Embed Size (px)
Citation preview

Scheffersomyces cryptocercus: a new xylose-fermenting yeast associated with the gutof wood roaches and new combinations in the Sugiyamaella yeast clade
Hector UrbinaRobert FrankMeredith Blackwell1
Department of Biological Sciences, Louisiana StateUniversity, Baton Rouge, Louisiana 70803
Abstract: The gut of wood-feeding insects is amicrohabitat for a specialized community of mi-crobes, including bacteria and several groups ofeukaryotes such as nematodes, parabasalids andfungi. The characterization of gut yeast communitiesfrom a variety of insects has shown that certain yeastsoften are associated with the insects. The gut of wood-feeding insects is rich in ascomycete yeasts and inparticular xylose-fermenting (X-F) and assimilatingyeasts have been consistently present in the gut oflignicolous insects. The objective of this study was thecharacterization of the yeast flora from the gut of thewood roach Cryptocercus sp. (Blattodea: Cryptocerci-dae). Five wood roaches were collected along theAppalachian Trail near the border between Tennes-see and North Carolina, USA. We isolated 18 yeaststrains from the wood roaches identified as Sugiya-maella paludigena and Sugiyamaella lignohabitans,xylose-assimilating yeasts, and Scheffersomyces crypto-cercus (NRRL Y-48824T 5 CBS 12658) a new species ofX-F yeast. The presence of X-F and certain non X-Fyeasts in the gut of the subsocial wood roachCryptocercus sp. extends the previous findings ofassociations between certain ascomycete yeasts andlignicolous insects. New combinations were made for13 asexual members of the Sugiyamaella clade.
Key words: RPB1, Saccharomycotina, XYL1, xy-lose reductase
INTRODUCTION
In the past decade new information about themicrobial communities inhabiting the gut of insectshas been accumulating (Kurtzman et al. 2011).Among insects, the bacterial gut communities oftermites, beetles and wood roaches have been studiedextensively but the fungal communities are less wellknown. Most studies of gut inhabitants of woodroaches, Cryptocercus (Blattodea: Cryptocercidae),
have focused on enumeration of bacteria, especiallyendosymbiontic bacteria, and protists without men-tion of fungi that may have been present (Sacchi et al.1998; Clark and Kambhampati 2003; Kitade 2004;Maekawa et al. 2005; Noda et al. 2006; Klass et al.2008; Berlanga et al. 2009; Carpenter et al. 2009;Ohkuma et al. 2009; Carpenter et al. 2010, 2011).One study, however, reported the isolation of a yeastspecies from Cryptocercus punctulatus (Prillinger et al.1996, Prillinger and Konig 2006).
Investigations of the gut of wood-feeding beetles(Passalidae) recently led to the discovery of severaldistinct clades of ascomycete yeasts that are consis-tently associated with these beetles, perhaps asadaptations of the host feeding behavior and habitat(Suh and Blackwell 2004, Suh et al. 2005, Nguyen et al.2006, Suh et al. 2006, Berkov et al. 2007, Grunwaldet al. 2010, Hughes et al. 2011, Calderon and Berkov2012). In particular members of the Sugiyamaella,Scheffersomyces and Spathaspora clades have beenfound in association with passalids (Zhang et al.2003, Suh et al. 2004, Nguyen et al. 2006, House-knecht et al. 2011), as well as other wood-feedinginsects and woody plants (Berkov et al. 2007,Grunwald et al. 2010, Hughes et al. 2011). Speciesof Sugiyamaella are xylose-assimilating yeasts, andmembers of the Scheffersomyces and Spathaspora cladeshave attracted attention because they possess the rareability to ferment D-xylose, which gives them econom-ic potential for production of bioethanol from plantwaste residues (Agbogbo and Wenger 2006, 2007;Agbogbo and Coward-Kelly 2008). Members of theseclades occupy a habitat rich in xylose, subunits of theplant cell wall carbohydrate hemicellulose.
The primary aim of this study was to investigate thepossibility that yeasts are present in the gut of adistinctive subsocial wood roach Cryptocercus sp. thatinhabits rotting wood in the Appalachian Mountainsof the United States. We isolated 18 yeast strainsidentified as Sugiyamaella palludigena, Sugiyamaellalignohabitans, and a previously undescribed X-F yeastScheffersomyces cryptocercus (NRRL Y-48824T 5 CBS12658). The species described was based on a multi-locus phylogenetic analysis, biochemical and mor-phological characterization, and the nucleotide com-parison of the single copy gene XYL1 that encodes forxylose reductase (XR). In addition, new combinationswere made for 13 asexual members of the Sugiya-maella clade.
Submitted 26 Mar 2012; accepted for publication 9 Oct 2012.1 Corresponding author. E-mail: [email protected]
Mycologia, 105(3), 2013, pp. 650–660. DOI: 10.3852/12-094# 2013 by The Mycological Society of America, Lawrence, KS 66044-8897
650

MATERIALS AND METHODS
Insect collection and yeast isolation.—Seven individuals ofCryptocercus sp. were collected along the Appalachian Trail atNewfound Gap near the border between Tennessee andNorth Carolina, USA. The roaches were kept alive until timeof dissection. Individual roaches were superficially disinfectedby washing with 70% alcohol (5 min), 5% bleach (5 min) andsterile water (10 min) before gut removal. Each roach thenwas placed on a flame-sterilized coverslip, and sterile forcepswere used to separate the exoskeleton to remove the gut.
Yeast isolation and culture.—The gut was divided into threeregions: foregut-midgut (F-M), anterior hindgut (AHG) andposterior hindgut (PHG). Each region was homogenized in500 mL 0.7% saline solution with 0.01% Tween 80. Onehundred microliters of each homogenized region wereplated on YPDM (0.3% yeast extract, 0.5% Bacto peptone,1% dextrose, 0.3% malt extract, 2% agar) mediumsupplemented with 0.05% calcium carbonate, vitaminsand salts (Boundy-Mills 2006, Kurtzman et al. 2011). Themedium was acidified with 0.6 mL concentrated HCl perliter, and 0.35 mg/L chloramphenicol was added to reducebacterial growth. After 3 d incubation at room temperature,18 yeast colonies were selected for additional study based oncultural characteristics. The isolates were purified bymultiple subcultures and stored on YPDM agar plates andbroth cultures at 280 C in 15% glycerol. Other culturetechniques and biochemical characterizations were carriedout by the methods recommended by Yarrow (1998),Barnett et al. (2000), and Kurtzman et al. (2011).
Amplification and sequencing of DNA.—Genomic DNA wasextracted with a WizardH Genomic DNA purification kit(Promega). The concentration, integrity and purity of totalextracted DNA were confirmed by gel electrophoresis in0.8% agarose in 0.53 Tris-Borate-EDTA (TBE) buffer. Initialrapid identification was carried out by PCR amplification andsequencing of , 600 bp of the D1/D2 region of the largesubunit rRNA gene (LSU) for use in BLAST queries(Kurtzman and Robnett 1998, Kurtzman et al. 2011). Toincrease the robustness of the phylogenetic analyses and theaccuracy of species recognition, PCR amplification of thesmall subunit of the rRNA gene (SSU , 1.6 Kbp) andinternal transcribed spacers 1 and 2 (ITS , 500 bp), andRPB1 loci were carried out in addition to the LSU (Whiteet al. 1990; Hibbett 1996; Schoch et al. 2009, 2012; Urbinaand Blackwell 2012). This multilocus dataset has been shownto generate a well supported phylogenetic reconstruction forthe Scheffersomyces clade (Urbina and Blackwell 2012). Thepurified PCR products were sequenced in both directions byBeckman Coulter Genomics (Danvers, Massachusetts).
RAPD-PCR fingerprinting.—RAPD–PCR analysis was per-formed with the oligonucleotide primer CDU (59–GCGATCCCCA–39) (Sullivan et al. 1995; Fadda et al.2004, 2010; Urbina and Blackwell 2012). Aliquots of 25 mLof amplified product were analyzed with electrophoresis on1.8% agarose gel in 13 TBE buffer with 13 SYBRH SafeDNA Gel Stain (Invitrogen, Grand Island, New York) at 70 Vfor 80 min. DNA fragments were viewed with a UV-lighttransilluminator and photographed with a Polaroid system.
Xylose reductase (XR) studies.—Use of the XYL1 locusrecently was recommended for rapid identification of crypticspecies in the Scheffersomyces clade (Urbina and Blackwell2012). The , 600 bp fragment of XYL1 was amplified withthe following degenerate primers, XYL1-forward (59–GGTYT-TYGGMTGYTGGAARSTC–39) and XYL1-reverse (59–AAW-GATTGWGGWCCRAAWGAWGA–39) and the PCR protocolas recommended (Urbina and Blackwell 2012).
Phylogenetic analysis.—Contig sequences and sequencingmanipulations were done with Se-AL 2.01a11 (http://tree.bio.ed.ac.uk/software/seal/) and Mesquite 2.74 (Maddisonand Maddison 2005). The sequences were aligned in theonline interface MAFFT 6.859 (http://mafft.cbrc.jp/alignment/software/) with different advanced alignmentstrategies per locus: LSU and RPB1, global homology (G-INS-i); ITS, one conserved domain (L-INS-i); and SSU,mitochondrial small subunit (MtSm) rRNA, and cyto-chrome oxidase II (COXII), secondary structure of RNA(Q-INS-i). In particular ITS loci were realigned with thesoftware SATe 2.1.2 (Liu et al. 2012), and ambiguoussequence alignment ends were eliminated in all thealignments. Maximum likelihood (ML) phylogenic inferencewas performed in RAxML-VI-HPC (Stamatakis 2006) with apartitioned multilocus matrix (each partition for SSU, LSU,ITS1, 5.8S, ITS2, COXII, MtSm, RPB1) under a general timereversible model with a gamma distribution of site ratevariation (GTRGAMMA). ML support was estimated with1000 bootstrap replicates. Alignments and trees were depos-ited in TreeBASE (accession number S13305).The sequencesfor RPB1 and XR were obtained with SEQUIN 11.9 (http://www.ncbi.nlm.nih.gov/Sequin/) with an alternative yeastnuclear codon bias (Ohama et al. 1993, Wohlbach et al.2011). Tree editing was done with FigTree 1.3.1 software(http://tree.bio.ed.ac.uk/software/figtree/). GenBank se-quence accession numbers are provided (TABLE I).
RESULTS
The yeast strains were identified initially with LSU andITS loci in BLAST queries (Altschul et al. 1990).Sugiyamaella paludigena and Su. lignohabitans wereidentified as members of the gut community ofCryptocercus sp. (FIG. 1). Sugiyamaella paludigenaoriginally was isolated from high moor peat nearMoscow, Russia (Golubev et al. 1981), and morerecently it was reported from rhizosphere soil of whitespruce (Lamarche et al. 2011). Sugiyamaella lignoha-bitans first was described from a decaying log of paperbirch and associated insect frass in USA (Kurtzman2007). In general Sugiyamaella species have beenfound in association with wood and insects, and Su.bullrunensis was described from the gut of wood-feeding beetle Odontotaenius disjunctus (Houseknechtet al. 2011) as was indicated by a molecular cloning studyof the same beetle (Zhang et al. 2003). Altogether, thesefindings suggest that Sugiyamaella species are widelydistributed in wood, soil and the gut of insects.
URBINA ET AL.: YEASTS FROM A WOOD ROACH 651

TABLE I. GenBank accession numbers of the nucleotide sequences used in this study
Species Code
Loci
LSU ITS COXII MtSm
B. illinoisensisT NRRL YB-1343 DQ442696 DQ898169 DQ443103 DQ442775B. malaysiensisT NRRL Y-6417 DQ442695 DQ898170 DQ443102 DQ442774T. ciferriT NRRL Y-10943 DQ442681 AY493435 DQ443088 DQ442760W. domercqiaeT NRRL Y-6692 DQ438240 DQ911462 DQ443084 DQ442756Z. hellenicusT NRRL Y-7136 DQ438216 DQ911464 DQ443047 DQ442719Su. americanaT NRRL YB-2067 DQ438193 HM208605 DQ443051 DQ442723Su. boreocaroliniensisT NRRL YB-1835 DQ438221 DQ911448 DQ443065 DQ442737Su. bullrunensisT EH008 HM208602 HM208601 – –Su. castrensisT NRRL Y-17329 DQ438195 DQ911441 DQ443053 DQ442725Su. chiloensisT NRRL Y-17643 DQ911433 – DQ911437 –Su. floridensisT NRRL YB-3827 DQ438222 DQ911442 DQ443066 DQ442738Su. grinbergsiiT NRRL Y-27117 DQ438199 DQ911443 DQ443057 DQ442729Su. japonicaT NRRL YB-2798 DQ438202 DQ911456 DQ443060 DQ442732Su. lignohabitansT NRRL YB-1473 DQ438198 HM208611 DQ443056 DQ442728Su. lignohabitans RG11-03-01-05-F5 JQ714004 JQ713960 – –
RG11-03-01-06-A4 JQ714007 JQ713963 – –RG11-03-01-06-A2 JQ714006 JQ713962 – –RG11-03-01-05-F2 JQ714003 JQ713959 – –RG11-03-01-05-M4 JQ714005 JQ713961 – –RG11-03-01-05-F1 JQ714002 JQ713958 – –
Su. marilandicaT NRRL YB-1847 DQ438219 DQ911445 DQ443063 DQ442728Su. marionensisT NRRL YB-1336 DQ438197 DQ911452 DQ443055 DQ442727Su. neomexicanaT NRRL YB-2450 DQ438201 DQ911447 DQ443059 DQ442731Su. novakiiT NRRL Y-27346 DQ438196 DQ911449 DQ443054 DQ442726Su. paludigenaT NRRL Y-12697 DQ438194 DQ911451 DQ443052 DQ442724Su. paludigena RG11-03-01-06-FM2 JQ714009 JQ713965 – –
RG11-03-01-05-A1 JQ714008 JQ713964 – –Su. pinicolaT NRRL YB-2263 DQ438200 DQ911453 DQ443058 DQ442730Su. smithiaeI NRRL Y-17850 DQ438218 HM208613 DQ443062 DQ442734Su. valdivianaT NRRL Y-7791 DQ438220 HM461646 DQ443064 DQ442736
SSU ITS LSU RPB1 XYL1C. bolitotheriT NRRL Y-27587 AY242142 FJ623599 AY242249 JN804828 –C. maxiiT NRRL Y-27588 AY242144 – AY242253 JN804830 –C. chickasawarumT NRRL Y-27566 AY242154 FJ172247 JQ025398 JN804829 –C. yuchorumT NRRL Y-27569 AY242169 JN943266 AY242278 JN804832 –C. terraborumT NRRL Y-27573 AY426956 FJ623596 AY309810 JN804831 –C. panamericanaT NRRL Y-27567 AY426960 FJ623601 AY309872 JN804835 –L. elongisporusT NRRL YB-4239 HQ876033 HQ876042 HQ876050 AY653537 –C. tropicalisT NRRL Y-12968 EU348785 AB437068 U45749 – –Sc. ergatensisT NRRL Y-17652 AB013524 EU343826 U45746 EU344098 JQ436926Sc. insectosaT NRRL Y-12854 AB013583 HQ652064 FM200041 JN804842 JQ235697Sc. lignosusT NRRL Y-12856 HQ651941 JN943262 U45772 JN804837 JQ235693Sc. segobiensisT NRRL Y-11571 AB054288 DQ409166 U45742 EF599429 JQ436925Sc. queiroziaeT NRRL Y-48722 – HM566445 HM566445 – –Sc. gosingicusT CBS 11433 HQ876040 HQ999978 HQ999955 – –Sc. spartinaeT NRRL Y-7322 FJ153139 HQ876044 U45764 – –Sc. stipitisT NRRL Y-7124 AB054280 JN943257 U45741 JN804841 JQ235696Sc. amazonensisT NRRL Y-48762 – JF826438 JF826438 – –Sc. shehataeT NRRL Y-12858 AB013582 JN943264 AF178049 JQ436927 JQ235691Sc. quercinusT NRRL Y-48825 JN940981 JN943260 JN703957 JN804838 JQ008829Sc. virginianusT NRRL Y-48822 JN940969 JN943259 JN703958 JN804839 JQ235695Sc. illinoinensisT NRRL Y-48827 JN940968 JN943261 JN703959 JN804840 JQ235694
652 MYCOLOGIA

TAXONOMY
REVISION OF THE SUGIYAMAELLA YEAST CLADE
We propose additional new combinations in the genusSugiyamaella by including clade members that previouslywere described as species of the polyphyletic asexual genusCandida based on a multilocus phylogenetic analysis with85% bootstrap support (FIG. 1) (see Knapp et al. 2011).
Sugiyamaella boreocaroliniensis (Kurtzman) H.Urbina & M. Blackw., comb. nov.
MycoBank MB564463Basionym: Candida boreocaroliniensis Kurtzman,
FEMS Yeast Res 7:1054, 2007
Sugiyamaella bullrunensis (S.O. Suh, Houseknecht &J.J. Zhou) H. Urbina & M. Blackw., comb. nov.
MycoBank MB564464Basionym: Candida bullrunensis S.O. Suh, House-
knecht & J.J. Zhou. Int J Syst Evol Microbiol 61:1752, 2011
Sugiyamaella castrensis (C. Ramirez & A.E. Gonzalez)H. Urbina & M. Blackw., comb. nov.
MycoBank MB564465Basionym: Candida castrensis C. Ramirez & A.E.
Gonzalez, Mycopathologia 87:178, 1984
Sugiyamaella floridensis (Kurtzman) H. Urbina & M.Blackw., comb. nov.
MycoBank MB564466Basionym: Candida floridensis Kurtzman, FEMS
Yeast Res 7:1054, 2007
Sugiyamaella grinbergsii (Kurtzman) H. Urbina & M.Blackw., comb. nov.
MycoBank MB564467Basionym: Candida grinbergsii Kurtzman, FEMS
Yeast Res 7:1056, 2007
Sugiyamaella lignohabitans (Kurtzman) H. Urbina &M. Blackw., comb. nov.
MycoBank MB564468Basionym: Candida lignohabitans Kurtzman, FEMS
Yeast Res 7:1057, 2007
Sugiyamaella marilandica (Kurtzman) H. Urbina & M.Blackw., comb. nov.
MycoBank MB564469Basionym: Candida marilandica Kurtzman, FEMS
Yeast Res 7:1059, 2007
Sugiyamaella marionensis (Kurtzman) H. Urbina &M. Blackw., comb. nov.
MycoBank MB564470Basionym: Candida marionensis Kurtzman, FEMS
Yeast Res 7:1059, 2007
Sugiyamaella neomexicana (Kurtzman) H. Urbina &M. Blackw., comb. nov.
MycoBank MB564471Basionym: Candida neomexicana Kurtzman, FEMS
Yeast Res 7:1060, 2007
Sugiyamaella novakii (G. Peter, Tornai-Lehoczki & T.Deak) H. Urbina & M. Blackw., comb. nov.
TABLE I. Continued
Species Code
Loci
LSU ITS COXII MtSm
Sc. cryptocercus NRRL Y-48824T; CBS12658;
RG11-03-01-06-P1
JQ714001 JQ713977 JQ714021 JQ713989 JQ714031
NRRL Y-48823;RG11-03-01-07-FM1
JQ713994 JQ713970 JQ714014 JQ713982 JQ714026
NRRL Y-48826;RG11-03-01-05-F7
JQ713998 JQ713974 JQ714018 JQ713986 JQ714029
RG11-03-01-05-P2 JQ713992 JQ713968 JQ714012 JQ713980 JQ714024RG11-03-01-05-P3 JQ713993 JQ713969 JQ714013 JQ713981 JQ714025RG11-03-01-05-A2 JQ713991 JQ713967 JQ714011 JQ713979 JQ714023RG11-03-01-06-FM1 JQ714000 JQ713976 JQ714020 JQ713988 JQ714022RG11-03-01-07-P2 JQ713995 JQ713971 JQ714015 JQ713983 JQ714027RG11-03-01-05-M2 JQ713999 JQ713975 JQ714019 JQ713987 JQ714030RG11-03-01-05-F3 JQ713997 JQ713973 JQ714017 JQ713985 JQ714028RG11-03-01-06-A1 JQ713996 JQ713972 JQ714016 JQ713984 –
Sequences generated in this work are in boldface.T 5 type strain; I 5 isotype strain.
URBINA ET AL.: YEASTS FROM A WOOD ROACH 653
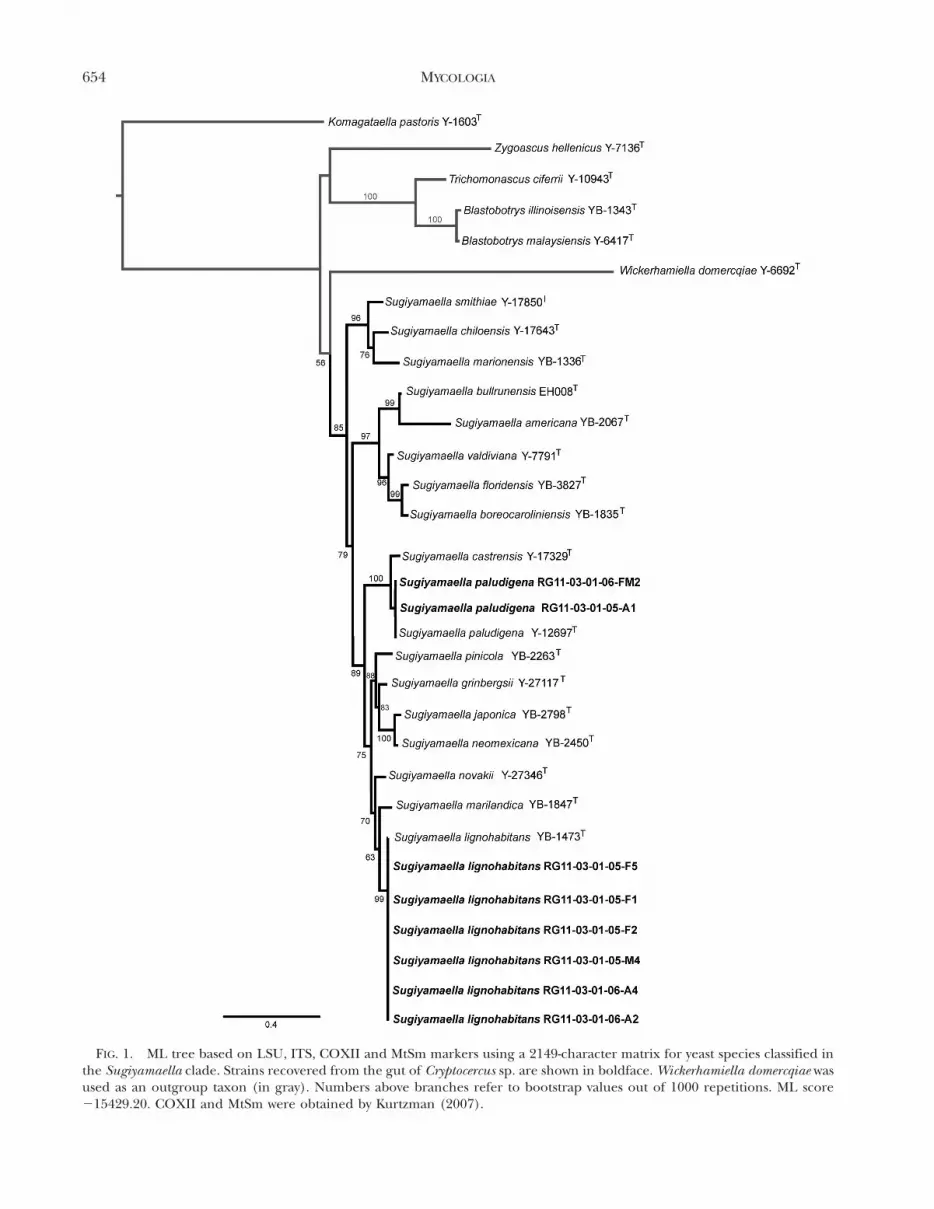
FIG. 1. ML tree based on LSU, ITS, COXII and MtSm markers using a 2149-character matrix for yeast species classified inthe Sugiyamaella clade. Strains recovered from the gut of Cryptocercus sp. are shown in boldface. Wickerhamiella domercqiae wasused as an outgroup taxon (in gray). Numbers above branches refer to bootstrap values out of 1000 repetitions. ML score215429.20. COXII and MtSm were obtained by Kurtzman (2007).
654 MYCOLOGIA
MycoBank MB564472Basionym: Candida novakii G. Peter, Tornai-Lehoczki
& T. Deak, Antonie van Leeuwenhoek 71:376, 1997
Sugiyamaella paludigena (Golubev & Blagod.) H.Urbina & M. Blackw., comb. nov.
MycoBank MB564473Basionym: Candida paludigena Golubev & Blagod.,
Int J Syst Bacteriol 31:92, 1981
Sugiyamaella pinicola (Kurtzman) H. Urbina & M.Blackw., comb. nov.
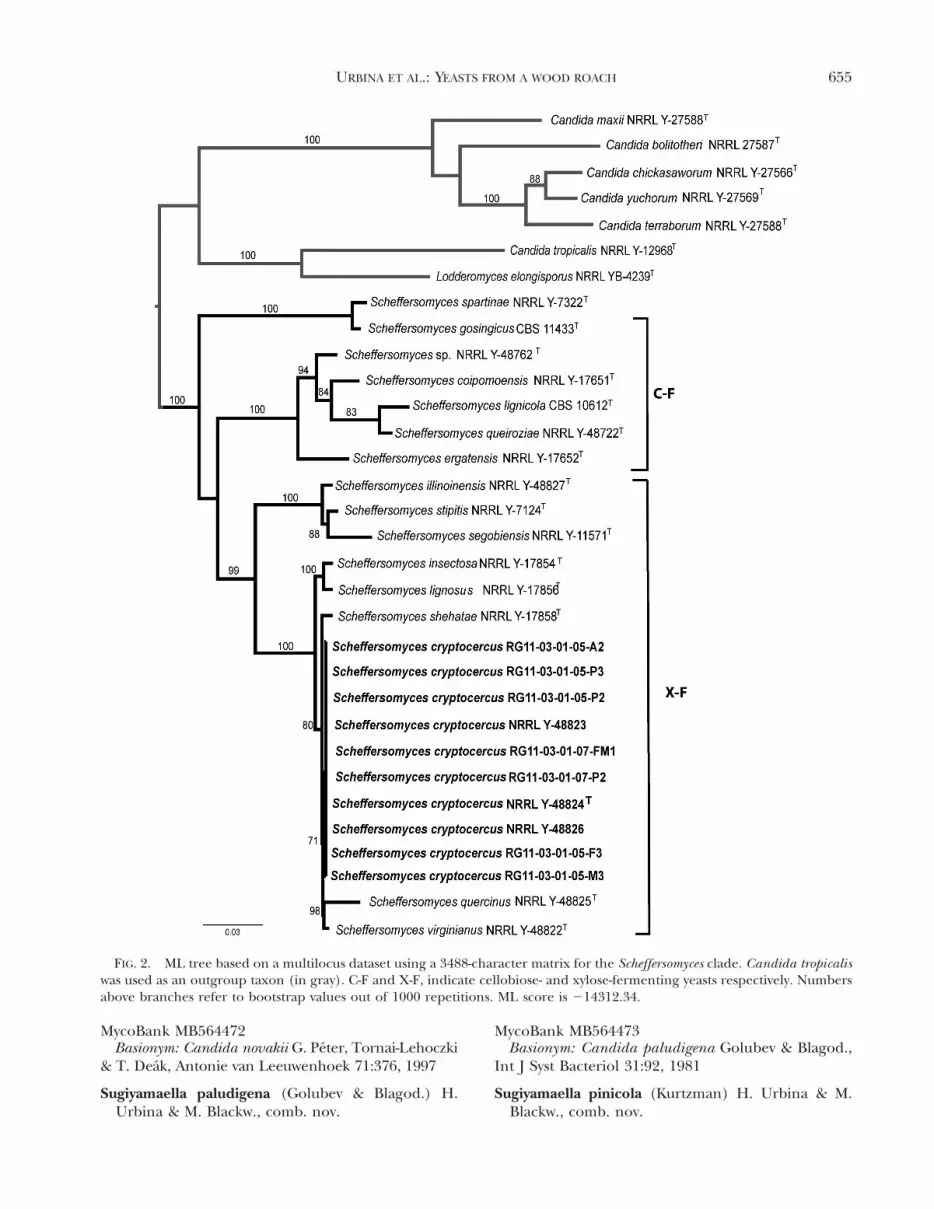
FIG. 2. ML tree based on a multilocus dataset using a 3488-character matrix for the Scheffersomyces clade. Candida tropicaliswas used as an outgroup taxon (in gray). C-F and X-F, indicate cellobiose- and xylose-fermenting yeasts respectively. Numbersabove branches refer to bootstrap values out of 1000 repetitions. ML score is 214312.34.
URBINA ET AL.: YEASTS FROM A WOOD ROACH 655

MycoBank MB564547Basionym: Candida pinicola Kurtzman, FEMS Yeast
Res 7:1061, 2007
Sugiyamaella valdiviana (Grinb. & Yarrow) H. Urbina& M. Blackw., comb. nov.
MycoBank MB564548Basionym: Candida valdiviana Grinb. & Yarrow,
Antonie van Leeuwenhoek 36:145, 1970In addition to the Sugiyamaella species isolated
from wood roaches, we also obtained strains of anundescribed X-F yeast species from each of the sevenroaches dissected. These isolates comprise a mono-phyletic group based on the multilocus phylogeneticanalysis (71% bootstrap support value), morpholog-ical characterizations, and a phylogenetic study usingXYL1 locus (FIGS. 2, 3, 4). The nucleotide differencesbetween the new strains and the close relative, Sc.shehatae, were 0 bp in LSU, 2 bp in SSU, 3 bp in ITS,11 bp in RPB1 and 20 bp in XYL1. In addition nomajor differences were found among strains of Sc.cryptocercus described below with CDU RAPD-PCRfingerprinting primer (FIG. 3).
Scheffersomyces cryptocercus H. Urbina & M.Blackw., sp. nov. FIG. 5
MycoBank MB564549Cells are subglobose to ellipsoidal (6–8.5 3 5–
7.5 mm) in yeast extract (0.5%), glucose (2%) brothafter 3 d at 25 C; asexual reproduction is by budding;cell clusters and pseudohyphae are present. Coloniesare cream and smooth with entire margins on YM
agar after 7 d at 25 C. Well developed true hyphae arepresent after 12 d on cornmeal agar in Dalmau plates.Asci and ascospores not observed. D-glucose, D-galactose,maltose, a-methyl-D-glucoside, a,a-trehalose and D-xylose are fermented; cellobiose, sucrose, melezitose,raffinose, inulin, starch, and lactose are not fermented.D-glucose, D-galactose, L-sorbose, N-acetyl-D-glucosamine,D-ribose, D-xylose, L-arabinose (weak), D-arabinose (var-iable), sucrose, maltose, a,a-trehalose, a-methyl-D-glucoside, cellobiose, salicin (variable), arbutin (variable),melibiose, melezitose, starch, ribitol, D-glucitol, D-manni-tol, D-glucono-1,5-lactone (variable), 2-keto-D-gluconate(fast) are assimilated. No growth occurs on L-rhamnose,lactose, raffinose, inulin, glycerol, erythritol, xylitol, L-arabinitol, galactitiol, myo-inositol, D-gluconate, D-glucuro-nate, DL-lactate, succinate, citrate, methanol, ethanol,propane-1,2-diol, butane-2,3-diol, quinic acid, D-glucarateand D-galactonate as sole carbon sources. Assimilation ofnitrogen compounds: positive for L-lysine, ethylamine-HCl, D-tryptophan, and cadaverine, and negative forimidazole, D-glucosamine, creatinine, creatine, nitrite,and nitrate. Growth is positive for myo-inositol, andnegative for other vitamin tests. Growth in amino acid-free medium is positive. Growth at 37 C is positive. Growthon YM agar with 10% sodium chloride is negative to slow.Growth in 50% glucose/yeast extract (0.5%) is negative.Growth is positive in 100 mg/mL cycloheximide. Diazo-nium blue b reaction is negative.
Type: NRRL Y-48824T (5 CBS 12658; RG11-03-01-06-P1) is preserved as a lyophilized preparation in theAgricultural Research Service Culture Collection(NRRL), Peoria, Illinois, USA. The habitat is the gutof the wood roach Cryptocercus sp. from the Appala-chian Trail at Newfound Gap near the borderbetween Tennessee and North Carolina, USA.
Etymology: The species name cryptocercus (N.L. gen.n.) refers to the host genus of wood roach,Cryptocercus, from which this species was isolated.
DISCUSSION
Several strains of a yeast closely related to Candidasake were found in the gut of the wood roach, C.punctulatus (Prillinger et al. 1996, Prillinger andKonig 2006). Candida sake also has been isolatedfrom food and liquid fermentations as well as soil andwater and clinical isolations. It is variable in fermen-tative ability, including weak or delayed fermentationof cellobiose but does not ferment xylose, althoughthe strains assimilate xylose. Several studies ofunidentified cockroaches collected in Panama andBlatta orientalis from Germany (Henninger andWindisch 1976, Nguyen et al. 2007) report differencesin the gut yeasts that probably reflect differences inbehavior and geographical distribution between
FIG. 3. Molecular comparison of Scheffersomyces crypto-cercus strains and related species using RAPD-PCR CDUfingerprinting primers.
656 MYCOLOGIA

cockroaches and Cryptocercus. Although the commonoccurrence of some gut microbes have been used tosupport a sister group relationship between Cryptocer-cus and termites, too few data support this contention(Legendre et al. 2008, Engel et al. 2009). In additionthe occurrence of the gut yeasts outside the insectsargues that the yeasts may be acquired from the habitat.
The isolation of the new X-F yeast isolated from thegut of wood roaches extends the association of yeastswith the rare biochemical ability to ferment D-xyloseand wood-feeding insects (Zhang et al. 2003, Suh et al.2004, Nguyen et al. 2006, Berkov et al. 2007,Grunwald et al. 2010). This finding is in agreementwith studies in which the X-F yeasts Sc. stipitis and a
FIG. 4. ML consensus tree based on XYL1 using a 489-character matrix of X-F members in the Scheffersomyces stipitissubclade. Scheffersomyces ergatensis was used as an outgroup taxon (in gray). Number above each branch refers to bootstrapvalues out of 1000 repetitions. ML score is 21813.42.
URBINA ET AL.: YEASTS FROM A WOOD ROACH 657

close relative of Sc. shehatae were the predominantascomycete yeasts in the gut of the wood-feedingbeetles, O. disjunctus, and long-horned beetles in thefamily Cerambycidae (Suh et al. 2003, Nardi et al. 2006,Grunwald et al. 2010). In addition to the commonassociation between Scheffersomyces spp. and wood-feeding insects, similar associations also occur betweenthe Sugiyamaella clade and the insects (Houseknecht etal. 2011). In addition the presence of Sc. cryptocercussuggests the possibility of specificity with an understud-ied group of wood-feeding insects. Furthermore, themethodology applied in this study, including a largenumber of isolations from different parts of the gut ofhost individuals, let us discover rare yeasts by overcom-ing the effect of the abundance of other species thatmay overshadow the presence of rare yeasts.
The phylogenetic analysis based on nucleotidesequences of the XYL1 locus alone agrees with theseparation of Sc. cryptocercus as a unique species asdetermined by the multilocus phylogenetic recon-struction (FIG. 4). The use of the easily amplifiedXYL1 has been used to recognize cryptic species inthe Scheffersomyces clade (Urbina and Blackwell 2012).
Despite the large number of XYL1 nucleotidedifferences between Sc. cryptocercus strains and thetype culture of Sc. shehatae, only four amino acidchanges were present at the N-terminal region in theputative XR, but none of these mutations changedthe polarity of the amino acids (data not shown). Themutations in the protein sequences surrounded theconserved domains, and these may have a role in theperformance of the enzyme as has been pointed out(Urbina and Blackwell 2012). The abundance ofsynonymous substitutions is an indication that selec-tion may act on this locus to maintain an invariableprotein sequence.
ACKNOWLEDGMENTS
We thank Christopher Carlton and Michael L. Ferro,Department of Entomology, Louisiana State University, forcollecting insect specimens used in this study. This work wassupported by the National Science Foundation (DEB-0732671), Department of Energy, contract to LawrenceLivermore National Laboratory (DE-AC52-07NA27344) andthe LSU Boyd Professor Research Fund.
LITERATURE CITED
Agbogbo FK. 2007. Production of ethanol from cornstover hemicellulose hydrolyzate using Pichia stipitis.J Ind Microbiol Biotechnol 34:723–727, doi:10.1007/s10295-007-0247-z
———, Coward-Kelly G. 2008. Cellulosic ethanol produc-tion using the naturally occurring xylose-fermentingyeast, Pichia stipitis. Biotechnol Lett 30:1515–1524,doi:10.1007/s10529-008-9728-z
———, Wenger KS. 2006. Effect of pretreatment chemicalson xylose fermentation by Pichia stipitis. BiotechnolLett 28:2065–2069, doi:10.1007/s10529-006-9192-6
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. 1990.Basic local alignment search tool. J Mol Biol 215:403–410, doi:10.1016/S0022-2836(05)80360-2
Barnett JA, Payne RW, Yarrow D. 2000. Yeasts: characteristicsand identification. Cambridge, UK: Cambridge Univ. Press.1139 p.
Berkov A, Feinstein J, Small J, Nkamany M. 2007. Yeastsisolated from Neotropical wood-boring beetles in SEPeru. Biotropica 39:530–538, doi:10.1111/j.1744-7429.2007.00280.x
Berlanga M, Paster BJ, Guerrero R. 2009. The taxophysio-logical paradox: changes in the intestinal microbiota ofthe xylophagous cockroach Cryptocercus punctulatus
FIG. 5. Morphological characterization of Scheffersomycescryptocercus. a. budding cells and (b) pseudohyphae, after7 d at 25 C in YM broth and agar. c, d. Budding cells grownon V8 agar at 17 C; arrow indicates a cell with multiplebudding. Bars 5 10 mm.
658 MYCOLOGIA

depending on the physiological state of the host. IntMicrobiol 12:227–236, doi:10.2436/20.1501.01.102
Boundy-Mills K. 2006. Methods for investigating yeastbiodiversity. In: Peter G, Rosa CA, eds. Biodiversityand ecophysiology of yeast. Berlin: Springer-Verlag.p 67–100.
Calderon O, Berkov A. 2012. Midgut and fat bodybacteriocytes in Neotropical cerambycid beetles (Cole-optera: Cerambycidae). Environ Entomol 41:108–117,doi:10.1603/EN11258
Carpenter KJ, Chow L, Keeling PJ. 2009. Morphology,phylogeny and diversity of Trichonympha (Parabasalia:Hypermastigida) of the wood-feeding cockroach Cryp-tocercus punctulatus. J Eukaryot Microbiol 56:305–313,doi:10.1111/j.1550-7408.2009.00406.x
———, Horak A, Chow L, Keeling PJ. 2011. Symbiosis,morphology and phylogeny of Hoplonymphidae (Para-basalia) of the wood-feeding roach Cryptocercus punc-tulatus. J Eukaryot Microbiol 58:426–436, doi:10.1111/j.1550-7408.2011.00564.x
———, ———, Keeling PJ. 2010. Phylogenetic position andmorphology of Spirotrichosomidae (Parabasalia): newevidence from Leptospironympha of Cryptocercus punc-tulatus. Protist 161:122–132, doi:10.1016/j.protis.2009.06.003
Clark JW, Kambhampati S. 2003. Phylogenetic analysis ofBlattabacterium, endosymbiotic bacteria from the woodroach, Cryptocercus (Blattodea: Cryptocercidae), includ-ing a description of three new species. Mol PhylogenetEvol 26:82–88, doi:10.1016/S1055-7903(02)00330-5
Engel MS, Grimaldi DA, Krishna AK. 2009. Termites(Isoptera): their phylogeny, classification and rise toecological dominance. Am Mus Novit 3650:1–27, http://hdl.handle.net/2246/5969, doi:10.1206/651.1
Fadda ME, Mossa V, Pisano MB, Deplano M, Cosentino S.2004. Occurrence and characterization of yeasts isolat-ed from artisanal Fiore Sardo cheese. Int J FoodMicrobiol 95:51–59, doi:10.1016/j.ijfoodmicro.2004.02.001
———, Viale S, Deplano M, Pisano MB, Cosentino S. 2010.Characterization of yeast population and molecularfingerprinting of Candida zeylanoides isolated fromgoat’s milk collected in Sardinia. Int J Food Microbiol136:376–380, doi:10.1016/j.ijfoodmicro.2009.10.007
Golubev WI, Blagodatskaya M, Suetin SO, Trotsenko R.1981. Pichia inositovora and Candida paludigena, twonew species of yeasts isolated from peat. Int J SystBacteriol 31:91–96, doi:0020-77 13/8 1/01009 1-06$02.00/0
Grunwald S, Pilhofer M, Holl W. 2010. Microbial associationsin gut systems of wood- and bark-inhabiting longhornedbeetles [Coleoptera: Cerambycidae]. Syst Appl Micro-biol 33:25–34, doi:10.1016/j.syapm.2009.10.002
Henninger W, Windisch S. 1976. Kluyveromyces blattae sp. n.,a new multispored yeast for Blatta orientalis (author’stransl). Arch Microbiol 109:153–156, doi:10.1007/BF00425128
Hibbett DS. 1996. Phylogenetic evidence for horizontaltransmission of group I introns in the nuclearribosomal DNA of mushroom-forming fungi. Mol Biol
Evol 13:903–917, doi:10.1093/oxfordjournals.molbev.a025658
Houseknecht JL, Hart EL, Suh SO, Zhou JJ. 2011. Yeasts inthe Sugiyamaella clade associated with wood-ingestingbeetles and the proposal of Candida bullrunensis sp. nov.Int J Syst Evol Microbiol 61:1751–1756, doi:10.1099/ijs.0.026427-0
Hughes GL, Allsopp PG, Webb RI, Yamada R, Iturbe-Ormaetxe I, Brumbley SM, O’Neill SL. 2011. Identifi-cation of yeast associated with the planthopper,Perkinsiella saccharicida: potential applications for Fijileaf gall control. Curr Microbiol 63:392–401, doi:10.1007/s00284-011-9990-5
Kitade O. 2004. Comparison of symbiotic flagellate faunaebetween termites and wood-feeding cockroach of thegenus Cryptocercus. Microbes Environ, 19:215–220.
Klass KD, Nalepa C, Lo N. 2008. Wood-feeding cockroachesas models for termite evolution (Insecta: Dictyoptera):Cryptocercus vs. Parasphaeria boleiriana. Mol PhylogenetEvol 46:809–817, doi:10.1016/J.Ympev.2007.11.028
Knapp S, McNeill J, Turland NJ. 2011. Changes topublication requirements made at the XVIII Interna-tional Botanical Congress in Melbourne: What doese-publication mean for you? Taxon 60:1498–1501.
Kurtzman CP. 2007. Eleven new species of Sugiyamaella andCandida from forest habitats. FEMS Yeast Res 7:1046–1063, doi:10.1111/j.1567-1364.2007.00224.x
———, Fell JW, Boekhout T. 2011. The yeasts, a taxonomicstudy. 5th ed. Amsterdam: Elsevier. p xxii + 1–289, xv +290–1335, xvi + 1336–2080.
———, Robnett CJ. 1998. Three new insect-associatedspecies of the yeast genus Candida. Can J Microbiol44:965–973.
Lamarche J, Stefani FO, Seguin A, Hamelin RC. 2011. Impactof endochitinase-transformed white spruce on soil fungalcommunities under greenhouse conditions. FEMS Mi-crobiol Ecol 76:199–208, doi:10.1111/j.1574-6941.2011.01041.x
Legendre F, Whiting MF, Bordereau C, Cancello EM, Evans TA,Grandcolas P. 2008. The phylogeny of termites (Dictyop-tera: Isoptera) based on mitochondrial and nuclearmarkers: implications for the evolution of the worker andpseudergate castes, and foraging behaviors. Mol Phylo-genet Evol 48:615–627, doi:10.1016/j.ympev.2008.04.017
Liu K, Warnow TJ, Holder MT, Nelesen SM, Yu J, StamatakisAP, Linder CR. 2012. SATe-II: very fast and accuratesimultaneous estimation of multiple sequence align-ments and phylogenetic trees. Syst Biol 61:90–106,doi:10.1093/sysbio/syr095
Maddison W, Maddison D. 2005. Mesquite: a modular systemfor evolutionary analysis. Evolution 62:1103–1118.
Maekawa K, Park YC, Lo N. 2005. Phylogeny of endosym-biont bacteria harbored by the woodroach Cryptocercusspp. (Cryptocercidae: Blattaria): molecular clock evi-dence for a late Cretaceous–early Tertiary split of Asianand American lineages. Mol Phylogenet Evol 36:728–733, doi:10.1016/J.Ympev.2005.03.033
Nardi JB, Bee CM, Miller LA, Nguyen NH, Suh SO,Blackwell M. 2006. Communities of microbes thatinhabit the changing hindgut landscape of a subsocial
URBINA ET AL.: YEASTS FROM A WOOD ROACH 659

beetle. Arthropod Struct Dev 35:57–68, doi:10.1016/j.asd.2005.06.003
Nguyen NH, Suh SO, Blackwell M. 2007. Five novel Candidaspecies in insect-associated yeast clades isolated fromNeuroptera and other insects. Mycologia 99:842–58,doi:10.3852/mycologia.99.6.842
———, ———, Marshall CJ, Blackwell M. 2006. Morpholog-ical and ecological similarities: wood-boring beetlesassociated with novel xylose-fermenting yeasts, Spathas-pora passalidarum gen. sp. nov. and Candida jeffriesii sp.nov. Mycol Res 110:1232–1241, doi:10.1016/j.mycres.2006.07.002
Noda S, Inoue T, Hongoh Y, Vongkaluang C, Kudo T,Ohkuma M. 2006. Identification and characterizationof ectosymbionts of distinct lineages in Bacteroidalesattached to flagellated protists in the gut of termitesand a wood-feeding cockroach. Environ Microbiol 8:11–20, doi:10.1111/j.1462-2920.2005.00860.x
Ohama T, Suzuki T, Mori M, Osawa S, Ueda T, Watanabe K,Nakase T. 1993. Non-universal decoding of the leucinecodon CUG in several Candida species. Nucleic AcidsRes 21:4039–4045, doi:10.1093/nar/21.17.4039
Ohkuma M, Noda S, Hongoh Y, Nalepa CA, Inoue T. 2009.Inheritance and diversification of symbiotic trichonym-phid flagellates from a common ancestor of termitesand the cockroach Cryptocercus. Proc Biol Sci 276:239–245, doi:10.1098/rspb.2008.1094
Prillinger P, Konig H. 2006. The intestinal yeasts. In: KonigH, Varma A, eds. Soil biology. Vol. 6. Intestinalmicroorganisms of soil invertebrates. Berlin, Heidel-berg: Springer-Verlag. p 319–334.
———, Messner R, Konig H, Bauer R, Lopandic K, MolnarO, Danget P, Weigang F, Kirisits T, Nakase T, Sigler L.1996. Yeasts associated with termites: a phenotypic andgenotypic characterization and use of coevolution fordating evolutionary radiations in asco- and basidiomy-cetes. System Appl Microbiol 19:265–283, doi:10.1016/j.bbr.2011.03.031
Sacchi L, Nalepa CA, Bigliardi E, Corona S, Grigolo A,Laudani U, Bandi C. 1998. Ultrastructural studies ofthe fat body and bacterial endosymbionts of Crypto-cercus punctulatus Scudder (Blattaria: Cryptocercidae).Symbiosis 25:251–269.
Schoch CL, Seifert KA, Huhndorf S, Robert V, Spouge JL,Levesque CA, Chen W, the Barcoding Consortium.2012. Nuclear ribosomal internal spacer (ITS) regionas a universal DNA barcode marker for fungi. Proc NatlAcad Sci USA 109: 6241– 6246, doi:10.1073/pnas.1117018109
———, Sung GH, Lopez-Giraldez F, Townsend JP, Miadli-kowska J, Hofstetter V, Robbertse B, Matheny PB, KauffF, Wang Z, Gueidan C, Andrie RM, Trippe K, CiufettiLM, Wynns A, Fraker E, Hodkinson BP, Bonito G,Groenewald JZ, Arzanlou M, de Hoog GS, Crous PW,Hewitt D, Pfister DH, Peterson K, Gryzenhout M,Wingfield MJ, Aptroot A, Suh SO, Blackwell M, HillisDM, Griffith GW, Castlebury LA, Rossman AY,Lumbsch HT, Lucking R, Budel B, Rauhut A, Dieder-ich P, Ertz D, Geiser DM, Hosaka K, Inderbitzin P,Kohlmeyer J, Volkmann-Kohlmeyer B, Mostert L,
O’Donnell K, Sipman H, Rogers JD, Shoemaker RA,Sugiyama J, Summerbell RC, Untereiner W, JohnstonPR, Stenroos S, Zuccaro A, Dyer PS, Crittenden PD,Cole MS, Hansen K, Trappe JM, Yahr R, Lutzoni F,Spatafora JW. 2009. The Ascomycota tree of life: aphylum-wide phylogeny clarifies the origin and evolu-tion of fundamental reproductive and ecological traits.Syst Biol 58:224–239, doi:10.1093/sysbio/syp020
Stamatakis A. 2006. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa andmixed models. Bioinformatics 22:2688–2690, doi:10.1093/bioinformatics/btl446
Suh SO, Blackwell M. 2004. Three new beetle-associatedyeast species in the Pichia guilliermondii clade. FEMSYeast Res 5:87–95, doi:10.1016/j.femsyr.2004.06.001
———, ———, Kurtzman CP, Lachance MA. 2006. Phyloge-netics of Saccharomycetales, the ascomycete yeasts. Myco-logia 98:1006–1017, doi:10.3852/mycologia.98.6.1006
———, Marshall CJ, McHugh JV, Blackwell M. 2003. Woodingestion by passalid beetles in the presence of xylose-fermenting gut yeasts. Mol Ecol 12:3137–3145, doi:10.1046/j.1365-294X.2003.01973.x
———, McHugh JV, Pollock DD, Blackwell M. 2005. Thebeetle gut: a hyperdiverse source of novel yeasts. MycolRes 109:261–265, doi:10.1017/S0953756205002388
———, White MM, Nguyen NH, Blackwell M. 2004. Thestatus and characterization of Enteroramus dimorphus: axylose-fermenting yeast attached to the gut of beetles.Mycologia 96:756–760, doi:10.2307/3762109
Sullivan DJ, Westerneng TJ, Haynes KA, Bennett DE,Coleman DC. 1995. Candida dubliniensis sp. nov.:phenotypic and molecular characterization of a novelspecies associated with oral candidosis in HIV-infectedindividuals. Microbiology 141:1507–1521, doi:10.1099/13500872-141-7-1507
Urbina H, Blackwell M. 2012. Multilocus phylogenetic studyof the Scheffersomyces yeast clade and molecularcharacterization of the N-terminal region of xylosereductase gene. PLoS ONE 7(6):e39128, doi:10.1371/journal.pone.0039128
White TJ, Bruns TD, Lee S, Taylor JW. 1990. Amplificationand direct sequencing of fungal ribosomal RNA genesfor phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ,White TJ, eds. PCR protocols: a guide to methods andapplications. San Diego: Academic Press. p 315–322.
Wohlbach DJ, Kuo A, Sato TK, Potts KM, Salamov AA, LabuttiKM, Sun H, Clum A, Pangilinan JL, Lindquist EA, LucasS, Lapidus A, Jin M, Gunawan C, Balan V, Dale BE,Jeffries TW, Zinkel R, Barry KW, Grigoriev IV, Gasch AP.2011. Comparative genomics of xylose-fermenting fungifor enhanced biofuel production. Proc Natl Acad Sci U SA 108:13212–13217, doi:10.1073/pnas.1103039108
Yarrow D. 1998. Methods for the isolation, maintenance andidentification of yeasts. In: Kurtzman CP, Fell JW, eds. Theyeasts, a taxonomic study. Amsterdam: Elsevier. p 77–100.
Zhang N, Suh SO, Blackwell M. 2003. Microorganisms inthe gut of beetles: evidence from molecular cloning. JInvertebr Pathol 84:226–33, doi:10.1016/j.jip.2003.10.002
660 MYCOLOGIA





























