Embed Size (px)
Citation preview

266
Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578 Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578 Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578 Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578
Therapeutics, Targets, and Chemical Biology
CancerResearch
Small-Molecule Inhibitors of Phosphatidylinositol 3-Kinase/Akt Signaling Inhibit Wnt/β-Catenin Pathway Cross-Talkand Suppress Medulloblastoma GrowthNinib Baryawno1, Baldur Sveinbjörnsson1,2, Staffan Eksborg1, Ching-Shih Chen3,Per Kogner1, and John Inge Johnsen1
Abstract
Authors' AWomen's a2Division of3Division ofThe Ohio S
Note: SupResearch O
CorresponUnit, Q6:05Sweden. Pbaryawno@
doi: 10.115
©2010 Am
Cancer R
DowDowDowDow
Activation of the β-catenin and receptor kinase pathways occurs often in medulloblastoma, the mostcommon pediatric malignant brain tumor. In this study, we show that molecular cross-talk between theβ-catenin and phosphatidylinositol 3-kinase (PI3K)/Akt signaling pathways is crucial to sustain medulloblas-toma pathophysiology. Constitutive activation of phosphoinositide-dependent protein kinase 1 (PDK1), Akt,and glycogen synthase kinase 3β (GSK-3β) was detected by immunohistochemistry in all primary medul-loblastomas examined (n = 41). Small-molecule inhibitors targeting the PI3K/Akt signaling pathway affectedβ-catenin signaling by inhibition of GSK-3β activity, resulting in cytoplasmic retention ofβ-catenin and reducedexpression of its target genes cyclin D1 and c-Myc. The PDK1 inhibitor OSU03012 induced mitochondrial-dependent apoptosis of medulloblastoma cells and enhanced the cytotoxic effects of chemotherapeutic drugsin a synergistic or additive manner. In vivo, OSU03012 inhibited the growth of established medulloblastomaxenograft tumors in a dose-dependent manner and augmented the antitumor effects of mammalian targetof rapamycin inhibitor CCI-779. These findings demonstrate the importance of cross-talk between the PI3K/Akt and β-catenin pathways in medulloblastoma and rationalize the PI3K/Akt signaling pathway as a thera-peutic target in treatment of this disease. Cancer Res; 70(1); 266–76. ©2010 AACR.
Introduction
Medulloblastoma, a primitive neuroectodermal tumor, isthe most common malignant pediatric brain tumor. Medul-loblastoma can originate from cerebellar granule neural pre-cursor cells located in the external granule layer (1, 2) andtypically develop in the cerebellum. Medulloblastoma cellsretain many features that resemble the progenitor cells ofthe embryonic brain (2), and more than half of these tu-mors display abnormal activation of the Hedgehog or Wntsignaling pathways (3), supporting a model of embryonaltumorigenesis.Wnt signaling plays a central role in modulating the bal-
ance between proliferation and differentiation of progenitorcells during embryonic central nervous system development(4). The canonical Wnt signaling pathway operates by stabi-lizing β-catenin. In the absence of Wnt/Wingless ligand
ffiliations: 1Childhood Cancer Research Unit, Department ofnd Children's Health, Karolinska Institutet, Stockholm, Sweden;Immunology, IMB, University of Tromsö, Tromsö, Norway; andMedicinal Chemistry and Pharmacology, College of Pharmacy,tate University, Columbus, Ohio
plementary data for this article are available at Cancernline (http://cancerres.aacrjournals.org/).
ding Author: Ninib Baryawno, Childhood Cancer Research, Astrid Lindgrens Children's Hospital, S-17176 Stockholm,hone: 46-851-777515; Fax: 46-851-773475; E-mail: ninib.ki.se.
8/0008-5472.CAN-09-0578
erican Association for Cancer Research.
es; 70(1) January 1, 2010
Researcon Septembecancerres.aacrjournals.org nloaded from
Researcon Septembecancerres.aacrjournals.org nloaded from
Researcon Septembecancerres.aacrjournals.org nloaded from
Researcon Septembecancerres.aacrjournals.org nloaded from
activation, β-catenin is sequestered in the cytoplasm by amultiprotein complex, which encompasses the adenomatouspolyposis coli protein, axin1, axin2/conductin, casein kinase1, and glycogen synthase kinase-3β (GSK-3β). In this state,β-catenin is phosphorylated at the NH2-terminal serine andthreonine residues by GSK-3β, which targets it for ubiquiti-nation and proteolytic degradation (5). Activation of Wnt sig-naling by binding of Wnt ligands to a Frizzled receptorinhibits the formation of the multiprotein complex andGSK-3β–mediated phosphorylation of β-catenin, resultingin an accumulation of hypophosphorylated β-catenin in thecytosol. Stabilized hypophosphorylated β-catenin eventuallytranslocates to the nucleus where it interacts with membersof the T-cell factor/lymphoid enhancer binding factor (TCF/LEF) family of transcription factors, leading to modulatedexpression of a broad range of genes, such as MYC andCCND1 (6, 7).Proteins regulating signaling through the phosphatidyli-
nositol 3-kinase (PI3K)/Akt pathway are frequently alteredin human cancer, including medulloblastoma (8, 9). Acti-vated Akt significantly augments Sonic-Hedgehog–inducedmedulloblastoma formation in mice (10). Activation ofthe PI3K/Akt signaling pathway is important for prolifera-tion of human medulloblastoma cells and cancer stemcells residing in the perivascular niche following irradiation(11, 12). Moreover, medulloblastomas frequently display re-duced expression of the Akt inhibitor PTEN (the tumorsuppressor phosphatase and tensin homologue deleted onchromosome 10) caused by promoter hypermethylation
h. r 14, 2015. © 2010 American Association for Cancerh. r 14, 2015. © 2010 American Association for Cancerh. r 14, 2015. © 2010 American Association for Cancerh. r 14, 2015. © 2010 American Association for Cancer

Inhibitors of PI3K/Akt-Wnt/β-Catenin Cross-Talk
Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578
and allelic losses on chromosome region 10q23 wherePTEN is localized (8, 13).PI3Ks are key components for the activation of Akt signaling
by catalyzing the generation of phosphatidylinositol-3,4,5-trisphosphate (PIP3) from phosphatidylinositol-4,5-bisphosphate(PIP2). PIP3 translocates Akt and phosphoinositide-dependentprotein kinase 1 (PDK1) from the cytoplasm to the inner cyto-plasmic membrane where Akt is activated through phosphoryla-tion by PDK1. This process is attenuated by PTEN, whichremoves the 3′-phosphate group of PIP3, resulting in the regener-ation of PIP2 and decreased Akt activity (8). Activated Aktphosphorylates a plethora of downstream substrates involvedin the regulation of cell survival, cell cycle progression, andcellular growth (10).In this study, we evaluated the importance of PI3K/Akt sig-
naling in medulloblastomas using small-molecule inhibitorsand show that molecular cross-talk between PI3K/Akt andWnt/β-catenin signaling is significant in medulloblastomatumorigenesis.
Materials and Methods
Tumor material and patient characteristics. Human tu-mor tissue samples were collected at the Karolinska Univer-sity Hospital, Sweden, between 1994 and 2005. Ethicalapproval was obtained by the Karolinska University HospitalResearch Ethics Committee. Tumor and patient characteris-tics are summarized in Supplementary Table S1.Immunohistochemistry and immunofluorescence. De-
tection of phospho-PDK1Ser241 (Abcam), phospho-AktSer473,phospho-AktThr308, phospho-GSK-3βSer9, and β-catenin inprimary tumor samples was performed using thecorresponding specific antibodies (Cell Signaling). Sectionsfrom xenograft tumors were incubated with the primary anti-bodies detecting Ki-67 (NeoMarkers) or active caspase-3(R&D Systems). As a secondary antibody, the horseradishperoxidase (HRP) SuperPicture Polymer detection kit andmatched isotype controls were used (Zymed Laboratories,Inc.). Proliferation (Ki-67) and apoptosis (cleaved caspase-3)were quantified as described (14).For immunofluorescence analysis, medulloblastoma cells
were treated with 1.5 to 3.0 μmol/L of OSU03012 for 6 hand then incubated overnight at 4°C with primary antibodydetecting β-catenin (Cell Signaling). Detection was per-formed using secondary biotinylated antibody and FITC-conjugated streptavidin (Zymed Laboratories).Chemicals. 2-Amino-N(4-5-(2-phenanthrenyl)-3-(trifluoro-
methyl)-1H-pyrazol-1-yl]phenyl)-acetamide (OSU03012),LY294002 (Cell Signaling), SB-216763, doxorubicin and vin-cristine (Sigma-Aldrich), and CHIR99021 (Stemgent) weredissolved in DMSO (Sigma-Aldrich). Rapamycin (sirolimus,LC Laboratories) and CCI-779 (temsirolimus, Wyeth) weredissolved in 99.5% ethanol. Temozolomide and cyclophos-phamide were supplied by the local pharmacy (ApoteketAB) and diluted in PBS to the appropriate stock concentra-tions. Sodium nitroprusside dihydrate (SNP; Sigma-Aldrich)was dissolved in distilled water. All inhibitors were furtherdiluted in Opti-MEM (Life Technologies) to the desired
www.aacrjournals.org
Researcon Septembecancerres.aacrjournals.org Downloaded from
in vitro concentration. For in vivo use of CCI-779, the stockwas diluted in 5% polyethylene glycol 400 (Sigma-Aldrich),5% Tween 20 (Bio-Rad), and 0.9% sterile saline, where-as OSU03012 for oral administration was prepared as asuspension in a vehicle fluid consisting of 0.5% (w/v)methylcellulose (Sigma-Aldrich) and 0.1% (v/v) Tween 80(Sigma-Aldrich) in sterile water.Cell lines and viability assays. Origin, culturing condi-
tions, and viability assays of human medulloblastoma and su-pratentorial primitive neuroectodermal (sPNET) cell lineswere as described (14).Fluorescence-activated cell sorting analysis. DNA con-
tent was assessed essentially as described (15). Briefly,D283 MED, D324 MED, PFSK-1, D458 MED, UW228-3, andMEB-MED-8A cells were treated with 3 μmol/L OSU03012for 24 and 48 h; stained with 4′,6-diamidino-2-phenylindole(DAPI; Sigma-Aldrich); and subjected to cell cycle analysisusing single-parameter DNA flow cytometry. The mitochon-drial transmembrane potential was assessed in D283 MED,D324 MED, PFSK-1, D458 MED, UW228-3, and MEB-MED-8Acells after 24-h incubation with 3 μmol/L OSU03012 usingtetramethylrhodamine ethyl ester (Molecular Probes) asdescribed (16).Protein fractionation, immunoprecipitation, and immu-
noblotting. Fractionated nuclear and cytosolic protein ly-sates were obtained using nuclear extraction buffer [20mmol/L Tris-HCl (pH 7.5), 420 mmol/L NaCl, 1.5 mmol/LMgCl2, 0.2 mmol/L EDTA, 25% glycerol, 1 mmol/L EDTA, 1mmol/L DTT, 0.1 mmol/L phenylmethylsulfonyl fluoride(PMSF)] or hypotonic lysis buffer [10 mmol/L HEPES(pH 7.9), 60 mmol/L KCl, 0.3% NP40, 1 mmol/L EDTA, 1mmol/L DTT, 0.1 mmol/L PMSF], respectively. Total cellprotein lysates were extracted from cells in radioimmuno-precipitation assay buffer [25 mmol/L Tris (pH 7.8), 2mmol/L EDTA, 20% glycerol, 0.1% NP40, 1 mmol/L DTT].Frozen tumors were disrupted with a rotor-stator homog-enizer in TNEN buffer [50 mmol/L Tris-HCl (pH 7.5), 250mmol/L NaCl, 0.1% NP40, 5 mmol/L EDTA] and subjectedto five freeze-thaw cycles (liquid nitrogen, 37°C, beforeclearing by centrifugation). All protein extraction bufferswere supplemented with MiniComplete protease inhibitorcocktail (Roche Diagnostic), 1 mmol/L NaF, and 1 mmol/LNaO3V4. Proteins were immunoprecipitated using axin1antibodies (Cell Signaling) coupled to agarose A beads fol-lowing the manufacturer's instructions (Sigma-Aldrich). Im-munoblotting was performed using antibodies detectingphospho-PDK1Ser241, phospho-AktSer473, phospho-AktThr308,phospho-S6K1Thr389, phospho-4E-BP1Ser65, phospho-GSK-3βSer9, phospho-β-cateninSer33/37/Thr41, Akt, β-catenin, cas-pase-9, caspase-3, poly(ADP-ribose) polymerase (PARP),the BH3-interacting domain death agonist (BID), cyclinD1, c-Myc (all from Cell Signaling), and β-actin (Sigma-Aldrich). Anti-mouse IgG and anti-rabbit IgG, conjugatedwith HRP (Pharmacia Biosciences), were used as secondaryantibodies. SuperSignal (Pierce) was used for chemilumi-nescent detection.Transfection, small interfering RNA knockdown, and
luciferase reporter gene assays. Cells were transfected with
Cancer Res; 70(1) January 1, 2010 267
h. r 14, 2015. © 2010 American Association for Cancer

Baryawno et al.
268
Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578
a LEF/TCF reporter kit, including negative and positivecontrols (SA Biosciences) and/or the full-length cDNA of β-catenin (pSPORTβ-cat; Open Biosystems) using Lipofectamine2000 (Invitrogen) for 48 h. Cells were then treated for 6 h with3 μmol/L OSU03012, and reporter gene activity was assessedusing a Dual-Luciferase Reporter Assay System (PromegaBiotech AB). GSK-3β knockdown was achieved using theSignalSilence GSK-3α/β kit (Cell Signaling) according tothe manufacturer's instructions.Xenografts and in vivo administration of OSU03012
and CCI-779. Four- to 6-wk-old female NMRI nu/nu micewere s.c. injected with 7 × 106 human D283 MED medullo-blastoma cells. Mice were randomly assigned into six treat-ment groups (eight mice in each treatment group) and thedrugs were given in daily doses as follows: (a) 7 mg/kgOSU03012 or (b) 70 mg/kg OSU03012 orally via a gastric feed-ing tube, (c) 20 mg/kg CCI-779 i.p., (d) 7 mg/kg OSU03012and 20 mg/kg CCI-779 i.p., (e) 70 mg/kg OSU03012 and20 mg/kg CCI-779 i.p., or (f) no treatment. Each mouse wastreated for 8 d and treatment was started on the appearanceof palpable tumors (mean, 0.13 mL). All animal experimentswere approved by the regional ethics committee for animalresearch (N234-05) in accordance with national regulations(SFS 1988:534, SFS 1988:539, and SFS 1988:541).Statistical analysis. Calculation of EC50 values and testing
for synergistic or additive effects of combination therapywere performed as previously described (15, 17). Mann-Whitney U test and the Kruskal-Wallis test (nonparametricANOVA) followed by Dunn's multiple comparison test wereused for analysis of statistical differences between two andseveral independent populations, respectively. All statisticaltests were two-sided.
Results
Key molecules in the PI3K/Akt signaling pathway areconstitutively activated in primary medulloblastomas. Byimmunohistochemical analysis of primary human medul-loblastoma and sPNET tumor samples (SupplementaryTable S1), specific phosphorylation of PDK1Ser241, AktThr308,and AktSer473 was detected in the cytoplasm of all primarytumor samples (Fig. 1). Interestingly, GSK-3β was also phos-phorylated Ser9 in all primary tumor samples analyzed(Fig. 1), whereas nuclear accumulation of β-catenin was de-tected in 27% of the cases. Thirty-one percent of the sampleshad mixed cytoplasmic/nuclear β-catenin staining (Fig. 1;Supplementary Table S1).Inhibition of PDK1 using the small-molecule inhibitor
OSU03012 potently suppresses medulloblastoma cell pro-liferation and clonogenic capacity. OSU03012, a third-generation celecoxib derivative, has been shown to affectPI3K/Akt signal transduction through inhibition of PDK1phosphorylation (18).As shown in Fig. 2A, treatment of medulloblastoma cells
with 3 μmol/L OSU03012 resulted in reduced phosphoryla-tion of PDK1Ser241, AktThr308, and AktSer473.All medulloblastoma cell lines showed a concentration-
dependent decrease in cell viability (Fig. 2B) and significant
Cancer Res; 70(1) January 1, 2010
Researcon Septembecancerres.aacrjournals.org Downloaded from
dose-dependent inhibition of colony formation (P < 0.001;Fig. 2C) after 48-hour exposure of OSU03012. Concentra-tions associated with 50% decrease in cell viability (biolog-ical EC50) ranged from 1.13 to 7.23 μmol/L (mean, 2.47μmol/L; Supplementary Table S2).OSU03012 induces S-phase arrest followed by apoptosis
in medulloblastoma cells. Cell cycle analysis revealed a pro-nounced S-phase arrest that was evident after both 24-hourand 48-hour incubation with 3 μmol/L OSU03012 in D283MED and D324 MED cells (see also Supplementary Fig.S1A–E). We also observed an accumulation of cells with a hy-podiploid DNA content (sub-G1). This was most pronouncedin MEB-MED-8A, D458 MED, and PFSK-1 cells incubatedwith 3 μmol/L OSU03012 (Supplementary Fig. S1C–E).Depolarization of the mitochondrial membrane potential
was detected in all medulloblastoma cell lines on treatmentwith OSU03012 (Supplementary Fig. S2A–E). Western blot-ting confirmed activation of caspase-9, caspase-3, and PARP,whereas no activation of BID was observed in any of the fivemedulloblastoma cell lines investigated (Fig. 2D).OSU03012 significantly augments the cytotoxic effect of
chemotherapeutic drugs in vitro. To further investigateOSU03012 as a potential drug in the treatment of medullo-blastoma, we studied the effects of OSU03012 in combinationwith mammalian target of rapamycin (mTOR) inhibitors orconventional chemotherapeutic drugs. D324 MED and D283MED cells were treated with increasing concentrations ofOSU03012, mTOR inhibitors, and chemotherapeutic drugs.The activities of the individual cytostatic drugs were deter-mined in initial experiments (data not shown). Fixed concen-tration ratios of the drugs were used with serial dilutions forcombination and single-drug treatments. The results aresummarized in Table 1, which shows the combination index(CI) at IC50. OSU03012 induced a synergistic or additive effectin D324 MED and D283 MED cells when used in combinationwith CCI-779 (IC50, 0.57–0.80; 0.78–0.94), rapamycin (IC50,0.57–0.83; 0.88–0.92), cyclophosphamide (IC50, 0.45–0.62;0.56–0.99), or doxorubicin (IC50, 0.97–1.12; 0.75–1.00). In con-trast, OSU03012 in combination with vincristine had an ad-ditive effect in D283 MED cells (IC50, 0.98–1.20), whereas anantagonistic effect was observed in D324 MED cells (IC50,1.01–1.35). OSU03012 in combination with temozolomidehad an antagonistic effect in both cell lines at the doses used(IC50, 1.18–1.38; 1.07–1.30).Furthermore, a combination of OSU03012 and the mTOR
inhibitor CCI-779 significantly augmented inhibition of theclonogenic capacity of D324 MED, PFSK-1, and UW228-3cells compared with single-drug treatment (P < 0.001; all celllines; Supplementary Fig. S3).Effects of OSU03012 on Akt downstream proteins in me-
dulloblastoma cells. The cell viability assays showed a syner-gistic induction of cell cytotoxicity when medulloblastomacells were treated with a combination of OSU03012 and themTOR inhibitors rapamycin or CCI-779 (Table 1). However, in-creased levels of AktSer473 phosphorylation were observed inbothD324MEDandD283MED cells treatedwith CCI-779. Thisactivation of Akt was inhibited with concomitant treatment ofthe cells with OSU03012 (Fig. 3A). Both OSU03012 and CCI-779
Cancer Research
h. r 14, 2015. © 2010 American Association for Cancer

Inhibitors of PI3K/Akt-Wnt/β-Catenin Cross-Talk
Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578
reduced the phosphorylation of the mTOR downstream tar-gets S6K1 and 4E-BP1 (Fig. 3B).OSU03012 has profound effects on the growth of estab-
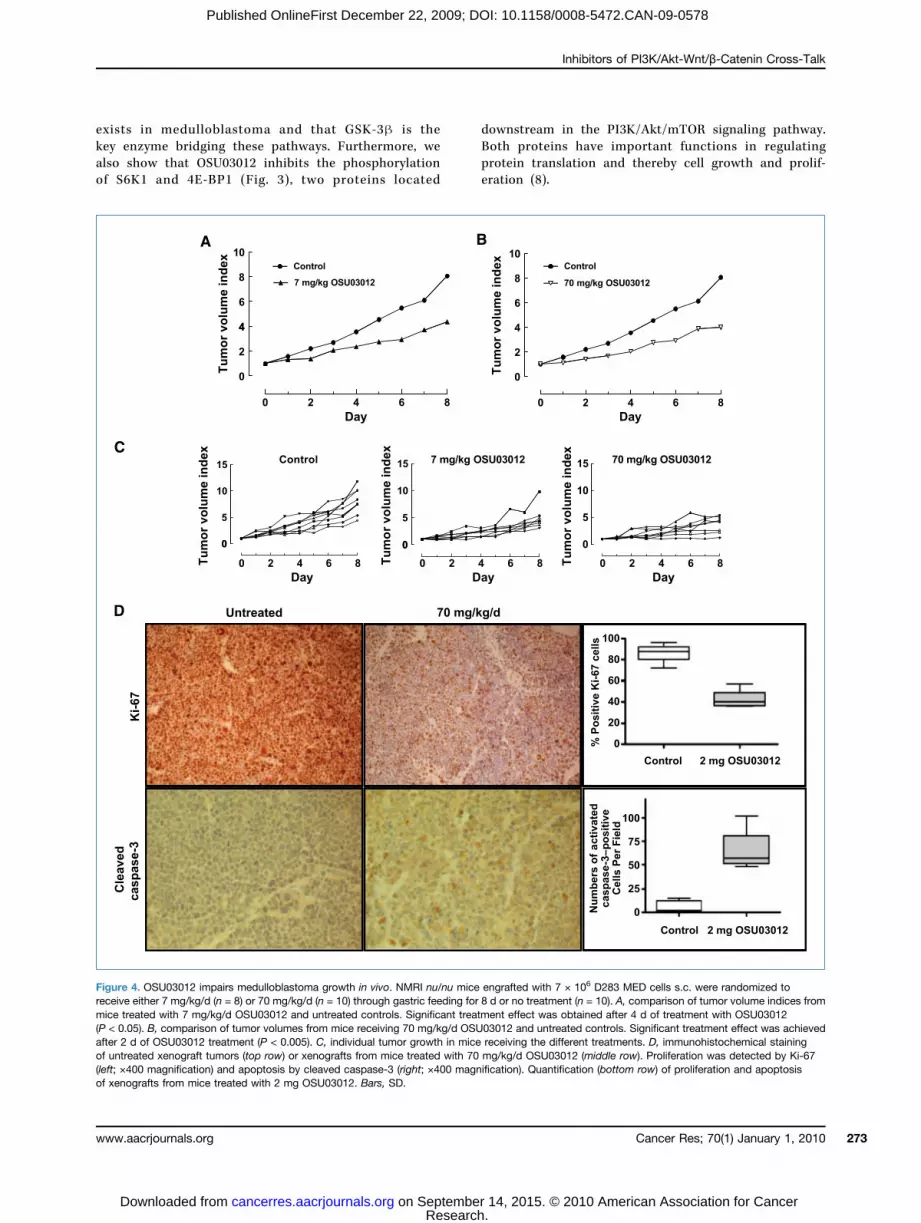
lished medulloblastoma xenografts. Athymic mice carryingestablished D283 MED xenografts were treated with either7 or 70 mg/kg of OSU03012 by daily oral gavages. Tumorgrowth was significantly inhibited after treatment for 2days with 70 mg/kg/d OSU03012 (P < 0.005) or 4 days with7 mg/kg/d OSU03012 (P < 0.05) compared with untreatedcontrols (Fig. 4A–C). At the end of treatment, tumor volumeswere reduced by 66% or 40% inmice receiving 70 or 7mg/kg/d,respectively, compared with controls. Significant elevatedactivated caspase-3 (P < 0.0001) expression and reducedexpression of the proliferation marker Ki-67 (P < 0.0001)were detected in tumors from mice treated with OSU03012(Fig. 4D).OSU03012 augments the effect of the mTOR inhibitor
CCI-779 on medulloblastoma growth in vivo. Because bothOSU03012 and CCI-779 inhibit components in the PI3K/Akt/mTOR signaling pathway and had synergistic cytotoxiceffects on medulloblastoma cells in vitro (Table 1), we ex-amined the in vivo activity of these drugs in combination.CCI-779 significantly inhibited the growth of establishedmedulloblastoma tumors from day 2 of treatment (P <0.05), and the tumor volume was reduced by 65% com-pared with controls at the end of treatment (Supplemen-tary Fig. S4A–C). However, CCI-779 in combination with 7
www.aacrjournals.org
Researcon Septembecancerres.aacrjournals.org Downloaded from
or 70 mg/kg/d of OSU03012 reduced the tumor growth by73% or 75%, respectively, compared with controls (Supple-mentary Fig. S4A and B).PDK1 inhibition by OSU03012 downregulates cyclin
D1 and c-Myc protein expression in medulloblastoma viaactivation of GSK-3β and inhibition of β-catenin function.Incubation of medulloblastoma cells with OSU03012 or thePI3K inhibitor LY294002 inhibited phosphorylation of GSK-3βSer9 (Fig. 5A; Supplementary Fig. S7).Subcellular protein fractionation and immunofluores-
cence cytology showed that medulloblastoma cell linescontained significant levels of β-catenin in the nucleus thatwas reduced in a dose-dependent manner on treatmentwith increasing concentrations of OSU03012 (Fig. 5B).OSU03012 induced β-catenin phosphorylation and reducedthe protein levels of c-Myc and cyclin D1 (Fig. 5A). Similarly,treatment of medulloblastoma cells with the PI3K inhibitorLY294002 also resulted in reduced expression of cyclin D1and c-Myc, whereas concomitant treatment of medulloblas-toma cells with OSU03012 and the mTOR inhibitor CCI-779resulted in an almost complete suppression of cyclin D1and c-Myc expression (Supplementary Figs. S6 and S7).OSU03012 significantly inhibited β-catenin–TCF/LEF trans-activation in D324 MED and D283 MED cells (P < 0.001;Fig. 5C). Forced overexpression of β-catenin by cotransfectionwith a β-catenin expression plasmid (pSPORTβ-cat) was alsoinhibited by OSU03012 (Fig. 5C).
Figure 1. Proteins in the PI3K/Akt signaling pathway are constitutively activated in primary medulloblastoma. Immunohistochemical analysis of primarymedulloblastoma tissue samples for phosphorylated PDK1Ser241, AktSer473, AktThr308, GSK-3βSer9, and β-catenin.
Cancer Res; 70(1) January 1, 2010 269
h. r 14, 2015. © 2010 American Association for Cancer

Baryawno et al.
270
Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578
To analyze the involvement of GSK-3β in the potentialcross-talk between PI3K/Akt signaling and nuclear translo-cation of β-catenin in medulloblastoma, we treated D283MED, D324 MED, and MEB-MED-8A (data not shown) cells
Cancer Res; 70(1) January 1, 2010
Researcon Septembecancerres.aacrjournals.org Downloaded from
with GSK-3β inhibitors or activators in combination withOSU03012. As shown in Fig. 5D, the GSK-3β inhibitorsCHIR99021 and SB-216763 significantly rescued medulloblas-toma cells from the toxic effects of OSU03012 (P < 0.001,
Figure 2. Inhibition of PDK1 signal transduction impairs the growth of medulloblastoma cells and induces apoptosis. A, OSU03012 reducesphosphorylation of PDK1Ser241, AktThr308, and AktSer473 in medulloblastoma cells. D324 MED, PFSK-1, D283 MED, and MEB-MED-8A were incubatedwith 3 μmol/L OSU03012 for 0.5, 6, or 24 h, and protein extracts were subjected to Western blot analysis to detect phosphorylated PDK1Ser241,AktThr308, AktSer473, and total Akt. β-Actin was used as a control for equal loading of samples. B, OSU03012 inhibits the growth of medulloblastoma cells.Medulloblastoma cells were treated with increasing concentrations of OSU03012 for 48 h, and cell survival was measured using an MTT assay. C,OSU03012 impairs the clonogenic capacity of medulloblastoma cells. D324 MED, PFSK-1, and UW228-3 cells were treated with 10, 100, or 500 nmol/Lof OSU03012 for 48 h. Cells were then incubated in drug-free medium for 10 d and colonies (>75 cells) with 50% plate efficiency were counted. D,medulloblastoma cells were treated with 3 μmol/L OSU03012 for 6 and 24 h, and protein extracts were subjected to Western blotting using antibodiesdetecting full-length and cleaved caspase-9, caspase-3, PARP, and BID. β-Actin was used to ensure equal protein loading.
Cancer Research
h. r 14, 2015. © 2010 American Association for Cancer

Inhibitors of PI3K/Akt-Wnt/β-Catenin Cross-Talk
Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578
CHIR99021; P < 0.001, SB-216763), whereas the GSK-3β acti-vator SNP significantly increased the toxic effects ofOSU03012 (P < 0.01). CHIR99021 and SB-216763 increasedGSK-3βSer9 phosphorylation and c-Myc protein expressionin OSU03012-treated medulloblastoma cells. SNP treatmentresulted in decreased GSK-3βSer9 phosphorylation and re-duced c-Myc expression (Supplementary Fig. S5A). Moreover,OSU03012 treatment of medulloblastoma cells resulted in in-creased binding of GSK-3β to axin1 compared with non-treated cells (Supplementary Fig. S5B). Similar smallinterfering RNA (siRNA) knockdown of GSK-3β inhibitedthe cytotoxic effect of OSU03012 and increased c-Myc ex-pression in D324 MED and D283 MED cells (Fig. 5D; Supple-mentary Fig. S5C).Finally, decreased phosphorylation of PDK1Ser241 and GSK-
3βSer9 and decreased levels of β-catenin, cyclin D1, andc-Myc proteins were observed in D283 MED xenograftstreated with OSU03012 (Supplementary Fig. S8).
Discussion
Recent studies indicate that numerous components of thePI3K/Akt signaling pathway are frequent targets for amplifi-cation, translocations, and mutations in cancer, resulting inactivation of the pathway (8). In medulloblastoma, activationof PI3K/Akt signaling seems to be a frequent event becausethe majority of primary medulloblastomas exhibit phosphor-ylation of AktSer473 independent of histologic staging (9, 19,20). In this study, we show that PDK1, a protein regulatingthe phosphorylation, and thereby Akt are constitutivelyactivated in primary medulloblastomas (Fig. 1). Moreover,GSK-3β, a protein that is constitutively active in restingcells (21), was found to be functionally inactivated by phos-phorylation in all of the primary medulloblastomas analyzed(Fig. 1). Similarly, phosphorylation of GSK-3βSer9 wasdetected by Western blot in three of six primary medullo-blastomas. However, no increased phosphorylation ofPDK1 and only weak activation of Akt were detected inthese samples (22).
www.aacrjournals.org
Researcon Septembecancerres.aacrjournals.org Downloaded from
Medulloblastoma cells exhibit increased expression ofseveral growth factor receptors that may transmit theirsignals through the PI3K/Akt signaling pathway, such asthe insulin-like growth factor-I (IGF-I) receptor (23), platelet-derived growth factor B (24, 25), tyrosine receptor kinaseB (26), the chemokine receptor (CXCR4; ref. 27), the epi-dermal growth factor (EGF) receptor (28), and c-KIT (29).The importance of PI3K/Akt signaling in cancer cells hasresulted in an intense search for agents that specificallyinhibit key components of this pathway. Several of thesecompounds show promising preclinical antitumor effects,and a few of them are already in clinical trials (30). Incontrast to PI3Ks or Akt for which several subunits orisoforms exist, only one single isoform of PDK1 has beenreported in humans. PDK1 has high specificity for phos-phorylation and activation of all three Akt isoforms(PBKα/Akt1, PBKβ/Akt2, and PBKγ/Akt3). PDK1 hypo-morphic mice display no deleterious phenotype, and theformation of PTEN-deficient tumors is abrogated whenPDK1 hypomorphic mice are crossed with PTEN heterozy-gote mice. Taken together, this makes PDK1 an attractivecandidate as an anticancer drug target (30, 31). Celecoxib,a drug designed to specifically inhibit the activity of cy-clooxygenase-2, has shown promising effects as an anti-cancer agent. One of the most prominent off-targeteffects of celecoxib is the specific inhibition of PDK1 ac-tivity possibly by competing with ATP for binding toPDK1 (32). OSU03012 was isolated by structure-based op-timization of celecoxib for compounds lacking cyclooxy-genase-2 activity but retaining PDK1-inhibiting activity(18). We have recently shown that celecoxib inhibitsmedulloblastoma growth both in vitro and in vivo (14).We therefore investigated the effects of OSU03012 on
medulloblastoma cells both at the molecular level andas a potential future drug in the treatment of this dis-ease. We observed that OSU03012 potently modulatedthe effect of key molecules in the PI3K/Akt signalingpathway (Fig. 2A). Cytotoxic effects of OSU03012 ona panel of medulloblastoma cell lines revealed thatOSU03012 was significantly more potent compared with
Table 1. Effect of OSU03012 in combination with chemotherapeutic drugs and mTOR inhibitors inmedulloblastoma cells
Combination
D324 MEDh. r 14, 2015. ©
D283 MED
n
Mean CI at IC50(95% confidence interval)
Effect n201
Mean CI at IC50
(95% confidence interval)
Cancer Res; 70(1) J
0 American Association fo
Effect
OSU03012 + CCI-779
6 0.69 (0.57–0.80) Synergistic 6 0.86 (0.78–0.94) Synergistic OSU03012 + rapamycin 6 0.70 (0.57–0.83) Synergistic 6 0.90 (0.88–0.92) Synergistic OSU03012 + vincristine 6 1.21 (1.01–1.35) Antagonistic 6 1.09 (0.98–1.20) Additive OSU03012 + doxorubicin 6 1.05 (0.97–1.12) Additive 6 0.87 (0.75–1.00) Additive OSU03012 + temozolomide 6 1.28 (1.18–1.38) Antagonistic 6 1.15 (1.07–1.30) Antagonistic OSU03012 + cyclophosphamide 6 0.53 (0.45–0.62) Synergistic 6 0.78 (0.56–0.99) Synergisticanuary 1, 2010 271
r Cancer

Baryawno et al.
272
Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578
the nonsteroidal anti-inflammatory drugs celecoxib ordiclofenac in inducing cytotoxicity of medulloblastomacells (Fig. 2B and C; Supplementary Table S2; ref. 14).The growth-inhibitory effect of OSU03012 was associatedwith induction of mitochondrial-dependent apoptosis ofmedulloblastoma cells (Fig. 2D; Supplementary Fig. S2).Cell cycle analysis showed an S-phase arrest in D283MED and D324 MED cells, whereas no evident S-phasearrest was observed for MEB-MED-8A, D458 MED, orPFSK-1 cells treated with OSU03012. The observed S-phase arrest may be due to OSU03012-mediated down-regulation of G1-S phase–specific cyclin D1 expression(Fig. 5A). Others have reported a combination of caspase-dependent and caspase-independent mechanisms forOSU03012-mediated induction of tumor cell death (33–35).Recently, OSU03012 was reported to induce autophagythrough accumulation of reactive oxygen species (ROS) inhepatocellular carcinoma (36). This is particularly interest-ing because Akt, in contrast to its ability to inhibit apopto-sis induced by chemotherapeutic agents, is not able toinhibit ROS-mediated apoptosis (37).Our results show that OSU03012 augmented the cytotoxic
effects of conventional chemotherapeutic drugs in a synergis-tic or additive manner (Table 1). Similarly, inhibition of thecatalytic PI3K p110α isoform enhances the effect of doxoru-bicin in medulloblastoma cells (38). Because the mTOR inhib-itor CCI-779 has been shown to have prominent effects onpreclinical medulloblastoma models and is currently under-going clinical testing in medulloblastoma patients (39, 40),we focused on this agent in more detail. A potential dilemmawhen designing anticancer therapies using agents that inhibitmTOR is that mTOR inhibitors have been shown to activateAkt through a feedback loop (15). However, our data showthat a combination of OSU03012 and CCI-779 inhibits Aktand acts synergistically to induce medulloblastoma cell cyto-toxicity in vitro (Table 1). Therefore, treatment with agents
Cancer Res; 70(1) January 1, 2010
Researcon Septembecancerres.aacrjournals.org Downloaded from
that simultaneously target key components in the PI3K/Akt/mTOR pathway may prove to be more efficacious.Nuclear transactivation and activity of β-catenin has been
directly related to signaling through theWnt pathway, and thiscoordinates a diverse array of developmental processes, in-cluding the proliferation and fate of neural progenitor cells(5). Wnt signaling has emerged as an important step in humanoncogenesis, and∼15% of sporadicmedulloblastomas containmutations in proteins of theWnt pathway (41–43). In contrast,∼25% of children with medulloblastoma have nucleopositiveβ-catenin (44). Clearly, there must be other mechanisms in-volved in regulating β-catenin activation. Indeed, it has beenshown that β-catenin–mediated transactivation of TCF/LEFcan be activated by growth factors such as EGF, hepatocytegrowth factor, IGF-I, IGF-II, and insulin (45–48).We found thattreatment of medulloblastoma cells with OSU03012 dimin-ished the amount of nuclear β-catenin and induced β-cateninphosphorylation of NH2-terminal serine and threonine resi-dues followed by decreased TCF/LEF transactivation. Treat-ment also reduced the expression of cyclin D1 and c-Myc(Fig. 5), two proteins that are important in medulloblastomatumorigenesis (49). Inhibition of nuclear β-catenin transloca-tion is likely to be due to the observed OSU03012-mediated in-hibition of Akt activity, leading to activation of GSK-3βfunction, because activated Akt has been shown to effectivelysuppress the role of GSK-3β in regulating the degradation ofβ-catenin (25, 50). This is supported by the findings that theGSK-3β activator SNP significantly increased the cytotoxic ef-fects of OSU03012, whereas the GSK-3β inhibitors CHIR99021and SB-216763 or siRNA knockdown of GSK-3β inhibitedOSU03012-mediated medulloblastoma cell death (Fig. 5D). At-tenuation of GSK-3β phosphorylation, growth, and survivalhas also been shown in medulloblastoma cells treated withthe IGF-I receptor inhibitor NVP-AEW541 (23).These results suggest that molecular cross-talk be-
tween the PI3K/Akt and the Wnt signaling pathways
Figure 3. PDK1 inhibition abolishes CCI-779–mediated increase of Akt phosphorylation and inactivates the function of S6K1 and 4E-BP1 inmedulloblastoma cells. A and B, D324 MED and D283 MED cells were incubated with 3 μmol/L OSU03012, 6 μmol/L CCI-779, or a combination of 3 μmol/LOSU03012 and 6 μmol/L CCI-779. Protein extracts were isolated after 6- and 24-h incubation and subjected to Western blotting using antibodies todetect phosphorylated AktSer473, total Akt, phosphorylated S6K1Thr389, and phosphorylated 4E-BP1Ser65. β-Actin was used to ensure equal protein loading.
Cancer Research
h. r 14, 2015. © 2010 American Association for Cancer
Inhibitors of PI3K/Akt-Wnt/β-Catenin Cross-Talk
Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578
exists in medulloblastoma and that GSK-3β is thekey enzyme bridging these pathways. Furthermore, wealso show that OSU03012 inhibits the phosphorylationof S6K1 and 4E-BP1 (Fig. 3), two proteins located
www.aacrjournals.org
Researcon Septembecancerres.aacrjournals.org Downloaded from
downstream in the PI3K/Akt/mTOR signaling pathway.Both proteins have important functions in regulatingprotein translation and thereby cell growth and prolif-eration (8).
Figure 4. OSU03012 impairs medulloblastoma growth in vivo. NMRI nu/nu mice engrafted with 7 × 106 D283 MED cells s.c. were randomized toreceive either 7 mg/kg/d (n = 8) or 70 mg/kg/d (n = 10) through gastric feeding for 8 d or no treatment (n = 10). A, comparison of tumor volume indices frommice treated with 7 mg/kg/d OSU03012 and untreated controls. Significant treatment effect was obtained after 4 d of treatment with OSU03012(P < 0.05). B, comparison of tumor volumes from mice receiving 70 mg/kg/d OSU03012 and untreated controls. Significant treatment effect was achievedafter 2 d of OSU03012 treatment (P < 0.005). C, individual tumor growth in mice receiving the different treatments. D, immunohistochemical stainingof untreated xenograft tumors (top row) or xenografts from mice treated with 70 mg/kg/d OSU03012 (middle row). Proliferation was detected by Ki-67(left; ×400 magnification) and apoptosis by cleaved caspase-3 (right; ×400 magnification). Quantification (bottom row) of proliferation and apoptosisof xenografts from mice treated with 2 mg OSU03012. Bars, SD.
Cancer Res; 70(1) January 1, 2010 273
h. r 14, 2015. © 2010 American Association for Cancer

Baryawno et al.
274
Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578
Because limited information about the in vivo antitu-morigenic effects of OSU03012 is available, we investigatedthe effects of OSU03012 on established human medullo-blastoma xenografts in nude mice. We found that
Cancer Res; 70(1) January 1, 2010
Researcon Septembecancerres.aacrjournals.org Downloaded from
OSU03012 effectively inhibited the growth of medulloblas-toma in vivo in a dose-dependent manner (Fig. 4A–C).This was accompanied by reduced proliferation and in-creased caspase-3 activity in OSU03012-treated tumors
Figure 5. PDK1 inhibition of medulloblastoma cells results in impaired β-catenin function and downregulation of cyclin D1 and c-Myc proteins. A, effect ofOSU03012 on proteins involved in Wnt pathway signaling in medulloblastoma cells. D324 MED, PFSK-1, D283 MED, and MEB-MED-8A medulloblastomacells were treated with 3 μmol/L OSU03012 for 0.5, 6, or 24 h. Immunoblotting was used to detect phospho-GSK-3βSer9, total GSK-3β, phospho-β-cateninSer33/37/Thr41, total β-catenin, cyclin D1, and c-Myc. β-Actin was used to ensure equal loading. B, PDK1 inhibition by OSU03012 prevents nucleartranslocation of β-catenin in medulloblastoma cells. D324 MED cells were incubated with increasing concentrations of OSU03012 for 6 or 24 h, andthe cellular location of β-catenin was detected using immunofluorescence (top) or Western blotting of subcellular fractionated protein extracts (bottom).Top, middle row, DAPI was used to enlighten the nucleus. C, OSU03012 suppresses β-catenin–mediated TCF/LEF transactivation. Comparison of TCF/LEFtransactivation activity in D324 MED and D283 MED medulloblastoma cells treated with OSU03012 or no treatment. Cotransfection with pSPORTβ-catinduced strong TCF/LEF transactivation, which is inhibited by OSU03012. D, effect of GSK-3β inhibitors, activators, and GSK-3β siRNA knockdown onOSU03012-mediated medulloblastoma cell toxicity. D324 MED and D283 MED cells were incubated with the GSK-3β activator SNP and the GSK-3βinhibitors CHIR99021 or SB-216763 as single treatment or in combination with OSU03012 for 48 h, and cell survival was measured using MTT assay. Cellstransfected with either scrambled or GSK-3β siRNA for 48 h were counted by trypan blue dye exclusion to evaluate cell viability.
Cancer Research
h. r 14, 2015. © 2010 American Association for Cancer

Inhibitors of PI3K/Akt-Wnt/β-Catenin Cross-Talk
Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578
(Fig. 4D). OSU03012 also augmented the antitumor effectof the mTOR inhibitor CCI-779 (Supplementary Fig.S4A–C).Taken together, our data show that molecular cross-talk
between two of the major molecular mechanisms control-ling embryonic development, the receptor tyrosine kinasePI3K/Akt signal transduction pathway and the canonicalWnt/β-catenin pathway, is important in medulloblastoma.We also show that small-molecule inhibitors targetingkey components in the PI3K/Akt signaling pathway mayrepresent an approach for the treatment of medulloblasto-ma and that the effect of OSU03012 should be furtherinvestigated in a clinical setting.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
www.aacrjournals.org
Researcon Septembecancerres.aacrjournals.org Downloaded from
Acknowledgments
We thank Lova Segerström, Helena Gleissman, AgnesRasmusson, Lotta Elfman, Anna-Maria Marino, and MikaelLerner for their assistance.
Grant Support
Swedish Children's Cancer Foundation, Swedish CancerSociety, Swedish Research Council, Märta and Gunnar VPhilipson Foundation, The Mary Béve Foundation,Dämman Foundation, and Norwegian Cancer Society.The costs of publication of this article were defrayed
in part by the payment of page charges. This article musttherefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.Received 2/13/09; revised 9/14/09; accepted 10/21/09;
published OnlineFirst 12/22/09.
References
1. Yang ZJ, Ellis T, Markant SL, et al. Medulloblastoma can be initiatedby deletion of Patched in lineage-restricted progenitors or stem cells.Cancer Cell 2008;14:135–45.
2. Schuller U, Heine VM, Mao J, et al. Acquisition of granule neuron pre-cursor identity is a critical determinant of progenitor cell competenceto form Shh-induced medulloblastoma. Cancer Cell 2008;14:123–34.
3. Hambardzumyan D, Becher OJ, Holland EC. Cancer stem cells andsurvival pathways. Cell Cycle 2008;7:1371–8.
4. Ille F, Sommer L. Wnt signaling: multiple functions in neural develop-ment. Cell Mol Life Sci 2005;62:1100–8.
5. Fodde R, Brabletz T. Wnt/β-catenin signaling in cancer stemnessand malignant behavior. Curr Opin Cell Biol 2007;19:150–8.
6. He TC, Sparks AB, Rago C, et al. Identification of c-MYC as a targetof the APC pathway. Science 1998;281:1509–12.
7. Tetsu O, McCormick F. β-Catenin regulates expression of cyclin D1in colon carcinoma cells. Nature 1999;398:422–6.
8. Cully M, You H, Levine AJ, Mak TW. Beyond PTEN mutations: thePI3K pathway as an integrator of multiple inputs during tumorigene-sis. Nat Rev Cancer 2006;6:184–92.
9. Hartmann W, Digon-Sontgerath B, Koch A, et al. Phosphatidylinosi-tol 3′-kinase/AKT signaling is activated in medulloblastoma cellproliferation and is associated with reduced expression of PTEN.Clin Cancer Res 2006;12:3019–27.
10. Manning BD, Cantley LC. AKT/PKB signaling: navigating down-stream. Cell 2007;129:1261–74.
11. Hambardzumyan D, Becher OJ, RosenblumMK, Pandolfi PP, Manova-Todorova K, Holland EC. PI3K pathway regulates survival of cancerstem cells residing in the perivascular niche following radiation inmedulloblastoma in vivo. Genes Dev 2008;22:436–48.
12. Johnsen JI, Kogner P, Albihn A, Henriksson MA. Embryonal neuraltumours and cell death. Apoptosis 2009;14:424–38.
13. Li J, Yen C, Liaw D, et al. PTEN, a putative protein tyrosine phospha-tase gene mutated in human brain, breast, and prostate cancer.Science 1997;275:1943–7.
14. Baryawno N, Sveinbjornsson B, Eksborg S, et al. Tumor-growth-promoting cyclooxygenase-2 prostaglandin E2 pathwayprovides medulloblastoma therapeutic targets. Neuro-oncol2008;10:661–74.
15. Johnsen JI, Segerstrom L, Orrego A, et al. Inhibitors of mammaliantarget of rapamycin downregulate MYCN protein expression andinhibit neuroblastoma growth in vitro and in vivo. Oncogene 2008.
16. Johnsen JI, Lindskog M, Ponthan F, et al. Cyclooxygenase-2 isexpressed in neuroblastoma, and nonsteroidal anti-inflammatorydrugs induce apoptosis and inhibit tumor growth in vivo. CancerRes 2004;64:7210–5.
17. Ponthan F, Wickstrom M, Gleissman H, et al. Celecoxib preventsneuroblastoma tumor development and potentiates the effect ofchemotherapeutic drugs in vitro and in vivo. Clin Cancer Res 2007;13:1036–44.
18. Zhu J, Huang JW, Tseng PH, et al. From the cyclooxygenase-2 in-hibitor celecoxib to a novel class of 3-phosphoinositide-dependentprotein kinase-1 inhibitors. Cancer Res 2004;64:4309–18.
19. Rao G, Pedone CA, Valle LD, Reiss K, Holland EC, Fults DW. Sonichedgehog and insulin-like growth factor signaling synergize toinduce medulloblastoma formation from nestin-expressing neuralprogenitors in mice. Oncogene 2004;23:6156–62.
20. Del Valle L,WangJY, LassakA, et al. Insulin-like growth factor I receptorsignaling system in JC virus T antigen-induced primitive neuroectoder-mal tumors-medulloblastomas. J Neurovirol 2002;8Suppl 2:138–47.
21. Cohen P, Frame S. The renaissance of GSK3. Nat Rev Mol Cell Biol2001;2:769–76.
22. Wlodarski PK, Boszczyk A, Grajkowska W, Roszkowski M, JozwiakJ. Implication of active Erk in the classic type of human medulloblas-toma. Folia Neuropathol 2008;46:117–22.
23. Urbanska K, Trojanek J, Del Valle L, et al. Inhibition of IGF-I receptorin anchorage-independence attenuates GSK-3β constitutive phos-phorylation and compromises growth and survival of medulloblasto-ma cell lines. Oncogene 2007;26:2308–17.
24. Gilbertson RJ, Clifford SC. PDGFRB is overexpressed in metastaticmedulloblastoma. Nat Genet 2003;35:197–8.
25. MacDonald TJ, Brown KM, LaFleur B, et al. Expression profiling ofmedulloblastoma: PDGFRA and the RAS/MAPK pathway as thera-peutic targets for metastatic disease. Nat Genet 2001;29:143–52.
26. Washiyama K, Muragaki Y, Rorke LB, et al. Neurotrophin and neuro-trophin receptor proteins in medulloblastomas and other primitiveneuroectodermal tumors of the pediatric central nervous system.Am J Pathol 1996;148:929–40.
27. Barbero S, Bajetto A, Bonavia R, et al. Expression of the chemokinereceptor CXCR4 and its ligand stromal cell-derived factor 1 in humanbrain tumors and their involvement in glial proliferation in vitro. Ann NY Acad Sci 2002;973:60–9.
28. Ray A, Ho M, Ma J, et al. A clinicobiological model predicting survivalin medulloblastoma. Clin Cancer Res 2004;10:7613–20.
29. Chilton-Macneill S, Ho M, Hawkins C, Gassas A, Zielenska M,Baruchel S. C-kit expression and mutational analysis in medullo-blastoma. Pediatr Dev Pathol 2004;7:493–8.
30. Garcia-Echeverria C, Sellers WR. Drug discovery approachestargeting the PI3K/Akt pathway in cancer. Oncogene 2008;27:5511–26.
31. Lawlor MA, Mora A, Ashby PR, et al. Essential role of PDK1 in
Cancer Res; 70(1) January 1, 2010 275
h. r 14, 2015. © 2010 American Association for Cancer

Baryawno et al.
276
Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578
regulating cell size and development in mice. EMBO J 2002;21:3728–38.
32. Grosch S, Maier TJ, Schiffmann S, Geisslinger G. Cyclooxygenase-2(COX-2)-independent anticarcinogenic effects of selective COX-2 in-hibitors. J Natl Cancer Inst 2006;98:736–47.
33. Johnson AJ, Smith LL, Zhu J, et al. A novel celecoxib derivative,OSU03012, induces cytotoxicity in primary CLL cells and trans-formed B-cell lymphoma cell line via a caspase- and Bcl-2-independentmechanism. Blood 2005;105:2504–9.
34. Yacoub A, Park MA, Hanna D, et al. OSU-03012 promotes caspase-independent but PERK-, cathepsin B-, BID-, and AIF-dependent kill-ing of transformed cells. Mol Pharmacol 2006;70:589–603.
35. Zhang S, Suvannasankha A, Crean CD, et al. OSU-03012, a novelcelecoxib derivative, is cytotoxic to myeloma cells and acts throughmultiple mechanisms. Clin Cancer Res 2007;13:4750–8.
36. Gao M, Yeh PY, Lu YS, et al. OSU-03012, a novel celecoxib deriva-tive, induces reactive oxygen species-related autophagy in hepato-cellular carcinoma. Cancer Res 2008;68:9348–57.
37. Nogueira V, Park Y, Chen CC, et al. Akt determines replicativesenescence and oxidative or oncogenic premature senescenceand sensitizes cells to oxidative apoptosis. Cancer Cell 2008;14:458–70.
38. Guerreiro AS, Fattet S, Fischer B, et al. Targeting the PI3K p110αisoform inhibits medulloblastoma proliferation, chemoresistance,and migration. Clin Cancer Res 2008;14:6761–9.
39. Fouladi M, Laningham F, Wu J, et al. Phase I study of everolimus inpediatric patients with refractory solid tumors. J Clin Oncol 2007;25:4806–12.
40. Geoerger B, Kerr K, Tang CB, et al. Antitumor activity of the rapamy-cin analog CCI-779 in human primitive neuroectodermal tumor/
Cancer Res; 70(1) January 1, 2010
Researcon Septembecancerres.aacrjournals.org Downloaded from
medulloblastoma models as single agent and in combination che-motherapy. Cancer Res 2001;61:1527–32.
41. Baeza N, Masuoka J, Kleihues P, Ohgaki H. AXIN1 mutations but notdeletions in cerebellar medulloblastomas. Oncogene 2003;22:632–6.
42. Eberhart CG, Tihan T, Burger PC. Nuclear localization and mutationof β-catenin in medulloblastomas. J Neuropathol Exp Neurol 2000;59:333–7.
43. Huang H, Mahler-Araujo BM, Sankila A, et al. APC mutations in spo-radic medulloblastomas. Am J Pathol 2000;156:433–7.
44. Ellison DW, Onilude OE, Lindsey JC, et al. β-Catenin status predictsa favorable outcome in childhood medulloblastoma: the United King-dom Children's Cancer Study Group Brain Tumour Committee. J ClinOncol 2005;23:7951–7.
45. Desbois-Mouthon C, Cadoret A, Blivet-Van Eggelpoel MJ, et al. In-sulin and IGF-1 stimulate the β-catenin pathway through two signal-ling cascades involving GSK-3β inhibition and Ras activation.Oncogene 2001;20:252–9.
46. Lu Z, Ghosh S, Wang Z, Hunter T. Downregulation of caveolin-1function by EGF leads to the loss of E-cadherin, increased transcrip-tional activity of β-catenin, and enhanced tumor cell invasion. CancerCell 2003;4:499–515.
47. Morali OG, Delmas V, Moore R, Jeanney C, Thiery JP, Larue L. IGF-IIinduces rapid β-catenin relocation to the nucleus during epitheliumto mesenchyme transition. Oncogene 2001;20:4942–50.
48. Polakis P. Wnt signaling and cancer. Genes Dev 2000;14:1837–51.49. Gilbertson RJ. Medulloblastoma: signalling a change in treatment.
Lancet Oncol 2004;5:209–18.50. Mulholland DJ, Dedhar S, Wu H, Nelson CC. PTEN and GSK3β: key
regulators of progression to androgen-independent prostate cancer.Oncogene 2006;25:329–37.
Cancer Research
h. r 14, 2015. © 2010 American Association for Cancer

Correction
Correction: Small-Molecule Inhibitors ofPhosphatidylinositol 3-Kinase/Akt SignalingInhibit Wnt/β-Catenin Pathway Cross-Talkand Suppress Medulloblastoma Growth
In this article (Cancer Res 2010;70:266–76), which was published in the January 1,2010 issue of Cancer Research (1), there are a number of errors. There are errorsin the third and fourth sentences of the abstract; the sentences should read asfollows:“Constitutive activation of phosphoinositide-dependent protein kinase 1
(PDK1), Akt, and phosphorylation of glycogen synthase kinase 3β (GSK-3β)was detected by immunohistochemistry in all primary medulloblastomas exam-ined (n = 41). Small-molecule inhibitors targeting the PI3K/Akt signalingpathway affected β-catenin signaling by activation of GSK-3β, resulting in cyto-plasmic retention of β-catenin and reduced expression of its target genes cyclinD1 and c-Myc.”In the final sentence of the fourth paragraph of the Results section, “median”
should appear instead of “mean”. In the paragraph “PDK1 inhibition by OSU03012downregulates cyclin D1 and c-Myc protein expression in medulloblastoma viaactivation of GSK-3β and inhibition of β-catenin function” on page 269, theSupplementary Figure referenced should be S6, not S7.The first two sentences of the Fig. 4 legend should read as follows:“OSU03012 impairs medulloblastoma growth in vivo. NMRI nu/nu mice engrafted
with 7 × 106 D283 MED cells s.c. were randomized to receive either 7 mg/kg/d (n = 8)or 70 mg/kg/d (n = 8) through gastric feeding for 8 d or no treatment (n = 8). A,comparison of tumor volume indexes from mice treated with 7 mg/kg/dOSU03012 and untreated controls.”The legend for Fig. 4D should read as follows:“D, immunohistochemical staining of untreated xenograft tumors (left column)
or xenografts from mice treated with 70 mg/kg/d OSU03012 (middle column).Proliferation was detected by Ki-67 (top row; ×400 magnification) and apoptosisby cleaved caspase-3 (bottom row; ×400 magnification). Quantification (rightcolumn) of proliferation and apoptosis of xenografts from mice treated with2 mg OSU03012. Bars, SD.”Finally, there are errors in Figs. 3 and 5D; the corrected figures appear below.
CancerResearch
Figure 3.
Cancer Res; 70(5) March 1, 20102138

Reference1. Baryawno N, Sveinbjörnsson B, Eksborg S, Chen C-S, Kogner P, Johnsen JI. Small-molecule
inhibitors of phosphatidylinositol 3-kinase/Akt signaling inhibit Wnt/β-catenin pathway cross-talkand suppress medulloblastoma growth. Cancer Res 2010;70:266–76.
©2010 American Association for Cancer Research.doi: 10.1158/0008-5472.CAN-10-0016
Figure 5D.
Published OnlineFirst 02/23/2010.©2010 American Association for Cancer Research.doi: 10.1158/0008-5472.CAN-10-0016
Cancer Res; 70(5) March 1, 2010www.aacrjournals.org 2139
Correction

2010;70:266-276. Published OnlineFirst December 22, 2009.Cancer Res Ninib Baryawno, Baldur Sveinbjörnsson, Staffan Eksborg, et al. Suppress Medulloblastoma Growth
-Catenin Pathway Cross-Talk andβSignaling Inhibit Wnt/Small-Molecule Inhibitors of Phosphatidylinositol 3-Kinase/Akt
Updated version
10.1158/0008-5472.CAN-09-0578doi:
Access the most recent version of this article at:
Material
Supplementary
http://cancerres.aacrjournals.org/content/suppl/2009/12/22/0008-5472.CAN-09-0578.DC1.html
Access the most recent supplemental material at:
Cited articles
http://cancerres.aacrjournals.org/content/70/1/266.full.html#ref-list-1
This article cites 49 articles, 20 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/70/1/266.full.html#related-urls
This article has been cited by 14 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on September 14, 2015. © 2010 American Association for Cancercancerres.aacrjournals.org Downloaded from
Published OnlineFirst December 22, 2009; DOI: 10.1158/0008-5472.CAN-09-0578





























