Embed Size (px)
Citation preview

http://jhc.sagepub.com/Journal of Histochemistry & Cytochemistry
http://jhc.sagepub.com/content/32/2/172The online version of this article can be found at:
DOI: 10.1177/32.2.6198353
1984 32: 172J Histochem CytochemP M Lansdorp, T H van der Kwast, M de Boer and W P Zeijlemaker
markers.Stepwise amplified immunoperoxidase (PAP) staining. I. Cellular morphology in relation to membrane
Published by:
http://www.sagepublications.com
On behalf of:
Official Journal of The Histochemical Society
can be found at:Journal of Histochemistry & CytochemistryAdditional services and information for
http://jhc.sagepub.com/cgi/alertsEmail Alerts:
http://jhc.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Feb 1, 1984Version of Record >>
by guest on November 23, 2014jhc.sagepub.comDownloaded from by guest on November 23, 2014jhc.sagepub.comDownloaded from

‘Supported in part by the Foundation for Medical Research FUNGO,
which is subsidized by the Netherlands Organization for the Advance-
ment of Pure Research (ZWO).
172
0022-1554/84/53.00
The Journal of Histochemistry and CytochemistryCopyright © 1984 by The Histochemical Society, Inc.
Vol. 32, No. 2, pp. 172-178, 1984Printed in U.S.A.
Stepwise Amplified Immunoperoxidase (PAP)Staining. I. Cellular Morphology in Relation toMembrane Markers’
PETER M. LANSDORP, THEO H. VAN DER KWAST, MARTIN DE BOER, and
WIM P. ZEULEMAKER
Central Laboratory of the Netherlands Red Cross Blood Transfusion Service and Laboratory for Experimental and Clinical Immunolo©y
ofthe Unizersity ofAmsterdarn. Amsterdam (P.M.L.: M.D.B. W.P.Z. and Department ofPathology. Erasmus University. Rotterdam
(T.H.z’.d.K.), The Netherlands
Received for publication January 31, 1983 and in revised form June 3, 1983; accepted July 27, 1983 (OA 83-108)
A novel procedure for the assay of monoclonal antibodiesis described. The technique is based on a combination of
three principles. Unlabeled (sheep) antiserum to mouseimmunoglobulin (Ig) and complexes of peroxidase with
mouse monoclonal antiperoxidase (monoclonal PAP corn-
plexes) are used as reagents in a variant of the unlabeledantibody enzyme (PAP) method, described by Sternberger.
The amount of peroxidase eventually bound to a mono-clonal antibody can be varied over a wide range by repe.
tition of incubation cycles with anti-mouse Ig and mono-
clonal PAP complexes. During the assay, incubations andwash steps are performed by immersion ofwhole slides. Theinfluence of repetitive incubation cycles with anti-mouseIg and monoclonal PAP complexes on background stain-
ing and detection of monoclonal antibodies at low con-centrations was quantitated in a model system. At a givenprimary antibody concentration, a linear relationship wasfound between peroxidase activity and the number of in-cubation cycles. Application of the technique to the de-
Introduction
Monoclonal antibodies are widely used in essentially all dis-
ciplines of biomedical sciences. To detect binding of mono-
clonal antibodies to cells, immunofluorescence in combination
with analysis of staining, e.g. , by flow cytofluorometry, is ex-
tremely useful. However, poor morphology and instability of
labeled preparations are major disadvantages of immunoflu-
orescence techniques, especially when the cells under study
are present as a minor population (Taylor and Chir, 1978;
Laurent et al., 1980). Mason et al. (1975) have shown that
tection of monoclonal antibodies bound to cell-surface an-tigens is described. Peripheral blood cells were labeled insuspension with monoclonal antibodies. Cytocentrifugepreparations of labeled cells were prepared, and such prep-arations were fixed before stepwise-amplified PAP stain-ing. Cells showed intense specific staining. Morphological
detail of stained and unstained cells is preserved, allowingmorphological analysis of labeled cells and rapid analysis
of monoclonal antibody specificity. Because the reagentsused in the assay can be produced in large quantities with
uniform quality, the technique can be readily automated.
This, together with the possibility to increase the sensi-tivity of antibody detection in a controlled, stepwise fash-ion to levels that cannot be reached with “single-step” tech-
niques, may further expand the applications of monoclonal
antibodies.
KEY WORDS: Monoclonal antibody detection; Monoclonal per-
oxidase-antiperoxidase (PAP) complexes; Immunoperoxidase
procedures; Immunocytochemistry; Immunocytology.
immunoperoxidase techniques can be used as an alternative
to immunofluorescence for staining of intracellular leukocyte
antigens in peripheral blood and bone marrow smears. In a
previous article, we described immunoperoxidase procedures
for the detection of monoclonal antibodies against cell-surface
antigens (Lansdorp et al., 1980). In that technique, the binding
ofmonoclonal antibodies to cells fixed in the wells of microtest
plates is assessed by a variant of the unlabeled anti-
body-enzyme method (Sternberger, 1975), with monoclonal
anti-horseradish peroxidase antibod ies complexed with per-
oxidase (PAP complexes) as a reagent.
The use of monoclonal PAP complexes in the unlabeled
antibody-enzyme method results in very low background
staining and intense specific staining. Furthermore, mono-
clonal PAP complexes can be produced in unlimited quantities
by guest on November 23, 2014jhc.sagepub.comDownloaded from

STEPWISE AMPLIFIED PAP STAINING 173
of constant quality, which results in high reproducibility. Al-
though analysis of individual cells in the microtest plates is
possible, the technique was originally developed for screening
of hybridoma supernatants and it is not readily applicable for
immunocytology. Two modifications were necessary to yield
satisfactory morphology in combination with clear immuno-
peroxidase staining. First, cells are labeled with monoclonal
antibodies in suspension before cytocentrifuge spreading and
fixation (Mason et al., 1975; Brown et al., 1979). This se-
quence is essential, because not all antigenic determinants re-
main after the relatively harsh fixation required to produce
satisfactory morphology. Second, the intensity of the perox-
idase staining is increased to give sufficient contrast between
stained and unstained cells. This is achieved by repeating the
incubation steps of the unlabeled antibody-enzyme method
used (Figure 1 ). The effect of repetitive incubation steps on
the peroxidase activity linked to limiting amounts of mono-
clonal antibody bound to cell-surface antigens was quantitated.
Application of the findings to immunoperoxidase staining of
peripheral blood cells labeled with monoclonal antibodies is
described.
Materials and Methods
Monoclonal Antibodies
D5, an 1gM monoclonal antibody defining a membrane-associateddeterminant that, within the hemopoietic system, seems to be confined
to the granulocytic cell lineage (Majdic et al., 1981), was a gift from
Dr. W. Knapp (Institute of Immunology, Vienna, Austria).
Figure 1. Schematic presentation of stepwise amplified irnmunope-
roxidase staining: (A) Primary (mouse) monoclonal antibody bound
tO antigen; (B) (sheep) antibody against mouse immunoglobulin; (C)
(mouse) monoclonal antibody against peroxidase complexed with per-
oxidase (PAP complexes). (1, 2, and 3) Number of incubation cycles.
(One incubation cycle represents incubation with anti-mouse 1g. wash-ing, incubation with PAP complexes, and washing again.)
63D3, an IgGI monoclonal antibody that shows strong binding to
adherent human peripheral blood mononuclear cells (Ugolini et al.,
1980), was obtained from BRL Molecular Diagnostics (Rockville, MD)
(cat. no. 9496 SM.
LICRILON Rb, an IgGI monoclonal antibody with specificity for
glycophorin A of human erythrocytes (Anstee and Edwards, 1982),
was a gift from Dr. PAW. Edwards (Ludwig Institute for Cancer
Research, Surrey, UK).
Leu 3a, an IgGI monoclonal antibody with specificity for human
T-helper/inducer cell antigen (Evans et al., 1981), was obtained from
Becton Dickinson (Sunny Vale, CA) (cat. no. 5320).OKIla, an IgG2 monoclonal antibody with specificity for HLA-
DR (Reinherz et al., 1979) was obtained from Ortho PharmaceuticalCorp. (Raritan, NJ).
W6/32, an IgG2A monoclonal antibody with specificity for a non-
polymorphic determinant present on HLA-A, HLA-B, and HLA-C
glycoproteins (Barnstable et al., l9�8), was obtained from Sera Lab.
(Sussex, UK) (cat. no. MASO15C).CLB GP IIIA/2 (C17), an IgGI monoclonal antibody specific for
a determinant Ofl platelet glycoprotein GP III A( Tetteroo et al. , 1 983),
was produced in our laboratory.
Anti-Mouse Ig
Antiserum against mouse immunoglobulins was prepared by injecting
sheep, at monthly intervals, with mouse myeloma 1gM )Bionetics) and
purified mouse IgG (prepared in our laboratory). As shown by mm-
munoelectrophoresis, this antiserum did not precipitate other mouse
serum proteins than IgG and 1gM (A. VIug, personal communication).
Before use, the antiserum was diluted 1:40 in Tris-buffered saline
(TBS, 0. 1 5 M NaC1, 0.05 M Tnis, pH 7.6) containing 5% (vlv) human
serum, 5% (v/v) normal sheep serum, and 0. 1 mg of thimerosallml
(Sigma Chemical Corp., St. Louis, MO) (cat. no. T5125).
Preparation and Analysis ofMonoclonal PAP
Complexes
Monoclonal IgGIK antibodies against horseradish peroxidase (HRP)
were purified from ascites of tumor-bearing mice by passage over
protein A-Sepharose (Pharmacia, Uppsala, Sweden), as described by
Ey et al. (1978). The purified antibody preparation (CLB-HRP-l) at
0.1 mg/mI in phosphate-buffered saline (PBS), was mixed with an
equal volume of PBS containing 1 mg of HRP/ml (Sigma, grade II),
10 mg of bovine serum albumin (BSA)/ml (Povite, Oss, The Neth-
erlands) and 0.2 mg of thimerosal/mI. This mixture was lyophmlized
or stored at 4#{176}Cfor up to 2 years without appreciable loss of activity.
The concentrations of peroxidase and antiperoxidase were chosen to
yield minimal dissociation of the immune complexes and minimal
nonspecific binding by free peroxidase. A suitable goat anti-mouse Ig
(cat. no. M1201) and the lyophilized PAP reagent (cat. no. Ml203)
may both be obtained from the Department of Immune Reagents of
the CLB (P. 0. Box 9190, 1006 AD Amsterdam, The Netherlands).
Agan electrophoresis was performed as described by Wieme ( 1976)Before use, PAP complexes were diluted I : 100 in Tris-buffered saline
containing 10% (v/v) normal sheep serum and 0. 1 mg of thimerosallml.
Cells
Granulocytes, monocytes, lymphocytes, erythrocytes, and platelets
were isolated from peripheral blood of a healthy individual by density
centnifugation and counterfiow-centrifugation elutriation, as described
by Ulmer and Flad (1979) and Figdor et al. (1981), respectively.
by guest on November 23, 2014jhc.sagepub.comDownloaded from

=..
A’B C
‘C
D
174 LANSDORP, VAN DER KWAST, DE BOER, ZEIJLEMAKER
Briefly, erythrocytes and granulocytes were first separated from other
cells by density centnifugation over Percoll (Pharmacia; d= 1.077 glcm5)
and subsequently from each other by counterfiow-centrifugation elu-
triation. Monocytes, lymphocytes, and blood platelets were separated
from each other by counterfiow-centrifugation elutriation. Purity of
the isolated cells, as determined by electronic sizing with a Coulter
counter ZF connected to a pnlse-height analyzer was higher than 90%.
For illustrative purposes, the cells were mixed at the following
ratios : granulocytes : monocytes : lymphocytes : erythrocytes : platelets
= 1: 1:2:6:8. Cells were washed once in ice-cold HF (Hank’s buffered
salt solution containing 25 mM N-2-hydroxyethylpiperazine-N’-2-
ethane-sulfonic acid (Hepes), 0. 1% (wlv) sodium azide, 0.38% (w/v)
tnisodium citrate, and 5% (v/v) fetal calf serum), then resuspended in
HF (0#{176}C)at a concentration of 10” nucleated cells/ml.
Labeling of Cells and Preparation of Slides
Cells were labeled with monoclonal antibodies for 45 mm at 0#{176}Cat
a concentration of 5 x 10� nucleated cells/mi. For each monoclonal
antibody the concentration was used that gave optimal labeling and
minimal background staining, namely, between 0.5 and 10 �.ig/ml for
all antibodies. Labeled cells were washed twice with HF at 0#{176}C.Sam-
pies of 50-100 x l0� cells were spun onto slides for 10 mm at 1000
rpm in a Shandon cytocentrifuge (Shandon Southern Products Ltd.,
Runcorn, Cheshire, UK).
Immunoperoxidase Procedures
Quantiation of the effect of repetitive incubation cycles.Terasaki test plates were coated with blood platelets as described
before (Lansdorp et al., 1982). W6/32 antibody at serial twofold di-lutions were added in 5 jil volumes to each well through a replicator
(Lansdorp et al., 1980). After incubation for 48 hr at 4#{176}C,the testplates were washed five times with PBS containing 0.2% (v/v) Tween
20 (PBS-T). To each well, 5 jil of sheep anti-mouse Ig was added.
Incubation at room temperature for 20 mm was followed by five
washes and incubation with monoclonal PAP complexes (5 jiL/well).
After incubation with PAP complexes for 20 mm at room temperature,
the test plates were washed five times with PBS-T. Next, the incu-
bation with anti-mouse Ig and PAP complexes was repeated on, al-
ternatively, the peroxidase substrate ortho-phenylene diamine was added.
Preparation of the substrate solution and quantitation of the peroxi-
dase reaction product after incubation at room temperature for 30
mm was performed as described (Lansdorp et al., 1980).
Immunoperoxidase staining of cells. The following protocolwas used for the labeling of individual cells (See Figure 5).
1. After cytocentrifugation, slides were placed in a microscope
slide rack. All incubations and washings were performed at
room temperature in staining jars by immersion ofwhole slides.
Air-dried slides (60 mm) were fixed with buffered formol ace-
tone (BFA; 20 mg NaHPO,1, 100 mg KH2PO.,, 45 ml acetone,
25 ml concentrated formalin, 30 ml distilled water, pH 7.0) for
30 sec and washed with distilled water as described by Mason
et al. (1975).
2. Blocking of endogenous peroxidase activity was achieved by
incubation for 30 mm with methanol containing 0.3% (w/v)
H202. Slides were washed twice by immersion (5 times) in jarscontaining TBS.
3. Incubation with anti-mouse Ig for 20 mm was followed by
washing (immersion 5 times in 4 jars containing TBS), incu-
bation with monoclonal PAP complexes for 20 mm, and wash-
ing again. The reagents were prepared as described previously.
4. Step 3 was repeated until five incubation cycles were completed.
5. The slides were then washed once with 0.05 M Tris, pH 7.5,and incubated for 8 mm with diaminobenzidine tetrahydro-
chloride (Sigma, grade II), 0.5 mg/mI, in 0.05 M Tris, pH 7.5,
containing 0.01% H2O2.
6. The slides were washed once with 0.05 M Tnis, pH 7,5, and
once with distilled water, counterstained with Mayer’s hema-
toxylin, dehydrated, and then mounted with malinol (Chroma,
Stuttgart, W. Germany) or DePeX (BDH Chemicals Ltd. Poole,
UK).
Results
Characteristics of monoclonal PAP complexes
Figure 2 shows the result of agar gel electrophoresis of anti-
peroxidase ascites, purified monoclonal antiperoxidase, mono-
clonal PAP complexes, and peroxidase. From the shift in elec-
trophoretic mobility of purified antibodies after addition of
(excess) peroxidase (Figure 2, lane C, arrow), it appears that
more than 95% of the immunoglobulin, recovered from pro-
tein A-Sepharose is capable of binding peroxidase and that,
under the conditions used, more than 95% of antiperoxidase
is completed with peroxidase. Excess peroxidase present in
the PAP reagent is visible as a faint band at the bottom of lane
C of Figure 2. Preliminary results (not shown) indicate that
diluted PAP can be used for at least 2 months.
Figure 2. Agar electrophoresis of undiluted ascites containing CLB-HRP-l (lane A); protein A-purified CLB-HRP-l, 8 mg/mI (lane B);monoclonal PAP complexes, 8 mg/ml (lane C), and horseradish per-
oxidase grade II, 8 mg/mI (lane D).
by guest on November 23, 2014jhc.sagepub.comDownloaded from
EC
Os
0
2x 4x 6x 8x lOx
number of incubation cycles
1.0
0.5
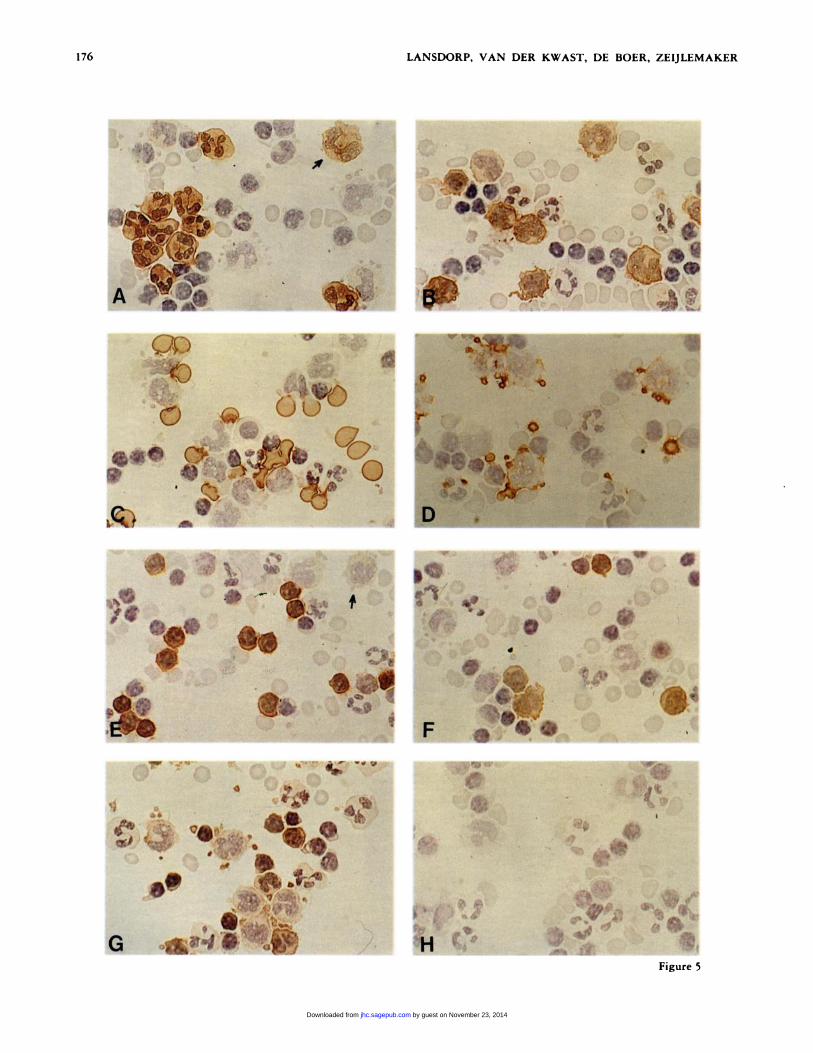
Figure 5. Stepwise amplified immunoperoxidase staining of periph-enal blood cells. Monoclonal antibodies used: (A) D5; (B) 63D3; (C)LICRJLON RiO; (D) CLB-GP lIlA; (E) Leu 3A; (F) OKIla; (G)
W6/32; (H) HF buffer only. To allow mutual comparison, all prepa-rations shown (one donor) were stained identically (five incubation
[w�i�J ng/mI cycles). Original magnification x 600.
STEPWISE AMPLIFIED PAP STAINING 175
Amplification of the Peroxidase Signal Linked to
Monodonal Antibody
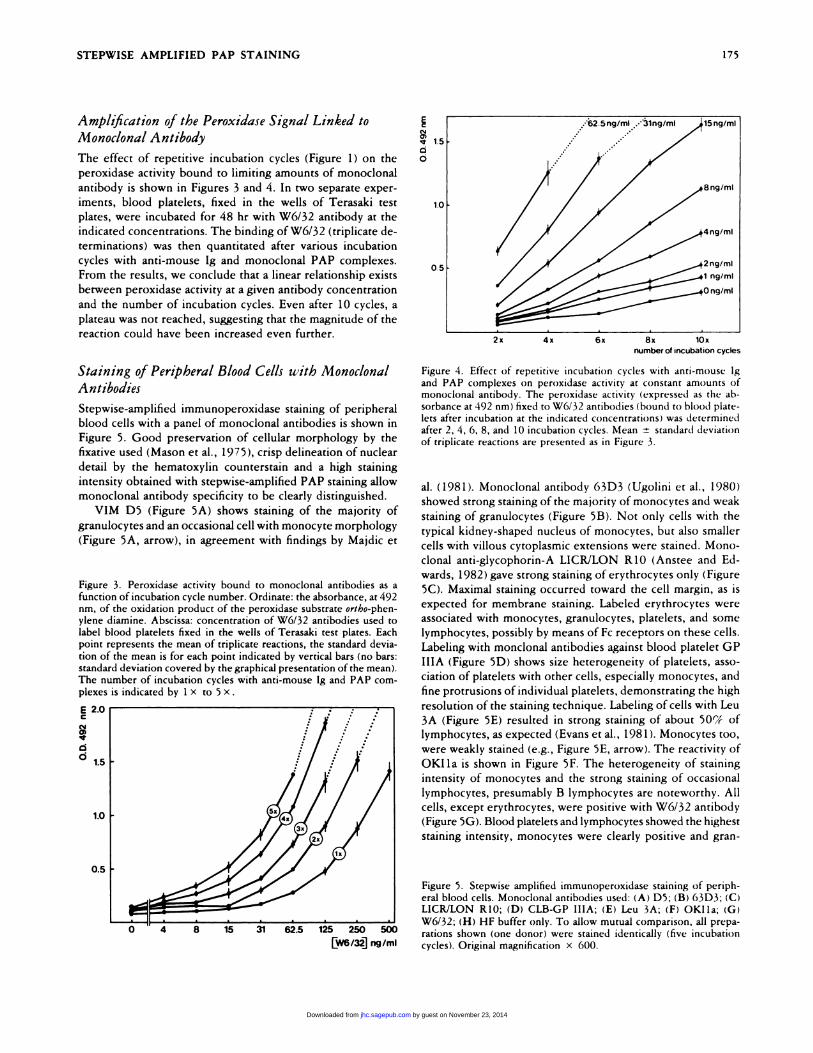
The effect of repetitive incubation cycles (Figure 1) on the
peroxidase activity bound to limiting amounts of monoclonal
antibody is shown in Figures 3 and 4. In two separate exper-
iments, blood platelets, fixed in the wells of Terasaki test
plates, were incubated for 48 hr with W6/32 antibody at the
indicated concentrations. The binding ofW6/32 (triplicate de-
terminations) was then quantitated after various incubation
cycles with anti-mouse Ig and monoclonal PAP complexes.
From the results, we conclude that a linear relationship exists
between peroxidase activity at a given antibody concentration
and the number of incubation cycles. Even after 10 cycles, a
plateau was not reached, suggesting that the magnitude of the
reaction could have been increased even further.
Staining of Peripheral Blood Cells with Monodonal
Antibodies
Stepwise-amplified immunoperoxidase staining of peripheral
blood cells with a panel of monoclonal antibodies is shown in
Figure 5. Good preservation of cellular morphology by the
fixative used (Mason et al., 1975), crisp delineation of nuclear
detail by the hematoxylin counterstain and a high staining
intensity obtained with stepwise-amplified PAP staining allow
monoclonal antibody specificity to be clearly distinguished.
VIM D5 (Figure 5A) shows staining of the majority of
granulocytes and an occasional cell with monocyte morphology
(Figure 5A, arrow), in agreement with findings by Majdic et
Figure 3. Penoxidase activity bound to monoclonal antibodies as afunction of incubation cycle number. Ordinate: the absorbance, at 492nm, of the oxidation product of the peroxidase substrate ortho-phen-
ylene diamine. Abscissa: concentration of W6/32 antibodies used to
label blood platelets fixed in the wells of Terasaki test plates. Eachpoint represents the mean of triplicate reactions, the standard devia-
tion of the mean is for each point indicated by vertical bans (no bars:
standard deviation covered by the graphical presentation of the mean).The number of incubation cycles with anti-mouse Ig and PAP corn-plexes is indicated by I x to 5 x.
E 2.0
I0 1.5
Figure 4. Effect of repetitive incubation cycles with anti-mouse Ig
and PAP complexes on peroxidase activity at constant amounts ofmonoclonal antibody. The peroxidase activity (expressed as the ab-
sorbance at 492 nm) fixed to W6/32 antibodies (bound to blood plate-lets after incubation at the indicated concentrations) was determinedafter 2, 4, 6, 8, and 10 incubation cycles. Mean ± standard deviation
of triplicate reactions are presented as in Figure 3.
al. (1981). Monoclonal antibody 63D3 (Ugolini et al., 1980)
showed strong staining of the majority of monocytes and weak
staining of granulocytes (Figure SB). Not only cells with the
typical kidney-shaped nucleus of monocytes, but also smaller
cells with villous cytoplasmic extensions were stained. Mono-
clonal anti-glycophorin-A LICR/LON RiO (Anstee and Ed-
wards, 1982) gave strong staining oferythrocytes only (Figure
SC). Maximal staining occurred toward the cell margin, as is
expected for membrane staining. Labeled erythrocytes were
associated with monocytes, granulocytes, platelets, and some
lymphocytes, possibly by means of Fc receptors on these cells.
Labeling with monclonal antibodies against blood platelet GP
lIlA (Figure SD) shows size heterogeneity of platelets, asso-
ciation of platelets with other cells, especially monocytes, and
fine protrusions ofindividual platelets, demonstrating the high
resolution of the staining technique. Labeling of cells with Leu
3A (Figure SE) resulted in strong staining of about 50% of
lymphocytes, as expected (Evans et al., 1981). Monocytes too,
were weakly stained (e.g. , Figure SE, arrow). The reactivity of
OKI1a is shown in Figure SF. The heterogeneity of staining
intensity of monocytes and the strong staining of occasional
lymphocytes, presumably B lymphocytes are noteworthy. All
cells, except erythrocytes, were positive with W6/32 antibody
(Figure SG). Blood platelets and lymphocytes showed the highest
staining intensity, monocytes were clearly positive and gran-
by guest on November 23, 2014jhc.sagepub.comDownloaded from
Q:�� i.�: ‘ #{149}�‘
�r,,� Cu
... .
�
0 ,,�, �
�‘:� . . � ,.
176 LANSDORP, VAN DER KWAST, DE BOER, ZEIJLEMAKER
. ,
I ‘ �
� �1-I �e
�0
�. � ... 14”
� �� ,� p,, ‘4
4”�, ‘: .
Figure 5
by guest on November 23, 2014jhc.sagepub.comDownloaded from

STEPWISE AMPLIFIED PAP STAINING 177
ulocytes were weakly stained, in agreement with findings by
Brown et al. (1979). When cells were not labeled with mono-
clonal antibodies (Figure SH), only hematoxylin-counten-
stained cells were seen.
Discussion
In this article, a new immunoperoxidase technique is described
that is based on a combination of three principles. These prin-
ciples are 1 ) application of monoclonal PAP complexes in the
unlabeled antibody-enzyme method, 2) amplification of stain-
ing intensity by repetitive incubation with the unlabeled an-
tibody-enzyme method, and 3) immersion ofwhole slides for
incubation and washing of cell preparations. The advantages
and disadvantages of immunoperoxidase techniques in corn-
panison with immunofluorescence techniques have been dis-
cussed by Taylor and Chir (1978) and Sternberger (1979). The
major advantage of the antibody-enzyme (PAP) method over
methods with enzyme-labeled antibodies is the low back-
ground staining of the PAP procedure, resulting in a high
sensitivity. In 1980, we described the use of monoclonal an-
tibodies against horseradish peroxidase for the preparation of
(mouse) PAP complexes (Lansdorp et al., 1980), and we showed
that such monoclonal PAP complexes can be used in the un-
labeled antibody-enzyme method described by Sternbergen
(1979). A major advantage ofour method is that large amounts
of monoclonal PAP complex of reproducible quality can easily
be prepared. The usefulness of rnonoclonal PAP complexes
for immunocytochemistry was recently confirmed in a corn-
parative study by Mason et al. (1982).
Several variants ofthe unlabeled antibody-enzyme method
with respect to the number of incubation cycles (Figure 1)
have been described. Vacca et al. (1980) used a “double” PAP
technique, which was more sensitive than the usual “single”
PAP technique (Ordronneau et al., 1981). Halverson et al.
( 1 98 1 ) state that repeating “bridge PAP cycles” twice was
necessary to clearly reveal the presence ofthe enzyme terminal
transferase in BS-fixed and paraffin-embedded human tissue.
The use of an extended number of incubation cycles of the
PAP technique has, to our knowledge, not been described
before. Amplification of fluorescence staining by repetition of
incubation with fluorescein-labeled antibodies and antiflu-
orescein antibodies was described by Schmitz and Kampa (1979)
and Bauman et al. ( 198 1 ). These authors recommended a max-
imum ofthree amplification steps because ofincreases of back-
ground staining. A major advantage ofthe technique described
here is the possibility of choosing a desired staining intensity
by varying the number of incubation cycles, employing up to
ten repetitions (Figures 3, 4).
It is remarkable that there appears to be a linear, rather
than exponential, relationship between the number of incu-
bation cycles and the amount of peroxidase bound (Figures
3, 4). Factors that might limit the measured peroxidase activity
are: dissociation of anti-mouse Ig from mouse Ig, dissociation
of peroxidase from antiperoxidase, and impaired accessibility
of substrate within large complexes. No attempts were made
to discriminate between these possibilities.
To decide whether the detection limit of the technique is
improved by repetitive incubations, one may apply the defi-
nitions of efficiency (e.g., the signal to noise ratio achieved in
a stained preparation) and sensitivity (e.g. , the lowest amount
of antigen that can be distinguished from background) of im-
munocytochernical procedures as proposed by Petrusz et al.
(1980). Figure 4 shows that the background staining increases
with the number of incubation cycles. Consequently, staining
efficiency is not so strongly affected by stepwise amplification,
although some (up to 4-fold) increase was found in the ex-
peniments shown in Figures 3 and 4. Sensitiz’ity of staining
increased up to 30-fold in these experiments (data not shown),
if sensitivity was defined as the smallest amount of antibody
that yielded a significantly higher signal than background (e.g.,
no overlap of the means plus or minus two standard deviations).
When we compared the avidin-biotin-peroxidase complex
method (Hsu et al., 1981) with our stepwise-amplified im-
munoperoxidase staining, we found that repeating incubation
with anti-mouse Ig and PAP only once (Figure 3, 2 X ) resulted
in a higher signal to noise ratio and a higher absolute amount
ofperoxidase bound to monoclonal antibodies than was achieved
with the avidin-biotin-peroxidase complex method (Van den
Kwast et al., manuscript in preparation). The sensitivity of the
method is further illustrated by Figure 5. An example is the
weak staining of monocytes by Leu 3A (Figure SE, arrow),
which was not mentioned by Evans et al. ( 1 98 1 ), who described
the antibody and used an indirect immunofluorescence tech-
nique to detect binding of this antibody to cells, including
monocytes. That Leu-3A antibodies do bind to monocytes
was also found by Warner et al. (First International Workshop
on Human Leucocyte Differentiation Antigens, Paris, 1982),
who used a sensitive immunofluorescence technique.
A disadvantage of stepwise-amplified immunoperoxidase
staining is the amount of labor involved due to the multiple
incubation and wash steps. This limitation is substantially re-
duced, however, by immersion of whole slides into staining
jars. A similar approach to immunoperoxidase staining was
reported by Sofroniew and Schnell ( 1982). The use of an
automated device to perform the multistep protocol of our
technique would reduce the amount of human labor. Such a
device would optimally exploit the possibilities offered by the
relative simplicity, availability, and reproducibility of the re-
agents used in the assay.
An advantage of stepwise-amplified immunoperoxidase
staining is the preservation of morphological detail of labeled
and unlabeled cells, thus allowing rapid analysis of monoclonal
antibody specificity, as shown in Figure 5. Important in this
respect is the sequence in which incubation with monoclonal
antibodies and fixation of cells is performed as well as the
choice of fixative. Most fixatives that have little effect on an-
tigenicity (such as acetone or low concentrations of (para-)
formaldehyde or glutaraldehyde) result in poor morphology,
whereas fixation procedures giving good preservation of cel-
lular morphology usually have a harmful effect on antigenicity.
The problem can be circumvented, as described by Mason et
al. (1980) and illustrated in Figure 5, by incubation of viable
cells with monoclonal antibodies in suspension before cyto-
centrifugation (resulting in optimal display of cellular mor-
by guest on November 23, 2014jhc.sagepub.comDownloaded from

178 LANSDORP, VAN DER KWAST, DE BOER, ZEIJLEMAKER
phology) and fixation. An additional advantage of this pro-
cedure is that membrane and cytoplasmic staining are not likely
to be confused. Cytocentrifuge preparations of labeled cells
are fixed with buffered formol acetone (Mason et al., 1975),
which results in considerable destruction of antigenic deter-
minants, but preserves cellular morphology well. Apparently,
some antigenic determinants on mouse immunoglobulin sur-
vive the harsh fixation procedure, and these determinants can
be linked to similar determinants on (monoclonal) PAP corn-
plex by the anti-mouse Ig antibody (Figure 1 ). Because not
all monoclonal antibodies are fixed at their binding sites by
the formol-acetone fixation procedure (P.M. Knight, personal
communication), alternative fixation procedures may be nec-
essary for some monoclonal antibodies.
Application of stepwise amplified immunoperoxidase stain-
ing to tissue sections has not been described in this article.
However, the principles and findings reported here are prob-
ably also valid for staining of tissue sections. Apart from ap-
plications in irnrnunocytochernistry, the principle of stepwise
amplification may be applied in a variety of ways to increase
the sensitivity of monoclonal antibody detection.
Acknowledgment
The authors thank T. Wegmanforperforming the agar electrophoresis. and
R.C. Aalberse and D. Roos for critical!) reading the manuscript.
Literature Cited
Anstee DJ, Edwards PAW (1982): Monoclonal antibodies to human
enythnocytes. EurJ Immunol 12:228
Barnstable CJ, Bodmer WF, Bron G, Galfr#{233}G, Milstein C, Williams
AF, Ziegler A (1978): Production of monoclonal antibodies to groupA erythrocytes, HLA and other human cell surface antigens. Newtools for genetic analysis. Cell 14:9
BaumanJGJ, WiegantJ, van Duijn P (1981): Cytochemical hybnidi-sation with fluorochrome-labelled RNA. III. Increased sensitivity by
the use of anti-fluonescein antibodies. Histochemistry 73:18 1
Brown G, Biberfeld P, Christensson B, Mason DY (1979): The dis-tnibution of HLA on human lymphoid, bone marrow and peripheral
blood cells. Eur J Immunol 9:2 72
Evans R.L, Wall DW, Platsoucas CD, Siegal FP, Fiknig SM, Testa CM,Good RA (1981): Thymus-dependent membrane antigens in man:inhibition of cell-mediated lympholysis by monoclonal antibodies tothe TH2 antigen. Proc Natl Acad Sci USA 78:544
Ey PL, Pnowse SJ,Jenkins CR (1978): Isolation ofpure IgGl, IgG2a
and IgG2b immunoglobulins from mouse serum using protein-A Se-pharose. Immunochemistry 15:429
Figdor CG, Bont WS, Touw I, de Roos J, Roosnek EE, de Vries JE( 1982 ): Isolation of functionally different human monocytes by coun-
terfiow centrifugation elutniation. Blood 60:46
Halverson CA, Falini B, Taylor CR, PankenJW (1981): Detection of
terminal transferase in paraffin sections with the immunopenoxidase
technique. AmJ Pathol 105:241
Hsu SM, Raine L, Fauger H ( 1981): The use of avidin-biotin-pen-
oxidase complex (ABC) in mmmunoperoxidase techniques: a compan-
ison between ABC and unlabeled antibody (PAP) procedures. J His-tochem Cytochem 29:577
Lansdorp PM, Astaldi GCB, Oosterhof F, Janssen MC, Zeijlemaken
WP (1980): Immunopenoxidase procedures to detect monoclonal an-tibodies against cell-surface antigens. Quantitation ofbinding and staining
of individual cells. J Immunol Meth 39:393
Lansdorp PM, Oosterhof F, Astaldi GCB, Zeijlemaken WP (1982):
Detection of HLA antigens on blood platelets and lymphocytes by
means of monoclonal antibodies in an ELISA technique. Tissue An-
tigens 19:11
Laurent G, Gourdin MF, Reyes F ( 1980): Immunoperoxidase detec-
tion of immunoglobulins in cells of immunoprolifenative diseases. A
comparison between conjugate and nonconjugate (PAP) procedures.
AmJ Clin Pathol 74:265
Majdic 0, Liszka K, Lutz D, Knapp W (1981): Myeloid differentiationantigen defined by a monoclonal antibody. Blood 58:1127
Mason DY, Farrell C, Taylor CR (1975): The detection of intracellular
antigens in human leucocytes by immunoperoxidase staining. Br JHaematol 31:361
Mason DY, Leonard RCF, Laurent G, Gourdin MF (1980): Immu-
noperoxidase staining of surface and intracellular immunoglobulin in
human neoplastic lymphoid cells. J Clin Pathol 33:609
Mason DY, Condelljl, Abdulaziz Z, Naiem M, Bordenave G (1982):
Preparation of penoxidase:antipenoxidase (PAP) complexes for im-
munohistological labeling of monoclonal antibodies. J Histochem Cy-tochem 30:1114
Ordronneau P, Lindstr#{246}m PBM, Petrusz P (1981): Four unlabeled
antibody bridge techniques: a comparison. J Histochem Cytochem29: 1397
Petrusz P, Ordronneau P. Finley JCW ( 1980); Criteria of reliability
for light microscopic immunocytochemical staining. HistochemJ 12:333
Reinherz EL, Kung PC, PesandroJM, RitzJ, Goldstein G, SchlossmanSF ( 1979): Ia determinants on human T-cell subsets defined by mono-
clonal antibody.J Exp Med 150:1472
Schmitz H, Kampa D (1979): Amplified direct immunofluorescence
(AMDI) for detection of Epstein-Barr-virus nuclear antigen. J Im-munol Meth 26:17 3
Sofroniew MV, Schnell U (1982): Long-term storage and regular re-
peated use of diluted antisera in glass staining jars for increased sen-
sitivity, reproducibility, and convenience of single- and two-colon light
microscopic immunocytochemistry. J Histochem Cytochem 30:504
Sternberger LA (1979): Immunocytochemistry, 2nd ed. John Wiley
& Sons, New York
Taylor CR, Chin B ( 1978): Immunoperoxidase techniques. Practical
and theoretical aspects. Arch Pathol Lab Med 102: 1 13
Tetteroo PAT, Lansdorp PM, Leeksma OC, von dem Borne AEG Kr
( 1983): Monoclonal antibodies against human platelet glycoprotein
Lila. BrJ Haematol, in press
Ugolini V, Nunez G, Smith RG, Stasny P, CapnaJD (1980): Initialcharacterization of monoclonal antibodies against human monocytes.
Proc NatI Acad Sci USA 77:6764
Ulmer AJ, Flad HD ( 1979): Discontinuous density gradient separation
of human mononuclear leucocytes using Percoll as gradient medium.
J Immunol Meth 30:1
Vacca LL, Abrahams SJ, Naftchi NE (1980): A modified peroxidase-
antipenoxidase procedure for improved localization of tissue antigens:
localization of substance P in rat spinal cord. J Histochem Cytochem28:297
Wieme R (1976): Agan Gel Electrophonesis. Elsevier PubI Co,
Amsterdam.
by guest on November 23, 2014jhc.sagepub.comDownloaded from





























