Embed Size (px)
Citation preview

Chiroptera Neotropical 17(1), July 2011
963
Skull variation of the vampire bat Desmodus rotundus (Chiroptera:
Phyllostomidae): Taxonomic implications for the Cuban fossil vampire bat Desmodus puntajudensis
Johanset Orihuela 1*
1. 15921 SW 44th ST Miami, Fl 33185. * Corresponding author. Email: [email protected] Abstract The Cuban fossil vampire bats of the Neotropical genus Desmodus have been recently raised to specific rank. However, the dichotomous taxonomic history of the Cuban Desmodus fossils, plus the discovery of additional and more complete specimens prompted a reanalysis of the previous taxonomic arrangements. Here I document the range of skull variation in Desmodus rotundus geographical populations through a study of qualitative and quantitative osteological analyses to find the taxonomic affinity of the Cuban fossil Desmodus. It was found that the shape and size of D. rotundus skulls are widely variant, especially in the shape of the rostrum, palatal construction, zygomatic morphology, and occipital orientation; characters that have been often employed to separate the Cuban fossils from the extant form. This study demonstrates that the discrete morphology and measurements used to separate the Cuban fossils from the extant D. rotundus do not support the new taxonomic arrangement. The Cuban sample falls instead within the variation range observed in D. rotundus, implying that the fossil Desmodus sample from Cuba belong to the extant D. rotundus. Additionally, sexual dimorphism and minor geographical variation in the size of D. rotundus skulls was statistically supported. These results support the importance of quantitative and qualitative studies in fossil bat systematic taxonomy towards a better understanding of their past biogeography, diversity, and extinction. Keywords: Cranial variation, morphology, Desmodus, vampire bats, Cuba. Introduction
The vampire bat subfamily Desmodontinae is endemic to the New World Neotropics where they are integrated by three extant species: Diphylla ecaudata (Spix 1823), Diaemus youngi (Jentink 1893), and Desmodus rotundus (Geoffroy 1810) (Greenhall et al., 1988; Koopman, 1988; Simmons, 2005). Of these, the common vampire bat Desmodus rotundus is the most widespread. It has a geographical range that extends from Sonora and Tamaulipas in northern Mexico through all Middle America to northern Argentina and northern Chile, including the islands of Margarita and Trinidad in the Caribbean basin. The remaining two species share a similar distribution pattern, but are less common (Greenhall et al., 1983; 1988; Koopman, 1988; Simmons, 2005).
The genus Desmodus is polytypic. Two subspecies have been often recognized within D. rotundus: D. rotundus murinus in Mesoamerica and D. rotundus rotundus in South America (Greenhall et al., 1988; Martins, 2008). However, other authors consider these geographical races to represent a single form D. rotundus (Koopman, 1988; Simmons, 2005). In addition, three extinct species of vampire bats are known from North,
Central, and South American fossils (Morgan et.al., 1988; Greenhall and Schmidt, 1989; Czaplewskiet al., 2003; Pardiñas and Tonni, 2000). Two of these, Desmodus stocki and Desmodus dracualae, are essentially recognized by their much larger size and slight morphological characteristics (Morgan et al., 1988; Czaplewski and Peachey 2003). The third species, Desmodus archeodaptes, is from the late Pliocene or early Pleistocene of northern Florida. This Desmodus falls within the size range and shares morphological traits with Desmodus rotundus. D. archaeodaptes is the oldest vampire bat known thus far. For more detailed information on the morphology and systematics of these taxa see Jones (1958), Gut (1959), Hutchison (1967), Morgan et al. (1988), Greenhall et al. (1983), Rey et al.(1988), Greenhall and Schmidt (1988), Arroyo-Cabrales and Ray (1997), Czaplewski and Cartelle (1998), Czaplewski et al. (2003), Czaplewski and Peachy (2003).
In the Greater Antilles, the genus Desmodus is known only from a handful of fossil specimens from Cuban Quaternary cave deposits (Koopman, 1958; Morgan et al., 1988; Morgan, 1991, 2001; Jiménez et al., 2005; Suárez, 2005). Arredondo

Chiroptera Neotropical 17(1), July 2011
964
(1958) and Koopman (1958) reported the first specimen, a heavily calcified rostrum from a supposed late Pleistocene faunal assemblage from Cueva Lamas, western Havana city (opp. Cit.). Koopman considered this specimen as “clearly referable” to Desmodus rotundus, conclusion that Clayton Ray later revised and supported (Koopman, 1958; Ray et al., 1988). In 1974, a second specimen was discovered in a supposed Holocene deposit of Centenario de Lenin cave in Punta Judas, northern Sanctí Spirítus province, central Cuba. Woloszyn and Mayo (1974) described this incomplete cranium as a new endemic subspecies Desmodus rotundus puntajudensis. More recently, the Cuban fossil sample was re-described and elevated to specific taxonomic rank by Suárez (2005), based on new remains from a supposed late Pleistocene assemblage from Cueva de los Paredones, western Havana and the middle Holocene deposits of Cueva Blancas in southwestern Havana (Jiménez et al., 2005).
Historically, the taxonomy of the Cuban fossil Desmodus has been divisive, and far from conservative. Depending on the author, the specimens have been recognized as undistinguishable from the extant Desmodus rotundus (Koopman, 1958; Fischer, 1977), as an endemic subspecies (Woloszyn and Mayo, 1974), as synonymous of the extinct taxon D. magnus (Varona, 1974), or as recently, a new species (Suárez, 2005). The sources for the uncertainty of whether the Cuban fossils represent a new form or belong to the extant D. rotundus are due to the fragmentary nature of the fossil specimens, small sample size, and the availability of comparative morphologic studies based on the osteology of the extant Desmodus populations. For example, previous authors compared the Cuban fossils to geographically limited samples of less than 10 recent specimens, usually from Mexico (e.g., Woloszyn and Mayo 1974; Suárez 2005), while others made unjustified taxonomic arrangements (e.g. Varona, 1974).
The comparison of a geographically diverse neontological sample with the Cuban specimens that could help solve these dichotomies is timely. Previous observations and the discovery of two additional Desmodus specimens (Orihuela, in press), including an almost complete skull, prompted the reanalysis of the Cuban fossil Desmodus taxonomic arrangement. Here I report a quantitative and qualitative osteological study of a large, geographically varied, neontological sample of Desmodus rotundus skulls that documents its range of cranial variation in order to help find the taxonomic affinity of the Cuban Desmodus fossils.
Material and Methods The primary goal of this study was to evaluate
the range of variation of Desmodus rotundus skulls to find the taxonomic affinity of the Cuban Desmodus fossil sample. Secondary goals included finding significant limits that would provide metric or morphologic discriminates between the supposed geographical subspecies, sexes, and other fossil species. This data could help recognize the species limit of the extant and extinct Desmodus groups.
The results are based on the study of cranial morphology and the statistical analysis of osteological measurements of over 70 adult specimens of Desmodus rotundus and a small fossil sample (< 10 specimens; including three incomplete skulls) from Cuban Quaternary cave deposits (see Appendix 2). Juveniles were characterized by open sutures (sutura interpalatina and sutura intermaxilari), completely undeveloped zygoma, and deciduous dentition. Skins and postcranial elements (i.e., humerus and scapula) of over 60 specimens, including Diphylla ecaudata and Diaemus youngi were additionally studied. Both sexes were equally treated except for the sex oriented statistical analyses. Museum specimens, encompassing most of D. rotundus present geographical range (Mesoamerica, South America, Tobago and Margarita islands), were examined and compared at the American Museum of Natural History (AMNH) in New York, and the Florida Museum of Natural History (FLMNH) at the University Of Florida (UF) in Gainesville (see Appendix-2). Measurements were taken with digital calipers to the nearest 0.01, and the results are expressed in millimeters (mm). Measurement methodology used is illustrated in Figure-1. Osteological terminology followed Silva Taboada (1979), Suárez (2005), and Giannini et al. (2006) see figure 2.
I did not study the entire Cuban Desmodus fossils personally. Of the four presently known skulls, I only studied one. To use as a comparative sample that could help compensate for unavailable data, the measurements reported by Koopman (1958), Jones (1958), Husson (1962), Goodwin (1969), Linares (1969-1970), Woloszyn and Mayo (1974), Morgan et al., (1988), Arroyo-Cabrales and Álvarez (1990), and Suárez (2005) were merged and analyzed statistically as a separate group (see Table-1). This additional group was subjected to descriptive and analytical statistical tests, as described below, that helped to identify differences in measuring methodology, and if there were significant difference between both samples (i.e., the one I measured directly, and the one merged from published literature).

Chiroptera Neotropical 17(1), July 2011
965
All statistical tests were performed with STATISTICA (StatSoft1995). For these I used 13 linear measurements taken on 42 complete adult skulls to illustrate the cranial morphology of D. rotundus (Fig. 1). One-way and two-way ANOVA tests with post hoc Tukey’s Tests were used to compare between groups of unequal size samples greater than 10. I tested for significant measurements between Desmodus species, and the variance of the directly measured sample and the one merged from the literature. T-tests were used to find mean differences between the geographical
populations of Desmodus subspecies (i.e., rotundus and murinus) and sexual dimorphism. Only the data from the directly measured sample was used for the T- tests. Significance levels were set at p < 0.050 on all tests. I followed Koopman (1988) and Simmons (2005) in considering the extant D. rotundus populations monotypic, even though there is molecular evidence that hint at the probability of separate geographical races (Martins 2008). These subspecific subdivisions were considered only for the geographical analysis.
Figure 1. Measurement methodology from superior, ventral, and left lateral views of Desmodus rotundus skull (AMNH261767). Measurements are represented in major case and follow the illustrated sequence. Abbreviations: AL-Con length taken from anterior canine fossa to the most posterior part of the occipital condyle; AC-LM length taken from anterior most part of the canine to the last maxillary molar; AC-PMF length taken from the anterior most part of the canine to the postglenoid process; AI-LM length taken from the anterior most part of the incisors to the posterior part of the last molar; BIW maximum width at the base of the incisors; LPW least width taken at the postorbital constriction; Pl or palatal length, length taken from anterior most part of incisors to the palatal process; MBW maximum braincase width; MCW maximum width between maxillary canines; MHB maximum height of the braincase taken from the uppermost part of the skull to the basioccipital region; MWIF maximum width taken between the infraorbital foramen; TLS total length of the skull; ZyW maximum zygomatic width taken at the laterals of the glenoid fossa.
Canonical and Principal Component Analysis
(PCI) were performed in order to accentuate or visualize significant differences between groups, and identify relationships between the groups of variables from the dataset. These tests helped project and plot relationships between groups. The Canonical analysis was based on species membership variable against all other lineal measurement variables. PCI was based only on the measurement variables, where widths were tested
against lengths. The resulting roots were plotted (Fig. 11).
A Pearson’s index of skeweness and interquartile range (IQR) tests were used to test whether the TLS measurement in the directly measured sample was normally distributed and to find the outlier parameters. This measurement was selected because of its wide Standard Distribution.
Chiroptera Neotropical 17(1), July 2011
966
Results A considerable range of individual variation
was found on the skull morphology of over 70 adult museum specimens of Desmodus rotundus regardless of their geographical origin. The most variant discrete characters are described here in detail, contrasted to the differentiating characteristics of the Cuban Desmodus sample described by previous authors.
Comparative Qualitative evaluation Upper Incisors and Canines (I-C) are blade
like and thin. The crown and shaft are indistinguishable. They range from thick in width
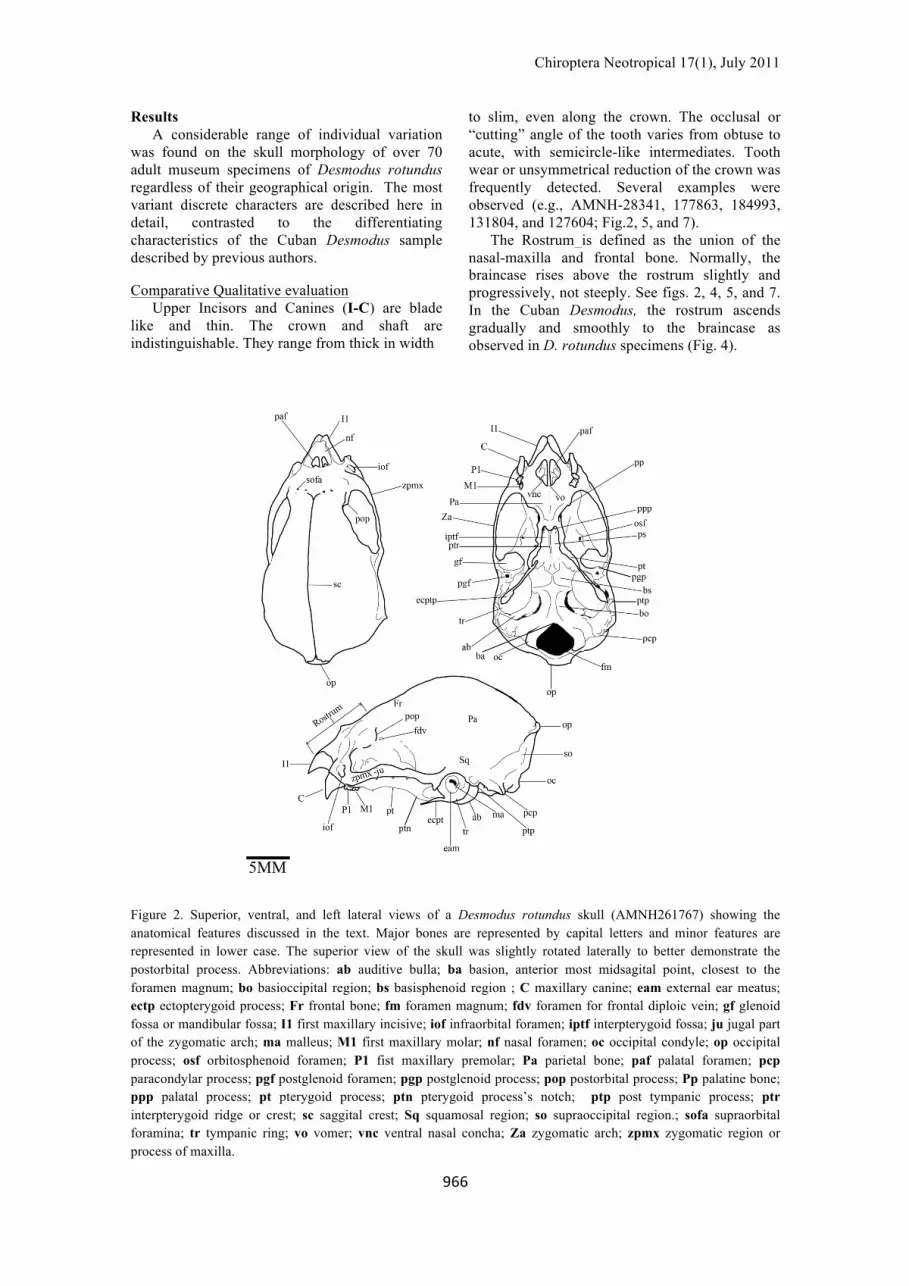
to slim, even along the crown. The occlusal or “cutting” angle of the tooth varies from obtuse to acute, with semicircle-like intermediates. Tooth wear or unsymmetrical reduction of the crown was frequently detected. Several examples were observed (e.g., AMNH-28341, 177863, 184993, 131804, and 127604; Fig.2, 5, and 7).
The Rostrum is defined as the union of the nasal-maxilla and frontal bone. Normally, the braincase rises above the rostrum slightly and progressively, not steeply. See figs. 2, 4, 5, and 7. In the Cuban Desmodus, the rostrum ascends gradually and smoothly to the braincase as observed in D. rotundus specimens (Fig. 4).
Figure 2. Superior, ventral, and left lateral views of a Desmodus rotundus skull (AMNH261767) showing the anatomical features discussed in the text. Major bones are represented by capital letters and minor features are represented in lower case. The superior view of the skull was slightly rotated laterally to better demonstrate the postorbital process. Abbreviations: ab auditive bulla; ba basion, anterior most midsagital point, closest to the foramen magnum; bo basioccipital region; bs basisphenoid region ; C maxillary canine; eam external ear meatus; ectp ectopterygoid process; Fr frontal bone; fm foramen magnum; fdv foramen for frontal diploic vein; gf glenoid fossa or mandibular fossa; I1 first maxillary incisive; iof infraorbital foramen; iptf interpterygoid fossa; ju jugal part of the zygomatic arch; ma malleus; M1 first maxillary molar; nf nasal foramen; oc occipital condyle; op occipital process; osf orbitosphenoid foramen; P1 fist maxillary premolar; Pa parietal bone; paf palatal foramen; pcp paracondylar process; pgf postglenoid foramen; pgp postglenoid process; pop postorbital process; Pp palatine bone; ppp palatal process; pt pterygoid process; ptn pterygoid process’s notch; ptp post tympanic process; ptr interpterygoid ridge or crest; sc saggital crest; Sq squamosal region; so supraoccipital region.; sofa supraorbital foramina; tr tympanic ring; vo vomer; vnc ventral nasal concha; Za zygomatic arch; zpmx zygomatic region or process of maxilla.

Chiroptera Neotropical 17(1), July 2011
967
Figure 3. Left lateral side views of the Zygomatic arch in Desmodus rotundus. A. AMNH131808, B. AMNH228226, C. AMNH127600, D. AMNH68568, E. AMNH183304, F. AMNH76087, G. AMNH248185, H. AMNH127604, I. AMNH71688, J. AMNH76087, K. AMNH76084, L. AMNH148659, M. AMNH2327, N. JRC600 represents a new D. rotundus specimen from Cuba (Orihuela, in Ed.) and O. Zygomatic arch of D. puntajudensis Holotype (IGACC7999/401) drawn from photographs in Woloszyn and Mayo (1974). Note the variability in depth, length, and shape of the zygomatic arch.
Figure 4. Skulls of three of the four Desmodus fossil specimens known from Cuba. Superior, left lateral, and inferior views of MNHNCu 76.4713 from Cueva de Paredones, Havana (left); IG ACC 7999/401 from Punta Judas, Sanctí Spirítus (middle), and JRC600 from Cueva de los Nesophontes, Palenque, Habana-Matanzas (right). These specimens were drawn from fotos in Woloszyn and Mayo (1974), Silva Taboada (1979), and Suárez (2005).
The Infraorbital foramen (iof) is regularly laterally compressed and thus not widely open. Woloszyn and Mayo (1974) and Suárez (2005) did not use this character. I found the Cuban specimens indistinguishable from D. rotundus.
Postorbital constriction: In D. puntajudensis, this structure was described as being wider and more excavated than in D. rotundus. I found that the lateral sides of the postorbital region range from restricted to widely parallel in D. rotundus.

Chiroptera Neotropical 17(1), July 2011
968
This makes the supraorbitals look inflated depending on the degree of constriction; the more constricted the postorbital region, the more inflated the supraorbitals appears. A postorbital process was observed in specimens with very prominent supraorbitals. This feature is, however, usually imperceptible in D. rotundus (Fig. 4-6). The postorbital foramina vary in size, and were present on all the specimens analyzed.
Nasal foramen (nf): This foramen is relatively uniform in shape. The shapes vary from ovoid to heart shaped. In the Cuban Desmodus, the nasals were described as depressed, although, this was also frequently observed in D. rotundus (Fig.4 and 5).
Zygomatic arch (=Zygoma, Za and zpmx): Woloszyn and Mayo (1974) used this rostrum bone to distinguish D. puntajudensis from D. rotundus. The endemic subspecies was described to differ from the last because its jugal arch was higher in his anterior portion. Contrastingly, I found that the zygomatic arch in D. rotundus showed a marked individual variation, both in morphology and size. It is usually found highest and thickest at the jugal-squamosal union, although, both portions of the zygomatic showed thickness variants. In some adult specimens, the zygoma was incomplete or underdeveloped (Fig. 3, 4, and 6). In juveniles, this feature is incomplete at both sides of the zygoma, not as seen in AMNH127600 (Fig.3 C).
Sagittal crest (sc): This is always present and visible, seldom very pronounced regardless of specimen size. It is usually well defined, as described for the Cuban Desmodus.
Braincase Shape: The shape of the braincase varies from globosely or bulbous to ovoid and rhomboid in outline. I found it globosely and ovoid in D. rotundus as was described for D. puntajudensis (Fig. 4-6). The height of the braincase elevates gradually, seen in a lateral aspect. However, specimens with superiorly bulging were also observed. This coronal uprising modifies the superior calvarium.
Pterygoid process and palatal processes (pt-pp): The pterygoid in D. puntajudensis was described as being small and laterally oriented. In D. rotundus it varied from internally oriented to open and laterally oriented as described for the Cuban Desmodus, although, this bone is typically found laterally oriented. (e.g., see AMNH 261767 in Fig.4).
The pterygoid process varied also in shape and in concavity of its fossae (iptf), ranging from
parallel to slightly convergent. The posterior of the pterygoid process’s notch (ptn) was prominent and hook-like with less frequent round or blunt ends. The inter-pterygoid crest (ptr) was always pronounced. The post-palatal process (ppp) or notch on the posterior part of the palatal varies from well developed and sharp to small and almost undetectable. The palatal notches (pp), on the poster-lateral sides of the palatine bone, are also very variable. Although this notch is typically prominently present, it was absent in a couple of specimens (not shown).
Palatine bone and Palatal foramina (Pa-paf): The general shape of the palatal bone is semi-oval. The palatal is wide and grooved shallowly in the midline. The palatal foramens are half-oval in outline. Although there are slight variations, these features are frequently as described above. The sutura intermaxilary was found visible in juvenile specimens only. In the Cuban Desmodus, the palatine bone was defined as being wide and concave, with no defined suture, diagnosis that matches the variation observed in D. rotundus (Fig. 4).
Glenoid fossa or mandibular fossa (gf): In the Cuban Desmodus, this feature was described to be flat and in a different plane. I did not find any “different plane” shift and no anatomical inclination in any of the published figures of D .puntajudensis. Nonetheless, in D. rotundus, this trait was found consistently round, flat or shallow, always oriented poster-laterally, and slightly inclined anteriorly.
Postglenoid process and foramen (pgf): This process was always present and visible in D. rotundus, varying between straight down or arched interiorly. The foramen was found in the posterior face of this process and was always visible. No differences were found in the Cuban fossil sample.
Basiosphenoid and basioccipital region (bs-bo): In Woloszyn and Mayo’s paper (1974), this area is named the basion (ba).These two regions unite at the synchondrosis interspehnoidalis, which is fussed in adults, and open in juvenile specimens. This forms an inconspicuous and gradual connection, usually concave. These pits are well excavated with a small distinct crest dividing the basiosphenoid fossae. These are shallow, but in specimens with a concave basioccipital, the basioesphinoid fossae seem deeper (Fig.3-6). Different morphology between the neontological and the Cuban fossil specimens was found.

Chiroptera Neotropical 17(1), July 2011
969
Figure 5. Desmodus skulls from AMNH with fossil specimens from Cuba. Superior A, inferior B, and left lateral views C. The last three are Cuban fossil specimens. The last specimen is JRC600. Note the variation in occipital process, paracondylar process, nasals, and canine angles.
Figure 6. Superior view of Desmodus skulls from the AMNH, FLMNH, and the specimens presently known from Cuba (on the far right). Note the variation in shape and size of the occipital process, braincase, and postorbital width. Original photograph is a courtesy of Dr. Adrian Tejedor (AMNH). Note: most of the AMNH specimens in this composite are not included in appendix 2.

Chiroptera Neotropical 17(1), July 2011
970
Figure 7. Left lateral side view of Desmodus rotundus skulls from AMNH. Some skulls have been rotated. Note the variation in the rostrum, zygomatic arch, and canine morphology.
Paracondylar process (pcp): This process varies from very prominent antero-posteriorly or laterally compressed with slight posterior inclination. The transition between this process and the postynpanic process (with the skull seen laterally) is usually gradual and not deeply concave. In the D. puntajudensis figures (as illustrated by Woloszyn and Mayo (1974) and Suárez (2005)), this transition falls within the observed range of D. rotundus by being gradual and not abrupt.
Postympanic process (ptp) is not prominent, and thus uniform. It is flattened antero-posteriorly. Usually ventrally straight, or slightly arched anterior-ventrally. No difference was noted between D. rotundus and the Cuban specimens.
Occipital condyle (oc) is usually round and visible laterally, projecting postero- ventrally.
The condyles vary in postero-ventral inclination, and antero-posterior width when seen laterally. No significant variation was noted in the Cuban sample.
Occipital process or occiput on supraoccipital region (op): This is another diagnostic characteristic previously used for the Cuban fossils. D. puntajudensis seemed to differ from D. rotundus in being “more” posteriorly projected; with supra occipitals greatly inflated along midline and occiput high and narrow, facing postero-ventrally. In this analysis, however, these features were observed within variation in D. rotundus. The occipital process varied from inflated posteriorly to ventrally straight. The more abrupt the process and protuberance seems, the deeper the depressions. The occiput or occipital notch can be prominent, small, or absent, sometimes ventrally inclined or horizontal and varying in shape from sharp to square (Fig. 4-7).
Foramen magnum (fm): The projection of the foramen magnum and its shape varies without relationship. The foramen magnum could be circular, semicircular, or ovoid outline. In general, projection varies slightly from ventrally inclined to caudal inclined. Woloszyn and Mayo (1974), and Suárez (2005) used this feature diagnostically, where the Cuban Desmodus was described to have the foramen magnum wider anteroposteriorly. I found no appreciable deviation from D. rotundus (Fig.4). This area also includes the occipital condyles and the notch for the rotation of the axis’ odontoid process. In some specimens, this notch is completely rounded-off or absent. In others the odontoid’s notch is noticeable.
Statistical
I found significant metrical variation between the sample I measured directly and the one merged from the literature. These differences were based on skull height (MHB) and anterior canine to occipital bone length (AL-Con). The Cuban Desmodus specimens were found to be slightly different from Desmodus rotundus only on two variables: total length of the skull (TLS) and height of the skull (MHB) as shown on Fig. 10. Reasons for this variation may be due to differences in measuring techniques between authors and approximated measurements as explained below. Sexual dimorphism was slightly supported statistically (p > 0.050) regardless of geographical origin.
Females were larger than males in total length of skull (TLS) and zygomatic width (ZyW), Fig. 9. These results further support the findings of Ralls (1976).

Chiroptera Neotropical 17(1), July 2011
971
Figure 8. Graphs showing significant variation in Desmodus rotundus geographical populations of Mesoamerica and South America and difference between samples. Variation in Canine to Occipital Length (AC-Con) between the author’s measured sample (A) and the merged sample from the literature (B). Variation in Height of the Skull length (MHS) among the author’s measured sample (A) an the merged sample from the literature (B). These significant differences might be due to differences in the measuring techniques as explained on text. Geographical variation found in Total Skull Length (TSL), and Total Dental Length (AI-LM) measurements: D. rotundus murinus (a) from Mesoamerica and D. rotundus rotundus (b) from South America.
Figure 9. Sexual variation found in directly measured Desmodus rotundus sample: Total Skull Length (TSL) and Zygomatic Width (ZyW) measurements between Female (F) and Male (M) specimens. See Table 1.
Contrary to expectations, minor geographical variation was detected between Mesoamerican and South American specimens. South American specimens were larger than the rest of Middle American specimens. However, morphological evidence (including an analysis of skins) that could support a taxonomic division between these two continuous populations of Desmodus rotundus
was not found. Such variations were detected in total length of skull (TLS) and total dental length (AI-LM) (Fig. 10). Martins (2008) reported similar morphological results. His morphological results, however, did not match the molecular data. Martins’ molecular results indicated that the South American populations could be separated taxonomically. (op. cit. pg.75).

Chiroptera Neotropical 17(1), July 2011
972
Table 1. Skull measurements of Desmodus rotundus from Cuban fossil specimens, museum collections, and those merged from literature (in mm). Sequence: N=number of specimens, M±SD= Mean ± Standard Deviation, and Range. *Measurements taken from Koopman (1958), Woloszyn and Mayo (1974), and Suárez (2005), and thr new specimen, JRC666, reported in Orihuela, (in press). **Specimens measured at AMNH and FLMNH. See Appendix 1, and Materials and Methods. ***Measurements taken from published literature and analyzed statistically as a group: Jones (1958), Koopman (1958), Husson (1962), Goodwin (1969), Linares (1969), Morgan et al., (1988), Arroyo-Cabrales and Alvarez (1990), and Suárez (2005).
Figure 10. Significant variation between Desmodus fossil and extant species in the following measurements: Total Skull Length (TLS), Height of the Skull (MHS), and Cerebral Width (MBW). Key: Desmodus rotundus (D. r.), Desmodus stocki (D. s.), Desmodus draculae (D. d.), Desmodus archaeodaptes (D. a.), and Desmodus from the Cuban Sample (D. p.). Specimen (JRC600) is included under the Cuban Sample, which in this case is represented as the maximum on this graph (12.06 mm). The differences on height of the skull between the Cuban Sample and the mainland sample (D.r.) might be due to differences in measuring techniques (see text).
Cuban fossil Desmodus specimens* Desmodus rotundus** Desmodus rotundus***
JRC666 IGACC7999/401
MNHNCu76.4713
MCZ4737 N M ±SD Range N M±SD Range
TLS 25.37 22.34 42 24.8 ±0.7 (23.32-26.93) 99 24.38 ±0.7 (22.7-25.6)
AI-LM 6.06 41 6.18 ±0.2 (5.67-6.58) 6 6.25 ±0.6 (5.2-6.86)
AC-LM 3.03 3.11 42 3.36 ±0.2 (2.97-3.68) 41 3.5 ±0.2 (3.15-4.0)
AC-PMF
11.67 41 12.07 ±0.5 (11.25-14.29)
PL 9.87 41 9.63 ±0.4 (8.86-10.48) 7 9.44 ±0.4 (9.0-9.81)
BIW 4.56 3.9 41 3.93 ±0.2 (3.57-4.56)
MCW 6.14 6.5 6.65 41 6.07 ±0.3 (5.55-6.78) 30 5.96 ±0.5 (5.18-6.75)
ZyW 11.83 12.44 42 11.9 ±0.4 (10.62-13.18) 104 12.10 ±0.5 (11.0-14.0)
LPW 5.51 5.1 5.2 5.1 42 5.49 ±0.3 (4.72-6.08) 115 5.49 ±0.3 (4.7-6.0)
MHS 12.06 11.4 11.7 41 13.07 ±0.5 (11.74-13.98)
MBW 12.46 12.0 12.1 41 12.09 ±0.3 (11.46-12.70) 70 12.09 ±0.4 (11.3-13.0)
MWIF 7.31 6.0 6.2 41 6.92 ±0.2 (6.23-7.59) 48 6.28 ±0.5 (5.4-7.1)
AC-Con 22.64 20.31 41 22.4±07 (20.69-24.05) 18 20.7 ±1.0 (18.78-22.43)

Chiroptera Neotropical 17(1), July 2011
973
The sample I measured directly showed mild
degrees of individual variation. The range of Standard Deviation varied from 0.06 to one SD indicating a small degree of spread in the data, which is supported by small Standard Error: 0.02-0.1. The most variant measurements were total length of skull (TLS) and anterior canine fossa to occipital condyle length (AC-Con.). The rest of the skull measurements maintained a low degree of Variation values of less than 0.5 points.
Pearson’s index of skeweness test indicated (PI= -0.00228) that the normal distribution of skull lengths is slightly skewed to the left of the distribution. The IQR test indicated an outlier range for (TSL) of 22.74 to 27.06 mm. These last two quantitative tests demonstrate that there are no outliers in total skull length in my directly measured sample, and that its distribution is most probably normal.
Figure 11. Canonical analysis plot-graph based on the first two roots of measurement variation in Desmodus. Desmodus rotundus murinus (■), D. rotundus rotundus (○), Cuban fossil sample (triangle), and Desmodus stocki (●). Analysis source: Species versus Total Skull Length (TLS). Discussion Quantitative aspects
The measurement comparisons between the Cuban Desmodus sample and the merged measurement sample are biased due to differences in measuring techniques and the use of approximated measurements. The total length of the skull given in Woloszyn and Mayo (1974) for D. r. puntajudensis (Holotype IGACCI7999/401) does not represent the real length of that skull because the skull is incomplete. This created an outlier in the statistical analyses and PCI (lowest IQR = 22.74mm). However, a similar measurement such as the length taken from the anterior canine alveoli to the occipital condyles (AC-Con) were compared (see Woloszyn and Mayo, 1974: pg. 255-257), the Cuban Desmodus fell within the size range of Desmodus rotundus. This indicates that, the outlier skull is not as small as it appears (Table-1, and Fig. 4).
The height of the skull (MHS) was another biased measurement. Woloszyn and Mayo (1974) probably measured their skull height from the innermost part of the basioesphenoidal-basioccipital region, while I measured it from outermost (Fig.1). Note that there are no (MHS) measurements from the merged sample from the literature. The general cranial traits previously described of “equal rank as those used to describe
Desmodus stocki and Desmodus draculae” (Suárez, 2005), are essentially and statistically insignificant and not morphologically supported by discrete characteristics.
Qualitative Aspects Originally, Woloszyn and Mayo (1974) stated
that morphological differences separating the Punta Judas specimen from recent populations was only of sub-specific nature and that its location within Desmodus rotundus was “beyond doubt” (pg. 260). Unfortunately, previous authors employed small and geographically restricted samples, usually comprised of less than 10 specimens from Mexico (in Woloszyn and Mayo 1974) or a Mexican sample of unknown number in Suárez (2005) to compare the Cuban fossil specimens.
Furthermore, discriminating characters that could justify a sub-specific division or distinguish the Middle American D. rotundus (“murinus”) from the South American D. rotundus (“rotundus”) morphologically where not observed. Minor geographical variation, however, was supported statistically, where southern specimens were slightly larger than the Central American ones (Fig. 8). This evidence, in part, supports Koopman’s (1988) and Simmons’ (2005)
-3
-2
-1
0
1
2
3
4
5
-4 -2 0 2 4 6 8

Chiroptera Neotropical 17(1), July 2011
974
decision in not delimiting any subspecies within D. rotundus.
Conclusions This study suggests that the fossil Desmodus
from Cuba fall within the range of variation observed in D. rotundus, and therefore should be synonymies. I found that the morphological characters previously used to define the Cuban fossil vampire bat specimens, currently known as D. puntajudensis, readily fall within the individual variation range observed in the extant vampire bat D. rotundus. This study suggests that the Cuban specimen should be systematically classified as Desmodus rotundus and not as a separate taxon. These results are important because the understanding of taxonomic limits within the Cuban fossils Desmodus could help clarify their paleobiogeography, diversity, and extinction. Acknowledgements
I thank my friend and colleague Adrian Tejedor for all the support and guidance, and for revising earlier drafts of this paper. For providing access to museum collections, I thank Chris Norris, Susan Bell, Judy Galkin, all at the AMNH, New York, and Richard Hulbert at the FLMNH, University of Florida. For always answering my questions and emails, I thank Gilberto Silva Taboada, William Suárez, and Osvaldo Jiménez at the National Museum of Natural History in Cuba, and the Gabinete de Arqueología, Havana, Cuba. Judy Chupasko and Bill Amaral at Museum of Comparative Zoology, Cambridge, for providing valuable information on their Cuban fossil specimen. For orientation, site logistics, and unlimited encouragement: Leonel Pérez Orozco, Amarilis León, Ádrian Tejedor, Tamara Castaño, Candido “Pachi” Santana, and Ricardo Viera Muñoz. For providing editorial revisions, I thank Adrian Tejedor, Tamara Castaño, Gary S. Morgan, Nicholas Czaplewski, the editorial staff at Chiroptera Neotropical, and two anonymous reviewers.
References Arredondo O. 1958. El vampiro cubano. Scout
10:6-7. Arroyo Cabrales J. & Ray C.E. 1997. Revisión de
los vampiros fósiles (Chiroptera: Phyllostomidae, Desmodontinae) de México. In Homenaje al Profesor Ticul Álvarez. Ed. J. Arroyo Cabrales, and O.J. Polaco, 69-86. Colección Científica, Instituto Nacional de Antropología e Historian, México, 357:1-391.
Arroyo-Cabrales J. & Álvarez T. 1990. Restos óseos de murciélagos procedentes de las excavaciones de las grutas Loltun. Colección Científica, Instituto Nacional de Antropología e
Historian, México, Distrito Federal, México, 103 pp.
Czaplewski N.J. & Cartelle C. 1998. Pleistocene Bats from Cave deposits in Bahia, Brazil. Journal of Mammalogy 79: 784-803.
Czaplewski N.J.; Krejca J. & Miller T.E. 2003. [A] Late Quaternary bats from Cebada Cave, Chiquibul cave Sistem, Belize. Caribbean Journal of Science 39:23-33.
Czaplewski N.J. & Peachey W.D. 2003. [B] Late Pleistocene bats from Arkenstone Cave, Arizona.The southwestern Naturalist 4(48): 593-609.
Fischer K. 1977. Quartare mikromammalia Cubas, vorwieged aus der Hohle San José de las Lamas, Santa Fé, provinz Habana. Zeitschrift für Geologische Wissenschaften. Berlin 5(2):213-255.
Giannini N.P.; Wible J.R. & Simmons N.B. 2006. On the cranial osteology of Chiroptera I. Pteropus (Megachiroptera: Pteropodidae). Bulletin of American Museum of Natural History 295: 134pp.
Greenhall A.M.; Joerman G.; Schmidt U. & Seidel M.R. 1983. Desmodus rotundus. Mammalian Species 202:1-6.
Greenhall A.M. & Schmidt U.1988. Natural History of Vamire Bats. CRC Press, Inc. Boca Raton, Florida. 240pp.
Goodwin G.G. 1969. Mammals from the state of Oaxaca, Mexico, in the American Museum ofNatural History. Bulletin of American Museum of Natural History141. Article 1.
Gut H.J. 1959. A Pleistocene vampire bat from Florida. Journal of Mammalogy 40: 534-538.
Husson A.M. 1962. Bats of Surinam. Zoo. Verhandel, Leiden 58:1-282.
Hutchison J.H. 1967. A Pleistocene vampire bat (Desmodus stocki) from Potter Creek Cave, Shasta County, California. Paleobios 3:1-6.
Jiménez O.; Condis M.M. & E. García. 2005. Vertebrados post-glaciales en un residuario fósil de Tyto alba scopoli (Aves: Tytonidae) en el occidente de Cuba. Revista Mexicana de Mastozoología, 9:84-111.
Jones J.K., Jr. 1958. Pleistocene bats from San Joseito Cave, Nuevo León, México. Univ. Kansas Publ. Mus. Nat. Hist. 9:389-396.
Koopman K.F. 1958. A fossil vampire bat from Cuba. Breviora 90:1-4.
Koopman K.F. 1988. Systematics and Distribution. Chap. 2. In Greenhall, A.M., and U. Schmidt (Eds). Natural History of Vamire Bats. CRC Press, Inc. Boca Raton, Florida. 240 pp.
Linares O.J. 1969. Quirópteros subfosiles encontrados en las cuevas venezolanas. Parte I. Boletín de la Sociedad Venezolana de Espeleología 1(2):119-145.

Chiroptera Neotropical 17(1), July 2011
975
Linares O.J. 1970- Parte III. Desmodus rotundus en la Cueva de la Brújula (Mi. 1) Miranda. Boletín de la Sociedad Venezolana de Espeleología 3:33-36.
Martins F.M. 2008. Filogeografia intraespecifica do morcego hemátofago Desmodus rotundus (Chiroptera, Phyllostomidae). Inedited Thesis.
Morgan G.S. 1991. Neotropical Chiroptera from the Pliocene and Pleistocene of Florida. Bulletin of American Museum of Natural History 206: 176-213.
Morgan G.S. 2001. Patterns of extinction in the West Indian Bats. In C.A. Woods and F.E. Sergile (Editors), Biogeography of the West Indies: Patterns And Perspectives: 369-406. Boca Raton, Florida: CRC press.
Morgan G.S.; Linares O.J. & Ray C.E. 1988. New species of vampire bats (Mammalia: Chiroptera: Desmodontinae) from Florida and Venezuela. Proceedings of Biological Society of Washington 101:912-928.
Orihuela J. In Press. Late Holocene fauna from a cave deposits in western Cuba: post-Columbian occurrence of the vampire bat Desmodus rotundus (Phyllostomidae: Desmodontinae). Caribbean Journal of Science 104.
Ralls K. 1976. Mammals in which females are larger than males. The Quarterly Review of Biology 51(2): 245-276.
Ray C.E.; Linares O.J. & Morgan G.S. 1988. Paleontology. Chapter 3, pp 19-30 in Greenhall, A.M., and U. Schmidt.eds.1988. Natural History of Vamire Bats. CRC Press, Inc. Boca Raton, Florida. 240pp.
Silva-Taboada G. 1979. Los murciélagos de Cuba. Editorial Academia, La Habana. 424pp.
Simmons N.B. 2005. Order Chiroptera. In D.E. Wilson and D.M. Reeder (Editors), Mammal species of the world: A Taxonomic and Geographic reference, 3rd. ed. Washington D.C. Smithsonian Institution Press.
Suárez W. 2005. Taxonomic Status of the Cuban Vampire Bat (Chiroptera: Phyllostomidae: Desmodontinae: Desmodus). Caribbean Journal of Science 41 (4): 761-767.
Varona L. 1974. Catalogo de los mamíferos vivientes y extinguidos de las Antillas. Academia de Ciencias de Cuba, La Habana. 139pp.
Woloszyn B.W. & Mayo N.A. 1974. Postglacial remains of a vampire bat (Chiroptera: Desmodus) from Cuba. Acta Zoologica Cracoviensia 19:253-265.
Appendix 1
Desmodus rotundus specimens studied from the neontological mammal collections of the Florida Museum of Natural History, University of Florida (UF), Gainesville, Florida, and American Museum of Natural History (AMNH) in New York. Total number of specimens directly studied from museum collections: 82.
Desmodus rotundus specimens examined at Florida Museum of Natural History (FLMNH),
University of Florida (UF). Oldest specimen examined was collected on July 4, 1922. Total examined: (7). MEXICO-Querétaro, Cadereyta, UF5988 (Adult Female); No locality: UF2612 (Juvenile-unknown sex) and UF11964 (Juvenile-unknown sex). VENEZUELA-Cojedes, Finca “La Coromoto”: UF13327 (Adult Male); Guárico, 40km South of Calabozo: UF16270 (Adult Male). PERU-Ambo: UF24847 (Adult Female). PARAGUAY-Chaco: UF20664 (Adult Female).
Desmodus rotundus specimens examined at American Museum of Natural History (AMNH). Oldest specimen examined was collected on May 6, 1906. Total specimens examined (skeleton and skin): 70. MEXICO-Chiapas, Pueblo Nuevo: AMNH203768 (Adult Female), AMNH172299 (Adult Female), Ocozocociultc: AMNH186900 (Adult Male); Veracruz: AMNH237373 (Adult Male); Rio Jaltepec: AMNH172296 (Adult Female). GUATEMALA-Puebla: AMNH68568 (Adult Male); Santa Rosa: AMNH235321 (Adult Male), AMNH235521 (Adult Male); Coban: AMNH174305 (Juvenile Female), AMNH174303 (Juvenile Male?), AMNH174305 (Juvenile Male); “Desmodus rufus” AMNH11298/9562. HONDURAS-La Flor: AMNH129754 (Adult Male), AMNH127604 (Adult Female), AMNH127599 (Adult Male); Archaga, La Cueva: AMNH228226 (Adult Male), Archaga, La Flor: AMNH127600 (Adult Male), AMNH127598 (Adult Male). EL SALVADOR- Coatepeque: AMNH177863 (Adult Male). NICARAGUA-Vol. de Chinandega: AMNH28341 (Adult Female), AMNH28340 (Adult Male). COSTA RICA-Turrialba: AMNH131807 (Adult Male), AMNH13808 (Adult Male); AMNH131806, AMNH131809.

Chiroptera Neotropical 17(1), July 2011
976
PANAMA- Rio Chaqres: AMNH184993 (Adult Male). ECUADOR-Boca: AMNH71688 (Adult Female), AMNH71689 (Adult Male). PERU- Depto. Junín, Tarma: AMNH230736 (Adult Male); Boca, Rio Urubamba: AMNH76084 (Adult Female), AMNH76087 (Adult Male). COLOMBIA-Boquerón: AMNH18728 (Adult Male). VENEZUELA-Isla de Margarita, Nova Esparta, Cueva de Quintín: AMNH uncataloged- Collected by Adrian Tejedor on May 19, 2004 on cave floor. FRENCH GUIANA-Paracou: AMNH267211 (Adult Female), AMNH267504 (Adult Female). BRAZIL-Costa Marques: AMNH209750 (Adult Male); Para: AMNH202327 (Adult Male); Catas Altas, Minas Gerais, uncataloged (Adult Male) collected by Adrian Tejedor on August 13, 2005. BOLIVIA-El Beni: AMNH261767 (Adult Female), AMNH215031 (Adult Female), AMNH215030 (Adult Female), AMNH910960 (Adult Male), AMNH210961 (Adult Male), AMNH248190 (Adult Female), AMNH248191 (Adult Female), AMNH248185 (Adult unknown sex), AMNH209748 (Adult Male), AMNH209755 (Adult Male), AMNH209749 (Adult Male), AMNH209735 (No sex), AMNH209758 (Adult Male), AMNH210957 (Adult Male), AMNH209758 (Adult Male), AMNH209736 (Adult Male), AMNH248181 (No sex), AMNH248182 (Adult Male), AMNH215031 (Adult Female), AMNH210961 (Adult Male); “Concepcioncita” Cave, Motacuzito, Santa Cruz, Adult Male not numbered collected by Dr. Adrian Tejedor on July 4, 2005; Provincia Caballero, “capitol” “ El Tunel” AMNH264904 (Adult Male). PARAGUAY-Boquerón, Pozo: AMNH234315 (Adult Male), AMNH234313 (Adult Male), AMNH234314 (Adult Female); Villarrica: AMNH148656 (Adult Female). ARGENTINA-St.Barbara: AMNH183304 (Adult Female); Laguna “La Brea” 25km West of Palma Sola: AMNH256983.
Diphylla ecaudata and Diaemus youngi specimens studied at AMNH. Total specimens examined: 5. MEXICO-Chiapas: AMNH203837 (Adult Male), AMNH203843 (Adult Female). TRINIDAD-Waller: AMNH A. M. field number 369 (Adult Male), AMNH 257104 (Adult Female). VENEZUELA-Ocampo: AMNH78285 (Adult Female). Oldest specimen was collected on 3April 1929.
There are four Desmodus fossil skulls known from the Cuban fossil record. These fossil specimens are deposited at Museo Nacional de Historia Natural (MNHNCu), the zoological collection of the Academia de Ciencias (CZACC), and Museum of Comparative Zoology at Harvard, Cambridge. Three of these, except JRC specimens, were examined only through published photographs and measurements (see Materials and Methods). The specimens in the JRC collection, however, were examined personally. CUBA- Habana, Caimito, Cueva de Paredones: MNHNCu76.4713, CZACC1.5546, CZACC1.5543, CZACC1.5547, CZACC1.5545, CZACC1.5544; Villa Clara, Punta Judas, Cueva Centenario de Lenin: IGACC7999/401(Desmodus rotundus puntajudensis Holotype, now uncataloged at CZACC); Matanzas-Habana, Loma del Palenque, Cueva de los Nesophontes: JRC600 (Skull) and JRC2389 (humeri) presently deposited at MNHN.







![WARTHRONE APPROVED ARMIES [WFB]VAMPIRE COUNTS](https://img.pdfslide.net/doc/110x75/63254c255c2c3bbfa8031ff1/warthrone-approved-armies-wfbvampire-counts.jpg)





















