Embed Size (px)
Citation preview

Eur. J. Biochem. 157,275-289 (1986) 0 FEBS 1986
Sequence-specific 'H-NMR assignments in rabbit-liver metallothionein-2 Gerhard WAGNER ', David NEUHAUS', Erich WORGOTTERI, Milan VASAK', Jeremias H. R. KAGI' and Kurt WUTHRICH' ' Institut fur Molekularbiologie und Biophysik, Eidgenossische Technische Hochschule-Honggerberg, Zurich
Biochemisches Institut der Universitat Zurich
(Received December 23, 1985) - EJB 85 1400
The complete sequence-specific assignment of the 'H nuclear magnetic resonance spectrum of a major subform of rabbit liver metallothionein-2 is presented. The sequential assignment procedures revealed a number of differences with regard to results obtained by earlier partial chemical sequencing of a preparation now known to be microheterogeneous. In particular, the present data indicate a polypeptide chain length of 62 amino acid residues as compared to the occurence of 61 amino acids in all other known mammalian metallothioneins. In the new sequence, which was also fully confirmed by chemical means, the additional amino acid residue was identified as Alas' inserted between Alas and Ala9 of the standard amino acid numeration. In addition to the predominant protein species all preparations contained a minor component, for which the two-dimensional 'H-nuclear magnetic resonance features are compatible with a chemically different, homologous metallothionein.
This is the third paper in a series of three publications describing the assignment of the 'H NMR spectrum of rabbit liver metallothionein-2. In the first paper we identified the cysteine spin systems in double-quantum-filtered two-dimen- sional correlated spectra (DQF-COSY) on the basis of the splitting of the P-proton resonances due to scalar coupling with the 'I3Cd nuclei [l]. The second paper reported on the identification of the 'H spin systems for all amino acid re- sidues according to residue type, using experimental tech- niques such as DQF-COSY, 2D double-quantum spectros- copy, 2D-relayed coherence transfer spectroscopy and 2D nuclear Overhauser enhancement spectroscopy (NOESY) [2]. The present paper describes the sequence-specific 'H reso- nance assignments for MT-2.
Sequence-specific resonance assignments provide the basis for determination of protein solution conformations [3]. Since in MT-2 all '13Cd2+ -Cys coordinative bonds were previously identified [4], it is now possible to locate the Cd2+-thiolate clusters relative to the polypeptide backbone and, in combina- tion with the intra-chain 'H-'H NOES obtained from NOESY, to determine the spatial arrangement of the polypeptide chain around the metal clusters of MT-2 [ 5 ] . Inde- pendent of this distance-geometry determination of the global conformation, the presently described sequence-specific reso- nance assignments enabled an empirical pattern recognition approach [6] to identify two 310 helices and a number of turn- like secondary structure elements [7].
For MT-2 the sequence-specific resonance assignments are of special interest also because a recently solved 0.23-nm crystal structure of a homologous rat metallothionein-2 has a largely different metal coordination topology, with only 9 of the total of 28 Cd-Cys bonds coinciding with those in rabbit
Correspondence to G. Wagner, Institut fur Molekularbiologie und Biophysik, Eidgenossische Technische Hochschule, CH-8093 Zurich- Honggerberg, Switzerland
Abbreviations. 1 D, one-dimensional ; 2D, two-dimensional; ppm, parts per million; COSY, 2D-correlated spectroscopy; DQF-COSY, double-quantum-filtered COSY; NOE, nuclear Overhauser effect; NOESY, 2D NOE spectroscopy; MT-2, metallothionein isoprotein 2 from rabbit liver.
MT-2 [8,9]. These coordinative bonds represent unambiguous evidence by which to compare the conformations of different metallothioneins. Technically speaking, the identification of the Cd-Cys bonds in the solution conformation depends on the results of heteronuclear 'H-lI3Cd COSY experiments [4] and on the resonance assignments documented in this paper, which clearly emphasizes the importance of these experiments.
Even though MT-2 is a small protein with a molecular mass of 6900 Da, the assignment of the 'H-NMR spectrum was, overall, very difficult. One reason for this is that h4T-2 has an unusual amino acid composition, containing only 12 of the 20 common amino acids, i.e. Val, Leu, Glu, Arg and the four aromatics are absent. Thus, except for one isoleucine, there are no residues with bulky side chains. Four residue types are abundant, i.e. there are nine alanines, eight serines, seven lysines and twenty cysteines.
In spite of this inherently difficult situation, the spin systems of all residues could be identified [2]. The spin system identification also revealed that the amino acid composition was at variance with the published primary structure [lo]. Compared to results in [lo], one Lys and one Ser were missing, and there were three additional spin systems of Ala, Asn and Pro. With regard to the sequential assignment techniques used in this paper [ l l , 121, this implicated the additional difficulty that there had to be several errors in the amino acid sequence.
A further complication arose because the 'H-NMR spectra of all the MT-2 preparations examined so far contained a number of extra signals belonging to a minor population of other protein species. Initially, it was imperative to distinguish these extra peaks properly from those of the major species and to clarify whether these other species repre- sented chemically different molecules or different con- formations of MT-2. A distinction from the major species was achieved by comparison of the spectra obtained from different protein preparations, which contained variable admixtures of the minor components under the same conditions of pH, ionic strength and temperature [2]. These observations further indicated that the extra peaks originated from a chemically different species, rather than from different conformations of MT-2, unless the latter were kinetically too stable for

276
thermodynamic equilibrium to be reached withn days. A direct identification as a chemically different protein was then obtained by sequential resonance assignments of a part of this minor species, which showed a primary structure homologous to, but not identical with MT-2.
Finally, a last difficulty was that the WX-2 preparations used so far had a limited lifetime. After l13Cd-enriched MT-2 was kept in solution for several days, new signals could be observed in the 1D '13Cd spectrum and in 2D 'H spectra. The origin of this instability is not yet fully understood. Overall, we thus faced the situation that MT-2 was a relatively short-lived protein with only partially known amino acid sequence, which had a poorly resolved 'H-NMR spectrum because of the uneven amino acid composition, and which had to be studied in mixed solutions containing a second, different protein with closely similar properties.
In Results and Discussion, each assignment step is indi- vidually described and documented. The main reasons which prompted us to choose this lengthy presentation are firstly the forementioned inherent difficulties in this project. Further- more, since crystallographic studies of a homologous protein [8, 91 revealed a largely different polypeptide-metal coordina- tion from that obtained on the basis of the present resonance assignments [4, 51, a complete documentation seemed advisible with regard to the possibility that there might be some controversy ahead.
MATERIALS AND METHODS
Metallothionein-2 (MT-2) was isolated from rabbit livers as described previously [lo, 131. Each preparation was characterized by amino acid analysis (Durrum 500) and by metal analysis using atomic absorption spectroscopy (In- strumentation Laboratory, model IL 157). To avoid a splitting of the Hfl resonances of cysteines by the J-coupling to the spin-1/2 nuclei '13Cd and '"Cd [l], the mixture of Zn, "'Cd, 12Cd and l1 3Cd metal ions found in MT-2 after the isolation
procedure was replaced with the NMR-inactive isotope "'Cd by reconstitution followed by gel filtration (Sephadex G75) [14]. For the NMR measurements, 8 - 12 mM solutions of the protein in H 2 0 or 2H20, with 20 mM [2H11]Tri~/HC1 (Merck and Sharp) and 20 mM KC1 were used. The dissolved samples were kept under argon atmosphere. The protein concentration was determined spectrophotometrically by measuring the absorbance of the apoprotein at 220nm in 0.01M HCI (cZZO = 47300M-' cm-l) [15].
All 2D-NMR spectra were recorded on a Bruker WM 500 spectrometer. DQF-COSY [16- 181 and NOESY [19,20] spectra were recorded as described previously. For all experi- ments quadrature detection was used in both dimensions, with time-proportional phase-incrementation of the first pulse [21], the carrier being placed in the center of the spectrum. For the spectra recorded in HzO, the spectral width was 5000 Hz in both dimensions. Experiments with 512 tl values were recorded; 2048 and 1600 data points along t2 were used for the COSY and NOESY spectra, respectively. Prior to Fourier transformation, the time domain data were multiplied with phase-shifted sine bell window functions [22], using phase shifts of 4 3 2 and n/64 in the t l and t 2 dimensions, respectively. With zero-filling the time domain data were then extended to 1024 points in t1 and 4096 points in t2 . In order to allow accurate comparison of cross-peak positions in the COSY and NOESY spectra, the two experiments were acquired in- terleaved, as described previously [2]. For the spectra recorded in 'H20, a spectral width of 2400 Hz was used in both
Fig. 1. Polypeptide backbone segment with identification of the sequential NOE connectiviiies daN, dNN and dBN. The dashed lines indicate the scalar coupling connectivities between protons within the same residue
dimensions. Otherwise the data handling was the same as for the spectra in H 2 0 .
RESULTS AND DISCUSSION
Assignment strategy
Following established procedures [3, 11, 121, sequential resonance assignments in MT-2 were obtained from NOEs between the amide proton of residue i + l and the a, amide and p protons of the preceding residue i (Fig. 1). The results obtained are listed in Table 1.
Since the amino acid sequence was not completely deter- mined for MT-2, it was an essential factor for successful sequential assignments that a complete identification of the amino acid spin systems to residue types was previously performed. Furthermore, since sequential connectivities rely almost exclusively on NOEs to backbone amide protons (Fig. l), it was essential that the spin system identifications included the connectivities of these resonances with the spin systems of the non-labile protons of the individual residues, and that the large majority of these connectivities relied on scalar couplings (Fig. 1). COSY provided this connectivity for 19 residues which have unique Ha chemical shifts. RELAYED-COSY allowed identification for an additional 30 residues (furthermore, 9 of the COSY assignments were confirmed). At this stage, after 49 NH signals had thus been connected to side-chain signals with COSY or RELAYED-COSY, unique COSY connectivities could be obtained for another 5 amide protons. Overall the amide protons of 54 residues were thus attached by scalar coupling connectivities. The remaining five NH signals were attached to their side-chain spin systems with NOESY experiments as described by Neuhaus et al. [2].
The assignment strategy used for MI'-2 was based on a somewhat different philosophy from that applied in the procedures used previously for other proteins [ll]. For the earlier assignments we recorded NOESY spectra with short mixing times of 80- 100 ms. This allowed only observation of NOE cross peaks corresponding to very short distances, i.e. 60.30 nm, while cross peaks corresponding to slightly longer distances had not reached their maximum intensity and were hidden in thermal noise, and in ' t l noise' arising from the strong residual diagonal peaks. There were two principal consequences of this. First, only incomplcte sets of all sequential connectivities were obtained, so that often only one sequential connectivity was observed for a given pair of
277
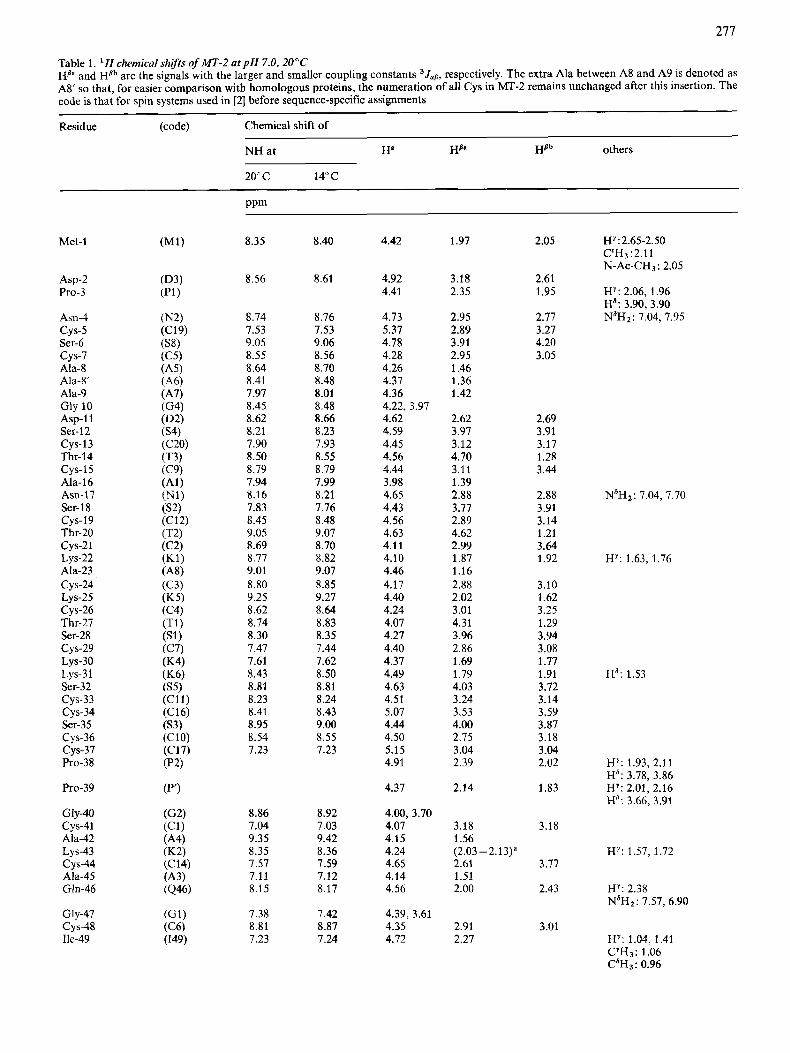
Table 1. ' H chemical shifrs of MT-2 at p H 7.0, 20°C HB" and HBb are the signals with the larger and smaller coupling constants 3Jallr respectively. The extra Ala between A8 and A9 is denoted as AS' so that, for easier comparison with homologous proteins, the numeration of all Cys in h4T-2 remains unchanged after this insertion. The code is that for spin systems used in [2] before sequence-specific assignments
Residue (code) Chemical shift of
NH at Ha HB" Hflh others
20" c 14°C
ppm
Met-1
Asp-2 Pro-3
Asn-4
Ser-6
Ala-8
Ala-9
cys-5
cys-7
Ah-8'
Gly- 10 Asp-1 1 Ser-12
Thr-14
Ala-16 Am17 Ser- 1 8
Thr-20
cys-13
cys-15
cys-19
cys-21 Lys-22
cys-24 Lys-25 CYS-26 Thr-27 Ser-28
Ala-23
cys-29 Lys-30
cys-33 cys-34
CYS-36 cys-37
LYS-3 1 Ser-32
Ser-35
Pro-38
Pro-39
Gly-40 cys-41
Lys-43 cys-44
Ala-42
Ala-45 Gln-46
Gly-47 CYS-48 Ile-49
8.35
8.56
8.74 7.53 9.05 8.55 8.64 8.41 7.97 8.45 8.62 8.21 7.90 8.50 8.79 7.94 8.16 7.83 8.45 9.05 8.69 8.77 9.01 8.80 9.25 8.62 8.74 8.30 7.47 7.61 8.43 8.81 8.23 8.41 8.95 8.54 7.23
8.86 7.04 9.35 8.35 7.57 7.11 8.15
7.38 8.81 7.23
8.40
8.61
8.76 7.53 9.06 8.56 8.70 8.48 8.01 8.48 8.66 8.23 7.93 8.55 8.79 7.99 8.21 7.76 8.48 9.07 8.70 8.82 9.07 8.85 9.27 8.64 8.83 8.35 7.44 1.62 8.50 8.81 8.24 8.43 9.00 8.55 7.23
8.92 7.03 9.42 8.36 7.59 7.12 8.17
7.42 8.87 7.24
4.42
4.92 4.41
4.13 5.37 4.18 4.28 4.26 4.37 4.36 4.22, 3.97 4.62 4.59 4.45 4.56 4.44 3.98 4.65 4.43 4.56 4.63 4.11 4.10 4.46 4.17 4.40 4.24 4.07 4.27 4.40 4.37 4.49 4.63 4.51 5.07 4.44 4.50 5.15 4.91
4.37
4.00, 3.70 4.07 4.15 4.24 4.65 4.14 4.56
4.39, 3.61 4.35 4.72
1.97
3.18 2.35
2.95 2.89 3.91 2.95 1.46 1.36 1.42
2.62 3.97 3.12 4.70 3.11 1.39 2.88 3.77 2.89 4.62 2.99 1.87 1.16 2.88 2.02 3.01 4.31 3.96 2.86 1.69 1.79 4.03 3.24 3.53 4.00 2.75 3.04 2.39
2.14
3.18 1.56
2.61 1.51 2.00
(2.03 -2.13)'
2.91 2.27
2.05
2.61 1.95
2.77 3.27 4.20 3.05
2.69 3.91 3.17 1.28 3.44
2.88 3.91 3.14 1.21 3.64 1.92
3.10 1.62 3.25 1.29 3.94 3.08 1.77 1.91 3.72 3.14 3.59 3.87 3.18 3.04 2.02
1.83
3.18
3.77
2.43
3.01
HY:2.65-2.50 C"Hj ~2.11 N-Ac-CH~: 2.05
HY: 2.06, 1.96 Ha: 3.90,3.90 N'H2: 7.04, 7.95
N'H2: 7.04, 7.70
H Y : 1.63, 1.76
Hd: 1.53
HY: 1.93, 2.11 Ha: 3.78, 3.86 HY: 2.01, 2.16 Ha; 3.66. 3.91
HY: 1.57. 1.72
HY: 2.38 N6Hz: 7.57, 6.90
HY: 1.04, 1.41
CdH3: 0.96 CYH3: 1.06
278
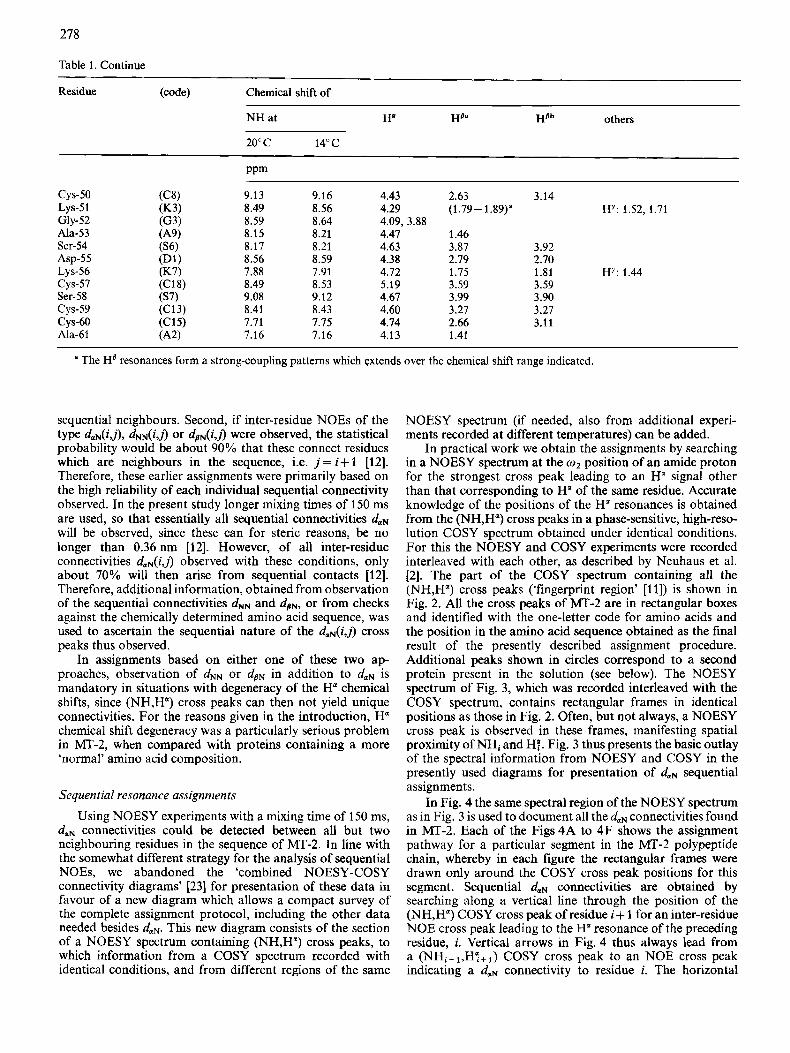
Table 1 . Continue
Residue (code) Chemical shift of
NH at
20" c 14°C
PPm
cys-50 Lys-51 Gly-52 Ala-53 Ser-54 ASP-55 LYS-56 cys-57
cys-59 Ser-58
CYS-60 Ala-61
9.13 8.49 8.59 8.15 8.17 8.56 7.88 8.49 9.08 8.41 7.71 7.16
9.16 8.56 8.64 8.21 8.21 8.59 7.91 8.53 9.12 8.43 7.75 7.16
4.43 2.63 3.14 4.29 (1.79-1.89)' HY: 1.52, 1.71 4.09, 3.88 4.47 1.46 4.63 3.87 3.92 4.38 2.79 2.70 4.12 1.75 1.81 HY: 1.44 5.19 3.59 3.59 4.67 3.99 3.90 4.60 3.27 3.27 4.74 2.66 3.11 4.13 1.41
a The HB resonances form a strong-coupling patterns which extends over the chemical shift range indicated.
sequential neighbours. Second, if inter-residue NOEs of the type duN(i,j), dNN(i,j) or dsN(i,i) were observed, the statistical probability would be about 90% that these connect residues which are neighbours in the sequence, i.e. j = i f 1 [12]. Therefore, these earlier assignments were primarily based on the high reliability of each individual sequential connectivity observed. In the present study longer mixing times of 150 ms are used, so that essentially all sequential connectivities duN will be observed, since these can for steric reasons, be no longer than 0.36 nm [12]. However, of all inter-residue connectivities d,N(i,j) observed with these conditions, only about 70% will then arise from sequential contacts [12]. Therefore, additional information, obtained from observation of the sequential connectivities d" and dSN, Or from checks against the chemically determined amino acid sequence, was used to ascertain the sequential nature of the duN(i,j) cross peaks thus observed.
In assignments based on either one of these two ap- proaches, observation of d" or dpN in addition to duN is mandatory in situations with degeneracy of the Ha chemical shifts, since (NH,H") cross peaks can then not yield unique connectivities. For the reasons given in the introduction, Ha chemical shift degeneracy was a particularly serious problem in MT-2, when compared with proteins containing a more 'normal' amino acid composition.
Sequential resonance assignments
Using NOESY experiments with a mixing time of 150 ms, duN connectivities could be detected between all but two neighbouring residues in the sequence of MT-2. In line with the somewhat different strategy for the analysis of sequential NOEs, we abandoned the 'combined NOESY-COSY connectivity diagrams' [23] for presentation of these data in favour of a new diagram which allows a compact survey of the complete assignment protocol, including the other data needed besides duN, This new diagram consists of the section of a NOESY spectrum containing (NH,H") cross peaks, to which information from a COSY spectrum recorded with identical conditions, and from different regions of the same
NOESY spectrum (if needed, also from additional experi- ments recorded at different temperatures) can be added.
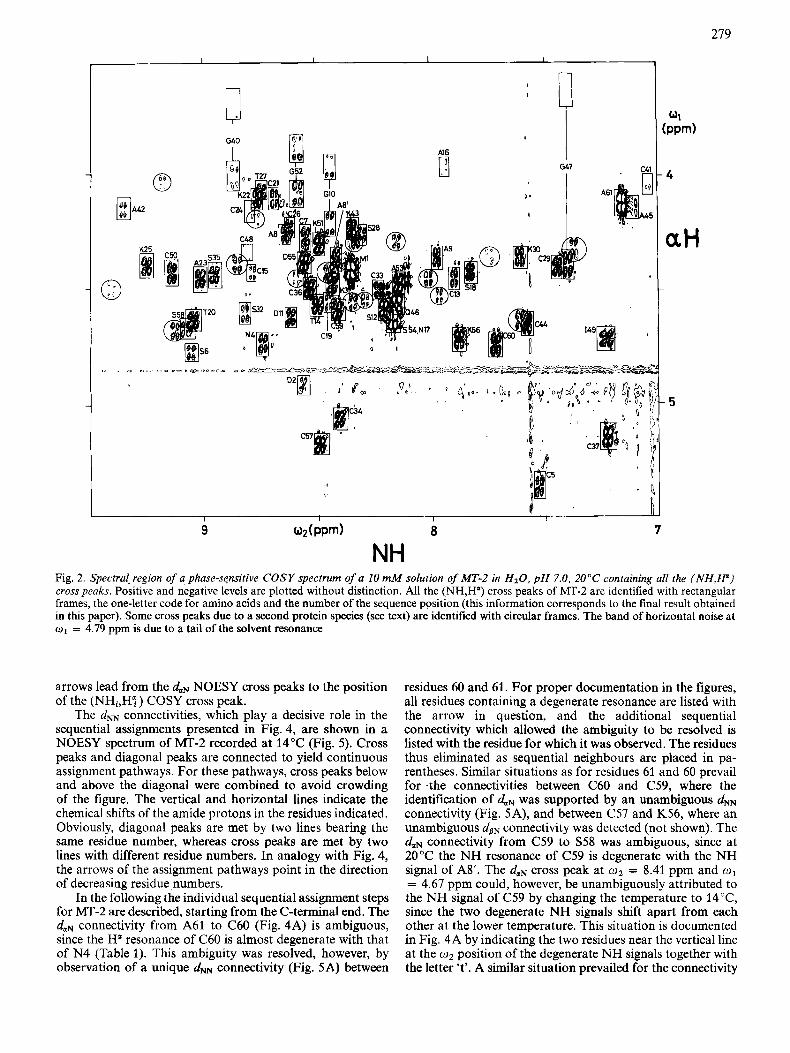
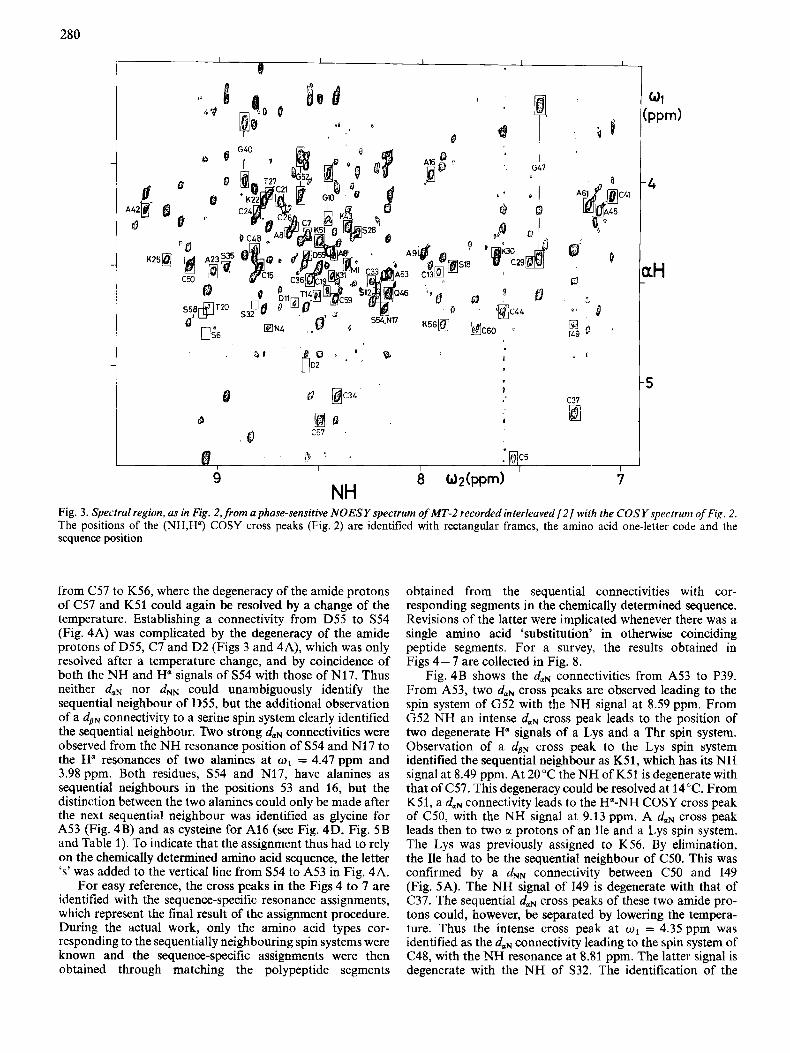
In practical work we obtain the assignments by searching in a NOESY spectrum at the o2 position of an amide proton for the strongest cross peak leading to an H" signal other than that corresponding to H" of the same residue. Accurate knowledge of the positions of the H a resonances is obtained from the (NH,H") cross peaks in a phase-sensitive, high-reso- lution COSY spectrum obtained under identical conditions. For this the NOESY and COSY experiments were recorded interleaved with each other, as described by Neuhaus et al. [2]. The part of the COSY spectrum containing all the (NH,H") cross peaks ('fingerprint region' [ll]) is shown in Fig. 2. All the cross peaks of MT-2 are in rectangular boxes and identified with the one-letter code for amino acids and the position in the amino acid sequence obtained as the final result of the presently described assignment procedure. Additional peaks shown in circles correspond to a second protein present in the solution (see below). The NOESY spectrum of Fig. 3, which was recorded interleaved with the COSY spectrum, contains rectangular frames in identical positions as those in Fig. 2. Often, but not always, a NOESY cross peak is observed in these frames, manifesting spatial proximity of NHi and Hq. Fig. 3 thus presents the basic outlay of the spectral information from NOESY and COSY in the presently used diagrams for presentation of duN sequential assignments.
In Fig. 4 the same spectral region of the NOESY spectrum as in Fig. 3 is used to document all the daN connectivities found in MT-2. Each of the Figs4A to 4F shows the assignment pathway for a particular segment in the MT-2 polypeptide chain, whereby in each figure the rectangular frames were drawn only around the COSY cross peak positions for this segment. Sequential daN connectivities are obtained by searching along a vertical line through the position of the (NH,H") COSY cross peak of residue i+ 1 for an inter-residue NOE cross peak leading to the H a resonance of the preceding residue, i. Vertical arrows in Fig. 4 thus always lead from a (NHi+l,H;+l) COSY cross peak to an NOE cross peak indicating a daN connectivity to residue i. The horizontal
279
1 G40
I
D*w . I ' ff,
c57
7
Fig. 2. Spectru[ region of a phase-sensitive COSY spectrum of a 10 m M solution of MT-2 in HzO, p H 7.0, 20°C containing all ihe ( N H , W ) cross peaks. Positive and negative levels are plotted without distinction. All the (NH,H") cross peaks of MT-2 are identified with rectangular frames, the one-letter code for amino acids and the number of the sequence position (this information corresponds to the final result obtained in this paper). Some cross peaks due to a second protein species (see text) are identified with circular frames. The band of horizontal noise at o1 = 4.79 ppm is due to a tail of the solvent resonance
arrows lead from the daN NOESY cross peaks to the position of the (NHi,HP ) COSY cross peak.
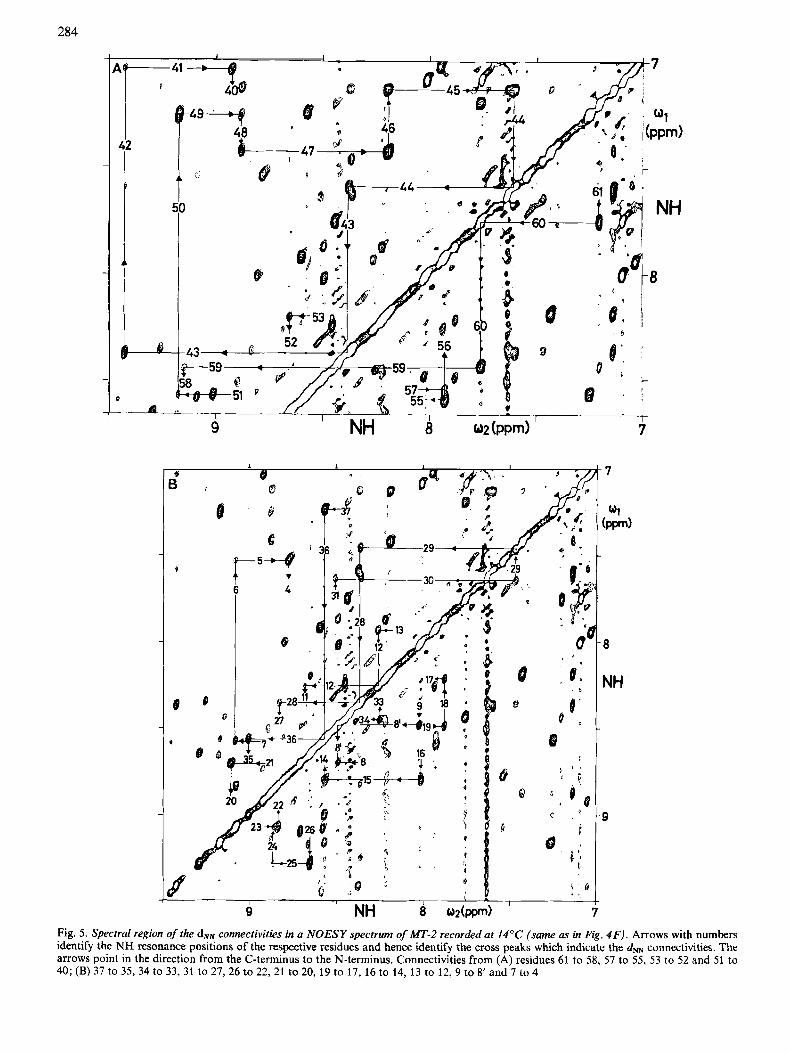
The dNN connectivities, which play a decisive role in the sequential assignments presented in Fig. 4, are shown in a NOESY spectrum of MT-2 recorded at 14°C (Fig. 5). Cross peaks and diagonal peaks are connected to yield continuous assignment pathways. For these pathways, cross peaks below and above the diagonal were combined to avoid crowding of the figure. The vertical and horizontal lines indicate the chemical shifts of the amide protons in the residues indicated. Obviously, diagonal peaks are met by two lines bearing the same residue number, whereas cross peaks are met by two lines with different residue numbers. In analogy with Fig. 4, the arrows of the assignment pathways point in the direction of decreasing residue numbers.
In the following the individual sequential assignment steps for h4T-2 are described, starting from the C-terminal end. The daN connectivity from A61 to C60 (Fig. 4A) is ambiguous, since the Ha resonance of C60 is almost degenerate with that of N4 (Table 1). This ambiguity was resolved, however, by observation of a unique d" connectivity (Fig. 5A) between
residues 60 and 61. For proper documentation in the figures, all residues containing a degenerate resonance are listed with the arrow in question, and the additional sequential connectivity which allowed the ambiguity to be resolved is listed with the residue for which it was observed. The residues thus eliminated as sequential neighbours are placed in pa- rentheses. Similar situations as for residues 61 and 60 prevail for .the connectivities between C60 and C59, where the identification of daN was supported by an unambiguous dNN connectivity (Fig. 5A), and between C57 and K56, where an unambiguous dpN connectivity was detected (not shown). The daN connectivity from (259 to S58 was ambiguous, since at 20°C the NH resonance of C59 is degenerate with the NH signal of A8'. The daN cross peak at o2 = 8.41 ppm and o1 = 4.67 ppm could, however, be unambiguously attributed to the NH signal of C59 by changing the temperature to 14"C, since the two degenerate NH signals shift apart from each other at the lower temperature. This situation is documented in Fig. 4A by indicating the two residues near the vertical line at the o2 position of the degenerate NH signals together with the letter 't'. A similar situation prevailed for the connectivity
280
a
Ad p5 "mc41
0' g
0
P - aH
" I I
9 NH
8 0 2 ( P P d '
t 5 c37 a
7
Fig. 3. Spectral region, as in Fig. 2, from a phase-sensitive NOESY spectrum of MT-2 recorded interleaved [ 2 / with the COSY spectrum of Fig. 2. The positions of the (NH,H") COSY cross peaks (Fig. 2) are identified with rectangular frames, the amino acid one-letter code and the sequence position
from C57 to K56, where the degeneracy of the amide protons of C57 and K51 could again be resolved by a change of the temperature. Establishing a connectivity from D55 to S54 (Fig. 4A) was complicated by the degeneracy of the amide protons of D55, C7 and D2 (Figs 3 and 4A), which was only resolved after a temperature change, and by coincidence of both the NH and Ha signals of S54 with those of N17. Thus neither daN nor dNN could unambiguously identify the sequential neighbour of D55, but the additional observation of a dSN connectivity to a serine spin system clearly identified the sequential neighbour. Two strong daN connectivities were observed from the NH resonance position of S54 and N17 to the Ha resonances of two alanines at o1 = 4.47 ppm and 3.98 ppm. Both residues, S54 and N17, have alanines as sequential neighbours in the positions 53 and 16, but the distinction between the two alanines could only be made after the next sequential neighbour was identified as glycine for A53 (Fig. 4B) and as cysteine for A16 (see Fig. 4D, Fig. 5B and Table 1). To indicate that the assignment thus had to rely on the chemically determined amino acid sequence, the letter 's' was added to the vertical line from S54 to A53 in Fig. 4A.
For easy reference, the cross peaks in the Figs 4 to 7 are identified with the sequence-specific resonance assignments, which represent the final result of the assignment procedure. During the actual work, only the amino acid types cor- responding to the sequentially neighbouring spin systems were known and the sequence-specific assignments were then obtained through matching the polypeptide segments
obtained from the sequential connectivities with cor- responding segments in the chemically determined sequence. Revisions of the latter were implicated whenever there was a single amino acid 'substitution' in otherwise coinciding peptide segments. For a survey, the results obtained in Figs 4- 7 are collected in Fig. 8.
Fig. 4B shows the daN connectivities from A53 to P39. From A53, two daN cross peaks are observed leading to the spin system of G52 with the NH signal at 8.59 ppm. From G52 NH an intense duN cross peak leads to the position of two degenerate H" signals of a Lys and a Thr spin system. Observation of a dsN cross peak to the Lys spin system identified the sequential neighbour as K51, which has its NH signal at 8.49 ppm. At 20°C the NH of K51 is degenerate with that of C57. This degeneracy could be resolved at 14°C. From K51, a duN connectivity leads to the H"-NH COSY cross peak of C50, with the NH signal at 9.13 ppm. A duN cross peak leads then to two CI protons of an Ile and a Lys spin system. The Lys was previously assigned to K56. By elimination, the Ile had to be the sequential neighbour of C50. This was confirmed by a d" connectivity between C50 and I49 (Fig. 5A). The NH signal of I49 is degenerate with that of C37. The sequential duN cross peaks of these two amide pro- tons could, however, be separated by lowering the tempera- ture. Thus the intense cross peak at o1 = 4.35ppm was identified as the daN connectivity leading to the spin system of C48, with the NH resonance at 8.81 ppm. The latter signal is degenerate with the NH of S32. The identification of the
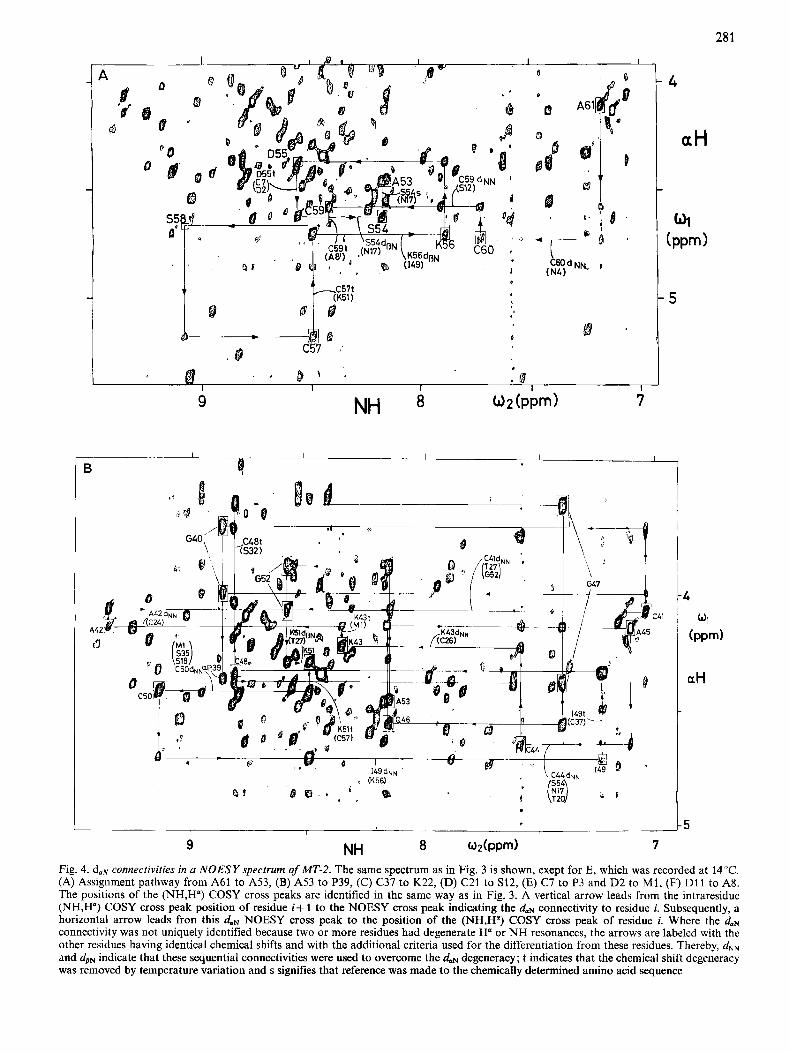
28 1
I ~~ I 4 I 1 1
Fig. 4. doN connectivities in u NOESY spectrum of MT-2. The same spectrum as in Fig. 3 is shown, exept for E, which was recorded at 14°C. (A) Assignment pathway from A61 to A53, (B) A53 to P39, (C) C37 to K22, (D) C21 to S12, (E) C7 to P3 and D2 to M i , (F) D11 to AS. The positions of the (NH,H") COSY cross peaks are identified in the same way as in Fig. 3. A vertical arrow leads from the intraresidue (NH,H") COSY cross peak position of residue i+ 1 to the NOESY cross peak indicating the doN connectivity to residue i. Subsequently, a horizontal arrow leads fron this daN NOESY cross peak to the position of the (NH,H? COSY cross peak of residue i. Where the (laN connectivity was not uniquely identified because two or more residues had degenerate H" or NH resonances, the arrows are labeled with the other residues having identical chemical shifts and with the additional criteria used for the differentiation from these residues. Thereby, dNN and dSN indicate that these sequential connectivities were used to overcome the daN degeneracy; t indicates that the chemical shift degeneracy was removed by temperature variation and s signifies that reference was made to the chemically determined amino acid sequence
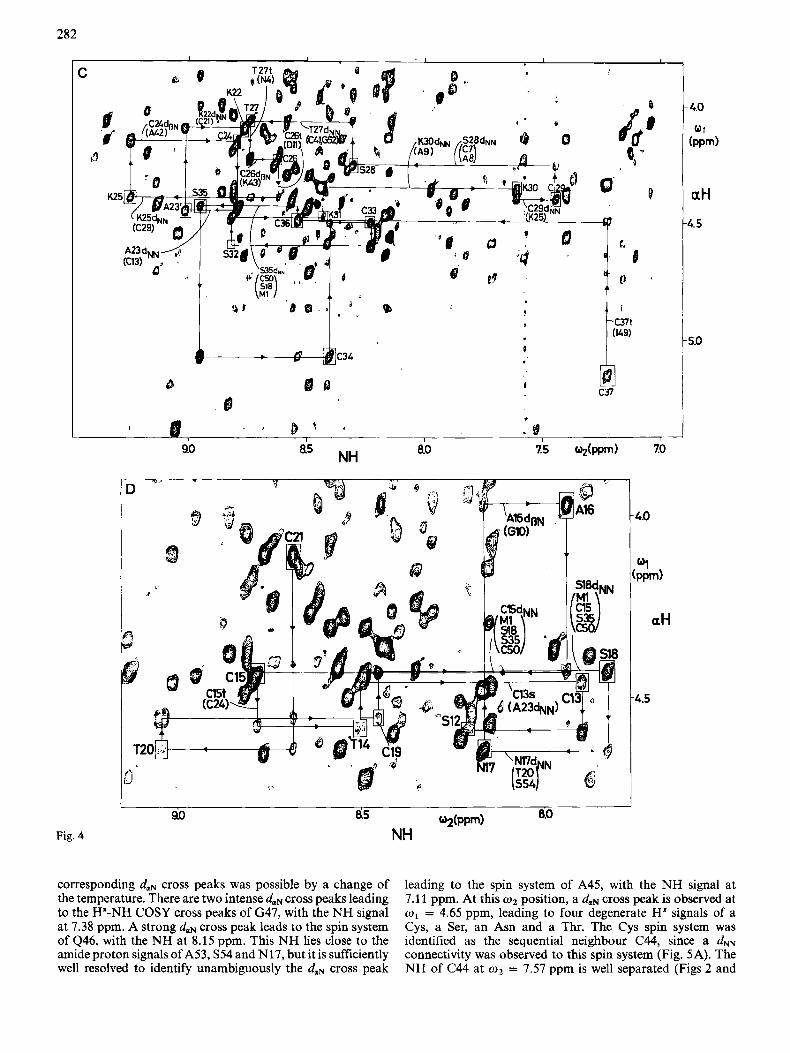
282
Fig. 4
"x,
D
3 p d
-4.5
- 4.0
01 ( P P d
aH
4.5
5.0
corresponding duN cross peaks was possible by a change of the temperature. There are two intense daN cross peaks leading to the H"-NH COSY cross peaks of G47, with the NH signal at 7.38 ppm. A strong daN cross peak leads to the spin system of 446, with the NH at 8.15 ppm. This NH lies close to the amide proton signals of A53, S54 and N17, but it is sufficiently well resolved to identify unambiguously the duN cross peak
leading to the spin system of A45, with the NH signal at 7.1 1 ppm. At this o2 position, a duN cross peak is observed at w1 = 4.65 ppm, leading to four degenerate H" signals of a Cys, a Ser, an Asn and a Thr. The Cys spin system was identified as the sequential neighbour C44, since a dNN connectivity was observed to this spin system (Fig. 5A). The NH of C44 at w2 = 7.57 ppm is well separated (Figs 2 and
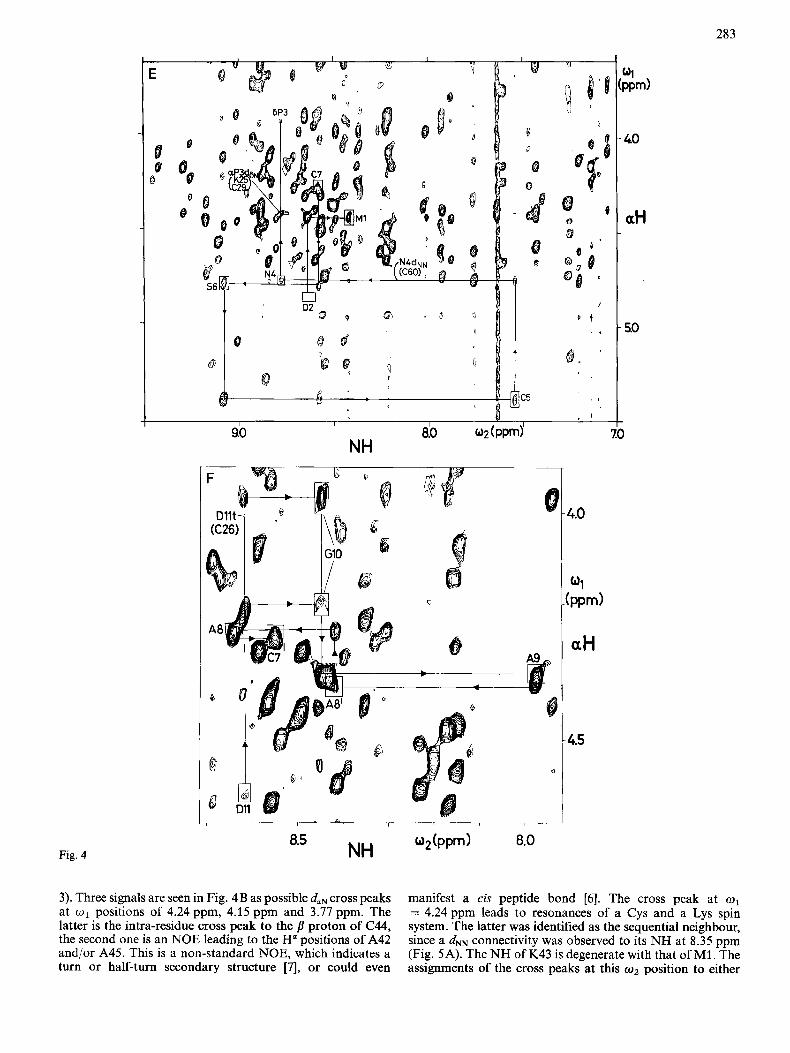
283
Fig. 4 o,(PPm) 8.0 NH 8.5
3). Three signals are seen in Fig. 4B as possible daN cross peaks at w1 positions of 4.24 ppm, 4.15 ppm and 3.77 ppm. The latter is the intra-residue cross peak to the p proton of C44, the second one is an NOE leading to the H a positions of A42 and/or A45. This is a non-standard NOE, which indicates a turn or half-turn secondary structure [7l, or could even
manifest a cis peptide bond [6] . The cross peak at o1 = 4.24 ppm leads to resonances of a Cys and a Lys spin system. The latter was identified as the sequential neighbour, since a d" connectivity was observed to its NH at 8.35 ppm (Fig. SA). The NH of IS43 is degenerate with that of M1. The assignments of the cross peaks at this w 2 position to either
284
P
42
0
7
7
"1 (m)
8
NH
9
Fig. 5. Spectral region of the d" connectivities in a NOESY spectrum of MT-2 recorded at 14°C (same as in Fig. 4F). Arrows with numbers identify the NH resonance positions of the respective residues and hence identify the cross peaks which indicate the &N connectivities. The arrows point in the direction from the C-terminus to the N-terminus. Connectivities from (A) residues 61 to 58, 57 to 55, 53 to 52 and 51 to 40; (B) 37 to 3 5 3 4 to 33, 31 to 27,26 to 22,21 to 20, 19 to 17, 16 to 14, 13 to 12,9 to 8' and 7 to 4
285
84 8.2 8.0 W2(PPm) 8.6
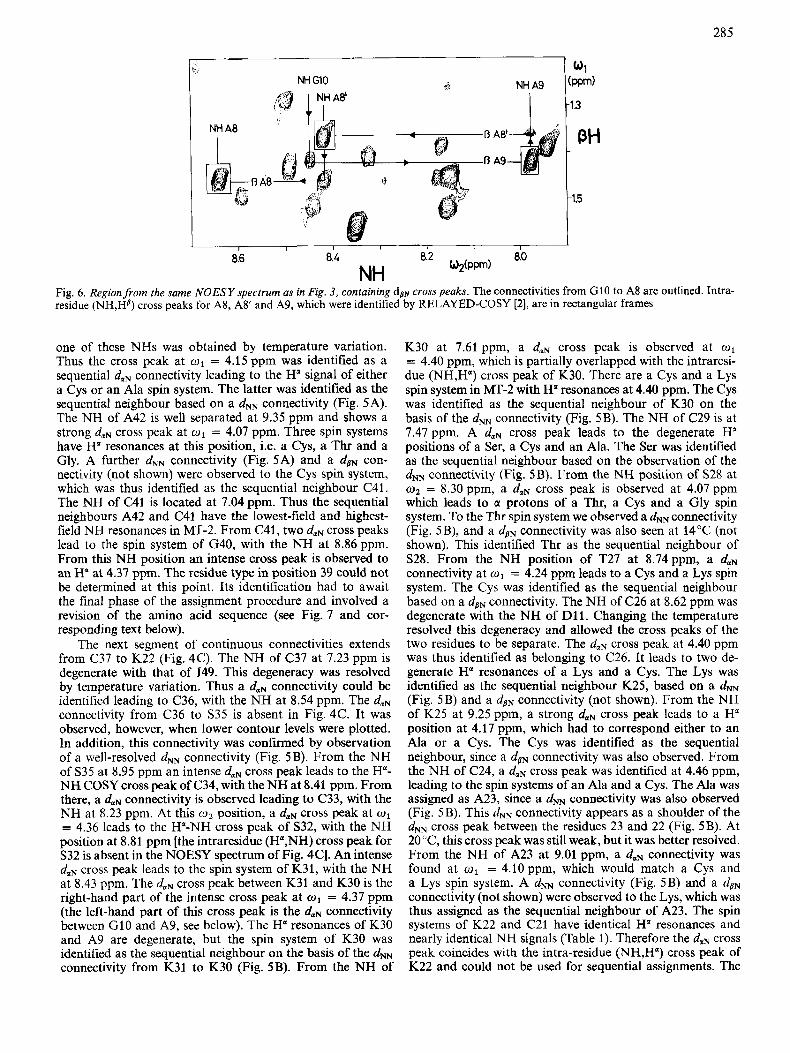
NH Fig. 6 . Region from the same NOESY spectrum as in Fig. 3, containing dp+ crosspeaks. The connectivities from G10 to A8 are outlined. Intra- residue (NH,HP) cross peaks for A8, A8' and A9, which were identified by RELAYED-COSY [2], are in rectangular frames
one of these NHs was obtained by temperature variation. Thus the cross peak at o1 = 4.15 pprn was identified as a sequential daN connectivity leading to the Ha signal of either a Cys or an Ala spin system. The latter was identified as the sequential neighbour based on a d" connectivity (Fig. 5A). The NH of A42 is well separated at 9.35 ppm and shows a strong dorN cross peak at w1 = 4.07 ppm. Three spin systems have Ha resonances at this position, i.e. a Cys, a Thr and a Gly. A further d" connectivity (Fig. 5A) and a dSN con- nectivity (not shown) were observed to the Cys spin system, which was thus identified as the sequential neighbour C41. The NH of C41 is located at 7.04 ppm. Thus the sequential neighbours A42 and C41 have the lowest-field and highest- field NH resonances in m - 2 . From C41, two daN cross peaks lead to the spin system of G40, with the NH at 8.86ppm. From this NH position an intense cross peak is observed to an Ha at 4.31 ppm. The residue type in position 39 could not be determined at this point. Its identification had to await the final phase of the assignment procedure and involved a revision of the amino acid sequence (see Fig. 7 and cor- responding text below).
The next segment of continuous connectivities extends from C37 to K22 (Fig. 4C). The NH of C37 at 7.23 ppm is degenerate with that of 149. This degeneracy was resolved by temperature variation. Thus a daN connectivity could be identified leading to C36, with the NH at 8.54 ppm. The dmN connectivity from C36 to S35 is absent in Fig. 4C. It was observed, however, when lower contour levels were plotted. In addition, this connectivity was confirmed by observation of a well-resolved d" connectivity (Fig. 5B). From the NH of S35 at 8.95 ppm an intense daN cross peak leads to the Ha- NH COSY cross peak of C34, with the NH at 8.41 ppm. From there, a daN connectivity is observed leading to C33, with the NH at 8.23 ppm. At this w 2 position, a daN cross peak at o1 = 4.36 leads to the Ha-NH cross peak of S32, with the NH position at 8.81 ppm [the intraresidue (Ha,NH) cross peak for S32 is absent in the NOESY spectrum of Fig. 4C]. An intense daN cross peak leads to the spin system of K31, with the NH at 8.43 ppm. The dolN cross peak between K31 and K30 is the right-hand part of the intense cross peak at o1 = 4.37 pprn (the left-hand part of this cross peak is the daN connectivity between G10 and A9, see below). The Ha resonances of K30 and A9 are degenerate, but the spin system of K30 was identified as the sequential neighbour on the basis of the dNN connectivity from K31 to K30 (Fig. 5B). From the NH of
K30 at 7.61 ppm, a daN cross peak is observed at w1 = 4.40 ppm, which is partially overlapped with the intraresi- due (NH,H") cross peak of K30. There are a Cys and a Lys spin system in MT-2 with H" resonances at 4.40 ppm. The Cys was identified as the sequential neighbour of K30 on the basis of the d" connectivity (Fig. 5B). The NH of C29 is at 7.47ppm. A daN cross peak leads to the degenerate H a positions of a Ser, a Cys and an Ala. The Ser was identified as the sequential neighbour based on the observation of the d" connectivity (Fig. SB). From the NH position of S28 at w 2 = 8.30 ppm, a daN cross peak is observed at 4.07 ppm which leads to a protons of a Thr, a Cys and a Gly spin system. To the Thr spin system we observed a d" connectivity (Fig. 5B), and a dsN connectivity was also seen at 14°C (not shown). This identified Thr as the sequential neighbour of S28. From the NH position of T27 at 8.74ppm, a daN connectivity at w1 = 4.24 ppm leads to a Cys and a Lys spin system. The Cys was identified as the sequential neighbour based on a dpfJ connectivity. The NH of C26 at 8.62 ppm was degenerate with the NH of D11. Changing the temperature resolved this degeneracy and allowed the cross peaks of the two residues to be separate. The daN cross peak at 4.40 ppm was thus identified as belonging to C26. It leads to two de- generate Ha resonances of a Lys and a Cys. The Lys was identified as the sequential neighbour K25, based on a dNN (Fig. 5B) and a dsN connectivity (not shown). From the NH of K25 at 9.25 ppm, a strong daN cross peak leads to a H" position at 4.17 ppm, which had to correspond either to an Ala or a Cys. The Cys was identified as the sequential neighbour, since a dSN connectivity was also observed. From the NH of C24, a daN cross peak was identified at 4.46 ppm, leading to the spin systems of an Ala and a Cys. The Ala was assigned as A23, since a d" connectivity was also observed (Fig. 5s). This dNN connectivity appears as a shoulder of the d" cross peak between the residues 23 and 22 (Fig. 5B). At 20°C, this cross peak was still weak, but it was better resolved. From the NH of A23 at 9.01 ppm, a daN connectivity was found at w1 = 4.10 ppm, which would match a Cys and a Lys spin system. A d" connectivity (Fig. 5B) and a dSN connectivity (not shown) were observed to the Lys, which was thus assigned as the sequential neighbour of A23. The spin systems of K22 and C21 have identical H a resonances and nearly identical NH signals (Table 1). Therefore the daN cross peak coincides with the intra-residue (NH,H") cross peak of K22 and could not be used for sequential assignments. The

286
N1
3.9 3.7
, E
Fig. 7. Regions of NOESY spectra containing cross peaks between hydrogen atoms of proline and ( A ) the w i d e protons of G4O and N4, ( B ) the amide proton of C37, ( C ) the u protons of 0 2 . P38 and C37. ( D ) the 6 protons of P3, P38 and P39. Intra-residue NOE cross peaks are in rectangular frames. (A), (B) and (D) are from the same NOESY spectrum of "2Cd-enriched h4T-2 in H20; (C) is part of a NOESY spectrum of "3Cd-enriched MT-2 in 'H,O; the intra-residue cross peak of C37 H" with the degenerate CBH2 is split along the w , axis due to the scalar coupling of Ha with 'I3Cd; (E) shows horizontal cross sections of (C) from w2 = 4.8-5.0 ppm a t the w1 positions indicated in the figure. To recognize the fine structure of the cross peaks, a Fourier transformation with high digital resolution (1.6 Hz/point) was used
8 9 10 a 17 20 23 39 -A-ADG-T- K-~-s- -
B -A-A-A-G-D--N-T-A- P-
Fig. 8. Summary of sequential 'H-NMR assignments ,for MT-2 and revisions of the primary structure. (A) Amino acid sequence of MT-2 including the modifications relative to the published primary structure. Above the sequence a survey is presented of all the sequential NOE connectivities (Fig. 1) observed in MT-2. (B) The upper trace corresponds to the published MT-2 sequence [lo], the lower trace indicates thc changes which resulted from the NMR studies and additional chemical investigations (see text)
rn 30 40 50 W A 8 8' 9 10 11 17 20 23 39 N)
AC -MDPNCSCAAAGDSC TCANSC TC KACKCT sc K K S C CSCCPPGCAKCAQGC I c KGA SD K C SCCA
d" cross peak would be too close to the diagonal to be recognized, but there was a d,,cross peak (not shown) which allowed the sequential connectivity to be established (Fig. 8).
Fig. 4D shows the daN connectivities from C21 to S12. From the NH resonance of C21 at 8.69 ppm, two cross peaks to Ha resonances are observed at 4.56 ppm and 4.63 ppm,
leading to a Cys and a Thr spin system, respectively. These spin systems were identified as C19 and T20. Thus there is, besides the sequential daN connectivity, also a daN(i,i+ 2) cross peak. The observation of the two cross peaks of comparable intensity (Fig. 4D) might indicate that the sequence order of the Cys and the Thr spin system could be interchanged. This

287
is not possible, however, since a strong sequential daN cross peak between the NH of the Thr and the Ha of the Cys is observed, but not between the NH of the Cys and the Ha of the Thr (Fig. 4D). This provides firm evidence for the assignment as C19-T20. A daN cross peak from the NH of C19 at 8.45 ppm leads to degenerate Ha resonances of two Cys, two Ser and one Met, One of the Ser spin systems was identified as the sequential neighbour of C19, since a dNN connectivity was also observed (Fig. 5B). From the NH position of S18 at 7.83 ppm, a daN cross peak at 4.65 ppm leads to three degener- ate H a signals of an Asn, a Thr and a Ser spin system. The Asn was identified as the sequential neighbour of S18 based on observation of a dNN cross peak (Fig. 5B) and a dPN connectivity. The NH of N17 at 8.16 ppm and its Ha overlap with the corresponding resonances of S54, as discussed above. Two daN cross peaks are observed at 4.47 ppm and 3.98 ppm. The former had been identified as the connectivity between S54 and A53 (Fig. 4A) and the latter is the connectivity leading from N17 to its sequential neighbour A16. The Ha of A16 is degenerate with one Ha of G10. Ala was identified as the sequential neighbour of N17 from observation of a dPN cross peak. From the NH of A16 at 7.94ppm, a daN cross peak to its sequential neighbour is observed at 4.44ppm. This H a signal is degenerate with four other Ha resonances (Fig. 4D). Among these, a Cys was identified as the sequential neighbour, since a dm cross peak was also observed (Fig. 5B). At 20"C, the NH of C15 is degenerate with that of C24. At 14"C, these two resonances shifted apart, so that the cross peaks belonging to the respective NH resonances could be distinguished and the cross peak at o1 = 4.56 ppm identified as the daN connectivity from C15 to T14. From the NH of TI4 at 8.50 ppm, a daN cross peak leads to the Ha signal of C13, which is degenerate with that of A23. As discussed above, A23 was sequentially assigned on the basis of a d" connectivity, so that the second cross peak with this a-proton chemical shift can now be attributed to C13. On the amino-terminal side, the assignment of C13 is neatly documented by unambiguous
The assignments for the segment from residues S12 to C7 called for major modifications of the amino acid sequence, as will be discussed in the following section. In the segment from C7 to M1 the Ha signals of N4 and S6 are bleached out by the irradiation of the solvent line at 20°C. Therefore Fig. 4E shows a NOESY spectrum recorded from the same sample at 14°C. From the NH of C7 at 8.55 ppm, a daN cross peak is observed at 4.78 ppm leading to the spin system of S6, with the NH resonance at 9.05 ppm. An intense daN cross peak at 5.37 ppm leads to the Ha of C5. The NH of C5 is at 7.53 ppm and shows a daN cross peak to N4. Independently, a continu- ous train of dNN connectivities was observed for this segment (Fig. 5B). At the NH position of N4 at 8.74 ppm, cross peaks are found at 4.41 ppm and 3.90 ppm. These are the H a and H6 positions of P3. The Ha of P3 is degenerate with those of K25 and C29, so that this sequential connectivity relies on the daN cross peak. Finally, a daN cross peak is observed between the previously identified unique methionine spin system [2] and an AMX spin system, which were attributed to positions 1 and 2 from reference to the sequence and from observation of sequential connectivities to P3 (see below).
daN and d" cross peaks with s12 (Figs 4D, 5B).
Modification of the chemically determined amino acid sequence and sequential assignments with proline residues
As mentioned in the introduction, the amino acid composition reflected in the 'H spin system identifications [2]
varied from that of the published sequence [lo] by the presence of three additional spin systems corresponding to Ala, Asn and Pro and by the absence of one each of Lys and Ser. From the sequential assignments discussed so far, we found that Lys in position 20 had to be replaced by Thr and that Thr in position 17 must be substituted by Asn (Fig. 4D). These changes keep the number of threonines constant and take account of the additional Asn and the missing Lys. The missing links in the sequential assignments, i.e. for the segments of residues 39 - 37 and 12 - 7 (Figs 4 and 5), could only be established in conjunction with further revisions of the sequence.
After identification of the sequential connectivities be- tween P3S and C37, G10 and A9, and AS and C7 (see below), we had, according to the published sequence [lo], established at least one sequential connectivity for all sequence positions except residue 39. For this single position, however, there remained two as-yet unassigned spin systems of Ala and Pro. A further NMR observation was that there were NOES from A9 and AS to the remaining Ala spin system, but not between A8 and A9. Chemical reinvestigation of the sequence by Hunziker et al. [24] then resulted in the identification of two peptide segments containing, respectively, Pro-Pro and Ala- Ala-Ala. Using this information, a consistent interpretation of the NMR data was obtained, with the previously unassigned Pro and Ala spin systems in positions 39 and 8', respectively, showing that MT-2 consists of 62 rather than 61 residues. The additional residue is between position 8 and 9 and is denoted 8'. The crucial sequential assignments are described in the following.
Starting from the Ha-NH COSY cross peak of S12 identified in Fig. 4D, a dNN connectivity was found to an aspartate spin system in position 11 (Fig. 5B). At 20°C the NH resonance of D11 is degenerate with that of C26, but the two lines could be separated at 14°C. Two daN cross peaks are observed between the NH of D11 and the Ha resonances of a Gly spin system, which was therefore assigned to position 10. From the NH of G10 at 8.45 ppm, further daN connectivities lead to three subsequent Ala spin systems, which we denote A9, A8' and A8; A8 is further linked with C7 by a daN connectivity. For A9 and A8, the intra-residue (NH,H") cross peaks are partially overlapped with the sequential dolN cross peaks leading to A8' and C7, respectively, but independently, nicely resolved sequential connectivities dPN confirmed the presence of the -Ala-Ala-Ala- segment. In Fig. 6, the intra- residue (NH,HP) cross peaks, identified by RELAYED-COSY are in rectangular frames, analogous to the intra-residue COSY cross peaks in Fig. 4. At the NH position of G10 at w2 = 8.45ppm, a dBN cross peak is observed at w2 = 1.42 ppm, which leads to the methyl resonance of A9. The
NH signal of A9 at o2 = 7.97 ppm shows a dBN cross peak at w1 = 1.36 ppm, which is the position of the P-methyl reso- nance of A8'. From the NH of A8' at w2 = 8.41 ppm, a further dPN connectivity leads to the methyl resonance of A8. Overall, we thus have sequential connectivities for a segment C7-A%A8'-A9-GlO-D11, whereas the corresponding segment in the published sequence is C7-A8-A9-DlO-G11.
In sequential assignments with Pro residues, the 6 protons of Pro take the role of the amide protons in other amino acid residues [l 1,121. The sequential connectivities dd are generally more dimcult to resolve and connecting the H' signals of Pro with HY, HB and Ha is usually more dificult than the attachment of the NH resonances to the side-chain spin systems in other residues. In MT-2 we had to use both COSY and NOESY to establish the intra-residue connectivities. For

288
P38, two (Ha,HB) cross peaks and one (HB,Hp) cross peak had been found in COSY [2]; in NOESY four (H6,Hy) cross peaks and one (Hd,HB) cross peak were found (see Fig. 7D and also Fig. 11 of [2]). Sequential NOES were observed between CdH2 of P38 and NH, Ha and CPH2 of C37 (Fig. 7B, C and D), so that the sequence-specific assignment of P38 is firmly estab- lished. For P3 and P39, the COSY connectivities within the fragments CaH-CBHz had been found [2]. daN and two dBN connectivities from each of these partial spin systems to N4 and G40, respectively, could now be observed (Fig. 7A). The CYH2-C6H2 fragments of these two prolines could be individu- ally assigned, even though the resonances of CdH2 of P3 and of one 6 proton of P39 partially overlap at 3.89 - 3.91 ppm. From this resonance position, NOE cross peaks were observed to one HB of P3 (Fig. 7D), to the NH of N4 (Fig. 7A) and to the Ha of D2 (Fig, 7C), which presents solid evidence that at least one &proton line of P3 is located near 3.90ppm. A further NOE cross peak leads from this resonance position to resonances near 4.92 ppm (Fig. 7C). This could correspond to the H" signals of either or both of the residues D2 and P38, so that this cross peak may contain the sequential dd connectivity between D2 and P3, between P38 and P39, or both. The cross peak at (wz = 4.91 ppm, w1 = 3.66 ppm) in Fig. 7C, which connects one H6 of Pro at 3.66 ppm with the H" of either D2 or P38, then provided the clue for the distinc- tion of the H6 and HY resonances of P3 and P39. For this we plotted horizontal cross sections through various cross peaks located at w2 = 4.92 ppm (Fig. 7E). The cross peak at w1 = 2.39 ppm is well isolated and connects Ha of P38 with its own HB. This cross section serves to recognize the line shape and the exact line position of P38 H". Obviously, it has the same shape and position as the peak at o1 = 3.66 ppm, but it is significantly different from the cross section at o1 = 2.61 ppm, which represents the line shape and position of D2
Ha in an intra-residue (H",HB) cross peak. Thus, the Hd proton at 3.66ppm was attributed to the sequential neighbour of P38, i.e. P39. The cross peak at o1 = 3.90 - 3.91 ppm consists mainly of the dad connectivity between D2 and P3; it may also contain, however, a minor contribution from a dad con- nectivity between P38 and P39.
Resonance assignments for 'protein impurities'
The COSY fingerprint of the MT-2 solution used for the resonance assignments (Fig. 2) and the other regions of COSY and NOESY spectra contain numerous cross peaks in addi- tion to those attributed to MT-2. Initially it was imperative properly to distinguish these extra peaks (which have in most instances lower intensities) from those of the major species. This was achieved by comparison of the spectra obtained from different protein preparations, which contained variable admixtures of the minor component (10-30%) if the conditions for the NMR experiments were the same (our unpublished results). We thus had evidence that more than one protein species was present in the preparations of MT-2. After the sequential assignments of the major species, smaller parts of the minor species could also be sequentially assigned. Because of the lower concentration and severe problems of overlap with the stronger signals of the major species, a complete assignment was not possible. The largest continuous segment that could be sequentially assigned is eight residues long and has an amino acid sequence identical to residues 39-45 of MT-2, except for replacements of P39 by alanine and A42 by threonine. Since this peptide segment does not
MT-2, we have evidence that the second species is a chemically different, homologous protein, and it can be ruled out that the second species is a different conformation of MT-2. With respect to the partial sequence A39 to A45, the minor species resembles MT-1, while it is identical to MT-2 with respect to its charge (which is -3 for MT-2 and -2 for MT-1). That the minor species is a homologous metallothionein species was further corroborated with heteronuclear COSY experiments using ' 13Cd-enriched samples. In the '' 3Cd spectrum, extra peaks for the minor species could be recognized. Hetero- nuclear COSY experiments as reported by Frey et al. [4] showed correlations between these extra '13Cd peaks and 'H resonances of the minor species. In particular, the spin systems C41 and C44 of the minor species both correlate with a '13Cd signal at slightly higher field than Cd6 in MT-2, which is bound to C41 and C44 [4].
Compared to MT-2, a smaller number of resolved IT-NH COSY cross peaks of the minor protein species is seen in Fig. 2. When plotting this spectral region with lower contour levels, 37 such cross peaks could be detected. The partial sequential assignments in the minor species indicate, however, that the H"-NH COSY cross peaks for the residues 46-61 coincide closely with those of MT-2, indicating that the minor species has a similar size as MT-2.
CONCLUSION
The sequential 'H-NMR assignments for MT-2 obtained with the experiments of the preceding section are summarized in Fig. 8A and the revisions of the primary structure which resulted from the present studies and from chemical re- investigation of the sequence [24] are surveyed in Fig. 8B. With regard to the latter, it is worth noting that the dipeptide segment G10-D11 has also been reported for human MT [25]. MT-2 is the first protein containing neither aromatic side chains nor regular secondary structure elements [5, 71 for which sequence-specific resonance assignments have been obtained. It should be reemphasized that the procedures de- scribed in this paper depend critically on the previous, complete identification of all amino acid spin systems, which was possible only with the use of advanced, high-resolution 2D-NMR experiments recorded in the phase-sensitive mode 121.
We thank Miss. M. Sutter for the preparation of the biological material and Mrs E.-H. Hunziker for the careful preparation of the illustrations. This research was supported by a fellowship from the Science and Engineering Research Council (UK) (Overseas post- doctoral fellowship to D.N.) and by the Schweizerischer Nationalfonds (projects 3.284-0.82 and 3.207-0.82). An EMBO fellowship to E.W. is gratefully acknowledged.
REFERENCES 1. Neuhaus, D., Wagner, G.,VaSkk, M., KIgi, J. H. R. & Wiithrich,
K. (1984) Eur. J . Biochem. 143,659-667. 2. Neuhaus, D., Wagner, G., VaSak, M., Klgi, J. H. R. & Wuthrich,
K. (1985) Eur. J . Biochem. 151,257-273. 3. Wiithrich, K., Wider, G., Wagner, G. & Braun, W. (1982) J. Mol.
Biol. 155, 311-319. 4. Frey, M. H., Wagner, G., VaSak, M., Ssrensen, 0. W., Neuhaus,
D., Worgotter, E., Kagi, J. H. R., Ernst, R . R. & Wiithrich, K. (1985) J. Am. Chem. SOC. 107,6847-6851.
5. Braun, W., Wagner, G., Worgotter, E., VaSak, M., Kagi, J. H. R. & Wiithrich, K. (1986) J. MoZ. Biol. 187, 125-129.
6. Wiithrich. K.. Billeter. M. & Braun. W. (1984) J. Mol. Biol. 180. , . coincide with any other part of the amino acid sequence of 7 1 5 - 740. '

289
7. Wagner, G., Neuhaus, D., Worgotter, E., VaSak, M., Kagi, J. H. R. & Wuthrich, K. (1986) J. Mol. Biol. 187,131 -135.
8. Melis, K. A., Carter, D. C., Stout, C. D. & Winge, D. R. (1983) J. Biol. Chem. 258,6255 -6257.
9. Furey, W. F., Robbins, A. H., Clancy, L. L., Winge, D. R., Wang, B. C. & Stout, C. D. (1986) Proceedings of 2nd International Meeting on Metallothionein (Kagi, J. H. R., ed.) Birkhauser, Basel, in the press.
10. Kimura, M., Otaki, N. & Imano, M. (1979) in Metallothionein (Kagi, J. H. R. & Nordberg, M., eds) pp. 163 - 168, Birkhauser, Basel.
11. Wagner, G. & Wiithrich, K. (1982) J. Mol. Bid. 155, 347-384. 12. Billeter, M., Braun, W. & Wiithrich, K. (1982) J. Mol. Biol. 155,
13. Kagi, J. H. R., Himmelhoch, S. R., Whanger, P. D., Bethune, J. L. & Vallee, B. L. (1974) J. Biol. Chem. 249,3537-3542.
14. VaSak, M., Nicholson, J. K., Hawkes, G. E. & Sadler, P. J. (1985) Biochemistry 24, 740 - 741.
15. Buhler, R. H. 0. & Kagi, J. H. R. (1979) in Metallothionein (Kagi, J. H. R. & Nordberg, M., eds) pp. 21 1 - 220, Birkhauser, Basel.
16. Piantini, 0. W., Serrensen, 0. W. & Ernst, R. R. (1982) J . Am. Chem. Soc. 104,6800-6801.
321 - 346.
17. Shaka, A. J. & Freeman, R. (1983) J. Mugn. Reson. 51, 169- 173.
18. Rance, M., Ssrensen, 0. W., Bodenhausen, G., Wagner, G., Ernst, R. R. & Wiithrich, K. (1984) Biochem. Biophys. Res. Commun. 117,479-485.
19. Jeener, J., Meier, B. H., Bachmann, P. & Ernst, R. R. (1979) J . Chem. Phys. 71,4546-4553.
20. Anil Kumar, Emst, R. R. & Wuthrich, K. (1980) Biochem. Bio- phys. Res. Commun. 95, 1 - 6.
21. Marion, D. & Wuthrich, K. (1983) Biochem. Biophys. Res. Commun. 113,967-974.
22. Wagner, G., Tschesche, H. & Wiithrich, K. (1978) Eur. J. Bio- chem. 86,67 - 76.
23. Wagner, G., Anil Kumar & Wiithrich, K. (1981) Eur. J. Biochem.
24. Hunziker, P. E., Kanzig, A., Kaur, P. & Kagi, J. H. R. (1986) Proceedings 2nd International Meeting on Metallothionein (Kagi, J. H. R., ed.) Birkhauser, Basel, in the press.
25. Kissling, M. M. & Kagi, J. H. R. (1977) FEBS Lett. 82, 247- 250.
114, 375-384.





























