Embed Size (px)
Citation preview

Exp Brain Res (1992) 88:213-218
E erimental Brain Research © Springer-Vertag t992
Synthesis and secretion of 2-macroglobulin by human glioma established cell lines R. Businaro 1, C. Fabrizi 2, L. Fumagalli 1, and G.M. Lauro 2
1 Dipartimento di Scienze Cardiovascolari e Respiratorie, and 2 Dipartimento di Biologia Cellulare e dello Sviluppo, Universitfi "La Sapienza", Via degli Apuli n ° 1, 1-00185 Roma, Italy
Received March 4, 1991 / Accepted August 15, 1991
Summary. Human a2-macroglobulin (a2M) is a high molecular weight plasma proteinase inhibitor exhibiting a broad specificity; in fact it is capable of binding en- dopeptidases from all known classes of proteases (Barret 1981). Two human glioma cell lines, namely an as- trocytoma and a glioblastoma, were found to synthesize and secrete in the culture medium a protein which re- sembles the serum a2M for immunological, biochemical and biological features. Using polyclonal antibodies to serum (~2 M, an a2M-like factor could be detected in the cytoplasm and in the culture medium of the tumor cells. Furthermore this factor accumulated in cytoplasmic granules if cells were incubated with monensin and its production was dramatically reduced following a treat- ment with cycloheximide. This protein behaved like the serum o~2M in immunoblotting analysis and exhibited the same antiproteolytic activity. Its role in human brain is unknown at present. Since interactions of proteinases and proteinase-inhibitors appear to influence the host- tumor immune response and to play a crucial role during the migration of metastasizing tumor cells, a2M ex- pression observed in these glioma cells could be involved in tumor cell proliferation and invasion.
Key words: Gtial cultures - Gliomas Tumor cells - Proteinase inhibitors - a2-macroglobulin - Human
Introduction
Increased levels of proteinase inhibitors were recently found in neoplastic tissues and sera of tumor-bearing patients (Harris et al. 1974; Cooper et al. 1976; Matsuda et al. 1983; Okumichi et al. 1984; Cheung and Lau 1986; Chawla et al. 1987; Sawaya et al. 1987). It is well known that the metastatic capacity may be influenced by the proteolytic activity present in the extracellular environ-
Offprint requests to: G.M. Lauro
ment, since many aspects of mitogenesis, migration and cell movement are controlled by proteases. It has been also shown that oncogene activation, which induces cell proliferation and malignant transformation, leads to the proteinase gene expression (Tryggvason et al. 1987). Fur- thermore interactions of proteinase and proteinase- inhibitors appear to play a crucial role during brain development, in cell migration (Guenther et al. 1985; Lindner et al. 1986) and neurite outgrowth (Pittman 1985; Hawkins and Seeds 1986; Monard 1988). In this connection, recent experiments by Mori et al. showed that ~2M derived from newborn rat astroglial cells in- duce neurite elongation in cultured neurons from rat central nervous system (Mori et al. 1990).
In the present study we decided to evaluate the presence of proteinase inhibitors in two human glioma established cell lines, namely an astrocytoma and a glio- blastoma. Occurrence of some of these inhibitors, such as c~2-macroglobulin (~2M), in human brain has been already reported (Dziegelewska et al. 1986) and its presence has been demonstrated immunohistochemically in human cerebral gliomas (Seitz and Wechster 1987). Its synthesis was observed in vitro in astrocyte primary cultures from newborn rats (Gebicke-Haerter et al. 1987). Moreover, several human melanoma and sarcoma cell lines synthesize and secrete ~zM (Saksela et al. 1984; Bizik et al. 1986; Grofova et al. 1988; Lizonova et at. 1990) and recently a glioblastoma multiforme was found to produce ~zM in an organ culture system (Keohane et al. t990).
Material and methods
Cell lines
Two human glioma cell lines, obtained from explants of a III WHO gemistocytic astrocytoma (T67; 27th-30th passage in culture) and from a glioblastoma (T70; 23rd-26th passage), were grown routine- ly in monolayers within 25-cm 2 flasks (Nunc, Denmark) containing HAM-F10 (Gibco, Paisley, UK) supplemented with 10% fetal calf

214
serum (FCS; Gibco, Paisley, UK) and gentamycin (40 gg/mt; Haz- leton, USA). Cells were cultured in a fully humidified atmosphere of 5 % CO2 in air and passaged with 0.05 % trypsin and 0.02 % EDTA (Flow, Irvine, UK) at a split ratio of 1 : 2.
For all experiments cells were washed twice with phosphate buffered saline (PBS) and then placed in serum free medium (SFM) for at least three days.
Both lines (T67, T70) were characterized by means of polyclonal and monoclonal antibodies directed against glial fibrillar acidic protein (GFAP), S-100 protein, fibronectin, factor VIII and vimen- tin (Dakopatts, Denmark) as previously described (Lauro et al. 1986; Cusimano et al. 1990).
Proteinase~inhibitors detection by immunofluorescence
T67 and T70 cells were grown on coverslips for five days in SFM, washed extensively in PBS, fixed 10 rain in acetone at - 2 0 ° C, washed again and then incubated overnight at 4 ° C in a moist chamber with the various rabbit polyclonal antibodies directed against: ctzM, 1 : 200; inter-c~-trypsin inhibitor, 1 : 200; a 1-antitryp- sin, 1 : 200; ctx-antichymotrypsin, 1:200 (Dakopatts, Denmark). Then they were treated for 1 h at room temperature with a mouse monoclonal anti-GFAP antibody diluted 1 : 10 (Boehringer Mann- heim, FRG). After removing the specific antiserum and several washings with PBS, the cells were treated, first with a rhodamine conjugated anti-rabbit IgG F(ab')2 antibody (30 min; diluted 1:50), and then with a ftuorescein conjugated anti-mouse IgG F(ab')2 antibody (30 min; diluted 1:50), both obtained from goat (Cappel, Cochranville, PA, USA). The cells were finally washed with PBS, mounted on glycerol-PBS and observed with a Leitz Dialux 20 fluorescence microscope under the appropriate wavelengths.
Monensin treatment
T67 and T70 cells, grown on coverslips, were maintained for five days in SFM, then replaced for 2 h with fresh SFM containing 10 gM monensin (Sigma, St.Louis, MO, USA); finally, they were washed in PBS, fixed and immunohistochemically processed as previously described.
Isolation and purification of c~zM
T67 cells were grown to confluency (85 cm 2 flasks; Nunc, Denmark) in F10, 10% FCS and antibiotic. After an incubation for 72 h in SFM, they were washed twice in PBS in order to remove residual serum proteins adsorbed to the cell layers and replaced with fresh SFM (20 ml per flask). After 72 h the culture medium was collected, passed through a 0.22 pm filter to remove cellular debris and stored at - 20 ° C.
Five hundred ml of culture medium were applied to an affinity cromatography column packed with Sepharose-4B by the method of Parikh et al. (1974). The Sepharose was activated with cyanogen bromide and then coupled to a rabbit antibody directed against azM (Dakopatts, Denmark). Bound protein was eluted with a linear gradient of 0 to 4.5 M MgC12. Fractions (2 ml) were collected and measured for opticat absorbance at 280 nm. The protein peaks were pooled and dialyzed against distilled water. The same procedure was used for purification of tIzM from human serum obtained from healthy donors. The concentration of azM was determined based on A2so nm 1% =9.0 (Barret 1981).
SDS-PAGE and Western blotting
a2M purified from glioma culture medium or from human serum was run in sodium dodecyl sulfate-polyacrylamide gel electro- phoresis (SDS-PAGE) according to Laemmli (1970) using 7.5%
resolving gels and 3 % stacking gels. Proteins were visualized using a silver stain procedure (Schoenle et al. 1984) or transferred to nitrocellulose membrane (Bio-Rad, CA, USA) for immunoblotting (Towbin et al. 1979). The membrane was incubated first with rabbit antibodies anti-azM (Dakopatts, Denmark) diluted 1:100, then with donkey biotinylated anti-rabbit Ig (Amersham, Bucks, UK) and finally with streptavidin-biotinylated horseradish peroxidase complex (Amersham, Bucks, UK). The bands were visualized using 4-chloro-l-naphthol (Sigma, St. Louis, Mo, USA) as substrate.
ELISA
Enzyme-linked immunosorbent assay (ELISA) was performed es- sentially as described by Engvall et aL (1971). Briefly, 96-well ELISA plates (Nunc, Denmark) were incubated with: a) a2M puri- fied from human serum, used as a positive control; b) medium obtained from cultures maintained in SFM for 72 h, as previously described. After incubation overnight at 4 ° C, plates were coated with 1% BSA in PBS, treated, first with polyclonal rabbit antibodies directed against ctzM, ch-antitrypsin or ctl-antichymotrypsin, then with donkey biotinylated anti-rabbit Ig (Amersham, Bucks, UK) and finally with streptavidin-biotinylated horseradish peroxidase complex (Amersham, Bucks, UK). A solution containing 0.04% o-phenylenediamine (Sigma, St.I,ouis, Mo, USA), 0.1 M citric acid, 0.2 M NazHPO4, and 0.012% H202 was used as substrate. The absorbance was read at 492 nm on an automated ELO 310 ELISA reader (Biotech, Cambridge, Ma). All plates were blanked against antigen coated wells which were incubateA with diluent only or rabbit normal IgG.
The unknown concentration of azM in the samples was deter- mined using a standard curve obtained with scalar dilutions of sermn ctzM as described by Kramer and Justus (1988).
Cycloheximide treatment
Cells were cultured in SFM for 72 h; then the culture medium was discarded and the cells incubated with cycloheximide (50 gg/ml; Sigma, St. Louis, Mo, USA) diluted in fresh SFM. In the controls, only SFM was added. After 3 or 6 h, medium was aspirated from the cultures, cells briefly trypsinized, diluted in PBS, and immediate- ly counted. Prior to counting, the cells were examined by light microscopy for morphological appearance and viability. As assess- ed by morphological criteria and dye (Trypan blue) exclusion, attached cells were > 95% viable.
Antbtrypsin activity assay
Since both 1:1 and 2:1 proteinase-uzM complexes can be formed, a double molar ratio trypsin-a2M was used to obtain a complete saturation of u2M. Tripsyn-ct/M complex was prepared by mixing 1 tool of uzM with 2 tool of trypsin (Sigma, St. Louis, Mo, USA) at 37 ° C for 30 rain. All proteins were dissolved in 0.1 M Tris HC1 pH 7.5, 2 mM CaCI2. l'*C-methylated casein (Sigma, St. Louis, Mo, USA) was added as substrate (0.08 ~g in 20 ~tl of 10 mM PBS) and incubated 30 rain at 37 ° C. The reaction was stopped at 4 ° C, the solution precipitated with thrycloracetic acid (20% w/v) and then centrifuged at 6000 g for 15 rain. The radioactivity present in the supernatant was determined using a liquid scintillation counter (Packard 360 C). The percentage of inhibition was calculated com- paring the values obtained in the presence of trypsin alone with those measured in the presence of the trypsin-azM complex.
Results
Cytological markers
Both T67 and T70 cell lines were positive for G F A P (which is a specific marker for astroglial cells), S-100 prote in and v iment in (which can be expressed by astro-

215
Fig. 1A, B. Double-immunofluorescence staining of T67 cells for GFAP A, and for ~2M B showing their colocalization in the same cells. Bar indicates 15 pm
Fig. 2. Immunocytochemicat detection of c~zM in T67 cells treated with monensin (10 gM) for 2 h: the positivity observed is granular and exclusively perinuclear. Bar indicates 10 lain
gtial cells); no positivity was detected for fibronectin (a typical marker for fibroblasts) and factor VIII (a specific marker of endothelial cells).
Gtioma derived proteinase inhibitors
Cells treated with anti-a2M antibodies showed in indirect immunofluorescence a strong positivity occurring in about 80% of the cells. Figure 1 shows the colocalization of GFAP (Fig. la) and ~2M (Fig. lb) in T67 cells; in particular, a2M related immunoreactivity was peculiarly granular and occurred in the perinuclear region and some elongations. When monensin was used the reaction was stronger and exclusively perinuclear (Fig. 2). Very similar findings, but of weaker intensity, were obtained using the T70 cell line. In both lines, no positivity was observed following incubation with antibodies directed against: inter-a-trypsin inhibitor, al-antitrypsin and al-anti- chymotrypsin.
Furthermore c~zM was detected in the culture medium by ELISA and its secretion could be prevented using an inhibitor of protein synthesis, cycloheximide (Fig. 3 a, b). Figure 4 depicts the a2M secretion during four days of culture: most of it occurred during the first 24 h, while only a little increment was observed in the following days (from 2nd to 4th). Similar to immunocytochemical data, T67 (astrocytoma derived cell line) seemed to produce larger amounts of this protein compared with T70 (glio- blastoma derived cell line), maintained in the same cul- ture conditions (Fig. 4). In particular, a2M detected in T67 cell line culture medium was about 5-fold compared with T70 cell line.
The culture supernatants assayed for the presence of other plasma proteinase-inhibitors were found to be negative for eq-antitrypsin and ch-antichymotrypsin.
The a2M-like molecule purified from T67 culture medium by affinity cromatography was analyzed by SDS-polyacrylamide gel electrophoresis and compared with a2M obtained from human serum. In both cases, only one prominent band of apparent molecular weight
1200-
1000-
~. 800-
600-
& 400-
200-
O-
1 2 0 0 I
[XX] cycloheximide 1
5 6
A T67 cells
E
c
[
1000-
800-
600-
400-
200-
0- 0
[XZ] cycloheximide [3
m control T70 cells
3 Time (hours)
Fig. 3A, B. Cycloheximide blocking effect on c~2M release by T67 (A) and TT0 (B) cells during serum-free maintenance. Cyclohexi- mide (50 gg/ml) was added to cells for a period of 3 or 6 hours. Each point refers to 5 x 105 cells and represents the mean value:ks.& from two flasks
of 180 Kd could be observed. For serum a2M some minor bands, probably representing degradation products (Barret 1981), were also present. The im- munoblotting analysis using anti-azM antibodies con- firmed these results (Fig. 5).
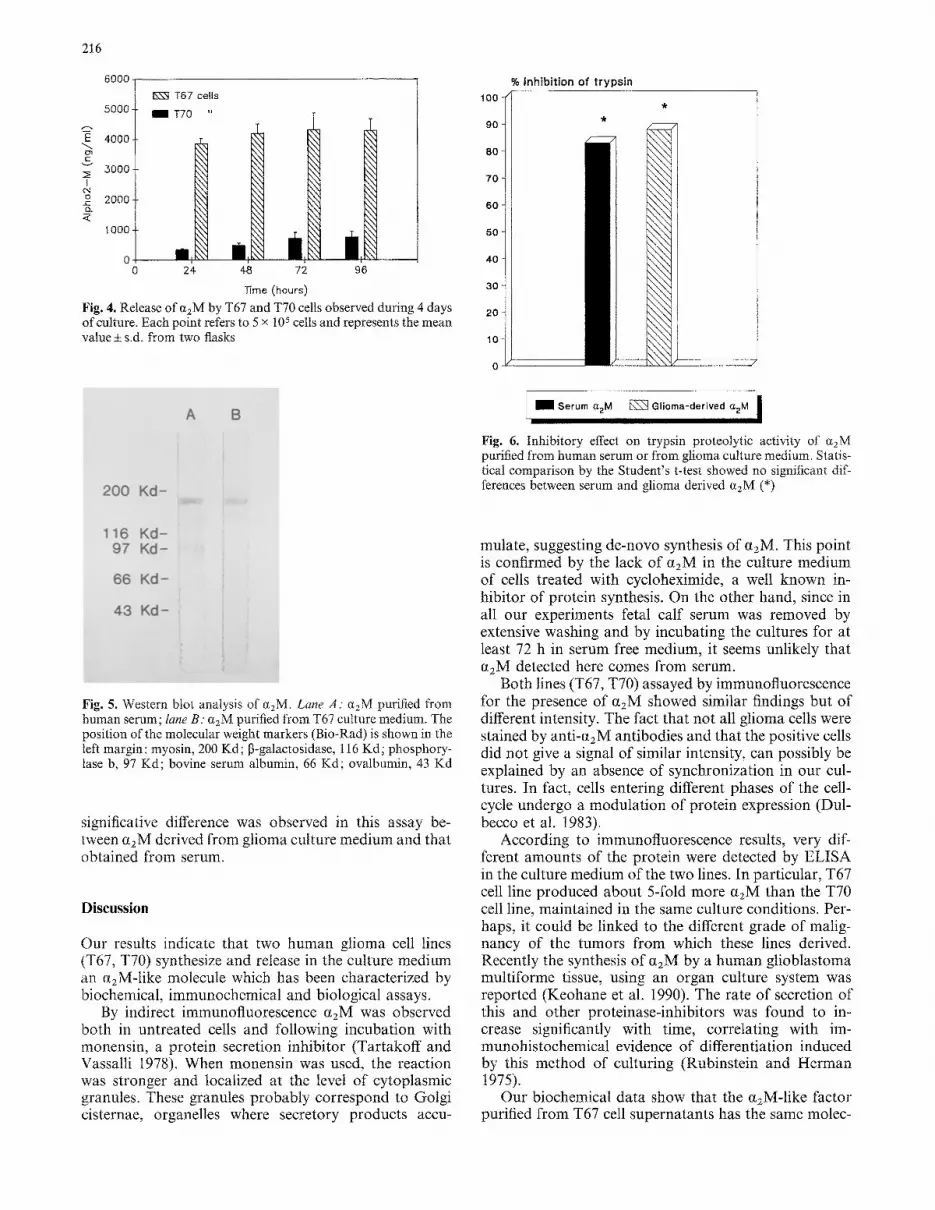
Successively, the anti-trypsin activity of the tumor derived a2M-like molecule was determined: it decreased the proteolytic effect of trypsin by 85% (Fig. 6). No
216
E
I t3
,<
6000-
5000.
4000"
3000.
2000.
1000"
O' 0
I ~ T67 cells
1 T70
• , , I I l l , 24 48 72 96
Time (hours)
Fig. 4. Release of a2M by T67 and T70 cells observed during 4 days of culture. Each point refers to 5 x 105 cells and represents the mean value :k s.d. from two flasks
A B
200 Kd-
100 -
90-
8 0
70-
60-
50-
4 0
30.
2 0
1 0
o-
% Inhibit ion of t rypsin
-k -k
1 Serum a2M [ ~ Glioma-derived a2M i
Fig. 6. Inhibitory effect on trypsin proteolytic activity of c~2M purified from human serum or from glioma culture medium. Statis- tical comparison by the Student's t-test showed no significant dif- ferences between serum and glioma derived ~zM (*)
116 Kd- 97 Kd-
66 K d -
43 Kd -
Fig. 5. Western blot analysis of ct2M. Lane A : c~2M purified from human serum; lane B: % M purified from T67 culture medium. The position of the molecular weight markers (Bio-Rad) is shown in the left margin: myosin, 200 Kd; [3-galactosidase, 116 Kd; phosphory- lase b, 97 Kd; bovine serum albumin, 66 Kd; ovalbumin, 43 Kd
significative difference was observed in this assay be- tween ~2M derived from glioma culture medium and that obtained from serum.
Discussion
Our results indicate that two human glioma cell lines (T67, TT0) synthesize and release in the culture medium an a2M-like molecule which has been characterized by biochemical, immunochemical and biological assays.
By indirect immunofluorescence a2M was observed both in untreated cells and following incubation with monensin, a protein secretion inhibitor (Tartakoff and Vassalli 1978). When monensin was used, the reaction was stronger and localized at the level of cytoplasmic granules. These granules probably correspond to Golgi cisternae, organelles where secretory products accu-
mulate, suggesting de-novo synthesis of Gt2M. This point is confirmed by the lack of a2M in the culture medium of cells treated with cycloheximide, a well known in- hibitor of protein synthesis. On the other hand, since in all our experiments fetal calf serum was removed by extensive washing and by incubating the cultures for at least 72 h in serum free medium, it seems unlikely that ~2M detected here comes from serum.
Both lines (T67, T70) assayed by immunofluorescence for the presence of a2M showed similar findings but of different intensity. The fact that not all gtioma cells were stained by anti-a2M antibodies and that the positive cells did not give a signal of similar intensity, can possibly be explained by an absence of synchronization in our cul- tures. In fact, cells entering different phases of the cell- cycle undergo a modulation of protein expression (Dul- becco et al. 1983).
According to immunofluorescence results, very dif- ferent amounts of the protein were detected by ELISA in the culture medium of the two lines. In particular, T67 cell line produced about 5-fold more a2M than the T70 cell line, maintained in the same culture conditions. Per- haps, it could be linked to the different grade of malig- nancy of the tumors from which these lines derived. Recently the synthesis of a2M by a human glioblastoma multiforme tissue, using an organ culture system was reported (Keohane et al. 1990). The rate of secretion of this and other proteinase-inhibitors was found to in- crease significantly with time, correlating with im- munohistochemical evidence of differentiation induced by this method of culturing (Rubinstein and Herman 1975).
Our biochemical data show that the a2M-like factor purified from T67 cell supernatants has the same molec-

217
ular weight as ~2M obtained f rom human serum (180 Kd). On the contrary, some Authors observed that ~zM derived f rom melanoma and sarcoma cell lines ex- hibited a molecular weight of 140 Kd, probably related to differences at the level o f glycosilation (Grofova et al. 1988). Both serum- and tumor-derived ~2M are bound by anti-a2M antibodies, as shown by ELISA and western blotting; moreover, the c%M like factor isolated from gtioma culture medium is functionally active; in fact it exhibits a strong inhibitory function against trypsin pro- teolytic activity, paralleling the effect of serum a2M. However, further molecular characterization of the gtioma derived protein is required to define its homology to serum % M . The role and significance of this tumor derived proteinase inhibitor are not known at present. Malignancy, and in particular invasiveness, have often been linked to increased proteolytic activity. Consider- able evidence has amassed suggesting that protease product ion by tumor cells may be required for invasive- ness (Tryggvason et al. 1987).
Conversely, evidence suggests that some proteinase inhibitors, such as a2M may modulate the host- tumor immune responses and have immunosoppressive effects on natural killing and ant ibody dependent cytotoxicity (Dickinson et al. 1985), polyclonal B-cell activation (Chang et al. 1983), and the mixed lymphocyte response (Hubbard et al. 1981). In this connection, a bad prog- nosis has been suggested for patients with ~2M contain- ing tumors (Tahara et al. 1984; Matoska et al. 1988; K a t a o k a et al. 1989).
Recently, serum % M was shown to bind to a wide range of cytokines, including nerve growth factor, sug- gesting a potentially important mechanism for the re- gulation of their activity (James 1990; Ronne et al. 1979). With reference to this, a modula tory effect of a2M on the nervous system cannot be excluded. Moreover, since interactions of proteinases and proteinase-inhibitors appear to play a crucial role in cell migration (Guenther et al. 1985; Lindner et al. 1986) and neurite outgrowth (Pit tman t985; Hawkins and Seeds 1986; Monard 1988), % M synthesis by human astrocytes could be involved in brain developmental events. Our preliminary results in- dicate that human fetal astrocytes exhibit intracytoplas- mic positivity for a2M. This finding suggest that a2M could have a physiological role during embryo develop- ment.
In conclusion, these glioma cell lines could be used as a model to try to elucidate the role of a2M, if any, in the growth and differentiation of the nervous cells.
References
Barret AJ (1981) %-macroglobulin. Methods Enzymol 80: 737765 Bizik J, Vaheri A, Saksela O, Kalkkinen N, Meri S, Grofova M
(1986) Human tumor cells synthesize and secrete %-macroglob- ulin in vitro. Int J Cancer 37:81-88
Chang JL, Ganea D, Dray S, Teodorescu M (1983) Blast trans- formation of B celts induced by an e~-macroglobulin associated lymphokine produced in crowed lymphoid cell cultures. J Im- munol 130: 267-273
Chawla RK, Lawson DH, Sarma PR, Nixson DW, Travis J (1987) Serum %-proteinase inhibitor in advanced cancer: mass va- riants and functionally inert forms. Cancer Res 47:1179-1184
Cheung A, Lau HKF (1986) Isolation and partial characterization of a proteinase inhibitor from human colorectal adenocar- cinoma. Biochim Biophys Acta 882:200-209
Cooper EH, Turner R, Geeike A (1976) Alphaglobulins in the surveillance of colorectal cancer. Biomedicine 24: 171-178
Cusimano G, Palladini G, Lauro GM (1990) MHC Class II ex- pression by human glioma cells after in vitro incubation with soluble antigens. Acta Neurol Scand 81:215-222
Dickinson AM, Shenton BK, Alomran AH, Donnelly PK, Proctor SJ (1985) Inhibition of natural killing and antibody dependent cell-mediated cytotoxicity by the plasma protease inhibitor %- macroglobulin (et2M) and ot2M protease complexes. Clin Im- munol Immunopathol 36: 259-265
Dulbecco R, Allen R, Okada S, Bowman M (1983) Functional changes of intermediate filaments in fibroblastic cells revealed by a monoclonal antibody. Proc Natl Acad Sci USA 80:1915-1918
Dziegielewska KM, Saunders NR, Schejter EJ, Zakert H, Zevin- Zonkin D, Zisling R, Soreq H (1986) Synthesis of plasma pro- teins in fetal, adult and neoplastic human brain tissue. Dev Biol 115:93-104
Engvall E, Perlmann P (1971) Enzyme linked immunosorbent assay (ELISA). Quantitative assay of immunoglobulin G. Im- munochemistry 8: 871-874
Gebicke-Haerter PJ, Bauer J, Brenner A, Gerok W (1987) %-mac- roglobulin synthesis in an astrocyte subpopulation. J Neu- rochem 49 : 1139--1145
Grofova M, Larsson E, Bengtsson A, Bizik J, Westermark B, Ponten J (1988) Localization of a2-macroglobutin in human primary sarcomas and synthesis in established cell lines. In vitro Cell Dev Biol 24:369-372
Guenther J, Nick HP, Monard D (1985) A gila-derived neurite- promoting factor with protease inhibitory activity. EMBO J 4:t963-1966
Harris CC, Primack A, Cohen MH (1974) Elevated %-antitrypsin serum levels in lung cancer patients. Cancer 34:280-281
Hawkins RL, Seeds NW (1986) Protease inhibition can influence the direction of neurite outgrowth by neonatal mouse sensory ganglia. J Neurosci (Abstr.) 6:1117
Hubbard WJ, Hess AD, Hsia S, Amos DB (1981) The effects of electrophoretically "slow" and "fast" %-macroglobulin on mixed lymphocyte cultures. J Immunol 126:292-299
James K (1990) Interactions beetween cytokines and %-macro- globulin. Immunology Today 11 : 163-166
Kataoka H, Nabeshima K, Komada N, Koono M (1989) New human colorectal carcinoma cell lines that secrete proteinase inhibitors in vitro. Virchows Arch [B] 57:157-165
Keohane ME, Scott WH, Scott RV, Gonias SL (1990) Secretion of %-macroglobulin, %-antiplasmin, and plasminogen activator inhibitor-1 by gliobtastoma multiforme in primary organ cul- ture. J Neurosurg 73:234~241
Kramer MD, Justus C (1988) The anti-proteolitic compound %- macroglobulin in human skin. Arch Dermatol Res 280:93-96
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685
Lauro GM, Di Lorenzo M, Grossi M, Maleci A, Guidetti B (1986) Prostaglandin E2 as an immunomodulating factor released in vitro by human glioma cells. Acta Neuropathol (Berl) 69 : 278-282
Lindner J, Guenther J, Nick HP, Zinser G, Antonicek H, Schachner M, Monard D (1986) Modulation of granule cell migration by a glia-derived protein. Proc Natl Acad Sci USA 83:4568-4571
Lizonova A, Bizik J, Grofova M, Vaheri A (1990) Coexpression of tumor-associated %-macroglobulin and growth factors in human melanoma cell lines. J Cell Biochem 43:31 ~325
Matoska J, Wahlstr6m T, Vaheri A, Bizik J, Grofova M (1988) Tumor-associated %-macroglobulin in human melanomas. Int J Cancer 41 : 359-363

218
Matsuda K, Ogawa M, Murata A, Kitahara T, Kosaki G (1983) Elevation of serum immunoreactive pancreatic secretory trypsin inhibitor contents in various malignant diseases. Res Commun Chem Pathol Pharmacol 40:301-305
Monard D (1988) Cell-derived proteases and protease inhibitors a s regulators of neurite outgrowth. TINS 11 : 541-544
Mori T, Iijima N, Kitabatake K, Kohsaka S (1990) ct2-macro- globulin is an astroglia-derived neurite-promoting factor for cultured neurons from rat central nervous system. Brain Res 527: 55-61
Okumichi T, Nishiki M, Takasugi S, Toki N, Ezaki H (1984) Isolation of urinary trypsin inhibitor-like inhibitor from human lung cancer tissue. Cancer Res 44: 2011-2015
Parikh I, March S, Cuatrecasas P (1974) Topics in the methodology of substitution reactions with agarose. Methods Enzymol 34: 77-102
Pittman RN (1985) Release of plasminogen activator and a cal- cium-dependent metalloprotease from cultured symphatetic and sensory neurons. Dev Biol 110:91-101
Ronne H, Anundi H, Rask L, Peterson PA (1979) Nerve growth factor binds to serum ~z2-macroglobulin. Biochem Biophys Res Commun 87:330-336
Rubinstein LJ, Herman MM (1975) Studies on the differentiation of human and experimental gliomas in organ culture systems. Recent Results Cancer Res 51:35-51
Saksela O, Vaheri A, Schleuning WD, Mignatti P, Barlati S (1984) Plasminogen activators, activation inhibitors and a2-macro- globulin produced by cultured normal and malignant human cells. Int J Cancer 33:609-616
Sawaya R, Zuccarello M, Highsmith R (1987) cq-antitrypsin in human brain tumors. J Neurosurg 67:258-262
Schoenle E J, Adams LD, Sammons DW (1984) Insulin-induced rapid decrease of a major protein in fat cell plasma membranes. J Biol Chem 259 :1211~12116
Seitz R J, Wechsler W (1987) Immunohistochemical demonstration of serum proteins in human cerebral gliomas. Acta Neuropathol 73:145-152
Tahara E, Ito H, Taniyama K, Yokozaki H, Hata J (1984) c~i-anti- trypsin, al-antichymotrypsin, and az-macroglobulin in human gastric carcinomas: a retrospective immunohistochemical study. Hum Pathol 15:957-964
Tartakoff A, Vassalli P (1978) Comparative studies of intracellutar transport of secretory proteins. J Cell Biol 102:2244-2253
Towbin H, Staehelin T, Gordon J (1979) Electrophoretic transfer of proteins from polyacrilamide gels to nitrocellulose. Procedure and some application. Proc Natl Acad Sci USA 76:4350-4354
Tryggvason K, H6yhtyfi M, Salo T (1987) Proteolytic degradation of extracellular matrix in tumor invasion. Biochim Biophys Acta 907:19t-2t7





























