Embed Size (px)
Citation preview

Hum Genet (1984) 67 : 72-85
© Springer-Verlag 1984
Satellited Y chromosomes: Structure, origin, and clinical significance
M. Schmid 1, T. Haaf 1, Eva Solleder 1, W. Schempp 2, M. Leipoldt 3, and H. Heilbronner 4
Institut f'tir Humangenetik, Universit~it Wfirzburg, Koellikerstr. 2, D-8700 Wtirzburg, Federal Republic of Germany 2 Institut ffir Humangenetik, Universitgt Freiburg, Albertstr. 11, D-7800 Freiburg i. Br., Federal Republic of Germany 3 Institut fiat Humangenetik, Universit/it G6ttingen, Nikolausberger Weg 5a, D-3400 G6ttingen, Federal Republic of Germany 4 Institut f'dr Humangenetik, Universit/it Ttibingen, Wilhelmstr. 27, D-7400 TiJbingen, Federal Republic of Germany
Summary. Three cases of inherited satellited Y chromosomes (Yqs) were analysed using several cytogenetic techniques. The cytogenetic data of the 14 cases of Yqs chromosomes described to date were reviewed. All Yqs chromosomes carry an active nucleolus organizer region (NOR) in their long arm and must have developed from translocations involving the short arms of the acrocentric autosomes. The structure of the heterochro- matic satellite region in the Yqs chromosomes shows conspic- uous inter-familial differences; this permits the reconstruction of the translocations from which the various Yqs were derived. Some causal factors leading to the development of Yqs chro- mosomes are considered: the specific localization of the four satellite DNAs and highly methylated DNA sequences in the karyotype, and some new experimental data on the spatial arrangement of heterochromatic regions in interphase nuclei. These provide distinct evidence for a preferential involvement of the autosomes 15 and 22 in the translocations with the Y heterochromatin. All clinical reports documenting Yqs males born with malformations were reviewed. It appears that the presence of an extra NOR and NOR-associated heterochroma- tin in the Yqs chromosomes does not cause any phenotypic abnormalities (as long as the Y euchromatin is intact). The possibility that a Yqs chromosome predisposes to non-disjunc- tion and/or to translocations of other chromosomes is dis- cussed.
Introduction
The so-called satellited Ychromosomes (Yqs) belong to the very rare structural variations in the human karyotype. In this type of structural rearrangement, parts of the short arm of an acro- centric autosome are translocated at the long arm of the Y chromosome, causing its satellited appearance. The first case of such a Yqs chromosome was described by Genest et al. (1967) in a patient with Down syndrome. The chromosome analysis of this patient demonstrated a trisomy 21 and "a Y chromosome the size of a group-F (19-20) autosome, with achromatic filaments terminating in euchromatic small knobs attached to the end of the long arms. These structures could not be differentiated morphologically from satellites of group D or G autosomes" (Genest et al. 1967). This description of the appearance of the rearranged Yqs chromosome can on the
Offprint requests to: M. Schmid
whole be used for all the other Yqs chromosomes found hence. Furthermore, this peculiar satellited Y chromosome was traced back to the great-grandfather of the patient (Family A of Genest et al. 1967, Table 1).
Four teen further cases of Yqs chromosomes have been de- scribed in the meantime (Tables 1 and 2). In 13 of these cases the Yqs chromosome occurred within the family line (S chmid 1969; Genest 1972, 1973, 1978; Genest et al. 1983; Hayata et al. 1975, 1977; Howard-Peebles and Stoddard 1976; Howell et al. 1978; Stella et al. 1980; Giraldo et al. 1981; Bordson and Varela 1982; Bayless-Underwood et al. 1983) and in one case the trans- location occurred de novo (Turleau et al. 1978). The detailed pedigree analysis of the Family B of Genest (1973, 1978) and Genest et al. (1983) led to the surprising result that this Yqs chromosome has been transmitted in patrilineary fashion for 315 years ! The presence of a distinct achromatic constriction and terminal satellites in the Yqs chromosomes suggested that in the course of the structural rearrangement the nucleolus organizer region (NOR) was translocated from the short arm of an acrocentric chromosome onto the Y chromosome. An indi- cation for this was provided by the many close associations observed between the Yqs chromosomes and the nucleolar constrictions in the short arms of the acrocentric autosomes. The definitive confirmation of the fact that the achromatic constrictions in the Yqs chromosomes are indeed NORs was provided by staining techniques which specifically label the NORs (N-banding, silver (Ag)-staining).
In the present study, three new Yqs chromosomes are introduced. Several special cytogenetic techniques were used in order to characterize these Yqs chromosomes. All cases with Yqs chromosomes previously described in the literature were reviewed and the minor or major malformations found in some probands with a satellited Y chromosome are discussed. The factors contributing to the origin of the human Yqs chro- mosomes are reviewed, and the NOR-bearing sex chromo- somes observed in other mammalian species are taken into consideration.
Case reports
In two of the current cases the satellited Y chromosomes were demonstrated in the metaphases of the amniotic fluid cells in tile course of intrauterine diagnosis. Prenatal diagnosis was performed because of the advanced age of the parents. There were no indications for a chromosomal analysis provided by

73
Fig. la-s. Yqs chromosomes of Case B showing conventional Giemsa staining (a-e), C-bands (d-f), Ag-stained NORs (g-i), distamycin A-induced undercondensation of the Y heterochromatin (k-m), distamycin A/DAPI fluorescence (n-p), and quinacrine fluorescence (q-s). The upper row presents the Yqs from six metaphases each, the two lower rows show partial metaphases where the Yqs chromosomes (large arrows) participate in satellite associations with acrocentric autosomes. Note the distinct satellites and nucleolar constrictions (a), the large amount ofheterochroma- tin (d) and the conspicuous Ag-stained NORs (g). In the Yqs chromosomes from distamycin A-treated lymphocytes (k) a small segment located at the distal end of the heterochromatin (short arrows) escapes the induced undercondensation. The heterochromatin of this Yqs chromosome fluoresces brightly with distamycin A/DAPI (n) and quinacrine (q)
the family histories. The Yqs chromosomes were also demon- strated in the metaphases from blood lymphocytes from the fathers of these two male fetuses (Cases B and W in this report).
The third case is a 17-year-old male patient with growth retardation (153 cm body length, 61kg body weight). Aside from the retarded growth, the clinical examination presented no abnormalities. A satellited Y chromosome was found in the metaphases of the lymphocytes both from the patient (Case K in this report) as well as from his father.
Materials and methods
The chromosome preparations of Cases B, K, and W as well as five additional, unrelated normal males without Yqs chromo- somes were obtained from the peripheral blood according to the method of Pfeiffer (1974). Chromosome medium la (Gibco Bio-Cult) was used for the conventional cultures as well as for the cultures treated with distamycin A and 5-azacytidine (0.5 ml blood per 3.5 ml medium). The staining methods were as follows:
C-banding (Figs. ld-f , 3b, 4n) was done according to the method of Sumner (1972), and silver (Ag)-staining of the NORs (Figs. lg-i , 3c, 5d-f) according to the method of Goodpasture and Bloom (1975).
Q-banding (Figs. lq-s , 3d, 4f). Quinacrine mustard was used according to the technique of Caspersson et al. (1970). The fluorescence photomicroscopy was performed using a Zeiss fluorescence microscope equipped with an HBO 50W/AC
lamp and a filter combination 450-490/FT 510/LP 520. Every fluorescing metaphase of which a karyotype was to be prepared was photographed with three different exposure times; this permits the detection of the very tiny satellites on the Yqs chromosomes in those photographs with long exposure times (Overton et al. 1976).
Distamycin A/DAPI-staining (Figs. in-p , 3e, 4g). A simple modification of the original technique of Schweizer et al. (1978) was performed successfully. The slides were flooded with distamycin A (0.1 mg/ml in McIlvaine buffer, pH 7.0) and covered with a coverslip. After 15-min incubation in the dark at room temperature, the slides were rinsed in McIlvaine buffer and stained with DAPI (0.2 gg/ml in McIlvaine buffer) for 10 min. Finally, the preparations were rinsed in McIvaine buf- fer for 10 s and mounted in a saturated saccharose solution. For microscopic analysis, a 2 FL filter (Zeiss) was used in the fluo- rescence microscope.
Bromodeoxyuridine (BrdU)-replication patterns (Fig. 4a-e, h-l , o, p). For the demonstration of the replication patterns on metaphase chromosomes, 10 gg BrdU/ml and 0.5 gg fluoro- deoxyuridine (FdU)/ml were added to the blood cultures 7 h before harvest. The metaphase chromosomes were differen- tially stained with the Hoechst-Giemsa technique according to the method of Schempp and Meer (1983) and with acridine orange (Couturier et al. 1973).
DistamyeinA-in vitro treatment(Figs. 1k-m, 2a-e). The conden- sation of the Y heterochromatin was prevented by treating the blood cultures with the oligopeptide antibiotic distamycin A according to the technique of Schmid (1979).

74
Fig. 2a-e. Giemsa-stained Yqs chromosomes (large arrows) of Case B from distamycin A-treated lymphocytes, a Metaphase with diplo-chromo- somes; the inset shows a higher magnification of the Yqs diplo-chromosome in satellite association with a G-group diplo-chromosome. Note that a small segment at the distal end of both Yqs chromosomes escapes the distamycin A-induced undercondensation of the heterochromatin (small arrows), b-e Partial metaphases with multiple satellite associations between the Yqs chromosome and two to six acrocentric autosomes
Fig. 3a-f. Yqs chromosomes of Case K showing Giemsa staining (a), C-bands (b), Ag-stained NORs (e), quinacrine fluorescence (d), distamycin A/DAPI fluorescence (e), and 5-azacytidine-induced undercondensations (13. Note the relatively small amount ofheterochromatin (b) (compare with Fig. la, d). The heterochromatin of this Yqs chromosome shows bright distamycin A/DAPI fluorescence but no bright quinacrine fluores- cence (d). In metaphases from 5-azacytidine-treated lymphocytes (13 distinct undercondensations can be observed in the heterochromatin of chromosomes 1, 9, 15, 16, and Y; the chromosomes were previously identified by quinacrine banding. The inset in (I3 shows the Yqs chromo- somes from three metaphases; the small arrows indicate the position of the satellites, g, h Giemsa-stained Y chromosomes and autosomes 1, 9,15, and 16 from 5-azacytidine-treated lymphocytes of normal male probands without Yqs chromosomes. Note that the undercondensed heterochro- matin of these normally structured Y chromosomes (g) is distinctly longer than that of the Yqs chromosomes (13
5-Azacytidine-in vitro treatment (Figs. 3f-h, 5a-c). The under- condensation of the heterochromatin in the chromosomes Y, 1, 9, 15, and 16 as well as the demonstration of the somatic pairings between these heterochromatic chromosome regions was achieved with the cytidine analogue 5-azacytidine (Viegas- P6quignot and Dutril laux 1976; Schmid et al. 1983). 5-Aza- cytidine (Serva) was dissolved in twice-distilled water and 100 gl each of this solution was added to the blood cultures at 5 x 10 -7 to 2 x 10 -7 (final concentration in the culture) 7h before fixation of the cells.
Results
The satellited Y chromosomes of the three probands B, K, and W could be recognized in the conventionally stained prepara-
tions (Figs. 1a-c, 3a, 4m). In all three cases the satellites were always demonstrated in the long arms of the Y chromosome; the short arms never showed any indication of satellites. In the middle metaphase stage the long arms of the Y chromosomes exhibited the usual degree of close chromatid apposition characteristic for the human Y chromosome (Schmid 1967). Without the satellites and satellite stalks (nucleolar constric- tions) these Y chromosomes were slightly shorter than the group-F chromosomes (Fig. 4a, Table 1). Regarding the size of the satellites and the length of the satellite stalks, there were definite inter-individual differences among the three Yqs chromosomes. Both the satellites as well as the satellite stalks were distinctly larger in the Yqs ofproband B than they were in the Yqs of probands K and W (compare Figs. la, 3a, 4m). The satellites and satellite stalks were least pronounced in proband

75
Fig. 4. Yqs chromosomes of the Case W. a Karyotype with R-bands after BrdU treatment and acridine orange staining. All autosomes and the X chromosome exhibit their normal replication patterns. The inset shows a photographic overexposure of the Yqs chromosome; note the satel- lites at the distal end of the Yqs chromosome (small arrow) which replicate earlier than the terminal Y heterochromatin, b, e Three examples each of the two types of replication patterns in the long arms of the Yqs chromosomes and in (d) and (e) the only type found in the normally structured Y chromosomes of control probands for comparison. The same differences were found after differential staining with the Hoechst-Giemsa te ch- nique (h-l); the small arrows in (e) and (i) point to the satellites, f Q uinacrine-stained Yqs chromosomes without bright heterochromatin fluores- cence, g Distamycin A/DAPI staining showing distinctly labeled heterochromatin (large arrow), m Conventional Giemsa staining, n C-bands; note the very small nucleolar constrictions, satellites, and heterochromatic regions (small arrow), o, p Examples of satellite associations between the Yqs chromosome (large arrows) and acrocentric autosomes from BrdU-treated lymphocytes (Hoechst-Giemsa staining)
Fig. 5. a Giemsa-stained metaphase from a 5-azacytidine-treated lymphocyte of a normal male without Yqs chromosome. The chromosomes were previously identified by quinacrine banding. Note the distinct undercondensations of the heterochromatin in the chromosomes 1, 9,15,16, and Y as well as the somatic pairing between the undercondensed heterochromatin of an autosome 15 and the Y chromosome (arrow). b Selected examples of somatic pairing configurations between the undercondensed heterochromatin ofautosomes 15 and normal Y chromosomes found in 5-azacytidine-treated lymphocytes, e Somatic pairings between the undercondensed heterochromatin ofautosomes 1, 9, 16, and Y chromo- somes. The position of the chromosome arms is illustrated by the camera lucida drawings beneath the photographic representations. The under- condensed heterochromatic regions are always turned toward each other and in some cases are in direct contact, d-f Three lymphocyte nuclei from a normal male proband possessing a normally structured Y chromosome prestained with quinacrine (left column) and restained with silver (right column). Note that the Y heterochromatin is always closely associated with a cluster of Ag-positive NORs of the acrocentric autosomes (a~,ows)

76
O
o
"0
o
%
+ ~ + I + ~ + ~ + + I
4- ~ + I + 4- + + 4- + + 4- 4-
+ ~ + + + + + + + + + + +
+ + + + + + + + + + + + + +
4- + + • 4- + + + I + 4- + + +
A A V / ~ V V / V A V V V V
" ~ t ~ " ~ o ~ .
0 ©
o
.o o~
I:~ , ~ , - ~
- d N ~
~ g
c~
N u . . ,
~ . ~ , .~ . . , ~ ~ + ~

77
W (Fig. 4m); in approximately 30% of the metaphases analys- ed, these could not be identified.
The specific Ag-labeling of the NORs was always localized within the satellite stalks of the Yqs chromosomes and in no case extended into the adjacent heterochromatic regions (Figs. 1g-i, 3 c). This was especially distinct in the preparations which were prestained with quinacrine and subsequently treated with silver. In the short arms of the normally structured acrocentric chromosomes, the dark silver-label is also always localized within the satellite stalks (Goodpasture et al. 1976).
The C-banding technique enabled the demonstration of a distinct constitutively heterochromatic region in the long arm (band Yq12) in the Yqs chromosomes of all three probands (Figs. 1d-f, 3b, 4n). However, the size of this heterochromatic band showed remarkable inter-individual differences. In the Yqs chromosome of proband B the heterochromatic region constituted approximately two-thirds of the long arm (Fig. ld- f). By comparison, it encompassed one-half of the long arm in the Yqs of proband K (Fig. 3b) and only one-third of the long arm in the Yqs ofproband W (Fig. 4n). Differences among the three Yqs chromosomes could also be observed with quin- acrine-banding and distamycin A/DAPI-staining. Thus in the Yqs of proband B the brightly fluorescing heterochromatin in the distal two-thirds of the long arm could be demonstrated both with quinacrine and with distamycin A/DAPI (Fig. 1n-s). The quinacrine-bright heterochromatin in this Yqs consisted of two closely adjacent bands (Fig. 1q-s), which are well known from observations made on normal Y chromosomes (for re- view see Btihler 1980). In the Yqs chromosomes of the pro- bands K and W, only the distamycin A/DAPI-bright hetero- chromatin was present (Figs. 3e, 4g). No remnants of the quinacrine-positive heterochromatin could be seen even after the photographs were over-exposed (Figs. 3 d, 4f). This finding can be explained if the position of the distamycin A/DAN- and quinacrine-bright heterochromatin in the normally structured Y chromosomes is taken into account. Whereas quinacrine stains the terminal portion of the Y heterochromatin, the region fluorescing brightly with distamycin A/DAPI is situat- ed further proximally (Fig. 7a; W. Schnedl, personal commu- nication). This indicates that in the translocation leading to the Yqs chromosomes of probands K and W, the break occurred within the distamycin A/DAPI-bright heterochromatin; this resulted in the loss of the terminal, quinacrine-bright hetero- chromatin (Fig. 7c). In contrast to this, the break leading to the development of the Yqs ofproband B must have been localized within the quinacrine-bright heterochromatin; this led to the loss of only a small portion of the Y chromosome (telomeric region), which is confirmed by the presence of both categories of heterochromatin (Fig. 7d).
The in vitro treatment of the lymphocytes with 100 lag dista- mycin A/ml for the last 24 h of culture prevents the normal condensation of the entire constitutive heterochromatin in the long arm of the Y chromosome (Prantera et al. 1979; Schmid 1979). This undercondensed heterochromatin appears as a thin chromatin thread suspended at the normally condensed euchromatic region. It was noticed that in the Yqs chromo- somes a small segment localized at the distal end of the consti- tutive heterochromatin escapes the distamycin A-induced undercondensation (Fig. lk-m). These segments appear par- ticularly distinctly in metaphases with diplo-chromosomes, which are quite common in distamycin A-treated cultures (Fig. 2a). It must be assumed that these distamycin A-insensi- tive segments are the centromere-adjacent portions (band p11) of the short arms of the acrocentric chromosomes, which were
translocated onto the Y chromosomes along with the NORs and the satellites. A schematic representation of such a trans- location and the Yqs chromosome resulting from it are shown in Fig. 7d.
The cytidine analogue 5-azacytidine induces very distinct undercondensations in the constitutive heterochromatin of the Y chromosome and the autosomes 1, 9,15, and 16 if applied in very low doses during the last hours of culture (Viegas- Pdquignot and Dutrillaux 1976; Schmid et al. 1983). The under- condensed heterochromatin appears as long and only weakly stainable threads; the euchromatin of these chromosomes condenses in a normal fashion (Fig. 3f-h). Just as in the Yqs chromosomes from distamycin A-treated lymphocytes, very small, normally condensed segments could be recognized at the distal end of the undercondensed heterochromatin in the 5-azacytidine-treated Yqs as well (Fig. 3f). In addition, the length of the undercondensed heterochromatin in the Yqs of the probands K and W was distinctly shorter than in the Y chromosomes of the control probands (compare Fig. 3 f and g). The latter can be explained by the decreased amount of consti- tutive heterochromatin in these Yqs. In the metaphases from the 5-azacytidine-treated lymphocyte cultures, the undercon- densed heterochromatid regions of the chromosomes 1, 9, 15, 16, and Y exhibit very conspicuous somatic pairings (Fig. 5b, c; Schmid et al. 1983). It can for several reasons be assumed that the somatic pairings between the heterochromatin of the chro- mosomes 15 and Y favor the origin of Yqs chromsomes (see Discussion).
A detailed study of the replication patterns after 5-BrdU incorporation followed by Hoechst 33258-Giemsa staining or acridine orange fluorescence staining, could be performed on the Yqs ofProband W. The replication patterns ofnon-satellit- ed Y chromosomes from two control probands were used for comparison. The analysis was done on early replication stages (corresponding to those of R-banded chromosomes) in pro- metaphasic chromosomes (Fig. 4a). Those chromosome re- gions substituted with 5-BrdU during DNA replication stain dull with acridine orange (pale blue with Giemsa), whereas those incorporating thymidine exhibit bright acridine orange fluorescence (dark red Giemsa staining). The replication pat- terns of the euchromatic portion of the Yqs chromosome (short arm and proximal region of the long arm) were the same as in the Y chromosomes of the control probands (compare Fig. 4b, c and d, e; Fig. 4h, i and k, 1). Between two and four early replicating bands could be demonstrated. The same bands were also found in an extensive study on the high resolu- tion replication patterns of the human Y chromosome (Schempp and Mfiller 1982). The patterns of replication of the constitutive heterochromatin in the long arm of the Yqs chro- mosome could be categorized into two different types. One type was the same as in the Y chromosomes of the control pro- bands: the entire distal heterochromatin in the long arm repli- cated late, as shown by the dull acridine orange fluorescence (Fig. 4b, d, e) or pale blue Giemsa staining (Fig. 4h, k, 1). In contrast to this, the second type of the Yqs showed small seg- ments at the distal end of the long arm which obviously belonged to an earlier replication than the heterochromatin and which thus exhibited a bright acridine orange fluorescence (Fig. 4a, c) or dark red Giemsa staining (Fig. 4i). It seems rea- sonable to interpret these segments as the satellites on the dis- tal end of the Yqs, which complete their DNA replication ear- lier than the Y heterochromatin and later than the Y euchro- matin. Furthermore, the analysis of the replication patterns in the prometaphasic stages permitted the exclusion of structural

78
changes in the autosomes and in the X chromosome (Fig. 4a). Since none of the ten autosomes presented an anomaly of the short arm, it can be assumed that the reciprocal translocation between the short arm of an acrocentric chromosome and the long arm of the Y occurred in one of the male ancestors of Proband W.
Like all the other Yqs chromosomes described to date, the Yqs in the present study also showed very conspicuous associa- tions of their satellited long arms with the satellited short arms of the acrocentric autosomes. A series of such satellite associa- tions as found in the differently stained preparations is shown in Figs. 1, 2, and 4. One hundred Giemsa-stained metaphases from conventional lymphocyte cultures from each of the three probands were scored for the presence or absence of satellite associations. The frequencies with which the three Yqs chro- mosomes participated in satellite associations were very differ- ent (Table 1). As has already been observed in earlier studies on the association frequencies of acrocentric autosomes (Schmid et al. 1974; Hayata et al. 1977), the association frequen- cies of the Yqs chromosomes are largely determined by the length of their nucleolar constrictions: the longer the nucleo- lax constriction, the higher the frequency of association. Thus the Yqs of the Probands B and K which had prominent nucleo- lar constrictions, were involved in satellite associations with the acrocentric autosomes in 18% and 21% of the metaphases, respectively. By comparison, the Yqs ofproband W whose nu- cleolar constriction was much less pronounced, associated in only 10% of the metaphases (Table 1). In situ hybridization experiments with radioactively labeled ribosomal RNA to mi- totic chromosomes have ascertained that the number of 18S and 28S ribosomal RNA genes in the nucleolar constrictions of the NOR-carrying chromosomes are variable (Henderson et al. 1973; Evans et al. 1974). Furthermore, a positive correlation was found between the content of ribosomal RNA genes of a given NOR-carrying chromosome (as estimated by in situ hybridization) and its association frequency (Warburton et al. 1976). For these reasons, it must be assumed that more riboso- mal RNA genes are localized in the longer nucleolar constric- tions of the Yqs chromosomes of the Probands B and K than in the Yqs of the Proband W.
It should be noted that the association frequencies of the NOR-carrying chromosomes are very highly influenced by the culture conditions. Among other things, there were distinctly increased association frequencies of all NOR-carrying chromo- somes in the distamycin A-treated cell cultures. Thus the ana- lysis of the association frequency of the Yqs in the distamycin A-treated cells ofProband B resulted in values twice that of the conventional cultures (Table 1). The multiple associations, involving three or more NOR-carrying chromosomes (Fig. 2b- e), were especially frequent in the distamycin A-treated cul- tures (30 of 100 associations). An increase in the frequencies of satellite associations by in vitro treatment with distamycin A was also noted in the cells from amniotic fluid (Jotterand-Bel- lomo 1983).
Discussion
1. Structure of the satellited Y-chromosomes
There is no doubt that the satellited Y chromosomes studied in the present paper as well as all of those described previously carry an active NOR in the long arm. Both the high frequencies of association of the Yqs with the acrocentric autosomes (Table 1) as well as the positive Ag-staining of their satellite
stalks are conclusive of this. The Ag-staining in mitotic meta- phase chromosomes only demonstrates those NORs which were active in the preceeding interphase nuclei (Miller et al. 1976a, b; Engel et al. 1977; Schmid et al. 1982). Cytochemical and biochemical analyses showed that the Ag-stained material consists ofnonhistone proteins rich in sulfhydril and disulfide groups (Ag-staining proteins B23 and C23) which rapidly asso- ciate with the newly transcribed ribosomal RNA (Buys and Osinga 1980; Busch et al. 1982). Residues of these proteins remain associated with the NORs up into the metaphase and can therefore be demonstrated with Ag-staining. Therefore it could be concluded with certainty that the satellited Y chro- mosomes have an active role in nucleolus organization.
The examined Yqs chromosomes of the three probands showed distinct differences regarding the size of the satellites, the nucleolar constrictions, and of the constitutive heterochro- matin in the long arms. Furthermore, differences between these Yqs chromosomes were determined following quin- acrine and distamycin A/DAPI staining. The cytogenetic data derived from the 12 other cases with Yqs chromosomes (Table 1) also show that the structure of satellited Y chromosomes is not uniform. These differences can be interpreted as follows: 1) The Yqs chromosomes developed from normally structured Y chromosomes by means of reciprocal translocations with the short arms of acrocentric chromosomes. The breaks which occurred in the course of this in the long arm of the Y chromo- somes were obviously localized in different sites within the heterochromatic band Yql2 (Fig. 7). Thus in the cases of Genest et al. (1983), Howard-Peebles and Stoddard (1976), Tur- leau et al. (1978), Giraldo et al. (1981), Bordson and Varela (1982), and in the present Case B, the breaks must have occurred in the distal region of the band Yql2, because the heterochromatin in these Yqs still constitutes approximately two-thirds of the long arms (Fig. 7d). In contrast to this, the small remainders of heterochromatin in the Yqs described by Stella et al. (1980), Shabtai et al. (1981), Bayless-Underwood et al. (1983), and the lack of distamycin A/DAPI-positive hetero- chromatin in the present Cases K and W can be explained by breaks in the proximal portion of the band q12 of the originally normally structured Y chromosome (Fig. 7c). Yqs chromo- somes where the entire C-band-positive heterochromatin in the long arm is lacking and where the nucleolar constriction is directly adjacent to the euchromatic band Yqll, are very rare (Fig. 7b). The very small Yqs described by Hayata et al. (1975, 1977) was such a heterochromatin deficient Yqs. 2) The remaining differences in the morphology of the differ- ent Yqs chromosomes can be explained by the heterogeneity of the chromosome segments translocated from the short arms of the acrocentric autosomes onto the Y chromosomes. It is well known that the length of the nucleolar constrictions, the size of the satellites, and the quinacrine fluorescence of the satellites in the short arms of the acrocentric autosomes are extremely variable and constitute a constant, characteristic feature of each individual autosome (e.g., McKenzie and Lubs 1975; Schned11978). The short arms of those acrocentric auto- somes which were involved in the reciprocal translocation with the Y chromosomes must have been structured in different ways. Prominent satellites with pronounced nucleolar con- strictions were translocated on the Yqs chromosomes de- scribed by Genest et al. (1967), Genest (1972, 1973, 1978), Howard-Peebles and Stoddard (1976), Hayata et al. (1975,1977), Stella et al. (1980), Giraldo et al. (1981), Bordson and Varela (1982), and Bayless-Underwood et al. (1983). This was also true for the Yqs of the present Probands B and K (Figs. la, 3 a, Table
79
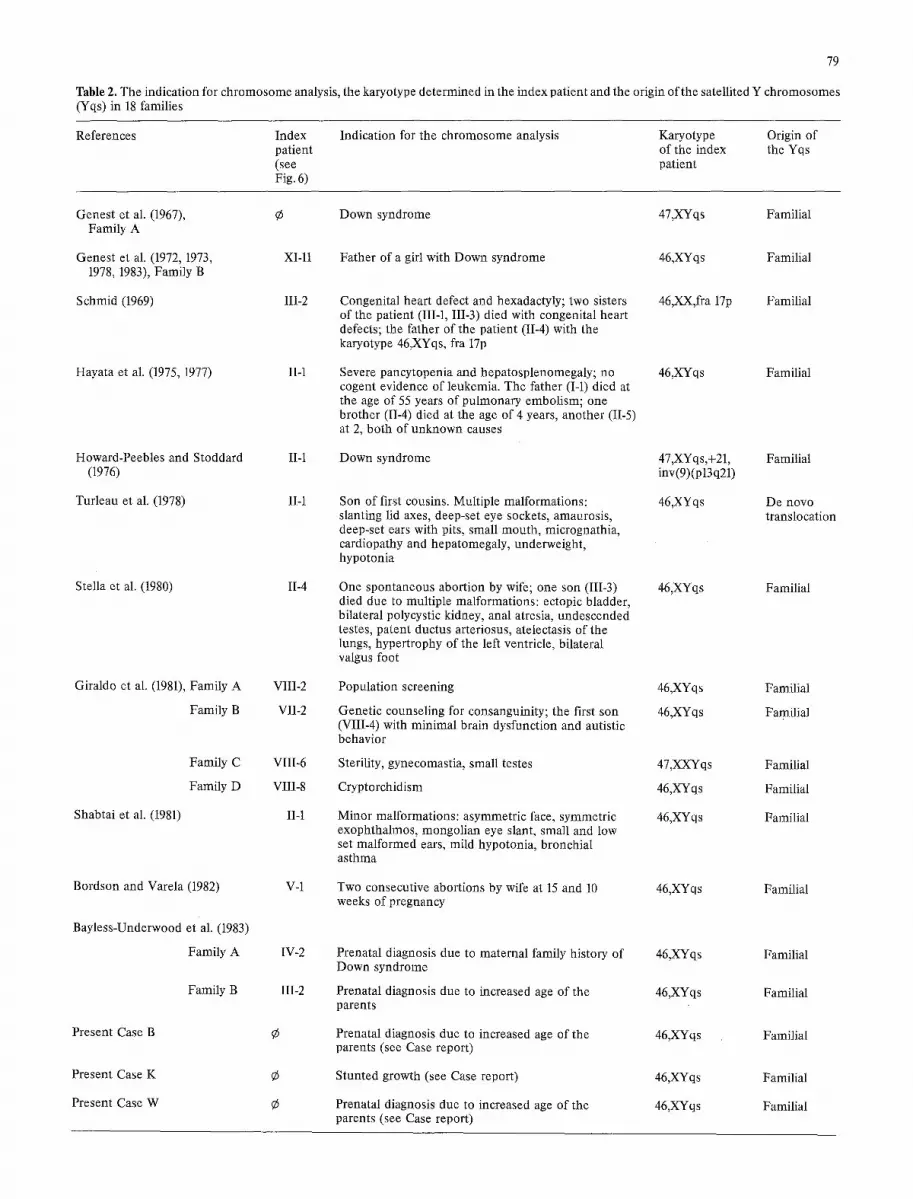
Table 2. The indication for chromosome analysis, the karyotype determined in the index patient and the origin of the satellited Y chromosomes (Yqs) in 18 families
References Index Indication for the chromosome analysis Karyotype Origin of patient of the index the Yqs (see patient Fig. 6)
Genest et al. (1967), ¢ Down syndrome 47,XYqs Familial Family A
Genest et al. (1972, 1973, XI-11 Father of a girl with Down syndrome 46,XYqs Familial 1978, 1983), Family B
Schmid (1969) III-2 Congenital heart defect and hexadactyly; two sisters 46,XX,fra 17p Familial of the patient (III-1, III-3) died with congenital heart defects; the father of the patient (II-4) with the karyotype 46,XYqs, fra 17p
Hayata et al. (1975, 1977) 1I -1 Severe pancytopenia and hepatosplenomegaly; no 46,XYqs Familial cogent evidence of leukemia. The father (I-l) died at the age of 55 years of pulmonary embolism; one brother (II-4) died at the age of 4 years, another (II-5) at 2, both of unknown causes
Howard-Peebles and Stoddard II-1 (1976)
Turleau et al. (1978) II-1
Stella et al. (1980) II-4
Giraldo et al. (1981), Family A VIII-2
Family B VII-2
Family C VIII-6
Family D VIII-8
II-1 Shabtai et al. (1981)
Bordson and Varela (1982)
Bayless-Underwood et al. (1983)
Present Case B
Present Case K
Present Case W
V-1
Family A IV-2
Family B III-2
¢
¢
Down syndrome 47,XYqs,+21, Familial inv(9)(p13q21)
Son of first cousins. Multiple malformations: slanting lid axes, deep-set eye sockets, amaurosis, deep-set ears with pits, small mouth, micrognathia, cardiopathy and hepatomegaly, underweight, hypotonia
One spontaneous abortion by wife; one son (111-3) died due to multiple malformations: ectopic bladder, bilateral polycystic kidney, anal atresia, undescended testes, patent ductus arteriosus, atelectasis of the lungs, hypertrophy of the left ventricle, bilateral valgus foot
Population screening
Genetic counseling for consanguinity; the first son (VIII-4) with minimal brain dysfunction and autistic behavior
Sterility, gynecomastia, small testes
Cryptorchidism
Minor malformations: asymmetric face, symmetric exophthalmos, mongolian eye slant, small and low set malformed ears, mild hypotonia, bronchial asthma
Two consecutive abortions by wife at 15 and 10 weeks of pregnancy
Prenatal diagnosis due to maternal family history of Down syndrome
Prenatal diagnosis due to increased age of the parents
Prenatal diagnosis due to increased age of the parents (see Case report)
Stunted growth (see Case report)
Prenatal diagnosis due to increased age of the parents (see Case report)
46,XYqs De novo translocation
46,XYqs Familial
46,XYqs Familial
46,XYqs Familial
47,XXYqs Familial
46,XYqs Familial
46,XYqs Familial
46,XYqs Familial
46,XYqs Familial
46,XYqs Familial
46,XYqs Familial
46,XYqs Familial
46,XYqs Familial

80
Genest et a1.(1973,1983)
Family B
i IF
I
Ill
~i I ,v
VI
VII
2 " I ; "x i S c h m i d (1969)
Gira ldo et al. (1981) F - -(?) . . . . . . . ~ . . . . . .
F m! J
, i I Fam.Cl~
/
Hayata et al. (1975,1977)
Stella et al. (1980) Shabtai et a1.(1982)
- - - - @ I i o ~ ~ I ~ I
#11 III
Bordson and Varela ~'I ~ (1982)I
J:~,, i ,,
~" ~ | IV IV
~" J;]v, li ~" Fam, D m~ vii
I--/ , i v,, /
~m m ~ ~ ~ ii /
Bayless-Underwood et al. (1983)
Turleau et al. (1978)
II /
Key:
[ ] 46,XYqs male V j ~ obligate 46,XYqs carrier
"[~" possible 46 XYqs carrier VI " ~ 46,XX,fra17p
H o w a r d - P e e b l e s and [ ] 46,XYqs, fra17p S toddard (1976) [ ] unbalanced karyotype
r]C)normal karyotype I ~ [ . [ ~ not examined
' • abortion II ~ r [ ~ B i ~ ) { ~ ( ~ ) ~ ) ~ ) ( ~ ) / index patient
7 --.9-- possible consanguinity • de novo Yqs
intrauterine diagnosis
Family A Family B
ill 2 ~ III
IV /
Fig. 6. Pedigrees of the previously reported families with Yqs chromosomes. In the pedigree described by Genest et al. (1973,1983) only the male descendants are registered. For the pedigree of Howard-Peebles and Stoddard (1976) the information supplied by Howell et al. (1978) was used also. The numbers at the bottom-left of some probands are refered to in the text and in Table 2

81
P12"
Eb
r-q 1' .
Fig.Ta-f. Schematic representation of different types of reciprocal translocations which can produce Yqs chromosomes, a Normally structured acrocentric autosome and Y chromosome. Dark sector: quinacrine bright heterochromatin. Hatched sector: distamycin A/ DAPI bright heterochromatin. Dotted line (band p12): NOR. Note that the bands pll and p13 in the acrocentric autosome are also hetero- chromatic. The wavey lines at the two chromosomes indicate the pre- sumed breakage points. For the sake of simplicity, only one of the possible breakage points (band pll) in the acrocentric autosome was taken into account, b Break at the boundary of euchromatic band Yqll and heterochromatic band Yql2. e Break within the distamycin A/DAPI bright region of the heterochromatic band Yql2. d Break within the quinacrine bright region of the heterochromatic band Yql2. Note the structural differences between the resulting Yqs chro- mosomes, e, fReciprocal translocations between an acrocentric auto- some lacking its satellite (band p13) and a normally structured Y chro- mosome: this produces a Yqs chromosome with nucleolus organizer region (satellite stalk) but without satellites in its long arm
1). By comparison, only small satellites were translocated from the acrocentric autosomes onto the Y chromosomes in the cases of Turleau et al. (1978) and in the present Case W (Fig. 4m). Finally, in the Yqs described by Shabtai et al. (1981), the satellites could only be surmised but were never clearly seen (Table 1). The only Yqs chromosome whose satellite showed a similarly bright fluorescence as the terminal hetero- chromatin of the band Yql2 was that of the present Case B (Fig. 1q-s); all other Yqs chromosomes had a satellite whose quinacrine fluorescence was approximately the same intensity as the proximal euchromatic band Yqll (Figs. 3d and 4f, Table 1).
It needs to be considered that acrocentric chromosomes lacking the satellites in their short arms are not rare. In these
I I~ III IV 5 "M d ' D 5 "A II 111 IV 5"M d'D 5-A
ttttt; 1i11!t!
Fig. 8. The localization of the four satellite DNAs (I-IV), 5-methyl- cytosine enriched DNA (5-M), distamycin A/DAPI bright hetero- chromatin (d-D), and 5-azacytidine-sensitive heterochromatin (5-A) in the Y chromosome, the five acrocentric autosomes and autosomes 1, 9, and 16. The satellite DNAs I-IV are distributed according to the satellite maps of Miklos and John (1979). Every chromosome was divided into equal sized segments, each corresponding to one-half the length of chromosome 21. The major sites of hybridization to such segments are shown as black sectors. The threshold values used in the quantitative evaluation of the autoradiographs were 1.5 grains/ segment for satellites I, II, and IlI, and 2.0 grains/segment for satellite IV. Chromosome 16 carries satellite II (hatched sector); however, its amount falls below the threshold values used by Miklos and John (1979). The hatched area in the acrocentric autosome 22 indicates a very low amount of highly methylated DNA as detected by immuno- fluorescence technique and antisera specific for 5-methylcytosine (Schnedl et al. 1975). Note that only the distamycin A/DAPI positive heterochromatin is 5-azacyticline-sensitive (compare with Figs. 3f and 5a)
acrocentrics, residual parts of the NOR can sometimes be seen in the form of stalks (Schmid et al. 1974). If one of these par- tially deleted short arms is translocated on a Y chromosome, it forms a Yqs with a NOR localized on the end of the long arm, but without a satellite (Fig. 7e, f). This kind of Yqs chromo- some, for which the Yqs observed by Shabtai et al. (1981) serves as a good example, can be easily overlooked in the metaphases if the preparations are not Ag-stained. There is therefore the possibility that Yqs chromosomes do occur in a greater fre- quency than can be concluded from the few cases observed to date. More extensive studies with the simple method of Ag- staining for NORs are necessary to determine the frequency of this structural variation of the Y chromosome in the popula- tion.
It must be concluded from the structural differences be- tween the different Yqs chromosomes occurring in family lines (Tables 1 and 2) that these Yqs are not of monophyletic origin, but instead developed several times independent of each other. The differences in ethnic origin of the 14 families also suggest this. Finally, de novo origin ofa Yqs could be un- equivocally demonstrated in the case reported by Turleau et al. (1978). A normally structured Y chromosome was found in the father of a child with Yqs born with multiple malformations (I-l, II-1 in Fig. 6). The paternity was confirmed by extensive analysis of red blood cell enzymes and serum types.

82
2. Y/Autosomal translocations and the origin of Yqs chromosomes
As was repeatedly presented, the exchanges between the long arms of the Y chromosomes and the short arms of the acro- centric autosomes are supposed to be reciprocal. It can there- fore be expected that portions of the Y heterochromatin will be translocated to the short arms of the acrocentric autosomes participating in these translocations (Fig. 7a-d). A consider- able number of acrocentric autosomes with large, quinacrine- bright short arms were found (see review by Smith et al. 1979). The proof that these brightly fluorescing short arms actually consisted ofY heterochromatin and not of enlarged autosomal heterochromatin was furthered by the identification of two Y- specific, repetitive DNA sequences (2.1 and 3.4 kilobases in length) using Hae III restriction endonuclease analysis of the DNA from female carriers of such acrocentrics (Kinross et al. 1978; Cooke and Noel 1979; Schmid et al. 1983) and/or by the specific undercondensation of the translocated Y heterochro- matin in distamycin A-treated lymphocytes (Schmid 1979; Cohen et al. 1981; Funderburk et al. 1982; Schmid et al. 1983). The reviews of the Y/autosomal translocations described to date show that in 70% of the cases the Y heterochromatin was translocated on the short arm of an acrocentric autos6me. Of these in turn, chromosome 15 is the most frequently involved with a rate of 52%, followed by chromosome 22 (33%), chromo- some 21 (7%), and chromosomes 13 and 14 (4% each) (Smith et al. 1979; Davis 1981). These numbers clearly show that the short arms of chromosomes 15 and 22 participate in transloca- tions with the Y chromosome much more frequently than expected, whereas chromosomes 13, 14, and 21 are underrepre- sented.
There are two mutually complementary explanations for the overproportionally frequent involvement of chromosomes 15 and 22 in translocations with the Y heterochromatin: (1) Very similar or identical DNA sequences are localized in the constitutive heterochromatin of the short arms of chromo- somes 15 and 22 and in the long arm of the Y chromosome, which promotes pairing and crossing over between these non- homologous chromosome regions. (2) In the interphase nu- clei, the heterochromatic regions of chromosomes 15, 22, and Y are arranged in very close spatial contact ("somatic pairing"), which makes a translocation between them more likely. As far as the DNA sequences are concerned, the studies with the in situ hybridization technique (for review see Gosden et al. 1975; Miklos and John 1979) have shown that high concentrations of all four satellite DNAs I - IV are localized in the heterochroma- tin of chromosomes 15 and Y; the heterochromatin of chromo- some 22 is enriched with the satellite DNAs I, III, and IV, whereas the amount of satellite DNA II found here falls below the threshold values used in the quantitative evaluation of the autoradiographs (Fig. 8). In the heterochromatin of the remaining acrocentric autosomes (Nos. 13, 14, and 21), only satellite IV is present in high concentrations. Using an immu- nofluorescence technique and antisera specific for 5-methyl- cytosine, it was furthermore possible to demonstrate that the heterochromatic regions of chromosomes 15 and Y and, to a lesser degree, chromosome 22 contain highly methylated DNA (Fig. 8) (Miller et al. 1974; Schnedl et al. 1975). These observa- tions show that the agreement of the highly repetitive DNA is in fact greater between the heterochromatin of chromosomes 15, 22, and Y than it is between the heterochromatin of chro- mosomes 13, 14, 21, and the Y chromosome.
Somatic pairings between the heterochromatin of chromo- somes 15 and Y have also been recently demonstrated success-
fully (Schmid et al. 1983a). The condensation of the hetero- chromatin in chromosomes 1, 9, 15, 16, and Y during the mitot- ic prophase is inhibited by the in vitro treatment of cell cul- tures with 5-azacytidine (Figs. 3g, h and 5a). As a consequence, the somatic pairings between these heterochromatic regions remain intact from the interphase nuclei up into metaphase and are thus susceptible to analysis (Fig. 5b, c). The examples shown in Fig. 5b illustrate the strength of the somatic pairings between the heterochromatin of chromosomes 15 and Y, and why these promote the development of Yqs chromosomes. The translocation only exchanges terminally located hetero- chromatin and a NOR. Since the heterochromatin is geneti- cally inert and as the number of active NORs is inter-individ- ually variable anyway, the gametes of the carrier of such a translocation are at no selective disadvantage. Both the Yqs chromosome as well as the chromosome 15 with the translocat- ed Y heterochromatin can be transmitted without effect on the phenotype of the progeny, even when they segregate indepen- dently of each other after the meiotic divisions; no imbalance of genetically relevant material can occur. The situation is completely different, however, when reciprocal translocations occur due to somatic pairings between the Y heterochromatin and the pericentromeric heterochromatin ofautosomes 1, 9, or 16 (Fig. 5c). In these cases, the entire arm of one of these auto- somes is translocated on the Y chromosome. The chromoso- mal balance of the progeny would only be secured if the re- arranged Y chromosome segregates in the same gamete to- gether with the rearranged autosome. For this reason, the translocations between the heterochromatin of the Y chromo- some and the non-acrocentric autosomes are subject to high postmeiotic and postzygotic selective pressure.
The mitotic condensation of the heterochromatin in the short arms of the acrocentric autosomes Nos. 13, 14, 21, and 22 is not inhibited by 5-azacytidine (Figs. 5a and 8). The somatic pairings between these acrocentrics and the Y heterochroma- tin can therefore not be analysed by in vitro treatment of the cultures with 5-azacytidine. However, there are definite indi- cations for the existence of somatic pairings between the short arms of all acrocentric autosomes and the Y heterochromatin in the interphase nuclei. It can thus be shown on quinacrine- and subsequently Ag-stained nuclei that the brightly fluor- escing Y heterochromatin (Y body) is always localized in the middle of all active, Ag-positive NORs (Fig. 5d-f). It can be stated in summary that both the common satellite DNAs in the heterochromatin of chromosomes 15, 22, and Y as well as the somatic pairings between the Y heterochromatin and the het- erochromatin of all acrocentric autosomes, constitute decisive factors for the development of Yqs chromosomes.
3. Clinical significance of the satellited Y chromosomes
As discussed in the previous section, the presence of a Yqs chromosome should cause no phenotypic abnormalities in the carriers. Most of the Yqs chromosomes described are indeed family marker chromosomes, transmitted in patrilineary fashion for several generations, which did not lead to progeny with specific phenotypic malformations in these families (Fig. 6). The most impressive example of this is the French- Canadian family described by Genest et al. (1973, 1983). In this family, the Yqs arose more than 300 years ago and is the oldest translocation chromosome yet reported in the human species. This Yqs was only discovered because one of the 16 male de- scendants in the eleventh generation (XI-11 in Fig. 6) had a child with trisomy 21.

Table 3. Summary of NOR-carrying sex chromosomes in mammals (according to Merry et al. 1983) a
83
Species NOR-carrying chromosomes b
Autosomes X Y
Hylobates concolor (whi te-cheeked gibbon)
Hylobates syndactylus (siamang)
Ammospermophilus harisii (antelope squirrel)
Nyctereutes procyonoides (raccoon dog)
Canisfamiliaris (domestic clog)
Callithrix ssp. (Brazilian marmosets)
Lagorchestes conspicillatus (hare wallaby)
Carollia castanea (Allen's fruit bat)
Carollia perspicillata (Seba's fruit bat)
Monodelphis domestica (gray short-tailed opossum)
q-
+
+
+
+
+
+
a The position of the NORs in the sex chromosomes is a constitutive property of the karyotypes of these species and not an occasionally occur- ring rearrangement in individual animals
b Demonstrated by in situ hybridization of radioactively labeled 18S and 28S ribosomal RNA to mitotic chromosomes and/or Ag-staining
Table 2 shows that the abnormal phenotypes diagnosed in some of the carriers of an inherited Yqs chromosome are of very heterogeneous nature. No specific anomaly can be corre- lated with the 46,XYqs karyotype. Neither was there an incrased incidence of infertility or a remarkable number of spontaneous abortions in the families of the index patients. This leaves the conclusion that the malformations found in some of the families with a satellited Y chromosome are inci- dental findings bearing no relation to this structural variation of the Y chromosome. This is confirmed by the fact that three of the inherited Yqs chromosomes were found by chance in prenatal diagnoses performed due to the advanced age of the parents (Family A of Bayless-Underwood et al. 1983; current Cases B and W), and another one in the course ofa cytogenetic population study on chromosomal polymorphisms (Family A of Giraldo et al. 1981).
The decision concerning a possible karyotype-phenotype correlation in the only case of de novo Yqs chromosome de- scribed to date (Turleau et al. 1978) poses more of a problem. In this case, a Yqs chromosome was demonstrated in a newborn male (II-1 in Fig. 6) with severe multiple malformations (Table 2), whereas the father (I-l, Fig. 6) possessed an inconspicuous phenotype and a normally structured Y chromosome. Illegiti- macy could be excluded with a high degree of certainty by the analysis of serologic markers. It was suggested that the translo- cation between the Y heterochromatin and the short arm of an acrocentric autosome took place during paternal meiosis. A postzygotic translocation seemed improbable as no mosaicism of 46,XY/46,XYqs was found and because none of the acro- centric autosomes in the 46,XYqs cells exhibited a morpholog- ic alteration in the short arms. The breakpoint in the paternal Y chromosome must have occurred in the distal portion of the heterochromatic band Yq12, because this heterochromatin was not visibly reduced (see Fig. 2c of Turleau et al. 1978). As the schematic diagram in Fig. 7d shows, no change in geneti- cally relevant euchromatin occurs in the Yqs chromosome in the course of such a translocation. Besides this, the symptoms shown by the patient we1"e much more severe than those occur- ring in an aberration involving the euchromatin in the long
arm of the Y chromosome (band Yqll) (see review by Davis 1981). For these reasons, it appears improbable that the de novo Yqs was the causal factor for the malformations found in the patient. It needs to be taken into account that the parents of the patient were first cousins; therefore, the possibility of an auto- somally recessively inherited d isease-as discussed by Turleau et al. (1978)-must also be considered.
The Ag-staining studies on karyotypes of normal male pro- bands have shown that the modal number of Ag-positive NORs per individual lies between 6 and 10 (Mikelsaar et al. 1977). It can already be recognized by this great inter-individual variability in the number of active NORs in diploid cells that no disadvantages can be expected for the carrier of an additional NOR in the Yqs chromosome. At this point, one should also consider the exceptional chromosomal position of the NORs in some other mammals. It has been shown for six species to date that aNOR is localized in the Y chromosomes ofal lmale individuals studied in addition to the NORs in the autosomes (Table 3). Apparently, NORs from autosomes were translocat- ed on the Y chromosomes in the ancestors of these species and these NOR-carrying Y chromosomes subsequently became established in all males by drift in small, inbreeding popula- tions. This produced a sex-specific difference in the NOR- number, which did not constitute a selective disadvantage for the further differentiation of these species. The NOR-carrying X chromosomes in some of these mammals can be assessed in a similar fashion (Table 3).
Some of the earlier studies on Yqs chromosomes already discussed the question of whether these structural variations in the Y chromosome could predispose to aberrations involv- ing other chromosomes. Thus the patient (Family A) of Genest et al. (1967) had both trisomy 21 and the Yqs chromo- some. The index patient of Howard-Peebles and Stoddard (1976) was a male with Yqs, trisomy 21, and inversion in the chromosome 9 (Table 2). The index patient of the Family B examined by Genest et al. (1973, 1983) (XI-11 in Fig. 6) was the father of a girl with trisomy 21. Klinefelter syndrome and a satellited Y chromosome (47,XXYqs) were determined in the index patient in the Family C of Giraldo et al. (1981) (VIII-6 in

84
Fig. 6) and in a case recorded by Genes t (1978); the latter is a pat ient whose ancestors came f rom Qu6bec and who were probably related to the French-Canadian Family B of Genes t et al. (1973, 1983). A very unusua l - -and with regard to the etio- logic invo lvement o f the Yqs the most i n t e r e s t i n g - c h r o m o - some aberrat ion was again found in the Family B o f G e n e s t et al. (1983) in a newborn male (XII-2 in Fig. 6) wi th craniosteno- sis and minor malformat ions . The banding analysis of the karyotype revealed that the terminal part of the long arm of a p resumed acrocentr ic au tosome No. 14 was translocated onto the satellite of the Yqs chromosome: t ( Y q s t e r : : 1 4 q 2 4 ~ 14q ter). The clinical picture of the patient was very similar to the phenotypes reported in four o ther cases of partial t r isomies for the terminal part of 14q. The t ranslocat ion mus t have ocurred de novo during paternal meiosis since the father of the pat ient (XI-8 in Fig. 6) had a typical Yqs chromosome. There was an interval of three centuries (nine generat ions) be tween the origin of the Yqs ch romosome in this family (probably in III-3 in Fig. 6) and the t ranslocat ion 14q/Yqs. Taking all un- balanced karyotypes found in the families with Yqs chromo- somes into account, it is not easy to decide whe ther the Yqs ch romosome played a role in the deve lopment of addit ional ch romosome aberrations or not. On the o ther hand, the observed tr isomies 21 and Klinefel ter syndromes are among the most f requent of all ch romosomal disorders and could therefore be just if iably considered incidental findings. On the o ther hand, the t ranslocat ion demonst ra ted be tween the satel- lites of a Yqs and a terminal part o f l 4 q (Genes t et al. 1983) war- rants caut ion with regard to trivializing the risk for unbalanced gametes in the carriers of Yqs chromosomes . W h e t h e r a causal relat ionship exists be tween the presence o f a Y q s ch romosome and the origin of chromosomal abnormali t ies during meiosis or not could be de te rmined by the analysis of ch romosome complements f rom the sperm of Yqs carriers by the fertiliza- t ion of hamster eggs (Rudak et al. 1978; Mart in 1983).
Acknowledgements. We thank Miss Beatrix Geile for technical assist- ance and Mrs. Jutta Jackowski for photographic documentation.
References
Bayless-Underwood L, Cho S, Ward B, Robinson A (1983) Two cases of prenatal diagnosis of a satellited Yq chromosome. Clin Genet 24 : 359-364
Bordson B, Varela M (1982) Yqs in an American family of Scottish descent. Hum Genet 60 : 387-388
Bfihler EM (1980) A synopsis of the human Y chromosome. Hum Genet 55 : 145-175
Busch H, Lischwe MA, Michalik J, Chan P-K, Busch RK (1982) Nucleolar proteins of special interest: silver-staining proteins B23 and C23 and antigens of human tumour nucleoli. In: Jordan EG, Cullis CA (eds) The nucleolus. Cambridge University Press, Cambridge, pp 43-7l
Buys CHCM, Osinga J (1980) Abundance of protein-bound sulf- hydril and disulfide groups at chromosomal nucleolus organizing regions-a cytochemical study on the selective silver staining of NORs. Chromosoma 77 : 1-11
Caspersson T, Zech L, Johannsson C, Modest EJ (1970) Identification of human chromosomes by DNA-binding fluorescent agents. Chromosoma 30 : 215-227
Cohen MM, Frederick RW, Balkin NE, Simpson SJ (1981) The iden- tification of Y chromosome translocations following distamycin A treatment. Clin Genet 19 : 335-342
Cooke H J, Noel B (1979) Confirmation ofY/autosome translocations using recombinant DNA. Hum Genet 50 : 39-44
Couturier J, Dutrillaux B, Lejeune J (1973) Etude des fluorescences sp6cifique des bandes R et des bandes Q des chromosomes humains. CR Seances Acad Sci [III] 276 : 339-342
Davis RM (1981) Localisation of male-determining factors in man: a thorough review of structural anomalies of the Y chromosome. J Med Genet 18 : 161-195
Engel W, Zenzes MT, Schmid M (1977) Activation of mouse ribo- somal RNA genes at the 2-cell stage. Hum Genet 38 : 57-63
Evans HJ, Buckland RA, Pardue ML (1974) Location of the genes coding for 18S and 28S ribosomal RNA in the human genome. Chromosoma 48 : 405-426
Funderburk S J, Klisak I, Sparkes RS, Carrel RE (1982) Familial Y-autosome translocation in two unrelated girls. Ann G6n6t (Paris) 25 : 119-122
Genest P (1972) An eleven-generation satellited Y chromosome. Lancet I : 1073
Genest P (1973) Transmission h6r6ditaire, depui 300 arts, d'un chro- mosome Y a satellites dans une lign6e familiale. Ann G6n6t (Paris) 16 : 35-38
Genest P (1978) Propos sur un chromosome Y a satellites. Ann G6n6t (Paris) 21 : 237-238
Genest P, Bouchard M, Bouchard J (1967) A satellited human Y chromosome. Lancet I : 1279-1280
Genest P, Genest FB, Gagnon-Blais D (1983) Un remaniement chro- mosomique inhabituel. Une translocation t61omerique autoso- mique sur un Y ~t satellites (Yqs) multicentenaire. Ann G6n6t (Paris) 26 : 86-90
Giraldo A, Martinez I, Guzmfin M, Silva E (1981) A family with a satellited Yq chromosome. Hum Genet 57 : 99-100
Goodpasture C, Bloom SE (1975) Visualization of nucleolar orga- nizer regions in mammalian chromosomes using silver staining. Chromosoma 53 : 37-50
Goodpasture C, Bloom SE, Hsu TC, Arrighi FE (1976) Human nucleolus organizers: the satellites or the stalks? Am J Hum Genet 28 : 559-566
Gosden JR, Mitchell AR, Buckland RA, Clayton RP, Evans HJ (1975) The location of four human satellite DNAs on human chromosomes. Exp Cell Res 92 : 148-158
Hayata I, Oshimura M, Kakati S, Sandberg AA (1975) Deletion of Y-heterochromatin and origin of brightly fluorescing satel- lites. Mam Chrom Newsletter 16 : 78
Hayata I, Oshimura M, Sandberg AA (1977) N-band polymorphism of human acrocentric chromosomes and its relevance to satellite association. Hum Genet 36 : 55-61
Henderson AS, Warburton D, Atwood KC (1973) Ribosomal DNA connectives between human acrocentric chromosomes. Nature 245 : 95-97
Howard-Peebles PN, Stoddard GR (1976) A satellited Yq chromo- some associated with trisomy 21 and an inversion of chromo- some 9. Hum Genet 34 : 223-225
Howell WM, Howard-Peebles PN, Block BM, Stoddard GR (1978) Silver stain reveals nucleolus organizer regions on a satellited Yq chromosome. Hum Genet 42 : 245-250
Jotterand-Bellomo M (1983) Les effets de la distamycine A sur les cellules du liquide amniotique cultiv6es in vitro. Ann G6n6t (Paris) 26 : 27-30
Kinross J, Fraccaro M, Scappaticci S, Tiepolo L, Zuffardi O, Pawlo- witzki IA, Jones KW (1978) Bsu restriction of DNA from cases exhibiting sex-chromosome abnormalities. Cytogenet Cell Genet 20 : 59-69
Martin RH (1983) A detailed method for obtaining preparations of human sperm chromosomes. Cytogenet Cell Genet 35 : 252-256
Martin Lucas MA, P6rez Castillo A, Abrisqueta JA (1983) Satellited Yq chromosome. A familial study. Clin Genet 23:237
Merry DE, Pathak S, VandeBerg JL (1983) Differential NOR activ- ities in somatic and germ cells ofMonodelphis domestica (Marsu- piala, Mammalia). Cytogenet Cell Genet 35 : 244-251
McKenzie WH, Lubs HA (1975) Human Q and C chromosomal variations: distributions and incidence. Cytogenet Cell Genet 14 : 97-115
Mikelsaar A-V, Schmid M, Krone W, Schwarzacher HG, Schnedl W (1977) Frequency of Ag-stained nucleolus organizer regions in the acrocentric chromosomes of man. Hum Genet 37 : 73-77
Miklos GLG, John B (1979) Heterochromatin and satellite DNA in man: properties and prospects. Am J Hum Genet 31 : 264-280
Miller O J, Schnedl W, Allen J, Erlanger I~F (1974) 5-Methylcytosine localised in mammalian constitutive heterochromatin. Nature 251 : 636-637

85
Miller DA, Dev VG, Tantravahi R, Miller OJ (1976a) Suppression of human nucleolus organizer activity in mouse-human somatic hybrid cells. Exp Cell Res 101 : 235-243
Miller OJ, Miller DA, Dev VG, Tantravahi R, Croce CM (1976b) Expression of human and suppression of mouse nucleolus orga- nizer activity in mouse-human somatic cell hybrids. Proc Natl Acad Sci USA 73 : 4531-4535
Overton KM, Magenis RE, Brady T, Chamberlin J, Parks M (1976) Cytogenetic darkroom magic: now you see them, now you don't. Am J Hum Genet 28 : 417-419
Pfeiffer RA (1974) Cell cultures from blood and bone marrow. In: Schwarzacher HG, Wolf U (eds) Methods in human cyto- genetics. Springer, Berlin Heidelberg New York, pp 1-37
Prantera G, Pimpinelli S, Rocchi A (1979) Effects of distamycin A on human leukocytes in vitro. Cytogenet Cell Genet 23 : 103-107
Rudak E, Jacobs PA, Yanagimachi R (1978) Direct analysis of the chromosome constitution of human spermatozoa. Nature 274 : 911-913
Schempp W, Meer B (1983) Cytologic evidence for three human X-chromosomal segments escaping inactivation. Hum Genet 63 : 171-174
Schempp W, Mtiller U (1982) High resolution replication patterns of the human Y chromosome. Intra- and interindividual varia- tion. Chromosoma 86 : 229-237
Schmid M (1979) Demonstration of Y/autosomal translocations using distamycin A. Hum Genet 53 : 107-109
Schmid M, Krone W, Vogel W (1974) On the relationship between the frequency of association and the nucleolar constriction of individual acrocentric chromosomes. Hum Genet 23 : 267-277
Schmid M, L6ser C, Schmidtke J, Engel W (1982) Evolutionary conservation of a common pattern of activity of nucleolus or- ganizers during spermatogenesis in vertebrates. Chromosoma 86 : 149-179
Schmid M, Grunert D, Haaf T, Engel W (1983a) A direct demon- stration of somatically paired heterochromatin of human chro- mosomes. Cytogenet Cell Genet 36 : 554-561
Schmid M, Schmidtke J, Kruse K, Tolksdorf M (1983b) Characteri- zation of a Y/15 translocation by banding methods, distamycin A treatment of lymphocytes and DNA restriction endonuclease analysis. Clin Genet 24 : 234-239
Schmid W (1967) Heterochromatin in mammals. Arch Julius Klaus- Stift Vererbungsforsch Sozialanthropol Rassenhyg 42 : 1-60
Schmid W (1969) Satellites on the long Y chromosome arm: a fami- lial Y/autosome translocation in man. Cytogenet Cell Genet 8 : 415-426
Schnedl W (1978) Structure and variability of human chromosomes analysed by recent techniques. Hum Genet 41 : 1-9
Schnedl W, Dev VG, Tantravahi R, Miller DA, Erlanger BF, Miller OJ (1975) 5-Methylcytosine in heterochromatic regions of chro- mosomes: chimpanzee and gorilla compared to the human. Chromosoma 52 : 59-66
Schweizer D, Ambros P, Andrle M (1978) Modification of DAPI banding on human chromosomes by prestaining with a DNA binding oligopeptide antibiotic, distamycin A. Exp Cell Res 111 : 327-332
Shabtai F, Eilam N, Elian E, Halbrecht I (1981) A new family with a satellited Y. Ann G6n6t (Paris) 24 : 223-225
Smith A, Fraser IS, Elliot G (1979) An infantile male with balanced Y;19 translocation: review of Y;autosome translocations. Ann G6n6t (Paris) 22 : 189-194
Stella M, Rossi R, Bonfante A, Rossi G (1980) A new case of human Y chromosome with satellites on the long arm. J G6n6t Hum 28 : 39-45
Sumner AT (1972) A simple technique for demonstrating centro- meric heterochromatin. Exp Cell Res 75 : 304-306
Turleau C, Chavin-Colin F, Seger J, Sorin M, Salet D, de Grouchy J (1978) Chromosome Y avec satellite (Yqs) et organisateur nu- cl6olaire survenu de novo. Ann G6n6t (Paris) 21 : 239-242
Viegas-P6quignot E, Dutrillaux B (1976) Segmentation of human chromosomes induced by 5-ACR (5-azacytidine). Hum Genet 34 : 247-254
Warburton D, Atwood KC, Henderson AS (1976) Variation in the number of genes for rRNA among human acrocentric chromo- somes: correlation with frequency of satellite associations. Cyto- genet Cell Genet 17 : 221-230
Received January 9, 1984





























