Embed Size (px)
Citation preview

a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 2 2 – 3 1
Soil predatory mite communities (Acari: Gamasina) inagroecosystems of Central Argentina
Jose Camilo Bedano a,*, Andrea Ruf b
aDepartamento de Geologıa, Universidad Nacional de Rıo Cuarto, Ruta 36, Km. 601, X5804 BYA Rıo Cuarto, Cordoba, ArgentinabDepartment of Ecology, University of Bremen, FB 2, UFT, Leobenerstrasse, Postbox 330440, D-28334 Bremen, Germany
a r t i c l e i n f o
Article history:
Received 19 April 2006
Received in revised form
3 October 2006
Accepted 21 November 2006
Keywords:
Gamasina
Acari
Agroecosystems
Disturbances
Land management
Argentina
a b s t r a c t
The objective of this study was to compare soil predatory mite communities over a gradient
of land use intensity in agroecosystems of Central Argentina. The study was conducted at La
Colacha basin, Cordoba, Argentina on coarse-loamy, illitic, thermic Typic Hapludoll. Four
sites with different management systems (natural grassland, cattle raising, arable and
mixed), but with the same Soil Series, similar geomorphological characteristics and the
same land use history until 50 years before, were sampled at four sampling dates. Thirty-
eight species, 19 genera and nine families were identified. Ninety-two percent of the
identified species are new to science. This shows that the taxonomic knowledge of Gama-
sina is poorly developed in Argentina. In this study, Gamasina community structure was
clearly influenced by the management type. This confirms our hypothesis that Gamasina
community structure significantly changes as material and energy inputs and mechanical
perturbations in the system increased from the natural soil to the conventional agricultural
sites. The effects of agricultural practices could be observed in the occurrence of the species,
the diversity and the dominance structure of the community, the maturity index, and also in
seven similarity measures and the multivariate canonical correspondence analysis (CCA).
Species number and diversity measures were highest in the cattle raising plot and very low
in the natural and arable site. So, in terms of species number, the arable field did not have an
impoverished community in comparison with the natural undisturbed grassland.
# 2006 Elsevier B.V. All rights reserved.
avai lable at www.sc iencedi rec t .com
journal homepage: www.e lsev ier .com/ locate /apsoi l
1. Introduction
Gamasina mites live in a wide range of terrestrial ecosystems
under very different environmental conditions. Most are top
predators and occupy a central position in the soil food web
(Koehler, 1999; Walter and Proctor, 1999). Thus, communities
have a significant role in regulating decomposition and
nutrient cycling since they influence population growth of
other organisms (Ruf, 1997; Koehler, 1999).
Soil Gamasina communities are sensitive to changes in
management practices and the type of land use is an especially
critical parameter (Buryn and Hartmann, 1992; Ruf and Beck,
* Corresponding author. Tel.: +54 358 4676198; fax: +54 358 4676200.E-mail address: [email protected] (J.C. Bedano).
0929-1393/$ – see front matter # 2006 Elsevier B.V. All rights reservedoi:10.1016/j.apsoil.2006.11.008
2005). It has been suggested that agricultural lands have an
impoverished Gamasina fauna compared with natural habitats
(Karg, 1967; Lagerlof and Andren, 1988; Koehler, 1999). Arable
lands inEuropeare dominated byfew opportunistic species that
are able to resist ploughing and other disturbances or can
rapidly recolonise disturbed habitats (Bolger and Curry, 1984;
Larink, 1997; Koehler, 1999; Kovac et al., 1999). However, the
predatory mite community of arable fields is generally not
reliably predictable due to the very small numbers of species
and individuals (Ruf and Beck, 2005). Additionally, data on soil
Gamasina in agricultural fields is largely lacking (Ruf and Beck,
2005; Beaulieu and Weeks, in press).
d.

a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 2 2 – 3 1 23
In Argentina there is neither information on Gamasina
community structure nor on the disturbance to the commu-
nities produced by agricultural practices in agroecosystems.
The objective of this study was to compare soil predatory mite
communities over a gradient of land use intensity in
agroecosystems of Central Argentina. We hypothesized that
as material and energy inputs (fertilizer and herbicide) and
mechanical perturbations in the system increased from zero
in a natural soil to elevated levels in conventional agricultural
sites, Gamasina community structure would significantly
change. We expect a shift from K- to r-selected species and
a more uneven dominance structure.
2. Material and methods
2.1. Site description
The study sites were located in La Colacha basin (Fig. 1),
Cordoba, Argentina (648390 and 648500W, and 328540 and
338030S) on soils developed from eolic sediments of Laguna
Oscura formation (Cantu, 1998). The soil was classified as
coarse-loamy, illitic, thermic Typic Hapludoll (Cantu, 1998)
following the USDA classification (Soil Survey Staff, 1998). The
climate is continental with annual rainfall of 800 mm and
mean annual temperature of 16.5 8C.
We selected four sites with the same Soil Series (according
to FAO and Soil Taxonomy classifications), which were in close
proximity to one another in an area of about 50 km2 and with
similar geomorphological characteristics: slope 1–2% and
elevation 700 m a.s.l. The four sites had the same land use
history until 50 years before our study (natural grasslands).
Since 1950, the land property was divided and agriculture was
started in the area (Cantu, 1998). In this area, we selected four
sites with different management systems: natural, cattle
raising, mixed and arable. The natural site (approximately
Fig. 1 – Map of the study area. La Colacha basin
5 ha) has been covered with natural pastures (dominated by
Stipa sp.) for the last 50 years. The cattle raising site
(approximately 45 ha) had been devoted to cattle raising with
intermediate livestock grazing pressure during the last 40
years. It has been seeded with alfalfa (Medicago sativa L.) that
had not been ploughed for the last 4 years.
The site managed under a mixed production system
(approximately 40 ha) had annual rotations of maize (Zea
mays L.) and sunflower (Helianthus annuus L.) crops and
pastures devoted to cattle grazing. It was managed using
conventional agricultural practices (including the application
of chemical fertilizers and herbicides). In October 1999, the
plot was chisel plowed and maize was planted, and then
harvested in August 2000. In February 2001, the plot was
plowed with a moldboard plow and in April oats (Avena sativa
L.) and clover (Trifolium sp.) were seeded without previous
herbicide treatment.
The conventional arable site (approximately 40 ha) has
been cultivated with peanut (Arachis sp.), maize and sunflower
for four decades using chemical fertilizers and herbicides and
conventional agricultural practices. In late 1999 maize was
planted, but due to a suspected low yield it was used as pasture
for cattle. In April 2000, oats and melilotus (Melilotus sp.)
pasture were sown.
2.2. Sampling, extraction and identification
Soil Gamasina were sampled four times: April and December
2000 and February and June 2001. On each sampling date, at
each of the four sites, we took six undisturbed soil cores (10 cm
diameter and 10 cm depth) using a hard plastic core sampler,
obtaining a total of 96 samples. Samples were randomly taken
in an area of approximately 1 ha within each site.
Mites were extracted for 10 days with a Berlese–Tullgren
apparatus (Southwood, 1980) and stored in a solution of 70%
alcohol. Specimens were mounted in permanent (using
, Rıo Cuarto, Cordoba Province, Argentina.

a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 2 2 – 3 124
Hoyer’s medium) and semi permanent (using lactic acid) slides
and were identified at the species level using the taxonomic
keys of Hirschmann (1960), Bregetova et al. (1977), Evans and
Till (1979), Hyatt (1980), Krantz and Ainscough (1990) and Karg
(1993).
2.3. Data analysis
2.3.1. Community parametersSix indices were computed for the predatory community in
each site: (1) species richness (S); (2) Simpson diversity index
(D); (3) Shannon index of diversity (H); (4) alpha diversity; (5)
evenness (J); (6) maturity index (Ruf, 1998) (MI). Species
richness: number of species present in each site. Simpson
diversity index (Simpson, 1949): D = 1 �P
( pi)2. Shannon–
Weaver (Shannon and Weaver, 1949): H = �P
i[pi log( pi)],
where pi = ni/N; ni is the number of individuals of the ith
species and N is the total number of individuals. Alpha
diversity index (Fisher et al., 1943). Evenness index: J = H/ln S,
where S is the number of species. Maturity index: weighted
mean of the r- and K-values assigned to the Gamasina families
(Ruf, 1997) as follows:
MI ¼PS
i¼1 KiPSi¼1 Ki þ
PSi¼1 ri
where S is the species number, K the K-value for the family of
species i, and r is the r-value for the family of species i.
2.3.2. Similarity measuresThe following similarity measures were evaluated:
Sample similarity measures for presence/absence data:
Jaccard coefficient:
SJac ¼a
aþ bþ c
Sorensen coefficient:
SSor ¼2a
2aþ bþ c
Sample similarity measures for abundance data:
Canberra metric:
dCanb ¼Xpj¼1
jx1 j � x2 jjx1 j þ x2 j
Euclidean distance:
dED ¼
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiXpj¼1
ðx1 j þ x2 jÞ2vuut
Bray–Curtis distance:
dBC ¼Pp
j¼1 jx1 j � x2 jjPpj¼1 x1 j þ x2 j
¼Pp
j¼1 jx1 j � x2 jjx1þ þ x2þ
Chi-square distance:
dx2 ¼
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiXpj¼1
1xþ j=xþþ
x1 j
x1þ�
x2 j
x2þ
� �2vuut
Manhattan metric:
dMan ¼Xpj¼1
jx1 j � x2 jj
In all cases: a is the number of species present in sample 1
only, b the number of species present in sample 2 only, c the
number of species present in both samples 1 and 2, X1j the
number of individuals of species j in sample 1, X2j the number
of individuals of species j in sample 2 and p is the number of
species in samples.
Similarity measures were computed using GINKGO soft-
ware (Universitat de Barcelona, 2005).
2.3.3. Multivariate approach
We used canonical correspondence analysis (CCA) (ter Braak,
1986) to simultaneously elucidate the main patterns of mite
community and environmental factors variations, and the
relationships of each of the species with respect to the
environmental variables (ter Braak, 1986; ter Braak and
Prentice, 1988). CCA was performed using CANOCO 4.53 (ter
Braak and Smilauer, 2004). We used the dominance data and
no other transformation was applied.
3. Results
3.1. Community composition
Thirty-eight Gamasina species were identified, only three of
them could be assigned to already existing morphospecies; 11
were considered to be distinct but closely related to European
species, and were denoted as affinis (aff.). Nineteen genera and
nine families were identified. Their abundance in each site is
given in Table 1.
The dominance structure of the community was clearly
different among the four sites. In the natural site Dendrolaelaps
aff. crassitarsalis was the dominant species (57%) and Hypoaspis
aff. angusta was the second dominant species, with 12%; the
other species occurred with less than 10% dominance. In soil
under cattle raising management also D. aff. crassitarsalis was
the most abundant predatory mite (26%) but three other
species were also important: Rhodacarus sp. (19%), Hypoaspis
sp. 1 (15%) and Asca aff. pini (14%). The mixed production
system was dominated by two species with 42 and 26%
dominance. The most dominant species was Protogamasellus
aff. primitivus (42%) followed by Hypoaspis aff. brevipellis (26%).
The arable site was dominated by six species with similar
numbers: A. aff. pini (18%), P. aff. primitivus (16%), Dendrolaelaps
sp. 1 (15%), Rhodacarus sp. (13%), Amblyseius aff. barkeri (12%)
and H. aff. brevipellis (11%).
Species from the families Halolaelapidae and Veigaiidae
were collected only from the natural soil while no phytoseiids
were observed in this site (Table 1).
The highest species richness was observed in the cattle
raising (24 spp.) and the lowest in the natural and arable site
(15 and 14 spp., respectively) (Table 1). Diversity was lowest
in the natural soil as measured by alpha, Simpson and
Shannon indices (Table 1). The highest values of alpha and
Shannon indices were in the cattle raising site and of Simpson
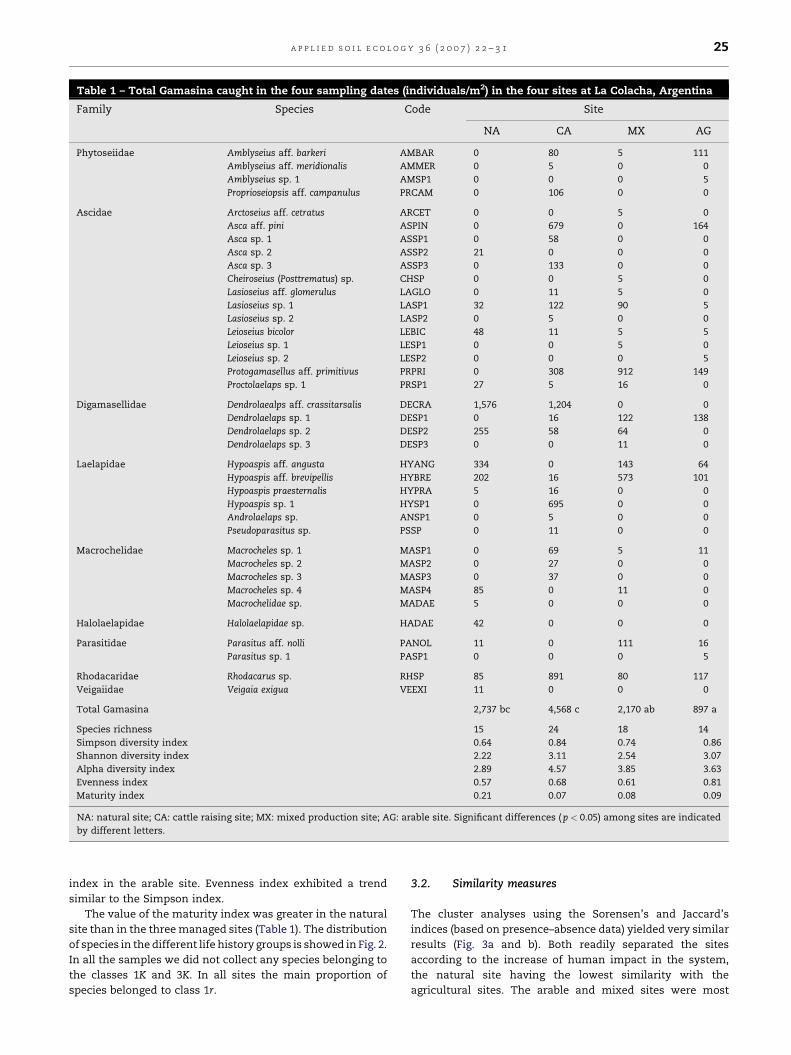
Table 1 – Total Gamasina caught in the four sampling dates (individuals/m2) in the four sites at La Colacha, Argentina
Family Species Code Site
NA CA MX AG
Phytoseiidae Amblyseius aff. barkeri AMBAR 0 80 5 111
Amblyseius aff. meridionalis AMMER 0 5 0 0
Amblyseius sp. 1 AMSP1 0 0 0 5
Proprioseiopsis aff. campanulus PRCAM 0 106 0 0
Ascidae Arctoseius aff. cetratus ARCET 0 0 5 0
Asca aff. pini ASPIN 0 679 0 164
Asca sp. 1 ASSP1 0 58 0 0
Asca sp. 2 ASSP2 21 0 0 0
Asca sp. 3 ASSP3 0 133 0 0
Cheiroseius (Posttrematus) sp. CHSP 0 0 5 0
Lasioseius aff. glomerulus LAGLO 0 11 5 0
Lasioseius sp. 1 LASP1 32 122 90 5
Lasioseius sp. 2 LASP2 0 5 0 0
Leioseius bicolor LEBIC 48 11 5 5
Leioseius sp. 1 LESP1 0 0 5 0
Leioseius sp. 2 LESP2 0 0 0 5
Protogamasellus aff. primitivus PRPRI 0 308 912 149
Proctolaelaps sp. 1 PRSP1 27 5 16 0
Digamasellidae Dendrolaealps aff. crassitarsalis DECRA 1,576 1,204 0 0
Dendrolaelaps sp. 1 DESP1 0 16 122 138
Dendrolaelaps sp. 2 DESP2 255 58 64 0
Dendrolaelaps sp. 3 DESP3 0 0 11 0
Laelapidae Hypoaspis aff. angusta HYANG 334 0 143 64
Hypoaspis aff. brevipellis HYBRE 202 16 573 101
Hypoaspis praesternalis HYPRA 5 16 0 0
Hypoaspis sp. 1 HYSP1 0 695 0 0
Androlaelaps sp. ANSP1 0 5 0 0
Pseudoparasitus sp. PSSP 0 11 0 0
Macrochelidae Macrocheles sp. 1 MASP1 0 69 5 11
Macrocheles sp. 2 MASP2 0 27 0 0
Macrocheles sp. 3 MASP3 0 37 0 0
Macrocheles sp. 4 MASP4 85 0 11 0
Macrochelidae sp. MADAE 5 0 0 0
Halolaelapidae Halolaelapidae sp. HADAE 42 0 0 0
Parasitidae Parasitus aff. nolli PANOL 11 0 111 16
Parasitus sp. 1 PASP1 0 0 0 5
Rhodacaridae Rhodacarus sp. RHSP 85 891 80 117
Veigaiidae Veigaia exigua VEEXI 11 0 0 0
Total Gamasina 2,737 bc 4,568 c 2,170 ab 897 a
Species richness 15 24 18 14
Simpson diversity index 0.64 0.84 0.74 0.86
Shannon diversity index 2.22 3.11 2.54 3.07
Alpha diversity index 2.89 4.57 3.85 3.63
Evenness index 0.57 0.68 0.61 0.81
Maturity index 0.21 0.07 0.08 0.09
NA: natural site; CA: cattle raising site; MX: mixed production site; AG: arable site. Significant differences ( p < 0.05) among sites are indicated
by different letters.
a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 2 2 – 3 1 25
index in the arable site. Evenness index exhibited a trend
similar to the Simpson index.
The value of the maturity index was greater in the natural
site than in the three managed sites (Table 1). The distribution
of species in the different life history groups is showed in Fig. 2.
In all the samples we did not collect any species belonging to
the classes 1K and 3K. In all sites the main proportion of
species belonged to class 1r.
3.2. Similarity measures
The cluster analyses using the Sorensen’s and Jaccard’s
indices (based on presence–absence data) yielded very similar
results (Fig. 3a and b). Both readily separated the sites
according to the increase of human impact in the system,
the natural site having the lowest similarity with the
agricultural sites. The arable and mixed sites were most
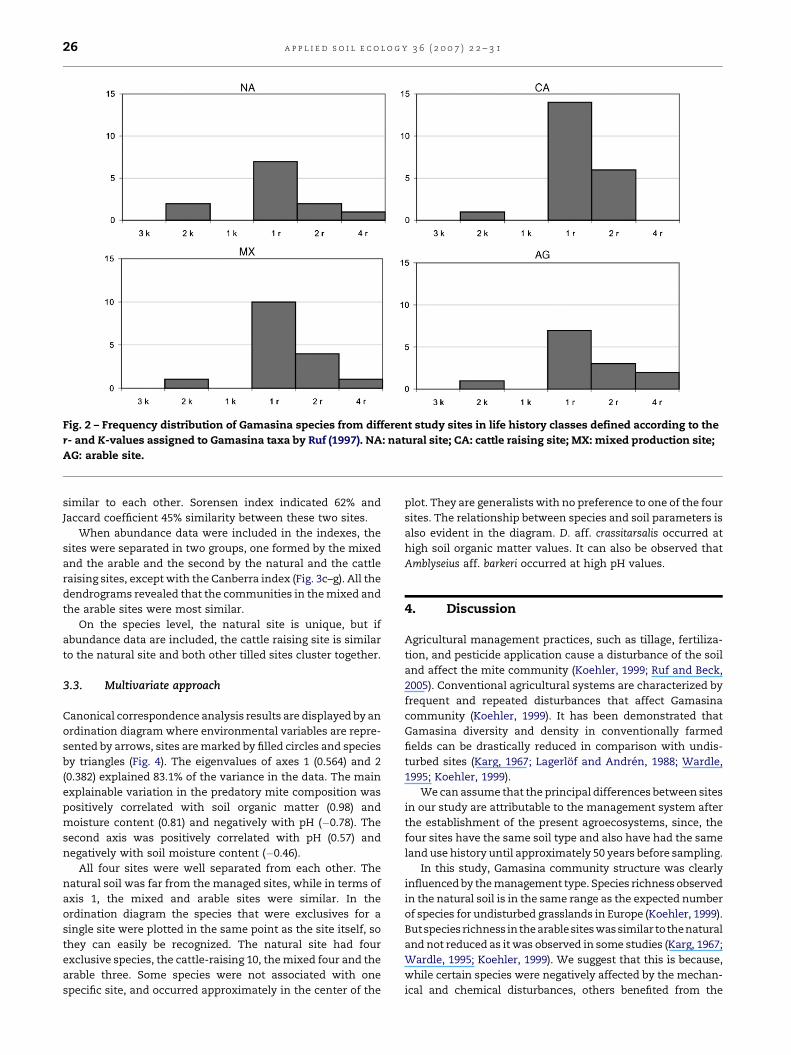
Fig. 2 – Frequency distribution of Gamasina species from different study sites in life history classes defined according to the
r- and K-values assigned to Gamasina taxa by Ruf (1997). NA: natural site; CA: cattle raising site; MX: mixed production site;
AG: arable site.
a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 2 2 – 3 126
similar to each other. Sorensen index indicated 62% and
Jaccard coefficient 45% similarity between these two sites.
When abundance data were included in the indexes, the
sites were separated in two groups, one formed by the mixed
and the arable and the second by the natural and the cattle
raising sites, except with the Canberra index (Fig. 3c–g). All the
dendrograms revealed that the communities in the mixed and
the arable sites were most similar.
On the species level, the natural site is unique, but if
abundance data are included, the cattle raising site is similar
to the natural site and both other tilled sites cluster together.
3.3. Multivariate approach
Canonical correspondence analysis results are displayed by an
ordination diagram where environmental variables are repre-
sented by arrows, sites are marked by filled circles and species
by triangles (Fig. 4). The eigenvalues of axes 1 (0.564) and 2
(0.382) explained 83.1% of the variance in the data. The main
explainable variation in the predatory mite composition was
positively correlated with soil organic matter (0.98) and
moisture content (0.81) and negatively with pH (�0.78). The
second axis was positively correlated with pH (0.57) and
negatively with soil moisture content (�0.46).
All four sites were well separated from each other. The
natural soil was far from the managed sites, while in terms of
axis 1, the mixed and arable sites were similar. In the
ordination diagram the species that were exclusives for a
single site were plotted in the same point as the site itself, so
they can easily be recognized. The natural site had four
exclusive species, the cattle-raising 10, the mixed four and the
arable three. Some species were not associated with one
specific site, and occurred approximately in the center of the
plot. They are generalists with no preference to one of the four
sites. The relationship between species and soil parameters is
also evident in the diagram. D. aff. crassitarsalis occurred at
high soil organic matter values. It can also be observed that
Amblyseius aff. barkeri occurred at high pH values.
4. Discussion
Agricultural management practices, such as tillage, fertiliza-
tion, and pesticide application cause a disturbance of the soil
and affect the mite community (Koehler, 1999; Ruf and Beck,
2005). Conventional agricultural systems are characterized by
frequent and repeated disturbances that affect Gamasina
community (Koehler, 1999). It has been demonstrated that
Gamasina diversity and density in conventionally farmed
fields can be drastically reduced in comparison with undis-
turbed sites (Karg, 1967; Lagerlof and Andren, 1988; Wardle,
1995; Koehler, 1999).
We can assume that the principal differences between sites
in our study are attributable to the management system after
the establishment of the present agroecosystems, since, the
four sites have the same soil type and also have had the same
land use history until approximately 50 years before sampling.
In this study, Gamasina community structure was clearly
influenced by the management type. Species richness observed
in the natural soil is in the same range as the expected number
of species for undisturbed grasslands in Europe (Koehler, 1999).
Butspecies richness inthe arablesites was similar to thenatural
and not reduced as it was observed in some studies (Karg, 1967;
Wardle, 1995; Koehler, 1999). We suggest that this is because,
while certain species were negatively affected by the mechan-
ical and chemical disturbances, others benefited from the

Fig. 3 – Clustering dendrograms of the four sites from La Colacha basin using ward linkage and different similarity
measures: (a) Jaccard; (b) Sorensen; (c) Canberra; (d) Euclidean; (e) Bray–Curtis; (f) Chi-square; (g) Manhattan. NA: natural site;
CA: cattle raising site; MX: mixed production site; AG: arable site.
a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 2 2 – 3 1 27
changes that these activities produce in the soil, like the
increase of the populations of some organisms that are prey of
predatory mites.
The high species richness and diversity observed in the
cattle raising system is thought to be related with the presence
of the cows and their activities. Many species are associated
with cattle dung some of them being phoretic on dung
dwelling insects (e.g. species of Macrochelidae and Ascidae)
(Karg, 1993; Krantz, 1998; Koehler, 1999). This particular
microhabitat could have allowed the establishment of some
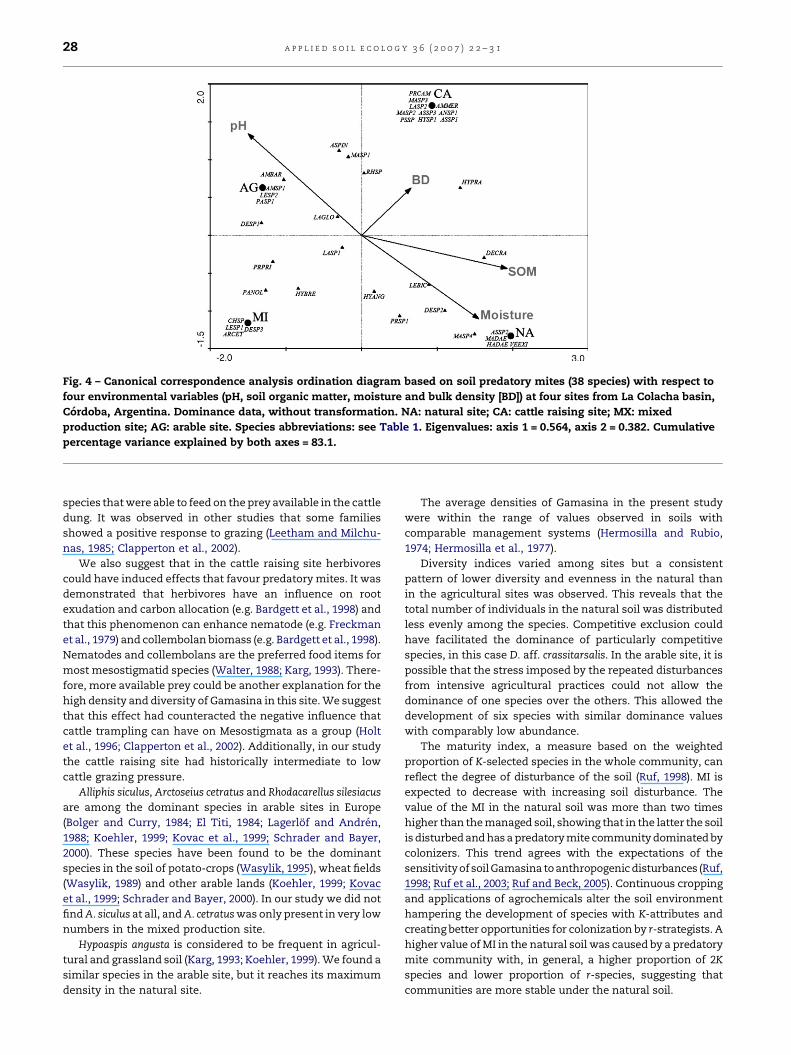
Fig. 4 – Canonical correspondence analysis ordination diagram based on soil predatory mites (38 species) with respect to
four environmental variables (pH, soil organic matter, moisture and bulk density [BD]) at four sites from La Colacha basin,
Cordoba, Argentina. Dominance data, without transformation. NA: natural site; CA: cattle raising site; MX: mixed
production site; AG: arable site. Species abbreviations: see Table 1. Eigenvalues: axis 1 = 0.564, axis 2 = 0.382. Cumulative
percentage variance explained by both axes = 83.1.
a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 2 2 – 3 128
species that were able to feed on the prey available in the cattle
dung. It was observed in other studies that some families
showed a positive response to grazing (Leetham and Milchu-
nas, 1985; Clapperton et al., 2002).
We also suggest that in the cattle raising site herbivores
could have induced effects that favour predatory mites. It was
demonstrated that herbivores have an influence on root
exudation and carbon allocation (e.g. Bardgett et al., 1998) and
that this phenomenon can enhance nematode (e.g. Freckman
et al., 1979) and collembolan biomass (e.g. Bardgett et al., 1998).
Nematodes and collembolans are the preferred food items for
most mesostigmatid species (Walter, 1988; Karg, 1993). There-
fore, more available prey could be another explanation for the
high density and diversity of Gamasina in this site. We suggest
that this effect had counteracted the negative influence that
cattle trampling can have on Mesostigmata as a group (Holt
et al., 1996; Clapperton et al., 2002). Additionally, in our study
the cattle raising site had historically intermediate to low
cattle grazing pressure.
Alliphis siculus, Arctoseius cetratus and Rhodacarellus silesiacus
are among the dominant species in arable sites in Europe
(Bolger and Curry, 1984; El Titi, 1984; Lagerlof and Andren,
1988; Koehler, 1999; Kovac et al., 1999; Schrader and Bayer,
2000). These species have been found to be the dominant
species in the soil of potato-crops (Wasylik, 1995), wheat fields
(Wasylik, 1989) and other arable lands (Koehler, 1999; Kovac
et al., 1999; Schrader and Bayer, 2000). In our study we did not
findA. siculus at all, andA. cetratuswas only present in very low
numbers in the mixed production site.
Hypoaspis angusta is considered to be frequent in agricul-
tural and grassland soil (Karg, 1993; Koehler, 1999). We found a
similar species in the arable site, but it reaches its maximum
density in the natural site.
The average densities of Gamasina in the present study
were within the range of values observed in soils with
comparable management systems (Hermosilla and Rubio,
1974; Hermosilla et al., 1977).
Diversity indices varied among sites but a consistent
pattern of lower diversity and evenness in the natural than
in the agricultural sites was observed. This reveals that the
total number of individuals in the natural soil was distributed
less evenly among the species. Competitive exclusion could
have facilitated the dominance of particularly competitive
species, in this case D. aff. crassitarsalis. In the arable site, it is
possible that the stress imposed by the repeated disturbances
from intensive agricultural practices could not allow the
dominance of one species over the others. This allowed the
development of six species with similar dominance values
with comparably low abundance.
The maturity index, a measure based on the weighted
proportion of K-selected species in the whole community, can
reflect the degree of disturbance of the soil (Ruf, 1998). MI is
expected to decrease with increasing soil disturbance. The
value of the MI in the natural soil was more than two times
higher than the managed soil, showing that in the latter the soil
is disturbed and has a predatory mite community dominated by
colonizers. This trend agrees with the expectations of the
sensitivity of soil Gamasina to anthropogenic disturbances (Ruf,
1998; Ruf et al., 2003; Ruf and Beck, 2005). Continuous cropping
and applications of agrochemicals alter the soil environment
hampering the development of species with K-attributes and
creating better opportunities for colonization by r-strategists. A
higher value of MI in the natural soil was caused by a predatory
mite community with, in general, a higher proportion of 2K
species and lower proportion of r-species, suggesting that
communities are more stable under the natural soil.

a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 2 2 – 3 1 29
There is only sparse data available of MI from agricultural
soils. Therefore, for a comparative purpose, we calculated this
index from published species list in agroecosystems. MI was in
all cases lower than 0.55. Kovac et al. (1999) investigated
Gamasina communities in two agricultural areas in Slovakia
in 12 sites on two soil types. The average MI of the
communities was 0.37. In other studies in agricultural sites
in Europe the MI was 0.27 and 0.29 (Grishina et al., 1995), 0.55
(Schulz, 1991), 0.4 (Karg, 1967), 0.45 (Wasylik, 1995), 0.44
(Wasylik, 1989) and 0.5 (Schrader and Bayer, 2000). But in that
case no undisturbed site was taken as a control that would
allow the assessment of the deviation of the values from the
natural condition. These values are clearly lower than values
calculated for forest soils in Europe (Ruf, 1998). In our study, we
can consider the value in the natural site as a reference value
and show the decrease of the index in the managed soils.
The MI in the natural soil was similar to some values
obtained in wet pastures in Europe (Ruf, unpublished data).
But higher values were also calculated for other grasslands
(Karg, 1967).
The communities in our sites were dominated by 1r
species, only one species with K attributes was found in the
managed sites and two in the natural. In a natural grassland in
Europe, Karg (1967) found nine species with 2K and 1 with 3K
attributes. But also in some European agricultural sites the
proportion of K-attributes species was higher than in our sites
(Karg, 1967; Wasylik, 1989, 1995; Schulz, 1991).
The MI values were extraordinarily low even for agricul-
tural sites. We suggest two likely explanations for that: a
biogeographical or an ecological effect. In the first case, the K-
selected species are simply not present in Argentinean soils
due to historical reasons. In the second, environmental
conditions are so unfavorable and unpredictable (climatic
conditions and/or management regime) that only r-selected
species could survive.
In the analysis of community similarity, the dendrograms
were useful to illustrate the changes of predatory mite
community composition among sites. The general patter of
sites separation observed did not change much according to
the similarity measure used. All tested indices showed that
the communities in the mixed and the arable sites were the
most similar.
The choice of similarity measure had a considerable effect
on site discrimination, this was also shown in other studies
(e.g. Cao et al., 1997, for river benthic macroinvertebrates). The
Jaccard and Sorensen indices seem better suited for this
particular study since both were the measures that corre-
sponded best with the management explanation. The unique-
ness of the natural soil community in terms of species
composition is clear in our study. When the abundance
information was included in the indices, the similarity
between the community in the natural soil and the cattle
raising site increased.
This also suggests that for a biological classification of
these sites, the evaluation of species composition is more
widely applicable than the evaluation of densities of predatory
mites. Density values are less stable than species composition,
especially in arable lands (Ruf and Beck, 2005) and this can
determine a poorer separation of sites. This was also taken
into account when the MI was adapted to predatory mites;
density information was not considered, and the index was
based only on the occurrence of species (Ruf, 1998).
CCA is a useful tool to examine the relationship between
mites and soil parameters. In our study soil organic matter
content, moisture content and pH were the main factors
explaining the mite community structure. They correlated
well with the first axis of CCA, therefore, the first axis of the
CCA could be interpreted as a gradient of soil organic matter
content, moisture content and pH. The community structure
of the natural soil was best explained by a high soil organic
matter and moisture content, while the low content of those
factors and the relatively high pH were the factors that
appeared to best explain the community structure in the
agricultural site.
This is in agreement with other studies showing that the
community of soil mites depends on a complex combination
of these factors (Andren and Lagerlof, 1983; Kovac et al., 1999;
Ruf and Rombke, 1999).
Also the relationship between species and soil parameters
can be nicely observed in the diagram. For example, the
tendency of D. aff. crassitarsalis to occur at sites with high soil
organic matter content. Even though Gamasina mites are not
directly dependent on dead organic matter as food (Coja and
Bruckner, 2003) this parameter can have an important
influence on their prey (mainly nematodes and collembolans).
There were marked differences in the Gamasina fauna
between the four sites with different land use types. This
confirms our hypothesis that Gamasina community structure
significantly changes as material and energy inputs and
mechanical perturbations in the system increased from the
natural soil to the conventional agricultural sites.
In terms of species number, the arable fields had not an
impoverished community of Gamasina in comparison with
the natural undisturbed grassland. The effects of agricultural
practices could be observed in other characteristics of the
community: the occurrence of the species, the diversity and
the dominance structure of the community, and the maturity
index. Differences could be detected by means of the similarity
measures approach and the multivariate CCA. The cattle
raising site constituted a special ecosystem since it had not
been ploughed 4 years before the samplings. It can be
considered a 4-year successional site with grazing.
Our results support the idea that in agricultural systems,
Gamasina communities indicate effects of agricultural practice.
As it was suggested by Koehler (1999), we observed that specific
assemblages were associated with the peculiar environmental
conditions of agricultural fields, but not with the same species
dominating the community as in European soils.
The taxonomy of Mesostigmata is poorly developed in
Argentina. This work constitutes the first time that a
community of soil Gamasina is analyzed at the species level.
This was corroborated with the fact that only 3 of the 38
species identified were species previously known for science.
Acknowledgements
Funding for this study was provided by Deutscher Akade-
mischer Austauschdienst (DAAD) and SECyT-UNRC. A
CONICET fellowship is gratefully acknowledged. We are

a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 2 2 – 3 130
also grateful to Dr. H.H. Koehler for helping with mite
identification.
r e f e r e n c e s
Andren, O., Lagerlof, J., 1983. Soil fauna (microarthropods,enchytraeids, nematodes) in Swedish agricultural croppingsystems. Acta Agric. Scand. 33, 33–52.
Bardgett, R.D., Wardle, D.A., Yeates, G.W., 1998. Linking above-ground and below-ground interactions: how plantresponses to foliar herbivory influence soil organisms. SoilBiol. Biochem. 30, 1867–1878.
Beaulieu, F., Weeks, A.R., in press. Free-living mesostigmaticmites in australia: their role in biological control andbioindication. Aust. J. Exp. Agric.
Bolger, T., Curry, J.P., 1984. Influences of pig slurry on soilmicroarthropods in grassland. Rev. Ecol. Biol. Sol. 21, 269–281.
Bregetova, N.G., Wainstein, B.A., Kadite, B.A., Koroleva, E.W.,Petrova, A.D., Tichomirov, G.I., Shcherbak, G.I., 1977. Key tosoil inhabiting mites. Mesostigmata. Akad. Nauk SSSR,Leningrad, 718 pp. (in Russian).
Buryn, R., Hartmann, P., 1992. Gamasid fauna (Acari,Mesostigmata) of a hedge and adjacent meadows in UpperFranconia (Bavaria, Germany). Pedobiologia 36, 97–108.
Cantu, M.P., 1998. Estudio Geocientıfico para la EvaluacionAmbiental y la Ordenacion Territorial de una cuencapedemontana. Ph.D. Thesis. Universidad Nacional de RıoCuarto, Argentina, 377 pp.
Cao, Y., Bark, A., Williams, W.P., 1997. A comparison ofclustering methods for river benthic community analysis.Hydrobiologia 347, 25–40.
Clapperton, M.J., Kanashiro, D.A., Behan-Pelletier, V.M., 2002.Changes in abundance and diversity of microarthropodsassociated with Fescue Prairie grazing regimes.Pedobiologia 46, 496–511.
Coja, T., Bruckner, A., 2003. Soil microhabitat diversity in atemperate Norway spruce (Picea abies) forest does notinfluence the community composition of gamasid mites(Gamasida, Acari). Eur. J. Soil Biol. 39, 79–84.
El Titi, A., 1984. The effects of three tillage systems on soilinhabiting Gamasina (Mesostigmata, Acarina) in arable soil.Pedobiologia 27, 79–88.
Evans, G.O., Till, W.M., 1979. Mesostigmatic mites of Britain andIreland (Chelicerata: Acari-Parasitiformes): an introductionto their external morphology and classification. Trans. Zool.Soc. Lond. 35, 139–270.
Fisher, R.A., Corbet, A.S., Williams, C.B., 1943. The relationbetween the number of species and the number ofindividuals in a random sample of an animal population. J.Anim. Ecol. 12, 42–58.
Freckman, D.W., Duncan, D.A., Larson, J.R., 1979. Nematodedensity and biomass in an annual grassland ecosystem. J.Range Manage. 32, 418–422.
Grishina, L.G., Nikolskij, V.V., Wasylik, A., 1995. Communities ofAcarina in the soils of potato fields of Western Siberia. Pol.Ecol. Stud. 21, 293–309.
Hermosilla, W., Rubio, I., 1974. Prospeccion preliminar de lafauna edafica de la estancia El Vecino (cuenca del Salado,Prov. Buenos Aires, Argentina). Physics 33, 259–265.
Hermosilla, W., Reca, A., Pujalte, J., Rubio, I., 1977. Efectos de lacompactacion del suelo sobre la fauna edafica en campospastoreados (Partido de Chascomus, Provincia de BuenosAires, Argentina). Physics 36, 227–236.
Hirschmann, W., 1960. Gangsystematik der Parasitiformes, Teil3, Die Gattung Dendrolaelaps Halbert 1915. Acarologie.Schriftenreihe fur Vergleichende Milbenkunde 3, 1–27.
Holt, J.A., Bristow, K.L., McIvor, J.G., 1996. The effects of grazingpressure on soil animals and hydraulic properties of twosoils in semi-arid tropical Queensland. Aust. J. Soil Res. 34,69–79.
Hyatt, K.H., 1980. Mites of the subfamily Parasitinae(Mesostigmata: Parasitidae) in the British Isles. Bull. Br. Mus.(Nat. Hist.) (Zool.) 38, 237–378.
Karg, W., 1967. Synokologische Untersuchungen vonBodenmilben aus forstwirtschaftlich und landwirtschaftlichgenutzten Boden. Pedobiologia 7, 198–214.
Karg, W., 1993. Acari (Acarina), Milben, Parasitiformes(Anactinochaeta), Cohors Gamasina Leach, Raubmilben. In:Dahl, F. (Ed.), Die Tierwelt Deutschlands, 59, Teil. 2ndrevised ed. G. Fischer, Jena, Stuttgart, New York, p. 523.
Koehler, H., 1999. Predatory mites (Gamasina, Mesostigmata).Agric. Ecosys. Environ. 74, 395–410.
Kovac, L., Schnitzerova, E., Miklisova, D., Mati, R., 1999.Gamasina communities (Acari, Parasitiformes) of arablesoils with two different soil types. Pedobiologia 43, 54–63.
Krantz, G.W., 1998. Reflections on the biology, morphology andecology of the Macrochelidae. Exp. Appl. Acarol. 22, 125–137.
Krantz, G.W., Ainscough, B.D., 1990. Acarina: Mesostigmata(Gamasida). In: Dindal, D.L. (Ed.), Soil Biology Guide. JohnWiley & Sons Inc., New York, pp. 583–665.
Lagerlof, J., Andren, O., 1988. Abundance and activity ofsoil mites (Acari) in four cropping systems. Pedobiologia 32,129–145.
Larink, O., 1997. Springtails and mites: important knots in thefood web of soils. In: Benckiser, G. (Ed.), Fauna in SoilEcosystems. Marcel Dekker, New York, pp. 225–264.
Leetham, J.W., Milchunas, D.G., 1985. The composition anddistribution of soil microarthropods in the shortgrasssteppe in relation to soil water, root biomass, and grazing bycattle. Pedobiologia 28, 311–325.
Ruf, A., 1997. Fortpflanzungsbiologie von Raubmilben undCharakterisierung von Boden—Ein Konzept zur Indikationvon Belastungszustanden von Boden. Abh. Ber.Naturkundemuseum Gorlitz 69, 209–216.
Ruf, A., 1998. A maturity index for predatory soil mites(Mesostigmata: Gamasina) as an indicator of environmentalimpacts of pollution on forest soil. Appl. Soil Ecol. 9, 447–452.
Ruf, A., Beck, L., 2005. The use of predatory soil mites inecological soil classification and assessment concepts, withperspectives for oribatid mites. Ecotox. Environ. Safety 62,290–299.
Ruf, A., Rombke, J., 1999. Beurteilung von Bodenqualitat mitHilfe von Bodentieren. In: Koehler, H., Mathes, K., Breckling,B. (Eds.), Bodenokologie interdisziplinar. Springer-Verlag,Berlin, pp. 119–132.
Ruf, A., Beck, L., Dreher, P., Hund-Rinke, K., Rombke, J., Spelda,J., 2003. A biological classification concept for theassessment of soil quality: biological soil classificationscheme (BBSK). Agric. Ecosyst. Environ. 98, 263–271.
Schrader, S., Bayer, B., 2000. Abundances of mites (Gamasinaand Oribatida) and biotic activity in arable soil affected bytillage and wheeling. Braunschweiger Naturkundl. Schr. 6,165–181.
Schulz, E., 1991. Die Milbenfauna (Acari: Mesostigmata undCryptostigmata) in Lebensraumen auf Kalkgestein:Populationsokologie, Sukzession und Beziehungen zumLebensraum. Ber. Forsch. zent. Waldokosyst. A79, 1–245.
Shannon, C.E., Weaver, W., 1949. The Mathematical Theory ofCommunication. University of Illinois Press, Urbana, 125 pp.
Simpson, E.H., 1949. Measurement of diversity. Nature 163, 688.Soil Survey Staff, 1998. Keys to Soil Taxonomy, 8th ed. USDA,
Natural Resources Conservation Service. U.S. GovernmentPrinting Office, Washington, DC, 326 pp.
Southwood, T.R.E., 1980. Ecological Methods, 2nd ed. Chapman& Hall, New York, 524 pp.

a p p l i e d s o i l e c o l o g y 3 6 ( 2 0 0 7 ) 2 2 – 3 1 31
ter Braak, C.J.F., 1986. Canonical correspondence analysis: a neweigenvector technique for multivariate direct gradientanalysis. Ecology 67, 1167–1179.
ter Braak, C.J.F., Prentice, I.C., 1988. A theory of gradientanalysis. Adv. Ecol. Res. 18, 271–317.
ter Braak, C.J.F., Smilauer, P., 2004. CANOCO for WindowsVersion 4.5.3. Trial Version. Biometrics. Plant ResearchInternational, Wageningen, The Netherlands.
Universitat de Barcelona, 2005. GINKGO User’s Manual, Version1.4. Universitat de Barcelona.
Walter, D.E., 1988. Nematophagy by soil arthropods from theshortgrass steppe, Chihuahuan desert, and Rocky
Mountains of the central United States. Agric. Ecosyst.Environ. 24, 307–316.
Walter, D.E., Proctor, H.C., 1999. Mites. Ecology, Evolution andBehaviour. CABI Publishing, Sydney, 313 pp.
Wardle, D.A., 1995. Impacts of disturbance on detritus foodwebs in agro-ecosystems of contrasting tillage and weedmanagement practices. Adv. Ecol. Res. 26, 105–185.
Wasylik, A., 1989. Communities of Acarina in soil of a wheatfield. Pol. Ecol. Stud. 15, 55–73.
Wasylik, A., 1995. Communities of Acarina in the soil of potato-crops and Acarina succession patterns. Pol. Ecol. Stud. 21,311–334.





























