Embed Size (px)
Citation preview

Journal of Ecology
2008,
96
, 1128–1140 doi: 10.1111/j.1365-2745.2008.01431.x
© 2008 The Author. Journal compilation © 2008 British Ecological Society
Blackwell Publishing Ltd
Spatial patterns of recruitment in Mediterranean plant
species: linking the fate of seeds, seedlings and
saplings in heterogeneous landscapes at different
scales
Lorena Gómez-Aparicio*
Instituto de Recursos Naturales y Agrobiología (IRNAS, CSIC), PO Box 1052, Sevilla 41080, Spain
Summary
1.
Plant recruitment is a multiphase process that takes place in environments that are heterogene-ous in space and time. In this work, I analyse how environmental heterogeneity in Mediterraneanforests affects dynamics of early recruitment at different scales, using the wind-dispersed tree
Aceropalus
subsp.
granatense
as a case study.
2.
Seed dispersal and viability, post-dispersal predation, seedling emergence and seedling andsapling survival were evaluated in different habitats (regional scale) and microhabitats (local scale).Simultaneously, a review of the literature on spatial dynamics of plant recruitment in Mediterra-nean systems was conducted to look for general patterns and investigate their fit to the
Acer
system.Nineteen woody and herbaceous species were included in the review.
3.
At the regional scale,
Acer
recruitment dynamics strongly converged among sites of the samehabitat. This was mainly due to large seedling emergence and survival differences among habitats.Although most of the studies reviewed analyzed only one site per habitat type, they also supportstrong regional variation (either site- or habitat-specific) in recruitment patterns.
4.
At the local scale,
Acer
recruitment was microhabitat-specific, a result shared by almost all thereviewed species independently of their life form and dispersal syndrome. This was mostly due tospatial differences in seed arrival (higher under conspecifics) and seedling survival (higher undernurse shrubs).
5.
Spatial discordance among seed rain and recruitment was found in 60% of the reviewed speciesat the regional scale, and in 67% at the local scale.
Acer
results supported this predominant lack ofconcordance. Discordance among seed rain and recruitment suggests that regeneration is largelylimited by safe sites than by seed availability. Because seedling survival was the limiting process witha larger impact on the magnitude and spatial pattern of recruitment, safe sites might be defined asthose where seedlings have a higher survival probability.
6.
Synthesis
: This study indicates that the influence of seed dispersal on the spatial patterns anddemography of plant species could be limited in heterogeneous and stressful environments (asare found in the Mediterranean), where recruitment is restricted to a small fraction of the landscape.If we are to preserve the distribution and abundance of Mediterranean species in the face ofenvironmental changes, we need to explicitly consider the strong patch-specificity that characterizestheir recruitment process at all scales.
Key-words:
Acer opalus
subsp.
granatense
, habitats, microhabitats, plant demography,recruitment dynamics, seed dispersal, seedling survival, spatial discordance, spatial heterogeneity
Introduction
Plant recruitment takes place in environments that areheterogeneous in space and time (Stewart
et al
. 2000).
Heterogeneity can therefore be defined as any factor ableto induce variation in demographic rates (Rees
et al
. 2000).This definition allows us to measure and compare the relevancefor demographic dynamics of different forms of heterogeneityacting at different scales organized in a hierarchical manner.At a regional scale, habitat-specific heterogeneity can be
*Correspondence author. E-mail: [email protected]

Spatial dynamics of recruitment at different scales
1129
© 2008 The Author. Journal compilation © 2008 British Ecological Society,
Journal of Ecology
,
96
, 1128–1140
caused by unique conditions related to topography, climate,or disturbance level (Traveset
et al
. 2003; Quintana-Ascencio
et al
. 2007; Zavaleta
et al
. 2007). At smaller spatial scales,heterogeneity can be the result of variations in microclimate(Canham
et al
. 1994; Nicotra
et al
. 1999; Puerta-Piñero
et al
.2007) or soil moisture and nutrient availability (Farley &Fitter 1999; Hutchings
et al
. 2003). Much of this variability isgenerated by organisms (Pickett
et al
. 2000). The extent towhich these sources of heterogeneity cause patch-specificrecruitment dynamics vary widely among species. Thus, theinfluence of environmental heterogeneity on species recruit-ment can be considered a continuum, from species that arevery specific in their environmental requirements (also calledniche- or microsite-limited species), to species with weakhabitat affinities and for which abundance is influencedmainly by seed availability (also called seed-limited species)(Münzbergová & Herben 2005; Moore & Elmendorf 2006).Explicit knowledge of where a species sits in this continuummay be crucial when predicting the consequences of environ-mental changes for species abundance and distribution atdifferent spatial scales.
Recruitment is a multiphase process involving severalsequential life-history stages (i.e. seeds, seedlings, saplings)connected by transitional processes (i.e. dispersal, emergence,survival) (Herrera
et al
. 1994; Clark
et al
. 1999; Wang &Smith 2002). Therefore, a full understanding of the impactof heterogeneity on recruitment requires an integrative‘linking-stages’ approach that explores how the effects onone stage cascade to the end of the cycle (Schupp & Fuentes1995). This is particularly true considering that processes inthe regeneration cycle can be spatially discordant, high-quality patches for some stages turning into low-qualitypatches for others (Houle 1992, 1994; Chambers 1995; Jordano& Herrera 1995; Clark
et al
. 1999; Rey & Alcántara 2000;Wenny 2000; Traveset
et al
. 2003; Gómez 2004). In highlyheterogeneous environments, strong spatial discordancemight cause a total collapse of regeneration even when anyprocess, in isolation, can be considered strongly limiting. Froma seed- vs. microsite-limitation framework, the absence of aspatial concordance between the initial seed template andwhere final recruitment occurs might be viewed as theresult of microsite-limitation processes. However, a strongconcordance across space would indicate prevalence, in plantpopulations, of seed-limitation processes.
In this work, I seek to analyze how environmental hetero-geneity in Mediterranean forests affects the spatial dynamicsand overall magnitude of recruitment at different scales(regional and local), using the wind-dispersed tree
Acer opalus
subsp
. granatense
Font Quer & Rothm (Aceraceae) as acase study. I used a demographic approach, consideringthe plant life cycle as a conjunction of consecutive stages(pre-dispersed seed, dispersed seed, seedling and sapling)connected by processes with specific transition probabilities(TPs). A TP is defined as the probability of moving fromone stage to the next, and thereby evaluates the losses ofpotential new individuals at each stage in the reproductivecycle. Considering TPs to be multiplicative rather than
additive, overall probabilities of recruitment (OPR) in eachspatial unit would be calculated as the product of every TP.Spatio-temporal patterns of individual TPs in
Acer
havebeen discussed elsewhere (see Gómez-Aparicio
et al
. 2007for seed dispersal and Gómez-Aparicio
et al.
2005a forseedling emergence and survival). By linking the seed,seedling and sapling stages, this article constitutes a broaderexpansion of the ideas developed in those previous articles,presenting a formal comparison of the relative importanceand spatial concordance of the processes that connect thesethree key life-stages.
For the study of TPs and OPR at the regional scale, Iselected four
Acer
populations occupying contrasting habitattypes (pine forests vs. stony slopes). By studying two sitesper habitat type, I ask whether recruitment dynamics aresite-specific or converge in sites with similar coarse-scalecharacteristics (i.e. similar habitats). This issue is still farfrom clear, because only one site per habitat type is usuallyconsidered in recruitment studies (i.e. Herrera
et al
. 1994;Vilá & Lloret 2000; Lázaro
et al
. 2006), which does notallow the separation of site- from habitat-specific patterns. Atthe local scale, I selected a number of microhabitats persite (4–7) differing in cover type, and asked whether TPsand OPR varied among conspecific and heterospecific trees,shrub species and open spaces without vegetation. In thisway, I meant to explore how plant interactions usuallyexplored for just one life-history stage (e.g. nurse effectson seedlings; Gómez-Aparicio
et al
. 2004, 2005a) operateat different stages that interact to affect final patterns ofsapling recruitment.
Specifically, I addressed the following questions: (1)At the regional scale, are
Acer
recruitment patterns site-dependent or habitat-dependent? (2) At the local scale, canmicrohabitats be classified as of high vs. low quality foroverall recruitment? If so, is such classification regionallyconsistent? (3) How spatially concordant are seed andrecruitment patterns at both the regional and local scales?The answers to these questions are discussed in the context ofthe results reported by similar studies conducted for otherMediterranean woody and herbaceous species. For this, Ireviewed the literature on ‘linking-stages’ analyses of recruit-ment patterns in Mediterranean plant species; a line ofresearch that has markedly advanced in recent decades,inspired by the seminal paper by Herrera
et al
. (1994).The results presented here constitute an effort to bothimprove and synthesize our current knowledge of thespatial patterns and limitations in the recruitment cycle ofMediterranean species.
Methods
STUDY
SPECIES
AND
S ITES
Acer opalus
subsp.
granatense
is the southernmost species of thegenus
Acer
in the Northern Hemisphere. Its distribution is composedof many small patches scattered throughout medium and highmountains (1100–2000 m a.s.l.) in the SE Iberian Peninsula, Balearic
1130
L. Gómez-Aparicio
© 2008 The Author. Journal compilation © 2008 British Ecological Society,
Journal of Ecology
,
96
, 1128–1140
Islands and northern Morocco (López-González 1994).
Acer opalus
subsp.
granatense
is catalogued as Vulnerable by the IUCN (2001)and included in the Red List of Threatened Vascular Plants ofAndalusia (Blanca
et al
. 2000). It is essentially dimorphic in gender,with protogynous individuals belonging to a mainly female class andmales and protandrous individuals, together, belonging to a maleclass (Gleiser
et al
. 2008). The indehiscent fruits are schizocarps,composed of two one-seeded samaroid mericarps. Seed viability israther low because of parthenocarpic abortion and post-dispersalpredation by
Bradybatus elongatulus
Boheman (Curculionidae). Anemo-chorous dispersal occurs during fall (September–December). Seedshave physiological and physical dormancy (Gleiser
et al
. 2004),germinating after winter. During this time, they may be consumedby rodents (mainly
Apodemus sylvaticus
). Seedlings are severely affectedby water stress during summer (Gómez-Aparicio
et al
. 2005a). Sap-lings are intensively damaged by ungulates (Gómez-Aparicio
et al
.2005b).
The study was conducted in two mountain ranges of southernSpain, Sierra Nevada (SN) and Sierra de Baza (SB), approximately80 km apart. The climate is Mediterranean, with rainfall concen-trated in late autumn and winter and hot dry summers. SierraNevada has much higher annual rainfall than does Sierra de Baza(826.6
±
91.3 mm vs. 520.8
±
60.9 mm; 1991–2004), although bothhave similar low summer rainfall (June–August: 40.7
±
8.7 mm and37.1
±
9.2 mm; 1991–2004). Winters are cold and snow is common,persisting up to 2 months, and frost occurs from November toApril. The study was conducted in 3 years with contrasting climaticconditions, whereas the first 2 years (2000 and 2001) had relativelydry summers with rainfall below the average for the site (2.0 mmand 10.0 mm in SN, 13.0 mm and 0 mm in SB), the last year (2002)had an infrequent wet summer (76.5 mm in SN, 57.2 mm in SB).
Two study sites were chosen per mountain range: one site consistedof maple intermingled with
Pinus sylvestris
forest (forest sites [F],hereafter) and the other a maple population situated on a stonyslope (stony-slope sites [SL], hereafter). The two habitat types(i.e. forests and stony-slopes) differed mainly in soil substrate andcanopy cover. Substrate in SL was composed largely of bare soilwith loose stones, whereas F presented a stable substrate and a highertree cover (Table 1). All sites were located between 1850–2000 ma.s.l., and had northern orientation and steep slopes (30–40
°
). Weidentified 4–7 microhabitats per site, a microhabitat being definedas a distinct homogeneous patch of habitat ranging from
<
1 m toseveral meters in diameter (Russell & Schupp 1998; García 2001).Microhabitats belonged to one of the following categories: (1) under
Acer
adults; (2) under the canopy of heterospecific trees or shrubs
>
1.5 m in height; (3) under shrubs
<
1.5 m in height; and (4) open inter-spaces between woody vegetation (Table 1). Tree and shrub specieswere chosen according to their availability at each site. Microhabitatcover was determined by means of 25
×
2 m transects (
n
=
10 persite), visually estimating cover percentages at each 1
×
2 m
2
.
SAMPLING
DESIGN
Seed rain and viability
Seed traps (0.045 m
2
aluminium trays) were placed at 30 samplinglocations per site and microhabitat combination, each locationcontaining two adjacent traps (
n
=
1260 traps). Traps were coveredwith 1.3-cm diameter mesh to avoid post-dispersal predation.Traps were sampled fortnightly during the dispersal period(September–December) of the years 2001 and 2002. The trapcontent was transported to the laboratory, where seeds were dissected
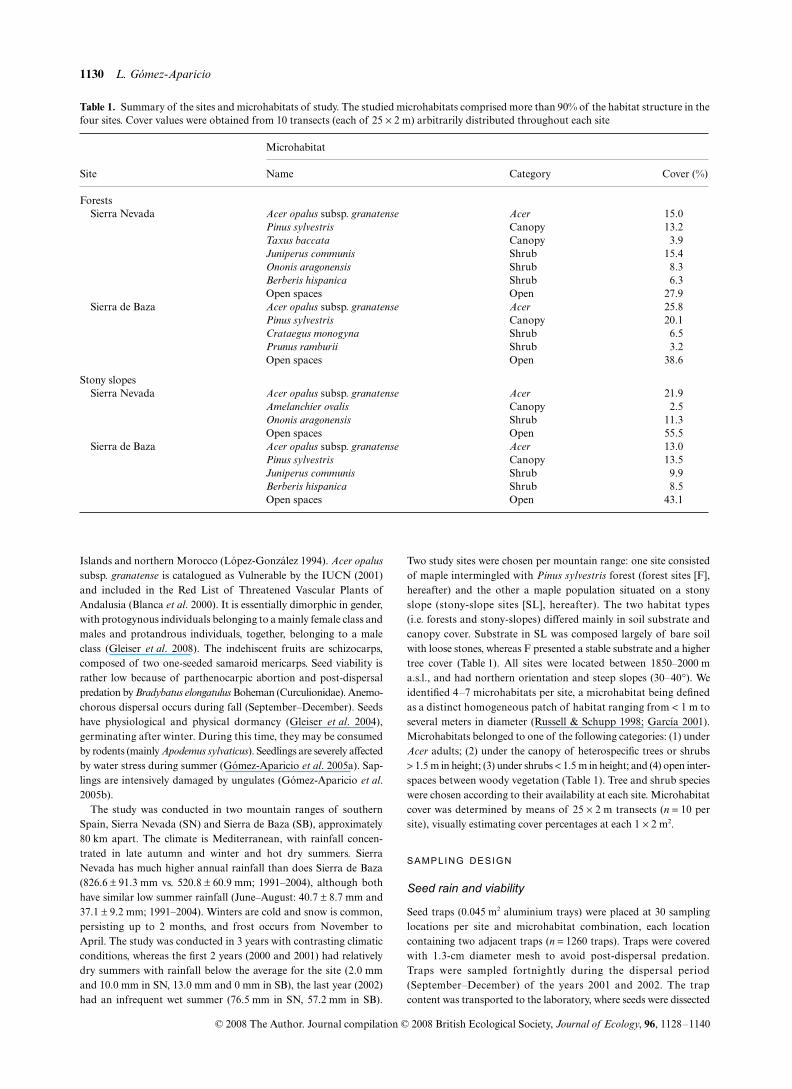
Table 1. Summary of the sites and microhabitats of study. The studied microhabitats comprised more than 90% of the habitat structure in thefour sites. Cover values were obtained from 10 transects (each of 25 × 2 m) arbitrarily distributed throughout each site
Site
Microhabitat
Name Category Cover (%)
ForestsSierra Nevada Acer opalus subsp. granatense Acer 15.0
Pinus sylvestris Canopy 13.2Taxus baccata Canopy 3.9Juniperus communis Shrub 15.4Ononis aragonensis Shrub 8.3Berberis hispanica Shrub 6.3Open spaces Open 27.9
Sierra de Baza Acer opalus subsp. granatense Acer 25.8Pinus sylvestris Canopy 20.1Crataegus monogyna Shrub 6.5Prunus ramburii Shrub 3.2Open spaces Open 38.6
Stony slopesSierra Nevada Acer opalus subsp. granatense Acer 21.9
Amelanchier ovalis Canopy 2.5Ononis aragonensis Shrub 11.3Open spaces Open 55.5
Sierra de Baza Acer opalus subsp. granatense Acer 13.0Pinus sylvestris Canopy 13.5Juniperus communis Shrub 9.9Berberis hispanica Shrub 8.5Open spaces Open 43.1

Spatial dynamics of recruitment at different scales
1131
© 2008 The Author. Journal compilation © 2008 British Ecological Society,
Journal of Ecology
,
96
, 1128–1140
for viability assessment (see Gómez-Aparicio
et al
. 2007 for furtherdetails). Viable seeds contain a yellowish embryo, which entirelyfills the seed locule, whereas aborted seeds bear an undevelopedembryo. Previous experiences showed that seeds estimated visuallyas filled correspond with viable seeds after tetrazolium test in morethan 90% of the cases (author’s unpublished data).
Post-dispersal seed predation
Losses to post-dispersal seed predation were evaluated at the samesampling locations as seed rain. A Petri dish (5 cm in diameter) waslocated at each location with two seeds inside (simulating dispersal ofan
Acer
fruit). To control for abiotic losses, 10 Petri dishes excludedfrom predators by a wire cage of 1.3 cm mesh (control treatment)were placed next to 10 out of the 30 exposed Petri dishes per micro-habitat. All the Petri dishes were fixed to the soil and camouflagedwith the corresponding substrate. The seeds used for the experimentwere collected in September 2001 from 5–6 parent trees per mountainrange. Seeds from the different trees were mixed to provide two seedpools, one for SN and another for SB, each used at the two siteswithin the corresponding mountain range. The number of survivingseeds in the dishes was recorded after 2, 7, 30 and 60 days. Theexperiment was set up twice, once in October 2001 (to quantify fallpredation) and once in April 2002 (to quantify spring predation).
Seedling emergence
The density of naturally emerged seedlings was sampled at the endof May in 2001 and 2002, coinciding with the peak of seedlingemergence in the study area. At each of the four study sites, thenumber of newly emerged seedlings was counted in 200–300 samplingquadrats (0.5 m
2
) randomly distributed per microhabitat (stratifiedrandom design).
The probability of seedling emergence was studied by conductinga seed sowing experiment at the same sampling locations where seedrain and post-dispersal seed predation were evaluated. Seeds usedin the sowing experiment belonged to the same pools used for thepredation experiments. The viability of the seeds in the pools wasassessed in the laboratory (12% viability in SN, 10% in SB). InDecember 2001, seeds were sown in groups of five at each samplinglocation (
n
=
3150 seeds). Within each group, seeds were sown 2 cmapart and at 2 cm in depth, and were protected with a wire cageto prevent seed predation by rodents and additional seed rain.Emergence was monitored fortnightly from the beginning of April tothe end of June 2002, when no additional emergence was registered(see further details in Gómez-Aparicio
et al
. 2005a).
Seedling and sapling survival
The probability of seedling survival was studied by individuallymarking 100–200 newly emerged seedlings per site and microhabitatcombination (
n
=
7200 seedlings) in three consecutive years (2000,2001 and 2002). In SL-SB, seedlings were marked only in 2001 and2002. Seedlings were censused twice a year, once after summer(September) and once after winter (April). Individuals survivingthe first summer were considered to be established saplings (seeGómez-Aparicio
et al
. 2005a for further details).To assess whether the long-term sapling distribution in the
Acer
populations mirrored the net results of the short-term processesanalyzed, a static survey of the abundance of saplings (i.e. 2–16-year-oldindividuals) was conducted in fall 2002. At each site, saplings were
sampled using 10 randomly positioned transects (25
×
2 m), and themicrohabitat in which they had established was recorded.
TRANSIT ION
PROBABIL IT IES
A total of seven process-specific TPs were calculated for eachcombination of site and microhabitat:
1.
Seed dispersal (D), the probability that a seed was dispersed to aparticular microhabitat (estimated as mean density of seeds dispersedin each microhabitat – average for 2001 and 2002 – relative to the sumof mean seed densities deposited in all microhabitats considered).
2.
Seed viability (V), the probability of a dispersed seed being viable(estimated as the proportion of viable seeds captured in the seedtraps; seed viability tested in the laboratory).
3.
Post-dispersal seed survival (S
s
), the probability of escapingpredation by rodents (estimated as the average survival percentage infall 2001 and spring 2002).
4.
Seedling emergence (E), the probability of emergence the firstspring after dispersal (estimated as the proportion of seedlings thatemerged in the sowing experiment multiplied by the viability rates ofthe seeds sown).
5.
Seedling survival (Sd
s), the probability of seedling survival afterthe first summer (estimated as an average for the three study years,except for SL-SB, for which only 2001 and 2002 data were available).6. Two-year old sapling survival (Sp1s), the probability of survival afterthe second summer (estimated as the average percentage of seedlingsfrom cohorts 2000 and 2001 that survived the second year of life).7. Three-year old sapling survival (Sp2s), the probability of survivalafter the third summer (estimated as the percentage of 2-year-oldsaplings from cohort 2000 that survived the third year of life; notcalculated for SL-SB).
The OPR in each microhabitat was calculated as the product of all sevenTPs. In addition, to quantify the overall quality of each microhabitatfor recruitment of a dispersed seed, a second index (overall quality,OQ) was calculated by multiplying all TPs except the probability ofseed dispersal (D). To extrapolate the microhabitat information tothe regional scale, TPs and OPR in each site were calculated byadding the products of the TPs and OPR (respectively) in eachmicrohabitat multiplied by the relative cover of each microhabitat.
DATA ANALYSIS
Most statistical analyses were conducted using Generalized LinearModels (SAS 2002). Different error distributions and link functionswere selected depending on the nature of the dependent variable. Thenumber of seeds dispersed per trap, number of natural emergencesper quadrat, and number of saplings per transect were fitted to aPoisson distribution with Log as the link function. The probabilityof seed viability, post-dispersal seed predation, seeding emergenceand seedling survival was analyzed using binomial error distributionsand Logit link functions. As there were no differences between sitesor microhabitats in abiotic looses (P > 0.05, one-way anovas), theywere not considered in the statistical analysis of seed-predation data.I first explored differences among sites in the processes studied(introducing Site as main factor) and then among microhabitatsseparately for each of the four sites (introducing Microhabitat asthe main factor). Concordance across sites and microhabitats in theprocess of recruitment was explored using TPs values and Kendallcoefficients of concordance solved by means of permutation tests(n = 9999 permutations, Legendre 2004). When data for severalyears or seasons were available, analyses were conducted on average
1132 L. Gómez-Aparicio
© 2008 The Author. Journal compilation © 2008 British Ecological Society, Journal of Ecology, 96, 1128–1140
values. Temporal variation in Acer demographic parameters hasbeen discussed elsewhere, and it has been shown to be generallylower than spatial variation (see Gómez-Aparicio 2004 for seedpredation, Gómez-Aparicio et al. 2007 for seed dispersal andGómez-Aparicio et al. 2005a for first-year seedling survival). Dataare reported as means ± 1 SE.
COMPILATION OF PUBLISHED STUDIES
I searched Science Citation Index Expanded (Web of Science,Thompson Scientific) for articles containing the words ‘Mediterranean’,‘spatial’, ‘recruitment’ and/or ‘regeneration’ in the title, keywords, orabstract. From the resulting set of articles, I selected all articlesthat investigated spatial patterns of transitional processes (i.e.dispersal, emergence, survival) among early life stages (seed,seedling, sapling) in Mediterranean terrestrial systems at least atone spatial scale (regional or local). Articles that covered only theseedling-sapling stages were excluded, unless they could bematched to a second article where the seed stage was analyzed.This resulted in a final number of 26 articles including 19 species oftrees, shrubs, herbs and grasses. For each study, I synthesized theinformation on (1) the number of habitat types were the earlyrecruitment cycle was analyzed; (2) number of sites per habitattype (replicate plots within the same site were not consideredas different sites); (3) number of microhabitats per site (whendifferent transitional processes were analyzed in a differentnumber of microhabitats, only the shared microhabitats wereconsidered); (4) critical TPs identified in the recruitment process;and (5) if there was spatial concordance among seed rain andrecruitment at regional (i.e. among habitats or sites) and local(i.e. among microhabitats) scales. A summary of these results isshown in Appendix S1 (see Supporting Information).
Results
SPATIAL PATTERNS OF A C E R RECRUITMENT AT THE REGIONAL SCALE
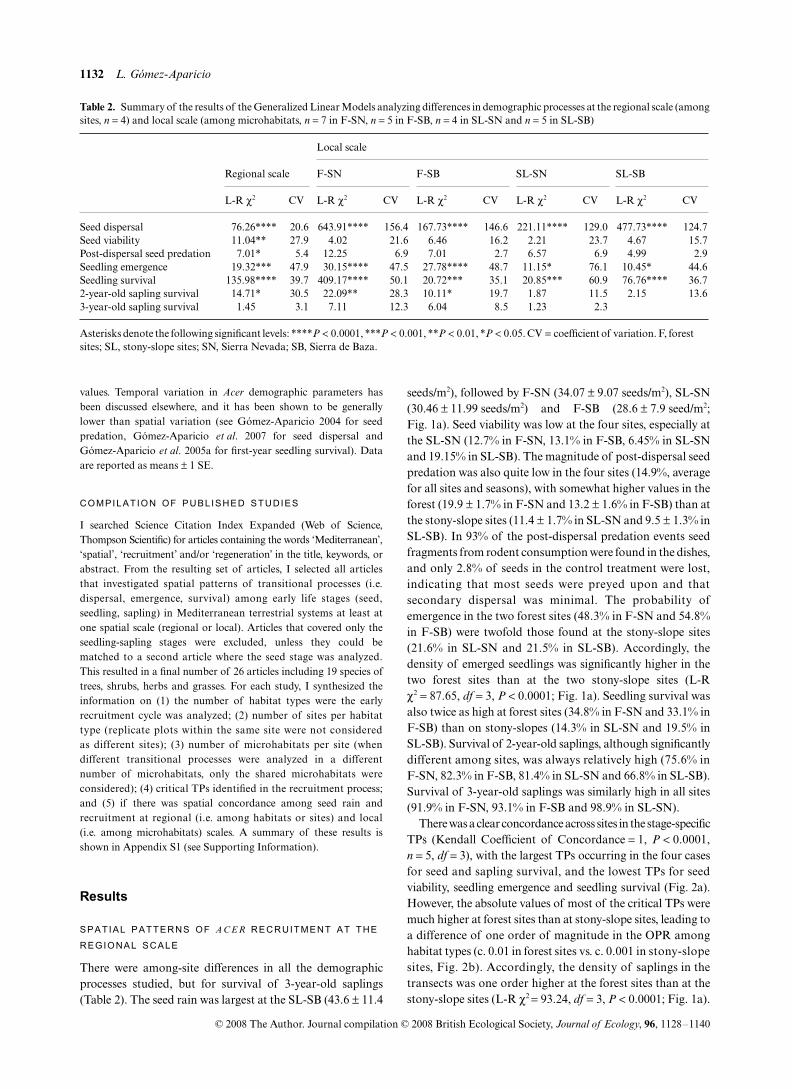
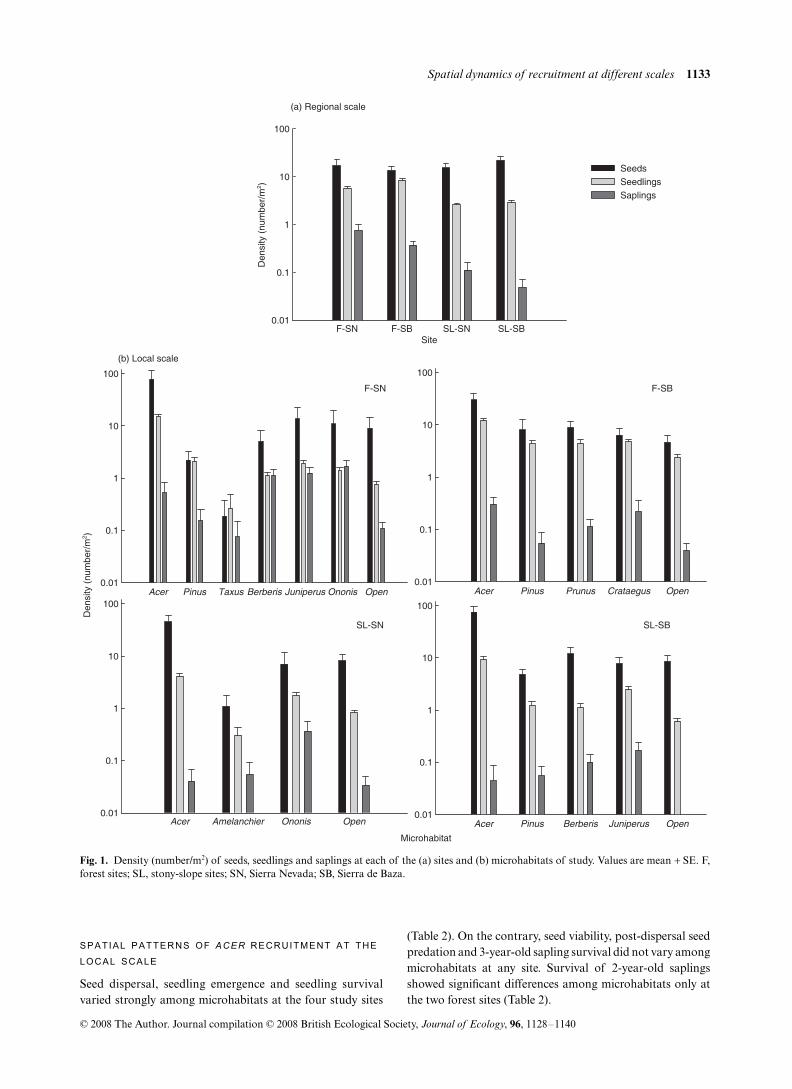
There were among-site differences in all the demographicprocesses studied, but for survival of 3-year-old saplings(Table 2). The seed rain was largest at the SL-SB (43.6 ± 11.4
seeds/m2), followed by F-SN (34.07 ± 9.07 seeds/m2), SL-SN(30.46 ± 11.99 seeds/m2) and F-SB (28.6 ± 7.9 seed/m2;Fig. 1a). Seed viability was low at the four sites, especially atthe SL-SN (12.7% in F-SN, 13.1% in F-SB, 6.45% in SL-SNand 19.15% in SL-SB). The magnitude of post-dispersal seedpredation was also quite low in the four sites (14.9%, averagefor all sites and seasons), with somewhat higher values in theforest (19.9 ± 1.7% in F-SN and 13.2 ± 1.6% in F-SB) than atthe stony-slope sites (11.4 ± 1.7% in SL-SN and 9.5 ± 1.3% inSL-SB). In 93% of the post-dispersal predation events seedfragments from rodent consumption were found in the dishes,and only 2.8% of seeds in the control treatment were lost,indicating that most seeds were preyed upon and thatsecondary dispersal was minimal. The probability ofemergence in the two forest sites (48.3% in F-SN and 54.8%in F-SB) were twofold those found at the stony-slope sites(21.6% in SL-SN and 21.5% in SL-SB). Accordingly, thedensity of emerged seedlings was significantly higher in thetwo forest sites than at the two stony-slope sites (L-Rχ2 = 87.65, df = 3, P < 0.0001; Fig. 1a). Seedling survival wasalso twice as high at forest sites (34.8% in F-SN and 33.1% inF-SB) than on stony-slopes (14.3% in SL-SN and 19.5% inSL-SB). Survival of 2-year-old saplings, although significantlydifferent among sites, was always relatively high (75.6% inF-SN, 82.3% in F-SB, 81.4% in SL-SN and 66.8% in SL-SB).Survival of 3-year-old saplings was similarly high in all sites(91.9% in F-SN, 93.1% in F-SB and 98.9% in SL-SN).
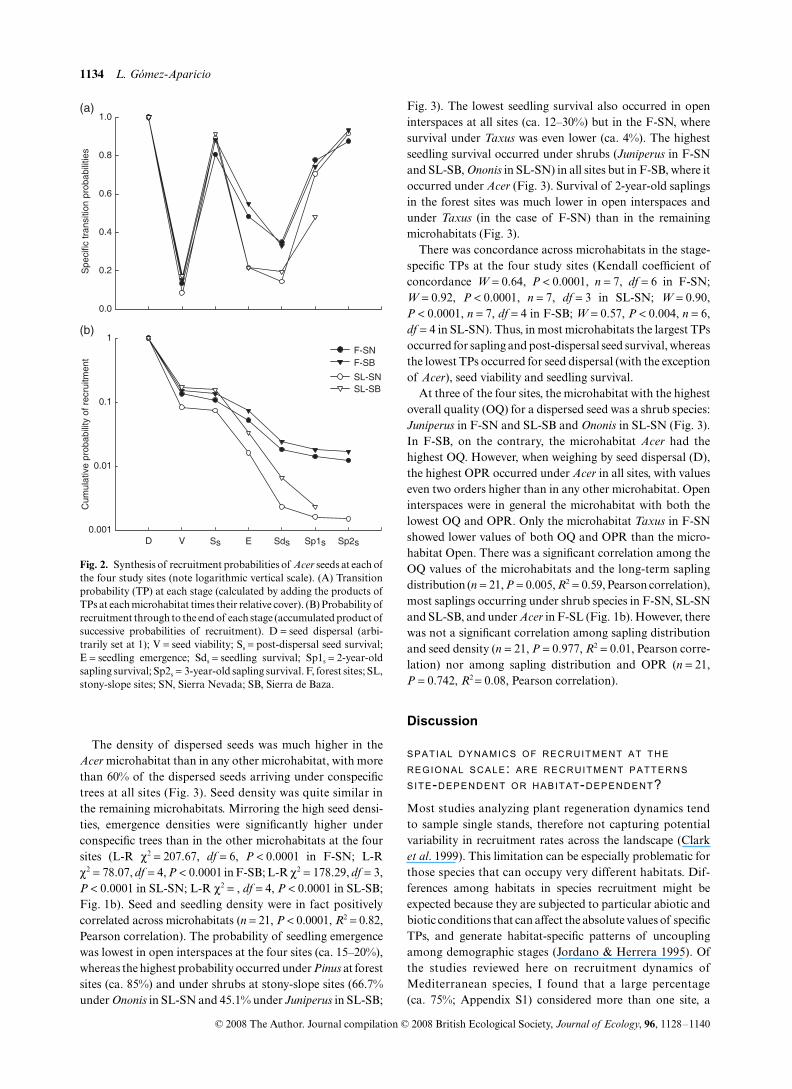
There was a clear concordance across sites in the stage-specificTPs (Kendall Coefficient of Concordance = 1, P < 0.0001,n = 5, df = 3), with the largest TPs occurring in the four casesfor seed and sapling survival, and the lowest TPs for seedviability, seedling emergence and seedling survival (Fig. 2a).However, the absolute values of most of the critical TPs weremuch higher at forest sites than at stony-slope sites, leading toa difference of one order of magnitude in the OPR amonghabitat types (c. 0.01 in forest sites vs. c. 0.001 in stony-slopesites, Fig. 2b). Accordingly, the density of saplings in thetransects was one order higher at the forest sites than at thestony-slope sites (L-R χ2 = 93.24, df = 3, P < 0.0001; Fig. 1a).
Table 2. Summary of the results of the Generalized Linear Models analyzing differences in demographic processes at the regional scale (amongsites, n = 4) and local scale (among microhabitats, n = 7 in F-SN, n = 5 in F-SB, n = 4 in SL-SN and n = 5 in SL-SB)
Regional scale
Local scale
F-SN F-SB SL-SN SL-SB
L-R χ2 CV L-R χ2 CV L-R χ2 CV L-R χ2 CV L-R χ2 CV
Seed dispersal 76.26**** 20.6 643.91**** 156.4 167.73**** 146.6 221.11**** 129.0 477.73**** 124.7Seed viability 11.04** 27.9 4.02 21.6 6.46 16.2 2.21 23.7 4.67 15.7Post-dispersal seed predation 7.01* 5.4 12.25 6.9 7.01 2.7 6.57 6.9 4.99 2.9Seedling emergence 19.32*** 47.9 30.15**** 47.5 27.78**** 48.7 11.15* 76.1 10.45* 44.6Seedling survival 135.98**** 39.7 409.17**** 50.1 20.72*** 35.1 20.85*** 60.9 76.76**** 36.72-year-old sapling survival 14.71* 30.5 22.09** 28.3 10.11* 19.7 1.87 11.5 2.15 13.63-year-old sapling survival 1.45 3.1 7.11 12.3 6.04 8.5 1.23 2.3
Asterisks denote the following significant levels: ****P < 0.0001, ***P < 0.001, **P < 0.01, *P < 0.05. CV = coefficient of variation. F, forest sites; SL, stony-slope sites; SN, Sierra Nevada; SB, Sierra de Baza.
Spatial dynamics of recruitment at different scales 1133
© 2008 The Author. Journal compilation © 2008 British Ecological Society, Journal of Ecology, 96, 1128–1140
SPATIAL PATTERNS OF ACER RECRUITMENT AT THE LOCAL SCALE
Seed dispersal, seedling emergence and seedling survivalvaried strongly among microhabitats at the four study sites
(Table 2). On the contrary, seed viability, post-dispersal seedpredation and 3-year-old sapling survival did not vary amongmicrohabitats at any site. Survival of 2-year-old saplingsshowed significant differences among microhabitats only atthe two forest sites (Table 2).
Fig. 1. Density (number/m2) of seeds, seedlings and saplings at each of the (a) sites and (b) microhabitats of study. Values are mean + SE. F,forest sites; SL, stony-slope sites; SN, Sierra Nevada; SB, Sierra de Baza.
1134 L. Gómez-Aparicio
© 2008 The Author. Journal compilation © 2008 British Ecological Society, Journal of Ecology, 96, 1128–1140
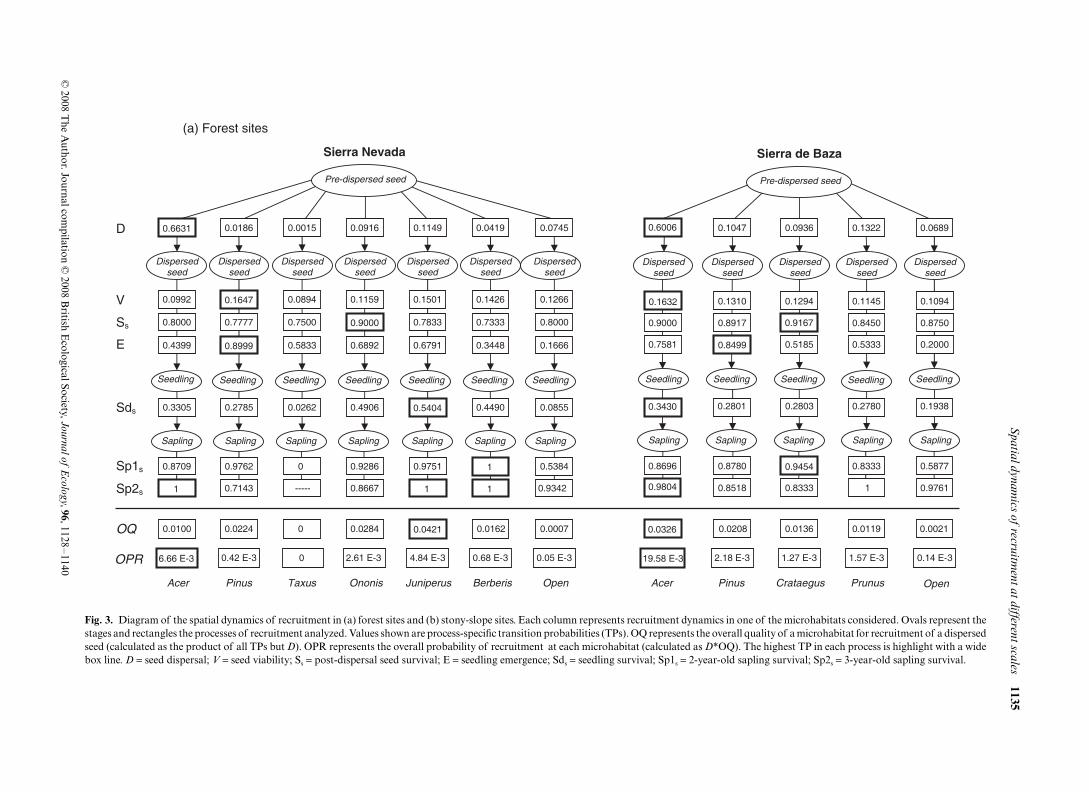
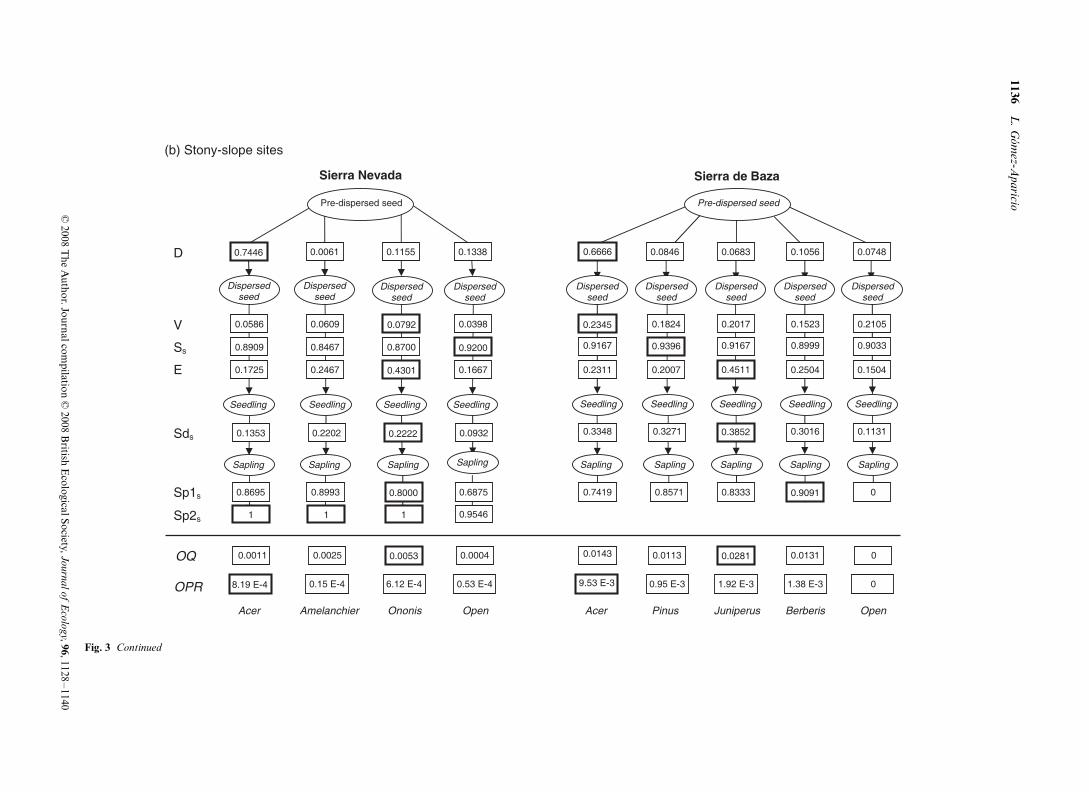
The density of dispersed seeds was much higher in theAcer microhabitat than in any other microhabitat, with morethan 60% of the dispersed seeds arriving under conspecifictrees at all sites (Fig. 3). Seed density was quite similar inthe remaining microhabitats. Mirroring the high seed densi-ties, emergence densities were significantly higher underconspecific trees than in the other microhabitats at the foursites (L-R χ2 = 207.67, df = 6, P < 0.0001 in F-SN; L-Rχ2 = 78.07, df = 4, P < 0.0001 in F-SB; L-R χ2 = 178.29, df = 3,P < 0.0001 in SL-SN; L-R χ2 = , df = 4, P < 0.0001 in SL-SB;Fig. 1b). Seed and seedling density were in fact positivelycorrelated across microhabitats (n = 21, P < 0.0001, R2 = 0.82,Pearson correlation). The probability of seedling emergencewas lowest in open interspaces at the four sites (ca. 15–20%),whereas the highest probability occurred under Pinus at forestsites (ca. 85%) and under shrubs at stony-slope sites (66.7%under Ononis in SL-SN and 45.1% under Juniperus in SL-SB;
Fig. 3). The lowest seedling survival also occurred in openinterspaces at all sites (ca. 12–30%) but in the F-SN, wheresurvival under Taxus was even lower (ca. 4%). The highestseedling survival occurred under shrubs (Juniperus in F-SNand SL-SB, Ononis in SL-SN) in all sites but in F-SB, where itoccurred under Acer (Fig. 3). Survival of 2-year-old saplingsin the forest sites was much lower in open interspaces andunder Taxus (in the case of F-SN) than in the remainingmicrohabitats (Fig. 3).
There was concordance across microhabitats in the stage-specific TPs at the four study sites (Kendall coefficient ofconcordance W = 0.64, P < 0.0001, n = 7, df = 6 in F-SN;W = 0.92, P < 0.0001, n = 7, df = 3 in SL-SN; W = 0.90,P < 0.0001, n = 7, df = 4 in F-SB; W = 0.57, P < 0.004, n = 6,df = 4 in SL-SN). Thus, in most microhabitats the largest TPsoccurred for sapling and post-dispersal seed survival, whereasthe lowest TPs occurred for seed dispersal (with the exceptionof Acer), seed viability and seedling survival.
At three of the four sites, the microhabitat with the highestoverall quality (OQ) for a dispersed seed was a shrub species:Juniperus in F-SN and SL-SB and Ononis in SL-SN (Fig. 3).In F-SB, on the contrary, the microhabitat Acer had thehighest OQ. However, when weighing by seed dispersal (D),the highest OPR occurred under Acer in all sites, with valueseven two orders higher than in any other microhabitat. Openinterspaces were in general the microhabitat with both thelowest OQ and OPR. Only the microhabitat Taxus in F-SNshowed lower values of both OQ and OPR than the micro-habitat Open. There was a significant correlation among theOQ values of the microhabitats and the long-term saplingdistribution (n = 21, P = 0.005, R2 = 0.59, Pearson correlation),most saplings occurring under shrub species in F-SN, SL-SNand SL-SB, and under Acer in F-SL (Fig. 1b). However, therewas not a significant correlation among sapling distributionand seed density (n = 21, P = 0.977, R2 = 0.01, Pearson corre-lation) nor among sapling distribution and OPR (n = 21,P = 0.742, R2 = 0.08, Pearson correlation).
Discussion
SPATIAL DYNAMICS OF RECRUITMENT AT THE REGIONAL SCALE: ARE RECRUITMENT PATTERNS SITE-DEPENDENT OR HABITAT-DEPENDENT?
Most studies analyzing plant regeneration dynamics tendto sample single stands, therefore not capturing potentialvariability in recruitment rates across the landscape (Clarket al. 1999). This limitation can be especially problematic forthose species that can occupy very different habitats. Dif-ferences among habitats in species recruitment might beexpected because they are subjected to particular abiotic andbiotic conditions that can affect the absolute values of specificTPs, and generate habitat-specific patterns of uncouplingamong demographic stages (Jordano & Herrera 1995). Ofthe studies reviewed here on recruitment dynamics ofMediterranean species, I found that a large percentage(ca. 75%; Appendix S1) considered more than one site, a
Fig. 2. Synthesis of recruitment probabilities of Acer seeds at each ofthe four study sites (note logarithmic vertical scale). (A) Transitionprobability (TP) at each stage (calculated by adding the products ofTPs at each microhabitat times their relative cover). (B) Probability ofrecruitment through to the end of each stage (accumulated product ofsuccessive probabilities of recruitment). D = seed dispersal (arbi-trarily set at 1); V = seed viability; Ss = post-dispersal seed survival;E = seedling emergence; Sds = seedling survival; Sp1s = 2-year-oldsapling survival; Sp2s = 3-year-old sapling survival. F, forest sites; SL,stony-slope sites; SN, Sierra Nevada; SB, Sierra de Baza.
Spatial dynam
ics of recruitment at different scales
1135
© 2008 T
he Author. Journal com
pilation © 2008 B
ritish Ecological Society, Journal of E
cology, 96, 1128–
1140
Fig. 3. Diagram of the spatial dynamics of recruitment in (a) forest sites and (b) stony-slope sites. Each column represents recruitment dynamics in one of the microhabitats considered. Ovals represent thestages and rectangles the processes of recruitment analyzed. Values shown are process-specific transition probabilities (TPs). OQ represents the overall quality of a microhabitat for recruitment of a dispersedseed (calculated as the product of all TPs but D). OPR represents the overall probability of recruitment at each microhabitat (calculated as D*OQ). The highest TP in each process is highlight with a widebox line. D = seed dispersal; V = seed viability; Ss = post-dispersal seed survival; E = seedling emergence; Sds = seedling survival; Sp1s = 2-year-old sapling survival; Sp2s = 3-year-old sapling survival.
1136L
. Góm
ez-Aparicio
© 2008 T
he Author. Journal com
pilation © 2008 B
ritish Ecological Society, Journal of E
cology, 96, 1128–
1140
Fig. 3 Continued

Spatial dynamics of recruitment at different scales 1137
© 2008 The Author. Journal compilation © 2008 British Ecological Society, Journal of Ecology, 96, 1128–1140
much larger proportion than the one found by Clark et al.(1999) (ca. 30%) in their review of recruitment limitation‘biased towards studies of temperate deciduous and borealforest’ (Clark et al. 1999, p. 3). Moreover, about half ofthe studies also included habitat as a source of variation,comparing recruitment patterns in forest vs. shrubland (e.g.Herrera et al. 1994; Gómez et al. 2003), shrubland vs.herbaceous vegetation (e.g. Calviño-Cancela 2002; Travesetet al. 2003), or mainland vs. island (e.g. Lázaro et al. 2006).All of these studies found habitat-specific patterns ofrecruitment, between-habitat differences in OPR being aslarge as 75-fold (Pulido & Díaz 2005). However, because onlyone of the studies considered more than one site per habitattype (Acácio et al. 2007), it is not possible to discern to whatextent patterns interpreted as habitat-specific were in factsite-specific.
Results from the Acer study clearly showed that recruit-ment dynamics had a strong convergence among sites of thesame habitat. Differential OPR among habitats was mostlythe consequence of large differences in the values of criticalTPs, and not the result of habitat-specific patterns of uncou-pling. In fact, the identity of the limiting TPs was stronglyconsistent at the regional scale, mainly seed viability, seedlingemergence and seedling survival. However, seed viability wassimilarly low at all sites, whereas seedling emergence andsurvival were much higher in forests, eventually leading to aOPR one order higher at forest sites (ca. 1%) than at stony-slope sites (ca. 0.1%; Fig. 1). These differences in seedlingestablishment at the regional scale are presumably the resultof a habitat-specific abiotic filter: substrate characteristics.Thus, the rocky and mobile substrate distinctive of the stonyslopes probably hampered seedling emergence because of apoor root-soil contact, implying an additional risk ofmortality for seedlings, whereas the well-developed soil offorest sites assured a higher emergence and survival success(Gómez-Aparicio et al. 2005a).
The remaining processes in the Acer recruitment cycle,related to seed and sapling survival, had a much smallerimpact on the magnitude and spatial patterns of recruitmentat the regional scale because of the combination of both highTPs and low inter-site variability. Post-dispersal seed survivalwas slightly lower in forest populations, in agreement with thereported preference of rodents for covered (e.g. forests) vs.open habitats (e.g. stony-slopes) (Webb & Willson 1985;Hulme 1994; Rey et al. 2002). However, it was above 80% atthe four sites. Similarly, survival was generally > 70% for2-year saplings, and > 85% for 3-year saplings. This resultimplies that seedlings that overcome the first year have a highprobability of persistence in the population, their risk ofmortality decreasing as they become older (see also De Steven1991; Herrera et al. 1994; Houle 1994; Rey & Alcántara2000). In summary, the low seedling emergence and survivalprobabilities that characterized stony-slope sites in compari-son with forest sites overrode other more subtle among-sitedifferences in the seed phase and cascaded through thesapling stage to the end of the recruitment cycle generating astrong habitat-specific overall recruitment success.
SPATIAL DYNAMICS OF RECRUITMENT AT THE LOCAL SCALE: ON THE IDENTITY OF LOW- AND HIGH-QUALITY MICROHABITATS FOR ESTABLISHMENT
The integrative analyses of the different processes comprisingthe Acer regeneration cycle indicates the existence of strongmicrohabitat-specific recruitment patterns. This result wasshared by 17 of the 18 species reviewed for which informationon recruitment patterns at the local scale was available (i.e. allspecies but Lavandula stoechas, which failed to recruit in allmicrohabitats). Microhabitat specificity therefore emergesas a common feature of recruitment dynamics in Mediterra-nean environments across life forms (trees, shrubs, herbs)and dispersal systems (zoochorous, anemochorous, myrme-cochorous, ballistic). However, not all processes in the recruit-ment cycle are similarly influenced by the microhabitat. Inmost species, including Acer, spatial patterns of recruitmentat the microhabitat scale were determined largely by differencesin three key processes: seed rain, seedling emergence andseedling survival.
In the reviewed studies, seed rain was repeatedly higherunder conspecifics than in any other microhabitat. However,this pattern was weaker in endozoochorous species because ofnon-random foraging by frugivorous birds. For these species,the microhabitats receiving the largest seed rain are usuallythe plant species contributing most fruits to the diet of majordispersers (i.e. Herrera et al. 1994, García et al. 2005). On theother hand, seedling emergence and survival were much lowerin open interspaces than under shrubs, consistently for the 10tree species reviewed (including Acer) and the two herbaceousspecies (Appendix S1). Moreover, this pattern of low- vs.high-quality microhabitats was fairly consistent at theregional scale (but see Garrido et al. 2007 for a case of inter-site variability). The positive effect of the vegetation cover -andespecially of shrubs- on seedling emergence and survival is awell-known facilitation interaction in Mediterranean forestsand shrublands, and it is related to the amelioration of stress-ful climatic conditions (mainly summer drought) by canopycover (Gómez-Aparicio et al. 2004; Padilla & Pugnaire2006). Among the 7 shrub species reviewed, however, therewas not such a consistent pattern in the identity of the bestmicrohabitats for recruitment. Indeed, for some shrub species,open spaces were either the best microhabitat for seedlingestablishment (Calviño-Cancela 2002) or showed qualitysimilar to that of covered microhabitats (Traveset et al. 2003;Lázaro et al. 2006; Rodríguez-Pérez & Traveset 2007). Thisresult is influenced by the fact that, in general, seedlingsurvival of shrub species was not so dependent on shade, andthat post-dispersal seed predation (also an important processfor several of these species) was higher in covered micro-habitats than in the open. Therefore, facilitation mechanismsclearly determined small-scale spatial patterns of recruitmentin trees and herbs but not in shrub species.
It is important to highlight that although shrubs were ingeneral high-quality microhabitats for establishment, not allshrub species had the same positive effect on recruitment. Forexample, the recruitment probability of an Acer seed dispersed

1138 L. Gómez-Aparicio
© 2008 The Author. Journal compilation © 2008 British Ecological Society, Journal of Ecology, 96, 1128–1140
under Berberis in F-SN was ca. two times lower than underOnonis and three times lower than under Juniperus. Similarly,in SL-SB recruitment probability under Berberis wasmuch lower than under Juniperus. Therefore, spiny deciduousshrubs (i.e. Berberis, Prunus, Crataegus) seem to have worsequality for Acer early recruitment than cushion-shapedshrubs (i.e. Juniperus or Ononis). The limited nurse capacityof spiny deciduous shrubs has been reported elsewhere forseedling survival (Castro et al. 2004a; Gómez-Aparicio et al.2004, 2005c), and results shown here suggest that it alsoapplies to the overall early recruitment cycle. The lack ofcushion-shaped shrubs in F-SB could explain why this sitewas the only one where the microhabitat with the largestoverall quality (OQ) was not a shrub species. Differentialrecruitment expectancies among shrub species were alsofound in some of the reviewed studies. For example, Herreraet al. (1994) found a higher probability of seed and seedlingsurvival under non-fleshy fruited shrubs than under fleshy-fruited shrubs, Pulido & Díaz (2005) found higher seedlingrecruitment under late-successional shrubs (Phyllireaangustifolia) than under early-successional shrubs (Cistusladanifer), and Rey & Alcántara (2000) and Rodríguez-Pérez& Traveset (2007) found Pistacia lentiscus to be a worse nursethan other shrub species such as Phyllirea latifolia orErica multiflora. The mechanistic explanation behind thesespecies-specific differences is unknown, and deserves furtherinvestigation.
IS THE PATTERN OF RECRUITMENT PREDICTABLE FROM THAT OF THE SEED RAIN AT REGIONAL AND LOCAL SPATIAL SCALES?
Concordance among seed rain and recruitment is predicted tooccur when seed rain is more heterogeneous than post-dispersal losses, and/or when survival expectancies arespatially coupled across stages (García et al. 2005). Strongheterogeneity in the spatial pattern of seed dispersal iscommon in plant species, so that it might frequently obscurevariation in later demographic processes, thereby promotingconcordance (Jordano & Herrera 1995). However, the secondof the conditions for concordance to occur – spatially coupledsurvival expectancies across stages – is much more unlikely,with conflicts between microhabitat qualities for differentprocesses usually taking place in plant populations, therebypromoting discordance (Schupp 1995). The result is apotential for a whole gradient of relationships among thespatial patterns of seed rain and recruitment, from totaldiscordance to total concordance. Moreover, the degree ofconcordance can change among scales (García et al. 2005).
At the regional scale, the review of 19 Mediterraneanspecies indicates the absence of a clear pattern of concordanceamong the spatial distribution of seed rain and recruitment.Thus, among the studies that considered more than one site,40% (6 of 15) found higher recruitment in those sites withhigher seed availability (spatial concordance), whereas theremaining 60% found no relationship (spatial discordance).Concordance occurred only when there were large differences
in species fecundity or cover among sites, whereas modestdifferences were easily overshadowed by later-acting proc-esses in the recruitment cycle, mainly acting at the seedlingstage. The Acer study is a clear example of this latter situation.If we consider a regional pool of Acer seeds, about 32% of theseed rain occurred in the SL-SB, followed by F-SN (25%),F-SB (22%) and SL-SN (21%). This variation, not related tothe type of habitat, was totally overshadowed by large among-habitat differences in seedling emergence and survival, andconsequently the OPR and sapling density was much higherin forest than at stony-slope sites.
At the local scale, spatial discordance was found for even ahigher proportion of the reviewed species (12 of 18 species;67%), indicating that the uncoupling of seed rain andrecruitment at small spatial scales is a frequent pattern inMediterranean systems across life forms and dispersalsyndromes. Two main causes of uncoupling were identifiedamong the reviewed studies: disruptions at the seed bank level(e.g. secondary dispersal, seed mortality) that uncoupledpatterns of seed rain and emerged seedlings (Herrera et al.1994; Calviño-Cancela 2002; Hampe 2004), and disruptionsat the seedling/sapling level because of low survival in highemergence microhabitats (Rey & Alcántara 2000; Travesetet al. 2003; Castro et al. 2004b). The Acer study would fit intothis second group of studies. Seed density was positivelycorrelated to density of emerged seedlings, but not to long-term sapling distribution (Fig. 1). Indeed, the largest seed andseedling density occurred under Acer adults, whereas long-term sapling recruitment occurred mainly under shrubs. Thefact that the Acer microhabitat did not show the highestsapling density despite its higher OPR values could derivedfrom an underestimation of seedling and sapling mortalityin this microhabitat. In fact, Acer survival under conspecificadults is more variable in time and through ontogeny thanin any other microhabitat. Survival under conspecifics ismuch lower in dry years (i.e. 2000 and 2001) than wet years(i.e. 2002; Gómez-Aparicio et al. 2005a), and for old (5–16years) than young (2–4 years) saplings (Gómez-Aparicioet al. 2005b). OPR values calculated over a longer timeperiod, and therefore including further dry years (which arethe rule in the Mediterranean) and older saplings, wouldprobably be lower than those obtained here for the Acermicrohabitat. On the contrary, the reported consistent highquality of shrubs for Acer recruitment through time andontogeny (Gómez-Aparicio et al. 2005a,b) supports the accu-racy of the large OPR values (second after Acer) obtained forthis microhabitat and explains why sapling distribution inthis species is strongly associated with shrubs in most sites.
Across scales, a lack of spatial concordance (or inconsist-ency; García et al. 2005) was found. Within the 10 species forwhich seed rain and recruitment patterns were concordant atleast at one scale, only two species (22%) showed spatialconcordance at both spatial scales (Taxus baccata and Buxusbalearica; see Appendix S1). Of the eight remaining species,one half showed concordance only at the regional scale andthe other half only at the local scale. As suggested by Garcíaet al. (2005), these inconsistencies may be the result of an

Spatial dynamics of recruitment at different scales 1139
© 2008 The Author. Journal compilation © 2008 British Ecological Society, Journal of Ecology, 96, 1128–1140
unbalanced distribution between scales of the total variancein seed-rain patterns, with concordance across scales (orconsistency) occurring only when variance in seed rain isproportionately distributed among scales. As suggested in thisstudy, this seems to be rarely the case in Mediterranean systems.
CONCLUSIONS
Results found for Acer opalus subsp. granatense and 19additional species clearly indicate that spatial patterns ofearly recruitment of Mediterranean trees, shrubs, herbs andgrasses are strongly affected by the heterogeneity of the environ-ment at different spatial scales, with habitat-specific andmicrohabitat-specific patterns of recruitment being the rulefor most species. Moreover, spatial patterns of recruitmentwere frequently not related to the spatial patterns of seeddispersal, especially at small spatial scales (among microhab-itats). This lack of concordance indicates that high speciesabundance and seed production at a specific location cannotbe considered an assurance of successful recruitment. Fromthe perspective of the debate of seed vs. microsite limitation, itsuggests that the recruitment process of many Mediterraneanspecies is limited largely by the availability of safe sites.Although microsite and seed limitation are almost certainlynever exclusive (Münzbergová & Herben 2005; Moore &Elmendorf 2006), the influence of the process of dispersal onthe spatial patterns and demography of plant populationscould be limited in stressful and heterogeneous environments(as Mediterranean systems) where environmental filters onlyallow recruitment in a small fraction of the landscape.
Safe sites for Mediterranean species might be defined asthose where seedlings have a larger probability of surviving.In fact, no other process had repeatedly (from species tospecies) as large an influence in determining spatial patternsand overall recruitment as seedling survival, because ithad both very low TPs and high spatial variability at bothregional and local scales. In the case of Acer, because of theoverwhelming importance of the seedling stage in determin-ing recruitment patterns, the long-term sapling distributiondid not resemble that of the seed rain, but that of the distri-bution of the best patches for seedling emergence andsurvival: forests at the habitat scale and shrubs at the localscale. This result seems to apply to Mediterranean speciesacross life forms and dispersal syndromes, although morestudies focusing on other than zoochorous woody species arenecessary for a proper assessment of the differences amongfunctional groups in spatial recruitment patterns. If we are topreserve the distribution and abundance of Mediterraneanspecies in the face of environmental changes, we need toexplicitly recognize and make use of the strong patch-specificity that characterizes the recruitment process ofthese species at all scales.
Acknowledgements
I am most indebted to Regino Zamora and Jose María Gómez for theirinvaluable intellectual and logistical support during this study. I thank theConsejería de Medio Ambiente (Junta de Andalucía) and the Directors of
the Sierra Nevada National Park and the Sierra de Baza Natural Park forpermission to do fieldwork. Many thanks also to Sergio de Haro, Elena Barazaand Jose Antonio Hódar for field assistance. David Nesbitt improved theEnglish version. This study was supported by a grant PFPU-MECD to L.G.A.,the Andalusian government grant (PAI) rnm-220 and by the coordinatedSpanish MEC projects HETEROMED (REN2002-04041-CO2-01/GLO) andDINAMED (CGL2005-05830-C03-03). This research is part of REDBOME(www.ugr.es/~redbome/) and GLOBIMED (www.globimed.net) networks onforest ecology.
References
Acácio, V., Holmgren, M., Jansen, P.A. & Schrotter, O. (2007) Multiple recruitmentlimitation causes arrested succession in Mediterranean cork oak systems.Ecosystems, 10, 1220–1230.
Blanca, G., Cabezudo, B., Hernández-Bermejo, J.E., Herrera, C.M., Molero,J., Muñoz, J. & Valdés, B. (2000) Libro Rojo de Flora Amenazada deAndalucía II: Especies Vulnerables. Consejería de Medio Ambiente, Junta deAndalucía, Sevilla.
Calviño-Cancela, M. (2002) Spatial patterns of seed dispersal and seedlingrecruitment in Corema album (Empetraceae): the importance of unspecializeddispersers for regeneration. Journal of Ecology, 90, 775–784.
Canham, C.D., Finzi, A.C., Pacala, S.W. & Burbank, D.H. (1994) Causes andconsequences of resource heterogeneity in forests: interspecific variation inlight transmission by canopy trees. Canadian Journal of Forest Research, 24,337–349.
Castro, J., Zamora, R., Hódar, J.A., Gómez, J.M. & Gómez-Aparicio, L.(2004a) Benefits of using shrubs as nurse plants for reforestation in Medi-terranean mountains: a 4-year study. Restoration Ecology, 12, 352–358.
Castro, J., Zamora, R., Hódar, J.A. & Gómez, J.M. (2004b) Seedling establishmentof a boreal tree species (Pinus sylvestris) at its southernmost distributionlimit: consequences of being in a marginal, Mediterranean habitat. Journalof Ecology, 92, 266–277.
Chambers, J.C. (1995) Relationships between seed fates and seedling establish-ment in an alpine ecosystem. Ecology, 76, 2124–2133.
Clark, J.S., Beckage, B., Camill, P., Cleveland, B., Hille Ris Lambers, J.,Lichter, J., Mclachlan, J., Mohan, J. & Wyckoff, P. (1999) Interpretingrecruitment limitation in forests. American Journal of Botany, 86, 1–16.
De Steven, D. (1991) Experiments on mechanisms of tree establishment inold-field succession: seedling survival and growth. Ecology, 72, 1076–1088.
Farley, R.A. & Fitter, A.H. (1999) Temporal and spatial variation in soilresources in a deciduous woodland. Journal of Ecology, 87, 688–696.
García, D. (2001) Effects of seed dispersal on Juniperus communis recruitmenton a Mediterranean mountain. Journal of Vegetation Science, 12, 839–848.
García, D., Obeso, J.R. & Martínez, I. (2005) Spatial concordance betweenseed rain and seedling establishment in bird-dispersed trees: does the scalematter? Journal of Ecology, 93, 693–704.
Garrido, J.L., Rey, P.J. & Herrera, C.M. (2007) Regional, local, and temporalvariation in seedling emergence, mortality, and recruitment of a perennialherb in a Mediterranean mountain habitat. Plant Ecology, 190, 109–121.
Gleiser, G., Picher, M.C., Veintimilla, P., Martínez, J. & Verdú, M. (2004) Seeddormancy in relation to seed storage behaviour in Acer. Botanical Journal ofthe Linnean Society, 145, 203–208.
Gleiser, G., Verdú, M., Segarra-Moragues, J.G., González-Martínez, S.C. &Pannell, J.R. (2008) Disassortative mating, sexual specialization and theevolution of gender dimorphism in heterodichogamous Acer opalus.Evolution, 62, 1676–1688.
Gómez, J.M. (2004) Importance of burial and microhabitat on Quercus ilexearly recruitment: non-additive effects on multiple demographic processes.Plant Ecology, 172, 287–297.
Gómez, J.M., García, D. & Zamora, R. (2003) Impact of vertebrate acorn- andseedling-predators on a Mediterranean Quercus pyrenaica forest. ForestEcology and Management, 180, 125–134.
Gómez-Aparicio, L. (2004) Papel de la heterogeneidad en la regeneracióndel Acer opalus subsp. granatense en la montaña mediterránea: implicacionespara la conservación y restauración de sus poblaciones. PhD Thesis.University of Granada, Granada, Spain.
Gómez-Aparicio, L., Zamora, R., Gómez, J.M., Hódar, J.A., Castro, J. &Baraza, E. (2004) Applying plant facilitation to forest restoration: ameta-analysis of the use of shrubs as nurse plants. Ecological Applications,14, 1128–1138.
Gómez-Aparicio, L., Gómez, J.M. & Zamora, R. (2005a) Microhabitats shiftrank in suitability for seedling establishment depending on habitat type andclimate. Journal of Ecology, 93, 1194–1202.
Gómez-Aparicio, L., Zamora, R. & Gómez, J.M. (2005b) The regeneration

1140 L. Gómez-Aparicio
© 2008 The Author. Journal compilation © 2008 British Ecological Society, Journal of Ecology, 96, 1128–1140
status of the endangered Acer opalus subsp. granatense throughout itsgeographical distribution in the Iberian Peninsula. Biological Conservation,121, 195–206.
Gómez-Aparicio, L., Valladares, F., Zamora, R. & Quero, J.L. (2005c)Response of tree seedlings to the abiotic heterogeneity generated by nurseshrubs: an experimental approach at different scales. Ecography, 28, 757–768.
Gómez-Aparicio, L., Gómez, J.M. & Zamora, R. (2007) Spatiotemporal patterns ofseed dispersal in a wind-dispersed Mediterranean tree (Acer opalus subsp.granatense): implications for regeneration. Ecography, 30, 13–22.
Hampe, A. (2004) Extensive hydrochory uncouples spatiotemporal patterns ofseedfall and seedling recruitment in a ‘bird-dispersed’ riparian tree. Journalof Ecology, 92, 797–807.
Herrera, C.M., Jordano, P., López-Soria, L. & Amat, J.A. (1994) Recruitmentof a mast-fruiting, bird-dispersed tree: bridging frugivore activity andseedling establishment. Ecological Monographs, 64, 315–344.
Houle, G. (1992) Spatial relationship between seed and seedling abundance andmortality in a deciduous forest of north-eastern North America. Journal ofEcology, 80, 99–108.
Houle, G. (1994) Spatiotemporal patterns in the components of theregeneration of four sympatric tree species: Acer rubrum, A. saccharum,Betula alleghaniensis and Fagus grandifolia. Journal of Ecology, 82, 39–53.
Hulme, P.E. (1994) Post-dispersal seed predation in grassland: its magnitudeand sources of variation. Journal of Ecology, 82, 645–652.
Hutchings, M.J., John, E.A. & Wijesinghe, D.K. (2003) Toward understandingthe consequences of soil heterogeneity for plant populations and communities.Ecology, 84, 2322–2334.
IUCN (2001) IUCN Red List Categories and Criteria v 3.1. IUCN, Gland.Jordano, P. & Herrera, C.M. (1995) Shuffling the offspring: uncoupling and
spatial discordance of multiple stages in vertebrate seed dispersal. Écoscience,2, 230–237.
Lázaro, A., Traveset, A. & Castillo, A. (2006) Spatial concordance at a regionalscale in the regeneration process of a circum-Mediterranean relict (Buxusbalearica): connecting seed dispersal to seedling establishment. Ecography,29, 683–696.
Legendre, P. (2004) Kendall Coefficient of Concordance: Global Test anda Posteriori Tests of Individual Judges – Program Kendall W User’s Guide.Département de Sciences Biologiques, Université de Montréal, Québec.
López-González, G. (1994) La Guía de Incafo de los Árboles y Arbustos de laPenínsula Ibérica. Editorial Incafo, Madrid.
Moore, K.A. & Elmendorf, S.C. (2006) Propagule vs. niche limitation: untan-gling the mechanisms behind plant species’ distributions. Ecology Letters, 9,797–804.
Münzbergová, Z. & Herben, T. (2005) Seed, dispersal, microsite, habitat andrecruitment limitation – identification of terms and concepts in studies oflimitations. Oecologia, 145, 1–8.
Nicotra, A.B., Chazdon, R.L. & Iriate, S.V.B. (1999) Spatial heterogeneity oflight and woody seedling regeneration in tropical wet forests. Ecology, 80,1908–1926.
Padilla, F.M. & Pugnaire, F.I. (2006) The role of nurse plants in the restoration ofdegraded environments. Frontiers in Ecology and the Environment, 4, 196–202.
Pickett, S.T., Cadenasso, M.L. & Jones, C.G. (2000) Generation of heteroge-neity by organisms: creation, maintenance and transformation. The Ecolog-ical Consequences of Environmental Heterogeneity (eds M.J. Hutchings, E.A.John & A.J.A. Stewart), pp. 33–52. Blackwell Science, Oxford.
Puerta-Piñero, C., Gómez, J.M. & Valladares, F. (2007) Irradiance and oakseedling survival and growth in a heterogeneous environment. ForestEcology and Management, 242, 462–469.
Pulido, F.J. & Díaz, M. (2005) Regeneration of a Mediterranean oak: awhole-cycle approach. Écoscience, 12, 92–102.
Quintana-Ascencio, P.F., Weekley, C.W. & Menges, E.S. (2007) Comparativedemography of a rare species in Florida scrub and road habitats. BiologicalConservation, 137, 263–270.
Rees, M., Mangel, M., Turnbull, L., Sheppard, A. & Briese, D. (2000) The effectsof heterogeneity on dispersal and colonization in plants. The EcologicalConsequences of Environmental Heterogeneity (ed. M.J. Hutchings, E.A. John& A.J.A. Stewart), pp. 237–266. Blackwell Science, Oxford.
Rey, P.J. & Alcántara, J.M. (2000) Recruitment dynamics of a fleshy-fruitedplant (Olea europaea): connecting patterns of seed dispersal to seedlingestablishment. Journal of Ecology, 88, 622–633.
Rey, P.J., Garrido, J.L., Alcántara, J.M., Ramírez, J.M., Aguilera, A., García,L., Manzaneda, A.J. & Fernández, R. (2002) Spatial variation in ant androdent post-dispersal predation of vertebrate-dispersed seeds. FunctionalEcology, 16, 773–781.
Rodríguez-Pérez, J. & Traveset, A. (2007) A multi-scale approach in the studyof plant regeneration: finding bottlenecks is not enough. Perspectives inPlant Ecology, Evolution and Systematics, 9, 1–13.
Russell, S.K. & Schupp, E.W. (1998) Effects of microhabitat patchiness on pat-terns of seed dispersal and seed predation of Cercocarpus ledifolius(Rosaceae). Oikos, 81, 434–443.
SAS (2002) SAS/STAT user’s Guide. Version 8.2. SAS Institute, Cary, NorthCarolina.
Schupp, E.W. & Fuentes, M. (1995) Spatial patterns of seed dispersal and theunification of plant population ecology. Écoscience, 2, 267–275.
Schupp, E.W. (1995) Seed-seedling conflicts, habitat choice, and patterns ofplant recruitment. American Journal of Botany, 82, 399–409.
Stewart, A.J.A., John, E.A. & Hutchings, M.J. (2000) The world is heterogeneous:ecological consequences of living in a patchy environment. The EcologicalConsequences of Environmental Heterogeneity (eds. M.J. Hutchings, E.A.John & A.J.A. Stewart), pp. 1–8. Blackwell Science, Oxford.
Traveset, A., Gulias, J., Riera, N. & Mus, M. (2003) Transition probabilitiesfrom pollination to establishment in a rare dioecious shrub species (Rhamnusludovici-salvatoris) in two habitats. Journal of Ecology, 91, 427–437.
Vilá, M. & Lloret, F. (2000) Seed dynamics of the mast seeding tussock grassAmpelodesmos mauritanica in Mediterranean shrublands. Journal ofEcology, 88, 479–491.
Wang, B.C. & Smith, T.B. (2002) Closing the seed dispersal loop. Trends inEcology and Evolution, 17, 379–385.
Webb, S.J. & Willson, M.F. (1985) Spatial heterogeneity in post-dispersalpredation on Prunus and Uvularia seeds. Oecologia, 67, 150–153.
Wenny, D.G. (2000) Seed dispersal, seed predation, and seedling recruitment ofOcotea endresiana (Lauraceae) in Costa Rica. Ecological Monographs, 70,331–351.
Zavaleta, E.S., Hulvey, K.B. & Fulfrost, B. (2007) Regional patterns ofrecruitment success and failure in two endemic California oaks. Diversityand Distributions, 13, 735–745.
Received 12 March 2008; accepted 16 July 2008Handling Editor: David Gibson
Supporting Information
Additional Supporting Information may be found in theonline version of this article:
Appendix S1. Summary of studies analyzing spatial patternsof early recruitment in Mediterranean plant species.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for the article.
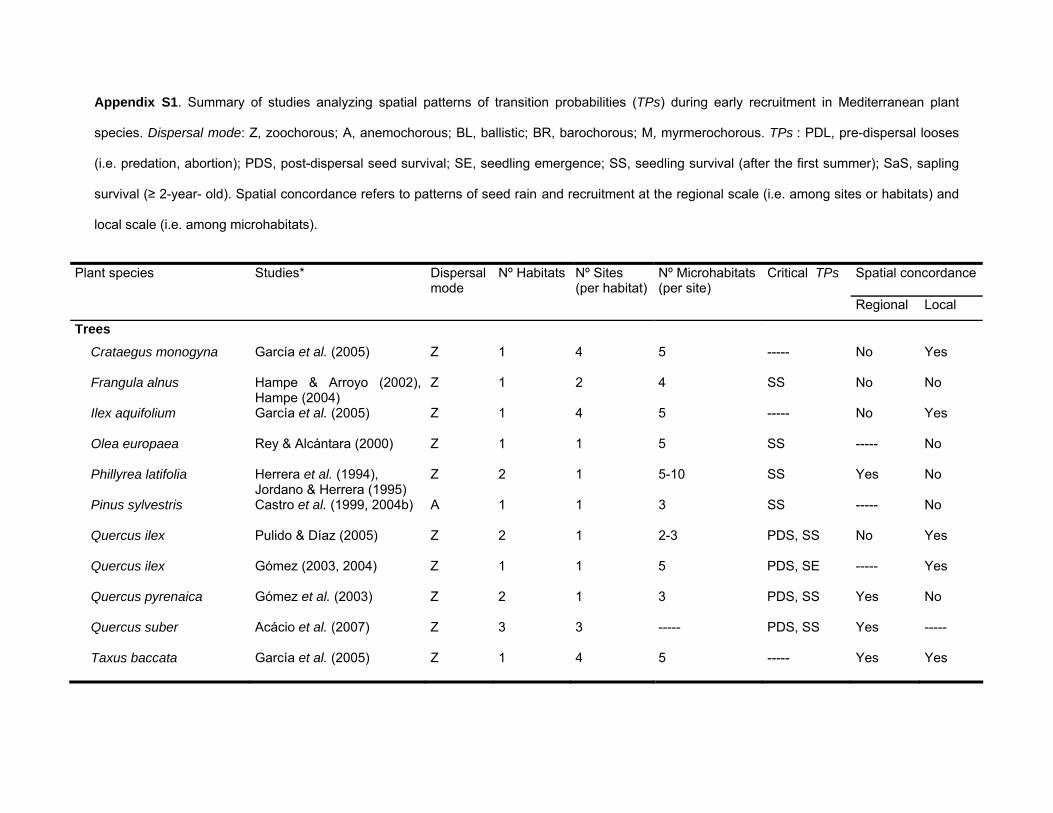
Appendix S1. Summary of studies analyzing spatial patterns of transition probabilities (TPs) during early recruitment in Mediterranean plant
species. Dispersal mode: Z, zoochorous; A, anemochorous; BL, ballistic; BR, barochorous; M, myrmerochorous. TPs : PDL, pre-dispersal looses
(i.e. predation, abortion); PDS, post-dispersal seed survival; SE, seedling emergence; SS, seedling survival (after the first summer); SaS, sapling
survival (≥ 2-year- old). Spatial concordance refers to patterns of seed rain and recruitment at the regional scale (i.e. among sites or habitats) and
local scale (i.e. among microhabitats).
Plant species Studies* Dispersal mode
Nº Habitats Nº Sites (per habitat)
Nº Microhabitats(per site)
Critical TPs Spatial concordance
Regional Local
Trees
Crataegus monogyna García et al. (2005) Z 1 4 5 ----- No Yes
Frangula alnus Hampe & Arroyo (2002), Hampe (2004)
Z 1 2 4 SS No No
Ilex aquifolium García et al. (2005) Z 1 4 5 ----- No Yes
Olea europaea Rey & Alcántara (2000) Z 1 1 5 SS ----- No
Phillyrea latifolia Herrera et al. (1994), Jordano & Herrera (1995)
Z 2 1 5-10 SS Yes No
Pinus sylvestris Castro et al. (1999, 2004b) A 1 1 3 SS ----- No
Quercus ilex Pulido & Díaz (2005) Z 2 1 2-3 PDS, SS No Yes
Quercus ilex Gómez (2003, 2004) Z 1 1 5 PDS, SE ----- Yes
Quercus pyrenaica Gómez et al. (2003) Z 2 1 3 PDS, SS Yes No
Quercus suber Acácio et al. (2007) Z 3 3 ----- PDS, SS Yes -----
Taxus baccata García et al. (2005) Z 1 4 5 ----- Yes Yes
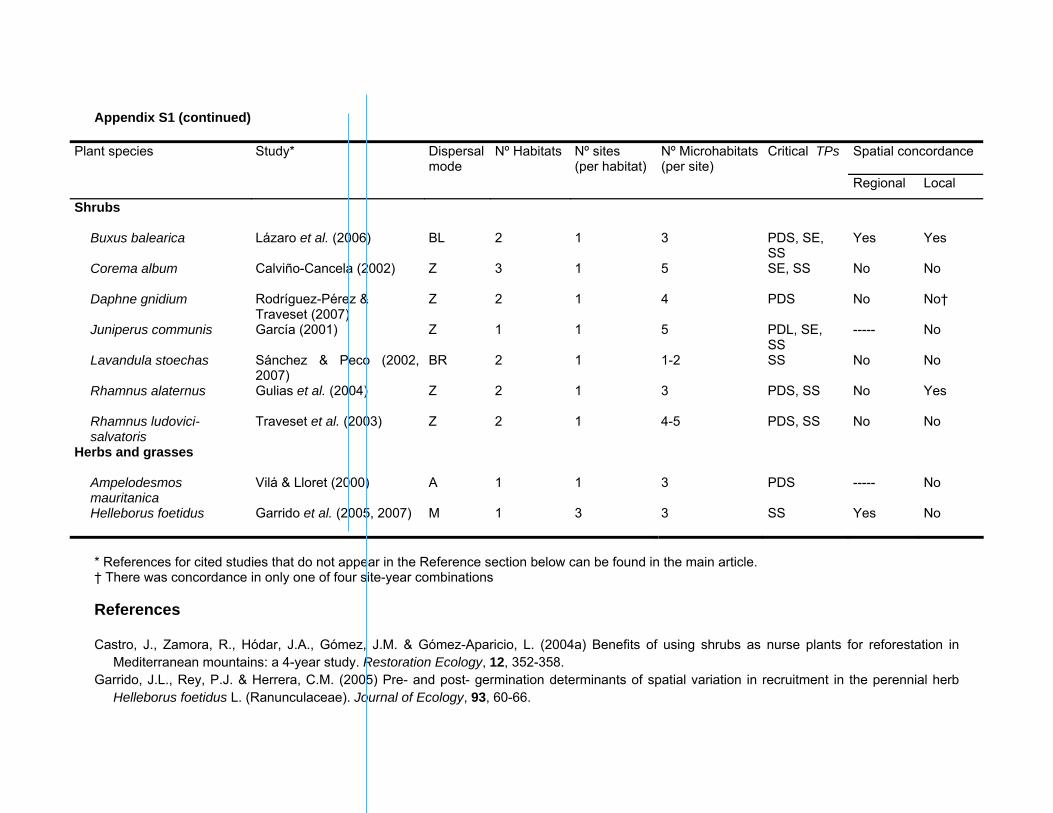
Appendix S1 (continued)
Plant species Study* Dispersal mode
Nº Habitats Nº sites (per habitat)
Nº Microhabitats(per site)
Critical TPs
Spatial concordance
Regional Local
Shrubs
Buxus balearica Lázaro et al. (2006) BL 2 1 3 PDS, SE, SS
Yes Yes
Corema album Calviño-Cancela (2002) Z 3 1 5 SE, SS No No
Daphne gnidium Rodríguez-Pérez & Traveset (2007)
Z 2 1 4 PDS No No†
Juniperus communis García (2001) Z 1 1 5 PDL, SE, SS
----- No
Lavandula stoechas Sánchez & Peco (2002, 2007)
BR 2 1 1-2 SS No No
Rhamnus alaternus Gulias et al. (2004) Z 2 1 3 PDS, SS No Yes
Rhamnus ludovici-salvatoris
Traveset et al. (2003) Z 2 1 4-5 PDS, SS No No
Herbs and grasses
Ampelodesmos mauritanica
Vilá & Lloret (2000) A 1 1 3 PDS ----- No
Helleborus foetidus Garrido et al. (2005, 2007) M 1 3 3 SS Yes No
* References for cited studies that do not appear in the Reference section below can be found in the main article. † There was concordance in only one of four site-year combinations References Castro, J., Zamora, R., Hódar, J.A., Gómez, J.M. & Gómez-Aparicio, L. (2004a) Benefits of using shrubs as nurse plants for reforestation in
Mediterranean mountains: a 4-year study. Restoration Ecology, 12, 352-358. Garrido, J.L., Rey, P.J. & Herrera, C.M. (2005) Pre- and post- germination determinants of spatial variation in recruitment in the perennial herb
Helleborus foetidus L. (Ranunculaceae). Journal of Ecology, 93, 60-66.

Gómez, J.M. (2003) Spatial patterns in long-distance dispersal of Quercus ilex acorns by jays in a heterogeneous landscape. Ecography, 26, 573-584.
Gulias, J., Traveset, A., Riera, N. & Mus, M. (2004) Critical stages in the recruitment process of Rhamnus alternus L. Annals of Botany, 93, 723-731.
Hampe, A. (2004) Extensive hydrochory uncouples spatiotemporal patterns of seedfall and seedling recruitment in a ‘bird-dispersed’ riparian tree. Journal of Ecology, 92, 797-807.
Sánchez, A.M. & Peco, B. (2002) Dispersal mechanisms in Lavandula stoechas subsp. pedunculata: autochory and endozoochory by sheep. Seed Science Research, 12, 101-111.
Sánchez, A.M. & Peco, B. (2007) Lack of recruitment in Lavandula stoechas subsp. pedunculata: a case of safe-limitation. Acta Oecologica, 31, 32-39.





























