Embed Size (px)
Citation preview

DIVERSITY OF MARINE MEIOFAUNA ON THE COAST OF BRAZIL
Saccocirridae (Annelida) from the southern and southeasternBrazilian coasts
Maikon Di Domenico & Alejandro Martínez &
A. Cecilia Z. Amaral & Paulo da Cunha Lana &
Katrine Worsaae
Received: 5 November 2013 /Revised: 3 February 2014 /Accepted: 4 February 2014# Senckenberg Gesellschaft für Naturforschung and Springer-Verlag Berlin Heidelberg 2014
Abstract Pharyngocirrus gabriellae (Du Bois-ReymondMarcus 1946) and Saccocirrus pussicus Du Bois-ReymondMarcus 1948 are redescribed based on newly collected mate-rial from the south and southeastern Brazilian coasts. Neo-types are designated, since no type material was formallyindicated in the original descriptions. New morphologicalinformation on the muscular ventral pharynx, ciliation, chae-tae and pygidium is provided. Pharyngocirrus gabriellae isadditionally characterised by the presence of mouth ciliarypatches, capillary lyrate chaetae with a single median toothand equally bifid medium chaetae with denticulate prongs.The novel diagnostic characters of Saccocirrus pussicus
include the absence of a pharynx and the presence of 1–3pygidial, adhesive ridges and 20 fertile segments. The twospecies live in very distinct interstitial habitats, with varyinghydrodynamics. Pharyngocirrus gabriellae is found at shel-tered intertidal and shallow areas in sediments with a redoxlayer, whereas Saccocirrus pussicus prefers the swash zone ofexposed, oxygenated, reflective beaches. Morphology andhabitat preferences of P. gabriellae and S. pussicus are consis-tent with other Saccocirridae species from the ‘Pharyngocirrus’and ‘Saccocirrus’ genera, respectively.
Keywords Araçá Bay . Southwest Atlantic . Interstitial .
Ecology .Meiofauna . Taxonomy
Introduction
Saccocirridae consists of 22 nominal species belonging toSaccocirrus Bobretzky 1872 and Pharyngocirrus DiDomenico, Martínez, Lana andWorsaae 2014, geographicallywidespread (Jouin 1971; Brown 1981; Jouin and Rao 1987;Purschke and Jouin 1988; Bailey-Brock et al. 2003; Jouin andGambi 2007; Di Domenico et al. 2014). The genusSaccocirrus consists of ten species and is defined by thesynapomorphies: presence of bilateral gonads, forked andspatulate chaetae, as well as absence of ventral muscularpharynx and ventral ciliation. The remaining 12 describedsaccocirrids, all in the genus Pharyngocirrus, have unilateralgonads, ventral muscular pharynx, anterior ventral ciliation, aswell as lyrate and bifid chaetae (Di Domenico et al. 2014).
Two species of saccocirrids are currently known from theBrazilian coast. Pharyngocirrus gabriellae (Du Bois-Reymond Marcus, 1946), with pharynx and ventral ciliation,is included in the ‘krusadensis’ clade, which one was describedas a new genus (Pharyngocirrus), based on a recent phyloge-netic analysis (Di Domenico et al. 2014). Saccocirrus pussicus
M. Di Domenico : P. da Cunha LanaBenthic Ecology Laboratory, Centre for Marine Studies, FederalUniversity of Paraná, Av. Beira-mar, s/n, P.O. Box 61,83255-976 Pontal do Sul, Pontal do Paraná, PR, Brazil
P. da Cunha Lanae-mail: [email protected]
M. Di Domenico :A. Martínez :K. WorsaaeMarine Biological Section, University of Copenhagen,Strandpromenaden 5, 3000 Helsingør, Denmark
A. Martíneze-mail: [email protected]
K. Worsaaee-mail: [email protected]
M. Di Domenico (*)Zoological Museum “Prof. Dr. Adão José Cardoso”, BiologicalInstitute, University of Campinas (UNICAMP), R Charles Darwins/n, Bloco N, P.O. Box 6109, 13083-863 Campinas, SP, Brazile-mail: [email protected]
A. C. Z. AmaralDepartamento de Biologia Animal (Zoologia), Instituto de Biologia,Universidade Estadual de Campinas, R. Monteiro Lobato, 255,13083-862 Campinas, SP, Brazile-mail: [email protected]
Mar BiodivDOI 10.1007/s12526-014-0208-5

Du Bois-Reymond Marcus, 1948 is included in the‘papillocercus’ clade, despite the record of a ventral pharynxin the original description (Di Domenico et al. 2014). Since theoriginal description, saccocirrids have only been recorded fromBrazil in three ecological surveys carried out at Paraná, SantaCatarina (Di Domenico et al. 2009) and Rio de Janeiro (Omenaet al. 2012). More recently the family was also recorded inUruguay (Rodriguez et al. 2013)
We redescribe herein P. gabriellae and S. pussicus basedupon newly collected material from a 4-year survey along2,000 km of the Brazilian coast. Several morphological detailsare added to the original description of P. gabriellae, regardingciliary patterns and chaetal morphology. The diagnosis ofS. pussicus is emended, since the original description unfortu-nately included a combination of characters of S. pussicussensuDuBois-ReymondMarcus, 1948 andP. gabriellae (pers.obs.). The habitat preferences of each species are described andneotypes are designated since type-material was not designatedor formally deposited in the original description.
Material and methods
Localities
Eleven sites were sampled between 20° S to 27° S, and 42° Wto 49° W in the Southwestern Atlantic (Fig. 1), along theBrazilian coastline. Material from the following beaches wasused for morphological description: Enseada de Zimbros (Coveof Zimbros), Estaleiro, Estaleirinho, and Barra Velha in SantaCatarina State; Mansa Beach in Paraná State; Araçá bay,Toque-Toque and Feiticeira in São Paulo, and Recreio dosBandeirantes, Barra da Tijuca and Leme in Rio de Janeiro State.The beaches Estaleiro, Estaleirinho, Barra Velha, Toque-Toque,Feiticeira, Recreio dos Bandeirantes and Leme are exposed orsemi-exposed to swells from the east and southeast. Theydisplay steep slopes, medium-coarse sandy sediments, absenceof redox layers and high hydraulic flux in the swash zone. Coveof Zimbros, Mansa beach and Araçá bay are sheltered, withshelly medium-coarse sandy sediments, gentle slopes, redoxlayer and low hydraulic flux. All the sampled localities areaffected by microtidal regimes with discontinuous semidiurnalperiods and mean amplitude of 0.8 m (Schettini et al. 1999;Carvalho et al. 1996). Tidal regimes are occasionally influencedby storms, which can raise the sea level ca. 1 m above astro-nomical tides (Carvalho et al. 1996; Schettini et al. 1999).
Routines for morphological observations
Sediment samples were collected by shovel in the intertidaland by SCUBA in the subtidal zone. Animals were extractedby MgCl2 decantation technique using a 63-μm mesh(Higgins and Thiel 1988). Live animals were photographed
and video recorded with an Olympus SZH microscope with amounted Canon PowerShot S45 camera. Material fixed formicroscopy were relaxed in MgCl2 and afterwards fixed ineither 2 % glutaraldehyde in cacodylate buffer (24 h, subse-quently transferred to 0.1 M cacodylate buffer), or fixed in2 % paraformaldehyde with 0.1 M sucrose (24 h at 4 °C,subsequently rinsed 6–7 times in PBS over 6 h and stored inPBS buffer with 0.2 M sucrose and 0.01 % NaN3) or fixed inBouin’s fixative (24 h, subsequently transferred to 70º ethanolthrough a gradient series).
Light microscopy (LM) observations were performed onspecimens fixed in Bouin’s fixative or in 2% paraformaldehyde.Specimens were permanently mounted in glycerol and exam-ined, measured and photographed with an Olympus DP71 cam-era mounted on an Olympus BX50 microscope at the MarineBiological Section, University of Copenhagen (MBS, UC).
Scanning electron microscopy (SEM) observations wereperformed on material fixed in 2 % glutaraldehyde. Materialwas post-fixed for 60 min in 1 % osmium tetroxide (in 0.1 Mcacodylate buffer), rinsed in distilled water, dehydratedthrough a graded ethanol-series, transferred to 100 % acetoneand critical-point dried. Dried specimens were then mountedon aluminium stubs, sputter-coated with platinum and exam-ined with a JEOL JSM-6335 F field emission scanning elec-tron microscope at the Natural History Museum of Denmark,University of Copenhagen (NHMD, UC). Type material wasdeposited at theMuseu de Zoologia daUniversidade Estadualde Campinas “Adão José Cardoso”, Brazil (ZUEC POL14053–14099); and at the Natural History Museum, Denmark(ZMUC-POL-2297-2299).
Sediment analyses
Sediment analyses were performed from 100-g sediment sam-ples collected at each locality. Samples were dried for 24 h at60ºC. Granulometric analyses were performed following stan-dard methods (Buchanan 1984). Sediment parameters wereobtained with the RYSGRAN package for R (R DevelopmentCore Team 2012), following the method of McCammon(1962). Classification is based on Wentworth (1922).
Additional material examined for morphological comparisons
Several species of Saccocirridae were examined for compar-ison at the United States National Museum/Smithsonian In-stitute (USNM), including type material of Pharyngocirrus(Saccocirrus) alanhongi Bailey-Brock, Dreyer and Brock,2003 (USNM POLY 1012494–1012497), P. eroticus Gray1969 (USNM POLY 36064–36066); P. jouinae Brown 1981(USNM POLY 62027–62033); Saccocirrus oahuensisBailey-Brock, Dreyer and Brock, 2003 (USNM POLY1012490–1012491); P. tridentiger Brown 1981 (USNMPOLY 62034–62040); S. waianaensis Bailey-Brock, Dreyer
Mar Biodiv

and Brock, 2003 (USNM POLY 1012492–1012493);P. sonomacus Martin, 1977 (USNM POLY 53050–53052).Additional material included newly collected specimens ofP. sonomacus Martin, 1977 from California, USA (collectedby K.W.); P. tridentiger Brown 1981 and P. jouinae Brown,1981 from New South Wales, Australia (courtesy of Greg W.Rouse); P. krusadensis Alikunhi, 1942, from New SouthWales, Australia and Phuket, Thailand (courtesy of Greg W.Rouse and Katharina Jörger); and S. papillocercus,Bobretzky,1871 from the Black Sea (courtesy of Elena Mastepanova).
Results
Morphological descriptions
Family SACCOCIRRIDAE Czerniavsky, 1881Genus Pharyngocirrus Di Domenico, Martínez, Lana and
Worsaae, 2014
See diagnosis after Di Domenico, Martínez, Lana andWorsaae, 2014
Pharyngocirrus gabriellae (Du Bois-Reymond Marcus,1946) emended
Figures 2, 3. Table 1.
Reference
Du Bois-Reymond Marcus, 1946 (Saccocirrus gabriellae);Wu and Yang 1962 (Saccocirrus major); Buzhinskaja 1967(Saccocirrus gabriellae); Scarlato et al. 1967 (Saccocirrusgabriellae); Sveshnikov 1967 (Saccocirrus gabriellae);Mastepanova 2004 (Saccocirrus gabriellae)
Neotype
Male, 6.0 mm long (whole mount ZUEC-Pol 14053), AraçáBay, São Sebastião, São Paulo, Brazil (23°48'56.05"S,45°24'31.51"W), intertidal between rocks in tidal pools.
Fig. 1 Map of study area showing sampled sandy beaches in the south and southeast of Brazil (Southwest Atlantic)
Mar Biodiv

Medium-coarse (Φ=0.34–1.14) poorly-moderately sorted(Φ=1.2–0.8) sandy sediments with redox layer. November07, 2012, Col: M. Di Domenico.
Additional material
Ten whole mount (ZUEC-Pol 14054–14063), Araçá Bay, SãoSebastião, São Paulo, Brazil (23°48'56.05"S, 45°24'31.51"
W), intertidally between rocks in tidal pools. Medium-coarse(Φ=0.34–1.14) poorly-moderately sorted (Φ=1.2–0.8) sandysediments with redox layer. Three specimens mounted onSEM stubs (ZMUC-Pol-2297), Enseada de Zimbros, SantaCatarina, Brazil (27°12'04.27"S, 48º29'57.32" W), shelteredbeach and between rocks in tidal pools, coarse (Φ=0.4-0.1)and moderately sorted (Φ=0.9–0.75) sand, redox layer pres-ent, intertidal to 3 m depth. Five specimens mounted on SEM
Fig. 2 Pharyngocirrus gabriellae light microscopy images. a Wholeanimal view, juvenile, dorsal view. b Anterior end, dorsal view. c Pha-ryngeal bulb, lateral view. d Lateral view of parapodium, showing chae-tae. e Pygidium, fixed paratype and whole-mounted, ventral view, dorsal
view. f Pharyngeal bulb, lateral view. Abbreviations: aog anterior gut, esoesophagus, ls longest chaeta, ic internal canal, mo mouth, pa palp, phpharynx, pp parapodium, pr prostomium, pyl pygidial lobe, pyp pygidiumadhesive ridges, se1 chaetiger 1
Mar Biodiv

stubs (ZMUC-Pol-2298), Farol Island in Mansa beach,Matinhos, Paraná, Brazil (25º51'02"S, 48º32'11.6" W), shel-tered beach and between rocks in tidal pools, medium tocoarse (Φ=0.34–1.14) poorly to moderately sorted (Φ=1.2-0.8) sand, redox layer present, intertidal. Five whole mount(ZUEC-Pol, 14064–14068), Ponta do Baleeiro beach SãoSebastião, São Paulo, Brazil (26°38'09"S 48°41'04"W), be-tween rocks in tidal pools, medium to coarse (Φ=0.2–1.30)poorly to moderately sorted (Φ=1.6-0.8) sand, redox layerpresent, intertidal to 3 m depth. Several days in 2011 to 2013,Col: M. Di Domenico, A.L. Prantoni.
Emended diagnosis
Brown body with 45–160 segments; last three smaller andachaetous. Prostomium with two pigmented eyes and longfiliform palps. Mouth surrounded by ciliary patches. Ventralmuscular pharynx present. Uniramous parapodia with threetypes of chaetae: (1) 1–2 long capillary chaetae, lyrate with asmall median tooth; (2) 2–3 medium bifid chaetae, with equallateral prongs and (3) 1–2 short simple notched chaetae.Females with unilateral ovaries at the right side of the gut,from segments 29–35 to 130–140, each ovary with 15–20oocytes and banana-shaped spermathecae. Males with unilat-eral seminal vesicles at the left side of the gut, each bearing anon-ciliated penial tube. Pygidial lobes with 6–10 transverseadhesive ridges.
Description
(Measurements from the neotype; ranges from adults inparentheses).
Body long, robust and brownish (Fig. 2a), 6 mm long (4–6 mm, n=6) and 150 μm wide (135–200 μm, n=6), up to 49segments (47–60, n=6) (Fig. 2a, b), last three smaller andachaetous.
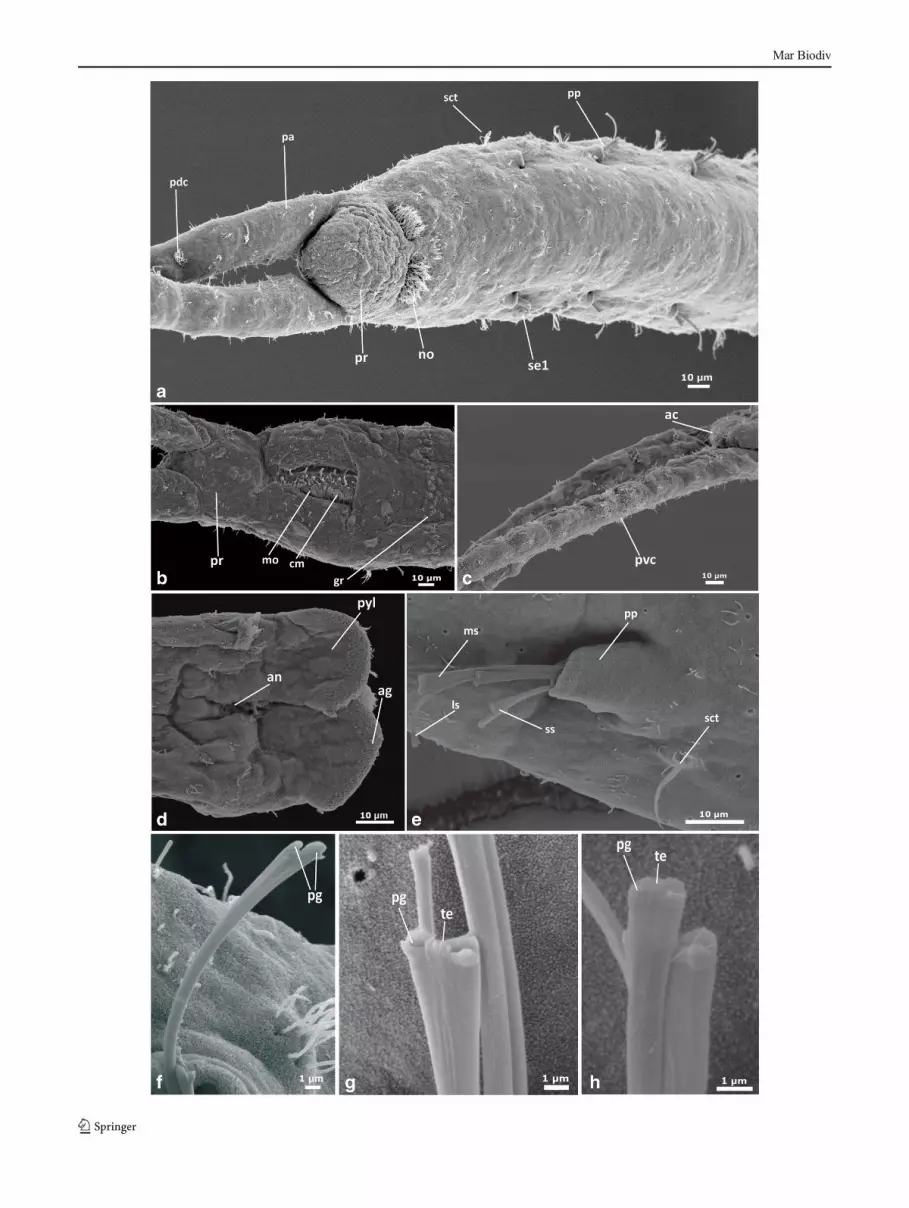
Prostomium rounded, 50 μm long (50–80, n=6) (pr,Fig. 2a), with two dorsal pigmented eyes. Palps 600 μm long(450–800 μm, n=6), hollow and slightly annulated, as long asthe first eight trunk segments. Palp internal channels connect-ed to oval tentacular ampulla, 90 μm long (80–120 μm, n=5)(Fig. 2), extending posteriorly. Palp ciliation consisting ofscattered non-motile tufts of short cilia, sometimes organisedas loose bands (pvc, pdc, Fig. 3c, d). Dorsal nuchal organsdensely ciliated, 20 μm long, arranged in furrows betweenprostomium and peristomium (Fig. 3a, b), (15–30 μm, n=5).Paired longitudinal ciliary bands, 10–15 μm long, extendingfrom the nuchal organs to the anterior end on each side of theprostomium (tc, Fig. 3b).
Peristomium 100 μm long (64–115, n=4) and 100 μmwide (90–170, n=4), with a ventral longitudinal slit-shapedmouth (54 μm, n=1, SEM). Mouth lined by two longitudinalciliary patches (mp, Fig. 3c) and provided with an internal
ciliated ridge (cm, Fig. 3b). Ciliation on the peristomium andtrunk otherwise consists of scattered tufts of short, non-motilecilia (length of cilia, 2 μm long, n=2). Mouth continues into aglandular oesophagus, extending along the three first trunksegments. Ventral muscular pharynx present, 200 μm long(180–240 μm, n=4, LM) (Fig. 2b).
Parapodia cylindrical, unirramous and retractile, projectinglaterally at each body segment (except last two segments), upto 50μm long (40–60, n=4) (pp, Figs. 2d, 3e). Each parapodiawith three types of chaetae: (1) 1–2 long capillary chaetae (ca.1.5 longer than the medium chaetae), thin and lyrate (2.5 μmwide at the tip n=2) (ls, Fig. 3i, h), with one short mediantooth only visible on the SEM (pg, Fig. 3i); (2) 2–3 bifidmedium chaetae, 3 μm wide terminally (n=2, SEM) withtwo equal lateral prongs (ms, Fig. 3f) provided with 2–3small denticles (te, Fig. 3f); (3) 1–2 thin short chaetae,notched terminally (1 μm wide, n=3, SEM) (ss, Fig. 3g).A small ciliary tuft with ca. 10–15 cilia (n=2, length ofthe cilia, 10 μm, n=2), ventrally on each parapodium (sct,Fig. 3a).
Pygidium with two lobes (pyl, Figs. 2e, 3j), 150 μm long(120–200, n=4, LM) and 55 μm wide (45–65, n=4), eachwith 6–10 ventral adhesive ridges (ag, Fig. 3j). Anus openingterminally between the lobes (an, Figs. 2e, 3j).
Mature description sensu Du Bois-Reymond Marcus(1946). Females with ca. 30 fertile segments with ovaries fromsegment 29–35 to segment 130–140. Each segment with 15–20 mature oocytes, at left sides of the gut. Spermathecae arebanana-shaped. Seminal vesicle in the left side of the gut, withnon-ciliated penial tube.
Taxonomical remarks
Pharyngocirrus gabriellae is included into the Pharyngocirrusgenus (‘krusadensis’ clade) after the morphology of the chae-tae (longest chaetae lyrated, medium chaetae bifid), presenceof unilateral gonads, pharyngeal bulb and ventral ciliation.Within the genus, P. gabriellae resembles P. sonomacus andP. eroticus by the presence of equal bifid chaetae and ventralciliation limited to the mouth area. However, Pharyngocirrusgabriellae differs from these two species by the presence ofmature oocytes at the left side of the gut (oocytes at the rightside of the gut in P. sonomacus and P. eroticus) and lowernumber of fertile segments.
Du Bois-Reymond Marcus (1946) provided a diagno-sis for Pharyngocirrus gabriellae in her original de-scription, with all original diagnostic features confirmedby our own observations. Our description herein pro-vides novel information on the arrangement of ventralmouth ciliation and morphology of the chaetae (mediantooth in the long capillary chaetae, denticulation in themedium chaetae). We also provide further ecologicalinformation, highlighting the preference of P. gabriellae
Mar Biodiv

Mar Biodiv

for sheltered beaches and tidal areas, with medium tocoarse, in poorly to moderately sorted sandy bottomswith a marked redox layer.
Justification for the neotype
No type material was designated in the original description ofPharyngocirrus gabriellae (Du Bois-Reymond Marcus1946), a necessary procedure for the formal definition of aspecies (Article75, ICZN). Some of the original material maystill remain at the Zoological Museum of São Paulo andZoological Museum of Natural History of Montevideo, butdeposit numbers were not provided and so the collectionsrelated to the original material cannot be searched unequivo-cally. No further records of P. gabriellae from the type localityare available. Our sampling at the type locality did not yieldspecimens of P. gabriellae, possibly as a consequence of theanthropogenic modifications at the beach in the past 70 years.A neotype for the species, collected at Araçá Bay (SãoSebastião), 200km from the type locality is herein formallyerected. The redescription of the species will serve as a base-line against the sibling species of P. gabriellae (Wu and Yang1962; Buzhinskaja 1967; Scarlato et al. 1967; Sveshnikov1967; Mastepanova 2004), which can now formally be de-scribed and contrasted with the type species.
Genus Saccocirrus Bobretzky, 1872 See diagnosis afterBobretzky, 1872 Saccocirrus pussicus Du Bois-ReymondMarcus 1948 Emended
(Figures 4, 5 and Table 2)
References
(Du Bois-Reymond Marcus 1948; Di Domenico et al. 2009;Rodriguez et al. 2013)
Neotype
Female, 6.5 mm long (ZUEC-Pol 14069). Praia da Península,Barra Velha, Santa Catarina, Brazil (26°38'09"S, 48°41'04"W). Swash zone, medium-coarse sand (Φ=0.9–1.16), moder-ately well sorted (Φ=0.5–0.56), intertidal. Date: 10/09/2010Col: M. Di Domenico.
Additional material
Three specimens mounted on SEM stubs (ZMUC-Pol, 2299)and 10 whole mounts (ZUEC-Pol, 14070–14079), same lo-cality as neotype. 15 whole mounts (ZUEC-Pol, 14080–14094), Praia de Estaleiro, Santa Catarina, Brazil (27°01'49"S, 48°34'48"W), swash zone, medium to coarse (Φ=0.34–1.14) moderately to well-sorted sand (Φ=0.4–1.0), 0–1 mdepth. Five whole mounts (ZUEC-Pol, 14095–14099),Estaleirinho beach Santa Catarina, Brazil (27°02'59"S,48°35'12"W), swash zone, medium to coarse (Φ=0.34–1.14) moderately to well-sorted (Φ=0.4–1.0); 0–1 m depth.Specimens are identified and fixed from, Praia da Feiticeira,Ilha Bela, São Paulo, Brazil (23°50'43"S, 45°24'32"W), invery coarse (Φ=0.3–0.5) well-sorted (Φ=0.2–0.4) sand at 0–1 m depth in swash zones; Praia do Toque-Toque, SãoSebastião, São Paulo (23º51'04"S, 45º29'27" W). Reflectivebeach, medium sand (Φ=1.6–2.1) moderately well sorted(Φ=0.6–0.8), intertidal; Recreio dos Bandeirantes Río deJaneiro, Brazil (23°01'46"S, 43°27'57"W), swash zone,coarse sand (Φ=0.5 -0.2) and moderately well-sorted sand(Φ=0.5-0.6), intertidal; Barra da Tijuca, Rio de Janeiro, Brazil(23°00'51"S, 43°22'23"W), swash zone, medium sand (Φ=0.9–1.6) and moderately well-sorted sand (Φ=0.5-0.6), inter-tidal; and Leme, Rio de Janeiro, Brazil (22°57'51"S,43°10'01"W), swash zone, medium-coarse sand (Φ=0.7–1.3), moderately sorted (Φ=0.75-0.9). Several days in 2010–2012, Col: M. Di Domenico, R. Barroso and C.A.M. Barboza.
Table 1 Measurements ofPharyngocirrus gabriellae fromneotype and whole mounted typematerial, compared with mea-sures from the literature. Average(Aver), maximum (Max), mini-mum (Min), number of individ-uals (N), Du Bois-ReymondMarcus (1946) (Marcus)
Marcus Neotype Aver Max Min N
Number of segment 160 49 60 90 47 6
Length (mm) 30 6 5 6 4 6
Width (µm) 400 150 140 200 135 6
Palps (µm) 1,000 600 600 800 450 6
No. of chaetigerous segments – 46 50 60 40 6
No. of segment without chaetae – 3 2 3 2 6
Prostomium length (um) – 50 60 80 50 6
�Fig. 3 Pharyngocirrus gabriellae, scanning electron microscopicimages. a Anterior end animal view, dorsal view. b Lateral view of andprostomium. c Ventral view of the palps and anterior end of prostomium.d Ventral view of prostomium and palps. e Lateral view of parapodium,showing chaeta. f Details of medium chaeta. g Details of short chaeta. hDetails of longest chaeta. i Details of longest chaeta. j Pygidium, dorsalview. Abbreviations: ac apical cilia, ag adhesive glands, an anus, cmciliated mouth, cp palp cilia, gr glandular ridge, ls longest chaetae, msmedium chaetae, mo mouth open, mp mouth ciliary patches, no nuchalorgan, pa palp, pc palp canal, pdc palp dorsal ciliation, pg prong, ppparapodium, pr prostomium, pvc palp ventral ciliation, pyl pygidiumlobe, se1 chaetiger 1, s setae, sct segmental ciliary tuff, ss short chaetae,tc transversal ciliation, te teeth
Mar Biodiv

Fig. 4 Saccocirrus pussicus lightmicroscopic images. a Wholeanimal view of neotype, male,dorsal view. b Anterior end,dorsal view. c Glandularoesophagus, fixed paratype andwhole-mounted, internal view. dContent in midgut, fixed paratypeand whole-mounted, internalview. e Bilateral seminal vesiclein a male (segments 35–38),dorsal view. f Seminal vesiclewith sperm and penis on liveparatype, dorsal view. g Bilateraleggs in a female on live paratype,dorsal view. h Pygidium, fixedparatype and whole-mounted,dorsal view. Arrows indicatesepta. Abbreviations: ag anteriorgut, an anus, eg egg, mg midgut,gc gut contents, go glandularoesophagus, oc oceli, p penis, papalp, pc palp canal, prprostomium, py pygidium, ssetae, sct segmental ciliary tuff,se21 setiger 21, se35 setiger 35,se38 setiger 38, sr supportingrods, sv seminal vesicle, sv1 firstsegment with seminal vesicle,sv20 20th segment with seminalvesicle
Mar Biodiv

Emended diagnosis
Body brownish, with 40–80 segments; last 8 smaller andachaetous. Prostomium with two pigmented eyes and longfiliform palps. First segment without ciliary patches. Pharynxabsent. Parapodia with three types of chaetae: (1) 1–2 longchaetae, robust and forked with equal prongs; (2) 2–3 mediumspatuled chaetae, with 4 minute teeth; (3) 2–3 short spatuledchaetae, with notched apex. Paired pygidial lobes with 1–3transverse adhesive ridges. Females with bilateral ovaries insegments 19–50 (along 78 segments), each with 2–3 largeoocytes. Spermathecae absent. Males with bilateral seminalvesicles in segments 21–40 (along 70 segments).
Emended description
(Measurements provided from neotype; ranges of all adultparatypes in parentheses).
Body long, robust and brownish (Fig. 4a), 6.5 mm long (4–14 mm, n=19) and 295 μm wide (140–300 μm, n=19), up to56 segments (39–78, n=19). Last 8 segments (6–12 segmentsin adults, n=19) smaller and achaetous.
Prostomium rounded, 110 μm long (45–125, n=25) and125 μmwide (60–140, n=7), with two dorsal pigmented eyes(Fig. 4b), 20 μm in diameter (15–25, n=04). Paired palps,annulated and hollow, 1.03 mm long (560–1,200 μm, n=20),as long as the first six trunk segments. Palp internal channelsconnected to oval tentacular ampullae, 105 μm long (80–120 μm, n=5), extending posteriorly along the peristomium.Palp without motile cilia, but abundant non-motile ciliary tufts(cp, Fig. 5a) scattered on the entire palp surface. Paired ovalnuchal organs, densely ciliated, dorsally between the prosto-mium and peristomium (no, Fig. 5a), 45 μm long (36–50 μm,n=5). A short ciliary band, 10–15 μm long, extendstransversally 5–10 μm apart from the nuchal organs in thedorso-anterior surface of the prostomium (pr, Fig. 5a).
Peristomium 105 μm long (65–115, n=4, LM) and 165 μmwide (95–175, n=4, LM), with a ventral slit-shaped mouth(Fig. 3b, mo). Mouth extends longitudinally (55 μm, n=1,SEM), lined by lateral and posterior lips (Fig. 5b) and provid-ed of an internal ciliated ridge (Fig. 5b, cm). Mouth cavitycontinues as a glandular oesophagus, extending along thethree first trunk segments. Muscular pharyngeal bulb absent(Fig. 4c). Ciliation on the peristomium limited to scatteredgroups of non-motile cilia.
Paired cylindrical retractile parapodia, projecting laterallyat each body segment (except the last seven), up to 55μm long(25–55, n=10) (pp, Fig. 5e). All parapodia uniramous, withthree types of chaetae: 1–2 longest, robust and forked chaeta(ls, Fig. 5e, f); 2–3 spatuled medium chaetae (ms, Fig. 5e, g)and 1–2 spatuled short chaetae (ss, Fig. 5e, h). Long chaetaethin and bifurcated in two short equal prongs, only visible onthe SEM (pg, Fig. 5f), 2.5 μmwide at the tip (n=2) and ca. 1.5
times longer than the medium chaetae. Medium chaetaespatulated, 3 μmwide terminally (n=2, SEM) with two lateralprongs equally long (1 μm wide, n=2), separated 1 μm apart.Four small teeth between the two lateral prongs (te, Fig. 5g,0.25 μm long, n=3; 0.10 μm wide, n=3, SEM), slightlylonger than the lateral prongs. Short chaetae thin, notchedterminally (1 μm wide, n=3, SEM) (Fig. 5h). Trunk ciliationrestricted to an individual tuft of ca. 10 cilia (length of thecilia, 10 μm, n=2), ventrally to each parapodia (sct, Fig. 5e)and small ciliary tufts of ca. 3–5 cilia, scattered on the entirebody surface (length of cilia, 2 μm; n=2). A band consistingof two rows ca. 25 groups of three cuticular small poresarranged close together (pores, ca. 1 μm in diameter; bands,5 μm wide, n=3), slightly raised from the body surface,extends ventrally on each segment (gr, Fig. 5b). These poresprobably correspond to epidermal glandular structures, thelatter but not the former identifiable with light microscopy.
Pygidium with two lobes (pyl, Figs. 4h, 5d), 120 μm long(90–130, n=4, LM) and 55 μmwide (45–65, n=4), each with1–3 ventral adhesive ridges (ag, Fig. 5d). Anus opening ter-minally between pygidial lobes, without distinct ciliation (an,Fig. 4h, 5d).
Females with ca. 20 fertile segments (max. 31, n=9), fromsegment 21 (20–23, n=9) to 41 (25–50, n=9). Each fertilesegment with 5–6 mature oocytes (eg, Fig. 4j), 100 μm indiameter (70–120, n=7), at both sides of the gut.Spermathecae absent. Males with 18 fertile segments, fromsegment 21 (18–23, n=10) to 37 (24–42, n=10). Seminalvesicle (sv, Fig. 4e, f), 100 μm diameter, at both sides of thegut, with terminal pencil-shaped penis (p, Fig. 4f), 500 μmlong (400–550, n=6, LM), 80 μm wide (70–90, n=6, LM)and provided with supporting rods (sr, Fig. 4f).
Juveniles 1-6 mm long (n=35) and 120–180 mmwide (n=35), up to 50 segments (last 7, smaller and achaetous). Matu-ration starts occurring when the juveniles extend 4 mm inlength.
Taxonomic remarks
Saccocirrus pussicus is included into the Saccocirrus genus(‘papillocercus’ clade) based on the morphology of the chae-tae (longest chaetae robust and forked, medium chaetaespatulated), presence of bilateral gonads and absence of pha-ryngeal bulb. Within the genus, it shows closest resemblanceto S. papillocercus Bobretzky, 1871 and S. heterochaetusJouin 1975. It differs from S. papillocercus (Jouin andGambi 2007) in the smaller number of segments, number ofpygidial ridges, fertile segments in both sexes; larger and lessabundant oocytes per segment and shorter body. Saccocirruspussicus differs from S. heterochaetus in the presence of largerand more abundant (six versus four) eggs per segment andabsence of hooks, characteristic of the fertile region ofS. heterochaetus.
Mar Biodiv
Mar Biodiv

In the original publication, Du Bois-Reymond Marcus(1948) describes Saccocirrus pussicus with 120 segments,muscular pharynx, 6–12 pygidial adhesive transversal ridgesand gonads from segments 24–36 to segments 45–73.
The specimens here identified as Saccocirrus pussicus(emended) differ from S. pussicus sensu Du Bois-Reymond Marcus, 1948 in the absence of muscular ventralpharynx, shorter body, smaller number of segments (78versus 120 segments in S. pussicus sensu Du Bois-Reymond Marcus 1948), smaller number of pygidial ridges(three versus 6–12 in S. pussicus sensu Du Bois-ReymondMarcus 1948), and less fertile segments (20 versus 36S. pussicus sensu Du Bois-Reymond Marcus 1948). Nev-ertheless, our specimens resemble S. pussicus sensu DuBois-Reymond. Marcus 1948 in the morphology of males’seminal vesicles, at both sides of the gut, with terminalpencil-shaped penis, number of oocytes (4–5) per segmentin females and the three types of chaetae (1–2 long, robustand forked chaetae, 2–3 spatulated medium chaetae and 1–2 thin spatulated short chaetae).
Further ecological information is also provided.Saccocirrus pussicus (papillocercus clade) occurs at ex-posed reflective beach, at swash zones until wave breaking
zone with medium to coarse sand and absence of redox (DiDomenico et al. 2014) similar to the habitat reported by DuBois-Reymond Marcus (1948).
Justification for the neotype
In the original description, Du Bois-Reymond Marcus attrib-uted to Sacoccirrus pussicus a combination of features fromboth S. pussicus (as here re-described) and S. gabriellae.Unfortunately, the type material from S. pussicus is lost,preventing revision of that species. However, after a intensive4-year survey around the localities and habitats reported in theoriginal description, none of the collected Saccocirrus spp.presented the same combination of characters attributed toS. pussicus by Du Bois-Reymond Marcus (1948), therebyjustifying the redescription of the species.
The neotype for Saccocirrus pussicus is designated fromBarra Velha, Santa Catarina, the locality which yielded mostof the material here reported, including abundant maturespecimens. Barra Velha is also in the vicinity of permanentresearch facilities, favouring future collections of fresh mate-rial for further research. The designation of a neotype willserve as a baseline from which the sibling species ofSaccocirrus pussicus can be formally described andcontrasted with the type species (Rodriguez et al. 2013).
Discussion
The results presented here confirm the presence of two speciesof Saccocirridae in the South and Southern coast of Brazil, aspreviously reported by Du Bois-Reymond Marcus more than60 years ago. The low diversity of saccocirrids in this areacontrasts with the abundance of species of other interstitial
Table 2 Measurements of Saccocirrus pussicus from neotype and whole-mounted type material. Average (Aver), maximum (Max), minimum (Min) andnumber of individuals (N) measured for males, females and juveniles
Males Females Juveniles
Neotype Aver. Max. Min. N. Aver. Max. Min. N. Aver. Max. Min. N.
Number of segments 56 59 70 39 10 61 78 44 9 35 60 20 35
Length (mm) 6.5 9 14 4 10 6 9 5 9 3 6 1 35
Width (µm) 295 217 300 170 7 206 300 140 7 140 222 100 19
Palps (µm) 1028 899 1200 630 10 869 1100 560 9 585 1188 270 35
No. of segments chaetigerous 48 50 63 28 10 52 71 38 9 28 53 16 35
No. of segment without chaeta 8 9 12 7 10 9 15 6 9 7 11 4 35
Gametes start 19 21 23 18 10 21 23 20 9 – – – –
Gametes end 35 37 42 24 10 41 50 25 9 – – – –
No. of fertile segments 16 18 21 7 10 20 31 2 9 – – – –
Prostomium length (µm) 112 90 130 80 10 100 126 78 9 60 80 47 35
�Fig. 5 Saccocirrus pussicus, scanning electron microscopic images. aAnterior end animal view, dorsal view. b Ventral view of the mouth andprostomium. c Latero-dorsal view of the palps and anterior end ofprostomium. d Ventral view of pygidium. e Lateral view ofparapodium, showing chaeta. f Details of long chaeta. g Details ofmedium chaeta. h Details of small chaeta. Abbreviations: ac apicalcilia, ag adhesive glands, an anus, cm ciliated mouth, cp palp cilia, grglandular ridge, mo mouth open, no nuchal organ, pa palp, ppparapodium, pr prostomium, pyl pygidium lobe, s setae, sct segmentalciliary tuff, se1 first setiger, se35 setiger 35, se38 setiger 38, sr supportingrods, sv seminal vesicle, sv1 first segment with seminal vesicle, sv20 20thsegment with seminal vesicle
Mar Biodiv

groups, such as Protodrilidae or Nerillidae (Di Domenico et al.2013; pers. obs.).
Despite their occurrence in the same overall geographicalarea, S. pussicus and P. gabriellae belong to very differentclades within Saccocirridae (Di Domenico et al. 2014). Basedon their morphological and molecular phylogeny, the closestrelative of S. pussicus is a new species from Italy that ismorphologically similar to S. major; whereas P. gabriellae isrelated to P. sonomacus from Chile and California (DiDomenico et al. 2014).
Within the same region, P. gabriellae and S. pussicus arenever found sympatrically, and both species exhibit clearlydistinct preferences for particular interstitial and hydrodynam-ic habitats. Pharyngocirrus gabriellae only occurs at shelteredbeaches, bays or coves, between rocks in tidal pools to 3 m, incoarse sandy sediments with a redox layer. In contrast,Saccocirrus pussicus exclusively inhabits the swash zone ofexposed reflective beach with medium to coarse sand and noredox layer (Di Domenico et al. 2009; Di Domenico,Martínez, Worsaae and Lana, pers. obs.).
This distinct habitat preference correlates with differentfeeding strategies. Pharyngocirrus gabriellae feeds on depos-ited phytoplankton, phytobenthos or macroalgal debris, oftenobserved along its gut (Du Bois-Reymond Marcus 1946),which they scrape with the mouth and muscular pharynx. Incontrast, S. pussicus is omnivorous and feeds on diatoms,copepods and debris, reported in the gut of most collectedspecimens (Du Bois-Reymond Marcus 1948, pers. obs.). In-terestingly, the habitat preferences and feeding strategies ofP. gabriellae and S. pussicus are shared by the remainingspecies in the each genera, respectively (Du Bois-ReymondMarcus 1946; 1948; Brown 1981; Bailey-Brock et al. 2003;Jouin and Gambi 2007; Di Domenico et al. 2014).
The biogeographic and ecological patterns exhibited bySaccocirridae, with major cosmopolitan clades exhibitingclear ecological preferences, is congruent with other familiesof interstitial annelids and indicates an early diversificationinto specific interstitial habitats (Di Domenico et al. 2013;Martínez et al. 2013). This specialisation into different inter-stitial microhabitats exhibited by several annelid familiesstresses the complexity of these habitats and highlights theimportance of ecology in the evolutionary history and diver-sification of interstitial lineages.
Acknowledgments We thank Alexandra Rizzo, Alvaro E. Migotto,Carlos Barboza, Cinthya S.G. Santos, Fabiane Gallucci, GustavoFonseca, Rômulo Barroso, and Tito C. M. Almeida for their generoushelp, advice and logistic support in the field trips. Alessandro L. Prantoni,André Pereira Cattani, Reginaldo Nunes andVeronica Oliveira weremosthelpful during field sampling. We thank our colleagues, technicians andstaff of the Marine Biological Section and Natural History Museum ofDenmark, University of Copenhagen for their generous help and advice.We further thank Greg W. Rouse and Katharina Jörger for providingcomparative material from Australia and Thailand, respectively. Thisstudy was supported by the Brazilian National Council for Technological
and Scientific Development (CNPq—Process 140611/2008-8), whichprovided the Ph.D. fellowship for M.D.D. São Paulo Research Founda-tion (FAPESP—Process 2012/08581-0, Process 2013/04358-7) providedpostdoctoral fellowships and grants for M.D.D.We are highly indebted toDr. JonNorenburg, KathrynAhlfeld and Cheryl Bright, from the NationalMuseum of Natural History/Smithsonian Institute, Washington DC, forkindly hosting M.D.D. at the museum to check the relevant type materialof Saccocirrus spp and Pharyngocirrus spp.
References
Bailey-Brock JH, Dreyer J, Brock RE (2003) Three new species ofSaccocirrus (Polychaeta: Saccocirridae) from Hawai’i. Pacific Sci57:463–478
Brown R (1981) Saccocirridae (Annelida: Archiannelida) from the cen-tral coast of New South Wales. Aust J Mar Freshw Res 32:439–456
Buchanan JB (1984) Sediment Analysis. In: Holme NA, McIntyre AD(eds)Methods to Study the Marine Benthos, vol 16, IPHHandbook.Blackwell Scientific Publications, London, pp 41–65
Buzhinskaja GN (1967) On the ecology of the polychaetous annelids ofthe Possjet Bay (Sea of Japan). Issled Fauny Morei 13:78–125 (InRussian)
Carvalho JLB, Klein AHF, Schettini CAF, Jabor PM (1996) Marésmeteorológicas em Santa Catarina: influência do vento nadeterminação de parâmetros de projetos para obras costeiras.Proceedings of III Simpósio sobre Oceanografia, São Paulo, p pp380
Di Domenico M, Lana PC, Garraffoni ARS (2009) Distribution patternsof interstitial polychaetes in sandy beaches of southern Brazil. MarEcol 30:47–62
Di Domenico M, Martínez A, Lana PC, Worsaae K (2013)Protodrilus (Protodrilidae, Annelida) from the southern andsoutheastern Brazilian coasts. Helgol Mar Res. doi:10.1007/s10152-013-0358-z
Di DomenicoM,Martínez A, Lana PC,Worsaae K (2014)Molecular andmorphological phylogeny of Saccocirridae (Annelida) reveals twocosmopolitan clades with specific habitat preferences. MolPhylogenet Evol. doi:10.1016/j.ympev.2014.02.003
Du Bois-Reymond Marcus E (1946) On a new archiannelid, Saccocirrusgabriellae, from Brazil. Comun Zool Mus Hist Nat Montev 37(2):1–11
Du Bois-Reymond Marcus E (1948) Further archiannelids from Brazil.Comun Zool Mus Hist Nat Montev 2:69–83
Gray JSA (1969) New species of Saccocirrus (Archiannelida) from theWest Coast of North America. Pac Sci 23:238–251
Higgins RP, Thiel H (1988) Introduction to study of meiofauna.Smithsonian Institution Press, Washington DC, pp 488
Jouin C (1971) Status of the knowledge of the systematics and ecology ofArchiannelida. Smithson. Contrib Zool 76:47–56
Jouin C, Gambi C (2007) Description of Saccocirrus goodrichi sp. nov.(Annelida: Polychaeta: Saccocirridae), a newMediterranean speciesand new data on the chaetae of S. papillocercus and S. major. CahBiol Mar 48:381–390
Jouin C, Rao GC (1987) Morphological studies on some Polygordiidaeand Saccocirridae (Polychaeta) from the Indian Ocean. Cah BiolMar 28:389–402
Martin GG (1977) Saccocirrus sonomacus sp. nov., a new archiannelidfrom California. Trans Am Microsc Soc 96:97–103
Martínez A, Di Domenico M, Jörger K, Norenburg J, Worsaae K (2013)Description of three new species of Protodrilus (Protodrilidae,Annelida) from Central America. Mar Biol Res. doi:10.1080/17451000.2013.765574
Mar Biodiv

Mastepanova EA (2004) Interstitial polychaetes of the seas of Russia. InvZool 1(1):59–64 (in Russian)
McCammonRB (1962) Efficiencies of percentile measures for describingthe mean size and sorting of sedimentary particles. J Geol 70(4):453–465
Omena EP, Lavrado HP, Paranhos R, Silva TA (2012) Spatial distributionof intertidal sandy beach Polychaeta along an estuarine andmorphodynamic gradient in an eutrophic tropical bay. Mar PollutBull 64(9):1861–1873
Purschke G, Jouin C (1988) Anatomy and ultrastructure of the ventralpharyngeal organs of Saccocirrus (Saccocirridae) andProtodriloides (Protodriloidae fam. n.) with remarks on the phylo-genetic relationships within Protodrilida (Annelida: Polychaeta). JZool 215:405–432
R Development Core Team (2012) R: a language and environment forstatistical computing. R Foundation for Statistical Computing, Vienna,Austria. ISBN 3-900051-07-0, URL http://www.R-project.org/
Rodriguez M, Venturini N, Di Domenico M, Martinez AG, Worsaae K(2013) First occurrence of the interstitial polychaete Saccocirrus
pussicus in exposed beaches of Uruguay. Panam J Aquat Sci 8(3):194–198
Scarlato OA, Golikov AN, Vasilenko SV, Tzvetkova NL, Grusov EN,Nesis KN (1967) Composition, structure and distribution of bottombiocenoses in the coastal waters of the Possjet Bay (Sea of Japan).Issled Fauny Morei 13:5–62 (In Russian)
Schettini CAF, Carvalho JL, Truccolo E (1999) Aspectos hidrodinâmicosda enseada da Armação de Itapocoroy, SC. Notas Técnicas daFACIMAR 3:99–109
Sveshnikov VA (1967) Larvae of archiannelids and polychaetes ofthe Possjet Bay (Sea of Japan). Exploration of the Fauna ofthe Seas. Biocoenoses of the Possjet Bay, the Sea of Japan.Akademia Nauk SSSR, Zool. Inst., Moscow, pp 125–159 (inRussian)
Wentworth CK (1922) A scale of grade and class terms for clasticsediments. J Geol 30:377–392
Wu BL, Yang DJ (1962) The taxonomy and distribution of the genusSaccocirrusBobretzky (Archiannelida). (Russian abstract). OceanolLimnol Sin 4:169–179
Mar Biodiv





























