Embed Size (px)
Citation preview

R
GJPPTa
b
c
d
e
f
a
ARRAA
KDELAF
1
msidwai
tD
Hf
(
h0
Virus Research 188 (2014) 15–26
Contents lists available at ScienceDirect
Virus Research
j ourna l h o mepa ge: www.elsev ier .com/ locate /v i rusres
ole of ERK1/2 signaling in dengue virus-induced liver injury
opinathan Pillai Sreekantha, Aporn Chuncharuneeb, Aunchalee Sirimontapornb,utatip Panaamponb, Chatchawan Srisawata, Atthapan Morchangc, Shilu Malakarc,eti Thuwajit c, Suwattanee Kooptiwutd, Aroonroong Suttitheptumronge,ucharee Songprakhone, Sansanee Noisakranf, Pa-thai Yenchitsomanuse,hawornchai Limjindapornb,e,∗
Graduate Program in Biochemistry, Department of Biochemistry, Faculty of Medicine Siriraj Hospital, Mahidol University, Bangkok, ThailandDepartment of Anatomy, Faculty of Medicine Siriraj Hospital, Mahidol University, Bangkok, ThailandGraduate Program in Immunology, Department of Immunology, Faculty of Medicine Siriraj Hospital, Mahidol University, Bangkok, ThailandDepartment of Physiology, Faculty of Medicine Siriraj Hospital, Mahidol University, Bangkok, ThailandDivision of Molecular Medicine, Department of Research and Development, Faculty of Medicine Siriraj Hospital, Mahidol University, Bangkok, ThailandMedical Biotechnology Unit, National Center for Genetic Engineering and Biotechnology, National Science and Technology Development Agency, Thailand
r t i c l e i n f o
rticle history:eceived 19 December 2013eceived in revised form 24 March 2014ccepted 24 March 2014vailable online 1 April 2014
eywords:engue virusRK1/2
a b s t r a c t
The liver is considered to be an important organ of dengue virus (DENV) replication and pathogenesis.However, molecular mechanisms of hepatic injury are still poorly understood. Modulation of MitogenActivated Protein Kinases (MAPKs) was previously shown to affect DENV-induced apoptosis of hepa-tocytes in vitro. However, the in vivo role of ERK1/2, a member of the MAPK family, and the questionwhether its activation can facilitate cell survival or cell death, has not been thoroughly investigated.Therefore, the role of ERK1/2 in a mouse model of DENV infection was examined. Our results show thatDENV induces phosphorylation of ERK1/2 and increases apoptosis. Inhibition of phosphorylated ERK1/2by the selective ERK1/2 inhibitor, FR180204, limits hepatocyte apoptosis and reduces DENV-induced
iver injurypoptosisR180204
liver injury. Clinical parameters, including leucopenia, thrombocytopenia, transaminases and histology,show improvements after FR180204 treatment. The expression of cell death genes was further identifiedusing real-time PCR array and Western blot analysis. Caspase-3 was significantly decreased in FR180204treated DENV-infected mice compared to the levels of untreated DENV-infected mice suggesting the roleof ERK1/2 signaling in immune-mediated liver injury during DENV infection.
. Introduction
Dengue virus (DENV) infection is one of the most importantosquito-borne viral diseases, which is endemic in tropical and
ub-tropical regions. Clinical manifestations of DENV infectionnclude dengue fever (DF), dengue hemorrhagic fever (DHF), andengue shock syndrome (DSS). Patients with DHF generally presentith hemorrhagic tendencies, plasma leakage, thrombocytopenia,
nd hemoconcentration. DSS may occur in cases of subsequentnfection with a different serotype of DENV (Halstead, 2007).
Hepatic dysfunction is a crucial feature seen in DENV infec-ion (Halstead, 2007). Transaminase levels are increased inENV-infected patients (Halstead, 1988; Kuo et al., 1992; Souza
∗ Corresponding author at: Department of Anatomy, Faculty of Medicine Sirirajospital, Mahidol University, Bangkok, Thailand. Tel.: +66 2419 7035;
ax: +66 2419 7035.E-mail addresses: [email protected], [email protected]
T. Limjindaporn).
ttp://dx.doi.org/10.1016/j.virusres.2014.03.025168-1702/© 2014 Elsevier B.V. All rights reserved.
© 2014 Elsevier B.V. All rights reserved.
et al., 2004) and in a murine model of DENV infection (Barth et al.,2006; Chen et al., 2004; Franca et al., 2010; Paes et al., 2005, 2009).Apoptosis of hepatic cells, which may be related to the pathogen-esis of DSS, has been observed both in vitro and in vivo (Couvelardet al., 1999; El-Bacha et al., 2007; Huerre et al., 2001; Limontaet al., 2007; Lin et al., 2008; Marianneau et al., 1998; Morchanget al., 2011; Nagila et al., 2011, 2013; Netsawang et al., 2010;Thongtan et al., 2004). DENV infection promotes apoptosis in thehepatoma cell line, HepG2, partly through the induction of TRAIL, amember of the death receptor pathway (Matsuda et al., 2005). TNF-� and Fas signaling also contribute to DENV-mediated apoptosis(Limjindaporn et al., 2007; Nagila et al., 2013). The mitochondriaof DENV-infected HepG2 cells exhibit functional and morpholog-ical defects, suggesting activation of the mitochondrial cell deathpathway (El-Bacha et al., 2007). Similarly, DENV infection of Huh-
7 cells, another hepatoma cell line, alters mitochondrial functionand expression of p53 (Nasirudeen and Liu, 2009; Nasirudeenet al., 2008). Therefore, both virus and cytokines contribute to
1 us Res
DG2
ickpHbcJOii(2loi
2
2
NAcaUt
2
lautowslniavaSiioavwbwip
2
(
6 G.P. Sreekanth et al. / Vir
ENV-induced liver injury (Chen et al., 2007; Costa et al., 2012;uabiraba et al., 2010, 2013; Renneson et al., 2011; Sung et al.,012).
Mitogen activated protein kinases (MAPKs) are a family of ser-ne/threonine kinases with three major types, including p38 MAPK,-Jun N-terminal kinase (JNK), and extracellular signal-regulatedinases (ERK1/2) (Raman et al., 2007). Following DENV infection,hosphorylated p38 MAPK is induced (Ceballos-Olvera et al., 2009;uerta-Zepeda et al., 2008; Lee et al., 2008; Nagila et al., 2013). Inhi-ition of p38 MAPK reduces DENV-mediated apoptosis of HepG2ell line (Nagila et al., 2013). DENV also induces phosphorylation ofNK and inhibition of JNK activity reduces DENV infection (Ceballos-lvera et al., 2010). However, the role of ERK1/2 during DENV
nfection remains unclear. Activation of ERK1/2 occurs during DENVnfection of liver cells, human endothelial cells and macrophagesCeballos-Olvera et al., 2010; Huerta-Zepeda et al., 2008; Lee et al.,008), but is blocked in the DENV-infected human alveolar epithe-
ial carcinoma cell line (Chang et al., 2012). To evaluate the rolef ERK1/2 in the pathogenesis of DENV infection, we examined thenfluence of ERK1/2 activation on DENV-mediated apoptosis in vivo.
. Materials and methods
.1. Animals
Eight week-old male Balb/c mice were obtained from theational Laboratory Animal Centre, Mahidol University, Thailand.ll animals were maintained at 23 ± 2 ◦C with a 12 h light/darkycle, autoclaved food and water ad libitum. The protocol waspproved by Siriraj Biosafety Risk Management Taskforce, Mahidolniversity (SI-2013-11) and Siriraj Animal Care and Use Commit-
ee, Mahidol University (SI-ACUP 004/2556).
.2. DENV infection and FR180204 treatment
DENV serotype 2 strain 16881 was propagated in mosquito celline C6/36. Viral titers were determined by focus forming unit (FFU)ssay (Jirakanjanakit et al., 1997). FR180204 (Tocris Bioscience) wassed to inhibit ERK1/2 activation, as FR180204 is previously showno inhibit ERK1/2 with more than 100-fold greater selectivity thanther kinases (Ohori et al., 2005). Two independent experimentsere performed; therefore, totally 48 mice were included in this
tudy. In each experiment, mice were infected intravenously in theateral tail vein with L-15 medium for the mock control (Group I,
= 6). Eighteen mice were infected with DENV. Six were infectedntravenously in the lateral tail vein with DENV in L-15 mediumt a dose of 4 × 105 FFU (Group II, n = 6). Six were infected intra-enously in the lateral tail vein with both DENV in L-15 mediumt a dose of 4 × 105 FFU and 2% DMSO (v/v) alone (Group III, n = 6).ix were infected intravenously in lateral tail vein with both DENVn L-15 medium at a dose of 4 × 105 FFU and FR180204 dissolvedn 2% DMSO at a dose of 50 mg/kg (Group IV, n = 6). Treatmentsf 2% DMSO or FR180204, dissolved in 2% DMSO were carried outn hour before and at an hour and 24 h post DENV infection. Theolume of all injections was 0.4 ml. At day 7 post infection, miceere euthanized by intra-peritoneal injection of sodium pentobar-
ital. Blood samples were processed immediately and liver tissuesere collected for subsequent analysis. Liver tissues were stored
n “RNA later” (Invitrogen) for subsequent RNA preparation androtein extraction.
.3. Quantification of DENV NS1 viral RNA by qRT-PCR assay
Total RNA was extracted from mock (n = 3), DENV-infected livern = 3), DENV-infected liver with 2% DMSO (n = 3) or DENV-infected
earch 188 (2014) 15–26
liver with FR180204 treatment (n = 3) using the Invitrap Spin Uni-versal RNA Mini Kit (Stratec Molecular) and quantified using aNanodrop machine. Equivalent amounts of RNA from each samplewere converted to cDNA with SuperScript® III First-Strand Syn-thesis System (Invitrogen) with a reverse primer, NS1-R 5′ GCCATC AAT GAG AAA GGT CTG G 3′. Amplification was performedusing the SYBR Green I reaction mix (Roche) in the presence ofNS1 specific primers including NS1-F 5′ CCG GCC AGA TCT GGAGAC ATC AAA GGA ATC 3′ and the NS1-R in a Roche Light Cycler480. In vitro transcription-derived DENV NS1 RNA with known copynumber served as a standard control for qRT-PCR. The Ct of viralRNA was measured and compared to the standard control. Resultswere obtained from three independent experiments and analyzedusing the GraphPad Prism 5 program.
2.4. Detection of host RNA by real-time PCR array
The Mouse Apoptosis RT2 ProfilerTM PCR Array (Qiagen) interro-gates 84 genes related to apoptotic pathways. RNA was extractedfrom mock, DENV-infected liver, DENV-infected liver with 2%DMSO or DENV-infected liver with FR180204 treatment using theInvitrap Spin Universal RNA Mini Kit (Stratec Molecular). Total RNAwas quantified using a Nanodrop machine; equivalent amountsof RNA from each sample were converted to cDNA using theSuperScript® III First-Strand Synthesis System (Invitrogen), mixedwith RT2qPCR mastermix containing SYBR Green (Qiagen), andequivalent volumes were aliquoted to each well of the real-timePCR arrays. The real-time PCR cycling program was run on a RocheLight Cycler 480 machine. The threshold cycle (Ct) of each gene wasdetermined and the data were analyzed using the web program athttp://www.pcrdataanalysis.sabiosciences.com/pcr/arrayanalysis.php.
Expression changes detected by the real-time PCR arrayswere validated by quantitative real-time PCR using different setof primers. Total RNA was extracted from mock (n = 3), DENV-infected liver (n = 3), DENV-infected liver with 2% DMSO (n = 3)or DENV-infected liver with FR180204 treatment (n = 3) with theInvitrap Spin Universal RNA Mini Kit (Stratec Molecular) andquantified using a Nanodrop machine. Equivalent amounts of RNAfrom each sample were converted to cDNA using SuperScript® IIIFirst-Strand Synthesis System (Invitrogen), and PCR amplificationwas carried out using TNF-�-specific primers including TNF-F 5′
CCCCCAGTCTGTATCCTTCT 3′ and TNF-R 5′ TTTGAGTCCTTGATG-GTGGT 3′, and GAPDH-specific primers including GAPDH-F5′
TGAATACGGCTACAGCAACA 3′ and GAPDH-R 5′ AGGCCCCTCCT-GTTATTATG 3′, respectively. Amplification was performed usingthe SYBR Green I reaction mix (Roche) in a Roche Light Cycler480. The Ct of mRNA of TNF-� and GAPDH were measured andthe differences between their �Ct were calculated. The relativeexpression values (2−��Ct) were then determined. Results wereobtained from three independent experiments and analyzed usingthe GraphPad Prism 5 program.
2.5. Western blot analysis
Total protein was extracted from mock (n = 3), DENV-infectedliver (n = 3), DENV-infected liver with 2% DMSO (n = 3), DENV-infected liver with FR180204 treatment (n = 3) in RIPA buffercontaining protease inhibitor (Roche) and subjected to Westernblot analysis (Towbin et al., 1979). Phosphatase inhibitor (Roche)was also added to maximize detection of phosphorylated proteins.Protein concentrations of the lysates were determined using the
Bradford protein assay (Bio-Rad) and adjusted to equivalent con-centrations prior to mixing with loading dye and heating at 95 ◦Cfor 5 min. Samples were separated by SDS-PAGE and blotted ontonitrocellulose membrane. The membrane was blocked in 5% BSA
G.P. Sreekanth et al. / Virus Research 188 (2014) 15–26 17
Fig. 1. Quantification of viral NS1 and viral titers from liver tissues of DENV- infected Balb/c mice. Mice were infected with L-15 medium (Mock) or DENV at a dose of 4 × 105
FFU in a volume at 0.4 ml of L-15 medium. Seven days after DENV infection, liver tissues were collected and stored in RNA later. Total viral RNA was isolated for viral NS1quantification and viral supernatant from the liver homogenate for the FFU assay. (A) Standard curve plotted by qRT-PCR, from the threshold cycle numbers (Ct) of tenfold serially diluted cDNA generated from DENV NS1 RNA standard with known copies (the dots in the figure corresponds to each ten fold dilutions) (B) Representativeamplification plot for the NS1 copies of Mock-infected (blue) and DENV-infected viral RNA (red) (C) Viral NS1 copies in 1 �g total RNA of mock-infected and DENV infectedliver tissue (D) Viral titers expressed from liver tissue homogenate in FFU per milligram (FFU/mg) of mock-infected and DENV-infected mice. All the results were obtainedf one bya be sl
ivb((aaciHEa
rom three animals (n = 3) from each group. For (C) and (D), statistical analysis was ds mean ± SEM. The asterisks show the level of significance (p < 0.05 considered toegend, the reader is referred to the web version of the article.)
n TBST for phosphoprotein detection, 5% skim milk in PBST foriral protein, and 5% skim milk in TBST for other proteins. Mem-ranes were washed and hybridized with mouse anti-total ERK1/2Santa Cruz Biotechnology), mouse anti-phosphorylated ERK1/2Santa Cruz Biotechnology), mouse anti-TNF-� (Abcam), mousenti-DENV E (Gentry et al., 1982; Henchal et al., 1985), mousenti-�-actin (Santa Cruz Biotechnology) or rabbit anti-cleavedaspase-3 (Cell signaling). The membranes were again washed and
ncubated with HRP-conjugated secondary antibodies, includingRP-conjugated rabbit anti-mouse IgG for ERK1/2, phosphorylatedRK1/2, TNF-�, DENV E and actin (Dako), or HRP-conjugated swinenti-rabbit IgG for cleaved caspase-3 (Dako). Immune complexesan unpaired t-test using GraphPad Prism 5 program and the data were representedtatistically significant). (For interpretation of the references to color in this figure
were detected by enhanced chemiluminescence (Pierce). Bandintensities were quantified for at least 3 mice from each group usingthe ImageJ program and normalized to respective housekeepinggenes.
2.6. Hematology and transaminases
Whole blood was obtained from mock (n = 12), DENV-infected
liver (n = 12), DENV-infected liver with 2% DMSO (n = 12) or DENV-infected liver with FR180204 treatment (n = 12) and maintainedin EDTA vaccutainer and suddenly processed for hematology. Thecomplete blood count was quantified using a CELL-DYNTM 3700
18 G.P. Sreekanth et al. / Virus Research 188 (2014) 15–26
Fig. 2. Detection of DENV E antigen from the liver tissues of DENV- infected Balb/cmice. (A) Western Blot analysis, using antibody to DENV E and normalized to GAPDH(B) Densitometry analysis of DENV E protein normalized to GAPDH by Western blotanalysis Results were obtained from three animals (n = 3) from each group. Statisticalad(
htta
2
liit
2
oD
Fig. 3. DENV induced liver injury in Balb/c mice. Mice were infected with L-15medium or DENV at a dose of 4 × 105 FFU in a volume at 0.4 ml of L-15 medium.Seven days after DENV infection, blood samples were processed for serum prepa-ration and subsequent analysis of liver enzymes, ALT (A) and AST (B). The resultswere obtained from twelve mice in each group and the exact values of enzymesare expressed in scatter plot. The line represents the average value from the totaltwelve mice from each group Statistical analysis was done by an unpaired t-test
Fotc
nalysis was done by an unpaired t-test using GraphPad Prism 5 program and theata were represented as mean ± SEM. The asterisk shows the level of significancep < 0.05 considered to be statistically significant).
ematological analyzer (Abbott, USA). Blood samples were allowedo clot for preparing the serum. Serum alanine (ALT) and aspar-ate (AST) aminotransferases were quantified using an automaticnalyzer (Model 902, Hitachi Company, Japan).
.7. Histology
Liver tissues were obtained from mock (n = 3), DENV-infectediver (n = 3), DENV-infected liver with 2% DMSO (n = 3) or DENV-nfected liver with FR180204 treatment (n = 3), harvested and fixedn 10% formalin in PBS. Fixed tissues were paraffin embedded, sec-ioned and stained with hematoxylin and eosin (H&E).
.8. Determination of virus titers in DENV-infected mice
At day 7 post DENV infection, mice were euthanized and 20 mgf liver tissues from mock (n = 3), DENV-infected liver (n = 3),ENV-infected liver with 2% DMSO (n = 3) or DENV-infected liver
using GraphPad Prism 5 program and the data were represented as mean ± SEM.The asterisks show the level of significance (p < 0.05 considered to be statisticallysignificant).
ig. 4. Histological analysis for DENV-induced liver injury in Balb/c mice. Mice were infected with L-15 medium or DENV at a dose of 4 × 105 FFU in a volume at 0.4 mlf L-15 medium. Seven days after DENV infection, liver tissues were collected in 10% Formalin in PBS for histological examination (H&E staining). (A) Mock-infected liverissue which shows the normal pathology (B) DENV-infected liver tissue, which shows the classical liver injury induced by DENV, including ballooning of the hepatocyte,ytoplasmic vacuolization, and cellular necrosis. The data shown are representatives for more than three independent experiments.

G.P. Sreekanth et al. / Virus Research 188 (2014) 15–26 19
Fig. 5. DENV altered hematological parameters in Balb/c mice. Mice were infectedwith L-15 medium or DENV at a dose of 4 × 105 FFU in a volume at 0.4 ml of L-15medium. Seven days after DENV infection, blood samples were processed imme-diately for hematological analysis including white blood cells (A), platelets (B) andhematocrit (C). The results obtained from two independent experiments with n = 6ffg
wcpsri5
3
3
a2
Table 1Fold changes in the gene expression profiling of DENV-infected mice compared tocontrol mice.
Gene name Gene description Fold changes
Tnf Tumor necrosis factor 4.9933Tnfrsf11b Tumor necrosis factor receptor superfamily,
member 11b3.7581
Cd40lg CD40 ligand 3.2266Bnip3l BCL2/adenovirus E1B interacting protein 3-like 3.1821Tnfsf10 Tumor necrosis factor (ligand) superfamily,
member 103.1167
Pycard PYD and CARD domain containing 3.0525Cd70 CD70 antigen 2.7895Bnip3 BCL2/adenovirus E1B interacting protein 3 2.6945Bcl2a1a B-cell leukemia/lymphoma 2 related protein
A1a2.3950
Fas Fas (TNF receptor superfamily member 6) 2.2501Traf1 Tnf receptor-associated factor 1 2.0994Nod1 Nucleotide-binding oligomerization domain
containing 11.9185
Dad1 Defender against cell death 1 1.9053Casp12 Caspase 12 1.8790Bcl10 B-cell leukemia/lymphoma 10 1.8404Bax Bcl2-associated X protein 1.8277Bcl2l1 Bcl2-like 1 1.8150Cd40 CD40 antigen 1.8150Bid BH3 interacting domain death agonist 1.7532Gadd45a Growth arrest and DNA-damage-inducible 45
alpha1.7532
Diablo Diablo homolog (Drosophila) 1.7291Apaf1 Apoptotic peptidase activating factor 1 1.7171Bad BCL2-associated agonist of cell death 1.7171Bak1 BCL2-antagonist/killer 1 1.6472Bnip2 BCL2/adenovirus E1B interacting protein 2 1.5801Casp14 Caspase 14 1.5369
or individual group. The results were expressed as mean ± SEM from twelve animalsrom each group. The asterisks indicate statistically significant differences betweenroups (p < 0.05).
ith FR180204 treatment (n = 3), were homogenized in RPMI andentrifuged for 10 min at 14,000 rpm to obtain supernatant. Theresence of infectious viral particles was then determined in theupernatants by the FFU assay (Jirakanjanakit et al., 1997) and rep-esented as FFU/mg of tissue. Results were obtained from threendependent experiments and analyzed using the GraphPad Prism
program.
. Results and discussion
.1. DENV infection induces liver injury in Balb/c mice
Balb/c mice are susceptible to DENV and provide a convenientnimal model of DENV infection (Barth et al., 2006; Paes et al.,005, 2009). In the present study, intravenous infection of DENV
Casp4 Caspase 4 1.5263
in Balb/c mice resulted in detected levels of viral RNA, viral pro-teins and infectious viral particles. To calculate the copy number ofviral RNA, a standard control was firstly generated using DENV NS1RNA derived from in vitro transcription with known copy numberas a template (Fig. 1A). Secondly, qRT-PCR assay was performedusing RNA extracted from mock or DENV-infected liver, respec-tively. Totally, 1.5 × 108 copies of DENV NS1 RNA were detectedin 1 �g of total RNA from liver of DENV-infected mice, but nonewas detected in control (Fig. 1B and C). In addition, viral titersin liver extracts were determined to demonstrate the presence ofreplicative viral particles after DENV infection. Mice were eutha-nized at day 7 after DENV infection and liver tissues from mock orDENV-infected mice were homogenized and centrifuged to obtainsupernatant. The presence of infectious viral particles in super-natants was then measured by the FFU assay. Totally, 1.4 × 105 FFUof infectious viral particles were detected in 1 mg of liver of DENV-infected mice, but none was detected in mock control (Fig. 1D).Western blot analysis was subsequently performed to measurethe amount of viral proteins using an antibody specific to DENVenvelope protein (DENV E). DENV E was detected in livers of DENV-infected mice, but none was detected in control (Fig. 2A and B). Theresult from this study and from others suggest that intravenousinjection system is the good route of administration of DENV inBalb/c mice, as DENV antigens were detected, which is in contrast tothe subcutaneous injection system, in which DENV antigens couldnot be easily detected (Franca et al., 2010).
Balb/c mice have been used to study liver injury during DENVinfection (Barth et al., 2006; Paes et al., 2005, 2009). Liver damagewas more severe in the intravenous injection as compared to theintraperitoneal injection (Barth et al., 2006; Paes et al., 2005, 2009).
Elevation of liver enzymes showed a peak of both AST and ALT at the7th day post DENV infection (Paes et al., 2005, 2009). In the presentstudy, DENV was injected intravenously and could induce liver
20 G.P. Sreekanth et al. / Virus Research 188 (2014) 15–26
Fig. 6. TNF-� expression was increased in DENV-infected Balb/c mice. Mice wereinfected with L-15 medium or DENV at a dose of 4 × 105 FFU in a volume at 0.4 mlof L-15 medium. Seven days after DENV infection, liver tissues were collected, andRNA and protein was isolated. (A) The relative mRNA expression of TNF-� by RT-PCR; (B) Western bloting analysis of TNF-� using antibodies to TNF-�, normalizedto GAPDH; (C) Densitometric analysis of TNF-� at protein level and normalized toGAPDH by Western blot analysis. Results were obtained from three animals (n = 3)from each group. Statistical analysis was done by an unpaired t-test using GraphPadPrism 5 program and the data were represented as mean ± SEM. The asterisks showthe level of significance (p < 0.05 considered to be statistically significant).
iCihpiso(2rp2
sichpta(2
Fig. 7. DENV induced the ERK 1/2 phosphorylation and FR180204 treatmentinhibited ERK 1/2 phosphorylation. Protein was extracted from the liver tissue ofmock-infected, DENV-infected, DMSO treated DENV-infected and FR180204 treatedDENV-infected mice. A cock-tail containing phosphatase inhibitors was also addedto maintain the phospho proteins and allowed them to be visualized by Westernblot analysis. (A) Western blot analysis using antibodies to phosphorylated-ERK1/2(P-ERK 1/2) and total ERK1/2 (t-ERK 1/2) normalized to GAPDH (C) Densitometryanalysis of P-ERK 1, P-ERK 2 and t-ERK at protein level normalized to GAPDH byWestern blot analysis. Results were obtained from three animals (n = 3) from eachgroup. Statistical analysis was done by One-way ANOVA using GraphPad Prism 5program and the data were represented as mean ± SEM. The asterisks show the
including Bcl-2-L10, and Bcl-2 itself, were down-regulated about
njury in Balb/c mice at the 7th day post DENV infection (Fig. 3B and). Both AST and ALT levels in DENV-infected mice increased signif-
cantly compared to those of mock control. Moreover, AST level wasigher than ALT level, which is similar to those in DENV-infectedatients (Nguyen et al., 1997; Paes et al., 2005). Histological exam-
nation of liver tissues from DENV-infected mice in the presenttudy also confirmed the signs of liver injury including ballooningf the hepatocyte, cytoplasmic vacuolization, and cellular necrosisFig. 4A and B). Progressive necrosis in parenchyma (Franca et al.,010; Paes et al., 2005) and increased infiltrating monocytes sur-ounding liver portal area, which could induce liver injury, werereviously reported in DENV-infected mice (de-Oliveira-Pinto et al.,012; Sung et al., 2012).
DENV altered hematological parameters of Balb/c mice in thistudy. The number of white blood cells and platelets in DENV-nfected mice were reduced by about 60% and 25%, respectively,ompared to that of mock-infected mice (Fig. 5A and B). In contrast,ematocrit in DENV-infected mice was increased about 25% com-ared to that of mock-infected mice (Fig. 5C). Our result validateshe model that could mimic the leucopenia, thrombocytopenia,nd hemoconcentration found in other dengue models in C57BL/6Guabiraba et al., 2010) and in DENV-infected patients (Binh et al.,
009).level of significance (p < 0.05 considered to be statistically significant).
3.2. Apoptotic gene expression profile in liver of DENV-infectedBalb/c mice
To gain insight into the molecular basis of liver damage in thismurine DENV model, we examined changes in the expression ofapoptotic genes. Total RNA was prepared from liver tissues of mock-infected and DENV-infected mice and used to interrogate a MouseApoptosis RT2 ProfilerTM PCR Array (Qiagen). Table 1 lists thosegenes exhibiting increased expression (≥1.5 fold) in DENV-infectedmice compared to the mock control. TNF-�, TRAIL and Fas mRNAexpression increased 5-fold, 3- fold, and 2-fold, respectively; theseresults are similar to those obtained from in vitro analysis of apopto-sis in DENV-infected hepatic cell lines (Limjindaporn et al., 2007;Matsuda et al., 2005; Nagila et al., 2013). The pro-apoptotic pro-file of infected cells was also evident by the increased expressionof Bid, Bax and Bak, as well as members of the caspase proteo-lytic cascade, Apaf-1, caspases-4 and caspases-14. Interestingly,CD40L and CD40 mRNA expression also increased significantly inDENV-infected mice. A previous study also shows that CD40L stim-ulated DENV infection of immature dendritic cells, suggesting amechanism for T cell-mediated immunopathology during DENVinfection (Sun et al., 2006). Though the majority of up-regulatedgenes were pro-apoptotic, DAD1, defender against cell death 1, wasalso induced. This is in contrast to the role of DAD1 in yellow headvirus (YHV) infection. The transcriptional level of DAD1 declineddramatically after YHV challenge (Molthathong et al., 2008). Therole of DAD1 in DENV-induced apoptosis merits further investi-gation. As expected, the anti-apoptosis members of Bcl-2 family,
2-fold, and 1.5-fold, respectively.

G.P. Sreekanth et al. / Virus Research 188 (2014) 15–26 21
Fig. 8. Quantification of viral NS1 and viral titers from liver tissues of DENV- infected FR180204 treated Balb/c mice. DENV-infected mice were either treated with 2% DMSO(v/v) alone (n = 6) or treated with FR180204, dissolved in 2% DMSO at a dose of 50 mg/kg (n = 6). Treatments were given an hour before infection at a dose of 4 × 105 FFU andat one and 24 h post infection. At day 7 post infection, liver tissues were collected and stored in “RNA later”. Total viral RNA was isolated for viral NS1 quantification, andviral supernatant from the liver homogenate for the FFU assay. (A) Representative amplification plot for the NS1 copies of 2% DMSO (v/v) treated DENV-infected (pink) andFR180204 treated DENV-infected mice (green). (C) Viral NS1 copies in 1 �g total RNA of 2% DMSO (v/v) treated DENV-infected and FR180204 treated DENV-infected mice.(D) Viral titers expressed from liver tissue homogenate in FFU per milligram (FFU/mg) of 2% DMSO (v/v) treated DENV-infected and FR180204 treated DENV-infected mice.All the results were obtained from three animals (n = 3) from each group. For (B) and (C), statistical analysis was done by an un-paired t-test using GraphPad Prism 5 programa to co
qsiwataee(tisD
nd the data were represented as mean ± SEM. (For interpretation of the references
Expression changes detected using the PCR arrays were subse-uently confirmed by quantitative real-time PCR using differentet of primers. Total RNA was extracted from mock or DENV-nfected liver and equivalent amounts of RNA from each sample
ere converted to cDNA. Amplification was performed by TNF-�nd GAPDH-specific primers using the SYBR Green I reaction mix;he Ct of mRNA of the TNF-� and GAPDH control were measurednd the differences between their �Ct were calculated. The relativexpression values (2−��Ct) were then determined. TNF-�-mRNAxpression increased dramatically, 30-fold post DENV infectionFig. 6A). Western blot analysis was performed and the result shows
hat TNF-� protein expression was also up-regulated in DENV-nfected mice (Fig. 6B and C). Both DENV and TNF-� were previouslyhown to be critical for endothelium damage in a mouse model ofENV infection (Chen et al., 2007). Moreover, anti-TNF antibodylor in this figure legend, the reader is referred to the web version of the article.)
reduced mortality in experimental DENV infection (Atrasheuskayaet al., 2003).
3.3. Inhibition of phosphorylated ERK1/2 by FR180204 treatmentdecreases liver injury and improves clinical parameters inDENV-infected Balb/c mice
ERK1/2 signaling has been implicated in the apoptosis and liverinjury that occurs in rabbits infected with the rabbit hemorrhagicdisease virus (Garcia-Lastra et al., 2009). Following DENV infection,phosphorylated p38 MAPK is induced in vitro (Ceballos-Olvera et al.,
2010; Huerta-Zepeda et al., 2008; Lee et al., 2008; Nagila et al.,2013). Inhibition of phosphorylated p38 MAPK also reduces TNF-� production and apoptosis in DENV-infected HepG2 cells (Nagilaet al., 2013). Furthermore, DENV induces phosphorylation of JNK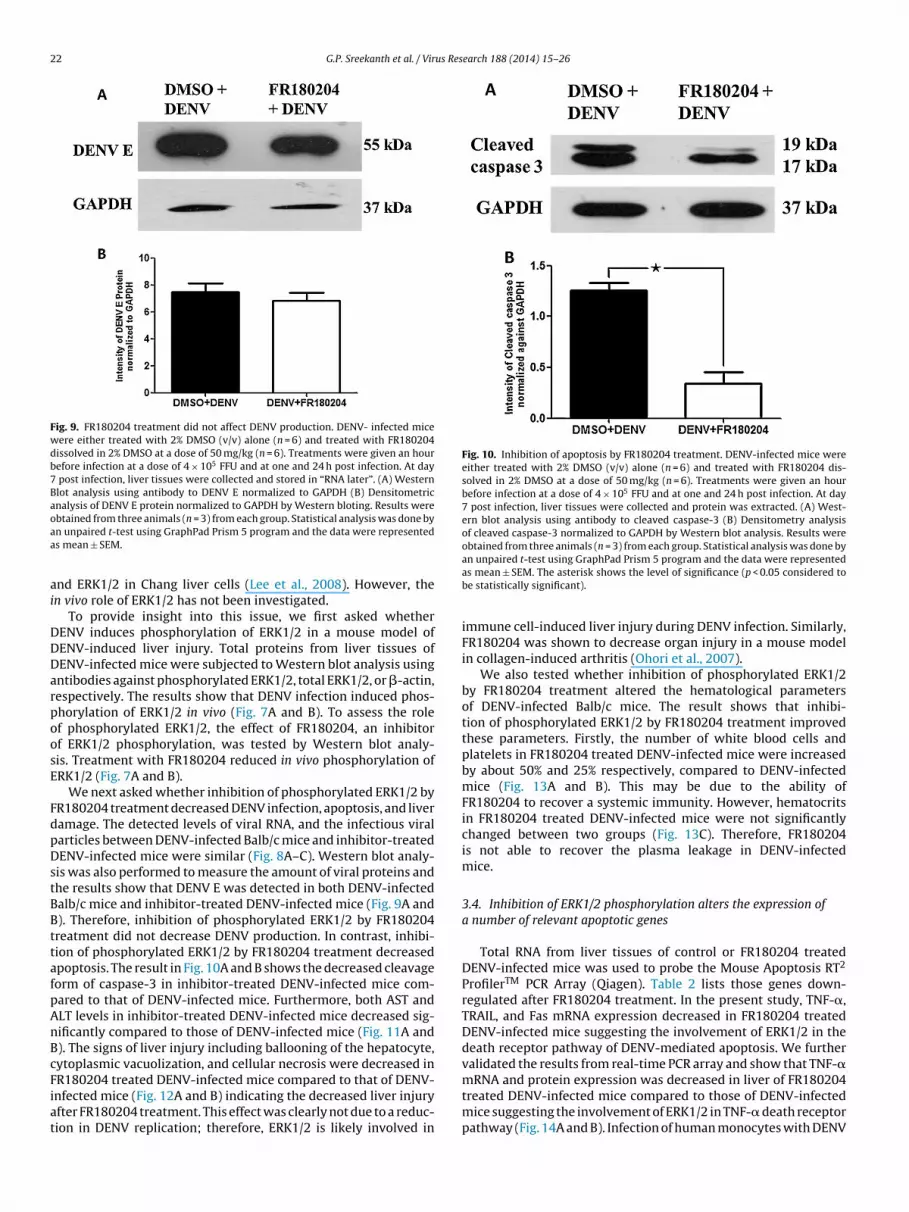
22 G.P. Sreekanth et al. / Virus Research 188 (2014) 15–26
Fig. 9. FR180204 treatment did not affect DENV production. DENV- infected micewere either treated with 2% DMSO (v/v) alone (n = 6) and treated with FR180204dissolved in 2% DMSO at a dose of 50 mg/kg (n = 6). Treatments were given an hourbefore infection at a dose of 4 × 105 FFU and at one and 24 h post infection. At day7 post infection, liver tissues were collected and stored in “RNA later”. (A) WesternBlot analysis using antibody to DENV E normalized to GAPDH (B) Densitometricanalysis of DENV E protein normalized to GAPDH by Western bloting. Results wereoaa
ai
DDDarpoosE
FdpDstBBttafpAnBcFiat
Fig. 10. Inhibition of apoptosis by FR180204 treatment. DENV-infected mice wereeither treated with 2% DMSO (v/v) alone (n = 6) and treated with FR180204 dis-solved in 2% DMSO at a dose of 50 mg/kg (n = 6). Treatments were given an hourbefore infection at a dose of 4 × 105 FFU and at one and 24 h post infection. At day7 post infection, liver tissues were collected and protein was extracted. (A) West-ern blot analysis using antibody to cleaved caspase-3 (B) Densitometry analysisof cleaved caspase-3 normalized to GAPDH by Western blot analysis. Results wereobtained from three animals (n = 3) from each group. Statistical analysis was done by
btained from three animals (n = 3) from each group. Statistical analysis was done byn unpaired t-test using GraphPad Prism 5 program and the data were representeds mean ± SEM.
nd ERK1/2 in Chang liver cells (Lee et al., 2008). However, then vivo role of ERK1/2 has not been investigated.
To provide insight into this issue, we first asked whetherENV induces phosphorylation of ERK1/2 in a mouse model ofENV-induced liver injury. Total proteins from liver tissues ofENV-infected mice were subjected to Western blot analysis usingntibodies against phosphorylated ERK1/2, total ERK1/2, or �-actin,espectively. The results show that DENV infection induced phos-horylation of ERK1/2 in vivo (Fig. 7A and B). To assess the rolef phosphorylated ERK1/2, the effect of FR180204, an inhibitorf ERK1/2 phosphorylation, was tested by Western blot analy-is. Treatment with FR180204 reduced in vivo phosphorylation ofRK1/2 (Fig. 7A and B).
We next asked whether inhibition of phosphorylated ERK1/2 byR180204 treatment decreased DENV infection, apoptosis, and liveramage. The detected levels of viral RNA, and the infectious viralarticles between DENV-infected Balb/c mice and inhibitor-treatedENV-infected mice were similar (Fig. 8A–C). Western blot analy-
is was also performed to measure the amount of viral proteins andhe results show that DENV E was detected in both DENV-infectedalb/c mice and inhibitor-treated DENV-infected mice (Fig. 9A and). Therefore, inhibition of phosphorylated ERK1/2 by FR180204reatment did not decrease DENV production. In contrast, inhibi-ion of phosphorylated ERK1/2 by FR180204 treatment decreasedpoptosis. The result in Fig. 10A and B shows the decreased cleavageorm of caspase-3 in inhibitor-treated DENV-infected mice com-ared to that of DENV-infected mice. Furthermore, both AST andLT levels in inhibitor-treated DENV-infected mice decreased sig-ificantly compared to those of DENV-infected mice (Fig. 11A and). The signs of liver injury including ballooning of the hepatocyte,ytoplasmic vacuolization, and cellular necrosis were decreased in
R180204 treated DENV-infected mice compared to that of DENV-nfected mice (Fig. 12A and B) indicating the decreased liver injuryfter FR180204 treatment. This effect was clearly not due to a reduc-ion in DENV replication; therefore, ERK1/2 is likely involved inan unpaired t-test using GraphPad Prism 5 program and the data were representedas mean ± SEM. The asterisk shows the level of significance (p < 0.05 considered tobe statistically significant).
immune cell-induced liver injury during DENV infection. Similarly,FR180204 was shown to decrease organ injury in a mouse modelin collagen-induced arthritis (Ohori et al., 2007).
We also tested whether inhibition of phosphorylated ERK1/2by FR180204 treatment altered the hematological parametersof DENV-infected Balb/c mice. The result shows that inhibi-tion of phosphorylated ERK1/2 by FR180204 treatment improvedthese parameters. Firstly, the number of white blood cells andplatelets in FR180204 treated DENV-infected mice were increasedby about 50% and 25% respectively, compared to DENV-infectedmice (Fig. 13A and B). This may be due to the ability ofFR180204 to recover a systemic immunity. However, hematocritsin FR180204 treated DENV-infected mice were not significantlychanged between two groups (Fig. 13C). Therefore, FR180204is not able to recover the plasma leakage in DENV-infectedmice.
3.4. Inhibition of ERK1/2 phosphorylation alters the expression ofa number of relevant apoptotic genes
Total RNA from liver tissues of control or FR180204 treatedDENV-infected mice was used to probe the Mouse Apoptosis RT2
ProfilerTM PCR Array (Qiagen). Table 2 lists those genes down-regulated after FR180204 treatment. In the present study, TNF-�,TRAIL, and Fas mRNA expression decreased in FR180204 treatedDENV-infected mice suggesting the involvement of ERK1/2 in thedeath receptor pathway of DENV-mediated apoptosis. We furthervalidated the results from real-time PCR array and show that TNF-�
mRNA and protein expression was decreased in liver of FR180204treated DENV-infected mice compared to those of DENV-infectedmice suggesting the involvement of ERK1/2 in TNF-� death receptorpathway (Fig. 14A and B). Infection of human monocytes with DENV
G.P. Sreekanth et al. / Virus Research 188 (2014) 15–26 23
Fig. 11. FR180204 treatment reduced the liver injury associated with ERK 1/2 phos-phorylation. DENV-infected mice were either treated with 2% DMSO (v/v) alone andtreated with FR180204 dissolved in 2% DMSO at a dose of 50 mg/kg. Treatmentswere given an hour before infection at a dose of 4 × 105 FFU and at one and 24 hpost infection. Seven days after DENV infection, blood samples were processed forserum preparation and subsequent analysis of liver enzymes, ALT (A) and AST (B).The results were obtained from twelve mice in each group and the exact values ofenzymes are expressed in scatter plot. The line represents the average value fromthe total twelve mice from each group. Statistical analysis was done by unpaired t-test using GraphPad Prism 5 program and the data were represented as mean ± SEM.The asterisks show the level of significance (p < 0.05 considered to be statisticallysignificant).
Table 2Ratio decreased in gene expression profiling of FR180204 treated DENV-infectedmice compared to DENV-infected mice.
Gene name Gene description Ratio decreased
Apaf1 Apoptotic peptidase activating factor 1 0.824441Cd40 CD40 antigen 0.779302Tnfsf10 Tumor necrosis factor (ligand)
superfamily, member 100.761844
Bid BH3 interacting domain death agonist 0.755140Bax Bcl2-associated X protein 0.739370Gadd45a Growth arrest and
DNA-damage-inducible 45 alpha0.724504
Bcl2l1 Bcl2-like 1 0.656121Bnip3 BCL2/adenovirus E1B interacting
protein 30.648897
Pycard PYD and CARD domain containing 0.641526Fas Fas (TNF receptor superfamily member
6)0.633968
Casp4 Caspase 4 0.613096Bad BCL2-associated agonist of cell death 0.599455Traf1 Tnf receptor-associated factor 1 0.520360Bak BCL2-antagonist/killer 1 0.506847Cd40lg CD40 ligand 0.485936Bnip2 BCL2/adenovirus E1B interacting
protein 20.478805
Diablo Diablo homolog (Drosophila) 0.467799Nod1 Nucleotide-binding oligomerization
domain containing 10.456612
Cd70 CD70 antigen 0.456596Dad1 Defender against cell death 1 0.441371Bcl10 B-cell leukemia/lymphoma 10 0.413602Casp12 Caspase 12 0.405413Tnfrsf11b Tumor necrosis factor receptor
superfamily, member 11b0.362708
Bcl2a1a B-cell leukemia/lymphoma 2 relatedprotein A1a
0.358298
Casp14 Caspase 14 0.283076
Fig. 12. FR180204 reduced pathology in DENV-induced liver injury. DENV infected micedissolved in 2% DMSO at a dose of 50 mg/kg (n = 6). Treatments were given an hour before iDENV infection, liver tissues were collected in 10% Formalin in PBS for histological examifrom liver injuries by FR180204 treatment. The data shown is representative for more tha
Tnf Tumor necrosis factor 0.220821Bnip3l BCL2/adenovirus E1B interacting
protein 3-like0.198927
is previously shown to increase apoptosis and expression of TNF-�(Espina et al., 2003). In addition, DENV induces endothelial cell pro-duction of reactive nitrogen and oxygen species and apoptotic celldeath, which is greatly enhanced by TNF-alpha (Yen et al., 2008).Inhibition of p38 MAPK in HepG2 cells also reduces TNF-� pro-duction and apoptosis during DENV infection (Nagila et al., 2013).
DENV infection induces apoptosis in HepG2 cells partly throughthe induction of TRAIL (Matsuda et al., 2005). Hepatitis C virusalso uses TRAIL to sensitize hepatocytes to apoptosis (Lan et al.,2008). However, TRAIL was previously shown to limit West Nilewere either treated with 2% DMSO (v/v) alone (n = 6) and treated with FR180204nfection at a dose of 4 × 105 FFU and at one and 24 h post infection. Seven days afternation (H&E staining). (A) The classical liver injury induced by DENV. (B) Recoveryn three independent experiments.

24 G.P. Sreekanth et al. / Virus Research 188 (2014) 15–26
Fig. 13. Hematological parameters in DENV-infected Balb/c mice with FR180204treatment. DENV-infected mice were either treated with 2% DMSO (v/v) alone andtreated with FR180204 dissolved in 2% DMSO at a dose of 50 mg/kg. Treatmentswere given an hour before infection at a dose of 4 × 105 FFU, and at one and 24 h postinfection. Seven days after DENV infection, blood samples were processed imme-diately for hematological analysis including white blood cells (A), platelets (B) andhematocrit (C). The results obtained from two independent experiments with n = 6ffg
v(eipeoaiDaa(
Fig. 14. TNF-� expression was decreased in DENV-infected Balb/c mice withFR180204 treatment. DENV-infected mice were either treated with 2% DMSO (v/v)alone (n = 6) and treated with FR180204 dissolved in 2% DMSO at a dose of 50 mg/kg(n = 6). Treatments were given an hour before infection at a dose of 4 × 105 FFU, andat one and 24 h post infection. Seven days after DENV infection, liver tissues werecollected, RNA and protein was isolated. (A) The relative mRNA expression of TNF-�by RT-PCR (B) Western blot analysis of TNF-� using antibody to TNF-�, normalizedto GAPDH (C) Densitometric analysis of TNF-� at protein level normalized to GAPDHby Western blot analysis. Results were obtained from three animals (n = 3) from each
or individual group. The results were expressed as mean ± SEM from 12 animalsrom each group. The asterisks indicate statistically significant differences betweenroups (p < 0.05).
irus infection of neural tissue and DENV infection of immune cellsShrestha et al., 2012; Warke et al., 2008). Thus, TRAIL may act asither an anti-viral protein to limit virus infection or mediate virus-nduced cell death at different stages of infection. The FasL/Fasathway is involved in DENV-mediated apoptosis of the vascularndothelial cells (Liao et al., 2010). Moreover, nuclear localizationf DENV capsid protein sensitizes HepG2 cells to Fas-mediatedpoptosis (Limjindaporn et al., 2007; Netsawang et al., 2010). Thenvolvement of ERK1/2 signaling in the mitochondrial pathway of
ENV-induced apoptosis is suggested by the decreased Bid, Bax,nd Bak, components of the apoptosome in DENV-infected liverfter FR180204 treatment. Apoptotic protease activating factor 1Apaf-1) was also significantly decreased by FR180204 treatment.group. Statistical analysis was done by an unpaired t-test using GraphPad Prism 5program and the data were represented as mean ± SEM. The asterisks shows thelevel of significance (p < 0.05 considered to be statistically significant).
These changes are consistent with the decrease in cleaved caspase-3 in FR180204 treated DENV-infected mice (Fig. 10) indicating arole of ERK1/2 signaling in DENV-induced apoptosis in vivo.
4. Conclusions
DENV induces phosphorylated ERK1/2 in vivo and inhibition ofERK1/2 by FR180204 treatment significantly reduces immune cell-induced liver injury during DENV infection.
Acknowledgements
This work was supported by Siriraj Research and DevelopmentGrant No. R015533001, Mahidol University, Thailand to TL. SGwas supported by Siriraj Graduate Thesis Scholarship. We appre-ciate the kind assistance from Dr. Amar Nagila for preparing theviral stock, Professor William A. Fonzi, Professor Guy Haegeman,for the critical reading and editing of this manuscript. TL and SNare Thailand Research Fund (TRF) Scholars and PY is a TRF-SeniorResearch Scholar.
References
Atrasheuskaya, A., Petzelbauer, P., Fredeking, T.M., Ignatyev, G., 2003. Anti-TNF anti-body treatment reduces mortality in experimental dengue virus infection. FEMSImmunol. Med. Microbiol. 35 (1), 33–42.

us Res
B
B
C
C
C
C
C
C
d
E
E
F
G
G
G
G
H
HH
H
H
J
K
L
G.P. Sreekanth et al. / Vir
arth, O.M., Barreto, D.F., Paes, M.V., Takiya, C.M., Pinhao, A.T., Schatzmayr, H.G.,2006. Morphological studies in a model for dengue-2 virus infection in mice.Mem. Inst. Oswaldo Cruz 101 (8), 905–915.
inh, P.T., Matheus, S., Huong, V.T., Deparis, X., Marechal, V., 2009. Early clinical andbiological features of severe clinical manifestations of dengue in Vietnameseadults. J. Clin. Virol. 45 (4), 276–280.
eballos-Olvera, I., Chavez-Salinas, S., Medina, F., Ludert, J.E., del Angel, R.M., 2010.JNK phosphorylation, induced during dengue virus infection, is important forviral infection and requires the presence of cholesterol. Virology 396 (1), 30–36.
hang, T.H., Chen, S.R., Yu, C.Y., Lin, Y.S., Chen, Y.S., Kubota, T., Matsuoka, M., Lin, Y.L.,2012. Dengue virus serotype 2 blocks extracellular signal-regulated kinase andnuclear factor-kappaB activation to downregulate cytokine production. PLoSONE 7 (8), e41635.
hen, H.C., Hofman, F.M., Kung, J.T., Lin, Y.D., Wu-Hsieh, B.A., 2007. Both virus andtumor necrosis factor alpha are critical for endothelium damage in a mousemodel of dengue virus-induced hemorrhage. J. Virol. 81 (11), 5518–5526.
hen, H.C., Lai, S.Y., Sung, J.M., Lee, S.H., Lin, Y.C., Wang, W.K., Chen, Y.C., Kao, C.L.,King, C.C., Wu-Hsieh, B.A., 2004. Lymphocyte activation and hepatic cellularinfiltration in immunocompetent mice infected by dengue virus. J. Med. Virol.73 (3), 419–431.
osta, V.V., Fagundes, C.T., Valadao, D.F., Cisalpino, D., Dias, A.C., Silveira, K.D.,Kangussu, L.M., Avila, T.V., Bonfim, M.R., Bonaventura, D., Silva, T.A., Sousa,L.P., Rachid, M.A., Vieira, L.Q., Menezes, G.B., de Paula, A.M., Atrasheuskaya, A.,Ignatyev, G., Teixeira, M.M., Souza, D.G., 2012. A model of DENV-3 infection thatrecapitulates severe disease and highlights the importance of IFN-gamma inhost resistance to infection. PLoS Negl. Trop. Dis. 6 (5), e1663.
ouvelard, A., Marianneau, P., Bedel, C., Drouet, M.T., Vachon, F., Henin, D., Deubel,V., 1999. Report of a fatal case of dengue infection with hepatitis: demonstra-tion of dengue antigens in hepatocytes and liver apoptosis. Hum. Pathol. 30 (9),1106–1110.
e-Oliveira-Pinto, L.M., Marinho, C.F., Povoa, T.F., de Azeredo, E.L., de Souza, L.A.,Barbosa, L.D., Motta-Castro, A.R., Alves, A.M., Avila, C.A., de Souza, L.J., da Cunha,R.V., Damasco, P.V., Paes, M.V., Kubelka, C.F., 2012. Regulation of inflammatorychemokine receptors on blood T cells associated to the circulating versus liverchemokines in dengue fever. PLoS ONE 7 (7), e38527.
l-Bacha, T., Midlej, V., Pereira da Silva, A.P., Silva da Costa, L., Benchimol, M., Galina,A., Da Poian, A.T., 2007. Mitochondrial and bioenergetic dysfunction in humanhepatic cells infected with dengue 2 virus. Biochim. Biophys. Acta 1772 (10),1158–1166.
spina, L.M., Valero, N.J., Hernandez, J.M., Mosquera, J.A., 2003. Increased apoptosisand expression of tumor necrosis factor-alpha caused by infection of culturedhuman monocytes with dengue virus. Am. J. Trop. Med. Hyg. 68 (1), 48–53.
ranca, R.F., Zucoloto, S., da Fonseca, B.A., 2010. A BALB/c mouse model shows thatliver involvement in dengue disease is immune-mediated. Exp. Mol. Pathol. 89(3), 321–326.
arcia-Lastra, R., San-Miguel, B., Crespo, I., Jorquera, F., Alvarez, M., Gonzalez-Gallego, J., Tunon, M.J., 2009. Signaling pathways involved in liver injury andregeneration in rabbit hemorrhagic disease, an animal model of virally-inducedfulminant hepatic failure. Vet. Res. 41 (1), 2.
entry, M.K., Henchal, E.A., McCown, J.M., Brandt, W.E., Dalrymple, J.M., 1982. Identi-fication of distinct antigenic determinants on dengue-2 virus using monoclonalantibodies. Am. J. Trop. Med. Hyg. 31 (3 Pt 1), 548–555.
uabiraba, R., Besnard, A.G., Marques, R.E., Maillet, I., Fagundes, C.T., Conceicao, T.M.,Rust, N.M., Charreau, S., Paris, I., Lecron, J.C., Renauld, J.C., Quesniaux, V., Da Poian,A.T., Arruda, L.B., Souza, D.G., Ryffel, B., Teixeira, M.M., 2013. IL-22 modulates IL-17A production and controls inflammation and tissue damage in experimentaldengue infection. Eur. J. Immunol. 43 (6), 1529–1544.
uabiraba, R., Marques, R.E., Besnard, A.G., Fagundes, C.T., Souza, D.G., Ryffel, B.,Teixeira, M.M., 2010. Role of the chemokine receptors CCR1, CCR2 and CCR4 inthe pathogenesis of experimental dengue infection in mice. PLoS ONE 5 (12),e15680.
alstead, S.B., 1988. Pathogenesis of dengue: challenges to molecular biology. Sci-ence 239 (4839), 476–481.
alstead, S.B., 2007. Dengue. Lancet 370 (9599), 1644–1652.enchal, E.A., McCown, J.M., Burke, D.S., Seguin, M.C., Brandt, W.E., 1985. Epitopic
analysis of antigenic determinants on the surface of dengue-2 virions usingmonoclonal antibodies. Am. J. Trop. Med. Hyg. 34 (1), 162–169.
uerre, M.R., Lan, N.T., Marianneau, P., Hue, N.B., Khun, H., Hung, N.T., Khen,N.T., Drouet, M.T., Huong, V.T., Ha, D.Q., Buisson, Y., Deubel, V., 2001. Liverhistopathology and biological correlates in five cases of fatal dengue fever inVietnamese children. Virchows Arch. 438 (2), 107–115.
uerta-Zepeda, A., Cabello-Gutierrez, C., Cime-Castillo, J., Monroy-Martinez, V.,Manjarrez-Zavala, M.E., Gutierrez-Rodriguez, M., Izaguirre, R., Ruiz-Ordaz, B.H.,2008. Crosstalk between coagulation and inflammation during dengue virusinfection. Thromb. Haemost. 99 (5), 936–943.
irakanjanakit, N., Sanohsomneing, T., Yoksan, S., Bhamarapravati, N., 1997. Themicro-focus reduction neutralization test for determining dengue and Japaneseencephalitis neutralizing antibodies in volunteers vaccinated against dengue.Trans. R. Soc. Trop. Med. Hyg. 91 (5), 614–617.
uo, C.H., Tai, D.I., Chang-Chien, C.S., Lan, C.K., Chiou, S.S., Liaw, Y.F., 1992. Liverbiochemical tests and dengue fever. Am. J. Trop. Med. Hyg. 47 (3), 265–270.
an, L., Gorke, S., Rau, S.J., Zeisel, M.B., Hildt, E., Himmelsbach, K., Carvajal-Yepes,M., Huber, R., Wakita, T., Schmitt-Graeff, A., Royer, C., Blum, H.E., Fischer, R.,Baumert, T.F., 2008. Hepatitis C virus infection sensitizes human hepatocytes toTRAIL-induced apoptosis in a caspase 9-dependent manner. J. Immunol. 181 (7),4926–4935.
earch 188 (2014) 15–26 25
Lee, Y.R., Lei, H.Y., Chen, S.H., Wang, J.R., Lin, Y.S., Yeh, T.M., Liu, C.C., Liu, H.S., 2008.Signaling pathways involved in dengue-2 virus infection induced RANTES over-expression. Am. J. Infect. Dis. 4 (1), 32–40.
Liao, H., Xu, J., Huang, J., 2010. FasL/Fas pathway is involved in dengue virusinduced apoptosis of the vascular endothelial cells. J. Med. Virol. 82 (8),1392–1399.
Limjindaporn, T., Netsawang, J., Noisakran, S., Thiemmeca, S., Wongwiwat, W., Sud-saward, S., Avirutnan, P., Puttikhunt, C., Kasinrerk, W., Sriburi, R., Sittisombut,N., Yenchitsomanus, P.T., Malasit, P., 2007. Sensitization to Fas-mediated apo-ptosis by dengue virus capsid protein. Biochem. Biophys. Res. Commun. 362 (2),334–339.
Limonta, D., Capo, V., Torres, G., Perez, A.B., Guzman, M.G., 2007. Apoptosis in tissuesfrom fatal dengue shock syndrome. J. Clin. Virol. 40 (1), 50–54.
Lin, C.F., Wan, S.W., Chen, M.C., Lin, S.C., Cheng, C.C., Chiu, S.C., Hsiao, Y.L., Lei, H.Y.,Liu, H.S., Yeh, T.M., Lin, Y.S., 2008. Liver injury caused by antibodies againstdengue virus nonstructural protein 1 in a murine model. Lab. Invest. 88 (10),1079–1089.
Marianneau, P., Steffan, A.M., Royer, C., Drouet, M.T., Kirn, A., Deubel, V., 1998. Differ-ing infection patterns of dengue and yellow fever viruses in a human hepatomacell line. J. Infect. Dis. 178 (5), 1270–1278.
Matsuda, T., Almasan, A., Tomita, M., Tamaki, K., Saito, M., Tadano, M., Yagita, H., Ohta,T., Mori, N., 2005. Dengue virus-induced apoptosis in hepatic cells is partly medi-ated by Apo2 ligand/tumour necrosis factor-related apoptosis-inducing ligand.J. Gen. Virol. 86 (Pt 4), 1055–1065.
Molthathong, S., Buaklin, A., Senapin, S., Klinbunga, S., Rojtinnakorn, J., Flegel, T.W.,2008. Up-regulation of ribophorin I after yellow head virus (YHV) challengein black tiger shrimp Penaeus monodon. Fish Shellfish Immunol. 25 (1–2),40–46.
Morchang, A., Yasamut, U., Netsawang, J., Noisakran, S., Wongwiwat, W.,Songprakhon, P., Srisawat, C., Puttikhunt, C., Kasinrerk, W., Malasit, P., Yen-chitsomanus, P.T., Limjindaporn, T., 2011. Cell death gene expression profile:role of RIPK2 in dengue virus-mediated apoptosis. Virus Res. 156 (1–2),25–34.
Nagila, A., Netsawang, J., Srisawat, C., Noisakran, S., Morchang, A., Yasamut, U., Put-tikhunt, C., Kasinrerk, W., Malasit, P., Yenchitsomanus, P.T., Limjindaporn, T.,2011. Role of CD137 signaling in dengue virus-mediated apoptosis. Biochem.Biophys. Res. Commun. 410 (3), 428–433.
Nagila, A., Netsawang, J., Suttitheptumrong, A., Morchang, A., Khunchai, S., Srisawat,C., Puttikhunt, C., Noisakran, S., Yenchitsomanus, P.T., Limjindaporn, T., 2013.Inhibition of p38MAPK and CD137 signaling reduce dengue virus-induced TNF-alpha secretion and apoptosis. Virol. J. 10 (1), 105.
Nasirudeen, A.M., Liu, D.X., 2009. Gene expression profiling by microarray analysisreveals an important role for caspase-1 in dengue virus-induced p53-mediatedapoptosis. J. Med. Virol. 81 (6), 1069–1081.
Nasirudeen, A.M., Wang, L., Liu, D.X., 2008. Induction of p53-dependent andmitochondria-mediated cell death pathway by dengue virus infection of humanand animal cells. Microbes Infect. 10 (10–11), 1124–1132.
Netsawang, J., Noisakran, S., Puttikhunt, C., Kasinrerk, W., Wongwiwat, W., Malasit,P., Yenchitsomanus, P.T., Limjindaporn, T., 2010. Nuclear localization of denguevirus capsid protein is required for DAXX interaction and apoptosis. Virus Res.147 (2), 275–283.
Nguyen, T.L., Nguyen, T.H., Tieu, N.T., 1997. The impact of dengue haemorrhagic feveron liver function. Res. Virol. 148 (4), 273–277.
Ohori, M., Kinoshita, T., Okubo, M., Sato, K., Yamazaki, A., Arakawa, H., Nishimura,S., Inamura, N., Nakajima, H., Neya, M., Miyake, H., Fujii, T., 2005. Identificationof a selective ERK inhibitor and structural determination of the inhibitor-ERK2complex. Biochem. Biophys. Res. Commun. 336 (1), 357–363.
Ohori, M., Takeuchi, M., Maruki, R., Nakajima, H., Miyake, H., 2007. FR180204, a noveland selective inhibitor of extracellular signal-regulated kinase, amelioratescollagen-induced arthritis in mice. Naunyn. Schmiedeberg’s Arch. Pharmacol.374 (4), 311–316.
Paes, M.V., Lenzi, H.L., Nogueira, A.C., Nuovo, G.J., Pinhao, A.T., Mota, E.M., Basilio-de-Oliveira, C.A., Schatzmayr, H., Barth, O.M., Alves, A.M., 2009. Hepatic damageassociated with dengue-2 virus replication in liver cells of BALB/c mice. Lab.Invest. 89 (10), 1140–1151.
Paes, M.V., Pinhao, A.T., Barreto, D.F., Costa, S.M., Oliveira, M.P., Nogueira, A.C., Takiya,C.M., Farias-Filho, J.C., Schatzmayr, H.G., Alves, A.M., Barth, O.M., 2005. Liverinjury and viremia in mice infected with dengue-2 virus. Virology 338 (2),236–246.
Raman, M., Chen, W., Cobb, M.H., 2007. Differential regulation and properties ofMAPKs. Oncogene 26 (22), 3100–3112.
Renneson, J., Guabiraba, R., Maillet, I., Marques, R.E., Ivanov, S., Fontaine, J., Paget, C.,Quesniaux, V., Faveeuw, C., Ryffel, B., Teixeira, M.M., Trottein, F., 2011. A detri-mental role for invariant natural killer T cells in the pathogenesis of experimentaldengue virus infection. Am. J. Pathol. 179 (4), 1872–1883.
Shrestha, B., Pinto, A.K., Green, S., Bosch, I., Diamond, M.S., 2012. CD8+ T cells useTRAIL to restrict West Nile virus pathogenesis by controlling infection in neu-rons. J. Virol. 86 (17), 8937–8948.
Souza, L.J., Alves, J.G., Nogueira, R.M., Gicovate Neto, C., Bastos, D.A., Siqueira, E.W.,Souto Filho, J.T., Cezario Tde, A., Soares, C.E., Carneiro Rda, C., 2004. Aminotrans-ferase changes and acute hepatitis in patients with dengue fever: analysis of
1585 cases. Braz. J. Infect. Dis. 8 (2), 156–163.Sun, P., Celluzzi, C.M., Marovich, M., Subramanian, H., Eller, M., Widjaja, S., Palmer, D.,Porter, K., Sun, W., Burgess, T., 2006. CD40 ligand enhances dengue viral infectionof dendritic cells: a possible mechanism for T cell-mediated immunopathology.J. Immunol. 177 (9), 6497–6503.

2 us Res
S
T
T
555–564.
6 G.P. Sreekanth et al. / Vir
ung, J.M., Lee, C.K., Wu-Hsieh, B.A., 2012. Intrahepatic infiltrating NK and CD8T cellscause liver cell death in different phases of dengue virus infection. PLoS ONE 7(9), e46292.
hongtan, T., Panyim, S., Smith, D.R., 2004. Apoptosis in dengue virus infected livercell lines HepG2 and Hep3B. J. Med. Virol. 72 (3), 436–444.
owbin, H., Staehelin, T., Gordon, J., 1979. Electrophoretic transfer of proteins frompolyacrylamide gels to nitrocellulose sheets: procedure and some applications.Proc. Natl. Acad. Sci. U.S.A. 76 (9), 4350–4354.
earch 188 (2014) 15–26
Warke, R.V., Martin, K.J., Giaya, K., Shaw, S.K., Rothman, A.L., Bosch, I., 2008.TRAIL is a novel antiviral protein against dengue virus. J. Virol. 82 (1),
Yen, Y.T., Chen, H.C., Lin, Y.D., Shieh, C.C., Wu-Hsieh, B.A., 2008. Enhancement bytumor necrosis factor alpha of dengue virus-induced endothelial cell productionof reactive nitrogen and oxygen species is key to hemorrhage development. J.Virol. 82 (24), 12312–12324.





























