Embed Size (px)
Citation preview

www.elsevier.com/locate/plantsci
Plant Science 171 (2006) 194–205
Screening for inhibitors of 2-oxoglutarate-dependent dioxygenases:
Flavanone 3b-hydroxylase and flavonol synthase§
Heidrun Halbwirth a, Thilo C. Fischer b, Karin Schlangen a, Wilhelm Rademacher c,Klaus-Jurgen Schleifer d, Gert Forkmann b, Karl Stich a,*
a Institut fur Verfahrenstechnik, Umwelttechnik und Technische Biowissenschaften, Technische Universitat Wien, Getreidemarkt 9/1665, A-1060 Wien, Austriab Lehrstuhl fur Zierpflanzenbau, Technische Universitat Munchen Weihenstephan, Am Hochanger 4, D-85350 Freising, Germany
c BASF Aktiengesellschaft, Agrarzentrum, D-67114 Limburgerhof, Germanyd BASF Aktiengesellschaft, Forschung Wirk und Effektstoffe, D-67056 Ludwigshafen, Germany
Received 17 November 2005; received in revised form 12 March 2006; accepted 22 March 2006
Available online 18 April 2006
Abstract
2-Oxoglutarate-dependent dioxygenases (2-ODDs) catalyze numerous steps in biosynthetic pathways of plants. Prohexadione-Ca is a known
inhibitor of such reactions, due to its structural similarity to 2-oxoglutarate. In apple (Malus domestica) and pear (Pyrus communis) leaves, the
transient inhibition of 2-ODDs flavanone 3b-hydroxylase (FHT) and flavonol synthase (FLS) by prohexadione-Ca results in distinct changes in the
flavonoid spectrum, which are responsible for an enhanced resistance against two major pome fruit diseases, fire blight (caused by Erwinia
amylovora) and apple scab (caused by Venturia inaequalis). We used recombinant apple and pear FHT and apple FLS for screening 23 structural
analogues of 2-oxoglutarate, mostly cyclohexanediones, pyridine dicarboxylic acids and N-heterocycles with carbonyl functions for other
dioxygenase inhibitors. Activations, which were also observed for some compounds, are interpreted as in vitro effects due to Fe2+-chelating ability.
Apart from structural similarity to 2-oxoglutarate, close structural similarity of cyclohexanediones and some pyridine dicarboxylic acids to
flavonoid substrates was identified. Beyond the competitive inhibition for the co-substrate 2-oxoglutarate, flavonoid converting 2-ODDs may also
be inhibited at the substrate binding site by these inhibitors. All compounds found to be active as inhibitors may prove useful for studying the
reaction mechanisms and substrate specificities of various 2-ODDs.
# 2006 Elsevier Ireland Ltd. All rights reserved.
Keywords: Pome fruits (apple, pear); Dioxygenase; Flavanone 3b-hydroxylase (FHT = F3H, EC 1.14.11.9); Flavonol synthase (FLS, EC 1.14.11.23);
Prohexadione-Ca; 2-Oxoglutarate analogous enzyme inhibitors
1. Introduction
Three enzymes of the flavonoid pathway in apple and pear
leaves are 2-oxoglutarate-dependent dioxygenases (2-ODDs):
flavanone 3b-hydroxylase (FHT, also F3H, EC 1.14.11.9),
flavonol synthase (FLS, EC 1.14.11.23), and anthocyanidin
Abbreviations: ANS, anthocyanidin synthase; DHK, dihydrokaempferol;
DHQ, dihydroquercetin; EGME, ethyleneglycol monomethylether; ERI, erio-
dictyol; FHT, flavanone 3-hydroxylase; FLS, flavonol synthase; NAR, narin-
genin; 2-ODD, 2-oxoglutarate-dependent dioxygenase§ The nucleotide sequences reported in this paper have been submitted to
GenBank under the accession numbers AY965339 (FHT Malus domestica cv.
Weirouge), AY965340 (FHT M. domestica cv. M9), AY965341 (FHT Pyrus
communis cv. Pyrodwarf), AY965342 (FHT P. communis cv. Conference) and
AY965343 (FLS M. domestica cv. M9).
* Corresponding author. Tel.: +43 1 58801 17320; fax: +43 1 58801 17399.
E-mail address: [email protected] (K. Stich).
0168-9452/$ – see front matter # 2006 Elsevier Ireland Ltd. All rights reserved.
doi:10.1016/j.plantsci.2006.03.014
synthase (ANS, EC 1.14.11.19) (Fig. 1). The acylcyclohex-
anedione prohexadione-Ca (CH1 in Fig. 2) – originally
developed as a plant growth retardant – is a structural analogue
of 2-oxoglutarate which competitively inhibits biosynthetic
steps catalyzed by 2-ODDs [1]. Several dioxygenases from
different pathways are affected. The inhibition of late steps of
gibberellin biosynthesis, leading to plants with a more compact
shoot, is of practical relevance in crop production [1]. Since
prohexadione-Ca is relatively short-lived and degrades into
natural compounds, it has favourable toxicological and
ecotoxicological features [1]. The transient inhibition of 2-
ODDs of the flavonoid pathway by prohexadione-Ca is of
interest, since it leads to significant changes in the flavonoid
spectrum of apple (Malus domestica) and pear (Pyrus
communis) leaves (Fig. 3). In particular, FHT inhibition leads
to the accumulation of its substrate eriodictyol, which is
subsequently converted to the 3-deoxyleucoanthocyanidin
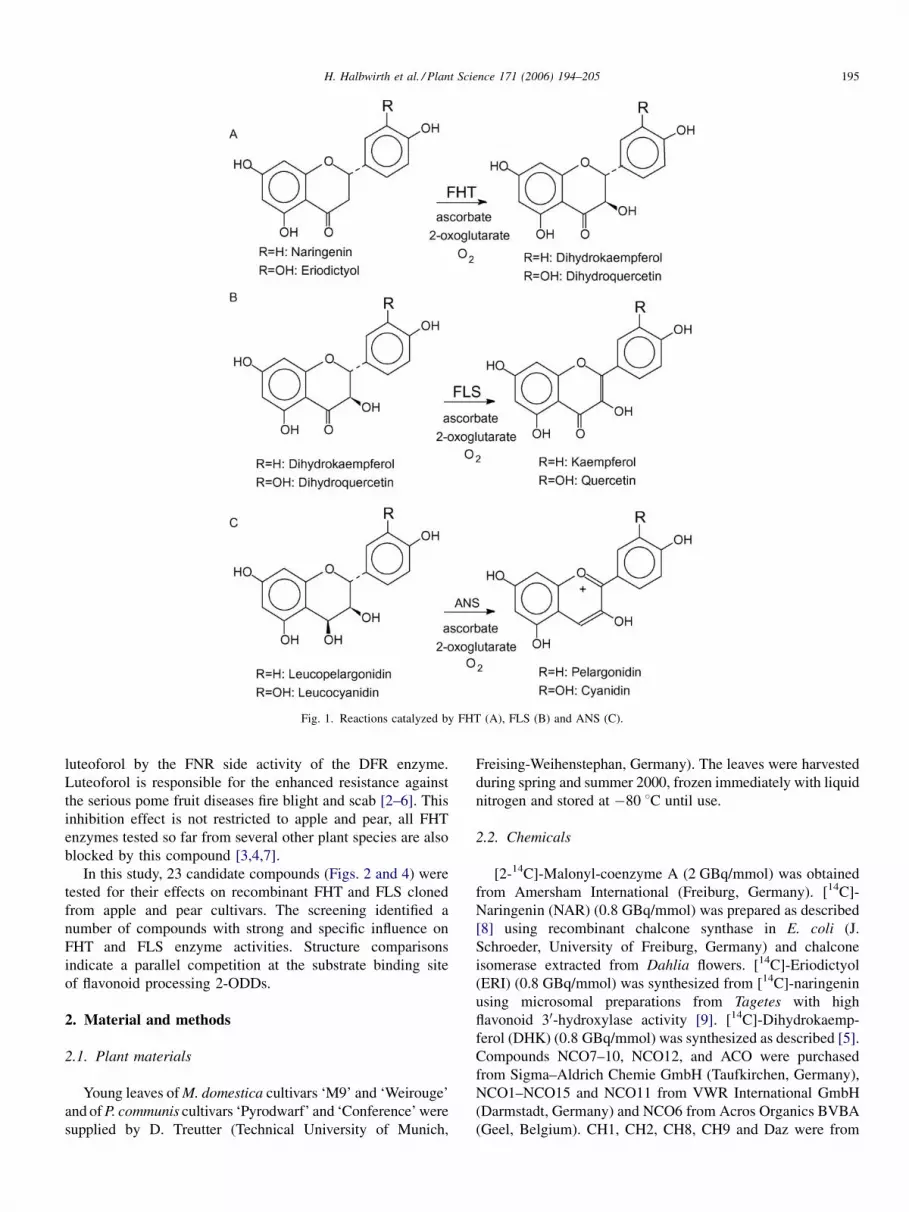
H. Halbwirth et al. / Plant Science 171 (2006) 194–205 195
Fig. 1. Reactions catalyzed by FHT (A), FLS (B) and ANS (C).
luteoforol by the FNR side activity of the DFR enzyme.
Luteoforol is responsible for the enhanced resistance against
the serious pome fruit diseases fire blight and scab [2–6]. This
inhibition effect is not restricted to apple and pear, all FHT
enzymes tested so far from several other plant species are also
blocked by this compound [3,4,7].
In this study, 23 candidate compounds (Figs. 2 and 4) were
tested for their effects on recombinant FHT and FLS cloned
from apple and pear cultivars. The screening identified a
number of compounds with strong and specific influence on
FHT and FLS enzyme activities. Structure comparisons
indicate a parallel competition at the substrate binding site
of flavonoid processing 2-ODDs.
2. Material and methods
2.1. Plant materials
Young leaves of M. domestica cultivars ‘M9’ and ‘Weirouge’
and of P. communis cultivars ‘Pyrodwarf’ and ‘Conference’ were
supplied by D. Treutter (Technical University of Munich,
Freising-Weihenstephan, Germany). The leaves were harvested
during spring and summer 2000, frozen immediately with liquid
nitrogen and stored at �80 8C until use.
2.2. Chemicals
[2-14C]-Malonyl-coenzyme A (2 GBq/mmol) was obtained
from Amersham International (Freiburg, Germany). [14C]-
Naringenin (NAR) (0.8 GBq/mmol) was prepared as described
[8] using recombinant chalcone synthase in E. coli (J.
Schroeder, University of Freiburg, Germany) and chalcone
isomerase extracted from Dahlia flowers. [14C]-Eriodictyol
(ERI) (0.8 GBq/mmol) was synthesized from [14C]-naringenin
using microsomal preparations from Tagetes with high
flavonoid 30-hydroxylase activity [9]. [14C]-Dihydrokaemp-
ferol (DHK) (0.8 GBq/mmol) was synthesized as described [5].
Compounds NCO7–10, NCO12, and ACO were purchased
from Sigma–Aldrich Chemie GmbH (Taufkirchen, Germany),
NCO1–NCO15 and NCO11 from VWR International GmbH
(Darmstadt, Germany) and NCO6 from Acros Organics BVBA
(Geel, Belgium). CH1, CH2, CH8, CH9 and Daz were from

H. Halbwirth et al. / Plant Science 171 (2006) 194–205196
Fig. 2. Chemical structures of cyclohexanediones (in order to improve clarity, the nomenclature used for CH1–CH9 has been made uniform with regard to numbering
of the carbon atoms and, therefore, does not always comply with IUPAC or CAS rules) and some related compounds tested as potential dioxygenase inhibitors. The
structures of the dioxygenase cofactors 2-oxoglutarate and ascorbic acid are shown for comparison. CH1, calcium 3-hydroxy-5-oxo-4-propionyl-cyclohex-3-ene-1-
carboxylate (prohexadione-Ca); CH2, 3-hydroxy-5-oxo-4-cyclopropanecarbonyl-cyclohex-3-ene-1-carboxylic acid ethyl ester (trinexapac-ethyl); CH3: 3-hydroxy-
5-oxo-4-propionyl-cyclohex-3-ene-1-carbothioic acid S-ethyl ester; CH4, 3-hydroxy-5-oxo-4-propionyl-cylohex-3-ene-1-carbaldehyde; CH5, 3-hydroxy-5-oxo-4-
butyryl-cyclohex-3-ene-1-carboxylic acid ethyl ester; CH6, 3-hydroxy-5-oxo-4-propionyl-clohex-3-ene-1-(2-dimethylamino)-thiazole; CH7, 3-hydroxy-5-oxo-4-
propionyl-cyclohex-3-ene-1-pentanoic acid; CH8, sodium 4,6-dioxo-2,2-dimethyl-5-(1-alloxyamino-butylidene)-cyclohexane-1-carboxylic acid methyl ester
(alloxydim-Na); CH9, 3-hydroxy-5-oxo-4-(1-ethoxyimino-butyl)-cyclohex-3-ene-1-(tetrahydro-thiopyran-3-yl) (cycloxydim); ACO, benzene-1,2,4,5-tetracar-
boxylic acid; Daz: succinic acid 2,2-dimethyl hydrazide (daminozide).

H. Halbwirth et al. / Plant Science 171 (2006) 194–205 197
Fig. 3. The flavonoid biosynthesis in apple and pear and the influence of prohexadione-Ca.
LGC Promochem GmbH (Wesel, Germany). The acylcyclo-
hexanediones CH3–CH7 were synthesized at BASF Main
Laboratory (Ludwigshafen, Germany), following standard
procedures. Technical details are given in [10–12].
2.3. PCR amplification of FHT and FLS cDNAs
mRNA was isolated from leaf material of M. domestica
and P. communis cultivars using the mMACS mRNA Isolation
Kit1 (Miltenyi Biotec, 752-01). Reverse transcription was
performed with the SuperScript II1 Reverse Transcriptase
(Gibco BRL, 18064) and an anchored oligo-dT-primer. For
RT-PCR primers were constructed for the respective
untranslated 50- and 30-regions. For Malus FHT sequences
were available [13,14] and the primers GAA CCA ACA AAT
TCG ACA CC and GGA AGG AAA AGG TAC AAG TAG C
were used for the amplification of FHT from M. domestica cv.
Weirouge and cv. M9. For the Pyrus FHT several combina-
tions of primers constructed from Malus FHT sequences were
tested and selected for the amplification of fragments of the
expected size range. Amplification was possible with the
primers GAA CCA ACA AAT TCG ACA CC (both Pyrus
cultivars, untranslated 50-regions) and CCA CAA AGA GCT
TTC AAG TGA CTG G (P. communis cv. ‘Pyrodwarf’,
untranslated 30-region) or GGA AGG AAA AGG TAC AAG
TAG C (P. communis cv. ‘Conference’, untranslated 30-region). For the Malus FLS RT-PCR the primers CGA ACG
TCC AAA GCC CTT C and CCT GGA ACA CAA ATT CAA
ACA C were used relying on an available sequence [13]. All
amplifications were done with the Taq/Pwo-Polymerase
system ‘Expand High Fidelity PCR System1 (Roche,
1732641).
2.4. Cloning and heterologous expression in yeast
Cloning and heterologous expression of M. domestica cv.
M9 FHT and FLS and P. communis cv. Pyrodwarf FHT was
carried out using the commercially available yeast expression
system pYES2.1 (TOPO TA Cloning1 Kit, Invitrogen K4150-
01). All constructs were transformed into the yeast strain
InvSc1 (Invitrogen V825-20) using the S.c. Easy CompTM
Transformation Kit (Invitrogen K5050-01). The enzymes were
prepared from galactose-induced yeast cultures as described by
Urban et al. [15]. Transformation of yeast and preparation of
yeast-derived enzymes was also done with empty pYES2-
vector to provide a negative control for enzyme activities.
Protein content was determined by a modified Lowry procedure
[16] using BSA as a standard.
2.5. Standard enzyme assay
In a final volume of 100 ml the reaction mixture contained
10 ml enzyme preparation (17 mg total protein, recombinant
FHT preparation; 69 mg total protein, recombinant FLS
preparation), 80 ml 0.1 M K2HPO4/KH2PO4 (0.6 M sorbitol,
0.4% Na-ascorbate, pH 7.25), 108 Bq [14C]-labeled substrate
(0.148 nmol NAR for FHT, 0.095 nmol DHK for FLS),
50 nmol 2-oxoglutarate (in 5 ml distilled water) and was
started by the addition of 10 nmol FeSO4�7H2O (in 5 ml
distilled water). After 15 min (FHT) or 30 min (FLS) at 30 8C

H. Halbwirth et al. / Plant Science 171 (2006) 194–205198
Fig. 4. N-Heterocycles tested as potential dioxygenase inhibitors. NCO1, pyridine-2,4-dicarboxylic acid; NCO2, pyridine-2,5-dicarboxylic acid; NCO3, pyridine-
2,6-dicarboxylic acid; NCO4, pyridine-3,4-dicarboxylic acid; NCO5, pyridine-3,5-dicarboxylic acid; NCO6, pyridine-2,6-dicarboxylic acid chloride; NCO7,
pyridine-2,3-dicarboxylic acid (quinolinic acid); NCO8, pyrrole-3,4-dicarboxylic acid diethyl ester; NCO9, pyrazole-3,5-dicarboxylic acid (hydrate); NCO10,
piperidine-2-carboxylic acid; NCO11, piperazine-1,4-bis-(2-ethane sulphonic acid); NCO12, piperidine-4-carboxylic acid.
the reaction was stopped with 10 ml glacial acetic acid. The
phenolic compounds were extracted with 70 ml EtOAc, the
organic phases were applied to a precoated cellulose plate
(1.0571.001, Merck, Darmstadt, Germany) and chromato-
graphed with chloroform/acetic acid/water (10:9:1) as a solvent
system. Heterologous expression of Malus DFR in yeast and
enzymatic reactions were performed as previously described
[5], but using the same buffer as required for the FHT and FLS
assays. Radioactivity was detected and quantified using a
Typhoon 8600 Imager (Amersham). Recovery of [14C]-labeled
substrates was 95%. HPLC analysis was performed as
described [17] using a Perkin-Elmer Series 200 system
equipped with a UV detector coupled with a 500TR Flow
Scintillation Analyzer for the detection of radiolabeled
substances.
2.6. Enzyme characterization
Determination of the pH-optimum was carried out as
described for the standard assay, but using 0.2 M KH2PO4–
K2HPO4 buffer (containing 0.4% sodium-ascorbate) with pH
values between 6.0 and 8.5. Dependence of the enzymatic
turnover on time was determined during a period of 60 min at
intervals of 5 min. Kinetic data were calculated from Line-
weaver–Burk plots. All data represent an average of at least
three independent experiments.
2.7. Inhibitor studies
FHTassays were performed with NAR, FLS and DFR assays
with DHK as substrates. The screening was performed with two
final inhibitor concentrations of 1 and 0.1 mM. All inhibitor
solutions were freshly prepared. NCO1, NCO8–10, ACO, CH2,
CH3, CH5–CH9 and Daz were dissolved in methanol, NCO2,
NCO4 and NCO7 in DMSO, and NCO3, NCO6 and CH4 in
EGME. NCO12 was dissolved in water and CH1 in 0.05 M
McIlvaine buffer (pH 4). NCO5 and NCO11 were dissolved in
0.05 M Tris/HCl (pH 8.5). For methanol soluble inhibitors,
30 ml (for FLS and FHT assays) or 15 ml (for DFR assays) of
the 3.33 mM solutions were transferred into 1.5 ml reaction

H. Halbwirth et al. / Plant Science 171 (2006) 194–205 199
tubes together with the substrates and the solvent was removed
by vacuum. For the other inhibitors, 5 ml (for FLS and FHT
assays) or 2.5 ml (for DFR assays) of 20 mM stock solutions
were transferred directly into the reaction tubes before starting
the assay. In these cases, the control also contained the
respective solvent. Determination of IC50 values was carried
out according to [18]. All data represent an average of at least
three independent experiments.
2.8. Structural comparisons
Three-dimensional structure superimpositions were carried
out using the software package SYBYL1 7.0, Tripos Inc., St.
Louis, MO, USA.
3. Results and discussion
FHT and FLS were chosen as two major targets of
prohexadione-Ca. We isolated the cDNAs of FHT from apple
and pear leaves as well as FLS from apple leaves, and used the
recombinant enzymes to discover other inhibitors of dioxy-
genases or of specific flavonoid enzymes.
Fig. 5. Alignment of FHT genes of Malus domestica (FHT-M) and Pyrus communis
(Lee et al., GenBank); M. sp., Malus species (Davies [14]); Wr, Weirouge; M9, cv.
homologous. Some exchanges seem to be specific for the genera.
3.1. Cloning and heterologous expression
The FHT-cDNA clones of the Malus cultivars ‘M9’ and
‘Weirouge’ were obtained via RT-PCR, based on the
sequence information present in the databases [13,14]. Both
cDNAs showed few exchanges on the amino acid level
(Fig. 5) and consequently only M9 FHT was used for
heterologous expression and inhibitor screening. No sequence
information was available for Pyrus, therefore, the isolation
of the respective cDNA clones was performed by using
different primer combinations derived from the untranslated
50- and 30- sequences of Malus FHT for RT-PCR. By this
approach the FHT cDNAs of the Pyrus cultivars ‘Pyrodwarf’
and ‘Conference’ were obtained and found to be identical at
amino acid sequence (Fig. 5). The Pyrus cDNAs also turned
out to be closely homologous to the Malus cDNAs, some
exchanges are obviously genera specific. The FLS cDNA of
the M. domestica cultivar ’M9’ was also cloned by RT-PCR
using the available sequence information from Fuji apple
[13]. There was one (A108G) amino acid exchange between
the cDNA of the M. domestica cultivar ‘Fuji’ and the M.
domestica rootstock ‘M9’.
(FHT-P) cultivars. Variable positions are shown in bold print. Cultivars: Fj, Fuji
M9; Pd, Pyrodwarf; Co, Conference. The amino acid sequences are all closely

H. Halbwirth et al. / Plant Science 171 (2006) 194–205200
3.2. Verification of the recombinant enzymes and
optimization of the enzyme assays
Final proofs that the isolated cDNA clones from Malus cv.
‘M9’ and Pyrus cv. ‘Conference’ encode FHT or FLS were
achieved by assays with the enzymes functionally expressed in
yeast. [14C]-Products were identified by HPLC co-chromato-
graphy with authentic reference substances (Rt DHK:
21.40 min; Rt DHQ: 19.94 min; Rt kaempferol: 26.93 min; Rt
quercetin: 25.23 min).
3.2.1. Flavanone 3b-hydroxylase (FHT)
Incubation of the flavanones [14C]-NAR or [14C]-ERI with
enzyme preparations of recombinant Malus and Pyrus FHT in
the presence of 2-oxoglutarate and Fe2+ led to the formation of
the dihydroflavonols [14C]-DHK and [14C]-DHQ, respectively
(Fig. 1). No activity was observed when control enzyme
preparations from yeast containing the empty expression vector
Table 1
Effects of potential inhibitorsa of 2-ODDs on FHT and FLS activities
Code Compound
CH1 Calcium 3-hydroxy-5-oxo-4-propionyl-cyclohex-
3-ene-1-carboxylate (prohexadione-Ca)
CH2 3-Hydroxy-5-oxo-4-cyclopropanecarbonyl-cyclohex-
3-ene-1-carboxylic acid ethyl ester (trinexapac-ethyl)
CH3 3-Hydroxy-5-oxo-4-propionyl-cyclohex-
3-ene-1-carbothioic acid S-ethyl ester
CH4 3-Hydroxy-5-oxo-4-propionyl-cylohex-
3-ene-1-carbaldehyde
CH5 3-Hydroxy-5-oxo-4-butyryl-cyclohex-
3-ene-1-carboxylic acid ethyl ester
CH6 3-Hydroxy-5-oxo-4-propionyl-clohex-
3-ene-1-(2-dimethylamino)-thiazole
CH7 3-Hydroxy-5-oxo-4-propionyl-cyclohex-
3-ene-1-pentanoic acid
CH8 Sodium 4,6-Dioxo-2,2-dimethyl-5-
(1-alloxyamino-butylidene)-cyclohexane-1-
carboxylic acid methyl ester (alloxydim-Na)
CH9 3-Hydroxy-5-oxo-4-(1-ethoxyimino-butyl)-cyclohex-
3-ene-1-(tetrahydro-thiopyran-3-yl) (cycloxydim)
ACO Benzene-1,2,4,5-tetracarboxylic acid
Daz Succinic acid 2,2-dimethyl hydrazide (daminozide)
NCO1 Pyridine-2,4-dicarboxylic acid
NCO2 Pyridine-2,5-dicarboxylic acid
NCO3 Pyridine-2,6-dicarboxylic acid
NCO4 Pyridine-3,4-dicarboxylic acid
NCO5 Pyridine-3,5-dicarboxylic acid
NCO6 Pyridine-2,6-dicarboxylic acid chloride
NCO7 Pyridine-2,3-dicarboxylic acid (quinolinic acid)
NCO8 Pyrrole-3,4-dicarboxylic acid diethyl ester
NCO9 Pyrazole-3,5-dicarboxylic acid (hydrate)
NCO10 Piperidine-2-carboxylic acid
NCO11 Piperazine-1,4-bis-(2-ethane sulphonic acid)
NCO12 Piperidine-4-carboxylic acid
Enzyme activities were measured in the presence of potential inhibitors using recom
Enzyme activities are expressed as % rel. compared to a control (without inhibitor).
protein for FLS and 2.6 mkat kg�1 for DFR. The latter served as a control for unspeci
(heteroanalogous) carboxyl functions; ACO, aromate with carbonyl functions; Daza In order to improve clarity, the nomenclature used for CH1–CH9 has been made
always comply with IUPAC or CAS rules.
pYES were used. The Malus FHT reaction was optimized for
the screening assay. An optimum at pH 7.25 was observed for
naringenin and eriodictyol as substrates. Highest reaction rates
were measured at 30 8C, where the formation of DHK was
linear with time up to 15 min and with protein concentration up
to 17 mg protein in the assay. The values for apparent Km and
Vmax were 10 mM and 117.5 mkat kg�1 protein for naringenin
(Vmax/Km = 11.8 mkat kg�1 mM�1) and 12 mM and 102.9 mkat
kg�1 protein for eriodictyol (Vmax/Km = 8.6 mkat kg�1 mM�1).
Hence, a slightly higher preference was observed for naringenin
compared to eriodictyol.
3.2.2. Flavonol synthase (FLS)
The identity of the putative FLS-cDNA clone isolated from
Malus cv. ‘M9’ was also confirmed by assays with the
recombinant enzyme. Incubation of the enzyme preparations
with [14C]-DHK or [14C]-DHQ in the presence of 2-
oxoglutarate and Fe2+ led to the expected formation of the
Concentration
(mM)
FHT
(% rel.)
FLS
(% rel.)
DFR
(% rel.)
1/0.1 10/33 11/29 104/105
1/0.1 9/38 50/77 105/99
1/0.1 15/40 33/90 103/105
1/0.1 33/75 56/82 86/101
1/0.1 36/68 48/88 99/103
1/0.1 25/56 44/73 81/103
1/0.1 13/75 23/64 97/108
1/0.1 89/115 78/84 105/121
1/0.1 100/100 104/100 118/87
1/0.1 100/97 95/98 110/113
1/0.1 108/108 97/104 98/110
1/0.1 7/21 3/13 114/108
1/0.1 320/132 3/11 127/116
1/0.1 220/186 150/144 115/100
1/0.1 98/101 86/85 127/116
1/0.1 99/99 128/102 100/101
1/0.1 196/194 120/105 104/102
1/0.1 205/120 171/115 115/101
1/0.1 113/100 113/103 95/100
1/0.1 97/100 22/79 103/97
1/0.1 75/84 91/94 112/113
1/0.1 96/101 115/102 95/99
1/0.1 106/107 103/102 97/99
binant enzymes. Inhibitor concentrations are final concentrations in the assay.
Hundred percent correspond to 3.3 mkat kg�1 protein for FHT, 0.13 mkat kg�1 -
fic effects. CH1–CH9, cyclohexanediones; NCO1–NCO12, N-heterocycles with
, daminozide.
uniform with regard to numbering of the carbon atoms and, therefore, does not

H. Halbwirth et al. / Plant Science 171 (2006) 194–205 201
Table 2
Influence of the inhibitor/2-oxoglutarate ratio on the inhibitory effect of
selected compounds
Ratio inhibitor/
2-oxoglutarate
Relative FHT
activity (%)
Relative FLS
activity (%)
Controls – 100 100
NCO1 5:1 11 7
1:1 12 8
1:2 25 18
1:5 76 68
CH3 5:1 16 38
1:1 35 62
1:2 72 86
1:5 98 100
CH7 5:1 14 26
1:1 38 40
1:2 72 75
1:5 95 95
Enzyme activities were measured at a fixed concentration of 0.5 mM inhibitor
and 2-oxoglutarate concentrations from 0.1 to 2.5 mM. Relative activities were
calculated against controls without inhibitor.
flavonols kaempferol and quercetin, respectively (Fig. 1).
Formation of these products was not observed with enzyme
preparations from control yeast carrying the empty expression
vector. The enzyme also required 0.6 M sorbitol in the assay
buffer for measurable enzyme activity. An optimum at pH 7.25
was observed for DHK and DHQ as substrates. Highest reaction
rates were achieved at 30 8C, where the reaction was linear with
time for up to 30 min and 210 mg protein in the assay. The values
for apparent Km and Vmax were 1 mM and 18.5 mkat kg�1 protein
for DHK (Vmax/Km = 18.5 mkat kg�1 mM�1) and 1.2 mM
and 21.5 mkat kg�1 protein for DHQ (Vmax/Km = 17.9 mkat
kg�1 mM�1).
3.3. Screening for dioxygenase inhibitors
Selection of candidate compounds was based on structural
similarity to 2-oxoglutarate and also included some published
dioxygenase inhibitors (NCO1, NCO2, NCO3) [20,21]. Two
series of substances were screened: (i) cyclohexanediones such
as prohexadione-Ca (Fig. 2), mostly acyl-substituted and (ii)
pyridine dicarboxylic acids and other N-heterocycles with
carboxyl functions (Fig. 4). The common feature of most
selected compounds is the presence of two or three carbonyl
groups or heteroanalogous carbonyl functions. Some of the
compounds used for screening are components of commer-
cially available plant growth regulators (prohexadione-Ca,
CH1 and trinexapac-ethyl, CH2) [1], which served as positive
controls for inhibition, whereas CH8 and CH9 are gramini-
cides inhibiting acetyl-CoA-carboxylase in fatty acid bio-
synthesis [21].
Results obtained with the heterologously expressed FHT
from Pyrus turned out to be identical to those of heterologously
expressed FHT from Malus and, therefore, are not shown
separately. Possible influences of pH and buffer components
could be excluded since FHT and FLS assays were performed
with the same buffer system. Dihydroflavonol 4-reductase
(DFR, EC number 1.1.1.219) assays performed under the same
conditions were also included (Table 1). No significant
influence on this NADPH-dependent oxidoreductase, which
represents a different enzyme class, was observed, so that the
possibility of unspecific effects on enzyme activities such as
enzyme denaturation can be ruled out. As a result of this
screening, several compounds with distinct effects on the
dioxygenases FHT and FLS were identified (Table 1).
Unexpectedly, some compounds had a stimulating effect on
enzymatic activity in the in vitro assay.
3.4. Cyclohexanediones and related structures
Of the cyclohexanediones tested, CH1–CH7 inhibited FHT
and FLS activities at a concentration as low as 0.1 mM.
However, the extent of inhibition, was variable and depended
on compound structures. For CH2, CH3 and CH5, it can be
expected that the effects are different in vitro and in planta due
to the action of plant esterases, which can hydrolyze the ester
function [1]. The inhibition effect was overcome by increasing
the 2-oxoglutarate concentrations, which demonstrated the
competitive nature of the FHT and FLS inhibition (Table 2). For
prohexadione-Ca the IC50 was 23 mM for FHT and 50 mM for
FLS (Fig. 6).
For anthocyanidin synthase (ANS), which is closely related
to FLS and FHT [22], the binding mode of the co-substrate
2-oxoglutarate was revealed by a crystallographic study of an
enzyme/Fe2+/2-oxoglutarate/substrate analog complex [23]. It
could be shown that Fe2+ in the active centre is complexed by
the C1-carboxylate and the oxo group of 2-oxoglutarate, an
aspartyl, two histidinyl residues and a H2O ligand. In addition,
the negative charge of the C5-carboxylate of 2-oxoglutarate
interacts with the positive charge of an arginine residue. In the
case of the acylcyclohexanedione inhibitors (CH1–CH7), one
of the respective oxo functions of the ring corresponds to the
oxo group of 2-oxoglutarate. Similarly, the carbonyl oxygen of
the acyl residues in positions 1 and 4 substitute for the
carboxylate groups in positions 1 and 5 of 2-oxoglutarate
(Fig. 7A).
The general structural requirements for effective inhibition
are demonstrated in Fig. 7A. All compounds possessing a
3-hydroxy-5-oxo-cyclohex-3-ene (or the tautomeric 3,5-dioxo-
cyclohexane) structure with a carbonyl substituent in position 1
(R side: e.g. carboxylic or carbothioic acid or ester, CH1, CH2,
CH3, CH5; carbaldehyde, CH4; or electronegative thiazole,
CH6) and a second substituent in position 4 (R0 side: carbonyl
function provided by propionyl or related substituents) are
strong inhibitors. These findings correspond with reports on
inhibitors of 2-ODDs involved in gibberellin biosynthesis
[1,19,20]. Carbonyls of the acylcyclohexanediones may
interact via their negative partial charges or the oxo functions
may form an enolate as a superior ligand. Obviously, the
heteroatoms of some of the active compounds can take on the
oxygen functions.
Apart from possessing a carbonyl (or a heteroanalogous
carbonyl) moiety, substitution at position 1 of the cyclohex-

H. Halbwirth et al. / Plant Science 171 (2006) 194–205202
Fig. 6. Concentration dependence of FHT and FLS inhibition by selected inhibitors of 2-ODDs. Recombinant FHT and FLS were incubated with increasing
concentrations of prohexadione-Ca, NCO1, NCO2 and NCO5, respectively. FHT (triangles, dashed lines) and FLS (squares, full lines) activities are expressed as % of
control (no added inhibitor). Hundred percent correspond to 3.1 mkat kg�1 protein for FHT and 0.15 mkat kg�1 protein for FLS.
anedione ring appears to be relatively flexible for inhibiting
FHT and FLS in vitro. Compounds with substituents clearly
different from CH1 and CH2, e.g. CH3, CH4, CH5, CH6, CH7
have still considerable activity. One may assume that generally
these molecules are stabilized by keto-enol tautomerism (three
oxygen functions at the R0 side) and would, therefore, give a
particularly good similarity to 2-oxoglutarate. In case of CH7
with the carbonyl at the end of the n-pentanoic acid substituent,
a ring conformation is possible that brings the carbonyl close to
the cyclohexanedione ring.
Fig. 7. Structural features of acylcyclohexanedione inhibitors. (A) Left: cyclohexane
For substituents R and R0 see text. Centre: their structure (blue) corresponds to the s
flavonoid structure (DHK). Center: structural overlapping of a typical acylcyclohexan
no 3-hydroxy function (cf. Fig. 1)]. Right: structure of prohexadione.
In contrast, the graminicidal compounds CH8 and CH9
generally showed a low or no inhibitory effect. This is in line
with their low structural similarity to 2-oxoglutarate and
prohexadione (Fig. 2). In particular, the structures lack a second
carbonyl or carbonyl-like substituent next to the 4-position of
the cyclohexanedione structure (equivalent to the 4-propionyl
moiety of prohexadione-Ca), which could mimic the C5-
carboxylate of 2-oxoglutarate. Another important structural
difference to 2-oxoglutarate is the presence of an N-oxime
function in the herbicidal molecules. Furthermore, neither the
diones with in vitro activity on FHT and FLS show common structural features.
tructure of 2-oxoglutarate (green). Right: structure of 2-oxoglutarate. (B) Left:
edione (prohexadione) and the flavonoid DHK [NAR is identical to DHK but has

H. Halbwirth et al. / Plant Science 171 (2006) 194–205 203
Fig. 8. Structural similarities of selected pyridine dicarboxylic acids to 2-oxoglutarate. The structure of 2-oxoglutarate is shown in green. Structures of NCO1 (left),
NCO2 (center), NCO 3 (right) are shown in light blue. The position of nitrogen is marked in dark blue.
Table 3
Effect of activating compounds on FHT and FLS activity in the absence or
presence of the cofactor 2-oxoglutarate
Addition of 0.5 mM
2-oxoglutarate
Addition
of 1 mM
Relative FHT
activity (%)
Relative FLS
activity (%)
No – 10 7
Yes – 100 100
No NCO2 25 n.d.
Yes NCO2 320 n.d.
No NCO3 18 10
Yes NCO3 220 145
No NCO6 17 9
Yes NCO6 198 128
No NCO7 21 12
Yes NCO7 209 171
2-Oxoglutarate and other low molecular weight compounds were removed by
gel chromatography from the enzyme preparation and enzyme activities were
determined in the absence or presence of 0.5 mM 2-oxoglutarate. All assays
contained 0.1 mM Fe2+ and 0.4% potassium-ascorbate in the buffer. Relative
activities were determined against a control without inhibitor. Hundred percent
correspond to 3.3 mkat kg�1 protein for FHT and 0.13 mkat kg�1 protein for
FLS. n.d., not determined.
benzene derivative ACO nor daminozide (Daz) affected FHT
or FLS activity (Table 1), although Daz inhibits other 2-
oxoglutarate-dependent dioxygenases involved in gibberellin
biosynthesis [1].
Structural analyses demonstrated that particularly the 1-
acyl-substituted cyclohexanediones show pronounced struc-
tural similarity not only to the co-substrate, 2-oxoglutarate, but
also to the flavonoid substrates DHK and NAR (Fig. 7B). These
compounds can obviously interact with flavonoid dioxygenases
in two different ways. They can affect both the substrate and the
co-substrate sites and this may explain their relatively high
degree of inhibition. Absence (NAR) or presence (DHK) of the
3-hydroxyl group in the flavonoid substrate could account for
differing effects of some inhibitors on both enzymes (CH2,
CH3, and CH6). This 3-hydroxyl group overlaps with part of
the acyl residue of the acylcylohexadiones in structural
comparison.
3.5. Pyridine dicarboxylic acids and other N-heterocycles
with carbonyl functions
In previous investigations, certain pyridine dicarboxylic
acids (NCO1, NCO2) have been reported to inhibit other 2-
ODDs, e.g. two gibberellin 2b-hydroxylases [19] and prolyl 4-
hydroxylase involved in collagen biosynthesis [20]. In our
study, NCO1 strongly inhibited FHT and FLS (Table 1, Fig. 6;
IC50 25 mM for FHT, 2.3 mM for FLS). In contrast, strong
activation was observed unexpectedly for NCO2, NCO3,
NCO6 and NCO7, which increased FHT activity up to three
times (Table 1). Moreover, significant differences were
observed in some cases for FHT and FLS. In particular
NCO2 and NCO9 acted as strong inhibitors of FLS (NCO2:
IC50 80 mM, NCO9: IC50 70 mM), but either enhanced (NCO2)
or did not affect (NCO9) FHT activity (Fig. 6, Table 1). The
observed activating effects are in obvious contrast to a
competitive inhibition that was expected because of the
structural similarities of NCO1, NCO2 and NCO3 to 2-
oxoglutarate. The nitrogen atom of the respective pyridine
overlaps with different oxygen functions of 2-oxoglutarate
(Fig. 8). In contrast, no oxygen function of 2-oxoglutarate
corresponds to the nitrogen atom of NCO3 (Fig. 8).
Compounds that enhanced FHT and FLS activities were
also tested in case they could act as additional co-substrates.
Enzyme assays, in which 2-oxoglutarate was removed from the
protein extract by gel chromatography, showed strongly
reduced FHT or FLS activity (Table 3). However, addition
of the respective activating compounds did not result in
restored or even enhanced enzyme activity as found in the
presence of 2-oxoglutarate. It is remarkable, that assays
performed without addition of 2-oxoglutarate still showed a
remaining activity of approximately 10% (Table 3). We
assume that this is due to enzyme bound 2-oxoglutarate, which
co-purified with the 2-oxoglutarate-dependent enzymes. Such
an effect had been observed for FHT before (Forkmann,
unpublished).
On the other hand it could be shown for NCO3 that lower
Fe2+ concentrations led to higher activation (Table 4). Several
interactions are possible between enzyme, enzyme-bound
Fe2+, Fe2+ in solution, flavonoid substrate, 2-oxoglutarate and
the carboxylated N-heterocycles. Some dicarboxypyridines
showed structural similarities to the flavonoid substrates

H. Halbwirth et al. / Plant Science 171 (2006) 194–205204
Fig. 9. Structural similarities of selected flavonoids to various pyridine dicarboxylic acids. Structures of NAR (above) and DHK (below) (both shown in grey) are
compared with structures of NCO1 (left), NCO2 (center), NCO3 (right) (shown in light blue). The positions of nitrogen and oxygen in the rings are marked in dark
blue and in red, respectively.
DHK and NAR (Fig. 9). However, it can be assumed that
competition with the flavonoid substrate would lead to
inhibition rather than activation.
It is likely that the carboxylated N-heterocycles chelate Fe2+
in analogue to 8-hydroxyquinoline, which forms stable
complexes with many transition metal ions [24]. This could
explain the inhibitory action of these compounds. In the
crystallographic study of ANS, a decrease of the electron
density of Fe2+ was observed with citrate as a chelator in the
crystallization solution [22,23]. However, chelation would
lower the available concentration of Fe2+ as enzyme cofactor
and hence would be expected to reduce enzyme activity.
Flavonoids can also chelate Fe2+ [25–27] and excessive Fe2+
might decrease the availability of flavonoid substrates.
Addition of the carboxylated N-heterocycles as chelators
would liberate the weaker flavonoid chelators from the
flavonoid/Fe2+ complex and augment the availability of
flavonoids as enzyme substrates. As expected, the activating
effect is much stronger for low Fe2+ concentrations (Table 4).
Moreover, NAR as the weaker chelator (no vicinal O functions)
in comparison to DHK (vicinal 3-hydroxy/4-oxo functions) is
obviously released more easily by the competing chelators and
hence FHT activity is increased more strongly than FLS activity
(Table 4). In addition, FHT had a much higher activity in this
Table 4
Influence of the Fe2+ concentration on the activating effect of NCO3
Fe2+concentrations
(mM)
Relative FHT
activity (%)
Relative FLS
activity (%)
1000 186 144
100 220 169
10 250 187
1 300 202
Relative enzyme activities were measured at a fixed concentration of 0.1 mM
NCO3 and Fe2+ concentrations from 1000 to 1 mM. Relative activities refer to
control assays in the absence of inhibitor.
assay than FLS. Hence, better substrate availability led to a
stronger effect in case of the FHT assays. It can be concluded
that competitive Fe2+ chelation by flavonoid substrates and
activating compounds provides a plausible explanation for the
observed in vitro activation effects.
4. Conclusions
The screening revealed several potent dioxygenase
inhibitors, which were predominantly acylcyclohexane-
diones, and demonstrated their effects on the flavonoid
enzymes FHT and FLS. The primary structural features of
these compounds necessary for competitive interaction have
been identified in vitro. Structural comparisons of acylcy-
clohexadiones with flavonoid substrates showed that for these
dioxygenases competitive inhibition can also be expected at
the substrate binding site, beyond the general effect on the
2-oxoglutarate co-substrate binding site common to all such
dioxygenases. The inhibiting compounds identified may now
be tested for their in vivo effects to explore their potential in
agronomic applications.
N-Heterocyclic compounds were tested with respect to
effects on FHT and FLS as a second group of structural
analogues of 2-oxoglutarate. Several of these compounds are
also potent inhibitors, but in some cases the inhibitory effect
seems to be counteracted by their action as Fe2+ chelators,
which resulted in higher free flavonoid substrate concentrations
in vitro and, hence, to a higher substrate turn-over.
In contrast to prohexadione-Ca, which shows comparable
inhibitory effects on FHT and FLS, some of the compounds
tested show a higher inhibition of FHT than FLS, e.g. CH2,
CH3. These inhibitors could be of practical interest, because
the beneficial changes in the flavonoid spectrum would be
achieved with lower unintended effects on dioxygenases
other than FHT. Other FHT enzymes (e.g. from leaves of
grape vine, strawberry, rose) show a similar inhibition by

H. Halbwirth et al. / Plant Science 171 (2006) 194–205 205
prohexadione-Ca [3,4,7], and almost identical results were
obtained in this study with recombinant FHT from apple and
pear, which show some variation in amino acid sequences.
Given the known effects of prohexadione-Ca on 2-ODDs of the
gibberellin pathway, it appears likely that the competitive and,
hence, inhibitory effects of the new compounds on FHT and
FLS activity are transferable to other 2-oxoglutarate-depen-
dent plant dioxygenases, irrespective of a putative additional
competition with the flavonoid substrates in case of flavonoid
enzymes. Identification of a 2-ODD gene function in
benzoxazinoid biosynthesis in wheat and maize is a recent
example of application of an inhibitor to other 2-ODDs [28].
Therefore, the new inhibitors can also serve as further tools
to study enzyme activities of the wide-spread family of
2-ODDs.
Acknowledgements
Part of this work was supported by the European Commission
(QLK5-CT-1999-01583). K. Stich and H. Halbwirth gratefully
acknowledge the support by Vienna University of Technology
(GZ 9006.10/006/2005). C. Statnik and R. Paltram are
acknowledged for excellent technical assistance and B. Ros
for performing work on molecular cloning. Special thanks go to
Jurgen Greiner for his extensive assistance during the inhibitor
studies. Finally, we would like to thank U. Hesse for support in
structure comparisons, W. Heller for helpful discussion and E.
Meggeneder for critically reading the manuscript.
References
[1] W. Rademacher, Growth retardants: effects on gibberellin biosynthesis
and other metabolic pathways, Annu. Rev. Plant Physiol. Plant Mol. Biol.
51 (2000) 501–531.
[2] S. Rommelt, N. Zimmermann, W. Rademacher, D. Treutter, Unusual
flavonoid routes induced by the dioxygenase inhibitor prohexadione-Ca in
apple (Malus domestica), Phytochemistry 64 (2003) 709–716.
[3] S. Rommelt, T.C. Fischer, H. Halbwirth, S. Peterek, K. Schlangen, J.B.
Speakman, D. Treutter, G. Forkmann, K. Stich, Effect of dioxygenase
inhibitors on the resistance-related flavonoid metabolism of apple and
pears: chemical, biochemical and molecular biological aspects, Eur. J.
Hortic. Sci. 68 (2003) 129–136.
[4] H. Halbwirth, T.C. Fischer, S. Rommelt, F. Spinelli, K. Schlangen, S.
Peterek, E. Sabatini, C. Messina, J.B. Speakman, C. Andreotti, W.
Rademacher, C. Bazzi, G. Costa, D. Treutter, G. Forkmann, K. Stich,
Induction of antimicrobial 3-deoxyflavonoids in pome fruit trees controls
fire blight, Z. Naturforsch. 58c (2003) 765–770.
[5] T.C. Fischer, H. Halbwirth, B. Meisel, K. Stich, G. Forkmann, Molecular
cloning, substrate specificity of the functionally expressed dihydroflavo-
nol 4-reductases from Malus domestica and Pyrus communis cultivars and
the consequences for flavonoid metabolism, Arch. Biochem. Biophys. 412
(2003) 223–230.
[6] F. Spinelli, J.B. Speakman, W. Rademacher, H. Halbwirth, K. Stich, G.
Costa, Luteoforol, a flavan 4-ol, is induced in pome fruits by prohex-
adione-calcium and shows phytoalexin-like properties against Erwinia
amylovora and other plant pathogens, Eur. J. Plant Pathol. 112 (2005)
133–142.
[7] C. Gosch, I. Puhl, H. Halbwirth, K. Schlangen, S. Rommelt, C. Andreotti,
G. Costa, T.C. Fischer, D. Treutter, K. Stich, G. Forkmann, Effect of
prohexadione-Ca on various fruit crops: flavonoid composition and sub-
strate specificity of their dihydroflavonol 4-reductases, Eur. J. Hortic. Sci.
68 (2003) 144–151.
[8] L. Britsch, W. Heller, H. Grisebach, Conversion of flavanone to flavone,
dihydroflavonol and flavonol with an enzyme system from cell cultures of
parsley, Z. Naturforsch. c 36 (1981) 741–750.
[9] H. Halbwirth, G. Forkmann, K. Stich, The A-ring specific hydroxylation
of flavonols in position 6 in Tagetes sp. is catalyzed by a cytochrome P450-
dependent monooxygenase, Plant Sci. 167 (2004) 129–135.
[10] H.G. Brunner, Cyclohexandion-carbonsaurederivate mit herbizider und
das Pflanzenwachstum regulierender Wirkung, European Patent 0,126,713
(1989).
[11] D. Jahn, M. Keil, D. Kolassa, U. Schirmer, R. Becker, J. Jung, W.
Rademacher, Mittel zur Regulierung des Pflanzenwachstums, European
Patent 0,210,445 (1996).
[12] M. Keil, U. Schirmer, J. Kast, D. Kolassa, B. Wurzer, N. Meyer, W.
Rademacher, J. Jung, D. Carlson, Cyclohexenonverbindungen, ein Ver-
fahren zu ihrer Herstellung sowie ihre Verwendung als Herbizide oder als
Pflanzenwachstum regulierende Mittel, European Patent 0,293,817
(1990).
[13] J.R. Lee, S.-T. Hong, Y.G. Yoo, S.-R. Kim, Unpublished, direct sequence
submission 1998 to GenBank: AF117270 (FHT), AF119095 (FLS).
[14] K.M. Davies, A cDNA clone for flavanone 3-hydroxylase from Malus,
Plant Physiol. 103 (1993) 291.
[15] P. Urban, C. Mignotte, M. Kazmaier, F. Delorme, D. Pompon, Cloning,
yeast expression, and characterization of the coupling of two distantly
related Arabidopsis thaliana NADPH-cytochrome P450 reductases with
P450 CYP73A5, J. Biol. Chem. 272 (1997) 19176–19186.
[16] H. Sandermann, J.L. Strominger, Purification and properties of C55-
isoprenoid alcohol phosphokinase from Staphylococcus aureus, J. Biol.
Chem. 247 (1972) 5123–5131.
[17] K. Vande Casteele, H. Geiger, C.F. Van Sumere, Separation of flavonoids
by reversed-phase high-performance liquid chromatography, J. Chroma-
togr. 240 (1982) 81–94.
[18] B.H. Wang, G.M. Polya, Selective inhibition of cyclic AMP-dependent
protein kinase by amphiphilic triterpenoids and related compounds,
Phytochemistry 41 (1996) 55–63.
[19] D.L. Griggs, P. Hedden, K.E. Temple-Smith, W. Rademacher, Inhibition
of gibberellin 2b-hydroxylases by acylcyclohexanedione derivatives,
Phytochemistry 30 (1991) 2513–2517.
[20] K. Majamaa, V. Gunzler, H.M. Hanauske-Abel, R. Myllyla, K.I. Kivir-
ikko, Partial identity of the 2-oxo-glutarate and ascorbate binding sites of
prolyl 4-hydroxylase, J. Biol. Chem. 261 (1986) 7819–7823.
[21] D. Herbert, K.A. Walker, L.J. Price, K.E. Pallett, S.M. Ridley, J.L.
Harwood, Acetyl-CoA carboxylase—a graminicide target site, Pestic.
Sci. 50 (1997) 67–71.
[22] J.J. Turnbull, J.I. Nakajima, R.W.D. Welford, M. Yamazaki, K. Saito, C.J.
Schofield, Mechanistic studies on three 2-oxoglutarate-dependent oxyge-
nases of flavonoid biosynthesis, J. Biol. Chem. 279 (2004) 1206–1216.
[23] R.C. Wilmouth, J.J. Turnbull, R.W.D. Welford, I.J. Clifton, A.G. Prescott,
C.J. Schofield, Structure and mechanism of anthocyanidin synthase from
Arabidopsis thaliana, Structure 10 (2002) 93–103.
[24] J. Falbe, M. Regitz (Eds.), Rompp-Lexikon Chemie, Band 1, Georg
Thieme Verlag, Stuttgart, New York, 1996, p. 780.
[25] I. Morel, G. Lescoat, P. Cillard, J. Cillard, Role of flavonoids and iron
chelation in antioxidant action, Meth. Enzymol. 234 (1994) 437–443.
[26] S.M. Kuo, P.S. Leavitt, C.P. Lin, Dietary flavonoids interact with trace
metals and affect metallothionein level in human intestinal cells, Biol.
Trace Element Res. 62 (1998) 135–153.
[27] L. Mira, M.T. Fernandez, M. Santos, R. Rocha, M.H. Florencio, K.
Jennings, Interactions of flavonoids with iron and copper ions: a
mechanism for their antioxidant activity, Free Radic. Res. 36 (2002)
1199–1208.
[28] M. Frey, K. Huber, W.J. Park, D. Sicker, P. Lindberg, R.B. Meeley, C.R.
Simmons, N. Yalpani, A. Gierl, A 2-oxoglutarate-dependent dioxygen-
ase is integrated in DIMBOA-biosynthesis, Phytochemistry 62 (2003)
371–376.











![Synthesis and monoamine transporter affinity of new 2β-carbomethoxy-3β-[aryl or heteroaryl]phenyltropanes](https://img.pdfslide.net/doc/110x75/6340412a3f6f6a2463099ada/synthesis-and-monoamine-transporter-affinity-of-new-2v-carbomethoxy-3v-aryl-or.jpg)

















