Embed Size (px)
Citation preview

REGULAR ARTICLE
Savanna soil fertility limits growth but not survivalof tropical forest tree seedlings
Ricardo A. G. Viani & Ricardo R. Rodrigues &
Todd E. Dawson & Rafael S. Oliveira
Received: 20 February 2011 /Accepted: 21 June 2011 /Published online: 14 July 2011# Springer Science+Business Media B.V. 2011
AbstractBackground and Aims Cerradão (Brazilian woodlandsavannas) and seasonally dry forests (SDF) fromsoutheastern Brazil occur under the same climate butare remarkably distinct in species composition. Theobjective of this study was to evaluate the role of soilorigin in the initial growth and distribution of SDFand Cerradão species.Methods We conducted a greenhouse experimentgrowing Cerradão and SDF tree seedlings over theirsoil and the soil of the contrasting vegetation type. Weevaluated soil nutrient availability and seedlingsurvivorship, growth and leaf functional traits.Results Despite the higher nutrient availability in SDFsoils, soil origin did not affect seedling survivorship. Thethree SDF species demonstrated home-soil advantage,
enhanced growth with increasing soil nutrient availabil-ity and had higher growth rates than Cerradão species,even on Cerradão soils. Growth of Cerradão seedlingswas not higher on Cerradão soil and, overall, was notpositively correlated with soil nutrient availability.Conclusions SDF species are fast-growing specieswhile Cerradão trees tend to be slow-growing species.Although savanna soil reduces growth of forestspecies, our findings suggest that soil chemicalattributes, alone, does not exclude the occurrence ofSDF seedlings in Cerradão and vice-versa.
Keywords Atlantic forest . Cerrado . Savanna-forestboundaries . Soil fertility . Nutrient availability . Growthstrategies
AbbreviationsLAR leaf area ratioLD leaf tissue densityLMR leaf mass ratioLT leaf thicknessNAR net assimilation rateRGR relative growth rateSDF seasonally dry forestSLA specific leaf area
Introduction
Savannas and forests are the two major vegetationtypes found in the tropics. In Brazil, the Cerradosavannas and the Atlantic Forest are the second and
Plant Soil (2011) 349:341–353DOI 10.1007/s11104-011-0879-7
Responsible Editor: Harry Olde Venterink.
R. A. G. Viani (*) : R. S. OliveiraDepartment of Plant Biology, Biology Institute,State University of Campinas,CP 6109,Campinas-SP 13083-970, Brazile-mail: [email protected]
R. R. RodriguesDepartment of Biological Sciences, “Luiz de Queiroz”College of Agriculture, University of São Paulo,CP 9,Piracicaba-SP 13418-900, Brazil
T. E. DawsonDepartment of Integrative Biology,University of California,Berkeley, CA 94720, USA

third largest biomes covering about 37% of thecountry. Due to their threatened status and highspecies diversity and endemism, both Cerrado andAtlantic Forest belong to the 25 world hotspots and assuch are considered the most important areas forbiodiversity conservation (Myers et al. 2000). Fre-quently, these two biomes form vast contact zones(Durigan and Ratter 2006) and this is particularly truein southeastern Brazil, where SDF, a vegetationphysiognomy of the Atlantic Forest, and Cerradão, awoodland of Cerrado biome, are among the dominantvegetation types (Kronka et al. 2005).
Climatically, part of the Brazilian Cerrado occursin areas that could support tropical forests. In fact,southeastern Brazil Cerradão and SDF occur undersimilar seasonal climates, characterized by a pro-nounced dry season during the winter, but they areremarkably distinct in structure and floristic com-position, with few species occurring in bothenvironments, suggesting species specialization todifferent habitats.
There are many forest-savanna contact zonesacross the world, in which factors such as fire(Ratter 1992; Hoffmann 2000; Hoffmann et al. 2003;Banfai and Bowman 2005), light regime (Hoffmannand Franco 2003), soil nutrient status (Ratter 1992;Bowman and Panton 1993; Bond 2010; Wang et al.2010; Lehmann et al. 2011), and rainfall seasonality(Lehmann et al. 2011) have been reported as driversor not of savanna-forest boundary dynamics. In theSDF-Cerradão boundaries, previous studies showedthat SDF occurs over soils with higher nutrientavailability, while Cerradão occurs over nutrient-poor soils (Furley and Ratter 1988; Ratter 1992;Ruggiero et al. 2002). Although these studiessuggest soil nutrient availability as the main factoraffecting the distribution of SDF and Cerradão insoutheastern Brazil contact zones, they were notbased on experimental work.
In fact, soil nutrient availability is a key factordetermining distribution of species and compositionof many tropical forests and woodlands (McGraw andChapin 1989; Bowman and Panton 1993; Theodoseand Bowman 1997; Oliveira-Filho et al. 2001; John etal. 2007). Soil-related habitat specialization has beendocumented in Amazon forests (Fine et al. 2004),dipterocarp rainforests (Palmiotto et al. 2004) and insandstones (ridges) and alluvium (valley) soils ofBorneo (Baltzer et al. 2005).
While there is evidence that soils are importantfor maintaining savanna-forest boundaries, therehas been no experimental examination of the rolethat soil nutrient availability may play in promotingthe existence of Cerradão and SDF species insoutheastern Brazil contact zones. A mechanisticunderstanding of how nutrient availability influen-ces the performance of forest and savanna treeseedlings is therefore essential for predicting biomedistribution under both current and future climaticconditions. To evaluate experimentally the role ofsoil origin on the growth and, consequently, thedistribution of SDF and Cerradão species, weconducted a greenhouse experiment using treeseedlings that were grown in their native soil andalso in the soil of the contrasting vegetation type.We then addressed the following questions: 1) Howdo Cerradão tree species perform in their nativesoil versus SDF soil and vice-versa? 2) Is soilnutrient availability a key factor affecting plantfunctional traits that may in turn limit the earlyestablishment of Cerradão and SDF tree species inthe field? 3) Do SDF and Cerradão species havedifferent growth responses in relation to contrastingsoil fertility? We hypothesized that if soil nutrientavailability (fertility) is a key factor limiting thedistribution of species from both vegetation types,then species will show lower survivorship and/orlower growth rates when grown on the soil of thecontrasting vegetation type.
Material and methods
Study area and species
Seeds from Cerradão and SDF species werecollected in Estação Ecológica de Assis (22°32-39′S, 50°22-24′W) and in Estação Ecológica dosCaetetus (22°22-26′S, 49°40-44′W), respectively.Both sites are protected areas located in the state ofSão Paulo, southeastern Brazil. Cerradão is bestdescribed as a dense woodland with abundantevergreen trees and shrubs. Cerradão vegetation hasa semi-closed canopy with tree cover varying from50–90% (Ribeiro and Walter 1998) and does nothave a remarkable grassy understory. The EstaçãoEcológica de Assis has a Cwa Koeppen’s climatetype, with a mean annual temperature of 22.1°C.
342 Plant Soil (2011) 349:341–353

Mean annual rainfall is 1,440 mm and elevation is505 m. The main soil type is a deep, well-drainedand sandy dystrophic red Latosol with low wateravailability during the driest months (Juhász et al.2006). SDF is a closed canopy vegetation type withabundant deciduous and semi-deciduous trees andshrubs. During the driest months, canopy trees maylose up to 50% of their leaf area (Veloso 1992). TheEstação Ecológica dos Caetetus has the sameclimate type (Cwa), but mean annual temperatureand rainfall of 21.3°C and 1,460 mm, respectively.Its mean elevation is 522 m. The main soil type isa well drained and sandy clay loam Ultisol. Inboth vegetation types, less than 25% of totalannual precipitation falls during April to Septem-ber, indicating a strong seasonality of rainfall andthe existence of a dry season during this period.Although only 100 km apart and possessingsimilar climates, the Cerradão and SDF sites havestructurally and floristically distinct vegetationtypes. The Cerradão site has almost the double ofstems per hectare compared to SDF but the SDFtrees have a mean diameter at breast height almostfour times higher. Total number of woody specieswith DBH>5 cm in the two study sites is 268, butonly 18 species are shared between them.
The seeds of all studied species were collected inthe field from Jul-2007 to Oct-2007. We selected threewoody species based on seed availability and ende-mism to one of the vegetation types. The chosenspecies are also widespread and abundant over theirrespective vegetation type (Oliveira-Filho 2006). Thethree SDF species were: Balfourodendron riedelianum(Engl.) Engl., Cariniana estrellensis (Raddi) Kuntze,Centrolobium tomentosum Guillemin ex Benth.), andthe Cerradão species: Dimorphandra mollis Benth.,Machaerium acutifolium Vogel, Stryphnodendronobovatum Benth. All species, except B. riedelianum(Rutaceae) and C. estrellensis (Lecythidaceae) fromSDF, are legumes with potential for associating withN2-fixing bacteria (Sylvester-Bradley et al. 1980, Fariaet al. 1984, SM Faria personal communication 2009).
Cerradão and SDF soils used as the growthmedia for the greenhouse experiment were collect-ed from the 0–20 cm depth layer, in six differentlocations at Estação Ecológica de Assis andEstação Ecológica dos Caetetus. To provide betterrepresentation of horizontal variation in soil charac-teristics from both vegetation types, each of the six
locations were treated as independent samples in ourexperiment. In both sites, the soil was collected atrandomly selected points, at least 500 m apart fromeach other. After the soil was collected, it wassieved to eliminate plant material such as foliage,branches and other large debris and placed intoopaque polyethylene pots 11 cm in diameter and25 cm deep.
Prior to the beginning of the greenhouse experi-ment, a subsample (ca. 500 g) of each soil collectedin the field was analyzed for its chemical propertiesat a private soil analysis laboratory (Pirasolo—Laboratório Agrotécnico Piracicaba), located inPiracicaba, SP, Brazil. To account for total soilinorganic N, N-NO3 and N-NH4 were extracted inKCl and determined by ultraviolet spectrophotome-try and by flow injection analysis (FIA) followedby spectrophotometry at 650 nm respectively. SoilpH was determined by potentiometry in CaCl2solution, Al was extracted in KCl and determinedby acid–base titulation, and H+Al was extracted inSMP buffer and determined by potentiometry (vanRaij et al. 2001). P, K, Ca, and Mg were extracted inion exchange resin; Cu, Fe, Zn and Mn wereextracted in DTFA and B was extract in BaCl2(EMBRAPA 1997). Soil cations were determined byspectrophotometry, P and B by colorimetry and S-SO4 by turbidimetry (van Raij et al. 2001). Organicmatter was determined by the method of Walkleyand Black (Allen 1989).
Greenhouse experiment
Seeds were sown into sand boxes and after seedlinggermination and emergence, each plant was trans-planted into pots containing either their home-soil orthe soil of the contrasting vegetation type. Trans-planting occurred between Nov-2007 and Dec-2007.All seedlings were 5–10 cm at the beginning.Evaluations were made at 5 and 9 months aftertransplantation of each species. Each species wastransplanted into 240 replicate pots, with each one ofthe 12 soil locations containing 20 seedlings. Theexperiment was performed in a greenhouse located inthe “Luiz de Queiroz” College of Agriculture,Piracicaba, Brazil; there was no temperature or lightcontrol but during the experiment, temperature withinthe greenhouse varied between 12°C and 34°C andrelative humidity varied between 35–100%. Water
Plant Soil (2011) 349:341–353 343

was supplied every day to insure constant wateravailability to the seedlings. No additional nutrientswere added to soil. Pots were randomly placed andregularly relocated to minimize the effect of potentialenvironmental heterogeneity inside the greenhouse.
Survivorship, growth and leaf traits measurements
Five months after transplanting, plants were evaluatedfor survivorship, total plant leaf area and dry mass.Survivorship per species within each soil location (%)was calculated dividing the number of survivingseedlings by the initial number of seedlings (20),and then multiplying this number by 100. For leafarea and dry mass quantification, five randomlyselected plants per soil location were harvested andseparated into leaves, stems and roots. For thecompound-leaved species (all except B. riedelianumand C. estrellensis), only the leaflets were included forthe leaf area and mass calculations. After themeasurement of leaf area with a digital portable areameter (Li-Cor, LI-3000A), all the parts were ovendried for at least 48 h at 60°C and weighed separatelyusing an analytical balance.
Nine months after transplantation, the seedlingswere evaluated again for total plant leaf area anddry mass. Additionally, LT, LD, SLA, LAR, LMR,root:shoot ratio and leaf nutrient concentration weredetermined. Total leaf area and plant dry mass wereassessed as mentioned above, except for M. acuti-folium, where the number of seedlings evaluated persoil location was reduced to only three plantsbecause of high mortality rates. LT was measuredon three fully expanded leaves per plant, usingdigital calipers, avoiding prominent veins. Forsimple-leaved species three thickness measurementsper leaf (lamina base, middle and apices) were made.For compound-leaved species, we measured themiddle portion of three leaflets per leaf, with eachleaflet representing a position on the leaf: base,middle or apices. LD was calculated by dividingleaf dry mass by the product of leaf area multipliedby LT. SLA was obtained by dividing plant leafarea by leaf dry mass. LAR was calculated bydividing plant leaf area by plant dry mass. LMRwas calculated as the ratio of leaf mass to totalplant biomass and root:shoot ratio as the ratio ofbelowground biomass (root biomass) to above-ground biomass (stem and leaf biomass). One leaf
nutrient concentration analysis per soil locationwas performed with a compound sample containing theleaves from all individuals harvested at 9 months. LeafN, P, K, Ca, Mg and S concentration were analyzedat the Soil Science Department, at “Luiz deQueiroz” College of Agriculture, University ofSão Paulo, Brazil. N was determined by Kjeldahldistillation, after sulfuric acid digestion. Othernutrients were determined by optical (P) or atomicabsorption spectrophotometry (K, Ca, Mg, SO4),after nitric perchloric digestion.
RGR was calculated as:
RGR ¼ ðM2�M1Þ=ðT2� T1Þ
where, M2 and M1 are plant dry mass at the secondand first harvest, and T2—T1 the interval in daysbetween first (5 months) and second (9 months)evaluation times. NAR, which represents the netresult of dry mass gained via photosynthesis and lostin respiration by unit of leaf area, was calculated as:
NAR ¼ ðM2�M1ÞðlnA2� lnA1Þ=ðT2� T1Þ=ðA2� A1Þ
where, A2 and A1 are plant leaf area at the second andfirst harvest, respectively, and the other are asdescribed above for RGR calculation.
Data analysis
Differences in soils attributes between the two vegeta-tion types were tested by an unpaired Student’st-test,using each soil location as a replicate of its respectivevegetation type. Within species, effect of soil originover plant traits was assessed by an unpaired Student’st-test, with each soil location being considered areplicate of its soil origin. Overall effect of soil origin(Cerradão and SDF) and species over plant survivor-ship, growth parameters, morphological traits andnutrient leaf concentration were analyzed by a two-factor Analysis of Variance, considering soil andspecies as factors. Then, a post-hoc analysis was madewithin species using Tukey’s test. Finally, we per-formed Pearson’s correlation analyses to test therelationship between growth parameters and the mostrelevant soil fertility attributes (inorganic N, P and K+Ca+Mg soil availability). All plant and soil data,except for RGR and NAR, were log-transformed prior
344 Plant Soil (2011) 349:341–353
to the Analysis of Variance and Student’st-test analy-ses, in order to achieve normality and homogeneity ofvariance assumptions.
Results
Soil characterization
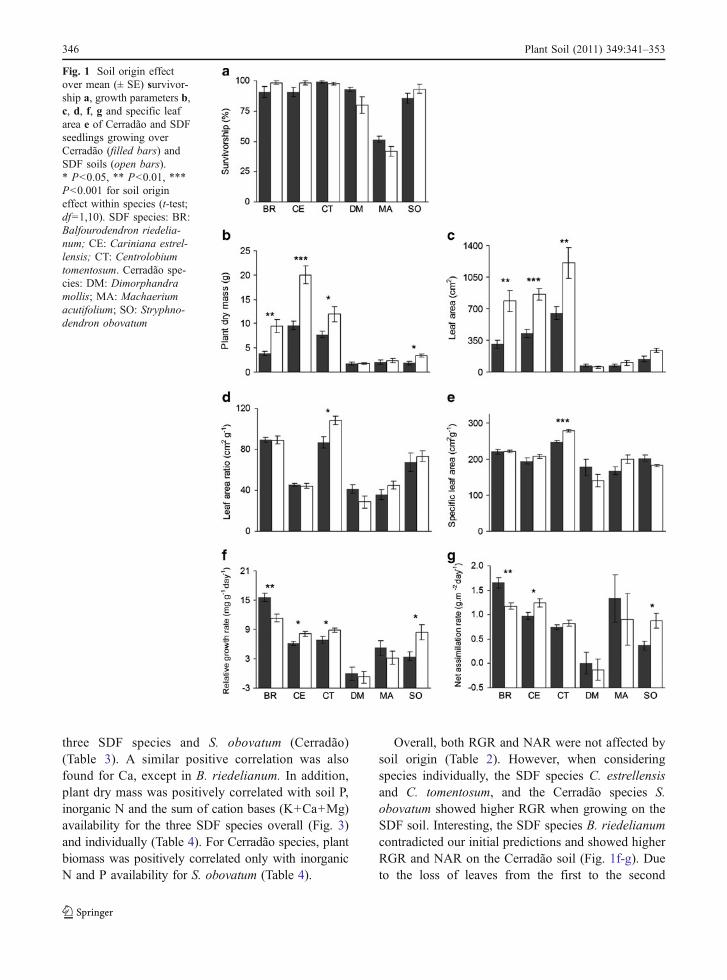
Cerradão and SDF soils are very different with respect tochemical properties and fertility (nutrient availability).SDF soils had lower Al concentrations and higherpH, organic matter, macronutrients, Zn and Mnavailability, cation exchange capacity and basesaturation than Cerradão soils (Table 1).
Seedlings performance
Seedling survivorship after 5 months, was significant-ly different among species (F 5,60=60.83; P<0.001)
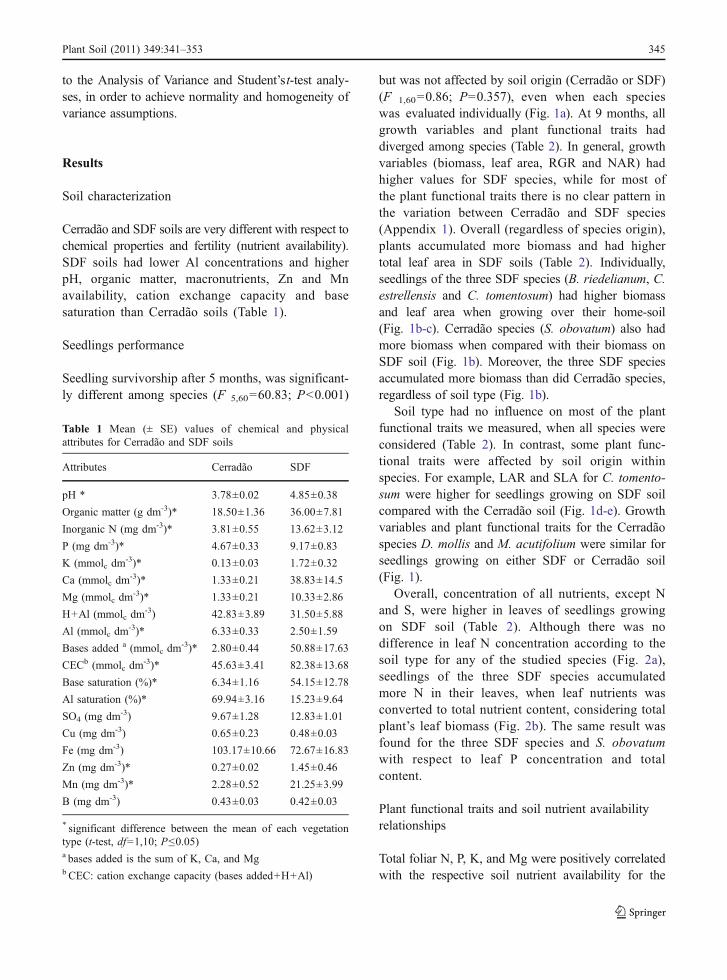
but was not affected by soil origin (Cerradão or SDF)(F 1,60=0.86; P=0.357), even when each specieswas evaluated individually (Fig. 1a). At 9 months, allgrowth variables and plant functional traits haddiverged among species (Table 2). In general, growthvariables (biomass, leaf area, RGR and NAR) hadhigher values for SDF species, while for most ofthe plant functional traits there is no clear pattern inthe variation between Cerradão and SDF species(Appendix 1). Overall (regardless of species origin),plants accumulated more biomass and had highertotal leaf area in SDF soils (Table 2). Individually,seedlings of the three SDF species (B. riedelianum, C.estrellensis and C. tomentosum) had higher biomassand leaf area when growing over their home-soil(Fig. 1b-c). Cerradão species (S. obovatum) also hadmore biomass when compared with their biomass onSDF soil (Fig. 1b). Moreover, the three SDF speciesaccumulated more biomass than did Cerradão species,regardless of soil type (Fig. 1b).
Soil type had no influence on most of the plantfunctional traits we measured, when all species wereconsidered (Table 2). In contrast, some plant func-tional traits were affected by soil origin withinspecies. For example, LAR and SLA for C. tomento-sum were higher for seedlings growing on SDF soilcompared with the Cerradão soil (Fig. 1d-e). Growthvariables and plant functional traits for the Cerradãospecies D. mollis and M. acutifolium were similar forseedlings growing on either SDF or Cerradão soil(Fig. 1).
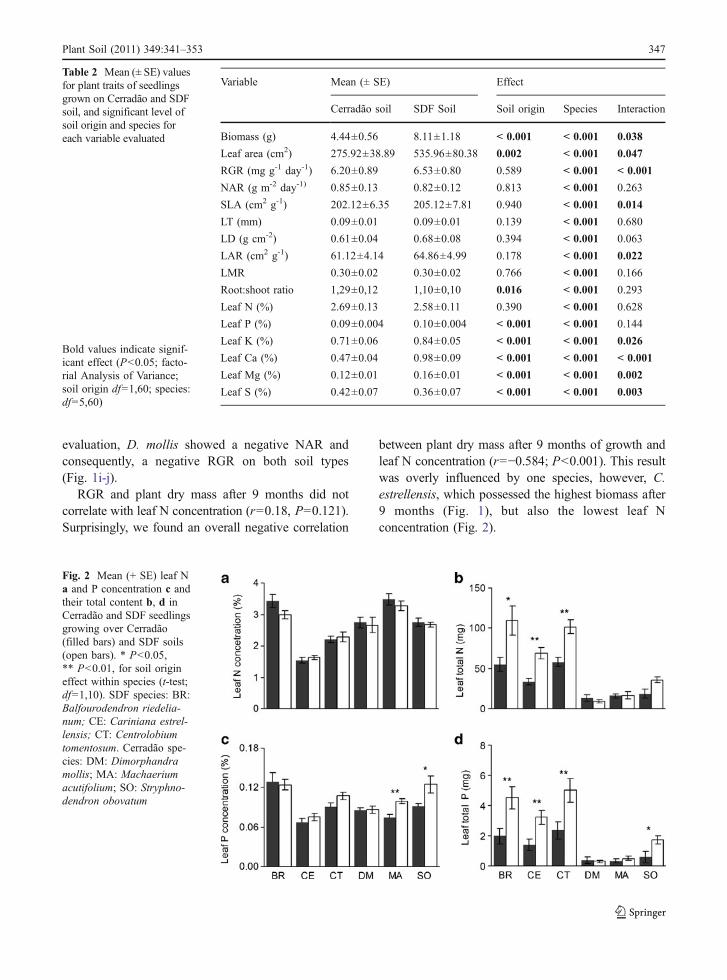
Overall, concentration of all nutrients, except Nand S, were higher in leaves of seedlings growingon SDF soil (Table 2). Although there was nodifference in leaf N concentration according to thesoil type for any of the studied species (Fig. 2a),seedlings of the three SDF species accumulatedmore N in their leaves, when leaf nutrients wasconverted to total nutrient content, considering totalplant’s leaf biomass (Fig. 2b). The same result wasfound for the three SDF species and S. obovatumwith respect to leaf P concentration and totalcontent.
Plant functional traits and soil nutrient availabilityrelationships
Total foliar N, P, K, and Mg were positively correlatedwith the respective soil nutrient availability for the
Table 1 Mean (± SE) values of chemical and physicalattributes for Cerradão and SDF soils
Attributes Cerradão SDF
pH * 3.78±0.02 4.85±0.38
Organic matter (g dm-3)* 18.50±1.36 36.00±7.81
Inorganic N (mg dm-3)* 3.81±0.55 13.62±3.12
P (mg dm-3)* 4.67±0.33 9.17±0.83
K (mmolc dm-3)* 0.13±0.03 1.72±0.32
Ca (mmolc dm-3)* 1.33±0.21 38.83±14.5
Mg (mmolc dm-3)* 1.33±0.21 10.33±2.86
H+Al (mmolc dm-3) 42.83±3.89 31.50±5.88
Al (mmolc dm-3)* 6.33±0.33 2.50±1.59
Bases added a (mmolc dm-3)* 2.80±0.44 50.88±17.63
CECb (mmolc dm-3)* 45.63±3.41 82.38±13.68
Base saturation (%)* 6.34±1.16 54.15±12.78
Al saturation (%)* 69.94±3.16 15.23±9.64
SO4 (mg dm-3) 9.67±1.28 12.83±1.01
Cu (mg dm-3) 0.65±0.23 0.48±0.03
Fe (mg dm-3) 103.17±10.66 72.67±16.83
Zn (mg dm-3)* 0.27±0.02 1.45±0.46
Mn (mg dm-3)* 2.28±0.52 21.25±3.99
B (mg dm-3) 0.43±0.03 0.42±0.03
* significant difference between the mean of each vegetationtype (t-test, df=1,10; P≤0.05)a bases added is the sum of K, Ca, and Mgb CEC: cation exchange capacity (bases added+H+Al)
Plant Soil (2011) 349:341–353 345
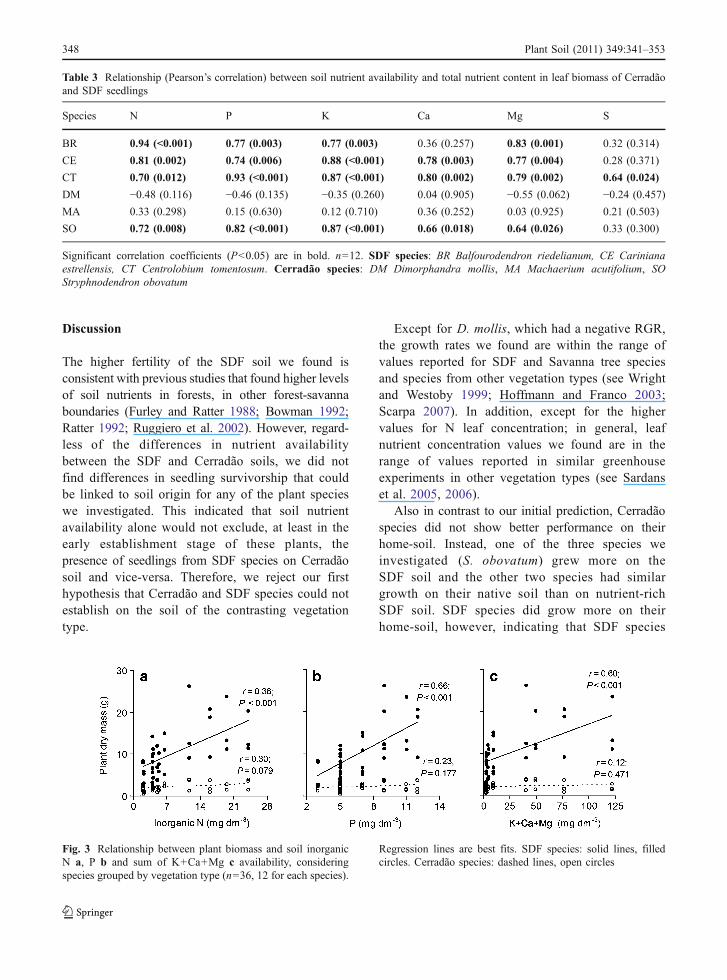
three SDF species and S. obovatum (Cerradão)(Table 3). A similar positive correlation was alsofound for Ca, except in B. riedelianum. In addition,plant dry mass was positively correlated with soil P,inorganic N and the sum of cation bases (K+Ca+Mg)availability for the three SDF species overall (Fig. 3)and individually (Table 4). For Cerradão species, plantbiomass was positively correlated only with inorganicN and P availability for S. obovatum (Table 4).
Overall, both RGR and NAR were not affected bysoil origin (Table 2). However, when consideringspecies individually, the SDF species C. estrellensisand C. tomentosum, and the Cerradão species S.obovatum showed higher RGR when growing on theSDF soil. Interesting, the SDF species B. riedelianumcontradicted our initial predictions and showed higherRGR and NAR on the Cerradão soil (Fig. 1f-g). Dueto the loss of leaves from the first to the second
Fig. 1 Soil origin effectover mean (± SE) survivor-ship a, growth parameters b,c, d, f, g and specific leafarea e of Cerradão and SDFseedlings growing overCerradão (filled bars) andSDF soils (open bars).* P<0.05, ** P<0.01, ***P<0.001 for soil origineffect within species (t-test;df=1,10). SDF species: BR:Balfourodendron riedelia-num; CE: Cariniana estrel-lensis; CT: Centrolobiumtomentosum. Cerradão spe-cies: DM: Dimorphandramollis; MA: Machaeriumacutifolium; SO: Stryphno-dendron obovatum
346 Plant Soil (2011) 349:341–353
evaluation, D. mollis showed a negative NAR andconsequently, a negative RGR on both soil types(Fig. 1i-j).
RGR and plant dry mass after 9 months did notcorrelate with leaf N concentration (r=0.18, P=0.121).Surprisingly, we found an overall negative correlation
between plant dry mass after 9 months of growth andleaf N concentration (r=−0.584; P<0.001). This resultwas overly influenced by one species, however, C.estrellensis, which possessed the highest biomass after9 months (Fig. 1), but also the lowest leaf Nconcentration (Fig. 2).
Variable Mean (± SE) Effect
Cerradão soil SDF Soil Soil origin Species Interaction
Biomass (g) 4.44±0.56 8.11±1.18 < 0.001 < 0.001 0.038
Leaf area (cm2) 275.92±38.89 535.96±80.38 0.002 < 0.001 0.047
RGR (mg g-1 day-1) 6.20±0.89 6.53±0.80 0.589 < 0.001 < 0.001
NAR (g m-2 day-1) 0.85±0.13 0.82±0.12 0.813 < 0.001 0.263
SLA (cm2 g-1) 202.12±6.35 205.12±7.81 0.940 < 0.001 0.014
LT (mm) 0.09±0.01 0.09±0.01 0.139 < 0.001 0.680
LD (g cm-2) 0.61±0.04 0.68±0.08 0.394 < 0.001 0.063
LAR (cm2 g-1) 61.12±4.14 64.86±4.99 0.178 < 0.001 0.022
LMR 0.30±0.02 0.30±0.02 0.766 < 0.001 0.166
Root:shoot ratio 1,29±0,12 1,10±0,10 0.016 < 0.001 0.293
Leaf N (%) 2.69±0.13 2.58±0.11 0.390 < 0.001 0.628
Leaf P (%) 0.09±0.004 0.10±0.004 < 0.001 < 0.001 0.144
Leaf K (%) 0.71±0.06 0.84±0.05 < 0.001 < 0.001 0.026
Leaf Ca (%) 0.47±0.04 0.98±0.09 < 0.001 < 0.001 < 0.001
Leaf Mg (%) 0.12±0.01 0.16±0.01 < 0.001 < 0.001 0.002
Leaf S (%) 0.42±0.07 0.36±0.07 < 0.001 < 0.001 0.003
Table 2 Mean (± SE) valuesfor plant traits of seedlingsgrown on Cerradão and SDFsoil, and significant level ofsoil origin and species foreach variable evaluated
Bold values indicate signif-icant effect (P<0.05; facto-rial Analysis of Variance;soil origin df=1,60; species:df=5,60)
Fig. 2 Mean (+ SE) leaf Na and P concentration c andtheir total content b, d inCerradão and SDF seedlingsgrowing over Cerradão(filled bars) and SDF soils(open bars). * P<0.05,** P<0.01, for soil origineffect within species (t-test;df=1,10). SDF species: BR:Balfourodendron riedelia-num; CE: Cariniana estrel-lensis; CT: Centrolobiumtomentosum. Cerradão spe-cies: DM: Dimorphandramollis; MA: Machaeriumacutifolium; SO: Stryphno-dendron obovatum
Plant Soil (2011) 349:341–353 347
Discussion
The higher fertility of the SDF soil we found isconsistent with previous studies that found higher levelsof soil nutrients in forests, in other forest-savannaboundaries (Furley and Ratter 1988; Bowman 1992;Ratter 1992; Ruggiero et al. 2002). However, regard-less of the differences in nutrient availabilitybetween the SDF and Cerradão soils, we did notfind differences in seedling survivorship that couldbe linked to soil origin for any of the plant specieswe investigated. This indicated that soil nutrientavailability alone would not exclude, at least in theearly establishment stage of these plants, thepresence of seedlings from SDF species on Cerradãosoil and vice-versa. Therefore, we reject our firsthypothesis that Cerradão and SDF species could notestablish on the soil of the contrasting vegetationtype.
Except for D. mollis, which had a negative RGR,the growth rates we found are within the range ofvalues reported for SDF and Savanna tree speciesand species from other vegetation types (see Wrightand Westoby 1999; Hoffmann and Franco 2003;Scarpa 2007). In addition, except for the highervalues for N leaf concentration; in general, leafnutrient concentration values we found are in therange of values reported in similar greenhouseexperiments in other vegetation types (see Sardanset al. 2005, 2006).
Also in contrast to our initial prediction, Cerradãospecies did not show better performance on theirhome-soil. Instead, one of the three species weinvestigated (S. obovatum) grew more on theSDF soil and the other two species had similargrowth on their native soil than on nutrient-richSDF soil. SDF species did grow more on theirhome-soil, however, indicating that SDF species
Table 3 Relationship (Pearson’s correlation) between soil nutrient availability and total nutrient content in leaf biomass of Cerradãoand SDF seedlings
Species N P K Ca Mg S
BR 0.94 (<0.001) 0.77 (0.003) 0.77 (0.003) 0.36 (0.257) 0.83 (0.001) 0.32 (0.314)
CE 0.81 (0.002) 0.74 (0.006) 0.88 (<0.001) 0.78 (0.003) 0.77 (0.004) 0.28 (0.371)
CT 0.70 (0.012) 0.93 (<0.001) 0.87 (<0.001) 0.80 (0.002) 0.79 (0.002) 0.64 (0.024)
DM −0.48 (0.116) −0.46 (0.135) −0.35 (0.260) 0.04 (0.905) −0.55 (0.062) −0.24 (0.457)
MA 0.33 (0.298) 0.15 (0.630) 0.12 (0.710) 0.36 (0.252) 0.03 (0.925) 0.21 (0.503)
SO 0.72 (0.008) 0.82 (<0.001) 0.87 (<0.001) 0.66 (0.018) 0.64 (0.026) 0.33 (0.300)
Significant correlation coefficients (P<0.05) are in bold. n=12. SDF species: BR Balfourodendron riedelianum, CE Carinianaestrellensis, CT Centrolobium tomentosum. Cerradão species: DM Dimorphandra mollis, MA Machaerium acutifolium, SOStryphnodendron obovatum
Fig. 3 Relationship between plant biomass and soil inorganicN a, P b and sum of K+Ca+Mg c availability, consideringspecies grouped by vegetation type (n=36, 12 for each species).
Regression lines are best fits. SDF species: solid lines, filledcircles. Cerradão species: dashed lines, open circles
348 Plant Soil (2011) 349:341–353

show some level of home-soil advantage. Thus, theresults we found with respect to growth on the twosoils, confirms only part of our initial predictionthat seedlings would perform better on theirhome-soil. This hypothesis was confirmed forSDF species, which performed better in theirmore fertile soils, but was rejected for savanna species,because no Cerradão species showed a home-soiladvantage.
For two of the three Cerradão species, ourfindings did not corroborate previous studies, whichdemonstrated that savanna species commonly re-spond positively to higher levels of nutrientavailability (Haridasan 2000; Barger et al. 2002;Kozovits et al. 2007). However, our results docorroborate the ones found for grasses in Africansavannas (Wang et al. 2010) and support thegeneral hypothesis that growth of wild species fromnutrient-poor habitats may not always respond tonutrient additions or higher soil fertility (Chapin etal. 1986). It is also possible that the physiologicaland morphological adaptations of Cerradão speciesfor being able to grow on nutrient-poor Cerradãosoils restrict them to respond when more soilnutrients are made available. Indeed, the ability ofa plant to tolerate stress is frequently inverselycorrelated with its competitive ability (Fine et al.2004; Liancourt et al. 2005; Brenes-Arguedas et al.2008).
Our results showed that SDF species have bothhigher growth rates than Cerradão species inCerradão soil and home-soil advantage (Fig. 1b).The higher growth rates of SDF species compared
to Cerradão species, even on Cerradão soils,corroborate other studies which found that whenother factors are controlled, species from morefertile soils tend to show a better growth responsenot only on their home-soil but also on contrastingsoils with lower nutrient availability (Fine et al.2004). These findings suggest that other factorsmay be interacting with soil fertility in order toprevent SDF species from outcompeting Cerradãospecies on savanna patches. Beyond soil nutrientavailability, other characteristics such as wateravailability (Bowman and Panton 1993; Hoffmannet al. 2004), light intensity (Hoffmann and Franco2003), fire occurrence (Hoffmann et al. 2003), andbiological interactions (Bowman and Panton 1993)have been suggested to be important factorscontrolling the establishment of seedlings and alsoinfluencing the dynamics of forest and savannaboundaries. However, the home-soil advantage ofSDF species does indicate that the lower nutrientavailability of the savanna soil reduces the growthof SDF seedlings, which in turn affects theirpotential to accumulate biomass at savanna sites.Therefore, our result may not lend support to theidea that soil nutrient availability does not inhibitdevelopment of forests on nutrient-poor soils, asproposed by Bond (2010).
One important point to be considered is that weevaluated only plants in their early stage of life.According to Aerts and Chapin (2000), fast-growing species, such as SDF trees, tend to havehigher biomass increments early in life comparedwith slow growing species, even in nutrient-poorenvironments. However, in the long-term andunder nutrient-limited conditions, species withlow nutrient loss rates can outcompete specieswith high nutrient loss rates, even when thesespecies have a greater competitive ability for nutrientuptake (Berendse 1994).
All leaf traits, including leaf nutrient concentration,varied significantly among species (Table 2, Appendix 1).Leaf functional traits are widely variable accordingto limitations imposed by the environment andhence commonly reflect plants resource investmentstrategies and aspects of species life history (Wrightet al. 2004; Poorter and Bongers 2006). Severalstudies have also reported that leaf functional traitscan be highly plastic when plants are grown inenvironments with different levels of resources
Table 4 Relationship (Pearson’s correlation) between soilnutrient availability and seedlings biomass after 9 months
Species Inorganic N P Sum of K+Ca+Mg
BR 0.96 (<0.001) 0.87 (<0.001) 0.89 (<0.001)
CE 0.81 (<0.001) 0.88 (<0.001) 0.7 (0.002)
CT 0.65 (0.023) 0.79 (0.002) 0.64 (0.024)
DM −0.25 (0.429) −0.13 (0.688) −0.22 (0.498)
MA 0.22 (0.494) −0.06 (0.844) −0.1 (0.747)
SO 0.72 (0.009) 0.74 (0.006) 0.54 (0.072)
Significant correlation coefficients (P<0.05) are in bold. n=12.SDF species: BR Balfourodendron riedelianum, CE Carinianaestrellensis, CT Centrolobium tomentosum. Cerradão species:DM Dimorphandra mollis; MA Machaerium acutifolium, SOStryphnodendron obovatum
Plant Soil (2011) 349:341–353 349

availability (Witkowski and Lamont 1991; Sultan2000; Hoffmann and Franco 2003, Kozovits et al.2007). In contrast, in our study most of the leaffunctional traits, including foliar N concentration,did not change in response to soil nutrient availabil-ity (two soil origins) (Table 2, Fig. 1). Despite thislack of plasticity in leaf functional traits, four ofthe six species we studied did increase their totalleaf area and accumulated more biomass whengrowing on the soil with higher nutrient avail-ability (SDF soil) (Table 2, Fig. 1). These datastrongly suggest that in some cases, the analysis ofonly a subset of leaf functional traits, such as SLA,LAR and foliar N concentration, may be not asinformative as an integrative analysis of plantgrowth performance that includes analysis of plantbiomass and total leaf area. In addition, speciesincreased their root:shoot ratio when were grown onthe nutrient-poor Cerradão soil, suggesting mor-phological plasticity of roots in response to varia-tion in nutrient availability (Jackson et al. 1990;Wang et al. 2006). However, we did not find aconsistent pattern of higher root:shoot ratio forsavanna species in comparison with forest species(Appendix 1), as found in other studies (Hoffmannet al. 2004).
Cerradão and SDF species have clearly con-trasting growth strategies. SDF species increasedtheir growth with increases in nutrient availability(Fig. 3). According to the growth strategies de-scribed by Grime (1977), SDF species behaved ascompetitors, rapidly accumulating abovegroundbiomass and nutrients. These species evolved ina more competitive environment with highernutrient and lower light availability, where fast-growing species with higher initial growth tend tobe favored. In our study we saw that Cerradãospecies growth strategies were more variable; S.obovatum behaved similarly to SDF species, in-creasing biomass when more nutrients were avail-able. On the other hand, D. mollis and M.acutifolium did not increase their growth withincreasing nutrient availability, suggesting a “stresstolerant” behavior (Fig. 3). In fact, the Cerradãoenvironment poses greater limitations for plantgrowth, such as lower nutrient availability andtherefore, species which can tolerate these condi-tions will be more successful. These results confirm, in
part, that species from nutrient-rich environments(in our case SDF) tend to be fast-growing specieswith higher relative growth rates and positiveresponse to increases in nutrient availability, where-as species from nutrient-poor habitats (Cerradão)tend to possess traits typical of slow-growing orstress-tolerant species (Aerts 1999; Aerts and Chapin2000).
In conclusion, our results demonstrated that Cerradãoand SDF soils are remarkably different withrespect to soil nutrient availability. We also foundthat SDF and Cerradão species tend to havedistinct growth strategies with respect to soilnutrient availability. Collectively, our results indi-cate that the chemical attributes of the upper soillayer do not exclude the occurrence of seedlings ofSDF woody species in savanna patches and vice-versa, at least in an initial stage of life. It ispossible that the mechanisms controlling establish-ment of Cerradão and SDF seedlings in contactzones may result from the interaction of soilchemical attributes with other site factors such asherbivory, soil water availability, fire and/or lightintensity. However, our findings also indicate thatthe upper soil layer has an effect on growthperformance of SDF species, with these speciesdemonstrating some home-soil advantage. Thissuggests that savanna soil reduces growth of forestspecies. Finally, in order to confirm our findingsand improve our knowledge about the filters thateffectively maintain contrasting floras in theseforest-savanna boundaries, we suggest reciprocaltransplanting and competitive field experimentstesting soil and its interaction with other factorssuch as herbivory, water availability, fire and/orlight intensity. A more detailed understanding ofthe interactions between vegetation, climate andedaphic factors is essential to better predict theimpact of future changes in climate in the forest/savanna distribution.
Acknowledgments This work was supported by a Coordena-ção de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)doctoral scholarship and by a Conselho Nacional de Desenvol-vimento Científico e Tecnológico (CNPq) “sandwich” pro-gram scholarship (200845/2008-0) from the Government ofBrazil, for R.A.G. Viani. Authors are thankful for PedroBrancalion for helpful comments in an earlier version of themanuscript.
350 Plant Soil (2011) 349:341–353

Appendix
References
Aerts R, Chapin FS III (2000) The mineral nutrition of wildplants revisited: a re-evaluation of processes and patterns.Adv Ecol Res 30:1–67
Aerts R (1999) Interspecific competition in natural plantcommunities: mechanisms, trade-offs and plant-soil feed-backs. J Ex Bot 50:29–37
Allen SE (1989) Chemical analysis of ecological materials, 2ndedn. Blackwell Scientific Publications, Oxford
Baltzer JL, Thomas SC, Nilus R, Burslem DFRP (2005)Edaphic specialization in tropical trees: physiologicalcorrelates and responses to reciprocal transplantation.Ecology 86:3063–3077. doi:10.1890/04-0598
Banfai DS, Bowman DMJS (2005) Dynamics of a savanna-forest in the Australian monsoon tropics inferred fromstand structures and historical aerial photography. Aust JBot 53:185–194. doi:10.1071/BT04141
Barger NN, D’Antonio CM, Ghneim T, Brink K, Cuevas E(2002) Nutrient limitation to primary productivity in asecondary Savanna in Venezuela. Biotropica 34:493–501
Berendse F (1994) Competition between plant populations atlow and high nutrient supplies. Oikos 71:253–260
Bond WJ (2010) Do nutrient-poor soils inhibit development offorests? A nutrient stock analysis. Plant Soil 334:47–60.doi:10.1007/s11104-010-0440-0
Bowman DMJS, Panton WJ (1993) Factors that controlmonsoon-rainforest seedling establishment and growth inNorth Australian Eucalyptus Savanna. J Ecol 81:297–304
Bowman DMSJ (1992) Monsoon Forests in North-westernAustralia. II* Forest-Savanna Transitions. Aust J Bot40:89–102
Brenes-Arguedas T, Ríos M, Rivas-Torres G, Blundo C, Coley PD,Kursar TA (2008) The effect of soil on the growth performanceof tropical species with contrasting distributions. Oikos117:1453–1460. doi:10.1111/j.2008.0030-1299.16903.x
Chapin FS III, Vitousek PM, Van Cleve K (1986) The nature ofnutrient limitation in plant communities. AmNat 127(1):48–58
Durigan G, Ratter JA (2006) Succesional changes in Cerradoand Cerrado/Forest Ecotonal vegetation in western SãoPaulo State, Brazil, 1962–2000. Edinb J Bot 63:119–130.doi:10.1017/S0960428606000357
EMBRAPA—Empresa Brasileira de Pesquisa Agropecuária(1997) Manual de métodos de análise de solo, 2nd edn.Centro Nacional de Pesquisa de Solos, Rio de Janeiro
Faria SM, Franco A, Menandro MS, Jesus RM, Baitello JB,Aguiar OT, Dobereiner J (1984) Levantamento da nodulação
Table 5 Mean (± SE) values for plant traits of tree seedlings grownon a greenhouse experiment with Cerradão and SDF soil. Withinlines, means followed by different letters indicate significantdifferences between species (P<0.05, two-way Anova, followed
by Tukey’s post-hoc test). SDF species: BR: Balfourodendronriedelianum; CE: Cariniana estrellensis; CT: Centrolobiumtomentosum. Cerradão species: DM: Dimorphandra mollis; MA:Machaerium acutifolium; SO: Stryphnodendron obovatum
Variables SDF species Cerradão Species
BR CE CT DM MA SO
Biomass (g) 6.51±1.04 c 14.76±1.89 a 9.81±1.04 b 1.79±0.17d 2.25±0.3d 2.54±0.33d
Leaf area (cm2) 534.61±89.47 b 637.45±76.45 ab 928.36±121.98 a 64.74±11.2d 85.39±16.19d 185.1±25.92 c
RGR (mg g-1 day-1) 13.45±0.85 a 7.09±0.43 b 7.85±0.5 b −0.4±0.89d 4.24±1.02 c 5.98±1.14 bc
NAR (g m-2 day-1) 1.42±0.09 a 1.11±0.07 ab 0.78±0.04 bc −0.06±0.15d 1.12±0.35 ab 0.62±0.12 c
SLA (cm2 g-1) 220.95±3.97 b 201.47±5.5 bc 263.61±5.09 a 159.48±14.38d 183.87±9.06 c 192.35±5.43 c
LT (mm) 0.075±0.001 d 0.157±0.002 a 0.115±0.006 b 0.073±0.003 de 0.091±0.003 c 0.067±0.002 e
LD (g cm-2) 0.67±2.13 b 0.32±1.32 c 0.36±4.69 c 1.05±3.87 a 0.69±3.29 b 0.78±4.93 b
LAR (cm2 g-1) 89.34±4.56 ab 44.94±2.97 c 97.67±6.44 a 34.91±4.27d 40.5±3.28cd 70.58±5.86 b
LMR 0.4±0.01 a 0.22±0.01 b 0.37±0.01 a 0.22±0.02 b 0.21±0.02 b 0.37±0.03 a
Root:shoot ratio 0.54±0.03 e 1.44±0.1 b 0.83±0.08d 1.11±0.06 c 2.12±0.23 a 1.12±0.14 c
Leaf N (%) 3.21±1.31 a 1.59±0.56d 2.25±0.86 c 2.7±1.45 bc 3.38±1.21 a 2.72±0.74 b
Leaf P (%) 0.13±0.08 a 0.07±0.04d 0.1±0.05 bc 0.09±0.03 c 0.09±0.05 c 0.11±0.08 b
Leaf K (%) 1.21±0.56 a 0.62±0.81 c 0.77±0.46 b 0.48±0.48d 0.5±0.25cd 1.07±0.68 a
Leaf Ca (%) 0.41±0.41 b 0.46±1.65 b 0.9±1.38 a 0.46±0.8 b 1.08±1.49 a 1.06±1.19 a
Leaf Mg (%) 0.18±0.23 a 0.19±0.17 a 0.14±0.13 b 0.05±0.03d 0.1±0.04 c 0.16±0.08 ab
Leaf S (%) 0.36±0.25 b 0.16±0.05d 0.12±0.07 e 0.16±0.28 de 0.28±0.33 c 1.28±0.52 a
Plant Soil (2011) 349:341–353 351

de leguminosas florestais nativas na região sudeste do Brasil.Pesq Agropecu Bras 19:143–153
Fine PVA, Mesones I, Coley PD (2004) Herbivores promotehabitat specialization by trees in Amazonian forests.Science 305:663–665. doi:10.1126/science.1098982
Furley PA, Ratter JA (1988) Soil resources and plant commu-nities of the central Brazilian Cerrado and their develop-ment. J Biogeogr 15:97–108
Grime JP (1977) Evidence for the existence of three primarystrategies in plants and its relevance to ecological andevolutionary biology. Am Nat 111:1169–1194
Haridasan M (2000) Nutrição mineral de plantas nativas docerrado. Rev Bras Fisiol Veg 12:54–64
Hoffmann WA (2000) Post-establishment seedling success inthe Brazilian cerrado: a comparison of savanna and forestspecies. Biotropica 32:62–69
HoffmannWA, Franco AC (2003) Comparative growth analysis oftropical forest and savanna woody plants using phylogenet-ically independent contrasts. J Ecol 91:475–484
Hoffmann WA, Orthen B, Nascimento PKV (2003) Compara-tive fire ecology of tropical savanna and forest trees. FunctEcol 17:720–726
Hoffmann WA, Orthen B, Franco AC (2004) Constrains toseedling success of savanna and forest trees across thesavanna-forest boundary. Oecologia 140:252–260.doi:10.1007/s00442-004-1595-2
Jackson RB, Manwaring JH, Caldwell MM (1990) Rapidphysiological adjustment of roots to localized soil enrich-ment. Nature 344:58–60
John R, Dalling JW, Harms KE, Yavitt JB, Stallard RF,Mirabello M, Hubbell SP, Valencia R, Navarrete H, VallejoM, Foster RB (2007) Soil nutrients influence spatialdistributions of tropical tree species. P Natl Acad Sci104:864–869
Juhász CEP, Cursi PR, Cooper M, Oliveira TC, Rodrigues RR(2006) Dinâmica físico-hídrica de uma toposseqüência desolos sob savana florestada (Cerradão) em Assis, SP. RevBras Cienc Solo 30:401–412
Kozovits AR, Bustamante MMC, Garofalo CR, Bucci S,Franco AC, Goldstein G, Meinzer FC (2007) Nutrientresorption and patterns of litter production and decompo-sition in a Neotropical Savanna. Funct Ecol 21:1034–1043. doi:10.1111/j.1365-2435.2007.01325.x
Kronka FJN, Nalon MA, Matsukuma CK, Kanashiro MSS,Pavao M, Shida CN, Joly CA, Couto HTZ, Baitello JB,Guillaumon JR (2005) Inventário florestal da vegetaçãonatural do Estado de São Paulo. Secretaria de Estado doMeio Ambiente, São Paulo
Lehmann CER, Archibald SA, Hoffmann WA, Bond WJ (2011)Deciphering the distribution of the savanna biome. NewPhytol. doi:10.1111/j.1469-8137.2011.03689.x
Liancourt P, Callaway RM, Michalet R (2005) Stress toleranceand competitive-response ability determine the outcome ofbiotic interactions. Ecology 86:1611–1618. doi:10.1890/04-1398
McGraw JB, Chapin FS III (1989) Competitive ability andadaptation to fertile and infertile soil in two Eriophorumspecies. Ecology 70:736–749
Myers N, Mittermeier RA, Mittermeier CG, Fonseca GAB,Kent J (2000) Biodiversity hotspots for conservationpriorities. Nature 403:853–858. doi:10.1038/35002501
Oliveira-Filho AT, Curi N, Vilela EA, Carvalho DA (2001)Variation in tree community composition and structurewith changes in soil properties within a fragment ofsemideciduous forest in South-eastern Brazil. Edinb J Bot58:139–158. doi:10.1017/S0960428601000506
Oliveira-Filho AT (2006) Catálogo das árvores nativas de MinasGerais: mapeamento e inventário da flora nativa e dosreflorestamentos de Minas Gerais. Editora UFLA, Lavras
Palmiotto PA, Davies SJ, Vogt KA, Ashton MS, Vogt D, AshtonP (2004) Soil-related habitat specialization in dipterocarprain forest tree species in Borneo. J Ecol 92:609–623
Poorter L, Bongers F (2006) Leaf traits are good predictors ofplant performance across 53 rain forest species. Ecology87:1733–1743
Ratter JA (1992) Transition between cerrado and forestvegetation in Brazil. In: Furley PA, Proctor J, Ratter JA(eds) Nature and dynamics of forest-savanna boundaries.Chapman and Hall, London, pp 417–429
Ribeiro JF, Walter BMT (1998) Fitofisionomias do BiomaCerrado. In: Sano S, Almeida S (eds) Cerrado: ambiente eflora. Embrapa-CPAC, Brasília, pp 89–166
Ruggiero PGC, Batalha MA, Pivello VR, Meirelles ST (2002)Soil-vegetation relationships in Cerrado (Brazilian savan-na) and semideciduous forest, Southeastern Brazil. PlantEcol 160:1–16
Sardans J, Peñuelas J, Roda F (2005) Changes in nutrient useefficiency, status and retranslocation in young post-fireregeneration Pinus halepensis in response to sudden N andP input, irrigation and removal of competing vegetation.Trees-Struct Funct 19:233–250. doi:10.1007/s00468-004-0374-3
Sardans J, Roda F, Peñuelas J (2006) Effects of a nutrient pulsesupply on nutrient status of the Mediterranean treesQuercus ilex subsp. ballota and Pinus halepensis ondifferent soils and under different competitive pressure.Trees 20:619–632. doi:10.1007/s00468-006-0077-z
Scarpa FM (2007) Estudo comparativo do crescimento deplântulas e dos atributos foliares em espécies do Cerrado eda Mata Atlântica. Campinas State University, Dissertation
Sultan SE (2000) Phenotypic plasticity for plant development,function and life history. Trends Plant Sci 5:537–542
Sylvester-Bradley R, Oliveira LA, Podesta-Filho JA, St. John TV(1980) Nodulation of legumes, nitrogenase activity of rootsand occurrence of nitrogen-fixing Azospirillum in representa-tive soils of Central Amazonia. Agro-Ecosystems 6:249–266
Theodose TA, Bowman WD (1997) Nutrient availability, plantabundance, and species diversity in two alpine tundracommunities. Ecology 78:1861–1872
van Raij B, Cantarella H, Quaggio JA, Andrade JC (2001)Análise Química para Avaliação da Fertilidade de SolosTropicais. Instituto Agronômico, Campinas
Veloso HP (1992) Manual técnico da vegetação brasileira.IBGE-Departamento de Recursos Naturais e EstudosAmbientais, Rio de Janeiro
Wang L, D'Odorico P, Ries L, Caylor K, Macko S (2010)Combined effects of soil moisture and nitrogen availabilityvariations on grass productivity in African savannas. Plantand Soil 328:95–108. doi:110.1007/s11104-11009-10085-z
Wang L, Mou PP, Jones RH (2006) Nutrient foraging viaphysiological and morphological plasticity in three plantspecies. Can J For Res 36:164–173. doi:10.1139/X05-239
352 Plant Soil (2011) 349:341–353

Wright IJ, Westoby M (1999) Differences in seedling growthbehaviour among species: trait correlations acrossspecies, and trait shifts along nutrient compared to raingradients. J Ecol 87:85–97
Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z,Bongers F, Cavender-Bares J, Chapin T, Cornelissen
JHC, Diemer M, Flexas J, Garnier E, Groom PK,Gulias J, Hikosaka K, Lamont BB, Lee T, Lee W,Lusk C, Midgley JJ, Navas ML, Niinemets U,Oleksyn J, Osada N, Poorter H, Poot P, Prior L,Pyankov VI, Roumet C, Thomas SC, Tjoelker MG,Veneklaas EJ, Villar R (2004) The worldwide leafeconomics spectrum. Nature 428:821–827. doi:10.1038/nature02403
Plant Soil (2011) 349:341–353 353
Witkowski ETF, Lamont BB (1991) Leaf specific mass confoundsleaf density and thickness. Oecologia 88:486–493





























