Embed Size (px)
Citation preview

REGULAR ARTICLE
Living between land and water – structural and functionaladaptations in vegetative organs of bladderworts
Markus S. Reut & Piotr Świątek &
Vitor F. O. Miranda & Bartosz J. Płachno
Received: 12 October 2020 /Accepted: 19 March 2021# The Author(s) 2021
AbstractAims The carnivorous Utricularia (Lentibulariaceae)has an anatomically simple and seemingly rootless veg-etative body. It occupies a variety of wetlands andinland waters and shows a broad range of life forms.Here, we aimed to elucidate structural and functionaltraits in various hydric conditions. Furthermore, weintended to evaluate morpho-anatomical adaptations incorrelation with life forms.Methods Morpho-anatomical characteristics typical forhydrophytes of all life forms were investigated by lightmicroscopy on 13 Utricularia taxa, compared to onePinguicula and two Genlisea taxa, and assessed bymultivariate analyses.Results Vegetative structures of Utricularia andGenlisea showed reduced cortical, supporting, and
vascular tissues. With increasing water table, leaveswere thinner, and narrower or dissected, and submergedorgans tended to contain chloroplasts in parenchymaticand epidermal cells. In some main stolons, an endoder-mis with Casparian strips was visible. Large gas cham-bers, including a novel ‘crescent’ and a special ‘hollow’aerenchyma pattern, were found in amphibious to free-floating taxa.Conclusions The evolutionary transfer of carnivoryfrom aerial to subterranean organs inGenlisea, and evenmore in Utricularia, coincides with a highly simplifiedanatomy, which is adapted to a broad variety of hydricconditions and compensates for structural innovations inthe uptake of nutrients.
Keywords Utricularia . Lentibulariaceae .
Hydrophyte . Carnivorous plants . Life forms .
Aerenchyma . Casparian strips . Anatomy
Introduction
Life forms of hydrophytes
Hydrophytes are plants living “in water or on a substratethat is saturated at a frequency and duration during thegrowing period sufficient to affect plant occurrence”(Tiner 2017). Water plants constitute about 1–2% ofthe angiosperms today, while vascular hydrophytes(macrophytes) evolved from terrestrial plants of hun-dreds of families and genera at several points in time(Cook 1999; Chambers et al. 2008).
Plant Soilhttps://doi.org/10.1007/s11104-021-04929-6
Responsible Editor: Hans Lambers.
M. S. Reut (*) :B. J. Płachno (*)Department of Plant Cytology and Embryology, Institute ofBotany, Faculty of Biology, Jagiellonian University in Kraków,Gronostajowa 9 St, 30-387 Kraków, Polande-mail: [email protected]: [email protected]
P. ŚwiątekInstitute of Biology, Faculty of Natural Sciences, University ofSilesia in Katowice, 9 Bankowa St.ID 40-007 Katowice, Poland
V. F. O. MirandaSchool of Agricultural and Veterinarian Sciences, Department ofApplied Biology, Laboratory of Plant Systematics, UNESP - SãoPaulo State University, Campus Jaboticabal, Jaboticabal CEP14884-900 SP, Brazil

The life forms (habits) of aquatic plants range fromterrestrial but occasionally inundated taxa, with anchor-age in the substrate and with leaves and reproductiveorgans exposed to the air, to unattached, free-floatingtaxa with all parts under water. Between the extremes,gradually different forms exist, contingent upon waterregimes and adaptive responses of the plants to environ-mental changes such as flooding and unflooding, pre-cipitation, or evaporation (cf. Sculthorpe 1967;Braendle and Crawford 1999; Tiner 2017). Many hy-drophytes have adapted to various habitats and henceshow more than just one life or growth form (cf.Sculthorpe 1967; Cook 1999).
Classification of hydrophytes
Authors engaged in the research of carnivorous plantsand the Lentibulariaceae in particular (e.g. Taylor 1989;Guisande et al. 2007; Reut and Płachno 2020) designatemost species within the Lentibulariaceae as “terres-trials”, although the majority colonizes wet, water-logged or temporarily submerged habitats. Accordingto Taylor (1989), “terrestrial” plants are those floweringwhen the soil is humid or saturated but being submergedin wet periods, which coincides with Cook’s (1999)definition of ‘tenagophytes’. Taylor (1989) delimits“terrestrials” from affixed aquatics and (freely)suspended (inflated) aquatics but in some instances alsofrom subaquatics. Affixed aquatics are mostly anchoredin the substrate (Guisande et al. 2007) and possiblyrepresent a transition towards suspended (free-floating)hydrophytes (Jobson et al. 2003). Subordinating sub-aquatic in “terrestrial” habits (e.g. Guisande et al. 2007)or subaquatic in affixed aquatic habits (e.g. Jobson et al.2018) leads to results which are difficult to compare.This attempt of a classification of life forms in carnivo-rous plants does not seem to be suitable for a classifica-tion of hydrophytes in general and needs to be adjusted.
In the current paper, we follow a simple classificationof habits of wetland and aquatic plants as outlined bySculthorpe (1967) and Hutchinson (1975), differentiat-ing between ‘anchored hydrophytes’ (encompassing‘emergent hydrophytes’, ‘floating-leaved hydrophytes’,and ‘submerged hydrophytes’) and ‘free-floating hydro-phytes’. However, since ‘amphiphytes’ are adapted to aterrestrial as well as a (periodically) submerged life, i.e.to aerobic and anaerobic conditions, we rank this groupamong anchored hydrophytes. They are often found inthe transitional zone between land and water where
flooding and unflooding occur (Braendle andCrawford 1999; Crawford 2008). Extending the scopeof hydrophytes by adding amphiphytes enables theidentification of ecological and evolutionary transects(i.e. various grades and combinations of morphologicaland anatomical specializations) within hydrophyticplants, and the concurrent delimitation of this groupfrom purely terrestrial plants (Barrett et al. 1993).
Morpho-anatomical characters
Early anatomical works on hydrophytes provide in-sights into structural characteristics of species and gen-era across various plant families (e.g. Schenck 1886;Arber 1920). In his book on aquatic vascular plants,Sculthorpe (1967) connects morpho-anatomical traitswith physiological and functional aspects. Experimentaland observational studies, mainly at the species level,led to the conclusion that the life in a transitional zonebetween land and water, and in fluctuating water condi-tions entails a variety of adaptive responses, and hence agreater phenotypic plasticity, especially in amphibiousand emergent hydrophytes (Barrett et al. 1993;Crawford 2008; Lusa et al. 2011). In these groups,heterophylly is widespread, exhibiting submerged andaerial leaf forms adapted to the respective milieu(Sculthorpe 1967; Mommer et al. 2005; Li et al.2019). In the relatively stable and homogeneous envi-ronment of the water column, however, the diversifica-tion of obligate hydrophytes is generally low (Barrettet al. 1993; Hidalgo et al. 2015). Overall, hydrophytesshow heterogeneous combinations of morphological,anatomical, and functional traits (Table 1) that differfrom those of strictly terrestrial plants.
Comparing species of various life forms within onegenus may uncover adaptations to habitats and hencealso evolutionary aspects rather than just phenotypicresponses to changes in the environment. However, todate, a comparative morpho-anatomical study on a hy-drophytic plant genus with a broad range of life forms ismissing. One reason may be that only few aquaticgenera offer a large variety of life forms (cf. Cook1999). One of these is the carnivorous Utricularia L.
Lentibulariaceae
Members of the bladderwort family (Lentibulariaceae:Lamiales) with its three genera Pinguicula L.,Genlisea A.St.-Hil. and Utricularia L. are among the
Plant Soil

most interesting herbaceous angiosperms because oftheir carnivorous nature, unique vegetative bodies,small genome sizes, and several more biological pecu-liarities (cf. Ibarra-Laclette et al. 2013; Rutishauser2020; Silva et al. 2020a).
Rosulate and rhizomatous growth forms are general-ly found in terrestrial or amphibious species of theLentibulariaceae, while stolonifery occurs within allhabits. In the current paper, we use the terms ‘rhizome’and ‘stolon’ group-specifically. Both are generally hor-izontally growing shoots, while rhizomes tend to beshorter, often having storage function and giving riseto roots. Stolons are considered to have longer inter-nodes. ‘Anchor stolons’ (‘rhizoids’, Taylor 1989) ofseveral Utricularia species are assigned to unbranchedand much shorter organs that grow geotropically posi-tive and generally into the soil (Reut and Płachno 2020).
The rosulate butterworts (Pinguicula) contain around100 species of which most of them are mesophytes,amphiphytes and lithophytes, and only a few are true
epiphytes (Fleischmann and Roccia 2018; Casper2019). Insects are trapped on the foliage leaves ofhomophyllous butterworts above ground throughoutthe year, while heterophyllous species are effectivelycarnivorous only during wet periods (e.g. Legendre2000; Fleischmann and Roccia 2018). Pinguicula spe-cies have rhizomes (Casper 2019) or short stems(Legendre 2000), and few produce runner stolons.
The corkscrew plants (Genlisea) comprise 31 spe-cies of amphiphytes and few emergent hydrophytes(Fleischmann 2012, 2018; Silva et al. 2020b).Genlisea is considered being heterophyllous, havingnon-carnivorous rosettes of green leaves aboveground and subterranean “rhizophylls” that functionlike an eel-trap (Reut 1993; Fleischmann 2018;Płachno and Muravnik 2018). Genlisea shows shortvertical stems or horizontal rhizomes (Fleischmann2018). Since the rhizomes in G. repens are elongated(Reut 1993), they will hereafter be called stolons inaccordance with Fleischmann (2018).
Table 1 Common morphological and anatomical characters and their functions in herbaceous hydrophytes
Morphological / anatomical adaptation Function
1) Reduced or absent roots in some (submerged) species BIO
2) Tendency to develop narrow leaves (submerged form) or dissected leaves (inseveral submerged or free-floating aquatics) increasing the ratio of surfaceto volume
ABS, FLE, LIG
3) Thin leaves without palisade parenchyma, and with reduced or nomesophyll, losing dorsiventrality
FLE, LIG
4) Epidermis of submerged organs/leaves with chloroplasts LIG
5) Delicate submerged organs with a thin cuticula ABS
6) Reduced xylem elements BIO
7) Prominent intercellular spaces (between spongy parenchyma in leaves) orlacunar / aerenchymatous anatomy
GAS
8) Occurrence of hydropotes or glandular types of trichomes in the epidermis,e.g. in submerged species of Potamogeton, Ceratophyllum, andCryptocoryne
ABS
9) Reduced or absent supporting tissue (sclerenchyma, collenchyma) FLE
10) Limitation of the vascular strand to a central cylinder in roots (and stems);endodermoid layer, channelizing the flow of water through reduced xylem
BAR, BIO
11) Reduction, inactivity, or absence of stomata (especially in submergedleaves); in some plants replaced by secretory structures
BIO, STO
12) Development of adventitious roots STA, ABS
Abbreviations of functions: ABS = Enhancement of absorption of nutrients, and dissolved CO2 and O2; GAS = Passage for an increaseddiffusion of gases, responding to gaseous pressure; buoyancy; BAR = Barrier to restrict radial oxygen loss (ROL) to the soil; BIO =Optimization of biomass investment (alternative structures for the uptake or transport of nutrients and dissolved gases); FLE = Flexibility inwater movements; LIG = Enhancement of the collection of light in lower light conditions and in every organ orientation; STA = Stabilizationof the plant (rosette) position in the water; STO =Redundancy of stomata (gas exchange in the air) in submerged organs; hydathodes forguttation (water secretion by root pressure) on floating leaves
Summarized from overviews by Sculthorpe (1967), Wetzel (1988), Wiegleb (1991), Jung et al. (2008), Colmer et al. (2011), Krähmer(2016), and Tiner (2017)
Plant Soil

Finally, the bladderworts (Utricularia) are the mostdiverse within the Lentibulariaceae. They consist ofabout 240 species and show the widest range of lifeforms within the family, from mesophytes to free-floating aquatics, in addition to rheophytes, lithophytes,and epiphytes (cf. Guisande et al. 2007; Jobson et al.2018; Jobson and Baleeiro 2020). Like Genlisea, “ter-restrial” Utricularia species have leaves for photosyn-thesis and subterranean leaf-like organs for carnivory. InUtricularia, the prey is actively captured by tiny suctionbladders, functioning like a mousetrap, while the tubulartrap of Genlisea is considered being passive. However,abundant water is required for an effective operation ofboth trap types (Guisande et al. 2007; Jobson et al. 2018;Płachno et al. 2019a, b). Most species ofUtricularia arestoloniferous (Taylor 1989; Jobson et al. 2003). Theterm “stolon” has established inUtricularia and prevailsfor reasons of consistency (Taylor 1989), also becausein some species it grows above ground and up to severalmeters in length.
Anatomy of vegetative organs in the Lentibulariaceae
Early anatomical examinations of vegetative parts ofcarnivorous plants were done e.g. by Kamieński(1891), Lang (1901) and Merl (1915), but they largelyfailed to imply comparative discussions on functionalaspects in a broader context of e.g. carnivory or adapta-tions to aquatic habitats. Later anatomical studies onvegetative organs in the Lentibulariaceae were mainlyfocusing on trap tissues (e.g. Lloyd 1942; Płachno et al.2014; Płachno et al. 2017) and the functional ultrastruc-ture of digestive glands (e.g. Płachno and Jankun 2004;Płachno et al. 2019a). Most recently, Płachno et al.(2020b) described structures of tubers in one Genliseaand two Utricularia species showing adaptation to abi-otic stress (e.g. seasonal drought or fire). Reut andPłachno (2020) examined the anatomy of stolons, blad-der stalks and leaves ofU. dichotoma sensu lato clones,originating from five amphibious to emergent popula-tions of Australia and New Zealand, and found simpleand lacunate characters, which are basically homoge-nous throughout the subterranean/submersed organsand across the sources.
Aims of the study
In the current study, we useUtricularia (Lentibulariaceae)as model genus to investigate differences and
commonalities of morpho-anatomical characters betweenspecies of several hydrophytic life forms, as this genusyields probably the greatest richness of habits amongsthydrophytes. We aim to elucidate by which structuralmeans Utricularia handles water, nutrients, and gases invarious hydric conditions. By applying multivariate anal-yses, we assess the clustering of life forms and the gradeof adaptations to the level of submergence, in correlationto combinations of morpho-anatomical characters of veg-etative organs of several amphibious, submerged, andfree-floating taxa of Utricularia and chosen taxa ofPinguicula andGenlisea. Results are discussed in relationto functional aspects of traits, the carnivorous syndrome,and the evolution of the Lentibulariaceae.
Materials and methods
Plant material and preparation
In our morpho-anatomical investigation, we selected atotal of 16 species from the Lentibulariaceae generaPinguicula, Genlisea and Utricularia (see Table 2 fora full description, including taxonomic ranking, growthform and sources).
The plant material was studied with respect to severalmorpho-anatomical characters as outlined in Table 1.Examinations were performed on leaves, roots(Pinguicula only), trap stalks, and several types of sto-lons (where present). The anatomy of leaves was inves-tigated on P. gigantea, G. hispidula, U. volubilis,U. paulineae, U. uniflora, U. delicatula, and U. stygia.Detailed data of all investigated Lentibulariaceae spe-cies with traits of leaves, stolons, roots, anchor stolons,and trap stalks is provided in the Supplementary Infor-mation (Online Resource) 1.
Plant samples used for light microscopywere fixed ina mixture of 2.5 % glutaraldehyde and 2.5% formalde-hyde in a 0.05 M cacodylate buffer (Sigma; pH 7.2)overnight or for up to four days. After fixation, thematerial was washed three times in a 0.1 M sodiumcacodylate buffer followed by post-fixation in 1% os-mium tetroxide solution at room temperature for 1.5 h.The samples were dehydrated in graded ethanol series to95%, infiltrated and embedded using an epoxy embed-ding medium kit (Fluka). After polymerisation at 60 °C,sections were cut using a Leica Ultracut UCT ultrami-crotome. Semi-thin sections (0.9-1.0 μm thick) wereprepared and stained with aqueous methylene blue /
Plant Soil

Table 2 Plant material used for anatomical examinations
GenusSubgenusSection
Species Habit,growth
Source
Pinguicula
Isoloba
Agnata Pinguicula gigantea Luhrs t (l), sto Ex origin Oaxaca, Mexico1*
Genlisea
Genlisea
Genlisea Genlisea hispidula Stapf a, ros Ex origin Africa1**
Genlisea repens Benj. a, sto Ex origin South America2**
Utricularia
Polypompholyx
Polypompholyx Utricularia multifida R.Br. a, ros Ex origin Western Australia3*
Pleiochasia Utricularia volubilis R.Br. (e-)s, ros Ex origin SWWesternAustralia4***
Utricularia paulineae A.Lowrie a, sto Ex origin SWWestern Australia5**
Utricularia oppositiflora R.Br.† a(-e), sto Ex origin Newcastle, New SouthWales, Australia4**
Utricularia dichotoma subsp. aquiloniaR.W.Jobson†
a, sto Ex origin Katoomba, New SouthWales, Australia1**
Utricularia tubulata F.Muell. f Ex origin Australia6****
Lasiocaules Utricularia uniflora R.Br. a, sto Ex origin Australia5**
Bivalvaria
Australes Utricularia delicatula Cheeseman a, sto Ex origin Whangamarino wetlands,New Zealand5**
Utricularia
Foliosa Utricularia tricolor A.St.Hil. a, sto Araraquara, São Paulo State, Brazil7
Utricularia Utricularia stygia Thor s, sto Treboň, Czech Republic8
Utricularia breviscapa Wright ex Griseb. f, sto, flo Araraquara, São Paulo State, Brazil7
Vesiculina Utricularia cucullata A.St.Hil. & Girard f, sto a) Itirapina, São Paulo State, Brazil;b) Delfinópolis, Minas GeraisState, Brazil9
Utricularia purpurea Walter (s-)f, sto Ex origin unknown4****
The taxonomy of genera, subgenera, sections and species follows Cieslak et al. (2005), Fleischmann (2012), Jobson et al. (2018), and Jobsonand Baleeiro (2020)
†Treated as U. dichotoma sensu Taylor (1989) in Reut and Płachno (2020), but subsequently underwent taxonomic revision by Jobson andBaleeiro (2020)
Habit of sample (alternative/usual life form in situ in brackets): amphiphyte (a); emergent hydrophyte (e); free-floating (suspended)hydrophyte (f); lithophyte (l); submerged hydrophyte (s); terrestrial plant (t). Growth form: with floats (flo); rosulate (ros); stoloniferous (sto)
Sources, ex situ: 1 Cultivation in the Botanic Garden of Jagiellonian University in Kraków, Poland. 2 Cultivation in the Department of PlantCytology and Embryology, Institute of Botany, Jagiellonian University in Kraków, Poland. 3 Collection of Kamil Pásek, Ostrava,Czech Republic, http://www.bestcarnivorousplants.net. 4 Collection of Dr. Lubomír Adamec, Institute of Botany of the Czech Academyof Sciences at Treboň, Czech Republic. 5 Collection of Nigel Hewitt-Cooper, UK, http://www.hccarnivorousplants.co.uk. 6 Collection ofCorin Gardiner, New Zealand. *Grown in humid soil. **Grown in wet to saturated soil. ***Grown under water in an aquarium, anchored insoil. ****Grown free-floating
Sources, collected in situ: 7 Farm near Araraquara, São Paulo State, Brazil; Fazenda Palmeiras, swamp c. 50m from the road; V.F.O.Mirandaet al. 2856. 8 Near Treboň, Czech Republic. 9 From two populations: (a) road Itirapina-Brotas, São Paulo State, Brazil; swamp c. 10 m fromthe road; V.F.O.Miranda et al. 2859; (b) Delfinópolis, Minas Gerais State, Brazil; flooded area c. 250 m from the road; V.F.O.Miranda et al.2877
Plant Soil

azure II (MB/AII) for 1–2 min (Humphrey and Pittman1974). Additional material was fixed as above, embed-ded in Technovit 7100 (Kulzer, Germany) and proc-essed as in Lustofin et al. (2020). Sections were exam-ined and photographed using a Nikon Eclipse E400microscope with a Nikon DS-Fi2 camera and NIS-Elements D 4.00.00 4.0 software (Nikon, Tokyo, Japan)and using an Olympus BX60microscope with an Olym-pus XC50 digital camera and cellSens Standard Soft-ware (Olympus, Tokyo, Japan).
Multivariate analyses
To quantitatively evaluate morpho-anatomical charactersand their correlationwith life forms in theLentibulariaceae,we selected characters 1–9 from Table 1, as only thoseprovided variability across the chosen taxa. The traits wererecorded from 14 species with comparable organs (leavesfor the morphological and stolons for anatomical datacollection). To visualize how the species can be optimallyclustered, if life forms can be suitably grouped, and whichmorpho-anatomical features contribute most to the group-ings, the data dimensionality was reduced through severalcomputing techniques while maintaining the most impor-tant information. We created three sets of variables formultivariate analyses on the 14 taxa. Dataset I contained‘life forms’ and ‘location of stolons’ in addition to the
morpho-anatomical characters 1–9. The trait ‘life forms’was selected to assess correlations of life forms with allother traits. ‘Location of stolons’ was included in dataset Ito evaluate how far the stolon related traits were influencedby the location of the stolons in the respective substrate. Indataset II we excluded ‘life forms’, and dataset IIIcontained the morpho-anatomical traits only. All biometrictraits and their values used in the analyses are presented inTable 3.
We ran a graph-based clustering with BioVinci(BioTuring Inc) version 3.0.9 to calculate the “best fit”dimensionality reduction method with highest Silhou-ette Score [-1, 1]. In addition, the tool identified suitabletraits supporting the evaluated hierarchical clusters in adecision tree by applying cost complexity pruningcriteria such as Gini index and Entropy. The samemethods were used when clusters were validated ac-cording to life forms (‘life form clustering’). We ranPrincipal Component Analyses (PCA) by exclusion vs.inclusion of pre-processing with scaling, and by apply-ing singular value decomposition (SVD) with imputa-tion to calculate principal components. Some PCA scat-ter plots were overlayed with vectors (traits), addinginformation on the correlations of traits (direction ofvectors) and the influence of traits (lengths of vectors).PCA statistics were taken from BioVinci (BioTuringInc) version 1.1.5. The computing and illustration of
Table 3 Biometric traits and their values used in multivariate analyses
Trait (variable) Value
L) Life form terrestrial plant (1), amphiphyte (2), submerged hydrophyte (4), free-floating(suspended) hydrophyte (5)
0) Stolon location above ground, aerial (1), subterranean, few mm to few cm in the soil (2),submersed, few cm under water - in soil or floating (3)
1) Anchoring organs roots present (1), anchor stolons (rhizoids) present (2),roots/rhizoids absent (3)
2) Leaf form large-fleshy obovate (1), small spathulate-circular (2), (small) narrow-linear (3),dissected (4)
3) Leaf thickness mesophyll layers: > 10 (1), 8–10 (2), 4–7 (3), < 4 (4)
4) Stolon chloroplasts none identified (1), in parenchyma (2), in parenchyma and epidermis (3)
5) Stolon diameter thickness, in mm: > 0.56 (1), 0.41–0.56 (2), 0.25–0.40 (3), < 0.25 (4)
6) Stolon xylem Xylem tracheary element (vessel), diameter in µm: > 20 (1), 14–20 (2),< 14 (3), none identified or xylem lacuna only (4)
7) Stolon intercellular spaces nil (1), slight intercellular spaces (2), moderate/extensive intercellular spaces towheel-shaped aerenchyma (3), crescent to hollow aerenchyma (4)
8) Stolon external glands absent (1), present (2)
9) Stolon supporting tissue present (1), absent (2)
Plant Soil

confidence ellipses (CI = 95%) of clusters in PCA weredone with ClustVis (Metsalu and Vilo 2015).
Results
Morpho-anatomy of leaves and main stolons
We characterized the anatomical traits of leaves andmain/runner stolons (see Table 4 for a fulldescription, including references to figures).
Additional data can be found in Table 5, which wasused for multivariate analyses.
Morpho-anatomy of other vegetative organs
The stoloniferous P. gigantea showed adventitious rootsarising from a short vertical stem, without branching orroot cap, but with unicellular hairs. Vegetative organs ofGenlisea and Utricularia did not exhibit external rootstructures and adventitious roots. SomeUtricularia spe-cies developed other anchoring organs (so called anchor
Table 4 Summary of morpho-anatomical characters of leaves and stolons
Leaves (Fig. 1a-e) Stolons (Figs. 1f-h and 2)
Organization Simple Simple
Cross-section Dorsiventral (but circular in trap bearingU. volubilisleaves)
Circular
Epidermis Uniseriate Uniseriate
Cuticle* Cuticularized Cuticle seemingly thin (e.g. Figs. 1f and g and 2e)but somewhat thicker in U. volubilis (Fig. 2d)
Stomata Stomatous (except for submersed leaves) No stomata found
Glands P. gigantea: Stalked and sessile glandsGenlisea and Utricularia: button-like glands
Genlisea and Utricularia: button-like glands abun-dant (Fig. 2e) with generally one head cell (butwith two head cells in U. breviscapa)
Mesophyll (leaf) /cortex (stolon)
Palisade parenchyma absent.P. gigantea: prominent water storage tissueGenlisea and Utricularia: Spongy parenchymaIntercellular spaces or lacunae present (Fig. 1c-e)
Pentagonal to polygonal, thin-walled and unlignifiedparenchymatic cells (more irregular cell shapes inP. gigantea, Fig. 1g).
Intercellular spaces: none (P. gigantea, Fig. 1g),small (e.g. G. repens, Fig. 1f), orfrequently aerenchymatic (Fig. 2a, c-i),predominantly lysigenous
No hypodermis/exodermis found
Chloroplasts P. gigantea: adaxially in mesophyll, abaxially inpatches, and in bundle sheaths; occasionallywith amyloplasts
Genlisea andUtricularia: in spongy parenchyma; insubmerged and free-floating Utricularia speciesalso in the epidermis (Fig. 1e)
None or very few to numerous in parenchyma; insome species also in the epidermis (Fig. 2i)
G. repens. Occasionally with amyloplasts (Fig. 1f)
Vascular tissue P. gigantea: bundle shape almost a closed circle,with 3 to >13 lignified xylem tracheary elements(vessels) scattered towards the centre and 3 to >13phloem groups towards the abaxial side(Fig. 1a, b)
Genlisea and Utricularia: 1–2 xylem tracheary ele-ments (vessels) and 1–3 phloem groups (e.g.Fig, 1c), but occasionally no xylem found(Fig. 1d, e)
Ectophloic central cylinder with unlignified pithcells and without pericycle. ≤4 lignified xylemtracheary elements and ≤12 or more phloemgroups (Figs. 1f-h and 2). Xylem vessels notfound in some stolons of U. stygia andU. breviscapa, or central (xylem) lacuna occa-sionally present (Fig. 2a, g). Phloem generallypresent but not identifiable in U. uniflora (Fig. 2f).
Supporting tissue Absent Generally absent, but present internal to parenchymasheath / endodermis in G. repens and U. tricolor(Fig. 1f, h)
*not conclusively demonstrated by the applied methods
Plant Soil

stolons or “rhizoids”) with geotropically positive growth(see also Table 5).
In the root of P. gigantea, parenchyma was denselypacked. Except for U. volubilis, all examined trap stalksand the anchor stolon of U. multifida showed spacious(‘hollow type’) aerenchyma, occupying nearly thewhole interior of the organ, and leaving only a bridgeof parenchyma between the vascular strand and theepidermis.
Anatomy of the central cylinder
The central cylinder was generally bordered by auniseriate parenchyma sheath in roots and in main sto-lons (Figs. 1f and 2d, e, g and i), in which an endodermis(Fig. 1h, 2a-c, h) with Casparian strips (Fig. 2b, h) wasvisible in U. tricolor, U. oppositiflora, U. delicatula,and U. stygia. The parenchyma sheath and the thin-walled endodermis with Casparian strips did not differgreatly in shape from cortex parenchyma. In the runner
stolon of P. gigantea, the sheath structure was lessapparent but implied by smaller parenchyma cells irreg-ularly surrounding and interlacing the vascularized cen-tral zone (Fig. 1g).
Multivariate analyses
The results of cluster analyses of datasets I-III are pre-sented in Table 6, showing Silhouette Scores, clusters ofspecies, and decisive traits for the branching (clusteringof species) for selected dimensionality reduction andclustering methods. A selection of multivariate analysesof datasets I-III is visualized in Fig. 3, illustrating cor-relations (distances) and clusters of species.
Even though UMAP is not an ideal method forcluster analysis, as the data tend to be uniformlydistributed, the separation of two clusters (terrestri-al-amphibious and submerged-free-floating) withgraph-based clustering of dataset I was distinctiveand received support by PCA (Table 6; Fig. 3a). The
Table 5 Data matrix of species and their values of biometric traits used in multivariate analyses
Dataset I
Dataset II
Dataset III
Species L 0 1 2 3 4 5 6 7 8 9
PGI 1 1 1 1 1 1 1 2 1 1 2
GRE 2 2 3 2 2 3 1 2 2 2 1
UVO 4 3 2 3 3 2 2 3 3 2 2
UPA 2 2 2 2 3 1 3 3 4 2 2
UOP 2 2 2 3 3 2 1 2 4 2 2
UDI 2 2 2 2 3 3 3 3 4 2 2
UTU 5 3 3 3 4 3 3 4 3 2 2
UUN 2 2 2 2 3 1 4 3 4 2 2
UDE 2 2 2 3 2 1 4 3 3 2 2
UTR 2 2 2 2 3 2 2 1 3 2 1
UST 4 3 2 4 4 2 2 2 3 2 2
UBR 5 3 3 4 4 3 1 4 3 2 2
UCU 5 3 3 4 4 3 3 3 3 2 2
UPU 5 3 3 4 4 3 2 3 3 2 2
Species (samples): G. repens (GRE); P. gigantea (PGI); U. breviscapa (UBR); U. cucullata (UCU); U. delicatula (UDE); U. dichotomasubsp. aquilonia (UDI);U. oppositiflora (UOP);U. paulineae (UPA);U. purpurea (UPU);U. stygia (UST);U. tricolor (UTR);U. tubulata(UTU); U. uniflora (UUN); U. volubilis (UVO)
Traits (variables, further defined in Table 3): Life form (habit) (L), stolon location (0), anchoring organ (1), leaf form (2), leaf thickness (3),stolon chloroplasts (4), stolon diameter (5), and stolon xylem (vessels / diameter) (6), stolon intercellular spaces (7), stolon external glands(8), stolon supporting tissue (9)
Plant Soil
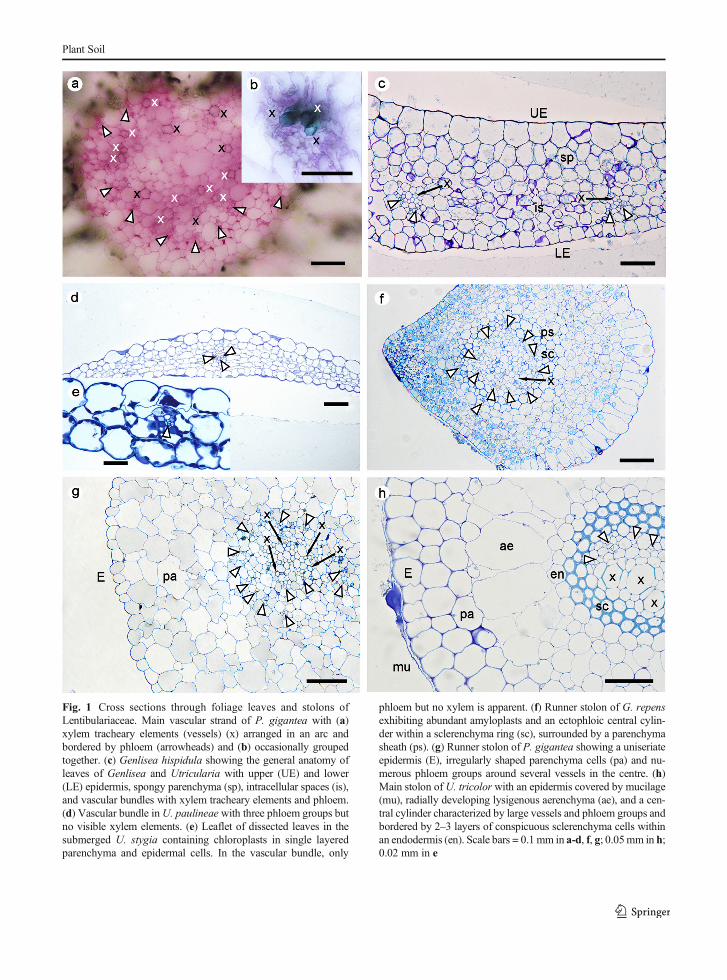
Fig. 1 Cross sections through foliage leaves and stolons ofLentibulariaceae. Main vascular strand of P. gigantea with (a)xylem tracheary elements (vessels) (x) arranged in an arc andbordered by phloem (arrowheads) and (b) occasionally groupedtogether. (c) Genlisea hispidula showing the general anatomy ofleaves of Genlisea and Utricularia with upper (UE) and lower(LE) epidermis, spongy parenchyma (sp), intracellular spaces (is),and vascular bundles with xylem tracheary elements and phloem.(d) Vascular bundle inU. paulineae with three phloem groups butno visible xylem elements. (e) Leaflet of dissected leaves in thesubmerged U. stygia containing chloroplasts in single layeredparenchyma and epidermal cells. In the vascular bundle, only
phloem but no xylem is apparent. (f) Runner stolon of G. repensexhibiting abundant amyloplasts and an ectophloic central cylin-der within a sclerenchyma ring (sc), surrounded by a parenchymasheath (ps). (g) Runner stolon of P. gigantea showing a uniseriateepidermis (E), irregularly shaped parenchyma cells (pa) and nu-merous phloem groups around several vessels in the centre. (h)Main stolon of U. tricolor with an epidermis covered by mucilage(mu), radially developing lysigenous aerenchyma (ae), and a cen-tral cylinder characterized by large vessels and phloem groups andbordered by 2–3 layers of conspicuous sclerenchyma cells withinan endodermis (en). Scale bars = 0.1 mm in a-d, f, g; 0.05mm in h;0.02 mm in e
Plant Soil

primary branching leading to these clusters were,however, mainly influenced by the two non-morpho-anatomical traits ‘life form’ and ‘stolon lo-cality’ (Table 6). While ‘life form’ as trait is obvi-ously a bias of life form clustering, ‘stolon location’has an affinity to the life form (by the milieu) but
also to growth and consequently to morphology. Asshown by the biplots (Fig. 3a, c), ‘stolon location’correlated to some extent to stolon related traits,namely to ‘chloroplast occurrence’ and ‘xylem di-ameter’, while the contribution of ‘occurrence ofexternal glands’ was weak (Fig. 3a, c). Hence, in
Plant Soil

submerged and free-floating species, main stolonstended to develop chloroplasts in parenchyma andepidermis, and smaller xylem tracheary elements.
The exclusion of the traits ‘life form’ and ‘stolonlocation’ led to a certain decrease of distances ofclusters in PCA (compare Fig. 3a, c, d), which be-came also evident by lower Silhouette Scores(Table 6). The correlation of species was not ex-tremely altered by excluding these traits from thedatasets, especially because ‘anchoring organ’, ‘leafform’ and ‘leaf thickness’ were directly correlated to‘life form’ and ‘stolon location’ and, therefore, like-wise contributed to the same (PC1) direction of dis-tribution. In the PCA of all datasets, this approxi-mately horizontal distribution followed a “life formtransect” from free-floating to submerged to amphib-ious species towards the terrestrial P. gigantea.
By considering the decisive characters across thePCA of datasets I-III, it appeared that the separation offree-floating and submerged species (with or withoutU. volubilis) from other clusters was strongly influenced
by ‘stolon location’ or ‘leaf thickness’, and that free-floating species split from submerged species mainlydriven by ‘anchoring organ’ and ‘chloroplast occur-rence’ (Table 6).
The vertical (PC2) distribution was generally relat-ed to ‘stolon thickness’ with some assistance from‘stolon intercellular spaces’ (Fig. 3a, c). This wasmainly apparent in the amphibious group and reflectedin the direction of the corresponding confidence ellip-se in Fig. 3d. Being transversal to the horizontaltransect, stolon thickness was not correlated to lifeforms. The vertical scattering of amphibious speciesrepresented a gradient of species with thin main sto-lons containing crescent to hollow aerenchyma (e.g.U. uniflora) to species with thick main stolons (e.g.G. repens). However, using graph-based clusteringwith datasets II and III generated a separation of agroup around U. delicatula from a diffuse grouparound U. tricolor (Fig. 3b-d; Table 6). The splitwas diminished when a few theoretical terrestrial spe-cies similar to P. gigantea were added and ‘stolonthickness’ was omitted from the datasets. In the PCAscatter plot of the accordingly modified dataset III,U. tricolor was much closer to U. delicatula, andU. oppositiflora was between U. dichotoma andU. paulineae/U. uniflora (not shown here).
By applying Kernel PCA (KPCA) in dataset II and III,the correlations of species in the datasets differed mainlyin the cluster assignment of U. volubilis (Table 6), whichaffiliated with U. stygia and the free-floating group orwith the terrestrial-amphibious group (Fig. 3b), respec-tively. When data were pre-processed by unit variancescaling (leading to more or less equally strong traits), thedensity of clusters was increased andU. oppositiflorawasmuch closer to the submerged taxa than to U. tricolor(e.g. Fig. 3d vs. a and c).
Discussion
Morpho-anatomical traits support life form clusteringof species in Lentibulariaceae
The applied multivariate analyses show that life formclustering is acceptably supported by adding someterrestrial species and discarding the interfering char-acter ‘stolon thickness’, even though the submergedgroup (U. stygia, U. volubilis) tends to overlap withthe free-floating and/or the amphibious group. The
�Fig. 2 Anatomical structures of stolons of Utricularia. a Mainstolon ofU. delicatulawith a uniseriate epidermis (E), aerenchyma(ae) between shrinking parenchyma cells (pa), and an endodermis(en) encompassing a vascular strand with two adjacent xylemtracheary elements (x) and a central lacuna (cl) in a phloem ring(arrowheads). b Close-up of the central cylinder of a, showinglignified Casparian strips (black arrowheads) in the endodermis.c Runner stolon of U. oppositiflora with half-moon shaped aeren-chyma in one sector, representing a unique distribution, designatedas “crescent” pattern. d Main stolon of U. volubilis containingmoderate intracellular spaces (is) and a central cylinder with theappearance of a two-part parenchyma sheath (ps), accommodatinga double vascular strand with one xylem tracheary element and 4phloem groups each. e Utricularia dichotoma subsp. aquiloniashowing extensive tangentially lysigenous aerenchyma dominat-ing the interior of the runner stolon. f Main stolon of U. unifloraexhibiting an advanced decay of cortex cells, forming ‘hollowtype’ aerenchyma. In the vascular tissue, phloem is indistinct andonly one xylem tracheary element is visible. g Main stolon ofU. tubulata with schizogenous aerenchyma and a central xylemlacuna (xl) in the vascular strand. Thick-walled cells are abundantwithin the pith. h Detailed view into the stele of a main stolon ofU. stygia revealing an endodermis with Casparian strips encirclingthe vascular cylinder. i Main stolon of U. cucullata showingplastids in the epidermis and in scanty parenchyma. The aeren-chyma pattern is wheel-shaped. The vascular bundle exhibits onexylem tracheary element and a sieve tube (arrowhead). Scalebars = 0.1 mm in a, c, f; 0.05 mm in d, e, g, i; 0.02 mm in h;0.01 mm in b
Plant Soil

datasets do not contain any emergent species. How-ever, U. volubilis and U. oppositiflora occur asemergent hydrophytes in some natural habitats(Taylor 1989; Jobson and Baleiero 2020). As expect-ed, both species show a strong correlation and haveaffinities with the amphibious as well as with thesubmerged habit.
Leaf traits
Our results demonstrate that leaves tend to be thinnerand narrower from terrestrial to free-floating taxa. Onone end of the transect, the anatomy of the fleshyP. gigantea leaf is largely consistent with the succulentleaves of several Mexican Pinguicula species of
Table 6 Cluster analyses of datasets
Dataset (variables) Method (Silhouette Score) Decision tree (cluster branching)Clusters (c)
Decisive traits forcluster separation
I (L, 0, 1–9) Graph-based clustering:UMAP (0.7), PCA (0.5)
Branching 1:S+F (c0),A+PGI (c1)
L (Gini or Entropy≤0.04) or 0 (Entropy>0.04)
Life form clustering:PCA (0.4)
Branching 1:S+F (c0),A+PGI (c1)Branching 2:S (c0a), F (c0b),A (c1a), PGI (c1b)
L/0
L/1/4Several
II (0, 1–9) Graph-based clustering:KPCA (0.7),
PCA (0.5)
Branching 1:S+F (c0),others (c1+c2)Branching 2:PGI, GRE, UTR, UOP (c1),UDI, UDE, UPA, UUN (c2)
0
5/6
Life form clustering:PCA (0.2)
Branching 1:S+F (c0),A+PGI (c1)Branching 2:S (c0a), F (c0b),A (c1a), PGI (c1b)
0
1/4Several
III (1–9) Graph-based clustering:KPCA (0.6),
PCA (0.5)
Branching 1:F+UST (c1),others (c0+c2)Branching 2:PGI, GRE, UTR, UOP, UVO (c0),UDI, UDE, UPA, UUN (c2)
3
5
Life form clustering:PCA (0.1)
Branching 1:F+UST (c1),others (c0+c2)Branching 2:F (c1a), UST (c1b),PGI (c0a), A+UVO (c0b)
3
1/4/6Several
Abbreviations: Kernel PCA (KPCA); Uniform Manifold Approximation and Projection (UMAP); Principal Components Analysis (PCA)
Species: G. repens (GRE); P. gigantea (PGI); U. delicatula (UDE); U. dichotoma (UDI); U. oppositiflora (UOP); U. paulineae (UPA);U. stygia (UST); U. tricolor (UTR); U. uniflora (UUN); U. volubilis (UVO)
Plant groups with the same life forms: amphiphytes (A); free-floating hydrophytes (F); submerged hydrophytes (S)
Clusters after branching, e.g.: S + F (c0) = S and F build together cluster 0. S (c0a), F (c0b) = S and F are two subclusters (a and b) of cluster 0
Decisive traits (further defined in Table 3): Life form (habit) (L), stolon location (0), anchoring organ (1), leaf thickness (3), stolonchloroplasts (4), stolon diameter (5), and stolon xylem (vessels / diameter) (6)
Traits, e.g.: 1/4 = traits 1 and 4 are more or less interchangeably decisive on the branching
Plant Soil

Fig. 3 Visualization of multivariate analyses with graph-basedclustering of stoloniferous Lentibulariaceae species. a PCA biplotof dataset I, showing two clearly separated clusters. The sub-merged UST and UVO are between amphibious and free-floating species. Variances PC1 = 58.71%, PC2 = 20.01%, PC1-3 = 86.4%. b Scatter plot of species according to KPCA (best fit)of dataset III with three clusters. c PCA biplot of dataset III withthe same clusters as in b. PC1 = 44.38%, PC2 = 28.30%, PC1-3 = 82.64%. d PCA scatter plot of dataset II with unit-variancescaling as pre-processing. Amphiphytes (amph) and free-floatinghydrophytes (free) are circumscribed by confidence (prediction)ellipses (CI = 95%). Both submerged species are imbedded in the
amphibious group. PC1 = 47.89 %, PC2 = 21.00 %, PC1-3 =82.61 %. No scaling was applied to analyses a-c. Species:G. repens (GRE); P. gigantea (PGI); U. breviscapa (UBR);U. cucullata (UCU); U. delicatula (UDE); U. dichotoma subsp.aquilonia (UDI); U. oppositiflora (UOP); U. paulineae (UPA);U. purpurea (UPU); U. stygia (UST); U. tricolor (UTR);U. tubulata (UTU); U. uniflora (UUN); U. volubilis (UVO).Traits (vectors, further defined in Table 3): Life form (habit) (L),stolon location (0), anchoring organ (1), leaf form (2), leaf thick-ness (3), stolon chloroplasts (4), stolon diameter (5), and stolonxylem (vessels / diameter) (6), stolon intercellular spaces (7),stolon external glands (8), stolon supporting tissue (9)
Plant Soil

subgenera Isoloba and Pinguicula that occupy wet andperiodically dry habitats and may use C4 carbon fixation(cf. Studnička 1991). On the other end, in line with e.g.Sculthorpe (1967) and Colmer et al. (2011), our obser-vations on Lentibulariaceae confirm that obligate hydro-phytes differ distinctively from emergent, amphibiousor terrestrial herbaceous plants by having thin leaves.
Free-floating aquatics are limited to sectionsUtricularia and Vesiculina (both of subgenusUtricularia)and U. tubulata of subgenus Polypompholyx (Taylor1989). The two former sections carry leaves (if present)divided into capillary segments, whilst the morphologi-cally unique U. tubulata shows verticillate and undividedleaves (Taylor 1989), reminding of e.g. the submergedand freely suspended Hydrilla verticillata (cf. Yeo et al.1984) or the carnivorous free-floatingAldrovandavesiculosa with terminal traps on its leaves (cf. Adamec2018). A combination of dissected leaves and a verticillatephyllotaxy is known from e.g. the free-floating (rarelysubmerged) Ceratophyllum, which is also considered tobe rootless (Sculthorpe 1967; Cook 1999).
Our results corroborate that narrow and thin leavesare common features in amphibious and emergent plantsas adaptive response to submergence, as shown in e.g.Rumex palustris (Mommer et al. 2005) and Melaleucacajuputi (Tanaka et al. 2011). In the aquatic environ-ment, the absorption of light and dissolved gases isenhanced by an increased surface area to volume ratio,which is accomplished by thinner, lobed, and dissectedleaves (e.g. Sculthorpe 1967; Colmer et al. 2011).
Chlorophyllous tissues
In accordance with our results, simple anatomical struc-tures without a palisade layer are a common character-istic of submerged leaves (e.g. Wiegleb 1991; Catianand Scremin-Dias 2015; Krähmer 2016), indicating or-gan flexibility and/or reduced light conditions (cf.Sculthorpe 1967; Gotoh et al. 2018). A diminishedphotosynthesis rate is partly compensated by chloro-plasts occurring in both parenchymatic and epidermalcells of submersed organs (Catian and Scremin-Dias2015), as indicated in our study by e.g. U. stygiaand U. cucullata. Results of our multivariate analysescorroborate that aquatic life forms relate to a higherabundance of chloroplasts in tissues of submerged run-ner stolons (cf. Sculthorpe 1967; Wetzel 1988; Colmeret al. 2011). However, responding to (periodic) submer-gence or shady habitats, chloroplasts are occasionally
observed in the epidermis of leaves (Genlisea sp.; Lang1901) or in runner stolons (e.g. G. repens) of amphibi-ous species, although inG. repens, plastids may developinto amyloplasts under drier conditions. In Pinguicula,the absence of a chloroplast-rich palisade parenchymamay provide more flexibility to the leaf for carnivorousmechanisms (Heslop-Harrison 2004).
Anchoring organs
In the Lentibulariaceae, Pinguicula still shows “typical”roots (although partly reduced in some species such asP. gigantea). InUtricularia, “rhizoids” (anchor stolons)seem to take on the anchoring function from roots. Ouranalyses show that the presence/absence of anchor sto-lons is a delimitation between submerged and free-floating life forms in Utricularia. However, the base ofpeduncles in some suspended Utricularia species (e.g.U. australis, U. gibba) develop rhizoids (Taylor 1989),which may anchor the plant when flowering.
Structures of absorption and secretion
In the absence of an effective root system, the surfacesof other organs adopt the uptake of water and nutrients(cf. Schenck 1886; Adlassnig et al. 2005; Babourina andRengel 2010). As in most submerged hydrophytes, thecuticle of organs of the examined species seems to begenerally thin, since the mechanical protection is of lessimportance in the water. On the outer surface of allvegetative organs across the examined Genlisea andUtricularia species, we found numerous button-liketrichomes, which were considered being hydropotes(Lloyd 1942; Płachno et al. 2005) with absorptive and/or secretory function, depending on their ontogeny(Fineran 1980). On traps of Utricularia and Genlisea,mature external glands seem to expel water to the sub-strate (Fineran 1985; Fineran and Reut unpublisheddata), while some glandular trichomes on leaves andsubterranean organs of Genlisea and Utricularia pro-duce mucilage, e.g. to protect from drying, as defensivecompound against herbivores, as source of nutrients formicro-organisms, or as lubricant for penetrating the soil(cf. Lang 1901; Płachno et al. 2005; Rivadavia et al.2013). Since only Pinguicula lacks external bottom-liketrichomes on vegetative organs, the occurrence of theseglands does only marginally contribute to the life formclustering in the present study.
Plant Soil

Further histochemical and ultrastructural work (in-cluding tracers) on the epidermis, cuticle, and externalglands of Genlisea and Utricularia of various habitswould be helpful to increase the knowledge on perme-ability and the various functions of external glands inrelation to their position on the organ and their devel-opmental stages.
Conductive tissues
A general simplification of xylem tissue in subterraneanand submersed organs was evident in the species stud-ied. However, in the aquatic environment, the reductionof xylem and lignified elements does not necessarilyimpede the acropetal flow of water and nutrients(Sculthorpe 1967; Pedersen and Sand-Jensen1993; Lusaet al. 2011). Moreover, in some main stolons ofUtricularia, we were able to identify an endodermiswith Casparian strips (CS), which directs and controlsthe passage of water and minerals (cf. Barberon 2017).Although an endodermis with CS mainly occurs inroots, it is not uncommon in aerial shoots and leaves(Lersten 1997; Seago 2020) but more often establishedin subterranean or submersed stems (Dalla Vecchia et al.1999). In the “rootless” bladderworts, the conductivefunction of the endodermis has been seemingly trans-ferred from roots into stolons that manifest dynamicdevelopmental processes for roots, shoots, and leaves(Rutishauser 2016, 2020; Reut and Płachno 2020).
Due to the thin cell walls of the parenchyma sheaths,the capability to visualize the CSwas limited. Therefore,even if the CS could not be detected, the presence of CScannot be conclusively ruled out for the remaining spe-cies of the study. Other staining and fluorescence tech-niques should be applied to improve the identification ofCS and to gain insights into their composition (see e.g.Dalla Vecchia et al. 1999; Naseera et al. 2012; Seago2020).
Aerenchyma
To enhance the exchange of gases within and betweenroots, stems, and leaves, plants develop intercellularcavities or chambers of various sizes (Takahashi et al.2014), whereby the enlarged space is called ‘aerenchy-ma’ (Seago et al. 2005; Jung et al. 2008), which addi-tionally facilitates buoyancy (Krähmer 2016). Althoughaerenchyma is typical for hydrophytes, it is also knownfrom some terrestrial plants caused by several forms of
stress (Evans 2003; Jung et al. 2008). The morpho-anatomical differences between emergent and sub-merged taxa studied reflect a big step from slight orperiodic inundation to complete and permanent submer-gence. Colonization of habitats with standing water (e.g.swamps, pools, lakes) has evolved with the abundanceof large air cavities in submerged organs, supporting anefficient diffusion of gases (cf. Wetzel 1988).
Lysigenous aerenchyma results from programmedcell death and cell collapse, which can be induced byhypoxia, nutrient deficiency and/or ethylene accumula-tion (Evans 2003; Striker 2012). Since lysigenous aer-enchyma formation is dominant in the material studied,these factors may all be present in the preferred habitatsof Genlisea and Utricularia, in which their carnivorousstrategy (partly) compensates the limited availability ofcertain nutrients (cf. Adamec 1997). Lysigenous aeren-chyma was also reported by Schweingruber et al. (2020)from stems of submerged Utricularia species and thecarnivorous free-floatingAldrovanda vesiculosa.
Our results indicate that the ‘wheel-shaped’ aeren-chyma pattern is abundant in main stolons of submergedand free-floatingUtricularia species, which is supportedby earlier observations on shoots of U. tenuicaulis (syn.U. australis) and U. gibba of section Utricularia byJung et al. (2008) and Chormanski and Richards(2012), respectively. Within the same order Lamiales,it was reported from the submerged Limnophilasessiflora by Jung et al. (2008).
The unique ‘crescent’ type aerenchyma, observed inrunner stolons ofU. oppositiflora, seems to be due to thedorsiventral symmetry of these stolons with organs aris-ing on nodes on the upper side of the runner (cf. Reutand Płachno 2020). The continuing decay ofparenchymatic cells (seemingly by tangential lysigeny)leads to a special ‘hollow type’ aerenchyma, which isevident in several subterranean organs of the examinedamphibious species of Genlisea (see also Lloyd 1942)and Utricularia subg. Polypompholyx.
Schizogeny, as visible in main stolons ofU. volubilisand U. tubulata of section Pleiochasia, is developed bycell differentiation, but molecular mechanisms substan-tial for this formation type are not yet known (Yamauchiet al. 2018).
Supporting tissues
Our results confirm earlier observations on Genlisea(Merl 1915) andUtricularia (Schenck 1886; Kamieński
Plant Soil

1891), whereupon the anatomy of thicker stems is char-acterized by a central cylinder and by an increasingnumber of vascular elements and the occurrence ofsupporting tissue, as indicated by sclerenchyma aroundthe pith in runner stolons of G. repens and U. tricolor.Sclerenchyma was also reported below the epidermis ofadhesive anchor stolons of the rheophyticU. neottioidesby Płachno et al. (2020a). However, since only twoamphibious species in our dataset showed sclerenchymaaround the pith in runner stolons, this trait did notcontribute greatly to the delimitation of life forms inthe multivariate analyses.
Evolutionary aspects of the Lentibulariaceae
With respect to the morpho-anatomical features, ourfindings reveal that the reduction of roots and conduc-tive tissues (especially xylem) are taxonomically com-mon characteristics of all genera of the Lentibulariaceae,which points to an ancestor that was adapted to at leastseasonally wet and/or periodically inundated habitats.The more free water is abundant, the less protection andstability of organs is required, which is apparentlyreflected in a loss of the root cap in some Pinguiculaspecies (cf. Rutishauser 2016, 2020), the palisade layerin leaves, and supporting tissues in subterranean/submersed organs within the Lentibulariaceae.
Apart from an abandonment of several developmen-tal processes for roots, Genlisea and Utricularia shareother more hydrophilic features, such as (in general)more delicate organs with reduced cuticle and button-like glandular trichomes on the surface of leaves andsubterranean/submersed organs, indicating a next levelin the adaptation to submergence but also a shift of thecarnivory from aerial leaves (in Pinguicula) to under-ground leaf-like organs (in Genlisea-Utricularia). Inother words, costs for innovative structures used forthe uptake of nutrients are compensated by less invest-ment in other structures (e.g. cortex, vascular elementsand supporting tissue), which seems to be facilitated in awatery milieu.
Most of the plants in our study were kept ex situ intheir preferred conditions, and hence revealed moreadaptive rather than reactive morpho-anatomicalfeatures. We recommend that future research onhydrophytes with a comparable plasticity takes the dif-ferentiation of emergent and submerged life forms intoconsideration. An experimental approach with variablesettings (e.g. water levels, light) could unlock which
morpho-anatomical changes are adaptations and whichones are responses.
Morpho-anatomical and functional adaptations to a lifebetween land and water
The key findings of our study are:
– Life forms along the hydric transect in theLentibulariaceae correlate directly to the locationof the stolon in the substrate and to the shape andthickness of leaves, which aims at enhancing theabsorption of sparse light, nutrients, dissolved gasesas well as the flexibility of leaves with increasingdepth of submergence of vegetative organs.
– Submerged and free-floatingUtricularia speciestend to develop chloroplasts in parenchymatic andepidermal cells of suspended main stolons, whichsupports the collection of light.
– Life forms in the Lentibulariaceae correspond withthe reduction of the typical root morphology andwith the presence and type of anchorage.
– Buoyancy and gas exchange are enhanced by largeintercellular cavities that are abundant in stems andshoots of amphibious to free-floating taxa studied.Novel ‘crescent’ to special ‘hollow’ aerenchymapatterns develop in dorsiventrally organized stemsand shoots of amphibious Genlisea and Utriculariaspecies.
– Morpho-anatomical characters in vegetative organsof Genlisea and Utricularia strongly show adapta-tions to the hydric environment, which they sharewith many other aquatic plants. However, some ofthese features may have been partly driven by theevolutionary shift of carnivory from aerial to sub-terranean organs and hence by an alternative uptakeof certain minerals in nutrient-poor and submergedhabitats.
Supplementary Information The online version contains sup-plementary material available at https://doi.org/10.1007/s11104-021-04929-6.
Acknowledgements This research was partially supported fi-nancially by the Ministry of Science and Higher Education ofPoland as part of the statutory activities of the Department of Plant
Plant Soil

Cytology and Embryology, Institute of Botany, Faculty of Biolo-gy, Jagiellonian University in Kraków (N18/DBS/000002). Thescientific stay of BJP in Brazil was financed by the JagiellonianUniversity within the project Integrate JU-Comprehensive Devel-opment Programme of the Jagiellonian University as part of theKnowledge Education Development 2014–2020 Operational Pro-gramme co-financed by the European Social Fund in the EuropeanUnion (POWR.03.05.00-00Z309/17 − 00).
This paper is dedicated to Brian A. Fineran, mentor, friend,excellent taxonomist, and expert in the ultrastructure of Utriculariaand parasitic plants, and in good memories of Christopher D. K.Cook, a profound researcher of aquatic plants. We are cordiallygrateful to Lubomír Adamec (Institute of Botany of the CzechAcademy of Sciences, Třeboň, Czech Republic), Kamil Pásek (BestCarnivorous Plants Store, Czech Republic), and Corin Gardiner formaterial from their collections. We thank the editor and reviewers fortheir valuable comments and helpful suggestions for the manuscript.
Declarations
Conflict of interest The authors declare that they have no con-flict of interest.
References
Adamec L (1997) Mineral nutrition of carnivorous plants: a re-view. Bot Rev 63:273–299. https://doi.org/10.1007/BF02857953
Adamec L (2018) Biological flora of Central Europe: Aldrovandavesiculosa L. Perspect Plant Ecol 35:8–21. https://doi.org/10.1016/j.ppees.2018.10.001
Adlassnig W, Peroutka M, Lambers H, Lichtscheidl IK (2005)The roots of carnivorous plants. Plant Soil 274:127–140
Arber A (1920) Water plants. A study of aquatic angiosperms.Cambridge University Press, Cambridge
Babourina O, Rengel Z (2010) Ion transport in aquatic plants. In:Mancuso S, Shabala S (eds) Waterlogging signalling andtolerance in plants. Springer, Berlin, pp 221–238.https://doi.org/10.1007/978-3-642-10305-6_11
Barberon M (2017) The endodermis as a checkpoint for nutrients.New Phytol 213:1604–1610. https://doi.org/10.1111/nph.14140
Barrett SCH, Eckert CG, Husband CG (1993) Evolutionary pro-cesses in aquatic plant populations. Aquat Bot 44:105–145
Braendle R, Crawford RMM (1999) Plants as amphibians.Perspect Plant Ecol 2:56–78
Casper SJ (2019) The insectivorous genus Pinguicula(Lentibulariaceae) in the Greater Antilles. In: Englera 35.Botanic Garden and Botanical Museum Berlin. FreieUniversität Berlin, Berlin
Catian G, Scremin-Dias E (2015) Phenotypic variations in leafanatomy of Nymphaea gardneriana (Nymphaeaceae) dem-onstrate its adaptive plasticity. J Torrey Bot Soc 142:18–26
Chambers PA, Lacoul P, Murphy KJ, Thomaz SM (2008) Globaldiversity of aquatic macrophytes in freshwater.
Hydrobiologia 595:9–26. https://doi.org/10.1007/s10750-007-9154-6
Chormanski TA, Richards JH (2012) An architectural model forthe bladderwort Utricularia gibba (Lentibulariaceae). JTorrey Bot Soc 139:137–148
Cieslak T, Polepalli JS,White A et al (2005) Phylogenetic analysisof ‘Pinguicula (Lentibulariaceae): chloroplast DNA se-quences and morphology support several geographically dis-tinct radiations. Am J Bot 92:1723–1736. https://doi.org/10.3732/ajb.92.10.1723
Colmer TD, Winkel A, Pedersen O (2011) A perspective onunderwater photosynthesis in submerged terrestrial wetlandplants. AoB Plants 11:1–15
Cook CDK (1999) The number and kinds of embryo-bearingplants which have become aquatic: a survey. Perspect PlantEcol 2:79–102
Crawford RMM (2008) Plants at the margin. Ecological limits andclimate change. Cambridge University Press, New York
Dalla Vecchia F, Cuccato F, La Rocca N et al (1999) Endodermis-like sheaths in the submerged freshwater macrophyteRanunculus trichophyllus Chaix. Ann Bot 83:93–97
Evans DE (2003) Aerenchyma formation. New Phytol 161:35–49Fineran BA (1980) Ontogeny of external glands in the bladderwort
Utricularia monanthos. Protoplasma 105:9–25Fineran BA (1985) Glandular trichomes in Utricularia: a review
of their structure and function. Israel J Bot 34:295–330.https://doi.org/10.1080/0021213X.1985.10677028
Fleischmann A (2012)Monograph of the genusGenlisea. RedfernNatural History, Poole
Fleischmann A (2018) Systematics and evolution ofLentibulariaceae. II. Genlisea. In: Ellison AM, Adamec L(eds) Carnivorous plants: physiology, ecology, and evolu-tion. Oxford University Press, New York, pp 81–88.https://doi.org/10.1093/oso/9780198779841.003.0007
Fleischmann A, Roccia A (2018) Systematics and evolution ofLentibulariaceae. I. Pinguicula. In: Ellison AM, Adamec L(eds) Carnivorous plants: physiology, ecology, and evolu-tion. Oxford University Press, New York, pp 70–80.https://doi.org/10.1093/oso/9780198779841.003.0006
Gotoh E, Suetsugu N, Higa T et al (2018) Palisade cell shapeaffects the light-induced chloroplast movements and leafphotosynthesis. Sci Rep 8:1472. https://doi.org/10.1038/s41598-018-19896-9
Guisande C, Granado-Lorencio C, Andrade-Sossa C, Duque SR(2007) Bladderworts. Funct Plant Sci Biotechnol 1:58–68
Heslop-Harrison Y (2004)Pinguicula L. J Ecol 92:1071–1118Hidalgo O, Garcia S, Garnatje T et al (2015) Genome size in
aquatic and wetland plants: fitting with the large genomeconstraint hypothesis with a few relevant exceptions. PlantSyst Evol 301:1927–1936. https://doi.org/10.1007/s00606-015-1205-2
Humphrey C, PittmanG (1974) A simple methylene blue-azure II-basic fuchsin for epoxy-embedded tissue sections. StainTechnol 49:9–14
Hutchinson GE (1975) A treatise on limnology. Vol III.Limnological botany. Wiley, New York
Ibarra-Laclette E, Lyons E, Hernández-Guzmán G et al (2013)Architecture and evolution of a minute plant genome. Nature498:94–98
Jobson RW, Baleeiro PC (2020) Radiations of fairy-aprons(Utricularia dichotoma, Lentibulariaceae) in Australia and
Plant Soil

New Zealand: molecular evidence and proposal of new sub-species. Aust Syst Bot 33:278–310. https://doi.org/10.1071/SB19003
Jobson RW, Playford J, Cameron KM, Albert VA (2003)Molecular phylogenetics of Lentibulariaceae inferred fromplastid rps16 intron and trnL-F DNA sequences: implicationsfor character evolution and biogeography. Syst Bot 28:157–171
Jobson RW, Baleeiro PC, Guisande C (2018) Systematics andevolution of Lentibulariaceae: III. Utricularia. In: EllisonAM, Adamec L (eds) Carnivorous plants: physiology, ecolo-gy, and evolution. Oxford University Press, NewYork, pp 89–104. https://doi.org/10.1093/oso/9780198779841.003.0008
Jung J, Lee SC, Choi H-K (2008) Anatomical patterns of aeren-chyma in aquatic and wetland plants. J Plant Biol 51:428–439. https://doi.org/10.1007/BF03036065
Kamieński F (1891) Lentibulariaceae. In: Engler A, Prantl KAE(eds) Die Natürlichen Pflanzenfamilien. Part IV, 3b. W.Engelmann, Leipzig, pp 108–123
Krähmer H (2016) Aquatic and wetland weeds. In: Krähmer H(ed) Atlas of weed mapping. Wiley-Blackwell, Chichester,pp 185–400
Lang FX (1901) ) Untersuchungen über Morphologie, Anatomieund Samenentwicklung von Polypompholyx und Byblisgigantea. Flora 88:149–206
Legendre L (2000) The genus Pinguicula L. (Lentibulariaceae): anoverview. Acta Bot Gallica 147:77–95. https://doi.org/10.1080/12538078.2000.10515837
Lersten NR (1997) Occurrence of endodermis with a Casparianstrip in stem and leaf. Bot Rev 63:265–272
Li G, Hu S, Hou H, Kimura S (2019) Heterophylly: phenotypicplasticity of leaf shape in aquatic and amphibious plants.Plants 8:420. https://doi.org/10.3390/plants8100420
Lloyd FE (1942) The carnivorous plants. Chronica BotanicaCompany, Waltham
Lusa MG, Torres Boeger MR, de Chiara Moço MC, Bona C(2011) Morpho-anatomical adaptations of Potamogetonpolygonus (Potamogetonaceae) to lotic and lentic environ-ments. Rodriguésia 62:927–936
Lustofin K, Świątek P, Miranda VFO, Płachno BJ (2020) Flowernectar trichome structure of carnivorous plants from thegenus butterworts Pinguicula L. (Lentibulariaceae).Protoplasma 257:245–259. https://doi.org/10.1007/s00709-019-01433-8
Merl EM (1915) Beiträge zur Kenntnis der Utricularien undGenliseen. Flora 108:127–200
Metsalu T, Vilo J (2015) Clustvis: a web tool for visualizingclustering of multivariate data using principal componentanalysis and heatmap. Nucleic Acids Res 43:W566–W570.https://doi.org/10.1093/nar/gkv468
Mommer L, Pons TL, Wolters-Arts M et al (2005) Submergence-induced morphological, anatomical, and biochemical re-sponses in a terrestrial species affect gas diffusion resistanceand photosynthetic performance. Plant Physiol 139:497–508
Naseera S, Leea Y, Lapierreb C et al (2012) Casparian stripdiffusion barrier in Arabidopsis is made of a lignin polymerwithout suberin. Proc Natl Acad Sci USA 109:10101–10106.https://doi.org/10.1073/pnas.1205726109
Pedersen O, Sand-Jensen K (1993) Water transport in submergedmacrophytes. Aquat Bot 44:385–406
Płachno BJ, Jankun A (2004) Transfer cell wall architecture insecretory hairs of Utricularia intermedia traps. Acta BiolCracov Bot 46:193–200
Płachno BJ, Muravnik LE (2018) Functional anatomy of carniv-orous traps. In: Ellison AM, Adamec L (eds) Carnivorousplants: physiology, ecology, and evolution. OxfordUniversity Press, New York, pp 167–179. https://doi.org/10.1093/oso/9780198779841.001.0001
Płachno BJ, Faber J, Jankun A (2005) Cuticular discontinuities inglandular hairs of Genlisea St.-Hil. in relation to their func-tions. Acta Bot Gallica 152:125–130. https://doi.org/10.1080/12538078.2005.10515463
Płachno BJ, Adamec L, Kamińska I (2014) Relationship betweentrap anatomy and function in Australian carnivorous bladder-worts (Utricularia) of the subgenus Polypompholyx. AquatBot 120:290–296
Płachno BJ, Kamińska I, Adamec L, Świątek P (2017) Vasculartissue in traps of Australian carnivorous bladderworts(Utricularia) of the subgenus Polypompholyx. Aquat Bot142:25–31
Płachno BJ, Świątek P, Adamec L et al (2019a) The trap architec-ture of Utricularia multifida and Utricularia westonii (subg.Polypompholyx). Front Plant Sci 10:336. https://doi.org/10.3389/fpls.2019.00336
Płachno BJ, Świątek P, Miranda VFO et al (2019b) The structureand occurrence of a velum in Utricularia traps(Lentibulariaceae). Front Plant Sci 10:302. https://doi.org/10.3389/fpls.2019.00302
Płachno BJ, Adamec L, Świątek P et al (2020a) Life in the current:Anatomy and Morphology of Utricularia neottioides. Int JMol Sci 21:4474. https://doi.org/10.3390/ijms21124474
Płachno BJ, Silva SR, Świątek P et al (2020b) Structural featuresof carnivorous plant (Genlisea, Utricularia) tubers as abioticstress resistance organs. J Mol Sci 21:5143. https://doi.org/10.3390/ijms21145143
Reut MS (1993) ) Trap structure of the carnivorous plantGenlisea(Lentibulariaceae). Bot Helv 103:101–111
Reut MS, Płachno BJ (2020) Unusual developmental morphologyand anatomy of vegetative organs in Utriculariadichotoma—leaf, shoot and root dynamics. Protoplasma257:371–390. https://doi.org/10.1007/s00709-019-01443-6
Rivadavia F, Gonella PM, Fleischmann A (2013) A new andtuberous species of Genlisea (Lentibulariaceae) from theCampos Rupestres of Brazil. Syst Bot 38:464–470.https://doi.org/10.1600/036364413X666679
Rutishauser R (2016) Evolution of unusual morphologies inLent ibular iaceae (bladderworts and al l ies) andPodostemaceae (river-weeds): a pictorial report at the inter-face of developmental biology and morphological diversifi-cation. Ann Bot 117:811–832. https://doi.org/10.1093/aob/mcv172
Rutishauser R (2020) EvoDevo: Past and future of continuum andprocess plant morphology. Philosophies 5:41. https://doi.org/10.3390/philosophies5040041
Schenck H (1886) Vergleichende Anatomie der submersenGewächse. Theodor Fischer, Cassel
Schweingruber F, Kučerová A, Adamec L, Doležal J (2020)Anatomic atlas of aquatic and wetland plant stems.Springer, Cham. https://doi.org/10.1007/978-3-030-33420-8
Sculthorpe CD (1967) The biology of aquatic vascular plants.Edward Arnold Ltd., London
Plant Soil

Seago JL Jr (2020) Revisiting the occurrence and evidence ofendodermis in angiosperm shoots. Flora 273(2020):151709.https://doi.org/10.1016/j.flora.2020.151709
Seago JL Jr, Leland C, Marsh LC et al (2005) A re-examination ofthe root cortex in wetland flowering plants with respect toaerenchyma. Ann Bot 96:565–579
Silva SR, Moraes AP, Penha HA et al (2020a) The terrestrialcarnivorous plant Utricularia reniformis sheds light on envi-ronmental and life-form genome plasticity. Int J Mol Sci 21:3. https://doi.org/10.3390/ijms21010003
Silva RS, Płachno BJ, Carvalho SGM, Miranda VFO(2020)Genlisea hawkingii (Lentibulariaceae), a new speciesfrom Serra da Canastra, Minas Gerais, Brazil. PLoS One 15:e0226337. https://doi.org/10.1371/journal.pone.0226337
Striker GG (2012) Flooding stress on plants: anatomical, morpho-logical and physiological responses. Botany 1:3–28
Studnička M (1991) Interesting succulent features in thePinguicula species from the Mexican evolutionary centre.Folia Geobot Phytotaxon 26:459–462
Takahashi H, Yamauchi T, Colmer T, Nakazono M (2014)Aerenchyma formation in plants. Plant Cell Monogr 21:247–265
Tanaka K, Masumori M, Yamanoshita T, Tange T (2011)Morphological and anatomical changes of Melaleucacajuputi under submergence. Trees 25:695–704. https://doi.org/10.1007/s00468-011-0547-9
Taylor P (1989) The genus Utricularia: a taxonomic monograph.HMSO, London
Tiner RW (2017) Wetland indicators: a guide to wetland identifi-cation, delineation, classification, and mapping. CRC Press,Boca Raton. https://doi.org/10.1201/9781420048612
Wetzel RG (1988) Water as an environment for plant life. In:Symoens J-J (ed) Vegetation of inland waters. Kluwer,Dordrecht, pp 1–30
Wiegleb G (1991) Die Lebens- und Wuchsformen dermakrophytischen Wasserpflanzen und deren Beziehungenzur Ökologie, Verbreitung und Vergesellschaftung derArten. Tuexenia 11:135–147
Yamauchi T, Colmer TD, Pedersen O, Nakazonoa M (2018) )Regulation of root traits for internal aeration and tolerance tosoil waterlogging-flooding stress. Plant Physiol 176:1118–1130
Yeo RR, Falk RH, Thurston JR (1984) The morphology ofHydrilla (Hydrilla verticillata (L. f.) Royle). J Aquat PlantManage 22:1–17
Publisher’s note Springer Nature remains neutral with regard tojurisdictional claims in published maps and institutionalaffiliations.
Plant Soil





























