Embed Size (px)
Citation preview

Structural Determination of Sphingomyelin byTandem Mass Spectrometry With ElectrosprayIonization
Fong-Fu Hsu and John TurkMass Spectrometry Resource, Division of Endocrinology, Diabetes, and Metabolism, Department of Medicine,Washington University School of Medicine, St. Louis, Missouri, USA
Alkaline metal adduct ions of sphingomyelin were formed by electrospray ionization inpositive ion mode. Under low energy collisionally activated dissociation (CAD), the production spectra yield abundant fragment ions representative of both long chain base and fatty acidwhich permit unequivocal determination of the structure. Tandem spectra obtained byconstant neutral loss scanning permit identification of sphingomyelin class and specific longchain base subclass in the mixture. The fragmentation pathways under CAD were proposed,and were further confirmed by source CAD tandem mass spectrometry. The total analysis ofsphingomyelin mixtures from bovine brain, bovine erythrocytes, and chicken egg yolk is alsopresented. (J Am Soc Mass Spectrom 2000, 11, 437–449) © 2000 American Society for MassSpectrometry
Sphingomyelin is an important constituent in ner-vous tissue and plasma membranes of higheranimals [1–4]. It is present in all mammalian cells
and has been shown to be the source of free ceramideand sphingosine in cell membranes. Both the free cer-amide and sphingosine found in membranes have beenshown to have powerful second messenger properties[5–7]. Sphingomyelins are the major lipid fraction iso-lated from bovine brain and are found in bovine milk[8]. Because sphingomyelin is a metabolic precursor ofceramides, which are considered to be among the mostimportant compounds for skin protection, it has beenproposed for use as an ingredient in cosmetics [9, 10].
Sphingomyelin contains a fatty acyl group bonded toa basic nitrogen atom of the long chain base as amide,and a phosphocholine residue esterified to the primaryalcohol residue of the long chain base. The majorsubclass of sphingomyelin, namely sphingosine longchain base phospholipid, is composed of a phosphoryl-choline ester-bound to a ceramide (Figure 1).
Because of the complexity of polar lipids, character-ization of sphingomyelin has been accomplished byseparation of the different residues of the lipids afterhydrolysis [1, 6]. Therefore, information on the molec-ular structure of the intact compounds were lost. Todetermine the structure, several direct mass spectromet-ric ionization techniques, such as chemical ionizationwith a moving-belt interface [11], thermospray [12],discharge-assisted thermospray (plasmaspray) [13], at-
mospheric pressure chemical ionization (APCI) [14],fast-atom bombardment (FAB) [15, 16], and electro-spray ionization (ESI) [17, 18] have been attempted.Recently, Karlsson et al. [18] reported the determinationof sphingomyelin structure by HPLC/mass spectrome-try with electrospray ionization (ESI) and HPLC/tan-dem mass spectrometry with APCI source collisional-activated decomposition (CAD). This method permitsassignment of the fatty acid constituents and long chainbase of the molecule by CAD of the protonated cer-amides generated by source CAD, in combination withtandem mass spectrometry.
Both glycerophosphatidylcholine (GPC) and sphin-gomyelin are choline-containing species, which pro-duce predominately protonated species under ESI inpositive ion mode. In the presence of alkaline metal ionssuch as Na1 however, they produce predominately[M 1 Na]1 ions [19]. ESI analysis of synthetic andnaturally derived phospholipids can be enhanced witha sensitivity 2 to 3 orders of magnitude greater than thatachieved by FAB-MS [17, 19], and thermospray [20]. Werecently demonstrated that CAD tandem mass spectraof lithiated molecular species of GPC [21] and triglyc-eride [22] provide structural information which permitsthe determination of the composition and the positionof fatty acids in the glycerol backbone. Here, we reportthe structural characterization of sphingomyelin by ESICAD tandem mass spectrometry using its alkalinemetal adduct ion. The utility of low energy CADtandem mass spectrometry with constant neutral loss(CNL) scanning for structural identification and itspotential application in the quantification of sphingo-myelin mixtures is also demonstrated.
Address reprint requests to Fong-Fu Hsu, Box 8127, Washington UniversitySchool of Medicine, 660 S Euclid, St. Louis, MO 63110. E-mail:[email protected]
© 2000 American Society for Mass Spectrometry. Published by Elsevier Science Inc. Received September 2, 19991044-0305/00/$20.00 Revised December 1, 1999PII S1044-0305(99)00150-6 Accepted December 3, 1999

ExperimentalChemicals and StandardsAll sphingomyelin standards: 1-phosphocholine-N-palmitoyl-sphingosine (d18:1/16:0-SM), 1-phosphocho-line-N-stearoyl-sphingomyelin (d18:1/18:0-SM), 1-phos-phocholine-N-tetradodecenoyl-sphingosine (d18:1/24:1-SM), and sphingomyelin mixtures from bovine brain,bovine erythrocyte, and chicken egg yolk were pur-chased from Sigma Chemical (St. Louis, MO). Thedesignation of sphingomyelin is in the form of d-LCB/FA, with d referring to the stereochemistry of the longchain base (LCB) and FA referring to fatty acid.
Mass SpectrometryESI/MS analyses were performed on a Finnigan TSQ-7000 triple stage quadrupole mass spectrometerequipped with an electrospray ion source and con-trolled by Finnigan ICIS software operated on a DECalpha station. Solution of standard sphingomyelin andsphingomyelin mixture were dissolved in chloroform/methanol (1/4) at final concentration of 5 pmol/mL,which yields intense alkaline metal adduct ion in thepresence of Li1, Na1, or K1. Samples were infused (1mL/min) into ESI source with a Harvard syringe pump.The electrospray needle and the skimmer were atground potential and the electrospray chamber and theentrance of the glass capillary at 4.5 kV. The heatedcapillary temperature was 250 °C. The lithiated molec-ular cations were selected in the first quadrupole andcollided with Ar (2.3 mtorr) in the rf only secondquadrupole using a collision energy of 30–55 eV. Forthe source CAD tandem mass spectrometry experiment,the skimmer voltage was set at 30–45 V to generateabundant fragment ions which were then selected in thefirst quadrupole and collided with Ar (2.3 mtorr) in thesecond quadrupole using a collision energy of 25–35 eV.Typically, a 1-min period of signal averaging wasemployed for scan spectrum and 1 to 5 min wasemployed for tandem spectrum.
Results and DiscussionMolecular Weight Determination by Alkaline-Metal Adduct IonIn positive ion mode, sphingomyelins yield abundantprotonated molecular species ([M 1 H]1) by electro-
spray ionization. In the presence of alkaline metal ionshowever, sphingomyelins produce alkaline metal cat-ions [M 1 Cat]1 where Cat 5 Li1, Na1, K1. The ESImass spectra of the sphingomyelin isolated from bovineerythrocytes are shown in Figure 2, which reveals thecomplexity of the mixture. The ESI ion abundanceprofiles of the mass spectra of the protonated ([M 1H]1) (panel A), lithiated ([M 1 Li]1) (panel B), sodiated([M 1 Na]1) (panel C), and potassiated (data notshown) sphingomyelin molecular species from bovineerythrocytes are identical, it may therefore be possibleto use alkaline-adduct molecular ions for semiquantita-tion of sphingomyelin in the mixture [19, 20]. Thesimilarity in the mass profiles of the [M 1 H]1 speciesand the [M 1 Li]1 species was further confirmed by thebovine brain sphingomyelin (panels D and E in Figure2) and chicken egg yolk (data not shown).
Characterization of Sphingomyelins by CADTandem Mass Spectra of the Lithiated Adduct([M 1 Li]1) Ions
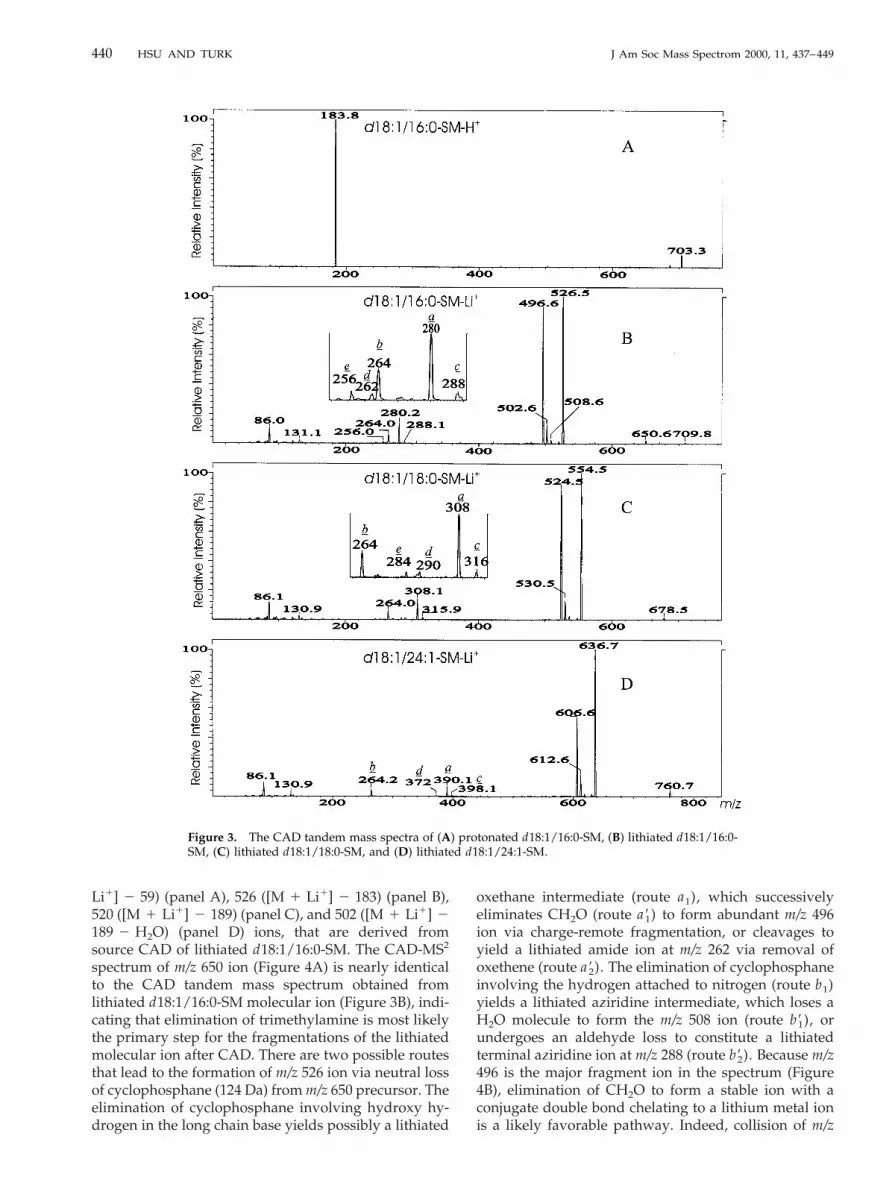
As it has been reported by FAB CAD [15, 16], the ESICAD tandem mass spectra of the [M 1 H]1 ions arisingfrom sphingomyelin and GPC yield one major fragmention at m/z 184, corresponding to protonated phospho-choline [15, 16, 21]. Ions reflecting the fatty acid constit-uents and the identities of the sphingosine base are notobserved (Figure 3A). In contrast, the product ionspectrum of the [M 1 Li]1 ions of sphingomyelin yieldsabundant structurally informative fragment ions thatare analogous to those previously reported for lithiatedGPC [21]. The product ion spectrum of the lithiated1-phosphocholine-N-palmitoyl-sphingosine (d18:1/16:0-SM) at m/z 709 yields fragment ions at m/z 650(Figure 3B), corresponding to neutral loss of trimethyl-amine ([M 1 Li]1 2 N(CH3)3) (route a), followed byneutral loss of cyclophosphane [(HO)P(O)(OCH2
CH2O)] or lithium cyclophosphanate [(LiO)P(O)(OCH2CH2O)], to form m/z 526 ([M 1 Li]1 2 183) and520 ([M 1 Li]1 2 189) ions, respectively. Ions at m/z131 and 86, representing lithiated cyclophosphane andCH2¢CH–N(CH3)3
1 ions, respectively, were also ob-served [21]. Further elimination of CH2O or H2O fromm/z 526 via charge-remote fragmentation yields ions atm/z 496 or 508, respectively. The ions at m/z 502 arisefrom a H2O loss from m/z 520, followed by neutral lossof the long chain base as a terminal conjugate diene orneutral loss of the fatty acid as a ketene to produce ionsat m/z 280 (a) and 264 (b), respectively. Because ions atm/z 264 and 280 reflect the long chain base and fattyacid in the molecule, and the intensity of the latter ionis more abundant than the former, the structure of thecompound can be easily determined. It appears thations at m/z 288 (c), 262 (d), and 256 (e) are alwayspresent in the spectrum along with ions at m/z 280. Thisprovides confirmation for the identities of the fatty acidin the molecule. A similar pattern of ions was also
Figure 1. Sphingomyelin.
438 HSU AND TURK J Am Soc Mass Spectrom 2000, 11, 437–449

observed for lithiated d18:1/18:0-SM (m/z 737, Figure3C) and lithiated d18:1/24:1-SM (m/z 819, Figure 3D).The two spectra contain abundant m/z 308 and 390 ions,representing stearic acid and tetradodecenic acid, re-spectively. The two spectra also contain ions at m/z 264,representing the d18:1 long chain base. The fragmenta-tion pathways (Scheme 1) and the structures of theabove ions were further investigated by source CAD-MS2.
The Fragmentation Pathways Confirmed by SourceCAD Tandem Mass Spectrometry (SourceCAD-MS2)
To investigate the genesis of the fragment ions in thetandem spectra, fragment ions generated by skimmerCAD were selected as the precursor ions, collisionallyactivated and analyzed by the third quadrupole. Figure4 illustrates the CAD-MS2 spectra of m/z 650 ([M 1
Figure 2. The ESI mass spectra of sphingomyelin isolated from bovine erythrocytes (panels A, B, andC), and bovine brain (panels D and E). (A) [M 1 H]1, (B) [M 1 Li]1, (C) [M 1 Na]1, (D) [M 1 H]1,and (E) [M 1 Li]1 adduct ions.
439J Am Soc Mass Spectrom 2000, 11, 437–449 STRUCTURAL DETERMINATION OF SPHINGOMYELIN
Li1] 2 59) (panel A), 526 ([M 1 Li1] 2 183) (panel B),520 ([M 1 Li1] 2 189) (panel C), and 502 ([M 1 Li1] 2189 2 H2O) (panel D) ions, that are derived fromsource CAD of lithiated d18:1/16:0-SM. The CAD-MS2
spectrum of m/z 650 ion (Figure 4A) is nearly identicalto the CAD tandem mass spectrum obtained fromlithiated d18:1/16:0-SM molecular ion (Figure 3B), indi-cating that elimination of trimethylamine is most likelythe primary step for the fragmentations of the lithiatedmolecular ion after CAD. There are two possible routesthat lead to the formation of m/z 526 ion via neutral lossof cyclophosphane (124 Da) from m/z 650 precursor. Theelimination of cyclophosphane involving hydroxy hy-drogen in the long chain base yields possibly a lithiated
oxethane intermediate (route a1), which successivelyeliminates CH2O (route a91) to form abundant m/z 496ion via charge-remote fragmentation, or cleavages toyield a lithiated amide ion at m/z 262 via removal ofoxethene (route a92). The elimination of cyclophosphaneinvolving the hydrogen attached to nitrogen (route b1)yields a lithiated aziridine intermediate, which loses aH2O molecule to form the m/z 508 ion (route b91), orundergoes an aldehyde loss to constitute a lithiatedterminal aziridine ion at m/z 288 (route b92). Because m/z496 is the major fragment ion in the spectrum (Figure4B), elimination of CH2O to form a stable ion with aconjugate double bond chelating to a lithium metal ionis a likely favorable pathway. Indeed, collision of m/z
Figure 3. The CAD tandem mass spectra of (A) protonated d18:1/16:0-SM, (B) lithiated d18:1/16:0-SM, (C) lithiated d18:1/18:0-SM, and (D) lithiated d18:1/24:1-SM.
440 HSU AND TURK J Am Soc Mass Spectrom 2000, 11, 437–449

496 ion under the same collision energy yields a spec-trum containing no fragment ions (data not shown).
Both m/z 520 and 502 ions are protonated precursors.When subjected to CAD, the tandem spectrum arisingfrom m/z 520 (Figure 4C) is nearly identical to thatobtained from m/z 502 (Figure 4D). Hence, the m/z 502ion arises from a water loss of m/z 520 (route c91) to forma protonated aziridine ion, which successively loses aketene (route c92) or a conjugate diene (route c93) to formm/z 264 and 280 ion, respectively. The formation of m/z256 ion most likely arises from the protonated oxethaneion, which yields a protonated amide ion via loss ofoxethene. This is also consistent with the earlier notionthat ion at m/z 262 in the tandem spectrum generatedfrom the lithiated counterpart precursor represents alithiated amide ion. The fragmentation pathways arefurther supported by the identification of the m/z 280ion. Figure 4E displays the source CAD-MS2 spectrumof m/z 280 ion, which contains the typical[(CH2CH¢N)CO(CH2)nCH¢CH2]1 series ions viacharge-remote fragmentation [23, 24], where n 5 0, 1, 2. . . , etc., reflecting the fatty acid chain and the remoteazirine charge site (Scheme 1). This is further confirmedby the tandem spectrum of the analogous ion at m/z 308
(data not shown) arising from source CAD of lithiatedd18:1/18:0-SM molecule. Therefore, an azirinelike inter-mediate (a9) may be formed, followed by constantneutral loss of terminal conjugate diene (route c93) toform a N-acyl-azirine ion (a). Abundant fragment ionsare observed in the m/z range from 30 to 150 in thetandem spectra of 502 and 520. A series of alkyl ions areobserved at m/z 43, 57, 71, etc., as well as unsaturatedion series at m/z 55, 69, 83, etc., at m/z 67, 81, 95, etc. andat m/z 79, 93, 107, etc. The formation of these ions ismost likely derived from charge-driven fragmentation[22, 25], while the lithiated precursors undergo charge-remote fragmentation [26].
The CAD Tandem Mass Spectra of Sodiated([M 1 Na]1) and Potassiated ([M 1 K]1)Adduct Ions
Under the same collision energy, the tandem massspectra of the [M 1 Na]1 ions of sphingomyelins alsocontain abundant fragment ions which can be used toidentify polar head group, fatty acyl chain, and longchain base in the molecule. As shown in Figure 5, the
Scheme 1
441J Am Soc Mass Spectrom 2000, 11, 437–449 STRUCTURAL DETERMINATION OF SPHINGOMYELIN

tandem mass spectrum of the m/z 725 precursor ions forsodiated d18:1/16:0-SM (panel A) yields abundant ionsat m/z 280 and 264, indicating the presence of palmitoylacyl and sphingosine base, respectively. However, ionscorresponding to sodiated aziridine (c series) and sodi-ated amide (d series) are absent. The tandem massspectrum also contains fragment ions at m/z 666 ([M 1Na]1 2 N(CH3)3), 542 ([M 1 Na]1 2 183), which arerelated to the loss of polar head group, along with ionsat m/z 524, 512, 502, 147, and 86, analogous to thoseobserved for [M 1 Li]1. The intensities of the ions at
m/z 147 (sodiated cyclophosphane) and m/z 86 becomepredominate, and Na1 appears at m/z 23. A similarspectrum is also observed for sodiated d18:1/24:0-SM(panel B). The dominance of fragment ions associatedwith alkaline metal becomes even more apparent in theproduct ion spectrum obtained from [M 1 K]1 ion(panel C), where ions at m/z 163 (potassiated cyclophos-phane), and m/z 39 (K1) are among the most abundant.These results demonstrate that the energy requirementfor release of alkaline metal ions is in this order K1 ,Na1 , Li1. Charge-remote fragmentation and loss of
Figure 4. The source CAD tandem mass spectra of (A) m/z 650, (B) m/z 526, (C) m/z 520, (D) m/z 502,and (E) m/z 280 ions generated by skimmer CAD of lithiated d18:1/16:0-SM molecule.
442 HSU AND TURK J Am Soc Mass Spectrom 2000, 11, 437–449

alkaline metal ion appear to be the two competingprocesses at both low energy CAD and high energyCAD [26]. Therefore, when formation of alkaline metalions becomes the major fragmentation pathway, struc-turally informative ions arising from charge-remotefragmentation become less abundant. This is consistentwith the notion that c and d series ions are absent in the
tandem spectra obtained with sodiated and potassiatedmolecular species. As described earlier, loss of trimethylamine is the primary step leading to the fragmentationfor [M 1 Li]1. The source CAD-MS2 spectrum of m/z666 (panel D), arising from sodiated d18:1/16:0-SM (m/z725) yields a spectrum similar to that obtained from m/z725 (panel A), but ions at m/z 147 become the most
Figure 5. The CAD tandem mass spectra of (A) sodiated d18:1/16:0-SM, (B) sodiated d18:1/24:0-SM,(C) potassiated d18:1/16:0-SM, and (D) source CAD-MS2 of m/z 666 ion produced from sodiatedd18:1/16:0-SM.
443J Am Soc Mass Spectrom 2000, 11, 437–449 STRUCTURAL DETERMINATION OF SPHINGOMYELIN

abundant and ions at m/z 86 is not observed (the m/z 86ion is also absent in Figure 4A). This is consistent withthe proposed structure for m/z 86, representing aCH2¢CH–N(CH3)3
1 ion which would not be present inthe CAD-MS2 spectra of the precursors where thetrimethylamine group had been lost (i.e., m/z 666 ion).For both CAD MS/MS and source CAD-MS2, the de-compositions of alkali-metal-cationized precursors pro-
gressively become dominated by release of metal cat-ions as lithium is replaced by sodium or potassiummetal ions. The competition between direct loss of themetal ion and remote decomposition leading to cation-ized fragment ions became so apparent that there arelittle discernible organic fragments and therefore struc-turally informative ions become less available for [M 1Na]1 and least available for [M 1 K]1 ion species.
Figure 6. The CAD tandem mass spectra of the sphingomyelin mixture of bovine erythrocytesobtained by (A) CNL scanning of 59, (B) CNL scanning of 183, (C) CNL scanning of 207, (D) CNLscanning of 213, and (E) precursor ion scanning of 184. Tandem spectra obtained from the above scansillustrate the profile of sphingomyelin phospholipid in the mixture.
444 HSU AND TURK J Am Soc Mass Spectrom 2000, 11, 437–449

Identification of Sphingomyelin Class via ConstantNeutral (CNL) Loss Scan
Constant neutral loss scanning has been previouslyreported for identification of GPC class in biologicalextract [21, 27]. Both GPC lipids and sphingomyelins
possess phosphocholine as polar head group. In theCAD tandem mass spectra, the major fragment ionsarising from CNL involve the participation of polarhead group [21] and are among the most importantfragmentation pathways for both GPC and sphingomy-elin. Figure 6 shows the tandem spectra of sphingomy-
Figure 7. The CAD tandem mass spectra of the sphingomyelin mixture of bovine erythrocytes(panels A, B, and C) and of chicken egg yolk (panels D and E). (A) CNL scanning of 429, (B) precursorion scanning of 264, (C) CNL scanning of 427, (D) CNL scanning of 213, and (E) CNL scanning of 429.CNL scanning of 429 and precursor ion scanning of m/z 264 classify sphingosine subclass. CNLscanning of 427 classify dehydrosphingosine subclass and CNL scanning of 213 classify sphingomy-elin phospholipid.
445J Am Soc Mass Spectrom 2000, 11, 437–449 STRUCTURAL DETERMINATION OF SPHINGOMYELIN

elin from bovine erythrocyte obtained from a CNL scanof 59 (panel A), 183 (panel B), 207 (panel C), or 213(panel D). The mass profiles obtained from these lossesare similar to its ESI mass spectrum (Figure 2B) and theparent ion spectrum of the protonated sphingomyelinobtained by scanning of m/z 184 (panel F). However, thespectra have been significantly simplified thus, facilitat-ing the identification of the sphingomyelin class in themixture. As discussed earlier, fragment ions at m/z 264(M 1 Li1 2 429) is characteristic of the sphingosinelong chain base (d18:1) subclass, which represents acombined neutral losses of lithium cyclophosphanate,H2O and sphingosine as a conjugated diene from [M 1Li1]. Therefore, identification of sphingomyelin mix-ture with the sphingosine long chain base subclass canalso be achieved by CNL scanning of 429. As shown inFigure 7, the mass profile of the CNL spectrum of 429(panel A) is equivalent to the parent ion spectrum of m/z264 (panel B), which clearly shows the class of 1-phos-phocholine-N-acyl-sphingosine molecules in the mix-ture. Sphingomyelins with d18:2 (dehydrosphingosine)or d18:0 (dihydrosphingosine) LCB in the mixture werefurther identified by CNL scanning of 427 (panel C) or431 (not shown), respectively. The CAD tandem spec-trum obtained from CNL scan of 427 yields a spectrumcontaining ions at m/z 735, 791, 817, 819, 845, and 847.This is consistent with the results obtained by CAD
product ion spectra of individual molecular species(Table 1). CNL scanning of 213 and 429 performed forchicken egg yolk (Figure 6D, E) and bovine brain (datanot shown) samples easily reveal sphingomyelin andidentify the sphingomyelin with sphingosine LCB in themixture.
When both Q1 and Q3 quadrupoles are operated atunit resolution for a CNL scanning, a tandem spectrumwith unit resolution can be obtained. As shown inFigure 6, tandem spectra obtained for bovine erythro-cyte by CNL scanning are all of unit resolution, but theparent ion profile of the protonated sphingomyelinobtained by scanning of 184 (panel E) is less resolved.At a collision energy of 52 eV, both the ion abundanceand mass profile of the tandem spectrum of the mixturearising from CNL loss of 213 (Figure 6D) is nearlyidentical to its ESI mass spectrum (Figure 2B). Thetandem spectrum arising from CNL of 183 at a collisionof 40 eV (Figure 6B) is also similar. However, thebackground ions of the two spectra have been signifi-cantly reduced and unit resolution retained. Becausethe ion intensities of both [M 1 Li1 2 213] and [M 1Li1 2 183] ions are the two most abundant under theapplied collision energy, spectra obtained by theselosses not only permit structural determination as de-scribed above, but also may provide a sensitive methodfor quantitation of the complex mixture. It appears that
Table 1. LCB-FA compositions in sphingomyelin as obtained by ESI CAD tandem mass spectrometry from bovine brain, bovineerythrocytes, and chicken egg yolk
Bovine brain Bovine erythrocyte Chicken egg yolk
MLi1Fattyacid LCB
Relativeabundance
(%) MLi1Fattyacid LCB
Relativeabundance
(%) MLi1Fattyacid LCB
Relativeabundance
(%)
681 14:0 18:1 ,1 681 14:0 18:1 2 681 14:0 18:1 1695 15:0 18:1 ,1 16:0 16:1 709 16:0 18:1 100709 16:0 18:1 13 695 16:0 17:1 4 723 17:0 18:1 ,1
18:0 16:1 709 16:0 18:1 45 737 18:0 18:1 9723 18:0 17:1 1 711 16:0 18:0 15 751 19:0 18:1 ,1735 18:0 18:2 6 723 17:0 18:1 5 765 20:0 18:1 1737 18:0 18:1 74 16:0 19:1 791 22:1 18:1 1751 19:0 18:1 1 18:0 17:1 793 22:0 18:1 3765 18:0 20:1 6 735 18:0 18:2 1 807 23:0 18:1 ,1
20:0 18:1 737 18:0 18:1 10 817 24:2 18:1 1791 22:1 18:1 8 765 20:0 18:1 2 819 24:1 18:1 3
20:1 20:1 791 22:0 18:2 10 821 24:0 18:1 2793 22:0 18:1 8 793 22:0 18:1 45805 23:1 18:1 6 805 23:0 18:2 3807 23:0 18:1 8 807 23:0 18:1 10819 24:1 18:1 100 817 24:1 18:2 11821 24:0 18:1 28 24:2 18:1833 25:1 18:1 11 819 24:0 18:2 78835 25:0 18:1 10 24:1 18:1847 26:1 18:1 7 821 24:0 18:1 100
83325:1 18:1
525:0 18:2
835 25:0 18:1 10845 26:1 18:2 4
84726:0 18:2
826:1 18:1
446 HSU AND TURK J Am Soc Mass Spectrom 2000, 11, 437–449

collision energy governs the intensities of the ions in thespectrum. Further studies in the application of CNLlosses to the quantitation of sphingomyelin mixture areunder investigation.
Identification of Molecular Structures inSphingomyelin Mixtures
While the mass profiles of the alkaline-metal adduct ofthe mixture provide the information on the molecularspecies, the CAD tandem mass spectra can be used toidentify individual sphingomyelin in the mixture. Thecompositions of the sphingomyelin mixture from bo-vine brain, chicken egg, and bovine erythrocyte, asidentified by CAD tandem mass spectrum of individual[M 1 Li]1 ion, are listed in Table 1. For bovine brain,bovine erythrocyte, and chicken egg yolk sphingomy-elins, 20, 26, and 12 molecules were identified, respec-tively, close to those reported by Karlsson et al. [18].However, discrepancies in molecular species were ob-served. For example, in bovine brain, 19:1-LCB was not
observed by us and others, but was reported by Karls-son et al.; ion at m/z 735 was identified as d18:2/18:0-SMby our method but this species was not observed byothers [11, 13, 17, 18]. These discrepancies in the com-position of minor components may arise from limit ofdetection by various methods and variation in thesamples. There are two isomers in the molecular speciesof m/z 709, 765, and 791 of the bovine brain sample. TheCAD product ion spectrum of m/z 709 (Figure 8A)clearly shows ions at m/z 280 and 308, reflecting 16:0and 18:0 fatty acid respectively, and ions at m/z 264 and236, representing 18:1 and 16:1 long chain base, respec-tively. Because the m/z 264/280 ion pair is observed ford18:1/16:0-SM (Figure 3B), the 236/308 ion pair can beeasily assigned as d16:1/18:0-SM. The structure is fur-ther confirmed by the source CAD-MS2 spectrum of them/z 502 ion (Figure 8B), which yields m/z 264/280 and236/308 ion pairs, reflecting d18:1/16:0-SM and d16:1/18:0-SM isomers, respectively. In bovine erythrocyte, atleast two isomers were observed for the molecular ionsat m/z 681, 723, 817, 819, and 847. The identification of
Figure 8. The CAD product ion mass spectrum of the lithiated sphingomyelin molecule of (A) m/z709 ions from bovine brain and (B) source CAD tandem mass spectrum of m/z 502 ions generated fromm/z 709. (C) The CAD product ion mass spectrum of the lithiated molecular species of m/z 819 frombovine erythrocytes. The tandem spectra clearly show two isomers for each mass.
447J Am Soc Mass Spectrom 2000, 11, 437–449 STRUCTURAL DETERMINATION OF SPHINGOMYELIN

isomeric composition of these molecules is exemplifiedby the CAD tandem spectrum of the m/z 819 ions(Figure 8C). The spectrum contains m/z 262/392 and264/390 ion pairs, representing the d18:2/24:0-SM andd18:1/24:1-SM isomers. The c-series ions at m/z 398 and400, d-series ions at m/z 372 and 374 and e-series ions atm/z 368 and 366 are also present in the spectrum thatfurther confirm the identities of the fatty acid constitu-ents. As compared to the m/z 264/390 ion pair observedfor lithiated d18:1/24:1-SM standard (Figure 3D), theisomer with a d18:2/24:0-SM structure can be easilydetermined. The major LCB constituents for the mixtureare 18:1 and 18:2 and whole array of fatty acid from 14:0to 26:0 were detected. The chicken egg yolk sphingo-myelin is uniformly composed of 18:1 LCB with 16:0and 18:0 as the major fatty acid constituents. This isconsistent with the results obtained from CNL scanningof 429 (Figure 7D), reflecting the sphingosine subclassin the mixture.
Conclusions
Electrospray ionization in combination with low energyCAD tandem mass spectrometry using product ion,precursor ion, and neutral loss scans provides a quickand reliable method for structural determination, andprofiling the sphingomyelin content and distribution inthe mixture. Because samples of biological origin maycontain impurities that may cause ion suppression,separation of sphingomyelin class by HPLC prior todirect infusion in combination with the above describedtandem mass spectrometry with CNL scanning mayhave great potential in the simultaneous quantificationand qualification of sphingomyelin mixtures.
AcknowledgmentsThis research was supported by US Public Health Service GrantsP41-RR-00954, R37-DK-34388, P60-DK-20579, and P01-HL-57-278and a grant (No. 996003) from the Juvenile Diabetes Foundation.
References1. Sweeley, C. C. Purification and partial characterization of
sphingomyelin from human plasma. J. Lipid Res. 1963, 4,402–406.
2. Osamu, M.; Tsubono, H.; Akiyama, M.; Sakagami, T. Sphin-gomyelins in human erythrocytes and plasma. J. Biochem.(Tokyo) 1967, 62, 618–620.
3. Samuelsson, K. On the occurrence and nature of free cer-amides in human plasma. Biochim. Biophys. Acta 1969, 176,211–213.
4. Lastennet, A.; Freysz, L.; Bieth, R. Distribution and metabo-lism of 2 types of sphingomyelins in various zones of thecentral nervous system. J. Physiol. (Paris) 1970, 62, 289.
5. Hannun, Y. A. Sphingolipid second messengers: tumor sup-pressor lipids. Adv. Exp. Med. Biol. 1997, 400A, 305–312.
6. Hannun, Y. A.; Obeid, L. M.; Wolff, R. A. The novel secondmessenger ceramide: identification, mechanism of action, andcellular activity. Adv. Lipid Res. 1993, 25, 43–64.
7. Chao, M. V. Ceramide: a potential second messenger in thenervous system. Mol. Cell Neurosci. 1995, 6, 91–96.
8. Morrison, W. R.; Hay, J. D. Polar lipids in bovine milk. II.Long-chain bases, normal and 2-hydroxy fatty acids, andisomeric cis and trans monoenoic fatty acids in the sphingo-lipids. Biochim. Biophys. Acta 1969, 202, 460–467.
9. Nyberg, L. In Phospholipids: Characterization, Metabolism andNovel Biological Applications; Cevc, G.; Paltauf, F., Eds.; AOCS:Champaign, IL, 1995; p 125.
10. Minami, H.; Nylander, T; Carlsson, A; Larsson, K. Incorpora-tion of proteins in sphingomyelin-water gel phases. Chem.Phys. Lipids 1996, 79, 65–70.
11. Jungalwala, F. B.; Evans, J. E.; McCluer, R. H. Composi-tional and molecular species analysis of phospholipids byhigh performance liquid chromatography coupled withchemical ionization mass spectrometry. J. Lipid Res. 1984,25, 738 –749.
12. Kim, H. Y.; Salem, N. Jr. Application of thermospray high-performance liquid chromatography/mass spectrometry forthe determination of phospholipids and related compounds.Anal. Chem. 1987, 59, 722–726.
13. Valeur, A.; Olsson, N. U.; Kaufmann, P.; Wada, S.; Kroon,C.-G.; Westerdahl, G; Odham, G. Quantification and com-parison of some natural sphingomyelins by on-line high-performance liquid chromatography/discharge-assistedthermospray mass spectrometry. Biol. Mass Spectrom. 1994,23, 313–319.
14. Byrdwell, W. C.; Borchman, D. Liquid chromatography/mass-spectrometric characterization of sphingomyelin and dihy-drosphingomyelin of human lens membranes. Ophtahl. Res.29, 191, 1997.
15. Murphy, R. C.; Harrison, K. A. Fast atom bombardment massspectrometry of phospholipids. Mass Spectrom. Rev. 1994, 13,57–75.
16. Ann, Q.; Adams, J. Collision-induced decomposition of sphin-gomyelins for structural elucidation. Biol. Mass Spectrom. 1993,22, 285–294.
17. Kerwin, J. L.; Tuininga, A. R.; Ericsson, L. H. Identification ofmolecular species of glycerophospholipids and sphingomye-lin using electrospray mass spectrometry. J. Lipid Res. 1994, 35,1102–1114.
18. Karlsson, A. A.; Michelsen, P.; Odham, G. Molecular species ofsphingomyelin: determination by high-performance liquidchromatography/mass spectrometry with electrospray andhigh-performance liquid chromatography/tandem mass spec-trometry with atmospheric pressure chemical ionization. J.Mass Spectrom. 1998, 33, 1192–1198.
19. Han, X.; Gross, R. W. Electrospray ionization mass spectro-scopic analysis of human erythrocyte plasma membrane phos-pholipids. Proc. Natl. Acad. Sci. USA 1994, 91, 10635–10639.
20. Kim, H. Y.; Wang, T.-C. L.; Ma, Y.-C. Liquid chromatography/mass spectrometry of phospholipids using electrospray ion-ization. Anal. Chem. 1994, 66, 3977–3982.
21. Hsu, F. F.; Bohrer, A.; Turk, J. Formation of lithiated adductsof glycerophosphocholine lipids facilitates their identificationby electrospray ionization tandem mass spectrometry. J. Am.Soc. Mass Spectrom. 1998, 9, 516–526.
22. Hsu, F. F.; Turk, J. Structural characterization of triacylglycer-ols as lithiated adducts by electrospray ionization mass spec-trometry using low-energy collisionally activated dissociationon a triple stage quadrupole instrument. J. Am. Soc. MassSpectrom. 1999, 10, 587–599.
23. Gross, M. L. Charge remote fragmentations: methods, mech-anism, and applications. Int. Mass Spectrom. Ion Processes 1992,118/119, 137–158.
24. Vetter, W.; Meister, W.; Oeaterhelt, G. 2-alkylimidazolinederivative to control fatty acid fragmentation upon electron
448 HSU AND TURK J Am Soc Mass Spectrom 2000, 11, 437–449

impact and electrospray ionization. J. Mass Spectrom. 1998, 33,461–472.
25. Murphy, R. C. In Handbook of lipid research; Plenum: New York,1993; p 190.
26. Adams, J.; Gross, M. L. Energy requirements for remotecharge site decompositions and structural information from
collisional activation of alkali metal cationized fatty alcohols.J. Am. Chem. Soc. 1988, 108, 6915–6921.
27. Domingues, P.; Amado, F. M. L.; Marques, M. G. O. S.;Ferrer-Corrcia, A. J. Constant neutral loss scanning for thecharacterization of glycerol phosphatidylcholine phospholip-ids. J. Am. Soc. Mass Spectrom. 1998, 9, 1189–1195.
449J Am Soc Mass Spectrom 2000, 11, 437–449 STRUCTURAL DETERMINATION OF SPHINGOMYELIN





























