los animales ni valores irregulares para pH (Krause y Oetzel, 2006) ni para N-NH3 (Okorie et al., 1977; Schaefer et al., 1980).
La degradabilidad de la MS de las harinas de soja fue mayor que la de los expellers (tabla 2), en coincidencia con las diferencias en solubilidad en KOH reportadas en la tabla 1. No obstante lo anterior, la degradabillidad para ambos ingredientes fue mayor a 90% de degradabilidad potencial (a+b). Estos resultados son coincidentes con lo reportado por Moretto (2015). Además, el contenido de PB-KOH en-contrado en las harinas de este trabajo estuvo acorde con lo esperado según la bibliografía para un tratamiento tér-mico correcto, mientras que en los expellers este indicador sugiere un tratamiento térmico de mayor intensidad, y con ello menor degradabilidad (Gallardo, 2008).
La mayor degradabilidad de las harinas de soja estuvo asociada con un menor contenido de EE y FC. Los lípidos son compuestos menos susceptibles al ataque enzimático de las bacterias ruminales (Jenkins, 1993), y que en exce-so pueden inhibir el crecimiento de los microorganismos con actividad fibrolítica (Palmquist y Jenkins, 1980). El con-tenido de EE de los expellers (6,3% - 7,8%) fue cercano al límite a partir del cual presentarían efectos tóxicos sobre el proceso de digestión microbiana. Hess et al. (2001) obser-varon un descenso lineal significativo de la digestibilidad total en respuesta a la inclusión de 2,9%-6,2% de EE para la ración total en novillos de carne alimentándose de heno de gramíneas y suplementados con aceite de soja.
De igual manera, el contenido de FC se correlaciona ne-gativamente con la degradabilidad y digestibilidad del ali-mento (NRC, 1989).
Para el caso de las harinas, la duración del proceso tér-mico mostró una relación lineal negativa sobre la fracción de MS inmediatamente soluble y positiva para la fracción b (MS potencialmente degradable), sugiriendo un “traspaso” de la fracción “a” hacia “b”.
La fracción “a” de los expellers fue mayor a la de las hari-nas, pero concomitantemente los expellers mostraron una menor fracción insoluble potencialmente degradable (pará-metro b) y menor tasa de degradación (parámetro c).
La degradabilidad in sacco de la PB de las harinas fue mayor a la de los expellers, independientemente del tra-tamiento térmico. Para los expellers, el 86% de la PB po-tencialmente degradable desapareció hacia las 16 horas de permanencia en el rumen, mientras que para la HS00 alcanzó el 95%.
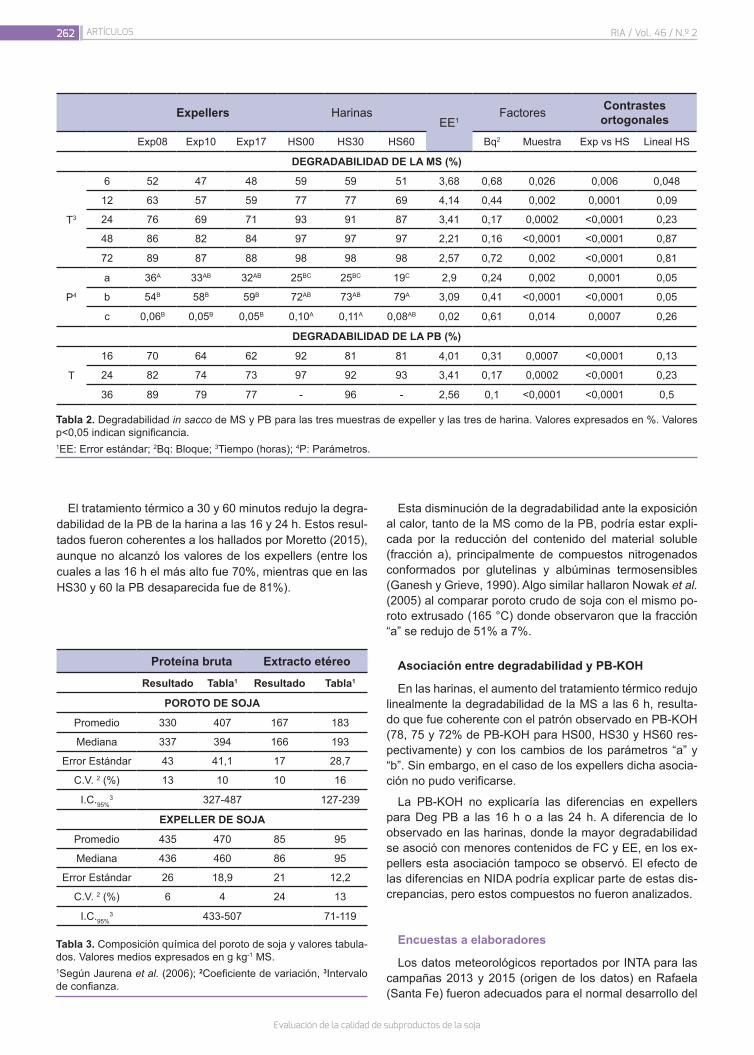
Evaluación de la calidad de subproductos de la soja
ARTÍCULOS RIA / Vol. 46 / N.º 2262262
El tratamiento térmico a 30 y 60 minutos redujo la degra-dabilidad de la PB de la harina a las 16 y 24 h. Estos resul-tados fueron coherentes a los hallados por Moretto (2015), aunque no alcanzó los valores de los expellers (entre los cuales a las 16 h el más alto fue 70%, mientras que en las HS30 y 60 la PB desaparecida fue de 81%).
Tabla 2. Degradabilidad in sacco de MS y PB para las tres muestras de expeller y las tres de harina. Valores expresados en %. Valores p<0,05 indican significancia.1EE: Error estándar; 2Bq: Bloque; 3Tiempo (horas); 4P: Parámetros.
Tabla 3. Composición química del poroto de soja y valores tabula-dos. Valores medios expresados en g kg-1 MS.1Según Jaurena et al. (2006); 2Coeficiente de variación, 3Intervalo de confianza.
Expellers HarinasEE1
Factores Contrastes ortogonales
Exp08 Exp10 Exp17 HS00 HS30 HS60 Bq2 Muestra Exp vs HS Lineal HS
DEGRADABILIDAD DE LA MS (%)
T3
6 52 47 48 59 59 51 3,68 0,68 0,026 0,006 0,048
12 63 57 59 77 77 69 4,14 0,44 0,002 0,0001 0,09
24 76 69 71 93 91 87 3,41 0,17 0,0002 <0,0001 0,23
48 86 82 84 97 97 97 2,21 0,16 <0,0001 <0,0001 0,87
72 89 87 88 98 98 98 2,57 0,72 0,002 <0,0001 0,81
P4
a 36A 33AB 32AB 25BC 25BC 19C 2,9 0,24 0,002 0,0001 0,05
b 54B 58B 59B 72AB 73AB 79A 3,09 0,41 <0,0001 <0,0001 0,05
c 0,06B 0,05B 0,05B 0,10A 0,11A 0,08AB 0,02 0,61 0,014 0,0007 0,26
DEGRADABILIDAD DE LA PB (%)
T
16 70 64 62 92 81 81 4,01 0,31 0,0007 <0,0001 0,13
24 82 74 73 97 92 93 3,41 0,17 0,0002 <0,0001 0,23
36 89 79 77 - 96 - 2,56 0,1 <0,0001 <0,0001 0,5
Proteína bruta Extracto etéreoResultado Tabla1 Resultado Tabla1
POROTO DE SOJA
Promedio 330 407 167 183
Mediana 337 394 166 193
Error Estándar 43 41,1 17 28,7
C.V. 2 (%) 13 10 10 16
I.C.95%3 327-487 127-239
EXPELLER DE SOJA
Promedio 435 470 85 95
Mediana 436 460 86 95
Error Estándar 26 18,9 21 12,2
C.V. 2 (%) 6 4 24 13
I.C.95%3 433-507 71-119
Esta disminución de la degradabilidad ante la exposición al calor, tanto de la MS como de la PB, podría estar expli-cada por la reducción del contenido del material soluble (fracción a), principalmente de compuestos nitrogenados conformados por glutelinas y albúminas termosensibles (Ganesh y Grieve, 1990). Algo similar hallaron Nowak et al. (2005) al comparar poroto crudo de soja con el mismo po-roto extrusado (165 °C) donde observaron que la fracción “a” se redujo de 51% a 7%.
Asociación entre degradabilidad y PB-KOH
En las harinas, el aumento del tratamiento térmico redujo linealmente la degradabilidad de la MS a las 6 h, resulta-do que fue coherente con el patrón observado en PB-KOH (78, 75 y 72% de PB-KOH para HS00, HS30 y HS60 res-pectivamente) y con los cambios de los parámetros “a” y “b”. Sin embargo, en el caso de los expellers dicha asocia-ción no pudo verificarse.
La PB-KOH no explicaría las diferencias en expellers para Deg PB a las 16 h o a las 24 h. A diferencia de lo observado en las harinas, donde la mayor degradabilidad se asoció con menores contenidos de FC y EE, en los ex-pellers esta asociación tampoco se observó. El efecto de las diferencias en NIDA podría explicar parte de estas dis-crepancias, pero estos compuestos no fueron analizados.
Encuestas a elaboradores
Los datos meteorológicos reportados por INTA para las campañas 2013 y 2015 (origen de los datos) en Rafaela (Santa Fe) fueron adecuados para el normal desarrollo del
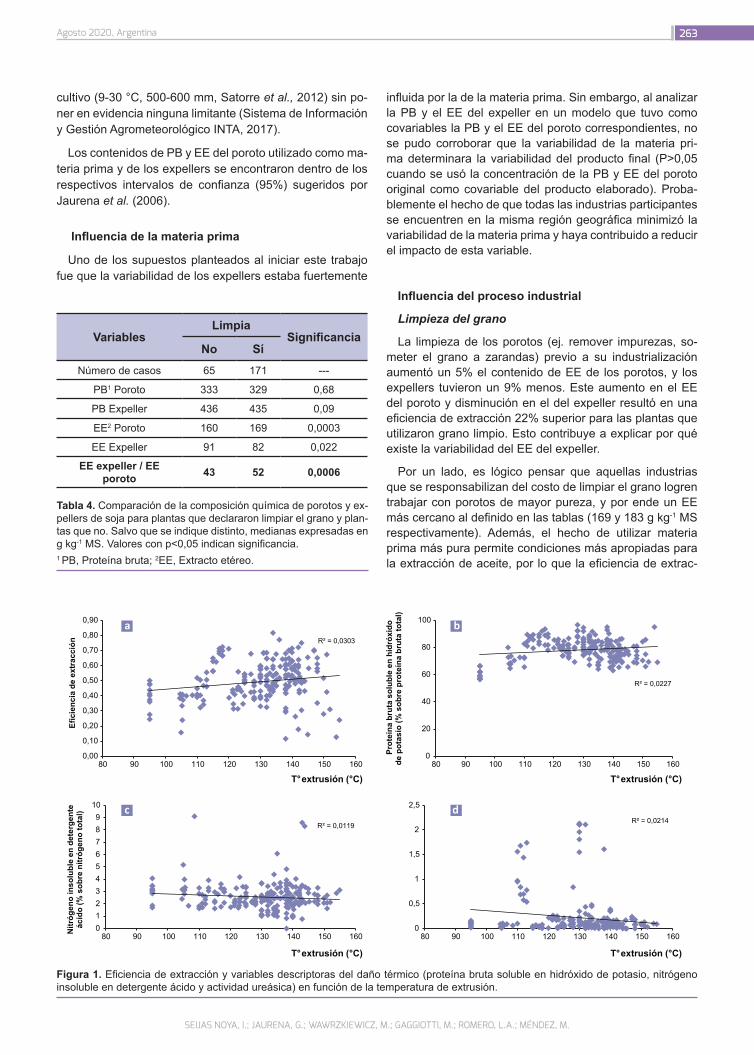
SEIJAS NOYA, I.; JAURENA, G.; WAWRZKIEWICZ, M.; GAGGIOTTI, M.; ROMERO, L.A.; MÉNDEZ, M.
Agosto 2020, Argentina 263263
cultivo (9-30 °C, 500-600 mm, Satorre et al., 2012) sin po-ner en evidencia ninguna limitante (Sistema de Información y Gestión Agrometeorológico INTA, 2017).
Los contenidos de PB y EE del poroto utilizado como ma-teria prima y de los expellers se encontraron dentro de los respectivos intervalos de confianza (95%) sugeridos por Jaurena et al. (2006).
Influencia de la materia prima
Uno de los supuestos planteados al iniciar este trabajo fue que la variabilidad de los expellers estaba fuertemente
Tabla 4. Comparación de la composición química de porotos y ex-pellers de soja para plantas que declararon limpiar el grano y plan-tas que no. Salvo que se indique distinto, medianas expresadas en g kg-1 MS. Valores con p<0,05 indican significancia.1 PB, Proteína bruta; 2EE, Extracto etéreo.
Figura 1. Eficiencia de extracción y variables descriptoras del daño térmico (proteína bruta soluble en hidróxido de potasio, nitrógeno insoluble en detergente ácido y actividad ureásica) en función de la temperatura de extrusión.
VariablesLimpia
SignificanciaNo Sí
Número de casos 65 171 ---
PB1 Poroto 333 329 0,68
PB Expeller 436 435 0,09
EE2 Poroto 160 169 0,0003
EE Expeller 91 82 0,022
EE expeller / EE poroto 43 52 0,0006
influida por la de la materia prima. Sin embargo, al analizar la PB y el EE del expeller en un modelo que tuvo como covariables la PB y el EE del poroto correspondientes, no se pudo corroborar que la variabilidad de la materia pri-ma determinara la variabilidad del producto final (P>0,05 cuando se usó la concentración de la PB y EE del poroto original como covariable del producto elaborado). Proba-blemente el hecho de que todas las industrias participantes se encuentren en la misma región geográfica minimizó la variabilidad de la materia prima y haya contribuido a reducir el impacto de esta variable.
Influencia del proceso industrial
Limpieza del grano
La limpieza de los porotos (ej. remover impurezas, so-meter el grano a zarandas) previo a su industrialización aumentó un 5% el contenido de EE de los porotos, y los expellers tuvieron un 9% menos. Este aumento en el EE del poroto y disminución en el del expeller resultó en una eficiencia de extracción 22% superior para las plantas que utilizaron grano limpio. Esto contribuye a explicar por qué existe la variabilidad del EE del expeller.
Por un lado, es lógico pensar que aquellas industrias que se responsabilizan del costo de limpiar el grano logren trabajar con porotos de mayor pureza, y por ende un EE más cercano al definido en las tablas (169 y 183 g kg-1 MS respectivamente). Además, el hecho de utilizar materia prima más pura permite condiciones más apropiadas para la extracción de aceite, por lo que la eficiencia de extrac-
R² = 0,0303
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
0,90
80 90 100 110 120 130 140 150 160
Efic
ienc
ia d
e ex
trac
ción
R² = 0,0119
0123456789
10
80 90 100 110 120 130 140 150 160Nitr
ógen
o in
solu
ble
en d
eter
gent
e ác
ido
(% s
obre
nitr
ógen
o to
tal)
T°extrusión (°C) T°extrusión (°C)
T°extrusión (°C) T°extrusión (°C)
R² = 0,0227
0
20
40
60
80
100
80 90 100 110 120 130 140 150 160
Prot
eína
bru
ta s
olub
le e
n hi
dróx
ido
de p
otas
io (%
sob
re p
rote
ína
brut
a to
tal)
R² = 0,0214
0
0,5
1
1,5
2
2,5
80 90 100 110 120 130 140 150 160
a b
c d
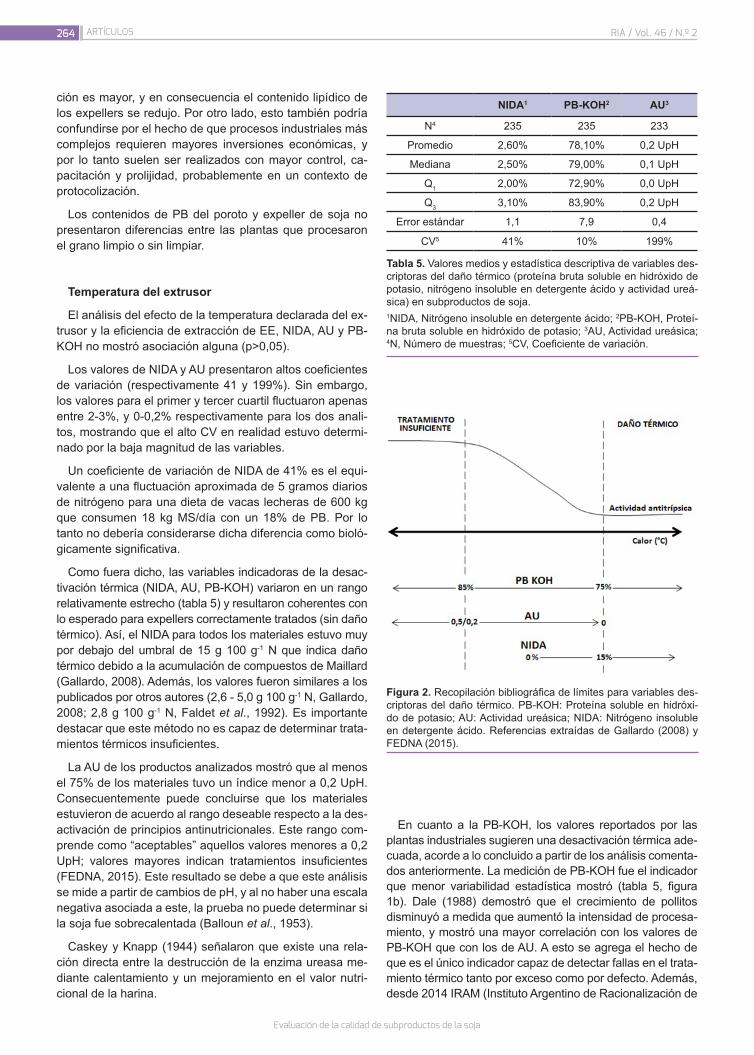
Evaluación de la calidad de subproductos de la soja
ARTÍCULOS RIA / Vol. 46 / N.º 2264264
ción es mayor, y en consecuencia el contenido lipídico de los expellers se redujo. Por otro lado, esto también podría confundirse por el hecho de que procesos industriales más complejos requieren mayores inversiones económicas, y por lo tanto suelen ser realizados con mayor control, ca-pacitación y prolijidad, probablemente en un contexto de protocolización.
Los contenidos de PB del poroto y expeller de soja no presentaron diferencias entre las plantas que procesaron el grano limpio o sin limpiar.
Temperatura del extrusor
El análisis del efecto de la temperatura declarada del ex-trusor y la eficiencia de extracción de EE, NIDA, AU y PB-KOH no mostró asociación alguna (p>0,05).
Los valores de NIDA y AU presentaron altos coeficientes de variación (respectivamente 41 y 199%). Sin embargo, los valores para el primer y tercer cuartil fluctuaron apenas entre 2-3%, y 0-0,2% respectivamente para los dos anali-tos, mostrando que el alto CV en realidad estuvo determi-nado por la baja magnitud de las variables.
Un coeficiente de variación de NIDA de 41% es el equi-valente a una fluctuación aproximada de 5 gramos diarios de nitrógeno para una dieta de vacas lecheras de 600 kg que consumen 18 kg MS/día con un 18% de PB. Por lo tanto no debería considerarse dicha diferencia como bioló-gicamente significativa.
Como fuera dicho, las variables indicadoras de la desac-tivación térmica (NIDA, AU, PB-KOH) variaron en un rango relativamente estrecho (tabla 5) y resultaron coherentes con lo esperado para expellers correctamente tratados (sin daño térmico). Así, el NIDA para todos los materiales estuvo muy por debajo del umbral de 15 g 100 g-1 N que indica daño térmico debido a la acumulación de compuestos de Maillard (Gallardo, 2008). Además, los valores fueron similares a los publicados por otros autores (2,6 - 5,0 g 100 g-1 N, Gallardo, 2008; 2,8 g 100 g-1 N, Faldet et al., 1992). Es importante destacar que este método no es capaz de determinar trata-mientos térmicos insuficientes.
La AU de los productos analizados mostró que al menos el 75% de los materiales tuvo un índice menor a 0,2 UpH. Consecuentemente puede concluirse que los materiales estuvieron de acuerdo al rango deseable respecto a la des-activación de principios antinutricionales. Este rango com-prende como “aceptables” aquellos valores menores a 0,2 UpH; valores mayores indican tratamientos insuficientes (FEDNA, 2015). Este resultado se debe a que este análisis se mide a partir de cambios de pH, y al no haber una escala negativa asociada a este, la prueba no puede determinar si la soja fue sobrecalentada (Balloun et al., 1953).
Caskey y Knapp (1944) señalaron que existe una rela-ción directa entre la destrucción de la enzima ureasa me-diante calentamiento y un mejoramiento en el valor nutri-cional de la harina.
NIDA1 PB-KOH2 AU3
N4 235 235 233
Promedio 2,60% 78,10% 0,2 UpH
Mediana 2,50% 79,00% 0,1 UpH
Q1 2,00% 72,90% 0,0 UpH
Q3 3,10% 83,90% 0,2 UpH
Error estándar 1,1 7,9 0,4
CV5 41% 10% 199%
Tabla 5. Valores medios y estadística descriptiva de variables des-criptoras del daño térmico (proteína bruta soluble en hidróxido de potasio, nitrógeno insoluble en detergente ácido y actividad ureá-sica) en subproductos de soja.1NIDA, Nitrógeno insoluble en detergente ácido; 2PB-KOH, Proteí-na bruta soluble en hidróxido de potasio; 3AU, Actividad ureásica; 4N, Número de muestras; 5CV, Coeficiente de variación.
Figura 2. Recopilación bibliográfica de límites para variables des-criptoras del daño térmico. PB-KOH: Proteína soluble en hidróxi-do de potasio; AU: Actividad ureásica; NIDA: Nitrógeno insoluble en detergente ácido. Referencias extraídas de Gallardo (2008) y FEDNA (2015).
En cuanto a la PB-KOH, los valores reportados por las plantas industriales sugieren una desactivación térmica ade-cuada, acorde a lo concluido a partir de los análisis comenta-dos anteriormente. La medición de PB-KOH fue el indicador que menor variabilidad estadística mostró (tabla 5, figura 1b). Dale (1988) demostró que el crecimiento de pollitos disminuyó a medida que aumentó la intensidad de procesa-miento, y mostró una mayor correlación con los valores de PB-KOH que con los de AU. A esto se agrega el hecho de que es el único indicador capaz de detectar fallas en el trata-miento térmico tanto por exceso como por defecto. Además, desde 2014 IRAM (Instituto Argentino de Racionalización de

SEIJAS NOYA, I.; JAURENA, G.; WAWRZKIEWICZ, M.; GAGGIOTTI, M.; ROMERO, L.A.; MÉNDEZ, M.
Agosto 2020, Argentina 265265
Materiales) incluye la solubilidad de PB-KOH como método estándar para la determinación de proteínas solubles en ha-rina de soja, harina de colza y pellets de girasol (IRAM 5614 / ISO 14244:2015). De lo anterior, se desprende que PB-KOH constituiría un indicador básico para tener en cuenta a la hora de evaluar los subproductos de soja.
CONCLUSIONES
Uno de los objetivos centrales de este trabajo fue ca-racterizar la calidad de los expellers de soja de la zona de Rafaela (Santa Fe). Se pudo concluir que la variabilidad en la calidad nutricional (PB, EE, Deg MS, Deg PB) de los subproductos de la soja no difirió de la reportada en la bi-bliografía nacional. Dicha variabilidad pudo ser explicada por los contenidos de EE y FC, y estuvo asociada con PB-KOH. En adición y en correspondencia con las caracterís-ticas composicionales antes señaladas, los expellers pre-sentaron menor degradabilidad que las harinas.
Entre los factores del procesamiento industrial que modi-ficaron la variabilidad del expeller, la limpieza de los granos previo a la extracción del aceite aumentó la concentración de lípidos en la materia prima, y se asoció con menores contenidos de EE en el expeller, generando una eficiencia de extracción un 22% mayor.
Por un lado, también se corroboró que el tratamiento tér-mico disminuyó la fracción a (inmediatamente soluble) y aumentó la fracción b (potencialmente degradable) de las harinas.
Por otro lado, no se halló correlación entre la temperatu-ra de extrusión declarada y las concentraciones de NIDA, AU o PB-KOH. Dichas variables se mantuvieron dentro de los límites recomendados según la bibliografía, de lo que se deduce que es probable que las plantas hayan utilizado tratamientos térmicos adecuados.
Este trabajo indica que uno de los procesos que puede aumentar la eficiencia del uso de los subproductos de la soja es trabajar con grano limpio, así como implementar controles de calidad sencillos y confiables como PB-KOH.
BIBLIOGRAFÍAAOAC International (formerly the Association of Official Analyt-
ical Chemists). 1990. Official Methods of Analysis. Arlington, VA: AOAC International, N.o 942.05. 962.09. 920.85.
AOAC International (formerly the Association of Official Analyt-ical Chemists). 1995. Official Methods of Analysis. Arlington, VA: AOAC International.
BALLOUN, S.; JOHNSON, E.; ARNOLD, L.K. 1953. Laboratory Estimation of the Nutritive Value of Soybean Oil Meals Poult Sci 32 (3): 517-527 pp.
CASKEY, C.D.; KNAPP, F.C. 1944. Method for detecting inadequate-ly heated soybean oil meal. Southern States Labs; Baltimore, Md. In-dustrial and Engineering Chemistry.Analytical.Vol.16.640-641 pp.
CHEFTEL, J.C.; CUQ, J.L.; LORIENT, D. 1989. Las proteínas de soja. Proteínas Alimentarias, Editorial Acribia S.A. Zaragoza, España. 257-275 pp.
CINQUE, F. 2011. Proyecto de inversión de una planta extrusado-ra de soja: Análisis económico y comercial. Trabajo Final. Facultad de Ciencias Agrarias, Universidad Católica Argentina. 16-17 pp.
COSCIA, A. 1982. Economía de las Oleaginosas. Ed. Hemisfe-rio Sur. 7-10 pp.
DALE, N. 1988. Solubilidad de la proteína: indicador del proce-sado de la harina de soja. Avicultura profesional, 5:122 p.
FALDET, M.A.; SON, Y.S.; SATTER, L.D. 1992. Chemical, in vitro, and in vivo evaluation of soybeans heat-treated by various processing methods. Journal of dairy science, 75(3), 789-795.
FUNDACIÓN ESPAÑOLA PARA EL DESARROLLO DE LA NU-TRICIÓN ANIMAL (FEDNA). 2015. (Disponible: http://www.funda-cionfedna.org/ ingredientes_para_piensos/harina-de-soja-47-pb verificado: 16 de abril de 2015).
GALLARDO, M. 2005. Soja: harinas de extracción para la ali-mentación del ganado. Instituto Nacional de Tecnología Agrope-cuaria (INTA).
GALLARDO, M. 2008. Concentrados y subproductos para la ali-mentación de rumiantes. xxi Curso Internacional de lechería para profesionales de América Latina. 153-162 pp.
GALLARDO, M. 2013. Utilización de subproductos agroindus-triales para formular y balancear dietas. Instituto Nacional de Tec-nología Agropecuaria (INTA). 3-9 pp.
GANESH, D.; GRIEVE, D.G. 1990. Effect of roasting raw soy-beans at three temperatures on in situ dry matter and nitrogen disap-pearance in dairy cows. Journal of dairy science, 73(11), 3222-3230.
HESS, B.W.; WHITNEY, M.B.; RULE, D.C. 2001. Site and extent of digestion by beef heifers fed medium-quality hay and supple-mental corn or soybean oil. Proc. West. Sect. Am. Soc. Anim. Sci., vol. 52, 469-472 pp.
INFOSTAT, 2009. Versión 3.0. Grupo Infostat. FCA. Universidad Nacional de Córdoba, Ed. Brujas. Córdoba, Argentina.
INTA. 2011. Seguimiento de la calidad del expeller de soja ela-borado por diferentes plantas productoras. Informe Interno del Proyecto Nacional Procesos Productivos Agroindustriales para Agregar Valor en Origen en forma Sustentable. 4 p.
INTA. 2017. Sistema de Información y Gestión Agrometeorológi-co. (http://siga2.inta.gov.ar/).
IRAM. 2015. Proceso de estandarización de una metodología analítica. De Argentina a ISO “Proteínas Solubles en harinas de soja”. (Disponible: asaga.org.ar/descargas/material/SESION_ AACI/AACI2_Fernandez.pdf verificado: 11 de junio de 2015).
JAURENA, G.; DANELÓN, J.L. 2006. Tabla de composición de alimentos para rumiantes de la región pampeana argentina. He-misferio Sur. 52 pp.
JAURENA, G. 2015. Determinación de Proteína Soluble en Hi-dróxido de Potasio (recopilación). Programa para el Mejoramiento de la Evaluación de Forrajes y Alimentos (PROMEFA).
JENKINS, T.C. 1993. Lipid Metabolism in the Rumen. Journal of Dairy Science 76:3851-3863.
KRAUSE, K.M.; OETZEL, G.R. 2006. Understanding and pre-venting subacute ruminal acidosis in dairy herds: A review. Animal Feed Science and Technology 126: 215.
LATIMORI, N.; KLOSTER, A.; GARIS, M. 2013. Uso de expeller de soja como sustituto de la harina de soja en dietas de engorde de bovinos a corral. Información para Extensión N.º 143. Proyecto Nacional: Procesos productivos agroindustriales para agregar va-lor en origen en forma sustentable. Instituto Nacional de tecnología Agropecuaria (INTA). 5 p.
MARTÍNEZ, A.; CUCA-GARCÍA, M.; MENDOZA-MARTÍNEZ, G.; HERRERA-HARO, J.G. 1996. Digestibilidad y valor nutritivo de aminoácidos de sorgo y soja en diversas formas de dietas para pollos de engorda. Arch. Latinoam. Prod. Anim. 4 (1):7.

Evaluación de la calidad de subproductos de la soja
ARTÍCULOS RIA / Vol. 46 / N.º 2266266
MEHREZ, A.Z.; ØRSKOV, E.R. 1977. A study of artificial fibre bag technique for determining the digestibily of feeds in the rumen. The Journal of Agricultural Science, Volume 88, Issue 3. 645-650 pp.
MORETTO, F. 2015. Efecto de la temperatura de extrusión del grano de soja sobre la degradación ruminal de la proteína del ex-peller. Universidad Nacional de Villa María, Córdoba. 20-28 pp.
NATIONAL RESEARCH COUNCIL (NRC). 1989. The nutri-ent requirements of dairy cattle. Sixth edition. National Academy Press, Washington, D. C.
NATIONAL RESEARCH COUNCIL (NRC). 2001. Nutrient Re-quirement of Dairy Cattle, seventh ed. National Research Council, National Academy Press, Washington, D. C.
NOWAK, W.; MICHALAK, S.; WYLEGALA, S. 2005. In situ eval-uation of ruminal degradability and intestinal digestibility of extrud-ed soybeans. Czech J. Anim. Sci, 50, 281-287.
OKORIE, A.U.; BUTTERY, P.J.; LEXIS, D. 1977. Amonnia con-centration and protein synthesis in the rumen. Abstracts of commu-nications. Proc. Nutr. Soc. 36. 38 p.
PALMQUIST, D.L.; JENKINS, T.C. 1980. Fat in lactation rations: review. Journal of Dairy Science, vol. 63. 1-14 pp.
SATORRE, E.; BENECH ARNOLD, R.; SLAFER, G.; DE LA FUENTE, E.; MIRALLES, D.; OTEGUI, M.; SAVIN, R. 2012. Producción de granos. Bases funcionales para su manejo. EU-DEBA.
SCHAEFER, D.M.; DAVIS, C.L.; BRYANT, M.P. 1980. Ammonia saturation constants for predominant species of rumen bacteria. Journal of Dairy Science, 63:1248-1263.
SISTEMA INTEGRADO DE INFORMACIÓN AGROPECUARIA. 2015. Ministerio de Agricultura, Ganadería y Pesca. (Disponible: http://www. siia.gob.ar/ verificado: 17 de septiembre de 2017).
WAWRZKIEWICZ, M.; JAURENA, G.; MARTÍNEZ, R.; CANTET, J.M. 2013. Protocolos de trabajo LASERNU y PROMEFA. Cáte-dra de Nutrición Animal, Facultad de Agronomía, Universidad de Buenos Aires.
WIENER® LAB. 2003. Protocolo de uso. UREMIA, del kit de uremia de laboratorios Wiener®.

BAILLERES, M.; CAMPESTRE, M.P.; ANTONELLI, C.J.; MELANI, G.; MENÉNDEZ, A.; RUIZ, O.A.
Agosto 2020, Argentina 267267
Promotion of Lotus tenuis and calf early weaning as a good management practice for breeding herds in marginal soils of the Flooding Pampa (Argentina)
ABSTRACTThe promotion of forage legumes in marginal environments for agriculture constitutes a major technological
challenge for livestock farming. The legume Lotus tenuis (Waldst. & Kit.) is a good example of beneficial na-turalization in the Salado River Basin, that is, in the largest area dedicated to bovine farming in the country. The aim of this study was to evaluate the promotion of L. tenuis in marginal areas as a valuable alternative for early weaning systems in livestock production. Two treatments were tested during three study periods. To evaluate the efficiency of the proposed system, average daily weight gain (DWG) and weight gain (WG) of conventional weaning (CW) and early weaning (EW) calves were determined and compared. EW calves grazing on L. tenuis gained more weight than CW calves during Study period 1 (P = 0.028), but no differences were observed between treatments during Study periods 2 and 3 (P ˃ 0.05). Body Condition Score (BCS) for dams in both treatments was evaluated. For all three study periods, BCS improvement was higher in cows from the EW treatment than in Lactating cows (LC) from the CW treatment. Thus, EW improved cow BCS and did not generate weight reductions in calves thanks to the Lotus-based diet. In conclusion, L. tenuis promotion in the summer season resulted higher yields in marginal areas than semi–natural pastures. It also provided an acceptable nutritional value to properly fed a considerable amount of calves on a small surface area, and enabled a better recovery of the dams in terms of body condition. The obtained results support the design of an easy-to-use strategy that facilitates adoption by producers.
Keywords: biomass, pastures, legumes, livestock.
RESUMEN
La promoción de leguminosas forrajeras en ambientes marginales para la agricultura constituye un impor-tante desafío tecnológico para la ganadería. Lotus tenuis (Waldst. & Kit.) es un buen ejemplo de naturalización beneficiosa en la Cuenca del Río Salado, es decir, en una de las más importantes regiones dedicada a la cría de bovinos en el país. El objetivo de este estudio fue evaluar la promoción de L. tenuis como una alternativa valiosa para los sistemas de destete temprano en la producción ganadera. Se probaron dos tratamientos du-rante tres períodos de estudio. Para evaluar los beneficios del sistema propuesto, se determinó y comparó la
1Instituto Nacional de Tecnología Agropecuaria (INTA), Chacra Experimental Integrada de Chascomús (CEICh). Chascomús, Buenos Aires, Argentina. 2Instituto Tecnológico de Chascomús (INTECh). Universidad Nacional San Martín (UNSAM), Consejo Nacional de Investigaciones Científi-cas y Técnicas de Argentina, (CONICET). Chascomús, Buenos Aires, Argentina. Correo electrónico: [email protected] 3Universidad de Buenos Aires (UBA), Facultad de Ciencias Exactas y Naturales, (FCEN-UBA). CABA, Buenos Aires, Argentina.4Instituto Nacional de Tecnología Agropecuaria (INTA), Instituto de Fisiología y Recursos Genéticos Vegetales (IFRGV) (CIAP). Córdoba, Argentina.
BAILLERES, M.1; CAMPESTRE, M.P.2; ANTONELLI, C.J.2; MELANI, G.1; MENÉNDEZ, A.3; RUIZ, O.A.2,4
Received September 10th 2019// Accepted May 28th 2020 // Published online November 11th 2020

Promotion of Lotus tenuis and calf early weaning as a good management practice for breeding herds in marginal soils of the Flooding Pampa (Argentina)
ARTÍCULOS RIA / Vol. 46 / N.º 2268268
ganancia de peso diario promedio y la ganancia del peso de terneros según se utilizaran procesos al pie de la vaca o con un destete temprano. Los terneros provenientes de un destete temprano que pastaban en promo-ciones de L. tenuis ganaron más peso que los terneros provenientes de un destete convencional durante el período de estudio 1 (P = 0.028), pero no se observó diferencia entre los tratamientos durante los períodos de estudio 2 y 3 (P ˃ 0.05). Se evaluó la puntuación de condición corporal para las madres en ambos tratamien-tos. Para los tres períodos de estudio la mejora de esta condición fue mayor en las vacas del tratamiento de destete temprano que en las vacas lactantes del tratamiento convencional. Por lo tanto, el destete temprano mejoró la condición corporal de la vaca, sin generar reducciones de peso en los terneros gracias a la dieta basada en L. tenuis. En conclusión, la promoción de esta leguminosa en la temporada de verano proporcionó mayores rendimientos en áreas marginales para la agricultura que las pasturas seminaturales. Asimismo, proporcionó una dieta con un valor nutricional aceptable para la alimentación adecuada de una cantidad con-siderable de terneros en una superficie pequeña, permitiendo al mismo tiempo, una mejor recuperación de las madres en términos de condición corporal. Los resultados obtenidos respaldan el diseño de una estrategia sencilla que facilita la adopción por parte de los productores.
Palabras clave: biomasa forrajera, pasturas, leguminosas, ganado vacuno.
INTRODUCTION
Allowing weaning to occur when calves reach an age of 6–8 months is a common practice in beef cow–calf opera-tions (Pate et al., 1985; Thrift and Thrift, 2004). On range-lands, however, this practice may be hampered by a rapid decline in the quantity and quality of forage resources as a consequence of lignification, temperature increase, or drought in mid– or late–summer (Mulliniks et al., 2013).
During this period, cow–calf pairs present their highest energy requirement due to lactation and cow pregnancy. Early weaning (EW, when the calf is 4–5 months old) is acknowledged as a viable alternative to counter summer quality forage shortage (Whittier, 1995; Merrill et al., 2008; Warner et al., 2015). It has been also shown that EW de-creases nutrient requirements of beef cows, enabling an improvement in cow Body Condition Score (BCS) (Merrill et al., 2008; Odhiambo et al., 2009; Martins et al., 2012; Waterman et al., 2012) and increasing pasture carrying ca-pacity (Johnson et al., 2015).
EW can also increase conception rates and shorten the in-terval from parturition to conception in calf heifers (Laster et al., 1973; Lusby et al., 1981). Despite the advantages of this practice, EW is a source of stress for the dam and for the calf. This effect is especially acute and prolonged for the calf, which at weaning is subjected to multiple stressors such as the loss of the mother and access to the udder and milk, and changes in the social and physical environment (Enríquez et al., 2011).
In recent years, researchers have reported the use of di-verse forage species such as Cynodon dactylon (L.), Chlo-ris gayana (Kunth.), Medicago sativa (L.), Trifolium repens (L.), or Zea mays (L.) silage (Castells et al., 2013; Terré et al., 2013; Corriher et al., 2014) to compensate for milk restriction to calves during weaning. Interestingly, Lotus species are forage legumes considered to have similar or even higher nutritional value than M. sativa and T. repens (Montes, 1988; Acuña, 1998; Blumenthal and McGraw,
1999; Cassida et al., 2000). Also, these species can either be grazed by livestock or can be used as hay or silage, unlike several other forage legumes that do not offer these possibilities (Blumenthal and McGraw, 1999).
Lotus tenuis (Waldst. & Kit.) is currently being tested as an alternative to overcome summer forage deficiency in the Flooding Pampa, the most important area for calf produc-tion in Argentina (Miñón et al., 1990; Escaray et al., 2012, 2014, 2016). An important advantage of L. tenuis when compared to M. sativa or other forage legumes is its great adaptability to marginal environments, where waterlogging and saline or alkaline soils often occurs (Escaray et al., 2012, 2014, 2016; Antonelli et al., 2016; 2019).
This type of soil condition predominates in vast extensions of the Flooding Pampa, where L. tenuis is naturalized. In summer, L. tenuis is already in reproductive state, but un-like C3 grasses, it continues to produce leaves with optimal nutritional quality, thus maintaining high levels of digestibility and crude protein (CP) (Hidalgo and Cauhépé, 2009) and reaches its maximum biomass (Colabelli and Miñon, 1994).
Due to these characteristics, Lotus promotion has been considered as an alternative to overcome forage deficiency during the summer season (Antonelli et al., 2016). However, L. tenuis shows low competitiveness during the first stages of implantation (Clua et al., 1997). This drawback can be re-verted by the application of low doses of herbicides during winter, a practice known as L. tenuis promotion. This prac-tice increases light and other resources for its implantation and leads to the dominance of this naturalized legume (An-tonelli et al., 2016; Nieva et al., 2016, 2018, 2019). This practice is currently carried out in small pasture areas so far considered marginal areas for agricultural activities, due to their low productive capacity as a result of their low fertility, recurrence of flooding and salt accumulation.
Research on cow–calf and weaning systems has been well documented in Australia (Tyler, 2012; Tyler et al., 2012),

BAILLERES, M.; CAMPESTRE, M.P.; ANTONELLI, C.J.; MELANI, G.; MENÉNDEZ, A.; RUIZ, O.A.
Agosto 2020, Argentina 269269
USA (Arthington et al., 2005; Vendramini et al., 2006), the neotropics (Moore and da Rocha, 1983; Betancourt–López et al., 2012), and Argentina (Pordomingo, 2002). However, none of these studies considered calves raised on a Lotus–based pastures. This study aimed at evaluating the effect on 1) Average daily weight gain (DWG) and weight gain (WG) of EW calves grazing on promoted L. tenuis paddocks com-pared to unweaned (UW) calves from conventional weaning (CW) treatment grazing on semi–natural pasture, and 2) the recovery of the cows from the EW treatment compared to lactating cows (LC) from the CW treatment in terms of BCS. The L. tenuis promotion increases the forage supply in con-strained areas characterized by its low nutritional value and legumes presence. This last situation could limit the perfor-mance of cows and calves (Minson, 1990). In this study, we inquire into whether promoted L. tenuis meets the biomass quantity and quality required for acceptable calf performance.
MATERIALS AND METHODS
Site and Pasture description
The study was conducted at the Chacra Experimental In-tegrada de Chascomús (CEICh–Ministerio de Agroindustrias de la Provincia de Buenos Aires–Instituto Nacional de Tec-nología Agropecuaria, Argentina, 35º45’27”S, 58°3’18”W), which is located in the Flooding Pampa region, during the pe-riods 2010–2011, 2011–2012 and 2012–2013 (Study periods 1, 2 and 3, respectively). This region has a temperate sub–humid climate with mean temperatures averaging 8.5 °C in winter and 21.5 °C in summer, and annual rainfall 850–1050 mm. Short floods of 5–7 cm depth occur at the beginning of almost every spring. Nonetheless, severe droughts may occur in early summer. Soils in the experimental area are Natraquoll type, characterized by an A1 horizon with 3.5% of organic matter and 0.22 mg kg–1 of extractable Fe, and by a natric B2t horizon at a depth of 17 cm, with 53.3% clay con-tent (Lavado and Taboada, 1988). Plant communities consist mainly of grasses and Compositae species. Native legumes are largely absent and semi–natural pastures are dominated by Festuca arundinacea, Thinopyrum ponticum, Cynodon dactylon, L. tenuis, and Sporobolus indicus.
Promoted L. tenuis pasture
Glyphosate (N–(phosphonomethyl) glycine; 3.5 l ha–1) was applied on a 2 hectares surface area, followed by two applications of 2,4 DB(4–(2,4–dichlorophenoxy) butyric acid, 1 l ha–1) and a single dose of Quizalofop–p–ethyl (Eth-yl(R) –2– [4– (6– chloro–2–quinoxalyloxy) phenoxy] propi-onate; 1.2 l ha–1), in six or seven annual cycles from June to August. After 4 or 5 years of herbicide application, plant species composition in promoted paddocks shifted, and L. tenuis became the dominant species (Nieva et al., 2016; 2018; 2019; Druille et al., 2017).
Weaning treatments
For comparative purposes, both groups of animals were simultaneously incorporated to the experimental lots: a)
use of herbicides to increase the presence and persistence of L. tenuis and b) without the use of herbicides (semi-nat-ural pasture). Regarding the load, 2 ha were assigned to the EW calves in the Lotus promoted paddocks. The dams from EW animal lots and cows-calf pairs from CW manage-ment (one animal per ha), grazed in the semi-natural pas-ture during 4-5 days, in parcels with availability of 1500 to 2000 kg DM/ha and were removed from parcels with rem-nants forage between 800 and 1200 kg DM/ha. This animal management protocol was optimized years previous to the EW and CW evaluation.
Two treatments were proposed: (1) CW: cow–calf pairs grazing on semi–natural pasture (LC n = 20; calves n = 20), and (2) EW: calves weaned at the age of 4–5 months, graz-ing on promoted L. tenuis paddocks (calves n = 20) and dams grazing on semi–natural pasture (n = 20).
Measurements were performed in the three growth peri-ods mentioned above: Study period 1, Study period 2, and Study period 3 from February to March according to for-age availability. All animals used in this research reached maturity and were included in the CEICh usual livestock productivity cycle.
Animal measurement
In each study period and treatment, Aberdeen Angus calves and dams were used to evaluate the Weight Gain (WG), average Daily Weight Gain (DWG), and Body Condi-tion Score (BCS). At the beginning of each study period and treatment, calf weight averaged 160 kg (initial weight). All calves from both treatments were reweighed (final weight) 25 days after the first weight register during the Study peri-od 1, 34 days later during the Study period 2 and 60 days later during Study period 3. This difference in the test time of each period was due to climatic or technical conditions. DWG was then calculated as [final weight–initial weight]/X days of the study period.
Initial BCS was determined for all LC and EW dams at the beginning of treatments. BCS refers to the relative amount of subcutaneous body fat or energy reserve in the cow. It was estimated by visual appraisal and scores were as-signed from 1 to 5, where 1 = a very thin cow and 5 = an ex-cessively fat one. At the end of each experiment, BCS de-termination was repeated on the same cows. BCS change in the period was calculated as final BCS–initial BCS.
Pasture measurement
Plant biomass was harvested from ten 0.25 m2 quadrats. Plant samples were collected six times during the growing season (October to March) for all three study periods, by clipping approximately 1 cm above the soil surface. Sam-ples were dried at 70 °C to constant weight, and dry bio-mass was calculated per hectare.
For forage quality determinations, L. tenuis plants were clipped in December, January, and February, with similar results. The biomass quantity and quality required for ac-
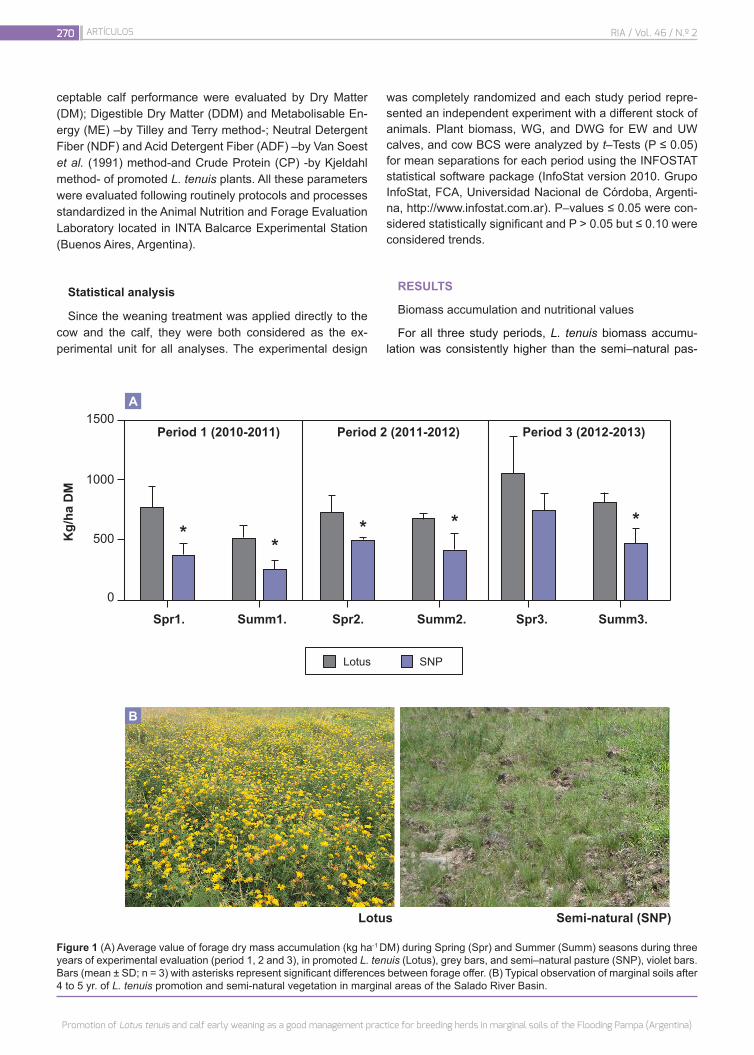
Promotion of Lotus tenuis and calf early weaning as a good management practice for breeding herds in marginal soils of the Flooding Pampa (Argentina)
ARTÍCULOS RIA / Vol. 46 / N.º 2270270
Figure 1 (A) Average value of forage dry mass accumulation (kg ha-1 DM) during Spring (Spr) and Summer (Summ) seasons during three years of experimental evaluation (period 1, 2 and 3), in promoted L. tenuis (Lotus), grey bars, and semi–natural pasture (SNP), violet bars. Bars (mean ± SD; n = 3) with asterisks represent significant differences between forage offer. (B) Typical observation of marginal soils after 4 to 5 yr. of L. tenuis promotion and semi-natural vegetation in marginal areas of the Salado River Basin.
ceptable calf performance were evaluated by Dry Matter (DM); Digestible Dry Matter (DDM) and Metabolisable En-ergy (ME) –by Tilley and Terry method-; Neutral Detergent Fiber (NDF) and Acid Detergent Fiber (ADF) –by Van Soest et al. (1991) method-and Crude Protein (CP) -by Kjeldahl method- of promoted L. tenuis plants. All these parameters were evaluated following routinely protocols and processes standardized in the Animal Nutrition and Forage Evaluation Laboratory located in INTA Balcarce Experimental Station (Buenos Aires, Argentina).
Statistical analysis
Since the weaning treatment was applied directly to the cow and the calf, they were both considered as the ex-perimental unit for all analyses. The experimental design
was completely randomized and each study period repre-sented an independent experiment with a different stock of animals. Plant biomass, WG, and DWG for EW and UW calves, and cow BCS were analyzed by t–Tests (P ≤ 0.05) for mean separations for each period using the INFOSTAT statistical software package (InfoStat version 2010. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argenti-na, http://www.infostat.com.ar). P–values ≤ 0.05 were con-sidered statistically significant and P ˃ 0.05 but ≤ 0.10 were considered trends.
RESULTS
Biomass accumulation and nutritional values
For all three study periods, L. tenuis biomass accumu-lation was consistently higher than the semi–natural pas-
1500
1000
500
0
Period 1 (2010-2011)
Lotus Semi-natural (SNP)
Spr1. Summ1. Spr2. Summ2. Spr3. Summ3.
Kg/
ha D
M
Period 2 (2011-2012) Period 3 (2012-2013)
Lotus SNP
* ** * *
A
B
BAILLERES, M.; CAMPESTRE, M.P.; ANTONELLI, C.J.; MELANI, G.; MENÉNDEZ, A.; RUIZ, O.A.
Agosto 2020, Argentina 271271
ture biomass, with an average value per growth period of 4525 kg ha–1 DM and 2824 kg ha–1 DM, respectively (Figure not shown). Moreover, as shown in figure 1.A, during the summer season (January–February–March), biomass was lower in the semi–natural pasture, while L. tenuis biomass
Nutritional quality Mean † ± SD ‡DM 245.3 ± 20.7
CP 203.3 ± 14.5
DDM 755.7 ± 32.0
ME 2.73 ± 1.2
NDF 299.0 ± 31.1
ADF 252.3 ± 30.8
Table 1. Forage nutritional quality of promoted L. tenuis evaluated as dry matter (DM, g kg–1), digestible dry matter (DDM, g kg–1 DM), metabolizable energy (ME, Mcal kg–1 DM), neutral detergent fiber (NDF, g kg–1 DM), acid detergent fiber (ADF, g kg–1 DM), and crude protein (CP, g kg–1 DM). † Values are averages from samples taken at three different times, from December to February. ‡ Standard deviation.
Table 2. Influence of weaning treatments and grazing source on calf performance expressed as (A) Daily weight gain (kg) and (B) Weight (kg).† Conventional–weaned treatment consisted of cow–calf pairs (n = 20 calves) grazing on semi–natural pastures while early–weaned treatment consisted of 4–5–month–old calves (n = 20) grazing on 2 hectares of promoted L. tenuis paddocks. ‡ Study period 1: (2010–2011) lasted 25 days, 2: (2011–2012) lasted 34 days and 3: (2012–2013) lasted 60 days. §Standard deviation.
was slightly higher, and even more so compared to semi–natural pasture in the grazing months of EW calves (Feb-ruary–March). Figure 1.B shows the typical coverage that each pasture reached in the summer season.
In addition to biomass accumulation, table 1 shows the values of forage nutritional quality in promoted L. tenuis plants during the summer season.
Influence of weaning treatment and promoted L. tenuis pasture on calf performance
Calf DWG was influenced by weaning treatment (table 2 A). DWG was higher during Study period 2 (2011–2012) (DWG = 0.65 kg day–1P ≤ 0.05) and presented a trend in favor of EW and grazing on promoted L. tenuis during Study period 1 (2010–2011) (DWG = 1.21 kg day-1 P≤ 0.10). How-ever, during Study Period 3 (2012–2013), there was a differ-ence in favor of UW calves grazing on semi–natural pastures (DWG = 0.70 kg day-1 P ≤ 0.05). Regarding WG, it is import-ant to note that all calves in both treatments weighed approx-imately the same (~160 kg) at the beginning of each study period (table 2 B). No WG differences were found between weaning treatments during Study period 2 and 3. However, there were WG differences during period 1 in favor of EW and grazing on promoted L. tenuis during 25 days.
Period ‡ Item Conventional-weaned† Sd § Early-weaned† Sd § P- value
A Daily weight gain (kg)
1 Final period 1.106 0.29 1.21 0.22 0.10
2 Final period 0.35 0.19 0.65 0.09 0.0025
3 Final period 0.708 0.315 0.35 0.083 0.0023
B Weight (kg)
1 Initial 160 6.07 164.1 12.47 0.155
Final 187.65 9.82 194.45 13.63 0.028
Period change 27.65 30.35
2 Initial 162.1 9.9 163.8 5.93 0.392
Final 174.1 11.76 186 3.81 0.26
Period change 12 22.2
3 Initial 154.23 14.14 156.63 17.48 0.288
Final 196.77 25.18 177.94 20.72 0.15
Period change 42.54 21.31
Promotion of Lotus tenuis and calf early weaning as a good management practice for breeding herds in marginal soils of the Flooding Pampa (Argentina)
ARTÍCULOS RIA / Vol. 46 / N.º 2272272
Influence of weaning treatment on cow performance
At the end of each assay, cow BCS (table 3) improved more in EW cows than in LC for Study Periods 1 and 3 (P< 0.05) but no differences were recorded for Study Period 2. It should be noted that in Study Period 2 EW cows started with a lower BCS (2.625) than LC (2.917). However, by the end of the 34–day treatment, BCS gain was 0.275 for EW cows but –0.084 for LC cows. At the same time, in all the three stud-ied periods, the period change value was greater for the EW cows compared to the period change value for the CW cows.
DISCUSSION
The grasslands of the Flooding Pampa are subject to the combined effects of grazing, floods, and droughts. Thus, a sustainable solution is needed for forage production en-hancement in marginal environments. L. tenuis promotion is an appealing alternative for cattle production in con-strained environments in this region. Our results show that L. tenuis promotion produced higher forage yields than the supply provided by the semi–natural pasture under these soil conditions (figure 1A).
L. tenuis not only contributes with quantity but also nutrition-al quality (table 1). L. tenuis values are comparable to those of other two Leguminosae: L. corniculatus (L). and T. repens (L). (Peiretti et al., 2016) with global commercial importance (Phelan et al., 2015) rich in high-quality protein content and highly digestible (Kaplan et al., 2009; Graves et al., 2012).
This study shows a clear improvement in BCS for EW cows. Similar results have been reported in other weaning studies conducted on pasture (Merrill et al., 2008; Martins et al., 2012; Johnson et al., 2015). The lower BCS of LC compared to EW
cows may be explained mainly by the continued demand for milk by UW calves (Johnson et al., 2015). Therefore, since UW calves are frequently maintained as cow–calf pair for approx-imately a month and a half longer than the period analyzed in this study, further BCS losses in LC would be expected. As reported by Whittier (1995), the long–term effect of lactation may delay pregnancy, so earlier weaning would improve BCS and increase pregnancy rate (Thrift and Thrift, 2004).
During Study Period 1, DWG did not differ between treat-ments and was higher than in Study Periods 2 and 3. In turn, differences were detected between treatments in the latter two periods, with higher DWG in EW than UW calves for Study Period 2, while the opposite occurred for Study Period 3. Such discrepancy could be given by the interval between days that the calves were found grazing Lotus in each study period. Hence, it would not be advisable to keep the calves grazing promoted L. tenuis for more than 34 days, probably due to the trampling effect generated by the calves on the pasture, unless the stocking density per hectare will be re-duced. However, to draw better conclusions further research analyzing additional sources of variation is required.
Weatherly (2008) reports that EW calves aged 3–6 months require 160 g kg–1 DM of CP for a 0.70 kg day–1 growth rate. Former information and our results showing CP values of 203.3 g kg–1 DM and DM of 245.3 g kg–1 for promoted L. tenuis suggest that it is highly suitable as for-age. In our study, this is reflected by the WG observed for animals grown on pastures promoted with predominance of L. tenuis during 25 days. The promotion of small areas of L. tenuis thus seems to be an important strategy enabling higher yields, taking into account that our results suggest that only 1 hectare is required to fully feed ten 4–5–month–old calves for a short period (table 2).
Table 3. Influence of weaning treatments on cow performance expressed as Body Condition Score (BCS). † Study period 1: (2010–2011) 25 days, 2: (2011–2012) 34 days and 3: (2012–2013) 60 days. ‡INITIAL BCS was measured in all cows at the beginning of both treatments (Conventional and Early weaning). § FINAL BCS was measured in all cows at the end of each Study period time. ¶ Standard deviation.
Period† Item Conventional-weaned Sd¶ Early-weaned Sd¶ P- value
BCS
1 Initial‡ 3.005 0.17 2.968 0.249 0.71
Final§ 3.085 0.12 3.421 0.334 0.0005
Period change 0.08 0.453
2 Initial‡ 2.917 0.25 2.625 0.294 0.056
Final§ 2.833 0.25 2.9 0.241 0.37
Period change -0.084 0.275
3 Initial‡ 2.984 0.146 2.976 0.251 0.88
Final§ 3.083 0.14 3.433 0.323 0.023
Period change 0.099 0.457

BAILLERES, M.; CAMPESTRE, M.P.; ANTONELLI, C.J.; MELANI, G.; MENÉNDEZ, A.; RUIZ, O.A.
Agosto 2020, Argentina 273273
ACUÑA, H.P. 1998. Varietal comparison of three species of genus Lotus (L. corniculatus L., L. uliginosus Cav. and L. tenuis Wald et Kit.) in soils with aptitude for cropping rice. Agric Tec Chile 58: 7-14.
ANTONELLI, C.J.; CALZADILLA, P.I.; ESCARAY, F.J.; BA-BUIN, M.F.; CAMPESTRE, M.P.; ROCCO, R.; BORDENAVE, C.D.; PEREA GARCÍA, A.; NIEVA, A.S.; LLAMES, M.E.; MA-GUIRE, V.; MELANI, G.; SARENA, D., BAILLERES, M.; CA-RRASCO, P.; PAOLOCCI, F.; GARRIZ, A.; MENÉNDEZ, A.; RUIZ 2016. Lotus spp: Biotechnological strategies to improve the bioeconomy of lowlands in te Salado River Basin (Argentina). AGROFOR International Journal, 1.
ANTONELLI, C.J.; CALZADILLA, P.I.; VILAS, J.M.; CAMPES-TRE, M.P.; ESCARAY, F.J.; RUIZ, O.A. 2019. Physiological and anatomical traits associated with tolerance to long-term partial sub-mergence stress in the Lotus genus: responses of forage species, a model and an interspecific hybrid. J. Agron Crop Sci, 205(1), 65-76.
ARTHINGTON, J.D.; SPEARS, J.W.; MILLER, D.C. 2005. The effect of early weaning on feedlot performance and measures of stress in beef calves. J Anim Sci 83: 933-939.
BETANCOURT–LÓPEZ, L.; PAREJA–MEJÍA, R.I.; CONDE–PULGARÍN, A.; CASTELLANOS, A.F.; MORENO–MARTÍNEZ, D. 2012. Manejo nutricional de terneros cebú comercial sometidos a amamantamiento restringido y destete precoz en el piedemonte de Casanare. Rev Cie Anim 5: 21-30.
BLUMENTHAL, M.J.; MCGRAW, R.L. 1999. Lotus adaptation, use and management. In: BEUSELINCK, P.R. (Ed.). Trefoil: The Science and Technology of Lotus, CSSA. 28: 97-119.
CASSIDA, K.A.; GRIFFIN, T.S.; RODRIGUEZ, J.; PATCHING, S.C.; HESTERMAN, O.B.; RUST, S.R. 2000. Protein degradability and forage quality in maturing alfalfa red clover, and birdsfoot tre-foil. Crop Sci 40: 209-215.
CASTELLS, L.; BACH, A.; ARIS, A.; TERRÉ, M. 2013. Effects of forage provision to young calves on rumen fermentation and development of the gastrointestinal tract. J DairySci96: 5226-5236.
CLUA, A.A.; GIMENEZ, D.O.; FERNANDEZ, L.V. 1997. Increase in forage yield in narrow leaf birdsfoot trefoil (Lotus tenuis Waldst & Kit ex Willd) in a permanent pasture with foliar applied gibberellic acid (GA3), and phosphorus. Plantgrowthregul 21: 223-228.
COLABELLI, M.; MIÑON, D. 1994. Rendimiento y cambios bo-tánicos de pasturas de Lotus tenuis puro y en mezcla bajo régimen de corte. AgriculturaTécnica, Chile 54: 39-45.
CORRIHER, A.; HILL, G.M.; ANDRAE, J.G.; FROETSCHEL, M.A.; MULLINIKS, B.G. 2014. Cow and calf performance on Coas-tal or Tifton 85 Bermudagrass pastures with aeschynomene creep-grazing paddocks. J Anim Sci 85: 2762-2771.
DRUILLE, M.; ACOSTA, A.; ACOSTA, G.; ROSSI, J.; GOLLUS-CIO, R.A.; BAILLERES, M. 2017. Response of beneficial soil fungi as-sociated with Lotus tenuis to the application of glyphosate. RIA 43: 3.
ENRÍQUEZ, D.; HÖTZEL, M.; UNGERFELD, R. 2011. Minimising the stress of weaning of beef calves: a review. Acta Vet Scand 53: 28.
ESCARAY, F.J.; MENENDEZ, A.B.; GÁRRIZ, A.; PIECKENS-TAIN, F.L.; ESTRELLA, M.J.; CASTAGNO, L.N.; CARRASCO, P.; SAN JUAN PINILLA, J.; RUIZ, O.A. 2012. Ecological and agronomic importance of the plant genus Lotus. Its application in grassland sustainability and the amelioration of constrained and contaminated soils. Plant Science, 182, 121-133.
ESCARAY, F.J.; PASSERI, V.; BABUIN, F.M.; MARCO, F.; CARRASCO, P.; DAMIANI, F., PIECKENSTAIN, F.L.; PAOLOC-CI, F.; RUIZ, O.A. 2014. Lotus tenuis x L. corniculatus interspecific hybridization as a means to breed bloat-safe pastures and gain insight into the genetic control of proanthocyanidin biosynthesis in legumes. BMC plant biology, 14(1), 40.
It is important to highlight that the promotion of L. tenuis would be one more link in the forage chain in a cow breed-ing enterprise, which would determine an advantage in a given period, which maybe be accompanied by other re-sources such as natural grassland or ryegrass promotions.
CONCLUSIONS
The technology of L. tenuis promotion that was designed and assayed by the Chacra Experimental Integrada de Chas-comús produced higher yields than those provided by semi–natural pastures. Also, L. tenuis has an acceptable nutritional value for adequately feeding a considerable number of calves on a small surface. L. tenuis has a high capacity for natural reseeding and can withstand the water deficit that often oc-curs from late spring through summer (Vignolio et al., 2011; Escaray et al., 2012). Forage supply is especially important in summer when breeding herd requirements are usually at their maximum. Ideally, this demand should be met by pasture quantity and quality and/or supplementary feed. Another point to consider is that in grasses and legumes the digestion rate is influenced by the increase in the lignin content at the be-ginning of the flowering period (Mahyuddin, 2008). L. tenuis, however, continues vegetating even after the flowering peri-od, with a wide overlap between vegetative and reproductive growth, as has been previously described for indeterminate growth species (Vignolio et al., 2016). Thus, EW of calves grazing on this forage is a useful management practice that reduces grazing pressure on semi–natural pastures, decreas-ing cow nutrient requirements. BCS improvement in EW cows could shorten the postpartum interval and improve pregnancy rate. This management practice can be potentially transferred to producers. Likewise, its adoption will allow us to increase the accuracy of the obtained results and its adaptation to other environments. It is also important to highlight the production of higher–quality meat, identifiable by its levels of unsaturated fatty acids, from animals fed on promoted L. tenuis compared to meat from animals fed in intensive fattening systems based on grains and balanced pelleted foods (Acosta et al., 2016). Therefore, we consider that this cow management practice could effectively improve the productivity in Argentina’s most important livestock breeding region.
ACKNOWLEDGMENTS
This work was financed by the following grants: PICT of Agencia Nacional de Promoción Científica y Tecnológica (ANPCYT, Argentina); Universidad Nacional de San Martin (UNSAM, Argentina) and Consejo Nacional de Investiga-ciones Científicas y Técnicas (CONICET, Argentina). We also appreciate the excellent technical assistance of the CEICh professional farm staff.
REFERENCESACOSTA, A.; ROSSI, J.L.; ACOSTA, G.; SCHOR, A.; GOLLUS-
CIO, R. 2016. Productive strategy of steers in a marginal area of the Pampean Region.vi Workshop of Lotus spp. Chascomús, Bue-nos Aires (Argentina).

Promotion of Lotus tenuis and calf early weaning as a good management practice for breeding herds in marginal soils of the Flooding Pampa (Argentina)
ARTÍCULOS RIA / Vol. 46 / N.º 2274274
GRAVES, M.E.; MCLEAN, N.; JONES, G.; MARTIN, R.C. 2012. Pasture and sheep performance response to sod-seeding red clover (Trifoliun pretense L.) or white clover (Trifolium repens L.) into natura-lized pastures in eastern Canada. Anim. Feed Sci. Technol. 177: 7-14.
HIDALGO, L.G.; CAUHÉPÉ, M.A. 2009. Argentine rangeland quality influences reproduction of yearling pregnant heifers? Grassl Sci 55: 74–78.
JOHNSON, P.S.; OLSON, K.C.;GATES, R.N.; PATTERSON, H.H.; HUBERT, M.; LANDBLOM, D.; KINCHELOE, J.J.; RICHTER, H.A.; GROVE, A. V. 2015. Early weaning reduces rangeland herbage disappearance. Crop, Forage and Turfgrass Manage. doi:10.2134/cftm2014.0090.
KAPLAN, M; ATALAY, A. I; MEDJEKAL, S. 2009.Potential nutri-tive value of wild birdsfoot trefoil (Lotus corniculatus) plants grown in different sites. Livest Res Rural Dev 21: 99.
LASTER, D.B.; GLIMP, H.A.; GREGORY, K.E. 1973.Effects of early weaning on postpartum reproduction of cows. J. Anim. Sci. 36:734.
LAVADO, R.S.; TABOADA, M.A. 1988. Water, salts and sodium dynamics in a Natraquoll in Argentina. Catena 15: 577-594.
LUSBY, K.S.; WETTEMANN, R.P.; TURMAN, E.J. 1981. Effects of early weaning calves from first-calf heifers on calf and heifer performance. J. AnimSci 53: 1193-1197.
MAHYUDDIN, P. 2008. Relationship between Chemical Compo-nent and In Vitro Digestibility of Tropical Grasses. Hayati J Biosci 15: 85-89.
MARTINS, P.G.M.A.; ARTHINGTON, J.D.; COOKE, R.F.; LAMB, C.G.; ARAÚJO, D.B.; TORRES, C.A.A.; GUIMARĀES, J.D.; MAN-CIO, A.B.. 2012. Evaluation of beef cow and calf separation sys-tems to improve reproductive performance of first-calf cows. Li-vestSci 150: 74-79.
MERRILL, M.L.; BOHNERT, D.W.; GANSKOPP, D.C.; JO-HNSON, D.D; FALCK, S.J. 2008. Effects of early weaning on cow performance, grazing behavior, and winter feed costs in the Inter-mountain West. Prof. Anim. Sci. 24: 29-34.
MINSON, D.J. 1990. Forage in Ruminant Nutrition. Academic Press, San Diego 483.
MIÑON, D.; SEVILLA, H.; MONTES, L.; FERNÁNDEZ, O.N. 1990. Lotus tenuis: leguminosa forrajera para la Pampa Deprimi-da. EEA Balcarce, INTA, Argentina. Boletín Técnico. 98: 15.
MONTES, L.1988. Lotus tenuis, Revista Argentina de Produc-ción Animal 8: 367-376.
MOORE, C.P.; CAMPOS DA ROCHA, C.1983. Reproductive performance of Gyr cows: the effect of weaning age of calves and postpartum energy intake. J. Anim. Sci. 57: 807-814.
MULLINIKS, J.T.; WATERMAN, R.C.; GEARY, T.W. 2013. Eco-nomics of early weaning in northern great plains beef cattle pro-duction system. Agric. Sci. 4: 219-233.
NIEVA, A.S.; BAILLERES, M.A.; CORRIALE, M.J.; LLAMES, M.E.; MENÉNDEZ, A.B.; RUIZ, O.A. 2016. Herbicide-mediated promotion of Lotus tenuis (Waldst. & Kit. ex Wild.) did not influence soil bacterial communities, in soils of the Flooding Pampa, Argenti-na. Applied soil ecology, 98, 83-91.
NIEVA, A.S.; BAILLERES, M.A.; LLAMES, M.E.; TABOADA, M.A.; RUIZ, O.A.; MENÉNDEZ, A. 2018. Promotion of Lotus te-nuis in the Flooding Pampa (Argentina) increases the soil fungal diversity. Fungal Ecology, 33, 80-91.
NIEVA, A.S.; VILAS, J.M.; GÁRRIZ, A.; MAIALE, S.J.; ME-NÉNDEZ, A.B.; ERBAN, A.; KOPKA, J.; RUIZ, O.A. 2019. The fungal endophyte Fusarium solani provokes differential effects on the fitness of two Lotus species. Plant Physiology and Biochemis-try, 144, 100-109.
ODHIAMBO, J.F.; RHINEHART, J.D.; HELMONDOLLAR, R.; PRITCHARD, J.Y.; OSBONE, P.I.; FELTON, E.E.; DAILEY, R.A. 2009. Effect of weaning regimen on energy profiles and reproducti-ve performance of beef cows. J. Anim. Sci. 87: 2428-2436.
PATE, F.M.; CROCKETT, J.R.; PHILLIPS, J.D.1985. Effect of calf weaning age and cow supplementation on cow productivity. J. Anim. Sci. 61: 343-348.
PEIRETTI, P.G.; GAI, F.; ALONZI, S.; TASSONE, S. 2016. Nu-tritive value and fatty acid profile of birdsfoot trefoil (Lotus cornicu-latus) and white clover (Trifolium repens) in Alpine pastures. Livest Res Rural Dev 28: 218.
PHELAN, P.; MOLONEY, A.P.; MCGEOUGH, E.J.; HUM-PHREYS, J.; BERTILSSON, J.; O´RIORDAN, E.G.; O´KIELY, P.. 2015. Forage legumes for grazing and conserving in ruminant pro-duction systems. Crit. Rev. Plant. Sci. 34: 281-326.
PORDOMINGO, A.J. 2002. Early weaning and length of supple-mentation effects on beef calves. J. Range Manage 55: 327-335.
TERRÉ, M.; PEDRALS, E.; DALMAU, A.; BACH, A. 2013. What do preweaned and weaned calves need in the diet: A high fiber content or a forage source? J. Dairy Sci. 96: 5217-5225.
THRIFT, F.A.; THRIFT, T.A. 2004. Review: Ramifications of wea-ning spring and fall born calves early or late relative to weaning at conventional ages. Prof. Anim. Sci. 20: 490-502.
TILLEY, J.M.; TERRY, R.A.1963. A two‐stage technique for the in vitro digestion of forage crops. Grass forage sci. 18: 104-111.
TYLER, R. 2012. Weaning management of beef calves - prac-tical guidelines for northern Australian beef producers. Meat and Livestock Australia Ltd, North Sydney, NSW, Australia.
TYLER, R.; ENGLISH, B.; SULLIVAN, M.; JACKSON, D.; MATTHEWS, R.; HOLMES, B.; MACDONALD, N.; OXLEY, T.; LEI-GO, S.;SMITH, P. 2012. Weaner management in northern beef herds. Meat and Livestock Australia Ltd, North Sydney, NSW, Australia.
VAN SOEST, P.J.; ROBERTSON, J.B.; LEWIS, B.A. 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysacchari-des in relation to animal nutrition. J. Dairy Sci. 74: 3583-3597.
VENDRAMINI, J.M.B.; SOLLENBERGER, L.E.; DUBEUX, J.C.B.; INTERRANTE, S.M.; STEWART, R.L.; ARTHINGTON, J.D. 2006. Concentrate supplementation effects on forage characte-ristics and performance of early–weaned calves grazing rye–rye-grass pastures. Crop Sci 46: 1595-1600.
VIGNOLIO, O.R.; CAMBARERI, G.S.; MACEIRA, N.O. 2011. Seed production of Lotus tenuis (Fabaceae), a forage legume: effects of row spacing, seeding date, and plant defoliation. Crop Pasture Sci 61: 1027-1035.
VIGNOLIO, O.R.; CAMBARERI, G.S.; PETIGROSSO, L.R.; MURILLO, N.; MACEIRA, N.O. 2016. Reproductive development of Lotus tenuis (Fabaceae) crop defoliated at different times and intensities. Am J Plant Sci 7: 1180.
WARNER, J.M.; JENKINS, K.H.; RASBY, R.J.; LUEBBE, M.K.; ERICKSON, G.E.; KLOPFENSTEIN, T.J. 2015. The effect of calf age at weaning on cow and calf performance and feed utilization by cow–calf pairs. Prof Anim Sci 31: 455-461.
WATERMAN, R.C.; GEARY, T.W.; PATERSON, J.A.; LIPSEY, R.J. 2012. Early weaning in Northern Great Plains beef cattle pro-duction systems: i. Performance and reproductive response in ran-ge beef cows. Livest Sci 148: 26-35.
WEATHERLY, E. 2008. Early Weaning of Beef Calves. AG1282 ISSN 1329-8062. (Available at: http://agriculture.vic.gov.au/agri-culture/livestock/beef/handling-and-management/early-weaning-of-beef-calves/ verified: September 08th 2017).
WHITTIER, J.C. 1995. Time of weaning and cow condition. In Range Beef Cow Symposium. 182.






























