Embed Size (px)
Citation preview

The Plant Cell, Vol. 13, 999–1010, May 2001, www.plantcell.org © 2001 American Society of Plant Physiologists
RESEARCH ARTICLE
slender
Rice, a Constitutive Gibberellin Response Mutant, Is Caused by a Null Mutation of the
SLR1
Gene, an Ortholog of the Height-Regulating Gene
GAI/RGA/RHT/D8
Akira Ikeda,
a,b,1
Miyako Ueguchi-Tanaka,
a
Yutaka Sonoda,
a,1
Hidemi Kitano,
a
Masaji Koshioka,
c
Yuzo Futsuhara,
b
Makoto Matsuoka,
a
and Junji Yamaguchi
a,1,2
a
BioScience Center and Graduate School of Bioagricultural Sciences, Nagoya University, Chikusa-ku, Nagoya 464-8601, Japan
b
Department of Agriculture, Meijo University, Tenpaku, Nagoya 468-8503, Japan
c
National Research Institute of Vegetables, Ornamental Plants, and Tea, 60 Kusawa, Ano, Age, Mie 514-2328, Japan
The rice
slender
mutant (
slr1-1
) is caused by a single recessive mutation and results in a constitutive gibberellin (GA)response phenotype. The mutant elongates as if saturated with GAs. In this mutant, (1) elongation was unaffected by
an inhibitor of GA biosynthesis, (2) GA-inducible
a
-amylase was produced by the aleurone layers without gibberellicacid application, and (3) endogenous GA content was lower than in the wild-type plant. These results indicate that theproduct of the
SLR1
gene is an intermediate of the GA signal transduction pathway.
SLR1
maps to
OsGAI
in rice andhas significant homology with height-regulating genes, such as
RHT-1Da
in wheat,
D8
in maize, and
GAI
and
RGA
in Ar-abidopsis. The
GAI
gene family is likely to encode transcriptional factors belonging to the
GRAS
gene superfamily. DNAsequence analysis revealed that the
slr1-1
mutation is a single basepair deletion of the nuclear localization signal do-main, resulting in a frameshift mutation that abolishes protein production. Furthermore, introduction of a 6-kb genomicDNA fragment containing the wild-type
SLR1
gene into the
slr1-1
mutant restored GA sensitivity to normal. These re-sults indicate that the
slr1-1
mutant is caused by a loss-of-function mutation of the
SLR1
gene, which is an ortholog of
GAI
,
RGA
,
RHT
, and
D8
. We also succeeded in producing GA-insensitive dwarf rice by transforming wild-type rice witha modified
SLR1
gene construct that has a 17–amino acid deletion affecting the DELLA region. Thus, we demonstrateopposite GA response phenotypes depending on the type of mutations in
SLR1
.
INTRODUCTION
Gibberellins (GAs) have an important role in the regulation ofmany physiologic processes in the growth and developmentof plants, such as seed germination, shoot/stem elongation,and flower development. Changes in both GA concentrationand tissue sensitivity to GA influence these processes. Themolecular mechanisms by which the GA signal is trans-duced into morphologic and biochemical changes in plants,however, are largely unknown.
Many studies on the detection of and response to GA arefocused on identifying the mutants that affect these pro-cesses. GA response mutants isolated from various plantspecies fall into two phenotypic categories: elongated
slen-
der
-type mutants and GA-unresponsive dwarf mutants(Hooley, 1994; Swain and Olszewski, 1996). The
slender
mutants have constitutive activation of their GA response,and the dwarf mutants are deficient in GA detection or sig-nal transduction. The
slender
mutant of barley, which is ho-mozygous for the recessive
sln1
alleles, is characterized bya rapid growth rate and long leaf sheaths (Foster, 1977).
slender
barley does not respond to growth retardants suchas ancymidol (Lanahan and Ho, 1988) and paclobutrazol(Croker et al., 1990). During germination, aleurone layers of
slender
barley synthesize
a
-amylase in the absence of ex-ogenous GA (Lanahan and Ho, 1988). The mutant contains asmaller amount of active GAs compared with wild-typeplants, indicating that GAs are not involved in the growth of
slender
barley. Thus, the
slender
mutation might be impor-tant for elucidating the GA signal transduction pathway.
In rice, many dwarf mutants have been characterized andclassified as dwarf and GA deficient, dwarf and GA insensi-tive, or dwarf due to other reasons (Mitsunaga et al., 1994).
1
Current address: Division of Biological Sciences, Graduate School ofScience, Hokkaido University, N10, W8, Sapporo 060-0810, Japan.
2
To whom correspondence should be addressed. E-mail [email protected]; fax 81-11-706-2737.

1000 The Plant Cell
Figure 1. Phenotype of slender Rice and Its Original Wild Type, Nipponbare.
(A) slender rice was isolated as a tall mutant from rice cv Nipponbare. The plants homozygous for the recessive slender (slr1-1) gene segregatedat a 1:3 ratio in 2-week-old plants.(B) Shoot elongation of wild type treated with (middle) or without (left) 10 mM GA3 and slender without GA3 treatment (right). The photograph wastaken 1 week after germination.(C) An elongated internode of a 2-week-old slender plant. The arrowhead indicates the third internode.(D) Root morphology of 1-week-old plants: wild type (left), slender (middle), and wild type treated with 10 mM GA3 (right).

SLR1
as a Rice GA Response Modulator 1001
A
slender
-type mutant has not been reported in rice, how-ever, except for the
awaodori
(
ao-1
) mutant (Nakamura,1992).
Several GA signaling intermediates have been identifiedby mutation analyses in a number of plant species (Taylor,1998), and some corresponding genes were recentlycloned. In Arabidopsis,
SPINDLY
(
SPY
) acts as a negativeregulator of the GA response. The deduced amino acid se-quence of SPY suggests that the protein is an
O
-linked
N
-acetylglucosamine transferase that might glycosylateother molecules involved in GA signaling (Thornton et al.,1999). A second GA signaling intermediate from Arabidopsisis encoded by the
GA-INSENSITIVE
(
GAI
) gene, which hasalso been cloned (Peng et al., 1997). The original
gai
allelecauses dwarfism and behaves genetically as a gain-of-func-tion mutation (Peng and Harberd, 1993). Recent molecularanalyses confirmed that this allele encodes a constitutivelyactive mutant protein that has apparently lost its ability torespond to GA (Peng et al., 1997). Additionally, extragenicsuppressors of GA mutants, defined by
rga
mutants, havebeen identified (Silverstone et al., 1997). RGA encodes aprotein similar to that of the
GAI
gene (Silverstone et al.,1998). The
gai
mutant allele that results in the GA-insensitivedwarf mutation contains a 51-bp in-frame deletion of 17amino acids in the DELLA domain (Peng et al., 1997). Peng etal. (1999) examined GA-insensitive dwarf mutants from otherspecies and demonstrated that the mutations of reducedheight (
RHT
)
–B1
and
RHT-D1
in wheat and
D8
in maize arecaused by an N-terminal truncation near the DELLA do-main. These results indicate that the DELLA domain is impor-tant to GA detection in the GA signal transduction pathway.
In wheat, some dwarf plants differ from tall plants in theirlack of GA response. These dwarf mutants have a lodging-resistant characteristic and have resulted in increasedwheat grain yields around the world since the 1960s. Theseso-called “green revolution” dwarf cultivars, which are de-rived from the Japanese variety Norin 10, have a phenotypewith a reduced response to GA that is caused by mutationsin the (
Rht-B1
and
Rht-D1
) gene, which is the ortholog of
GAI
(Peng et al., 1999).In this study, we isolated and characterized the
slender
-type mutant in rice (
slr1-1
). Biochemical analysis revealedthat the mutation results in a constitutive GA response.Consequently, we demonstrated that the
slr1-1
mutant con-tains a loss-of-function mutation in the
SLR1
gene, which isan ortholog of
GAI
and
RGA
in Arabidopsis,
RHT
in wheat,and
D8
in maize.
RESULTS
Isolation of
slender
Rice
A mutant with greatly accelerated extension growth was iso-lated from the rice cv Nipponbare after treatment with
g
irra-
diation. The phenotype of the mutant, designated
slender
(
slr1-1
), has very rapid extension growth in the seedling andis sterile. M2 progeny tests of heterozygotes yielded a seg-regation of 311 normal and 89
slender
plants (
x
2
[3:1]
5
1.47; 0.1
,
P
,
0.25), indicating monofactorial recessive in-heritance of the
slender
characteristic.
slender
Rice Behaves as if It Were Continually Saturated with GAs
One of the best known actions of GA is the stimulation ofshoot growth in rice (Murakami, 1968; Matsukura et al.,1998). Germinating seeds of progeny from the heterozygotesegregated a shoot length phenotype (Figure 1A). Shoots ofthe
slender
phenotype were more than twofold longer thanthose of the wild-type plant and were similar to those ofwild-type plants treated with gibberellic acid (GA
3
; Figure1B). Basal internodes in
slender
rice elongate concurrentlyduring seedling growth, which is usually observed in thewild-type plant treated with GA
3
(Figure 1C). The mutantalso had a reduced number and root length compared withthe wild-type plant (Figure 1D). Matsukura et al. (1998) re-ported that the promotion of leaf sheath growth by GA
3
isdue mainly to cell elongation. The average longitudinal andtransverse lengths of the epidermal cells at the apical por-tion of the second leaf sheath were approximately twofoldand 0.5-fold, respectively, the lengths of epidermal cells inthe wild type (Table 1). The overall results indicate that
slen-der
rice behaves as if it were continually saturated with GAs.The anatomic characteristics of
slender
rice are similarto those previously reported for the
ao-1
mutant in rice(Nakamura, 1992) and the
slender
mutant in barley (Favretand Malvarez, 1975; Foster, 1977).
slender
and wild-type plants were grown on agar plateswith or without uniconazole, a GA biosynthesis inhibitor(Izumi et al., 1984; Mitsunaga and Yamaguchi, 1993), for upto 15 days. The height of wild-type plants was decreasedsignificantly with uniconazole throughout the experimentalperiod, but the height of
slender
plants was unaffected (Fig-ure 2). This observation suggests that the elongation of
slen-der
is independent of endogenous GA levels.
Table 1.
Characteristics of Epidermal Cells in the Second Leaf Sheath of
slender
Rice and its Wild Typea
Position Plant Cell Length (mm) Cell Width (mm) Length/Width
Basal Wild type 69.3 6 11.8 19.7 6 1.9 3.5 slender 88.2 6 5.8 15.8 6 1.3 5.7
Apical Wild type 55.9 6 11.1 17.5 6 2.4 3.2 slender 107.9 6 10.3 9.2 6 0.7 11.7
a The second leaf sheath was dissected from 1-week-old slenderand wild-type plants and stained with Safranin. Values are means6SE (n 5 20).

1002 The Plant Cell
slender Mutation Results in Modulation of Endogenous GAs and a-Amylase Production
Endogenous levels of GAs were estimated using a combina-tion of HPLC purification and gas chromatography–massspectrometry analysis (Figure 3). The level of GA1, the activeGA molecule in vegetative tissues of rice, was lower in theshoots of slender rice than in the wild type. Similarly, thelevels of GA19 and GA20, both of which are inactive precur-sors of the GA1 molecule, were lower in slender than in thewild type (Figure 3), indicating that the mutation decreasesthe endogenous GA level in the shoot.
For further characterization of slender rice, agar plate as-says for amylases were conducted using the progeny seedsfrom selfing a plant heterozygous for the slender allele. Theembryoless half-seeds were placed on the starch plate withor without 1 mM GA3 for 2 days, and the starch was stainedwith iodine (Figure 4A). Production and secretion of a-amy-lase from wild-type embryoless half-seeds were observedas cleared zones (plaques) only on the plate containing GA3
(Figure 4A, 1GA3). Some slender half-seeds, however, pro-duced amylase even in the absence of exogenous GA3 (Fig-ure 4A, 2GA3). The ratio of the seeds requiring GA3 foramylase secretion to seeds requiring no GA3 was less than3:1 (30:7 actual values). This suggests that the M2 progenywere segregated into slender and normal characteristics fora-amylase production, because the aleurone layer consistsof triploid tissues. These results demonstrate that exoge-nous GA3 is not necessary for the synthesis and secretion ofamylases from the embryoless half-seeds of slender rice.
We confirmed that the secreted amylase was GA-induc-ible a-amylase (Figures 4B and 4C). Immunoblotting usingantisera against rice a-amylase (isoform A) revealed that theamylases secreted from the embryoless half-seeds of theslender mutant without GA3 application are identical tothose of wild-type seeds with GA3 application (Figure 4B,lanes 2 and 3). Isoform analysis using an isoelectric focusinggel revealed that the main amylase is an isoform A, the geneproduct of RAmy1A (Figure 4C; Mitsui et al., 1996; Yamaguchi,1998). The transcript of the rice a-amylase gene RAmy1A isabundant in aleurone layer cells during germination and it ispromoted by GA3 (Itoh et al., 1995; Sugimoto et al., 1998).These results indicate that a-amylase is produced indepen-dently of GAs from the aleurone layer cells of slender rice.
These findings (GA-mediated phenomena, shoot growth,and a-amylase production) suggest that slender rice (slr1-1)is a constitutive GA response mutant. A similar conclusionwas reported for the barley slender mutant (Chandler, 1988;Lanahan and Ho, 1988). Based on these results, it is likelyFigure 2. Effect of Uniconazole on Shoot Elongation in slender Rice
and Its Wild Type.
(A) Seed of wild-type (1 and 2) and slender rice (3 and 4) were germi-nated for 6 days with (2 and 4) or without (1 and 3) 6.9 mM unicona-zole.(B) Shoots of wild-type and slender rice were measured from 3 to 15days after germination. Results from wild-type (open squares) andslender rice (open circles) without uniconazole and wild-type (closed
squares) and slender rice (closed circles) with 6.9 mM uniconazoleare shown. Data are means (n 5 10). Variation in width was ,10% ofthe reported data.

SLR1 as a Rice GA Response Modulator 1003
that the SLR1 gene is an intermediate of the GA signalingpathway.
Molecular Cloning of the SLR1 Gene
Nipponbare (a Japonica rice) containing slr1-1 was crossedto Kasalath (an Indica rice), and segregation of slr1-1 andOsGAI (Ogawa et al., 2000) was examined to determine ifSLR1 and GAI are linked. GAI in Arabidopsis is an interme-diate of the GA signaling pathway. Direct sequencing wasperformed to determine the difference of the GAI homologbetween Nipponbare and Kasalath. Base differences at po-sition 1025 from the translation initiation site and their phe-notypes were as follows: A, Nipponbare with a slenderphenotype; G, Kasalath with a normal phenotype; and amixture of A and G in the heterozygous plant having a nor-mal phenotype. F2 progeny tests of the heterozygotesyielded a segregation of 49 normal and 16 slender plants,and genotypes based on the base change indicated 16 Nip-ponbare homozygous (Japonica), 33 heterozygous (Japon-ica plus Indica), and 16 Kasalath homozygous (Indica)plants. These results indicate tight linkage between the slen-der mutation (slr1-1) and OsGAI.
We determined the open reading frame (ORF) sequenceof the rice GAI homolog (data not shown). This sequence
was identical to that of OsGAI reported previously (Ogawaet al., 2000). We propose that OsGAI be renamed SLR1 af-ter its mutant phenotype and so use the term SLR1 in placeof OsGAI throughout this article. The deduced SLR1 proteinshares a high overall identity with RHT-D1a in wheat(77.2%), D8 in maize (80.3%), and RGA and GAI in Arabi-dopsis (41.2 and 47.2%, respectively). This protein contains
Figure 3. GA Content of Shoots in Wild-Type and slender Rice.
Ten-day-old shoots were measured for GA content by gas chroma-tography–mass spectrometry analysis. Ten-day-old shoots wereharvested from wild-type and slender rice and the GA content wasdetermined. Samples were performed on 10 independent plants.FW, fresh weight. Bars indicate 6SE.
Figure 4. a-Amylase Production from Embryoless Half-Seeds ofWild-Type and slender Rice.
(A) Embryoless half-seeds of wild-type and slender rice were placedon starch plates containing 1 mM GA3 (1GA3) or no GA3 (2GA3) for 2days, and starch was detected by staining with iodine.(B) and (C) Immunoblot and zymogram pattern of a-amylase ex-pressed in embryoless half-seeds of wild-type and slender rice. (B)shows immunochemical detection of a-amylase protein using a-amy-lase antibody. (C) shows zymogram pattern of a-amylase activity.Lanes 1, wild type without GA3 application; lanes 2, wild type withGA3 application; lanes 3, slender rice without GA3 application. Em-bryoless half-seeds of slender rice secreted isoform A (molecularmass z43 kD; pI 4.5), which is a GA3-inducible a-amylase in rice(Yamaguchi, 1998).
1004 The Plant Cell
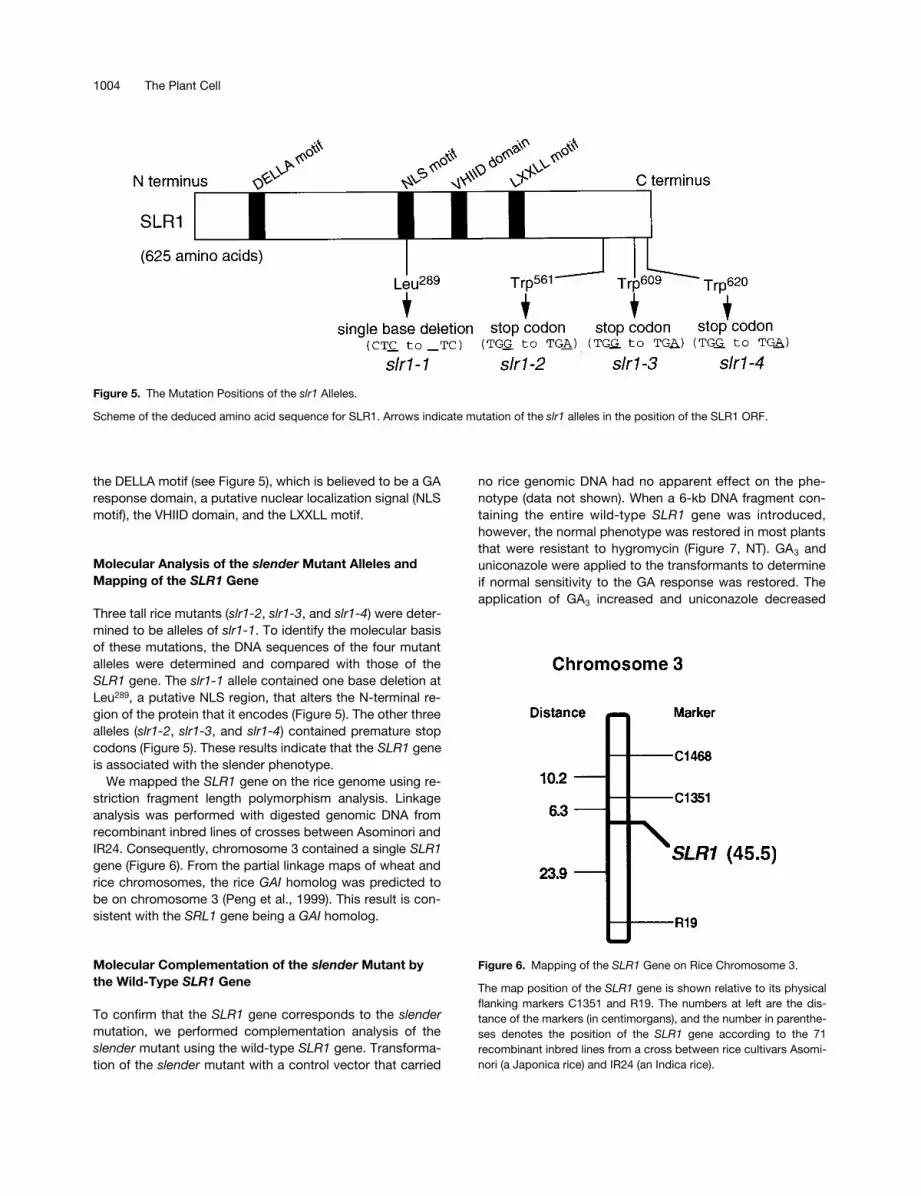
the DELLA motif (see Figure 5), which is believed to be a GAresponse domain, a putative nuclear localization signal (NLSmotif), the VHIID domain, and the LXXLL motif.
Molecular Analysis of the slender Mutant Alleles and Mapping of the SLR1 Gene
Three tall rice mutants (slr1-2, slr1-3, and slr1-4) were deter-mined to be alleles of slr1-1. To identify the molecular basisof these mutations, the DNA sequences of the four mutantalleles were determined and compared with those of theSLR1 gene. The slr1-1 allele contained one base deletion atLeu289, a putative NLS region, that alters the N-terminal re-gion of the protein that it encodes (Figure 5). The other threealleles (slr1-2, slr1-3, and slr1-4) contained premature stopcodons (Figure 5). These results indicate that the SLR1 geneis associated with the slender phenotype.
We mapped the SLR1 gene on the rice genome using re-striction fragment length polymorphism analysis. Linkageanalysis was performed with digested genomic DNA fromrecombinant inbred lines of crosses between Asominori andIR24. Consequently, chromosome 3 contained a single SLR1gene (Figure 6). From the partial linkage maps of wheat andrice chromosomes, the rice GAI homolog was predicted tobe on chromosome 3 (Peng et al., 1999). This result is con-sistent with the SRL1 gene being a GAI homolog.
Molecular Complementation of the slender Mutant by the Wild-Type SLR1 Gene
To confirm that the SLR1 gene corresponds to the slendermutation, we performed complementation analysis of theslender mutant using the wild-type SLR1 gene. Transforma-tion of the slender mutant with a control vector that carried
no rice genomic DNA had no apparent effect on the phe-notype (data not shown). When a 6-kb DNA fragment con-taining the entire wild-type SLR1 gene was introduced,however, the normal phenotype was restored in most plantsthat were resistant to hygromycin (Figure 7, NT). GA3 anduniconazole were applied to the transformants to determineif normal sensitivity to the GA response was restored. Theapplication of GA3 increased and uniconazole decreased
Figure 5. The Mutation Positions of the slr1 Alleles.
Scheme of the deduced amino acid sequence for SLR1. Arrows indicate mutation of the slr1 alleles in the position of the SLR1 ORF.
Figure 6. Mapping of the SLR1 Gene on Rice Chromosome 3.
The map position of the SLR1 gene is shown relative to its physicalflanking markers C1351 and R19. The numbers at left are the dis-tance of the markers (in centimorgans), and the number in parenthe-ses denotes the position of the SLR1 gene according to the 71recombinant inbred lines from a cross between rice cultivars Asomi-nori (a Japonica rice) and IR24 (an Indica rice).

SLR1 as a Rice GA Response Modulator 1005
plant height (Figure 7, 1GA3 and 1Unico), indicating thatthe introduction of the wild-type SRL1 gene complementedthe slender mutation. Based on these results, we concludedthat the slender mutant is caused by a loss-of-function mu-tation of the SLR1 gene, which is identical to the rice GAIhomolog.
Truncation of DELLA, a Putative GA Response Motif in the SLR1 Gene, Leads to the Dwarf Phenotype
In Arabidopsis, the GA-insensitive dwarf mutation allele gaicontains a 51-bp deletion of the GAI ORF. This in-frame de-letion results in the absence of 17 amino acid residues in theDELLA motif of the GAI protein. We attempted to produceGA-insensitive dwarf rice using the truncated SLR1 geneconstruct pSLRtr, which also contains a 17–amino acid de-letion (39-DELLAALGYKVRSSDMA-55) in the DELLA motif(Figure 8A). When the pSLRtr construct was introduced intothe normal Japonica rice (cv Nipponbare) to produce z200plants, .90% of transgenic rice resulted in a typical dwarfplant (Figure 8B, pSLRtr). The transgenic plant had retardedgrowth in the leaf sheath and blade and thickening in theroots, which are typical responses for a GA-deficient plant.The dwarf transgenic plant had no response, however, toexogenously applied GA3. Introduction of the truncated SLR1gene led to a wide range of plant heights, from severe, ex-tremely dwarf phenotypes to mild phenotypes (Figure 8C).
DISCUSSION
slender (slr1-1) Is a Constitutive GA Response Mutantin Rice
In this study, slender rice, which is caused by a single reces-sive mutation, was considered to be a constitutive GA re-sponse phenotype for the following reasons. The mutantshoot was more than twofold taller than the wild type, simi-lar to wild-type shoots treated with high amounts of GA3.The height of slender was unaffected by the application ofuniconazole, an inhibitor of GA biosynthesis, and endoge-nous levels of GAs were lower in slender than in the wildtype. Finally, GA-inducible a-amylase, the RAmy1A protein,was produced without GA3 application.
There are similar recessive slender-type mutants in barley(slender [sln1]; Chandler, 1988; Lanahan and Ho, 1988) andin pea (la crys; Potts et al., 1985). These mutants behave asif saturated with GAs and are not responsive to either exog-enous GAs or GA biosynthesis inhibitors. On the other hand,the SPY mutant in Arabidopsis (Jacobsen and Olszewski,1993) and the pro mutant in tomato (Jones, 1987) respondto applied GAs. These two types of mutants might be due tothe mutation of genes that have different functions or that
encode different intermediates in the GA signal transductionpathway.
Embryoless half-seeds of slender produced GA-induciblea-amylase without GA3, indicating that the SLR1 proteinmight be a negative regulator that blocks transcription of thea-amylase gene (see below). The induction of a-amylase byGAs in cereal grains represents a classic model system forstudying the mode of action of GAs (Jones and Jacobsen,1991). a-Amylase induction by GA occurs mainly at the tran-scriptional steps, which might be mediated through abscisicacid, Ca21 (Schuurink et al., 1996), cyclic GMP (Penson etal., 1996), or sugar signaling (Perata et al., 1997), and sig-nals communication among them (Toyofuku et al., 2000).GA-dependent transcriptional activation of the a-amylasegene requires the GA response cis-element complex andthe GAmyb protein as a trans-acting factor (Gubler et al.,1995). Further study is needed to clarify the targeting site ofthe SLR1 protein through the GA signaling pathway.
Figure 7. Complementation Analysis of the Slender Phenotype.
A 6-kb wild-type genomic DNA fragment containing the entire SLR1gene was used for transformation of the slender mutant (slr1-1). Thetransgenic plant had a normal phenotype without hormone treat-ment (NT). Application of 10 mM GA3 (1GA3) led to increased height,and treatment with 6.9 mM uniconazole (1Unico) led to reducedheight.

1006 The Plant Cell
The SLR1 Gene Is an Ortholog of the Height-Regulating Gene GAI/RGA/RHT/D8
We succeeded in the molecular cloning of the SLENDERgene (SLR1), which was originally derived from the rice GAIhomolog. Molecular analysis of slender mutant alleles re-vealed that four mutant alleles contain nonsense or stopcodon mutations in the SLR1 gene. Introduction of a 6-kbgenomic DNA fragment for the wild-type SLR1 gene into theslr1-1 mutant restored the normal phenotype and sensitivityto the GA response.
GAI, SLR1, D8, RHT-1Da, and RGA seem to be transcrip-tional factors belonging to the GRAS gene superfamily,which has a putative NLS, an Ser/Thr-rich repeat region,and the LXXLL motif. Indeed, in a transient assay using on-ion epidermal cells, RGA–GFP (green fluorescent protein)and OsGAI–GFP fusion proteins were localized exclusivelyin the nucleus (Silverstone et al., 1998; Ogawa et al., 2000).Transactivation assays indicate that the OsGAI protein has atranscriptional activator or coactivator (Ogawa et al., 2000).
Sequence comparison among members of the GAI familyidentified a putative Src homology 2 (SH2) phosphotyrosinebinding domain (Peng et al., 1999). The SH2 domain ispresent in a family of transcription factors called STATs (sig-nal transducers and activators of transcription) in animals(Darnell, 1997). The function of this domain is to mediate thebinding of STATs to various receptor tyrosine kinases. TheSTATs are then activated by the receptor kinase and trans-locate from the cytoplasm to the nucleus. If this putativeSH2 domain is functional in plants, then tyrosine phosphor-ylation might have a role in GA signaling (Silverstone andSun, 2000).
The slr1-4 mutation was detected at the extreme C-termi-nal end (Trp620) of the SLR1 protein (deduced ORF of 625amino acids). This mutant did not have any phenotypic dif-ferences from the other alleles. These results indicate thateven the loss of six amino acid residues at the C-terminalend, which are not a conserved motif among family mem-bers, abolishes the protein function in spite of the presenceof consensus motifs NLS, LXXLL, and SH2.
Figure 8. Truncation of the DELLA Motif in SLR1 Leads to a DwarfPhenotype.
(A) The construct pSLRtr was used for the transformation of riceplants. The Actin1 promoter (pAct1) was recovered from vectorpBI101-Hm3. Transformation with the SLRtr sequence resulted inthe DELLA motif truncation form (39-DELLAALGYKVRSSDMA-55) ofthe SLR1 ORF. nos, nopaline synthase.(B) Construct pSLRtr (A) was introduced into the normal Japonicarice Nipponbare (Control). Introduction of the truncated SLR1 generesulted in the typical dwarf plant (pSLRtr). The transgenic plantshows retarded growth in the leaf sheath and blade.(C) Introduction of the truncated SLR1 gene resulted in a range ofplant heights. Construct pSLRtr (A) was introduced into the wild-type cv Nipponbare (Control). The pSLRtr transgenic plants show awide range of plant heights, from weak to severe dwarf phenotypes.

SLR1 as a Rice GA Response Modulator 1007
SLR1 Is a Negative Regulator Involved in the GA Signaling Pathway
The slr1-1, slr1-2, and slr1-3 mutations truncate the protein,suggesting that these mutations are loss-of-function muta-tions. Therefore, SLR1 protein must be a negative regulatorof the GA signaling process, because lack of its negativeregulation results in a constitutive GA response phenotype(Figure 9).
Consistent with the similarity of the phenotypes of Rht-B1b, Rht-D1b, and D8 to the semidominant gai mutant, thegenetic lesions in these mutants are also similar to those ingai (Peng et al., 1999). The lesions in three D8 mutant alleleshave been identified. Both D8-1 and D8-2023 contain smallinternal deletions in the DELLA domain. The D8-Mpl allelehas a 330-bp deletion at the N terminus that begins in the59–untranslated amino acid region and extends throughVal84. Because of the gain-of-function (semidominant) na-ture of this mutant, it is proposed that it still encodes a trun-cated D8 protein that might be initiated at Met106 (Peng etal., 1999). Rht-B1b and Rht-D1b both contain a base substi-tution that introduces a stop codon in the DELLA domain.Because there is a methionine a short distance after thesestop codons, it is possible that translational reinitiation oc-curs to synthesize a protein lacking the original N terminus(Peng et al., 1999). These results support the hypothesisthat the DELLA domain in the N terminus of the GAI proteinfamily is responsible for modulating the activity of these pro-teins in response to the GA signal. Presumably, the effect ofeither the internal deletions or the N-terminal truncations isto lock the protein into a conformation that can no longer, orcan only weakly, respond to the GA signal (Silverstone andSun, 2000).
The Distinct Manner of Mutation on the GAI Ortholog Leads to Opposite Phenotypes in Terms ofGA Sensitivity
In Arabidopsis, two orthologous proteins, GAI and RGA,have a redundant GA response; therefore, even if one of theproteins is inactive, the plant still has a normal phenotype(Figure 9). In this case, a dominant negative mutation suchas the DELLA truncation is the only way to produce an ab-normal phenotype, such as the GA-insensitive dwarf. Thereis a similar redundancy in wheat, with its gene duplication,because wheat possesses three sets of chromosomes (A,B, and D genomes).
In rice, the slender mutant results in a loss-of-functionmutation of the SLR1 gene (Figure 9). Because the SLR1gene is likely to be nonredundant, the loss-of-functionmutation results in clearly abnormal phenotypes in termsof the GA response, that is, the slender and constitutiveGA response phenotype. We demonstrated that the dis-tinct manner of mutation on the orthologous genes leadsto opposite phenotypes in terms of the GA response:slender, a constitutive GA response, and a GA-insensitivedwarf.
Dwarfism in Rice and the Alternative Green Revolution
World wheat grain yields increased substantially in the1960s and 1970s because farmers rapidly adopted the newvarieties and cultivation methods of the so-called green rev-olution. The new varieties are shorter (dwarf or semidwarf)and increase grain yield by means of lodging resistance.These wheat cultivars are short because they contain theDELLA-truncated RHT gene (Peng et al., 1999).
With rice, a similar green revolution was realized by the in-troduction of the distinct gene sd-1 from GAI/RHT. IR8 wasestablished by crossing Dee-geo-woo-gen (sd-1) from aChinese cultivar to Peta from an Indonesian cultivar at theInternational Rice Research Institute (Los Baños, Philip-pines; Hargrove et al., 1980). In Japan, the semidwarf cul-tivar Reimei (d-49; allelic to sd-1), which was derived fromcv Fujiminori through g irradiation, was released in 1966(Futsuhara et al., 1967). Reimei was grown over wide areasof the northern part of Japan. Furthermore, it has been usedas a parent strain for cross-breeding, taking advantage of itslodging resistance and high yields. We demonstrated thatthe introduction of the truncated SLR1 gene results in arange of plant heights (Figure 8C). DNA gel blot analysis re-vealed that these plants possess a single gene introductionand that the diversity in height is due mainly to the positionaleffect of the transgene insertion (data not shown), suggest-ing the potential for a plant height–regulating system. Be-cause the SLR1 gene is distinct from sd-1 (the rice greenrevolution gene), it presents an alternative means to pro-duce dwarf rice plants. Further investigation is needed toevaluate this hypothesis.
Figure 9. Model of the GA Signal Transduction Pathway in Rice, Ar-abidopsis, and Wheat.
Arrows indicate the GA signal transduction pathway from GA recep-tion (GA) to various actions of GA, such as shoot elongation, a-amy-lase induction, and so on. SLR1 in rice, GAI and RGA in Arabidopsis,and RHT-A1, -B1, and -D1 proteins in wheat negatively regulate theGA signaling pathway.

1008 The Plant Cell
METHODS
Isolation of slender Rice
A recessive mutant, slender1-1, was isolated from the progeny ofrice seeds (Oryza sativa cv Nipponbare, a Japonica rice) irradiatedwith 200 Gy of g-rays (dose rate 2 Gy/min). Surface-sterilized seedsof wild-type and mutant plants were soaked in water for 3 days andthen placed in artificial soil for 11 days and grown in a greenhouse.Mutant segregants were distinguished from normal segregants bythe extremely extended phenotype. The other slender alleles (slr1-2,1-3, and 1-4) were independently isolated.
Measurement of Shoot Elongation
Shoot elongation was quantified by a modification of the method de-scribed by Matsukura et al. (1998). Seeds of wild-type and mutantplants (n 5 25) were surface sterilized for 30 min with a 3% NaClOsolution, washed three times with sterile distilled water, soaked in thedistilled water for 24 hr in the presence or absence of 6.9 mM unicon-azole, and then placed in sterile distilled water for an additional 24 hr.The seeds were then placed on a 1% agar plate and grown under flu-orescent lamps at 308C. For the surface anatomy, the second leafsheath was dissected from third leaf–stage plants and stained withSafranin (Wako Chemical, Osaka, Japan).
Agar Plate Assay, Immunoblotting, and Zymographyof a-Amylase
The agar plate assay was performed essentially as described byLanahan and Ho (1988). Seeds were cut transversely, and the half-seed containing the embryos were planted to determine their pheno-types. The embryoless half-seeds were surface sterilized with 3%NaClO for 15 min and washed six times with sterile water. These half-seeds were then placed on 2% agar plates containing 10 mM so-dium acetate and 2 mM CaCl2 at pH 5.3. GA plates were made byadding 1 mM gibberellic acid (GA3) to the cooled agar. To detect se-creted a-amylase activity, we added soluble potato starch (0.2%) tothe agar before autoclaving. Agar plates were developed by incubat-ing the plates in I2 gas. Half-seeds that synthesized and secreteda-amylase had transparent halos around them resulting from the di-gestion of the starch by amylases.
Proteins to be examined were separated using SDS-PAGE or iso-electric focusing, transferred to a nitrocellulose membrane using theNovablot Protein Transfer Kit, and analyzed using the anti-a-amylaseantibody (Mitsunaga and Yamaguchi, 1993). An alkaline phos-phatase–labeled secondary antibody was used to detect the immu-noreactive band.
The crude extract was examined by isoelectric focusing usingPharmacia broad range (pH 3.5 to 9.5) Ampholine Pageplates. Iso-electric focusing was performed for 1.5 hr according to the manufac-turer’s instructions. Samples (15 mL of crude extract) were applied toapplication paper placed 3 cm distant from the cathode. The appli-cation paper was removed after 45 min of electrophoresis, and therun was continued for an additional 45 min. To visualize the bands ofamylolytic activities, the gel was incubated for 1 hr in 50 mM sodiumacetate buffer, pH 5.2, containing 10 mM CaCl2 and 1% boiled solu-
ble starch. After washing with distilled water, the gel was stained with0.6% I2 and 6% KI solution (Perata et al., 1992).
Quantitative Analyses of Endogenous GAs
GAs from each sample were extracted and purified according to themethod of Hisamatsu et al. (1998). As internal standards, 70 ng of17,17,2H2-GA1, 100 ng of 17,17,2H2-GA19, and 50 ng of 17,17,2H2-GA20
were added to each sample. After several HPLC purification steps,fractions that had GA activity on a rice assay (Nishijima et al., 1992)were methylated and trimethylsilylated in glass tubes. The derivedsamples were analyzed using gas chromatography–mass spectrom-etry. The concentrations of GA1, GA19, and GA20 in the tubes werecalculated from the ratios of peak areas at mass-to-charge ratios of506:508, 434:436, and 418:420, respectively, according to themethod of Gaskin and MacMillan (1991).
Screening of the SLR1 Gene from Rice Genomic Libraries
To isolate the SLR1 gene, we used the rice GAI homologous ex-pressed sequence tag clone (accession number D39460). GenomicDNA libraries were constructed from rice. Nuclear genomic DNA wasisolated from 2-week-old seedlings. The DNA was partially digestedwith Sau3AI and enriched for fragments of z20 kb on a sucrose gra-dient. The fragments were cloned into the BamHI site of EMBL3(Stratagene, La Jolla, CA).
Screening by hybridization was performed in 50% formamide, 6 3SSC (1 3 SSC is 0.15 M NaCl and 0.015 M sodium citrate), 5 3 Den-hardt’s solution (1 3 Denhardt’s solution is 0.02% Ficoll, 0.02%polyvinylpyrrolidone, and 0.02% BSA), 0.5% SDS, and 0.1 mg/mLsalmon sperm DNA at 428C for 14 hr using the GAI homologous ex-pressed sequence tag clone as a probe.
Mapping of the SLR1 Gene in Rice Recombinant Inbred Lines
To map the SLR1 gene, 71 recombinant inbred lines from a cross be-tween two rice cultivars, Asominori (a Japonica rice) and IR24 (an In-dica rice), were used. Restriction fragment length polymorphismanalysis was performed with a probe specific for the rice GAI homol-ogous expressed sequence tag clone. DNA gel blot hybridizationwas performed as described by Church and Gilbert (1984) exceptthat membranes were hybridized at high stringency (688C). Thelinkage analysis was performed using the MAPMAKER program(Whitehead Institute for Biomedical Research/Massachusetts Insti-tute of Technology Center for Genome Research, Wilmington, MA).
Genomic DNA Isolation and Direct Genome Sequencing
Rice genomic DNA was isolated with the ISOPLANT DNA isolation kit(Nippon Gene Co., Tokyo, Japan). The coding region of the SLR1gene was amplified by polymerase chain reaction (PCR) using spe-cific primers S6 (59-TCGTCGTCCTCATCGTCGTC-39) and A6 (59-GCAGCCGGTGCAGCTCGAAC-39) from 300 ng of rice genomic DNA.PCR products were gel purified, and the nucleotide sequences weredetermined using the dideoxynucleotide chain termination methodwith an automated sequencing system (ABI373A; Applied Biosys-tem, Inc., Foster City, CA). Analysis of the nucleotide sequences was

SLR1 as a Rice GA Response Modulator 1009
performed using DANSIS computer software (Hitachi Software Engi-neering, Tokyo, Japan).
Rice Transformation and Complementation of the slr1-1 Mutant
A genomic clone, pAI1, including the entire coding region and the 59
and 39 flanking regions, was inserted as a 6-kb restriction fragmentbetween the SmaI sites of the hygromycin resistance binary vectorpBI101-Hm3 (Sato et al., 1999). The pAI1 gene construct was intro-duced into the slr1-1 mutant as described above. Control plantswere transformed using the vector pBI-cont.
pSLRtr was made as follows. A 123-bp fragment was amplified byPCR from 6 kb of genomic DNA containing the SLR ORF using a 59
primer containing an additional XbaI site (59-TCTAGAATGAAGCGC-GAGTA-39) and a 39 primer containing an additional SalI site (59-GTCGACGTCCTCCTCCTCCC-39). A 1722-bp fragment was ampli-fied by PCR from 6 kb of genomic DNA containing the SLR ORF us-ing a 59 primer containing an additional SalI site (59-GTCGAC-GTCGCGCAGAAGCT-39) and a 39 primer containing an additionalSmaI site (59-CCCGGGTCACGCCGCGGCGA-39). These PCR prod-ucts were cloned into a pBIAct1nos vector (Yamamuro et al., 2000).pBIAct1nos containing the actin (Act1) promoter (Zhang et al., 1991)and the nopaline synthase terminator was prepared by substitutingthe Act1 promoter in the hygromycin resistance binary vector be-tween the XbaI and SmaI sites. pBI-cont, which contains no insert,was used as a control vector. pSLR1tr and the control constructwere introduced into rice cv Nipponbare by Agrobacterium tumefa-ciens–mediated transformation, as described by Hiei et al. (1994).
ACKNOWLEDGMENTS
We are grateful to Dr. Pierdomenico Perata for critically reviewing themanuscript and Professor Kenzo Nakamura (Nagoya University) forproviding us with the pBI101-Hm3 vector. This work was supportedby Grants-in-Aid for Scientific Research (Grant Nos. 09660004 and10460003) from the Ministry of Education, Science, Sports, and Cul-ture, Japan. A.I. acknowledges a Research Fellowship of the JapanSociety for the Promotion of Science for Young Scientists (2000–2001). We thank Dr. T. Sasaki of the Japanese Rice Genome Pro-gram for the expressed sequence tag clones.
Received October 23, 2000; accepted March 7, 2001.
REFERENCES
Chandler, P.M. (1988). Hormonal regulation of gene expression inthe “slender” mutant of barley (Hordeum vulgare L.). Planta 175,115–120.
Church, G., and Gilbert, W. (1984). Genomic sequencing. Proc.Natl. Acad. Sci. USA 81, 1991–1995.
Croker, S.J., Hedden, P., Lenton, J.R., and Stoddart, J.L. (1990).Comparison of gibberellins in normal and slender barley seed-lings. Plant Physiol. 94, 194–200.
Darnell, J.E., Jr. (1997). STATs and gene regulation. Science 277,1630–1635.
Favret, E.A., and Malvarez, E.M. (1975). Genetic regulatory mecha-nisms for seedling growth in barley. Barley Genet. 3, 37–43.
Foster, C.A. (1977). Slender: An accelerated extension growthmutant of barley. Barley Genet. Newsl. 7, 24–27.
Futsuhara, Y., Toriyama, K., and Tsunoda, K. (1967). Breeding ofthe new rice variety, “Reimei,” through gamma-ray irradiation.Jpn. J. Breed. 17, 85–90.
Gaskin, P., and MacMillan, J. (1991). Quantitative analysis. In GC-MSof the Gibberellins and Related Compounds: Methodology and aLibrary of Spectra, P. Gaskin and J. MacMillan, eds (Bristol, UK:Cantock’s Enterprises), pp. 117–124.
Gubler, F., Kalla, R., Roberts, J.K., and Jacobsen, J.V. (1995).Gibberellin-regulated expression of a myb gene in barley aleuronecells: Evidence for Myb transactivation of a high-pI a-amylasegene promoter. Plant Cell 7, 1879–1891.
Hargrove, T.R., Coffman, W.R., and Cabanilla, V.L. (1980). Ances-try of improved cultivars of Asian rice. Crop Sci. 20, 721–727.
Hiei, Y., Ohta, S., Komari, T., and Kumashiro, T. (1994). Efficienttransformation of rice (Oryza sativa L.) mediated by Agrobacteriumand sequence analysis of the boundaries of the T-DNA. Plant J. 6,271–282.
Hisamatsu, T., Koshioka, M., Kubota, S., King, R.W., andMander, L.N. (1998). Isolation and identification of GA12 (12b-hydroxy-GA12) in Matthiola incana. Phytochemistry 47, 3–6.
Hooley, R. (1994). Gibberellins: Perception, transduction andresponses. Plant Mol. Biol. 26, 1529–1555.
Itoh, K., Yamaguchi, J., Huang, N., Rodriguez, R.L., Akazawa, T.,and Shimamoto, K. (1995). Developmental and hormonal regula-tion of rice a-amylase (RAmy1A)-gusA fusion genes in transgenicrice seeds. Plant Physiol. 107, 25–31.
Izumi, K., Yamaguchi, I., Wada, A., Oshio, H., and Takahashi, N.(1984). Effects of a new plant growth retardant (E)-1-(4-chlorophe-nyl)-4,4-dimethyl-2-(1,2,4-triazol-1-yl)-1-penten-3-ol (S-3307) onthe growth and gibberellin content of rice plants. Plant Cell Phys-iol. 25, 611–617.
Jacobsen, S.E., and Olszewski, N.E. (1993). Mutations at theSPINDLY locus of Arabidopsis alter gibberellin signal transduc-tion. Plant Cell 5, 887–896.
Jones, M.G. (1987). Gibberellins and the procera mutant of tomato.Planta 172, 280–284.
Jones, R.L., and Jacobsen, J.V. (1991). Regulation of synthesisand transport of secreted proteins in cereal aleurone. Int. Rev.Cytol. 126, 49–88.
Lanahan, M.B., and Ho, T.-H.D. (1988). Slender barley: A constitu-tive gibberellin-response mutant. Planta 175, 107–114.
Matsukura, C., Itoh, S., Nemoto, K., Tanimoto, E., and Yamaguchi,J. (1998). Promotion of leaf sheath growth by gibberellic acid in adwarf mutant of rice. Planta 205, 145–152.
Mitsui, T., Yamaguchi, J., and Akazawa, T. (1996). Physicochemi-cal and serological characterization of rice a-amylase isoformsand identification of their corresponding genes. Plant Physiol.110, 1395–1404.

1010 The Plant Cell
Mitsunaga, S., and Yamaguchi, J. (1993). Production of a-amylaseis repressed by uniconazole, an inhibitor of the biosynthesis ofgibberellin, in a dwarf mutant of rice, Waito-C. Plant Cell Physiol.34, 243–249.
Mitsunaga, S., Tashiro, T., and Yamaguchi, J. (1994). Identifica-tion and characterization of gibberellin-insensitive mutants selectedfrom among dwarf mutants of rice. Theor. Appl. Genet. 87, 705–712.
Murakami, H. (1968). A new rice seedling test for gibberellins,‘microdrop method’ and its use for testing extracts of rice andmorning glory. Bot. Mag. Tokyo 81, 33–43.
Nakamura, I. (1992). A rice mutant showing accelerated internodeovergrowth. Rice Genet. Newsl. 9, 61–62.
Nishijima, T., Koshioka, M., and Yamaji, H. (1992). A non-dwarfrice seedling bioassay for gibberellins. Plant Physiol. 98, 962–965.
Ogawa, M., Kusano, T., Katsumi, M., and Sano, H. (2000). Ricegibberellin-insensitive gene homolog, OsGAI, encodes a nuclear-localized protein capable of gene activation at the transcriptionallevel. Gene 245, 21–29.
Peng, J., and Harberd, N.P. (1993). Derivative alleles of the Arabi-dopsis gibberellin-insensitive (gai) mutation confer a wild-typephenotype. Plant Cell 5, 351–360.
Peng, J., Carol, P., Richards, D.E., King, K.E., Cowling, R.J.,Murphy, G.P., and Harberd, N.P. (1997). The Arabidopsis GAIgene defines a signaling pathway that negatively regulates gibber-ellin responses. Genes Dev. 11, 3194–3205.
Peng, J., et al. (1999). ‘Green revolution’ genes encode mutant gib-berellin response modulators. Nature 400, 256–261.
Penson, S.P., Schuurink, R.C., Fath, A., Gubler, F., Jacobsen,J.V., and Jones, R.L. (1996). cGMP is required for gibberellicacid–induced gene expression in barley aleurone. Plant Cell 8,2325–2333.
Perata, P., Pozueta-Romero, J., Akazawa, T., and Yamaguchi, J.(1992). Effect of anoxia on starch breakdown in rice and wheatseeds. Planta 188, 611–618.
Perata, P., Matsukura, C., Vernieri, P., and Yamaguchi, J. (1997).Sugar repression of a gibberellin-dependent signaling pathway inbarley embryos. Plant Cell 9, 2197–2208.
Potts, W.C., Reid, J.B., and Murfet, I.C. (1985). Internode length inPisum: Gibberellins and the slender phenotype. Physiol. Plant. 63,357–364.
Sato, Y., Sentoku, N., Miura, Y., Hirochika, H., Kitano, H., andMatsuoka, M. (1999). Loss of function mutations in the ricehomeobox gene OSH15 affect the architecture of internodesresulting in dwarf plants. EMBO J. 18, 992–1002.
Schuurink, R.C., Chan, P.V., and Jones, R.L. (1996). Modulation ofcalmodulin mRNA and protein levels in barley aleurone. PlantPhysiol. 111, 371–380.
Silverstone, A.L., and Sun, T. (2000). Gibberellins and the greenrevolution. Trends Plant Sci. 5, 1–2.
Silverstone, A.L., Mak, P.Y.A., Casamitjana-Martinez, E., andSun, T.-p. (1997). The new RGA locus encodes a negative regula-tor of gibberellin response in Arabidopsis thaliana. Genetics 146,1087–1099.
Silverstone, A.L., Ciampaglio, C.N., and Sun, T.-p. (1998). TheArabidopsis RGA gene encodes a transcriptional regulatorrepressing the gibberellin signal transduction pathway. Plant Cell10, 155–159.
Sugimoto, N., Takeda, G., Nagato, Y., and Yamaguchi, J. (1998).Temporal and spatial expression pattern of a-amylase gene dur-ing germination in rice and barley. Plant Cell Physiol. 39, 323–333.
Swain, S.M., and Olszewski, N.E. (1996). Genetic analysis of gib-berellin signal transduction. Plant Physiol. 112, 11–17.
Taylor, C.B. (1998). GA signaling: Genes and GTPases. Plant Cell10, 131–133.
Thornton, T.M., Swain, S.M., and Olszewski, N.E. (1999). Gibber-ellin signal transduction presents . . . the SPY who O-GlcNAc’dme. Trends Plant Sci. 4, 424–428.
Toyofuku, K., Loreti, E., Vernieri, P., Alpi, A., Perata, P., andYamaguchi, J. (2000). Glucose modulates the abscisic acid–inducible Rab16A gene in cereal embryos. Plant Mol. Biol. 42,451–460.
Yamaguchi, J. (1998). Analysis of embryo-specific a-amylase usingisolated mature rice embryos. Breeding Sci. 48, 365–370.
Yamamuro, C., Ihara, Y., Wu, X., Noguchi, T., Fujioka, S., Takatsuto,S., Ashikari, M., Kitano, H., and Matsuoka, M. (2000). Loss offunction of a rice brassinosteroid insensitive1 homolog preventsinternode elongation and bending of the lamina joint. Plant Cell12, 1591–1606.
Zhang, W., McElroy, D., and Wu, R. (1991). Analysis of rice Act1 59
region activity in transgenic rice plants. Plant Cell 3, 1155–1165.

DOI 10.1105/tpc.13.5.999 2001;13;999-1010Plant Cell
Futsuhara, Makoto Matsuoka and Junji YamaguchiAkira Ikeda, Miyako Ueguchi-Tanaka, Yutaka Sonoda, Hidemi Kitano, Masaji Koshioka, Yuzo
GAI/RGA/RHT/D8 Gene, an Ortholog of the Height-Regulating Gene SLR1 Rice, a Constitutive Gibberellin Response Mutant, Is Caused by a Null Mutation of the slender
This information is current as of August 8, 2015
References http://www.plantcell.org/content/13/5/999.full.html#ref-list-1
This article cites 45 articles, 22 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists






![[Flanders versus The Netherlands: focus on differences between depressive symptoms in men and women measured on the basis of CES-D8]](https://img.pdfslide.net/doc/110x75/634763316e55ee279302b9bd/flanders-versus-the-netherlands-focus-on-differences-between-depressive-symptoms.jpg)






















