Embed Size (px)
Citation preview

The European Journal of Heart Failure 5(2003)229–239
1388-9842/03/$30.00� 2003 European Society of Cardiology. Published by Elsevier Science B.V. All rights reserved.doi:10.1016/S1388-9842Ž03.00010-2
Role of nitric oxide in posthypoxic contractile dysfunction of diabeticcardiomyopathy
Magdi M. El-Omar , Roger Lord , Nick J. Draper , Ajay M. Shah *a b a c,
Department of Cardiology, University of Wales College of Medicine, Heath Park, Cardiff CF4 4XN, UKa
Discipline of Surgery, University of Tasmania, Hobart, Australiab
Guy’s, King’s & St. Thomas’s School of Medicine, King’s College London, Bessemer Road, London SE5 9PJ, UKc
Received 16 August 2002; received in revised form 18 October 2002; accepted 12 November 2002
Abstract
We investigated the role of nitric oxide synthase(NOS) in the contractile dysfunction of diabetic cardiomyopathy, comparingstreptozotocin-treated(60 mgykg) diabetic Wistar rats with matched non-diabetic controls. Isolated isovolumic heart function wasstudied during normoxia and in response to brief hypoxia-reoxygenation. Diabetic hearts had significantly lower left-ventricularpressure and slower isovolumic relaxation than controls(relaxation time constant,T 40.2"2.3 vs. 27.7"0.9 ms;P-0.05) and ablunted response to hypoxia. These abnormalities were unaffected by NOS inhibition. Upon reoxygenation after brief hypoxia,diabetic hearts exhibited substantial worsening of LV relaxation compared to normal hearts(T 69.1"3.3 vs. 56.6"7.9 ms;P-0.05). This post-hypoxic diastolic dysfunction was significantly attenuated either by the non-selective NOS inhibitor L-NAME,the iNOS inhibitor L-NIL, or the reactive-oxygen-species(ROS) scavenger thiourea. Only diabetic hearts expressed iNOS protein,whereas eNOS expression was similar in both groups. In conclusion, diabetic hearts exhibit markedly abnormal post-hypoxicrelaxation, which is attributable to both ROS and NO derived from iNOS.� 2003 European Society of Cardiology. Published by Elsevier Science B.V. All rights reserved.
Keywords: Diastole; Nitric oxide synthase(NOS); Hypoxia; Relaxation; Reactive oxygen species
1. Introduction
Diabetes mellitus is associated with a specific cardio-myopathy, independent of hypertension, coronary arterydisease or hyperlipidaemia, as evidenced by clinicalstudies and experimental animal models, such as strep-tozotocin-induced diabetesw1–3x. Prominent early fea-tures of diabetic cardiomyopathy include impairedmyocardial relaxation and increased diastolic stiffness(i.e. diastolic dysfunction), abnormalities which areexacerbated following even brief ischaemiaw1–3x. Moreadvanced cardiomyopathy is characterised by systolicand autonomic dysfunction. The underlying mechanismsof diabetic cardiomyopathy remain incompletely under-stood. Intrinsic abnormalities of excitation–contractioncoupling and the myofilaments, as well as metabolicdysfunction, are likely to contributew1–3x.
*Corresponding author. Tel:q44-207-346-3865; fax:q44-207-346-4771.
E-mail address: [email protected](A.M. Shah).
Previous work from our laboratory and others hasestablished that nitric oxide(NO) is an importantmodulator of myocardial functionw4,5x. Endothelium-derived NO modulates myocardial relaxation, diastolictone and oxygen consumption in preparations rangingfrom single myocytes and isolated hearts to the intactheart in vivo w5,6x. NO may also be generated physio-logically within cardiac myocytes by an endothelial-typenitric oxide synthase(eNOS), and can modulateb-adrenergic inotropic response. In certain pathologicalsettings, the inducible form of nitric oxide synthase(iNOS) is expressed in cardiac myocytes and endothelialcells, as well as in other cell types. Expression of iNOSin dilated cardiomyopathy and endotoxic shock is sug-gested to contribute to myocardial dysfunctionw4,5x. Inthe context of ischaemia-reperfusion or hypoxia-reoxy-genation, the reaction of NO with superoxide radicalsto form peroxynitrite may induce deleterious effects,depending upon the level of peroxynitrite generatedw6,7x.
by guest on Novem
ber 21, 2013http://eurjhf.oxfordjournals.org/
Dow
nloaded from

230 M.M. El-Omar et al. / The European Journal of Heart Failure 5 (2003) 229–239
The possible role of eNOS or iNOS in the develop-ment of diabetic cardiac dysfunction is unclear. How-ever, many recent studies have reported alterations inthe expression of NOS isoforms andyor NO bioactivityin several diabetic tissues. For example, iNOS is report-ed to be expressed in the diabetic kidneyw8x, vascularsmooth musclew9x and retinaw10x. Vascular endothelialdysfunction, in part due to reduced bioavailability ofNO, is known to be a feature of diabetesw7,11x. Theaims of this study were to examine:(a) the potentialroles of NOS in baseline cardiac dysfunction in thestreptozotocin-treated rat, an experimental model ofinsulin-dependent diabetic cardiomyopathy independentof coronary atherosclerosisw2,3x; and (b) the responseof the diabetic heart to brief hypoxia-reoxygenation, aninsult that exacerbates diastolic dysfunction.
2. Methods
2.1. Experimental animals
Experiments were performed in accordance with theUK Animals (Scientific Procedures)Act 1986 andinstitutional guidelines. Diabetes was induced in maleWistar rats(200–300 g)by intraperitoneal streptozoto-cin injection (60 mgykg in 0.1 M citrate buffer).Diabetes was confirmed on the third day post-strepto-zotocin by hyperglycaemia()20 mmolyl, Glucometer).Approximately 15% of rats had glucose levels-20mmolyl and were excluded from the study. Weeklyurinalysis was performed for ketonuria. At killing 4–6weeks later, bloodyserum were collected for estimationof glycosylated haemoglobin A1(HbA1C) and randomglucose. A total of 64 diabetic animals were studied.The control group comprised 55 age- and sex-matched
non-diabetic litter mates, which were studied contem-poraneously. All studies were comparisons of diabeticanimals against matched non-diabetic(control)animals.
2.2. Isolated perfused hearts
Animals were terminally anaesthetised with intraper-itoneal sodium pentobarbitone(60 mgykg). Hearts wererapidly excised and mounted on a non-recirculatingLangendorff apparatus. The perfusion solution com-prised (in mM): NaCl 117; KCl 5.7; NaHCO 4.4;3
NaH PO 1.2; CaCl 1.25; MgCl 1.7; hydroxyethylpi-2 4 2 2
perazine ethanesulfonic acid(HEPES) 20; and glucose10; pH 7.4, 378C. The pH was adjusted to 7.4 withNaOH. This was gassed with 100% O during normoxic2
perfusion (pO f700 mmHg) and 100% N during2 2
hypoxic periods(pO f40 mmHg). Coronary flow rate2
was adjusted to give a mean coronary perfusion pressure(CPP) of 80 mmHg, and was then maintained constant.Hearts were paced at 10% above intrinsic rate by a rightatrial electrode at 10% above threshold voltage. Left
ventricular pressure(LVP) was monitored using anintraventricular balloon connected to a Statham(P23XL)pressure transducer. LV end-diastolic pressure(LVEDP)was set to 10 mmHg. Pressure data were sampled via aMacLab data module(AD Instruments, UK). LV dPydt was obtained from the first derivative of the LVPmax
signal. The exponential time-constant of isovolumic LVrelaxation, T, was calculated as previously describedw12x.
2.3. Experimental protocol
LV function of diabetic and control hearts was com-pared during normoxia(after at least 20 min of equili-bration) and during exposure to brief hypoxia(10 min,pO f40 mmHg) and reoxygenation. We studied the2
effects of the following interventions, each in a separategroup of diabetic or non-diabetic hearts(nG6, exceptns4 for control qL-NIL group): (a) no additionalagents added;(b) a non-selective NOS inhibitor, L-NAME (50 mM); (c) a selective iNOS inhibitor, L-NIL(10 mM); (d) a cyclooxygenase inhibitor, indomethacin(10 mM); (e) a lipooxygenase inhibitor, diethylcarba-mazine(0.25 mM); (f) a scavenger of reactive oxygenspecies(ROS), thiourea(10 mM); and(g) insulin 0.01IUyml. In hypoxia-reoxygenation experiments, test com-pounds were added at least 5 min prior to hypoxia andremained present subsequently. At the end of experi-ments, hearts were blotted dry and weighed.
2.4. Western analyses for eNOS and iNOS
Cardiac myocytes were isolated from perfused heartsby collagenase digestionw13x. After passage through a2% BSA cushion, they comprised)95% rod-shapedstriated cells with no significant endothelial cell contam-ination. Myocytes were lysed in 1 ml of boiling 1%SDS with 10 mM Tris(pH 7.4). Protein concentrationwas measured using a micro BCA kit(Pierce). Equalamounts of protein(100 mgylane) were run on 8.5%SDS-polyacrylamide gels and electroblotted onto nitro-cellulose membrane. Membranes were incubated eitherwith a rabbit anti-iNOS polyclonal antibody(Calbio-chem, La Jolla, CA) or a rabbit anti-eNOS polyclonalantibody(Alpha Diagnostics, TX). The secondary anti-body was a horseradish peroxidase-linked donkey anti-rabbit antibody(Amersham, UK). Blots were assessedby densitometry(Bio Rad GS-700 densitometer, Her-cules, CA).
2.5. Measurement of total nitriteynitrate (NO )x
Plasma samples from control and streptozotocin-treat-ed rats were stored aty20 8C. Total plasma NO wasx
measured using a 2,3-diamino-napthalene fluorometricassay, adapted for use in a 96-well plate, after treating
by guest on Novem
ber 21, 2013http://eurjhf.oxfordjournals.org/
Dow
nloaded from

231M.M. El-Omar et al. / The European Journal of Heart Failure 5 (2003) 229–239
Table 1Baseline characteristics of control and diabetic groups
Parameter Control Diabetic
Blood glucose(mmolyl) 11.3"0.3 40.8"2.6*
HbA1C (%) 4.0"0.03 11.9"0.5*
Body weight(g) 386"10.2 184.6"8.9*
Heart weight(g) 1.4"0.04 0.87"0.02*
Heartybody weight(gyg) 0.0036"0.0001 0.0048"0.0001*
Spontaneous heart rate(bpm) 264.3"8.3 232"8.8*
Paced heart rate(bpm) 286.6"7.8 257.4"6.6*
LVDP (mmHg) 78.0"6.7 59.5"4.3*
LV dPydt (mmHgys)max 2120.9"120.0 1255.1"94.0*
T (ms) 27.7"0.9 40.2"2.3*
CPP(mmHg) 83.0"0.7 80.7"0.5Coronary flow per gram heart tissue(mlymin g) 7.3"0.6 7.8"0.8
Values are expressed as mean"S.E.M.;n)30 per group.P-0.05 C vs. STZ.*
Fig. 1. eNOS and iNOS protein expression in diabetic and control myocardium.(a) Western blot showing isolated cardiac myocyte protein probedfor eNOS: lanes a–c, control myocytes, lanes d–f, diabetic myocytes, lane g, eNOS positive control(Alpha Diagnostics, TX). Similar resultswere obtained in two independent experiments.(b) Isolated cardiac myocyte protein probed for iNOS: lanes a–e, diabetic myocytes, lanes f–j,control myocytes, lane k, iNOS positive control(Calbiochem). Similar results were obtained in two experiments.
with nitrate reductase to convert nitrate to nitritew14x.Total NO concentration was assessed from the 450-nmx
fluorescence emission after excitation at 375 nm, usingsodium nitrite and sodium nitrate to generate standardcurves.
2.6. Materials
Unless stated otherwise, all agents were obtained fromSigma Chemical Co(UK) and were prepared fresh inphysiological buffer, apart from indomethacin, whichwas initially dissolved in dimethylsulfoxide(DMSO).The final DMSO concentration(-0.01%) had no sig-nificant effects.
2.7. Data analysis
Results are presented as mean"standard error.Responses to hypoxia-reoxygenation, as well as baseline
effects of compounds, were compared by two-wayrepeated-measures ANOVA, followed by a Student–Neumann–Keul test to isolate differences. Comparisonof single effects among groups was by one-way ANO-VA, followed by a Student–Neumann–Keul test toisolate differences.P-0.05 was considered significant.
3. Results
3.1. Baseline characteristics
Streptozotocin-treated rats had severe hyperglycaemia,elevated HbA1C, and reduced body and heart weightscompared to controls(Table 1). The relative loss ofheart weight was less than that of body weight, asreflected in higher heartybody weight ratios(Table 1).None of the animals had any clinical evidence ofinfection either before sacrifice or at post-mortem. In asubset of eight diabetic animals, rectal temperatures
by guest on Novem
ber 21, 2013http://eurjhf.oxfordjournals.org/
Dow
nloaded from
232 M.M. El-Omar et al. / The European Journal of Heart Failure 5 (2003) 229–239
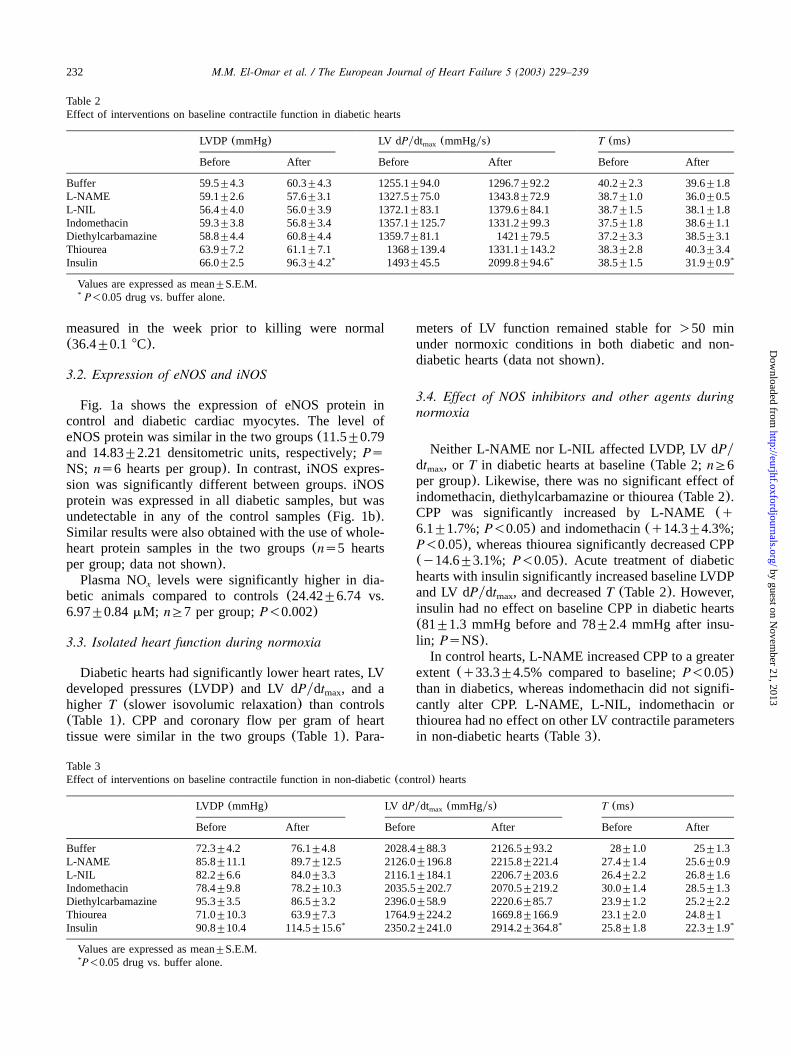
Table 2Effect of interventions on baseline contractile function in diabetic hearts
LVDP (mmHg) LV dPydt (mmHgys)max T (ms)
Before After Before After Before After
Buffer 59.5"4.3 60.3"4.3 1255.1"94.0 1296.7"92.2 40.2"2.3 39.6"1.8L-NAME 59.1"2.6 57.6"3.1 1327.5"75.0 1343.8"72.9 38.7"1.0 36.0"0.5L-NIL 56.4"4.0 56.0"3.9 1372.1"83.1 1379.6"84.1 38.7"1.5 38.1"1.8Indomethacin 59.3"3.8 56.8"3.4 1357.1"125.7 1331.2"99.3 37.5"1.8 38.6"1.1Diethylcarbamazine 58.8"4.4 60.8"4.4 1359.7"81.1 1421"79.5 37.2"3.3 38.5"3.1Thiourea 63.9"7.2 61.1"7.1 1368"139.4 1331.1"143.2 38.3"2.8 40.3"3.4Insulin 66.0"2.5 96.3"4.2* 1493"45.5 2099.8"94.6* 38.5"1.5 31.9"0.9*
Values are expressed as mean"S.E.M.P-0.05 drug vs. buffer alone.*
Table 3Effect of interventions on baseline contractile function in non-diabetic(control) hearts
LVDP (mmHg) LV dPydt (mmHgys)max T (ms)
Before After Before After Before After
Buffer 72.3"4.2 76.1"4.8 2028.4"88.3 2126.5"93.2 28"1.0 25"1.3L-NAME 85.8"11.1 89.7"12.5 2126.0"196.8 2215.8"221.4 27.4"1.4 25.6"0.9L-NIL 82.2"6.6 84.0"3.3 2116.1"184.1 2206.7"203.6 26.4"2.2 26.8"1.6Indomethacin 78.4"9.8 78.2"10.3 2035.5"202.7 2070.5"219.2 30.0"1.4 28.5"1.3Diethylcarbamazine 95.3"3.5 86.5"3.2 2396.0"58.9 2220.6"85.7 23.9"1.2 25.2"2.2Thiourea 71.0"10.3 63.9"7.3 1764.9"224.2 1669.8"166.9 23.1"2.0 24.8"1Insulin 90.8"10.4 114.5"15.6* 2350.2"241.0 2914.2"364.8* 25.8"1.8 22.3"1.9*
Values are expressed as mean"S.E.M.P-0.05 drug vs. buffer alone.*
measured in the week prior to killing were normal(36.4"0.1 8C).
3.2. Expression of eNOS and iNOS
Fig. 1a shows the expression of eNOS protein incontrol and diabetic cardiac myocytes. The level ofeNOS protein was similar in the two groups(11.5"0.79and 14.83"2.21 densitometric units, respectively;PsNS; ns6 hearts per group). In contrast, iNOS expres-sion was significantly different between groups. iNOSprotein was expressed in all diabetic samples, but wasundetectable in any of the control samples(Fig. 1b).Similar results were also obtained with the use of whole-heart protein samples in the two groups(ns5 heartsper group; data not shown).Plasma NO levels were significantly higher in dia-x
betic animals compared to controls(24.42"6.74 vs.6.97"0.84mM; nG7 per group;P-0.002)
3.3. Isolated heart function during normoxia
Diabetic hearts had significantly lower heart rates, LVdeveloped pressures(LVDP) and LV dPydt , and amax
higher T (slower isovolumic relaxation) than controls(Table 1). CPP and coronary flow per gram of hearttissue were similar in the two groups(Table 1). Para-
meters of LV function remained stable for)50 minunder normoxic conditions in both diabetic and non-diabetic hearts(data not shown).
3.4. Effect of NOS inhibitors and other agents duringnormoxia
Neither L-NAME nor L-NIL affected LVDP, LV dPydt , or T in diabetic hearts at baseline(Table 2;nG6max
per group). Likewise, there was no significant effect ofindomethacin, diethylcarbamazine or thiourea(Table 2).CPP was significantly increased by L-NAME(q6.1"1.7%;P-0.05) and indomethacin(q14.3"4.3%;P-0.05), whereas thiourea significantly decreased CPP(y14.6"3.1%; P-0.05). Acute treatment of diabetichearts with insulin significantly increased baseline LVDPand LV dPydt , and decreasedT (Table 2). However,max
insulin had no effect on baseline CPP in diabetic hearts(81"1.3 mmHg before and 78"2.4 mmHg after insu-lin; PsNS).In control hearts, L-NAME increased CPP to a greater
extent (q33.3"4.5% compared to baseline;P-0.05)than in diabetics, whereas indomethacin did not signifi-cantly alter CPP. L-NAME, L-NIL, indomethacin orthiourea had no effect on other LV contractile parametersin non-diabetic hearts(Table 3).
by guest on Novem
ber 21, 2013http://eurjhf.oxfordjournals.org/
Dow
nloaded from

233M.M. El-Omar et al. / The European Journal of Heart Failure 5 (2003) 229–239
3.5. Isolated heart function during brief hypoxia-reoxygenation
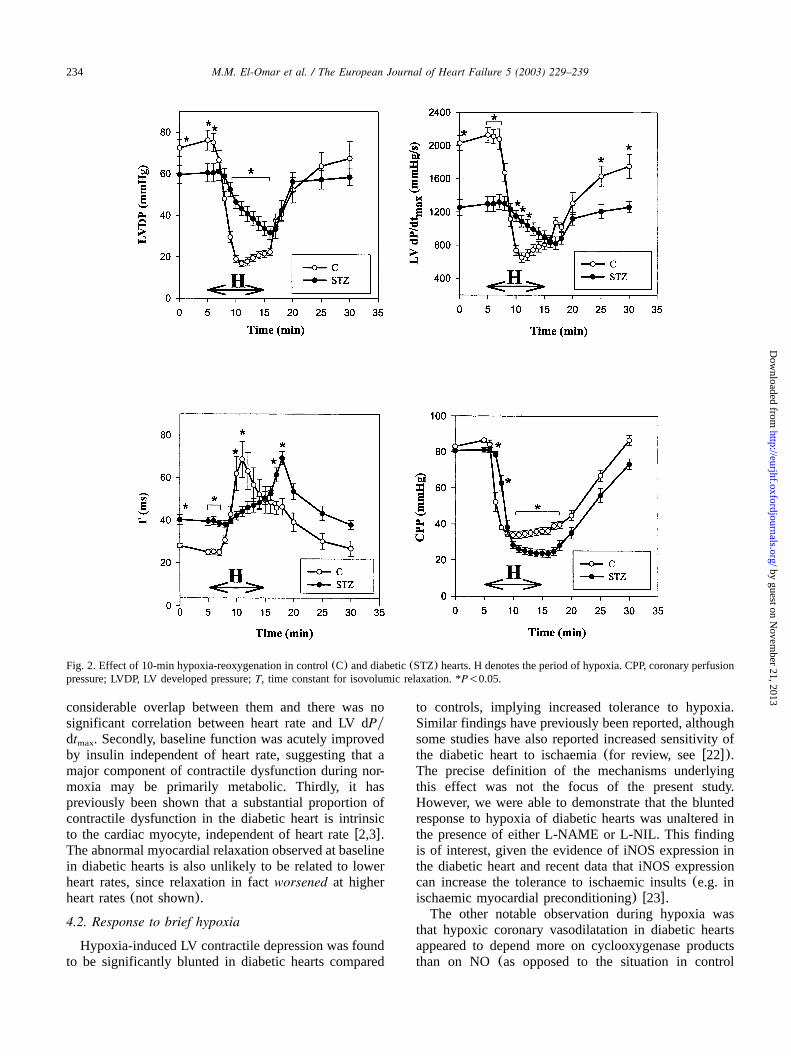
Fig. 2 shows the effects of 10-min hypoxia-reoxygen-ation in control and diabetic hearts. In controls, hypoxiacaused marked reduction in LVDP, LV dPydt andmax
CPP, and prolongedT within 1–2 min. In markedcontrast, in diabetic hearts there was a much slowerdecline in LVDP and LV dPydt , and a more gradualmax
prolongation ofT. Thus, end-hypoxic levels of LVDPand LV dPydt were higher in diabetic than controlmax
hearts, despite significantly lower pre-hypoxic values.However, the reduction in CPP in diabetic hearts wassimilar to controls, reaching a nadir slightly lower thanthe controls.Upon reoxygenation of control hearts, all indices
recovered fairly rapidly. There was a minor deteriorationin T soon after reoxygenation(average 3.2"1.0 ms)(Fig. 2). By 15 min after reoxygenation, LVDP, CPPandT were not significantly different from pre-hypoxiclevels, but LV dPydt was still slightly lower than themax
pre-hypoxic level.In diabetic hearts, however, upon reoxygenation there
was a striking further prolongation inT (average18.6"3.0 ms or 50.2"9.3% of the baseline value),which gradually recovered over the next 10–15 min(Fig. 2). In contrast, LVDP, LV dPydt and CPP inmax
diabetic hearts recovered rapidly following reoxygena-tion. Thus, reoxygenation was accompanied by a tran-sient, but profound, further slowing of LV relaxation inthe diabetic group.
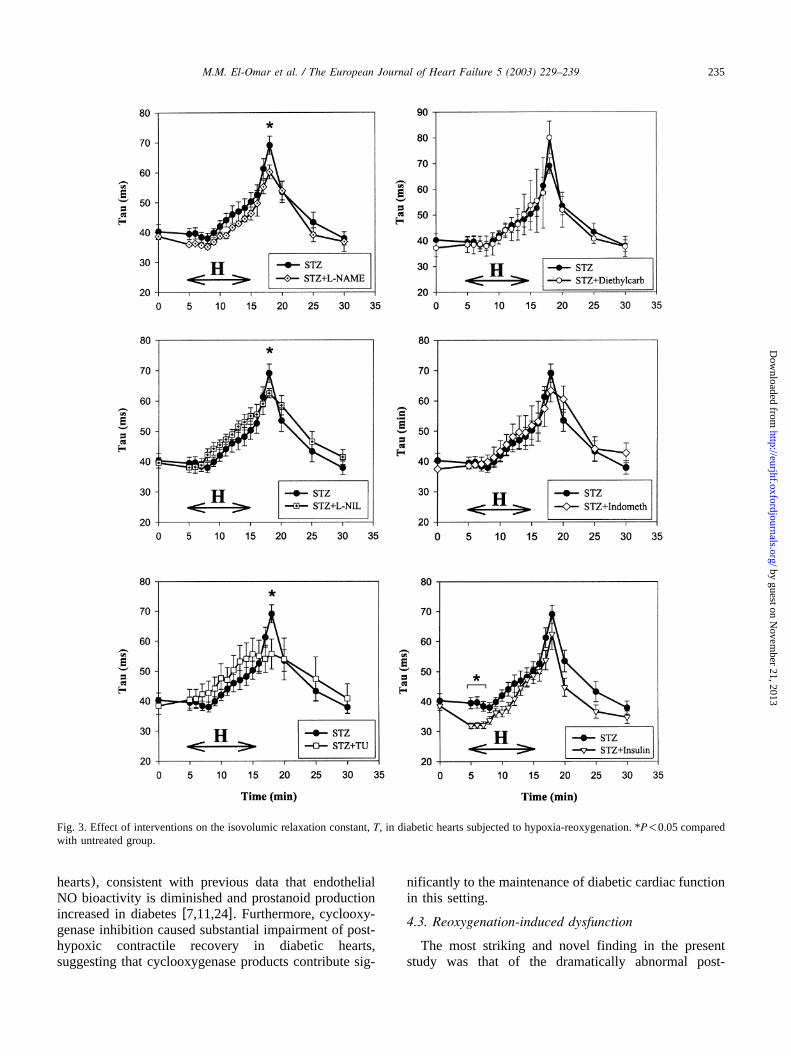
3.6. Effects of NOS inhibitors and other agents on theresponse to hypoxia-reoxygenation
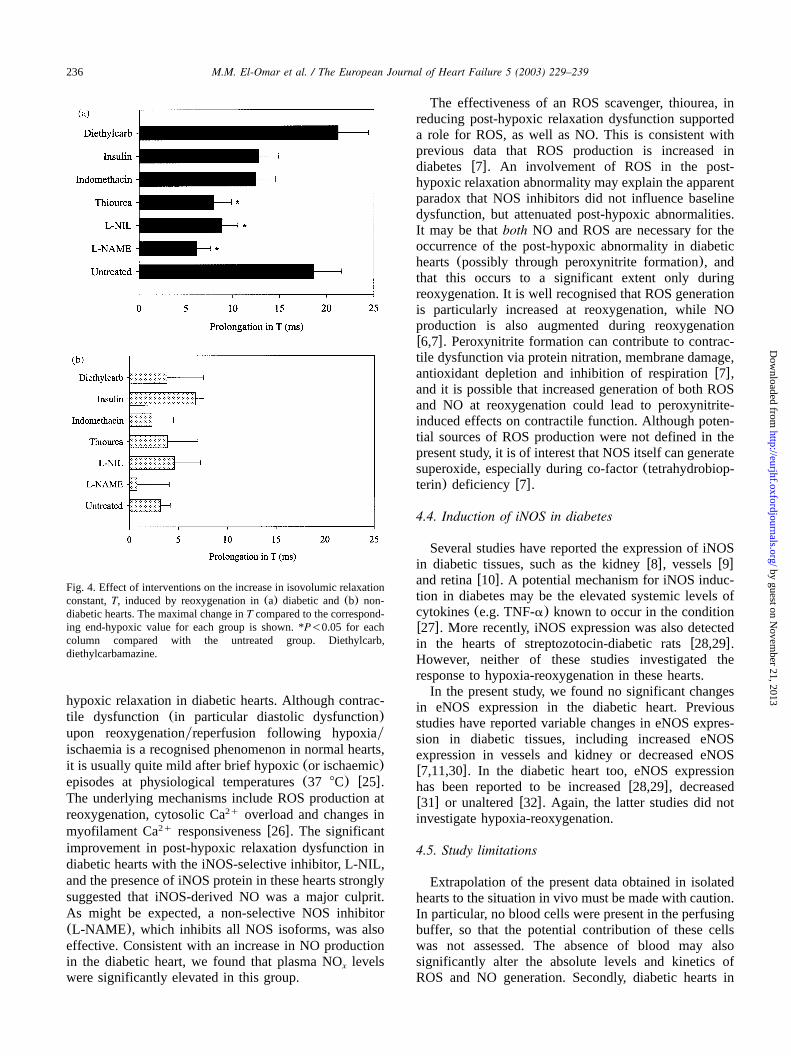
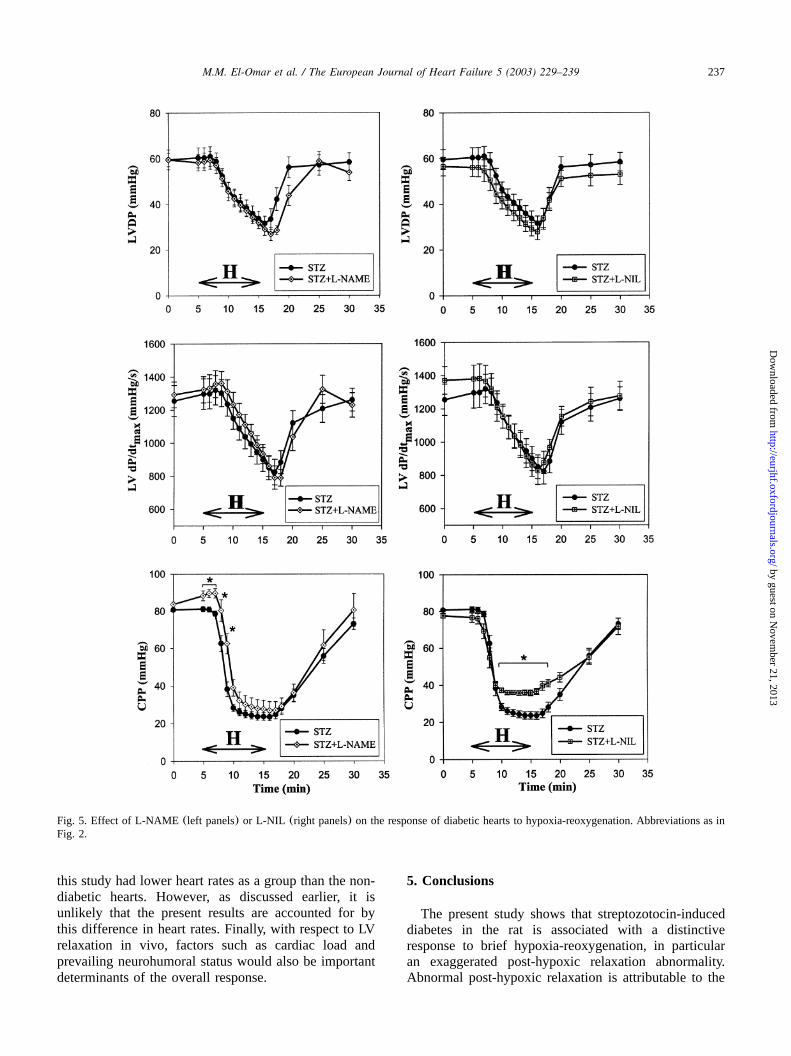
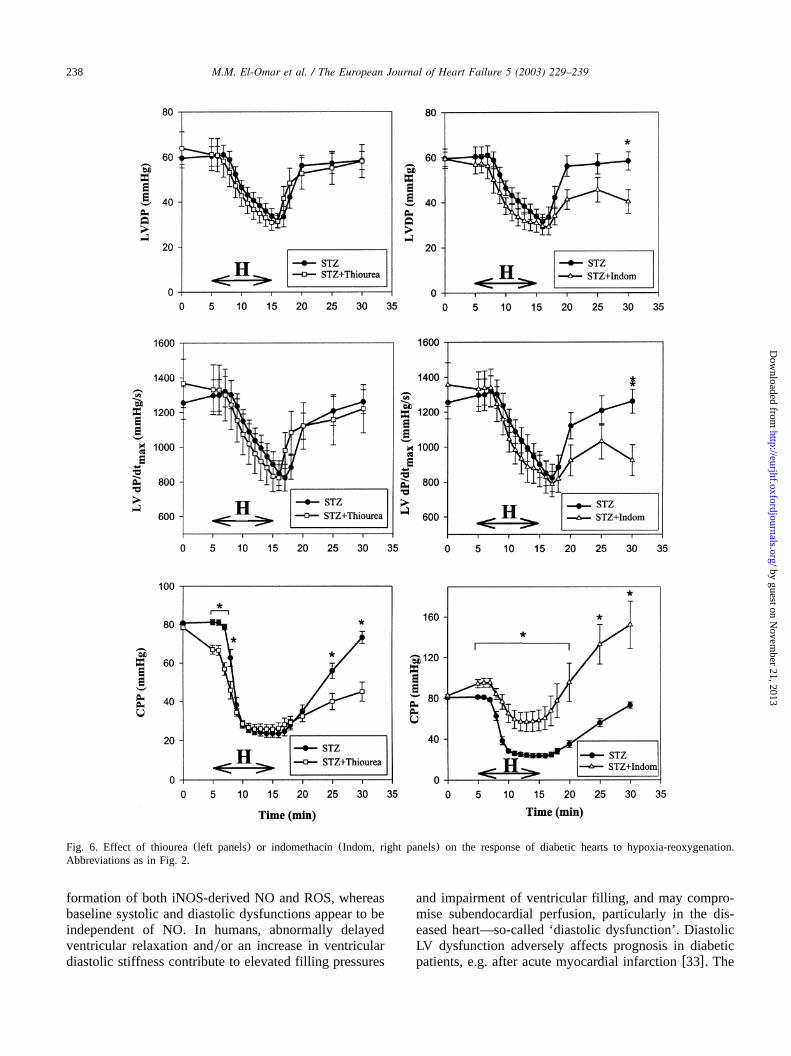
In the presence of L-NAME, L-NIL or thiourea, themarked prolongation ofT upon reoxygenation of dia-betic hearts was significantly attenuated(Figs. 3 and 4).The improvement in LV relaxation with these agentsoccurred in the absence of any significant effects onLVDP or LV dPydt (Figs. 5 and 6). None of themax
other agents tested significantly affected the post-hypox-ic relaxation abnormality of diabetic hearts(Figs. 3 and4). Of note, post-hypoxic prolongation ofT was unal-tered in the insulin-treated diabetic group(Fig. 3), andremained significantly greater than non-diabetic(con-trol) hearts despite the improvement in pre-hypoxic LVrelaxation. Interestingly, in the presence of indometha-cin, hypoxic coronary vasodilatation was significantlyblunted in diabetic hearts, and this was associated withimpaired post-hypoxic recovery of LVDP and LV dPydt (Fig. 6).max
In control non-diabetic hearts, L-NAME, L-NIL or
thiourea had no significant effect onT (Fig. 4) or onLV contractile parameters upon reoxygenation(data notshown).
4. Discussion
The main new findings of this study were that:(1)streptozotocin-induced diabetic hearts exhibited a pro-found (although transient) post-hypoxic relaxationabnormality, which was attributable to both NO derivedfrom iNOS and to ROS; and(2) baseline dysfunctionof the diabetic heart(systolic impairment, reduced heartrate and slow LV relaxation) was not influenced byNOS inhibitors or a ROS scavenger. In addition, LVcontractile depression during hypoxia was blunted indiabetic hearts, a response again unaffected by NOSinhibition.
4.1. Baseline dysfunction in diabetes and role of NO
The baseline dysfunction observed in this study issimilar to that previously reported in many other studiesof streptozotocin-induced diabetesw2,3x. Abnormal myo-cardial relaxation is an early feature of diabetic cardio-myopathy, and several different underlying mechanismshave been postulated including: abnormalities of cardiacrepolarisation w15x, Ca handling w16x, Na yH2q q q
exchangew17x, myofilament Ca responsivenessw3x2q
and metabolismw18x.In the present study, we showed that, despite increased
expression of iNOS in the diabetic heart, the overpro-duction of neither NO nor ROS significantly contributedto baseline contractile dysfunction, at least acutely.Similarly, neither the cyclooxygenase nor lipooxygenasepathways were involved, since indomethacin and die-thylcarbamazine also had no effects on LV function.The lack of effect of NOS inhibitors on baseline dys-function, despite documented iNOS expression, mightappear surprising, but is consistent with several studiesin vitro and in humans in vivo, where NO derived fromiNOS was found to have little or no effect on basalfunction w4,5,19–21x. In some of these studies, aninfluence of NO onb-adrenergic inotropic responsive-ness was nevertheless demonstratedw5,19,20x. The pos-sibility that L-arginine availability may be rate-limitingis not consistent with the finding that NOS inhibitorsaltered CPP and that they had demonstrable effectsduring reoxygenation.It is notable that heart rates in the diabetic group
were slightly but significantly lower than the controlgroup. The possibility that this might have contributedto reduced systolic function in diabetic hearts shouldtherefore be considered. We believe this is unlikely forseveral reasons. First, although diabetic and controlhearts had different rates as a group, there was a
by guest on Novem
ber 21, 2013http://eurjhf.oxfordjournals.org/
Dow
nloaded from
234 M.M. El-Omar et al. / The European Journal of Heart Failure 5 (2003) 229–239
Fig. 2. Effect of 10-min hypoxia-reoxygenation in control(C) and diabetic(STZ) hearts. H denotes the period of hypoxia. CPP, coronary perfusionpressure; LVDP, LV developed pressure;T, time constant for isovolumic relaxation. *P-0.05.
considerable overlap between them and there was nosignificant correlation between heart rate and LV dPydt . Secondly, baseline function was acutely improvedmax
by insulin independent of heart rate, suggesting that amajor component of contractile dysfunction during nor-moxia may be primarily metabolic. Thirdly, it haspreviously been shown that a substantial proportion ofcontractile dysfunction in the diabetic heart is intrinsicto the cardiac myocyte, independent of heart ratew2,3x.The abnormal myocardial relaxation observed at baselinein diabetic hearts is also unlikely to be related to lowerheart rates, since relaxation in factworsened at higherheart rates(not shown).
4.2. Response to brief hypoxia
Hypoxia-induced LV contractile depression was foundto be significantly blunted in diabetic hearts compared
to controls, implying increased tolerance to hypoxia.Similar findings have previously been reported, althoughsome studies have also reported increased sensitivity ofthe diabetic heart to ischaemia(for review, seew22x).The precise definition of the mechanisms underlyingthis effect was not the focus of the present study.However, we were able to demonstrate that the bluntedresponse to hypoxia of diabetic hearts was unaltered inthe presence of either L-NAME or L-NIL. This findingis of interest, given the evidence of iNOS expression inthe diabetic heart and recent data that iNOS expressioncan increase the tolerance to ischaemic insults(e.g. inischaemic myocardial preconditioning) w23x.
The other notable observation during hypoxia wasthat hypoxic coronary vasodilatation in diabetic heartsappeared to depend more on cyclooxygenase productsthan on NO (as opposed to the situation in control
by guest on Novem
ber 21, 2013http://eurjhf.oxfordjournals.org/
Dow
nloaded from
235M.M. El-Omar et al. / The European Journal of Heart Failure 5 (2003) 229–239
Fig. 3. Effect of interventions on the isovolumic relaxation constant,T, in diabetic hearts subjected to hypoxia-reoxygenation. *P-0.05 comparedwith untreated group.
hearts), consistent with previous data that endothelialNO bioactivity is diminished and prostanoid productionincreased in diabetesw7,11,24x. Furthermore, cyclooxy-genase inhibition caused substantial impairment of post-hypoxic contractile recovery in diabetic hearts,suggesting that cyclooxygenase products contribute sig-
nificantly to the maintenance of diabetic cardiac functionin this setting.
4.3. Reoxygenation-induced dysfunction
The most striking and novel finding in the presentstudy was that of the dramatically abnormal post-
by guest on Novem
ber 21, 2013http://eurjhf.oxfordjournals.org/
Dow
nloaded from
236 M.M. El-Omar et al. / The European Journal of Heart Failure 5 (2003) 229–239
Fig. 4. Effect of interventions on the increase in isovolumic relaxationconstant,T, induced by reoxygenation in(a) diabetic and(b) non-diabetic hearts. The maximal change inT compared to the correspond-ing end-hypoxic value for each group is shown. *P-0.05 for eachcolumn compared with the untreated group. Diethylcarb,diethylcarbamazine.
hypoxic relaxation in diabetic hearts. Although contrac-tile dysfunction (in particular diastolic dysfunction)upon reoxygenationyreperfusion following hypoxiayischaemia is a recognised phenomenon in normal hearts,it is usually quite mild after brief hypoxic(or ischaemic)episodes at physiological temperatures(37 8C) w25x.The underlying mechanisms include ROS production atreoxygenation, cytosolic Ca overload and changes in2q
myofilament Ca responsivenessw26x. The significant2q
improvement in post-hypoxic relaxation dysfunction indiabetic hearts with the iNOS-selective inhibitor, L-NIL,and the presence of iNOS protein in these hearts stronglysuggested that iNOS-derived NO was a major culprit.As might be expected, a non-selective NOS inhibitor(L-NAME), which inhibits all NOS isoforms, was alsoeffective. Consistent with an increase in NO productionin the diabetic heart, we found that plasma NO levelsx
were significantly elevated in this group.
The effectiveness of an ROS scavenger, thiourea, inreducing post-hypoxic relaxation dysfunction supporteda role for ROS, as well as NO. This is consistent withprevious data that ROS production is increased indiabetes w7x. An involvement of ROS in the post-hypoxic relaxation abnormality may explain the apparentparadox that NOS inhibitors did not influence baselinedysfunction, but attenuated post-hypoxic abnormalities.It may be thatboth NO and ROS are necessary for theoccurrence of the post-hypoxic abnormality in diabetichearts(possibly through peroxynitrite formation), andthat this occurs to a significant extent only duringreoxygenation. It is well recognised that ROS generationis particularly increased at reoxygenation, while NOproduction is also augmented during reoxygenationw6,7x. Peroxynitrite formation can contribute to contrac-tile dysfunction via protein nitration, membrane damage,antioxidant depletion and inhibition of respirationw7x,and it is possible that increased generation of both ROSand NO at reoxygenation could lead to peroxynitrite-induced effects on contractile function. Although poten-tial sources of ROS production were not defined in thepresent study, it is of interest that NOS itself can generatesuperoxide, especially during co-factor(tetrahydrobiop-terin) deficiencyw7x.
4.4. Induction of iNOS in diabetes
Several studies have reported the expression of iNOSin diabetic tissues, such as the kidneyw8x, vesselsw9xand retinaw10x. A potential mechanism for iNOS induc-tion in diabetes may be the elevated systemic levels ofcytokines(e.g. TNF-a) known to occur in the conditionw27x. More recently, iNOS expression was also detectedin the hearts of streptozotocin-diabetic ratsw28,29x.However, neither of these studies investigated theresponse to hypoxia-reoxygenation in these hearts.In the present study, we found no significant changes
in eNOS expression in the diabetic heart. Previousstudies have reported variable changes in eNOS expres-sion in diabetic tissues, including increased eNOSexpression in vessels and kidney or decreased eNOSw7,11,30x. In the diabetic heart too, eNOS expressionhas been reported to be increasedw28,29x, decreasedw31x or unalteredw32x. Again, the latter studies did notinvestigate hypoxia-reoxygenation.
4.5. Study limitations
Extrapolation of the present data obtained in isolatedhearts to the situation in vivo must be made with caution.In particular, no blood cells were present in the perfusingbuffer, so that the potential contribution of these cellswas not assessed. The absence of blood may alsosignificantly alter the absolute levels and kinetics ofROS and NO generation. Secondly, diabetic hearts in
by guest on Novem
ber 21, 2013http://eurjhf.oxfordjournals.org/
Dow
nloaded from
237M.M. El-Omar et al. / The European Journal of Heart Failure 5 (2003) 229–239
Fig. 5. Effect of L-NAME (left panels)or L-NIL (right panels) on the response of diabetic hearts to hypoxia-reoxygenation. Abbreviations as inFig. 2.
this study had lower heart rates as a group than the non-diabetic hearts. However, as discussed earlier, it isunlikely that the present results are accounted for bythis difference in heart rates. Finally, with respect to LVrelaxation in vivo, factors such as cardiac load andprevailing neurohumoral status would also be importantdeterminants of the overall response.
5. Conclusions
The present study shows that streptozotocin-induceddiabetes in the rat is associated with a distinctiveresponse to brief hypoxia-reoxygenation, in particularan exaggerated post-hypoxic relaxation abnormality.Abnormal post-hypoxic relaxation is attributable to the
by guest on Novem
ber 21, 2013http://eurjhf.oxfordjournals.org/
Dow
nloaded from
238 M.M. El-Omar et al. / The European Journal of Heart Failure 5 (2003) 229–239
Fig. 6. Effect of thiourea(left panels) or indomethacin(Indom, right panels) on the response of diabetic hearts to hypoxia-reoxygenation.Abbreviations as in Fig. 2.
formation of both iNOS-derived NO and ROS, whereasbaseline systolic and diastolic dysfunctions appear to beindependent of NO. In humans, abnormally delayedventricular relaxation andyor an increase in ventriculardiastolic stiffness contribute to elevated filling pressures
and impairment of ventricular filling, and may compro-mise subendocardial perfusion, particularly in the dis-eased heart—so-called ‘diastolic dysfunction’. DiastolicLV dysfunction adversely affects prognosis in diabeticpatients, e.g. after acute myocardial infarctionw33x. The
by guest on Novem
ber 21, 2013http://eurjhf.oxfordjournals.org/
Dow
nloaded from

239M.M. El-Omar et al. / The European Journal of Heart Failure 5 (2003) 229–239
possible contribution of myocardial iNOS expression toischaemic and post-ischaemic contractile dysfunction indiabetic patients merits investigation.
Acknowledgments
Supported by the British Heart Foundation and theUK Medical Research Council. ME was supported by aBritish Heart Foundation(BHF) Junior Research Fel-lowship and NJD by a UK Medical Research CouncilPhD studentship. AMS holds the BHF Chair of Cardi-ology at King’s College London. We are very gratefulto Dr Phil James for undertaking the plasma NO assays.x
References
w1x Sowers JR, Epstein M, Frohlich ED. Diabetes, hypertension,and cardiovascular disease: an update. Hypertension2001;37:1053–9.
w2x Francis GS. Diabetic cardiomyopathy: fact or fiction? Heart2001;85:247–8.
w3x Dhalla NS, Liu X, Panagia V, Takeda N. Subcellular remodel-ing and heart dysfunction in chronic diabetes. Cardiovasc Res1998;40:239–47.
w4x Kelly RA, Balligand JL, Smith TW. Nitric oxide and cardiacfunction. Circ Res 1996;79:363–80.
w5x Shah AM, MacCarthy PA. Paracrine and autocrine effects ofnitric oxide on myocardial function. Pharmacol Ther2000;86:49–86.
w6x Xie YW, Wolin MS. Role of nitric oxide and its interactionwith superoxide in the suppression of cardiac muscle mito-chondrial respiration. Involvement in response to hypoxiayreoxygenation. Circulation 1996;94:2580–6.
w7x Li H, Forstermann U. Nitric oxide in the pathogenesis ofvascular disease. J Pathol 2000;190:244–54.
w8x Cosenzi A, Bernobich E, Bonavista M, Trevisan R, Bellini G,Campanacci L. Early effects of diabetes on inducible nitricoxide synthase in the kidney. Acta Diabetol 2002;39:91–6.
w9x Bardell AL, MacLeod KM. Evidence for inducible nitric-oxidesynthase expression and activity in vascular smooth muscle ofstreptozotocin-diabetic rats. J Pharmacol Exp Ther2001;296:252–9.
w10x El-Asrar AM, Desmet S, Meersschaert A, Dralands L, Missot-ten L, Geboes K. Expression of the inducible isoform of nitricoxide synthase in the retinas of human subjects with diabetesmellitus. Am J Ophthalmol 2001;132:551–6.
w11x Hink U, Li H, Mollnau H, et al. Mechanisms underlyingendothelial dysfunction in diabetes mellitus. Circ Res2001;88:E14–E22.
w12x Weiss JL, Frederiksen JW, Weisfeldt ML. Hemodynamic deter-minants of the time-course of fall in canine left ventricularpressure. J Clin Invest 1976;58:751–60.
w13x Shah AM, Mebazaa A, Yang ZK, et al. Inhibition of myocardialcrossbridge cycling by hypoxic endothelial cells: a potentialmechanism for matching oxygen supply and demand? CircRes 1997;80:688–98.
w14x Misko TP, Schilling RJ, Salvemini D, Moore WM, Currie MG.A fluorometric assay for the measurement of nitrite in biolog-ical samples. Anal Biochem 1993;214:11–6.
w15x Wang DW, Kiyosue T, Shigematsu S, Arita M. Abnormalitiesof K and Ca currents in ventricular myocytes from ratsq 2q
with chronic diabetes. Am J Physiol 1995;269:H1288–H1296.w16x Pierce GN, Russell JC. Regulation of intracellular Ca in the2q
heart during diabetes. Cardiovasc Res 1997;34:41–7.w17x Feuvray D. The regulation of intracellular pH in the diabetic
myocardium. Cardiovasc Res 1997;34:48–54.w18x Stanley WC, Lopaschuk GD, McCormack JG. Regulation of
energy substrate metabolism in the diabetic heart. CardiovascRes 1997;34:25–33.
w19x Hare JM, Givertz MM, Creager MA, Colucci WS. Increasedsensitivity to nitric oxide synthase inhibition in patients withheart failure: potentiation of beta-adrenergic inotropic respon-siveness. Circulation 1998;97:161–6.
w20x Paulus WJ, Kastner S, Pujadas P, Shah AM, Drexler H,Vanderheyden M. Left ventricular contractile effects of induc-ible nitric oxide synthase in the human allograft. Circulation1997;96:3436–42.
w21x Cotton JM, Kearney MT, MacCarthy PA, et al. Effects of nitricoxide synthase inhibition on basal function and the force–frequency relationship in the normal and failing human heartin vivo. Circulation 2001;104:2318–23.
w22x Feuvray D, Lopaschuk GD. Controversies on the sensitivity ofthe diabetic heart to ischemic injury: the sensitivity of thediabetic heart to ischemic injury is decreased. Cardiovasc Res1997;34:113–20.
w23x Bolli R, Manchikalapudi S, Tang XL, et al. The protectiveeffect of late preconditioning against myocardial stunning inconscious rabbits is mediated by nitric oxide synthase. Evi-dence that nitric oxide acts both as a trigger and as a mediatorof the late phase of ischemic preconditioning. Circ Res1997;81:1094–107.
w24x Rosen P, Schror K. Increased prostacyclin release from perfusedhearts of acutely diabetic rats. Diabetologia 1980;18:391–4.
w25x Weisfeldt ML, Armstrong P, Scully HE, Sanders CA, DaggettWM. Incomplete relaxation between beats after myocardialhypoxia and ischemia. J Clin Invest 1974;53:1626–36.
w26x Piper HM, Garcia-Dorado D, Ovize M. A fresh look atreperfusion injury. Cardiovasc Res 1998;38:291–300.
w27x Rabinovitch A. An update on cytokines in the pathogenesis ofinsulin-dependent diabetes mellitus. Diabetes Metab Rev1998;14:129–51.
w28x Smith JM, Paulson DJ, Romano FD. Inhibition of nitric oxidesynthase by L-NAME improves ventricular performance instreptozotocin-diabetic rats. J Mol Cell Cardiol 1997;29:2393–402.
w29x Stockklauser-Farber K, Ballhausen T, Laufer A, Rosen P.Influence of diabetes on cardiac nitric oxide synthase expres-sion and activity. Biochim Biophys Acta 2000;1535:10–20.
w30x Onozato ML, Tojo A, Goto A, Fujita T, Wilcox CS. Oxidativestress and nitric oxide synthase in rat diabetic nephropathy:effects of ACE and ARB. Kidney Int 2002;61:186–94.
w31x Felaco M, Grilli A, De Lutiis MA, et al. Endothelial nitricoxide synthase(eNOS) expression and localization in healthyand diabetic rat hearts. Ann Clin Lab Sci 2001;31:179–86.
w32x Joffe II, Travers KE, Perreault-Micale CL, et al. Abnormalcardiac function in the streptozotocin-induced non-insulin-dependent diabetic rat: non-invasive assessment with Dopplerechocardiography and contribution of the nitric oxide pathway.J Am Coll Cardiol 1999;34:2111–9.
w33x Jacoby RM, Nesto RW. Acute myocardial infarction in thediabetic patient: pathophysiology, clinical course and prognosis.J Am Coll Cardiol 1992;20:736–44.
by guest on Novem
ber 21, 2013http://eurjhf.oxfordjournals.org/
Dow
nloaded from





























