Embed Size (px)
Citation preview

Vaccine 22 (2004) 4390–4396
Strong local and systemic protective immunity induced in the ferret modelby an intranasal virosome-formulated influenza subunit vaccine
Rob Lambkina, John S. Oxforda, Seb Bossuyta, Alex Manna, Ian C. Metcalfeb,Christian Herzogb, Jean-Franc¸ois Viretb, Reinhard Gluckb,∗
a Department of Medical Microbiology and Retroscreen Virology, St. Bartholomew’s and the Royal London, Queen Mary School of Medicineand Dentistry, University of London, 327 Mile End Road, London E1 4NS, UK
b Berna Biotech Ltd., Rehhagstr. 79, CH-3018 Berne, Switzerland
Received 4 July 2003; accepted 14 October 2003
Abstract
erally, haveb f virosomesa the efficacyo ng in nasals n ferret andh e adjuvantedw ologousv accinations,p©
K
1
ttctetflcii
U
ablene-ns,
tionim-se byys re-sssing
ag-yines.to re-
enzare-
cts
0d
The proliferation of influenza viruses causes costly, recurrent, annual epidemics. Current vaccines, mainly administered parenteen shown to be suboptimal in terms of efficacy, particularly where local IgA responses are concerned. Recent investigations os delivery systems for viral HA and NA antigens have demonstrated an improved immune response. This paper investigatesf a novel virosome-based intranasal influenza vaccine by its ability to reduce disease symptoms and its effect on viral sheddiecretions of immunised ferrets. The use of ferrets in the study of influenza vaccines is based on the good comparability betweeuman response to the disease. Intranasal, as opposed to parenteral, administration of a trivalent virosome-based subunit vaccinith HLT provides an almost total prevention of virus shedding combined with a high level of immunological protection against homirus challenge. The ease of application of an intranasal vaccine may have positive repercussions in the adoption of influenza varticularly in ‘at-risk’ groups.2004 Elsevier Ltd. All rights reserved.
eywords:Local and systemic immunity; Intranasal; Virosome; Influenza
. Introduction
Influenza, a contagious disease of the upper respiratoryract, is a costly socio-economic burden[1], whilst medicallyhe virus has a high morbidity in young children, immuno-ompromised individuals and the elderly[2–4]. Immunisa-ion, with current inactivated vaccines given parentally, isffective at reducing mortality, attenuating the symptoms of
he disease and minimising the sequelae associated with in-uenza. It has also been proposed that immunisation of younghildren, identified as enhancing ongoing viral transmissionn communities[5], might significantly reduce the spread ofnfluenza virus in the community. Therefore, the develop-
∗ Corresponding author. Tel.: +41 31 980 6500; fax: +41 31 980 6785.E-mail address:[email protected] (R. Gluck).
RL: http://www.retroscreen.com/.
ment of an efficacious and highly tolerable vaccine, suitfor all members of a community would be of great befit. The majority of current vaccines have several limitatioincluding non-biodegradability, inflammation and induraat the site of injection and either a weak, or no, cellularmune response. Attempts to increase antibody responincreasing the antigen content per dose have not alwasulted in improved immunogenicity[6]. These deficienciehave led to the search for alternative adjuvants possegreater efficacy and tolerability.
The majority of healthy individuals produce serum hemglutinin (HA)-specific immunoglobulin G (IgG) antibodwhen treated with the current parenteral influenza vaccSubcutaneous vaccines have also been documentedduce hospitalisation and deaths significantly during influepidemics through inducing a strong systemic immunesponse[7]. However, only a minimal number of subje
264-410X/$ – see front matter © 2004 Elsevier Ltd. All rights reserved.oi:10.1016/j.vaccine.2003.10.054

R. Lambkin et al. / Vaccine 22 (2004) 4390–4396 4391
present a significant rise in HA-specific nasal IgA antibodies[8].
This reduced ability of parenteral vaccines to elicit an im-munological response at the mucosa, the site of virus entryand propagation, may be an important determinant of the sub-optimal protective capacity of these vaccines. Stimulation ofboth local and systemic antibody responses may improve theimmunogenicity and clinical efficacy of vaccines. The firstline of defence for the host is mucosal immunity, which con-stitutes a major component of the immunological humoraland cell-mediated response in the upper and lower respira-tory tract.
An efficient intranasal vaccine applied to the site of virusentry would be expected to stimulate humoral and cellular im-mune responses at both mucosal and systemic levels[9,10].A further advantage of an intranasal influenza vaccine is theneedle-free administration that may improve vaccine cover-age due to greater patient acceptance particularly in ‘at-risk’groups.
The use of virosomes, or immunopotentiating reconsti-tuted influenza virosomes (IRIVs), as vaccine delivery sys-tems have been shown to greatly potentate immune responsesin animals and in humans[11–13]. Two parenteral virosome-based vaccines, against hepatitis A virus (Epaxal®) and in-fluenza (Inflexal V®), have been licensed.
ainv neu-r ayer[ gicp t in-flv dvan-t tainsn y andh s ands ex-i nes[ inesh
hu-m tiona l-u rmso le-s to bea ther n,t , thatt em-b tedt imalm nic-i acy[
par-e heir
efficacy at preventing influenza induced symptoms and virusshedding in nasal secretions of immunised ferrets.
2. Materials and methods
2.1. Test animals
Female ferrets (either albino or polecat), approximately1-year-old and of a body weight of 700–800 g, were obtainedfrom Foxfield Farms UK Ltd., Foxfield, UK. Animals wereidentified by an electronic chip, which was inserted underthe skin. This yielded a unique nine-digit identification num-ber for each ferret, and allowed tracking of any single an-imal throughout the experiment. Animals were maintainedunder controlled ferret diet (Diet F; Special Diet Services,Witham, UK). All animal work was conducted according toH.M. Home Office guidelines.
2.2. Immunisation
The vaccines consisted of virosomal preparations contain-ing 15�g of influenza virus hemagglutinin from each of thethree viral strains, namely A(H1N1) (A/Beijing/262/95like),A(H3N2) (A/Sydney/5/97), and B (B/Harbin/7/94), recom-m 999.T l ad-jt
ani-m lt pe-c roup2t notr eent umana
2
vP
2
ero-l eptord torso ayst cifica
dailyf fer-r tilled
Virosomes are manufactured by inserting the mirus antigenic determinants, the haemagglutinin andaminidase surface glycoproteins, into a liposome bil14]. These formulations exhibit enhanced immunoloroperties when used as active ingredients in a trivalenuenza vaccine[15] or as antigen delivery systems[16]. Airosome-based adjuvanted delivery system provides aages over alternatives in that it is bio-degradable, cono detergents or preservatives, has a greater tolerabilitas been shown to produce higher seroconversion rateignificantly greater protection rates when compared tosting commercial formula whole virion and subunit vacci17]. The properties and achievements of virosomal vaccave been recently reviewed[18].
Influenza infection in the ferret model closely mimicsan influenza, with regard both to the sensitivity to infecnd the clinical response[19]. The results of a study evaating cold-recombinant strains of influenza virus in tef clinical manifestations, viral titres and histopathologicions in turbinates and lungs of ferrets were also foundgood comparison of the results obtained from testing
ecombinants in humans[20]. In addition, it has been showhrough the observation of sequence changes in HA1he transmission of type A influenza in ferrets closely resles human transmission[21]. These studies have valida
he use of the ferret model as the most appropriate anodel to quantify infectiousness, virulence or pathoge
ty of influenza viruses and to investigate vaccine effic22].
In the present study, we have compared nasal andnteral formulations of a virosomal influenza vaccine for t
ended by the WHO for the influenza season 1998–1he intranasal vaccine contained in addition a mucosa
uvant (Escherichia coliheat-labile toxin; 4�g for the fullwo-doses immunisation scheme).
The ferrets were divided into three groups of sevenals: Group 1 received a daily dose of 200�l of intranasa
est vaccine (100�l per nostril) on days 0 and 7, using a sial spray device developed for human immunisation; Greceived the intramuscular test vaccine (15�g, injected into
he hind quarters) on day 0; Group 3 was a control groupeceiving vaccine. The differences in administration betwhe nasal and parenteral groups corresponded to the hdministration procedures.
.3. Challenge
On day 21 ferrets were challenged with 106.5 TCID50 ofirus strain A/Sydney/5/97 (H3N2) administered in 500�lBS (250�l per nostril).
.4. Sample collection and analysis
Blood samples were taken at day 0 and day 28 for sogical analysis. Serum samples were treated with recestroying enzyme (RDE) to remove non-specific inhibif haemagglutination. Haemagglutinin inhibition (HI) ass
o detect pre and post immunisation levels of virus-spentibody were conducted on antisera.
Temperatures and nasal washes were taken oncerom day 21 to day 28. To obtain the nasal mucus, theets were first anaesthetised; 1.0 ml PBS was then ins
4392 R. Lambkin et al. / Vaccine 22 (2004) 4390–4396
into each nostril followed by collection of the mucus run-ning from the nose. Samples from nasal lavages were mixedwith Trypan blue and mounted on a haemocytometer in or-der to determine inflammatory cell counts. Influenza viruswas titrated on MDCK cells from a second aliquot of nasallavage sample; each virus titration assay was conducted inquadruplicate from which the TCID50 was calculated usingthe Kaber calculation according to:
log TCID50 endpoint= −L −[(
S
100− 0.5
)× logd
]
whereL is the log of highest concentration used,Sthe sum ofpercentage infected wells at each dilution andd is the dilution.
On day 28, the ferrets were culled, the lungs were removedand one lung was fixed in formalin for later histological anal-ysis.
A one-way between groups analysis of variance was con-ducted to explore the impact of the type of vaccine adminis-tered on the various clinical parameters following challengewith influenza A virus. The degree of significant differencewas estimated at theP < 0.05 level. Post-hoc comparisonswere then conducted using the Tukey HSD test[23].
F2oa
3. Results
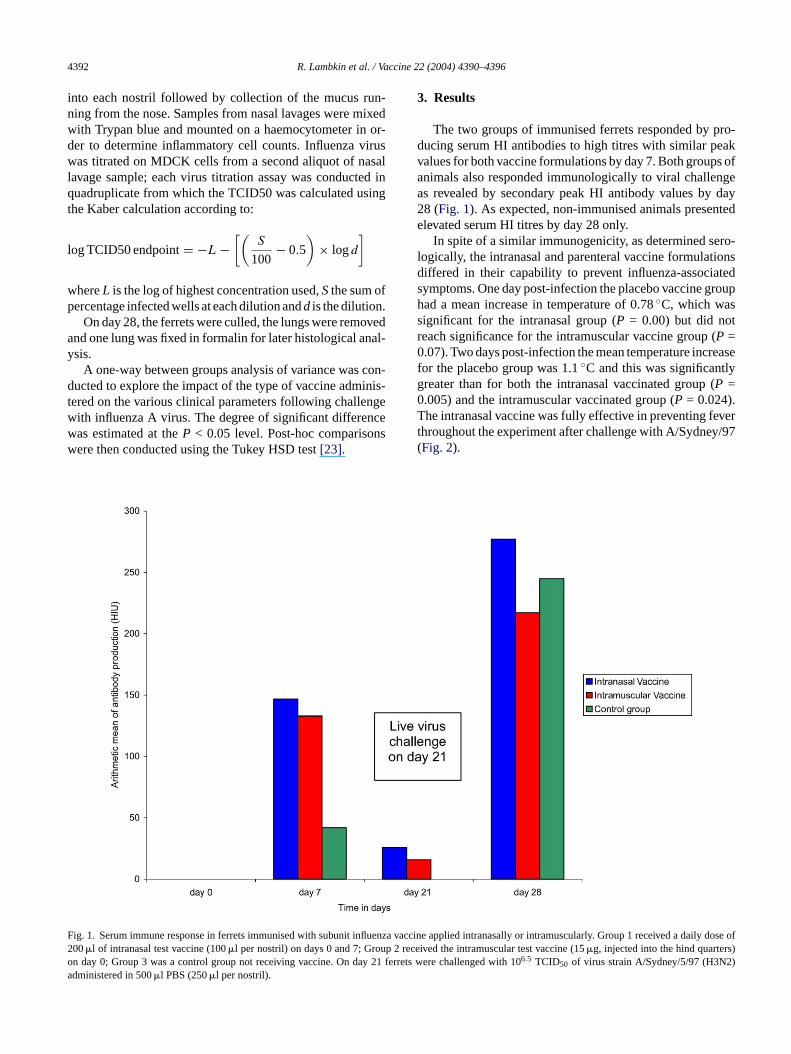
The two groups of immunised ferrets responded by pro-ducing serum HI antibodies to high titres with similar peakvalues for both vaccine formulations by day 7. Both groups ofanimals also responded immunologically to viral challengeas revealed by secondary peak HI antibody values by day28 (Fig. 1). As expected, non-immunised animals presentedelevated serum HI titres by day 28 only.
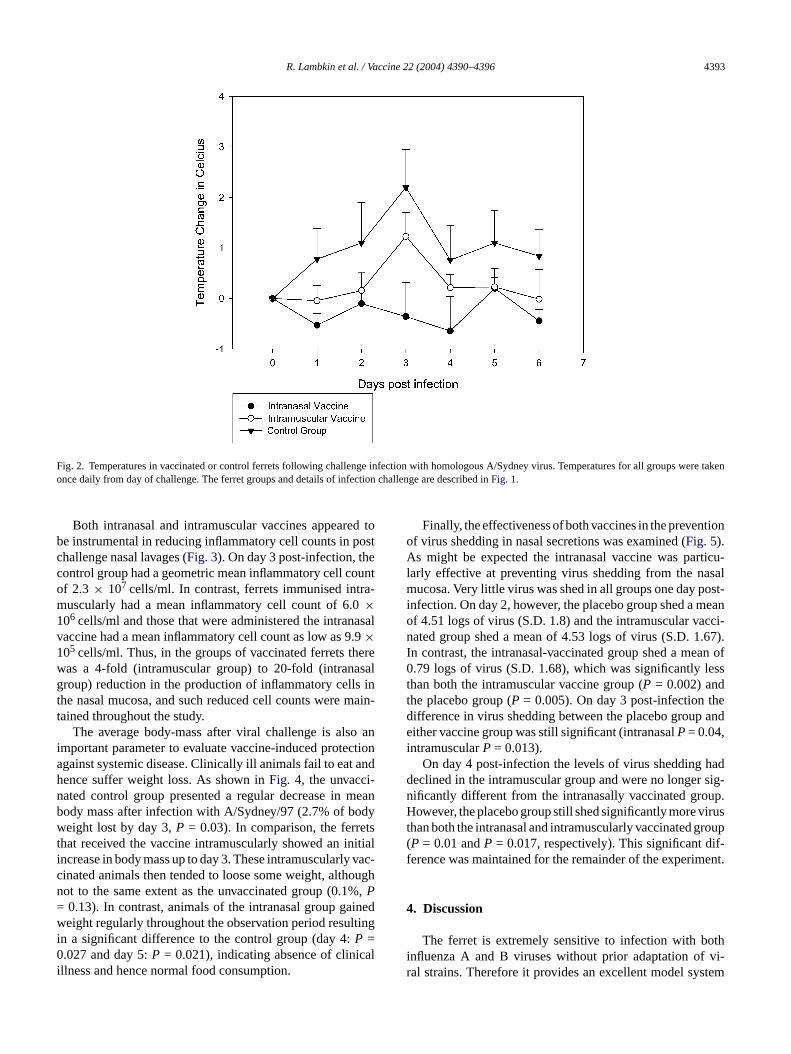
In spite of a similar immunogenicity, as determined sero-logically, the intranasal and parenteral vaccine formulationsdiffered in their capability to prevent influenza-associatedsymptoms. One day post-infection the placebo vaccine grouphad a mean increase in temperature of 0.78◦C, which wassignificant for the intranasal group (P = 0.00) but did notreach significance for the intramuscular vaccine group (P =0.07). Two days post-infection the mean temperature increasefor the placebo group was 1.1◦C and this was significantlygreater than for both the intranasal vaccinated group (P =0.005) and the intramuscular vaccinated group (P = 0.024).The intranasal vaccine was fully effective in preventing feverthroughout the experiment after challenge with A/Sydney/97(Fig. 2).
ig. 1. Serum immune response in ferrets immunised with subunit influenza00�l of intranasal test vaccine (100�l per nostril) on days 0 and 7; Group 2 rn day 0; Group 3 was a control group not receiving vaccine. On day 21 fedministered in 500�l PBS (250�l per nostril).
vaccine applied intranasally or intramuscularly. Group 1 received a daily dose ofeceived the intramuscular test vaccine (15�g, injected into the hind quarters)rrets were challenged with 106.5 TCID50 of virus strain A/Sydney/5/97 (H3N2)
R. Lambkin et al. / Vaccine 22 (2004) 4390–4396 4393
Fig. 2. Temperatures in vaccinated or control ferrets following challenge infection with homologous A/Sydney virus. Temperatures for all groups were takenonce daily from day of challenge. The ferret groups and details of infection challenge are described inFig. 1.
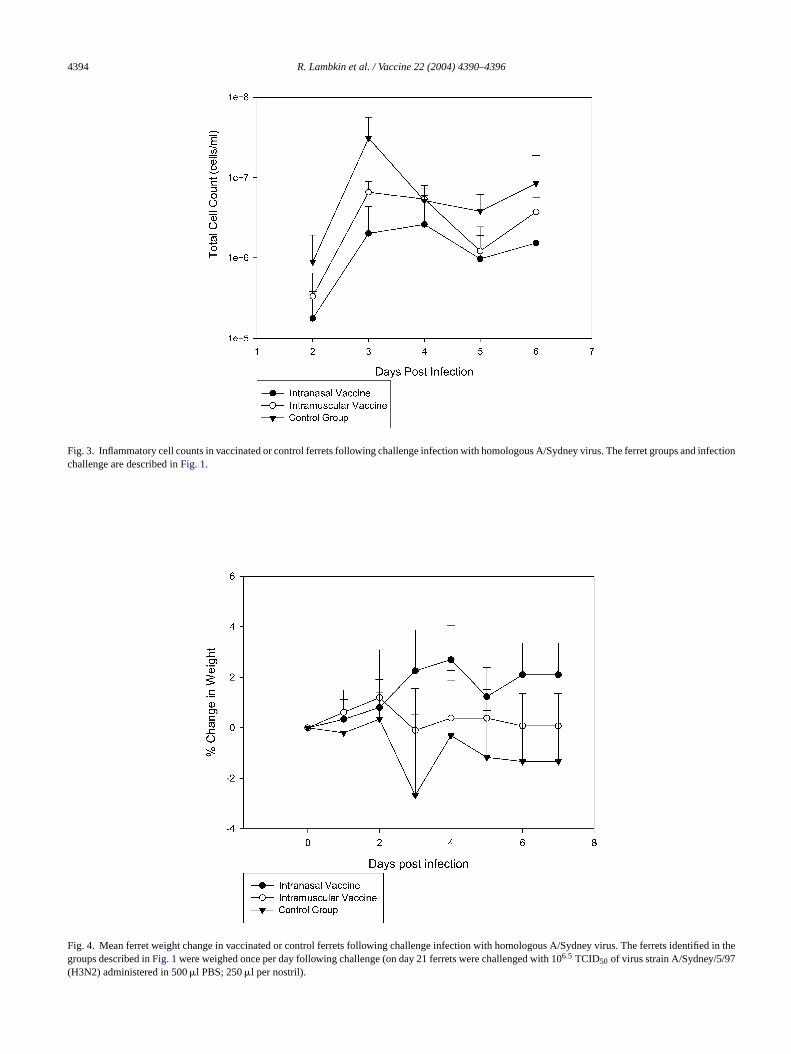
Both intranasal and intramuscular vaccines appeared tobe instrumental in reducing inflammatory cell counts in postchallenge nasal lavages (Fig. 3). On day 3 post-infection, thecontrol group had a geometric mean inflammatory cell countof 2.3 × 107 cells/ml. In contrast, ferrets immunised intra-muscularly had a mean inflammatory cell count of 6.0×106 cells/ml and those that were administered the intranasalvaccine had a mean inflammatory cell count as low as 9.9×105 cells/ml. Thus, in the groups of vaccinated ferrets therewas a 4-fold (intramuscular group) to 20-fold (intranasalgroup) reduction in the production of inflammatory cells inthe nasal mucosa, and such reduced cell counts were main-tained throughout the study.
The average body-mass after viral challenge is also animportant parameter to evaluate vaccine-induced protectionagainst systemic disease. Clinically ill animals fail to eat andhence suffer weight loss. As shown inFig. 4, the unvacci-nated control group presented a regular decrease in meanbody mass after infection with A/Sydney/97 (2.7% of bodyweight lost by day 3,P = 0.03). In comparison, the ferretsthat received the vaccine intramuscularly showed an initialincrease in body mass up to day 3. These intramuscularly vac-cinated animals then tended to loose some weight, althoughnot to the same extent as the unvaccinated group (0.1%,P= 0.13). In contrast, animals of the intranasal group gainedw ltingi0 ali
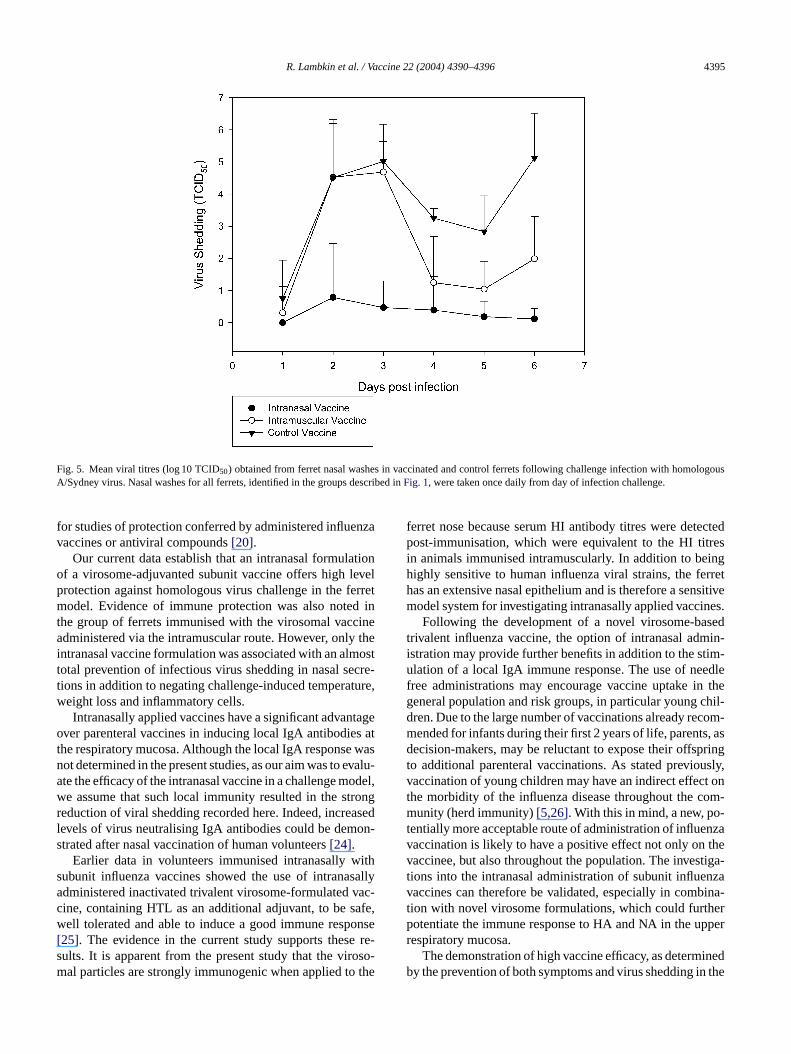
Finally, the effectiveness of both vaccines in the preventionof virus shedding in nasal secretions was examined (Fig. 5).As might be expected the intranasal vaccine was particu-larly effective at preventing virus shedding from the nasalmucosa. Very little virus was shed in all groups one day post-infection. On day 2, however, the placebo group shed a meanof 4.51 logs of virus (S.D. 1.8) and the intramuscular vacci-nated group shed a mean of 4.53 logs of virus (S.D. 1.67).In contrast, the intranasal-vaccinated group shed a mean of0.79 logs of virus (S.D. 1.68), which was significantly lessthan both the intramuscular vaccine group (P = 0.002) andthe placebo group (P = 0.005). On day 3 post-infection thedifference in virus shedding between the placebo group andeither vaccine group was still significant (intranasalP= 0.04,intramuscularP = 0.013).
On day 4 post-infection the levels of virus shedding haddeclined in the intramuscular group and were no longer sig-nificantly different from the intranasally vaccinated group.However, the placebo group still shed significantly more virusthan both the intranasal and intramuscularly vaccinated group(P = 0.01 andP = 0.017, respectively). This significant dif-ference was maintained for the remainder of the experiment.
4. Discussion
othi vi-r stem
eight regularly throughout the observation period resun a significant difference to the control group (day 4:P =.027 and day 5:P = 0.021), indicating absence of clinic
llness and hence normal food consumption.
The ferret is extremely sensitive to infection with bnfluenza A and B viruses without prior adaptation ofal strains. Therefore it provides an excellent model sy
4394 R. Lambkin et al. / Vaccine 22 (2004) 4390–4396
Fig. 3. Inflammatory cell counts in vaccinated or control ferrets following challenge infection with homologous A/Sydney virus. The ferret groups and infectionchallenge are described inFig. 1.
Fig. 4. Mean ferret weight change in vaccinated or control ferrets following challenge infection with homologous A/Sydney virus. The ferrets identified in thegroups described inFig. 1were weighed once per day following challenge (on day 21 ferrets were challenged with 106.5 TCID50 of virus strain A/Sydney/5/97(H3N2) administered in 500�l PBS; 250�l per nostril).
R. Lambkin et al. / Vaccine 22 (2004) 4390–4396 4395
Fig. 5. Mean viral titres (log 10 TCID50) obtained from ferret nasal washes in vaccinated and control ferrets following challenge infection with homologousA/Sydney virus. Nasal washes for all ferrets, identified in the groups described inFig. 1, were taken once daily from day of infection challenge.
for studies of protection conferred by administered influenzavaccines or antiviral compounds[20].
Our current data establish that an intranasal formulationof a virosome-adjuvanted subunit vaccine offers high levelprotection against homologous virus challenge in the ferretmodel. Evidence of immune protection was also noted inthe group of ferrets immunised with the virosomal vaccineadministered via the intramuscular route. However, only theintranasal vaccine formulation was associated with an almosttotal prevention of infectious virus shedding in nasal secre-tions in addition to negating challenge-induced temperature,weight loss and inflammatory cells.
Intranasally applied vaccines have a significant advantageover parenteral vaccines in inducing local IgA antibodies atthe respiratory mucosa. Although the local IgA response wasnot determined in the present studies, as our aim was to evalu-ate the efficacy of the intranasal vaccine in a challenge model,we assume that such local immunity resulted in the strongreduction of viral shedding recorded here. Indeed, increasedlevels of virus neutralising IgA antibodies could be demon-strated after nasal vaccination of human volunteers[24].
Earlier data in volunteers immunised intranasally withsubunit influenza vaccines showed the use of intranasallyadministered inactivated trivalent virosome-formulated vac-cine, containing HTL as an additional adjuvant, to be safe,w onse[ e re-s oso-m the
ferret nose because serum HI antibody titres were detectedpost-immunisation, which were equivalent to the HI titresin animals immunised intramuscularly. In addition to beinghighly sensitive to human influenza viral strains, the ferrethas an extensive nasal epithelium and is therefore a sensitivemodel system for investigating intranasally applied vaccines.
Following the development of a novel virosome-basedtrivalent influenza vaccine, the option of intranasal admin-istration may provide further benefits in addition to the stim-ulation of a local IgA immune response. The use of needlefree administrations may encourage vaccine uptake in thegeneral population and risk groups, in particular young chil-dren. Due to the large number of vaccinations already recom-mended for infants during their first 2 years of life, parents, asdecision-makers, may be reluctant to expose their offspringto additional parenteral vaccinations. As stated previously,vaccination of young children may have an indirect effect onthe morbidity of the influenza disease throughout the com-munity (herd immunity)[5,26]. With this in mind, a new, po-tentially more acceptable route of administration of influenzavaccination is likely to have a positive effect not only on thevaccinee, but also throughout the population. The investiga-tions into the intranasal administration of subunit influenzavaccines can therefore be validated, especially in combina-tion with novel virosome formulations, which could furtherp pperr
inedb n the
ell tolerated and able to induce a good immune resp25]. The evidence in the current study supports thesults. It is apparent from the present study that the viral particles are strongly immunogenic when applied to
otentiate the immune response to HA and NA in the uespiratory mucosa.
The demonstration of high vaccine efficacy, as determy the prevention of both symptoms and virus shedding i

4396 R. Lambkin et al. / Vaccine 22 (2004) 4390–4396
ferret challenge model, and consistent high immunogenicityin humans will justify the further evaluation of virosomalformulations in the development of nasal influenza vaccines.
References
[1] Nicholson KG. Socioeconomics of influenza and influenza vaccina-tion in Europe. Pharmacoeconomics 1996;9:75–8.
[2] Kingston BJ, Wright Jr CV. Influenza in the nursing home. Am FamPhysician 2002;65:75–8 and 72.
[3] Marshall BC, Henshaw C, Evans DA, Bleyl K, Alder S, Liou TG.Influenza vaccination coverage level at a cystic fibrosis center. Pe-diatrics 2002;109:E80.
[4] Redding G, Walker RE, Hessel C, et al. Safety and tolerabilityof cold-adapted influenza vaccine in children and adolescents withasthma. Pediatr Infect Dis J 2002;21:44–8.
[5] Reichert TA, Sugaya N, Fedson DS, et al. The Japanese experiencewith vaccinating school children against influenza. N Engl J Med2001;344:889–96.
[6] Couch RB, Keitel WA, Cate TR. Improvement of inactivated in-fluenza vaccines. J Infect Dis 1997;176:38–44.
[7] Fleming DM, Watson JM, Nicholas S, Smith GE, Swan AV. Studyof the effectiveness of influenza vaccination in the elderly in theepidemic of 1989–1990. Epidemiol Infect 1995;115:581–9.
[8] Clements ML, Betts RF, Tierney EL, et al. Serum and nasal washantibodies associated with resistance to experimental challenge withinfluenza A wild type virus. J Clin Microbiol 1986;24:157–60.
[9] Holmgren J. Mucosal immunity and vaccination. FEMS Microbiol
[ ent:3.
[ nts.
[ lentiatric
[13] Garcon NM, Six HR. Universal vaccine carrier. J Immunol1991;146:3697–702.
[14] Mischler R, Metcalfe IC. Inflexal V a trivalent virosome sub-unit influenza vaccine: production. Vaccine 2002;20(Suppl. 5):B17–23.
[15] Gluck R, Mischler R, Finkel B, Que JU, Scarpa B, Cryz Jr SJ. Im-munogenicity of new virosome influenza vaccine in elderly people.Lancet 1994;334:160–3.
[16] Cryz Jr SJ, Gluck R. Immunopotentiating reconstituted influenzavirosomes as a novel antigen delivery system. Dev Biol Stand1998;92:219–23.
[17] Gluck R. Preclinical and clinical evaluation of a new intranasal in-fluenza vaccine. Int Congr Ser 2001;1219:969–78.
[18] Moser C, Metcalfe IC, Viret JF. Virosomal adjuvanted antigen de-livery systems. Expert Rev Vac 2003;2(2):189–96.
[19] Sweet C, Smith H. Pathogenicity of influenza virus. Microbiol Rev1980;44:303–30.
[20] Maassab HF, Kendal AP, Abrams GD, Monto AS. Evaluation ofa cold-recombinant influenza virus vaccine in ferrets. J Infect Dis1982;146:780–90.
[21] Herlocher ML, Elias S, Truscon R, Harrison S, Mindell D, Simon C,et al. Ferrets as a transmission model for influenza: sequence changesin HA1 of type A (H3N2) virus. J Infect Dis 2001;184:542–6.
[22] Fenton RJ, Clark A, Potter CW. Immunity to influenza in ferrets.XIV. Comparative immunity following infection or immunisationwith live or inactivated vaccine. Br J Exp Pathol 1981;62:297–307.
[23] Hochberg Y, Tamhane AC. Multiple Comparison Procedures. JohnWiley & Sons; 1987.
[24] Durrer P, Gluck U, Spyr C, Lang AB, Zurbriggen R, Herzog C,et al. Mucosal antibody response induced with a nasal virosome-
[ onj,ated
[ ism,cina-
Immunol 1991;89:1–10.10] McGhee JR, Kiyono H. New perspectives in vaccine developm
mucosal immunity to infections. Infect Agents Dis 1993;2:55–711] Allison AC, Gregoriadis G. Liposomes as immunological adjuva
Nature 1974;252:252–5.12] Conne P, Gauthey L, Vernet P, et al. Immunogenicity of triva
subunit versus virosome-formulated influenza vaccines in gerpatients. Vaccine 1997;15:1675–9.
based influenza vaccine. Vaccine 2003;21(27–30):4328–34.25] Crovari P, Ansaldi F, Bruzzone BM, Garbarino E, Di Pietroant
Herzog C, et al. Safety and immunogenicity of mucosal, inactivvirosome-formulated influenza vaccine withEscherichia coliheatlabile toxin as an adjuvant. J Pre Med Hyg 1999;40:83–8.
26] Nettleman MD, White T, Lavoie S, Chavin C. School absenteeparental work loss, and acceptance of childhood influenza vaction. Vaccine 2001;321:178–80.





























