Embed Size (px)
Citation preview

John D. HortonTrudy L HortonRobin DzialoIan GravenorRalph MinterPamela RitchieLanier GartlandMartin D, WatsonMax D, Cooper
T-ceU and natural killer celldevelopmentin thymectomized Xenopus
Autfiors' addresses
Jo!in Horton'.Trudy Horton', Robin Dzio!o', !on Gravenor̂
Ra!p!i Minter', Pamela Ritchti;', Lanier Gari!and^
Martin IVutson'. Max Cooper̂
'Department of Biological Sciences. University
of Durham. Durham, UK.
'Division of Developmental and Clinieal
Immunology and the Howard Hughes Medical
Institute at the University of Alabama at
Birmingham. Birmingham. Alabama, USA.
Correspondence to:
Jolin D. Horton
Department of Bio!ogieal Sciences
University of Durham
South Road
Durham DHl 3LE
UK
Fax: 44 1913 742417
e-mail: j,[email protected],uk
Acknowledgements
This research was supported by project grants
from the Leverhulme Trust (JDH) and the
University of Durham (JDH and MDW) and
dirough BBSRC studentsliips (IG and RM). MDC
is a Howard Hughes Medica! institute
investigator. We are indebted to
Dr. Louis Du Pasquier for his generosity in
supplying molectilar probes for identifying
Xenopus T ceUs and for the Xenoptis tumour eel! !ines
used in our NK stndies.
Immunohgical Reviews 1998
Vol. 166; 245-258Primed in Denmark. All rights reserved
Copyriglit © Mtinksgaard 1998
Immunological ReviewsISSNOIOS-2896
Stjmmary: The Xenopus ear!y-thymectomy model system is used to investi-gate the extent to which the thymus controls T-cell development and toprobe the evolution of natural killer (NK) ceils. Loss of T-cell function fol-lowing thymectomy, together with the paucity of cells expressing mono-clonal antibody-defined T-cel! surface markers, and greatly reducedexpression of T-cell receptor p transcripts in spleen, Hver and intestine,indicate that T-cell development is minimal in the absence ofthe thymus.Our findings therefore mitigate against the idea tbat a substantial extrathy-mic pathway of T-cell development exists in early vertebrate evolution.Rather, they suggest that in this amphibian representative T cells are pre-dominately thymus dependent. In vitro studies with control and thymecto-mized Xenopus splenocytes reveal that a non-T/non-B population and alsotwo T-cell subsets all display natural cytotoxicity towards aUogeneic thy-mus lymphoid tumour cells (which are deficient in MHC antigen expres-sion). Since Xenopus thymectomized early in larval development are perma-nently deficient in T cells, tbey may provide a useful phylogenetic modelfor the study of NK cells.
Introduction
The immune system ofthe clawed frog, Xenopus, has been char-acterised in depth (1). These and other phylogenetic studieshave helped confirm a high degree of similarity hetweenImmune systems of all jawed vertehrates and have heen usefulin identifying novel immune system components of likelyimportance in mammals. One such example of this is the recentdiscovery of a developmentally regulated type-I transmem-brane protein, called CTX, on cortical thymocytes in Xenopus(2). Gene homologues for CTX, a receptor believed to play animportant role in thymocyte development, have since beenidentified in birds (3), mice and humans (L. Du Pasquier, per-sonal communication). likewise, studies on fish have indicatedevolutionary conservation of natural killer (NK)-cell receptors:thus catfish non-specific cytotoxic cells (NCC) and human NK-cell killing are hlocked by monoclonal antibodies (mAbs)against a vimentin-like molecule on catfish NCC (4).
Xenopus offers an advantage over mammalian model systemsfor investigating the role of the thymus in immune systemdevelopment in that it is possihle to remove the Xenopus thymus
245

Horton et al • T-cell and NK development in Tx Xenopus
at a rudimentary stage of T-cell development (5-7), The first
part of this review describes experiments using the Xenopus
early-thymectomy model to investigate the extent to which the
thymus controls T-cell development. We then focus attention
on candidate NK cells in control and thymectomised (Tx) Xeno-
pus in a search revealing that several populations of natural cyto-
toxic cells exist at this level of evolution.
Effect of thymectomy on T-cell development
Background
Although the thymus plays a special role in T-cell development,the possibility of a substantial extrathymic T-cell maturationalpathway is still a topical, controversial issue. Considerable evi-dence exists for some degree of extrathymic T-cell develop-ment, in which the intestine of euthymic rodents is thought toplay a major role (8-12), Thus in mice it has been suggestedthat gut endoderm attracts lymphoid precursors and inducesthese to undergo dt novo T-cell development within the intestinalepithelium (the so called intraepithelial lymphocytes (IELs))(10, 12), Evidence for such extrathymic development includesthe finding of recombination-activating gene-1 (Rag-1) tran-scripts and DNA deletion circles (containing T-cell receptor(TCR) VDJ genes) within IELs, the Utter frequently expressingthe phenotype TCRY5/CD8aa or TCRap/CD8aa (8, 10, 13).However, other evidence indicates that most IELs in eutbymicmice are in fact derived from the thymus, or are dependent onthymus-derived factors (14, 15). Evidence that the liver isanother site of extratbymic T-ceil development emanates fromstudies characterising TCR molecules on liver lymphocytes(16-18), especially during early ontogeny, prior to tbymusdevelopment (19, 20). Involvement of tbe bone marrowmicroenvironment in supporting the differentiation of T-cellprecursors into CD4'̂ and CD8^ T cells has also recently beenreported (21). Altbough tbe emergence of some lympbocytesexpressing TCR's in congenitally athymic rodents is cited as evi-dence for extratbymic T-cell maturation (22), sucb T-celldevelopment could well be promoted by vestigial tbymus tissuefound in nude mice (23, 24). Significant extrathymic develop-ment of IELs appears unlikely in sheep, since thymectomy intitero is followed by persistent depletion of intestinal lympho-cytes (25). In birds, a comprehensive search has failed to pro-vide evidence for either TCRaP or TCRyS extratbymic T cells(26-29).
The suggestion that extra thymic T cells occupy an inter-mediate pbylogenetic position between NK ceUs and thymus-derived T cells (30) and might reflect a more primitive patb-way of T-ceil development (3!) led us to search for thymus-
independent T-cell development at the amphibian level of ver-
tebrate evolution. In Xenopus, tbe tbymic anlage can be easily and
completely removed between 5 and 7 days of development (6),
At 5 days of age (stage 47), the tbymus comprises <500 cells
(32), including epithelial cells of tbe tbymic anlage and a few
lymphoid precursors, which began to enter the anlage during
the fourth day of larval development (33). Even by 7 days (stage
48), when thymocyte nimibers have reached approximately
4,000, fewer than 20% of these cells express tbe 120 kDa Xenopus
T-lympbocyte antigen (XTLA-1) (34, 35) and CD8 expression
is not yet identifiable (36). Tbe earliness of tbe thymectomy
operation, coupled with the fact that thymectoniized Xenoptis are
free of both tbe complex genetic defects inherent in nude
(athymic) rodents and direct maternal influence on tbe devel-
oping embryo, provide an unambiguous test model for assess-
ing tbe role of tbe thymus in supplying T cells to the rest of the
body. The question of whether or not T cells can be generated
in athymic Xmopus can now be more directly assessed witb the
acquisition of reliable markers for Xenopus T cells. Monoclonal
antibodies (mAbs) have been generated against Xenopus CD5
(37) and CD8 (38, 39) homologues, and other mAbs identify
less well characterised T-cell epitopes (34, 38). Additionally,
tbe Xenopus TCRp chain gene has recently been cloned (40).
Initial characterisation of lymphocyte development
in thymectomised Xenopus
Tbe effect of early (5-7 day) thymectomy on lymphocytedevelopment was initially examined in tbe spleen, a majorperipberal lymphoid tissue in Xenopus, which lacks lymphnodes. Altbough die overall growth of control and Tx Xenopuslaevis siblings over the 14-montb experimental period was com-parable (Fig. I), splenocyte numbers were reduced in Tx ani-mals to approximately 3 0-5 0% of control levels in all four agegroups examined (Table 1). Tbe athymic frogs routinely remainin excellent bealth at least until 9 months of age. However, wehave noted incidents of wasting or subcutaneous bydrops insome batches of older frogs. Both of these conditions, whichare of unknown aetiology, were more frequent in Tx animalstban in control siblings. Early tbymectomy carried out inanotber laboratory (41) indicated tbat most Tx Xenopus frogletsbecame infected by nematodes (Capillaria), and the 'flaky skin'disease caused by tbis parasite was corrected by tbymusimplantation. Tbus while most athymic Xenopus survive remark-ably well, lack of a thymus can lead to serious immunologicdeficiencies and susceptibility to certain pathogens.
When splenocytes were analysed for T and B-cell surfacemarkers by flow cytometry (Fig, 2), the mean percentage ofcells expressing the pan-T-celi marker CD5 was found to grad-
246 Immimoiogicol Reviews 166/ J 998

Horton et al • T-cell and NK development in Tx Xenopus
Fig. 1. The Xenopus thymectomy model system.A. At S weeks of age the paired, piginentedthymus gland (T) is readily visible through thetransparent skin of the dorsal pharyngealregion of a control larva, lying jusl posterior tothe eyes. B. Note absence of thymus in the 5-week-old larva thai had been thymectomized(Tx) ai 5 days of age. prior to T-lymphocytedevelopment. Blood vessels (red/black incolour) and nerves (white) are seen traversingthe pharyngeal region, the black abdominalregions can be seen posteriorly. Followingmetamorphosis al 7-8 weeks, conirol (1C) andTx (ID) siblings (here 6 months old) continueto display comparable growth rates, despiteabsence of bonu fide T cells. Absence ofthymus inTx frogs is confirmed by dissection andhisiological analysis of the ihymic regions.Magnification: x4 (A, B); xO.7 (CD).
ually increase with age in normal (control) X. laevjs. from 28%
in one-month-old larvae (age group I) to 62% at 10-14months of age (group IV). (Xenopus B cells do not constitutivelyexpress CD5 (37).) The percentage of CD8* splenocytes like-wise increased from 11% in larvae to 27% in 10—14-month-old adults. Approximately 20% larval and 35% adult spleencells from controls expressed the XTLA-1 antigen. In the Txanimals, however, XTLA-1*, CD5^ and CD8-̂ (F17mAb) sple-nocytes were virtually undetectable at all ages (Fig. 2; data notshown), in keeping with an earlier study revealing loss ofXTLA-1 + cells from diverse lymphoid tissue sites of early-TxXenopus (35). In contrast, the B-cell percentages were consis-tently higher in Tx animals, as expected from the initial char-
acterisation of B cells and Ig levels in such frogs (42). Typical
staining patterns for splenocytes from 10-14-month control
and Tx frogs are shown in Fig, 3.
Search for T cells in diverse lymphoid tissues following
th/mectomy
The 10—14-month-old control and early-Tx X. luevis were alsoanalysed for T-cell development in the liver and intestine - sitesof putative extra thymic T-cell development in mammals. Lym-phocyte numbers obtained from Xenopus liver and gut variedappreciably from one preparation to another, making reliablecomparison of absolute numbers of these cells in control andTx frogs impractical. Mean percentages of mAb-stained lym-
Immunologicdl Reviews 166/1998 247

HHorton et a! • T-cell and NK development in Tx Xenopus
CD5*
III IV I II III IV
Age groups
III IV
Fig, 2. Ontogeny of T and B cells in the spleens of control andthymectomised Xenopus. Mean percentages of sp!enocytes (±SD) stainingfor CD5, CDS, or IgM are shown in control (•) and thymectomized ( o)XenopQs kevis. Exp!anation of age groups I-JV, the number of experimentsperformed oti each group, and the mean absolute splenoeyte numbers inthese animals are shown in Table I, Splenocytes, prepared as describedelsewhere (5!), were washed in amphibian strength PBS (APBScontaining 0.1% NaN; and 0,1% BSA) before incubation with either F17(anti-CD8) (38) or D8 (anti-IgM) (38) primary mAb, After washing, thecells were incubated with FITC-labelled, rabbit anti-mouse
immunoglobuiin, adsorbed with 5% Xenopus serum. Cells were thenwashed with AFBS contait:iing 1% normal mouse sertim and counter-stained with PE-labelled, 2B1 mAb (anti-CD5) (37), The control mouseantibody for indirect FITC stainings was CT3, an anti-chicken CD3 (87);the phycoerythrin control was mouse IgG conjugated to phycoerythrin.Five to ten thousand lymphocytes were analysed using Becton DickinsonFACScan or Coulter XL flow cytometers. Gates were set by forward andside scatter to exclude erythrocytes, granulocytes and dead cells fromanalysis, the latter being confirmed by propidium iodide staining.
phocytes in spleen, liver and gut (intestine) of control and Tx
froglets are given in Table 2, Representative staining patterns are
shown in Fig, 4,
Mean percentages of CD5+ lymphocytes recorded in
spleens of control frogs (70%) were similar to those found in
gut (75%) and liver (59%). AbouE half of these Tcells co-
expressed CD8, although the proportion of intestinal Tcells
expressing the CD8 epitope recognised by mAb AM22 tended
to be somewhat higher. The levels of splenic and hepatic lym-
phocytes staining with the D4 mAb, which identifies an 80 kDa
homodimer on Xenopus Tcells (38), were consistently just
below the total percentage of CD5+ T cells. In contrast, dual
Table 1. Effect of thymectomy on splenoeyte development Table 2. Percentages of mAh-stained lymphocytes in spleen, liver andgut of control and thymectomised Xenopus
Splenoeyte numbers' at different ages
IV
(1 month) {2-5 months) (fr-9 months) (10-14 months)
Control
Tx0.3
0.1
5.4±3.5
2,4±1.5
13,7±5.8
5.7±4.0
22.4±15,6
12.6±10.8
'Values represent mean cell numbers+1 standard deviation (xiO'), At 1 montb(age group I), 4-8 larval spleens were pooled; thereafter 5-7 individual spleenswere examined at each time point. Thymectomy was performed at 5-7 days of ageand animals reared at 23°C,
Control
Tx
{±S.D.) of mAb-stained cells
Organ
Spleen
Liver
Gut
Spleen
Liver
Gut
2B1
70±13
59 ±375+3
6±3
21+11
32+10
AM22
33±9
31 ±4
49±8
5±3
20±8
27+7
F17
33±7
3O±3
35+8
2±0
4±2
5±5
D4
68±13
56±3
17±11
2±0
5±4
4+3
l^arkers were set to exclude from positive analyses 98% of cells stained witb con-trol mouse antibody. Mean percentages of lymphocytes (± standard deviation)expressing CD5 (mAb 2B1), CD8 (mAbs AM22 & F17), and the 80 kDa epitoperecognised by mAb D4 (specificity unknown} are given, Spleen, liver and gut wereexamined from 5 animals (10-14 months old) in each experimental group. Thymec-tomy performed at 5-7 days of age,
248 Immtmological Reviews !66 / !998

Horton et al • T-cell and NK development in Tx Xenopus
CONTROL
35% 29% 52%
: ,1
''••'W.. . , n.Tif
40%
Fig. 3. Representative flow cytometricanalysis of mAb stained splenocytes fromcontrol and thymectomised adult Xenopus,Splenocytes from 1 -year-old X. laevis werestained sequentially with anti-CD8 (FITC) andthen anti-CD5 (PE) mAhs or anti-IgM (FITC)followed by anti-CD5 (PE) and surfacefluorescence evaluated by flow cytometry. as inFig. 2. Percentages of CD5'CDS"lymphocytes(putative helper T cells), CD5*CD8+lymphocytes (cytotoxic T cells), and CD5-IgM+ B cells are indicated. Quadrants set toexclude 98% cells stained with control reagentsfrom positive analysis.
THYMECTOMIZED
:
<1%
' 1 •
(1%
' ' 'J ' • ' • • " •
84%
CD8
colour flow cytometry revealed that a significant population of
CD 5+ T cells in the gut were D4".
In Tx Xenopus, the epitopes recognised hy the Fl 7 anti-CD8
mAb and the D4 mAh were not found on splenocytes, and were
often undetectable in liver and gut. However, splenocytes
expressing low levels of CD5 and the AM22 anti-CD8 epitopes
were relatively abundant in the Tx frogs. (The AM22 mAb may
detect a CD8a epitope and the F17 mAb may detect a CDBp
epitope. but this has not yet been proven.) Approximately 30%
of the lymphocytes from liver and intestine of these Tx frogs
were CDS'" and AM22i", but none expressed the distinctly
higher fluorescence intensities of these markers typical of con-
trol T cells (Fig. 4). Compared with controls, elevated propor-
tions of IgM'̂ B cells were found in the spleen, liver and gut of
these Tx frogs. The XT-1 mAb was not used in these studies,
because we have recently found the XTLA-1 antigen is not iden-
tifiable in some batches of control X. lams (43).
Histological analysis of the spleen from 1-year-old control
frogs indicated that CDS''T cells were concentrated in the zone
near the boundary region of the white and red pulp. CD5+ cells
were also scattered throughout the white pulp follicles
(Fig. 5A), which are rich in IgM+ B lymphocytes (44, 45). The
distrihution of the CD8+ (AM22+) subpopulation of cells was
similar to that of the total population of CD5+ T lymphocytes.
Splenic CDS+ and CD8+ cells were not identifiable in splenic
sections from Tx siblings (Fig. SB). CD5+and CD8+ (AM22+)
lymphocytes were found throughout the intestine of control
Xenopus (Figs SC & SD), both in the lamina propria, where nod-
ular lymphoid tissue and more diffuse lymphocyte collections
are seen, and in the intestinal epithelium (the IELs), near the
hasement memhrane. These CDS+ and CDS"'lymphocytes typ-
ically displayed a circumferential "rim" pattern of staining. In
contrast, the CD5'". CD8''' cells identified by FACS in the intes-
tine of Tx frogs were not histologically visihle, even with
hiotin-avidin enhancement of the mAb staining (Fig. SE).
Search for TCR transcripts following thymectomy
The most stringent evidence for the existence of T cells would
he the detection of TCR gene rearrangement and expression.
Anti-Xenoptis TCR antibodies are not yet available, but complete
TCRp cDNA clones have recently been isolated and sequenced
(40), This information was used to design consensus Vp and Cp
ImmunoJogico] Reviews 166/1998 249
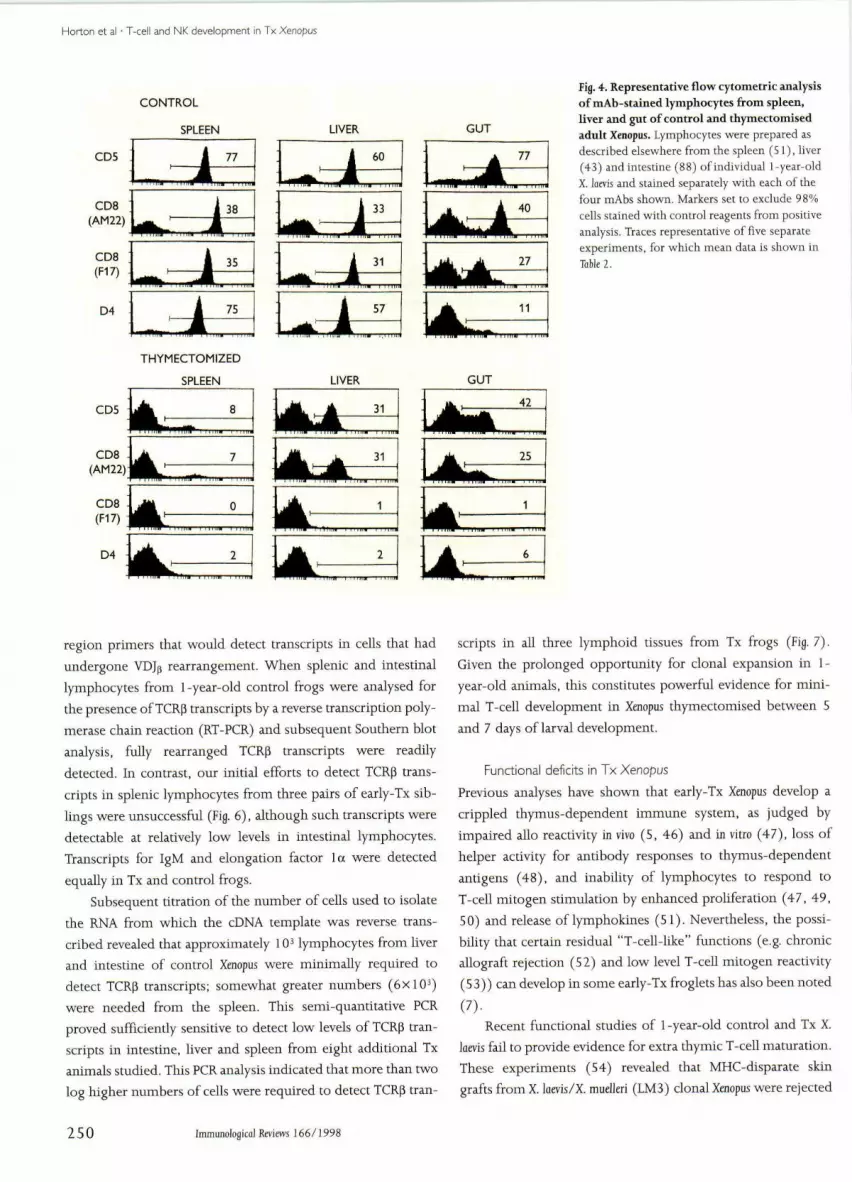
Horton et al • T-cell and NK development in Tx Xenopus
CONTROL
SPLEEN
CD5
CD8(AM22)
77
38
D4
35
75
THYMECTOMIZED
SPLEEN
CD5
CDS(AM 22)
CDS(F17)
D4
LIVER GUT
LIVER
31
31
GUT
' - '''
1 A
42
25
1
"- -
6
Fig. 4. Representative flow cytometric analysisof mAb-stained lymphocytes from spleen,liver and gut of control and thymectomisedadnlt Xenopus. Lymphocytes were prepared asdescribed elsewhere from the spleen (51), hver(43) and intestine (88) of individual i-year-oldX, laevis and stained separately with each of thefour mAbs shown. Markers set to exclude 98%cells stained with control reagents from positiveanalysis. Traces representative of five separateexperiments, for which mean data is shown inTable 2,
region primers tbat would detect transcripts in cells tbat hadundergone VDJp rearrangement. When splenic and intestinallympbocytes from 1-year-old control frogs were analysed forthe presence of TCRp transcripts by a reverse transcription poly-merase chain reaction (RT-PCR) and subsequent Soutbern blotanalysis, fully rearranged TCRp transcripts were readilydetected. In contrast, our initial efforts to detect TCR|3 trans-cripts in splenic lympbocytes from tbree pairs of early-Tx sib-lings were unsuccessful (Fig, 6), altbough sucb transcripts weredetectable at relatively low levels in intestinal lympbocytes.Transcripts for IgM and elongation factor la were detectedequally in Tx and control frogs.
Subsequent titration of the number of cells used to isolatethe RNA from which tbe cDNA template was reverse trans-cribed revealed tbat approximately 10̂ lymphocytes from hverand intestine of control Xenopus were minimally required todetect TCRp transcripts; somewhat greater numbers (6x10^)were needed from the spleen. Tbis semi-quantitative PCRproved sufficiently sensitive to detect low levels of TCRp tran-scripts in intestine, liver and spleen from eight additional Txanimals studied. This PCR analysis indicated that more than twolog higher numbers of cells were required to detect TCRP tran-
scripts in all three lympboid tissues from Tx frogs (Fig, 7),
Given tbe prolonged opportunity for clonal expansion in 1-
year-old animals, tbis constitutes powerful evidence for mini-
ma! T-cell development in Xenopus thymectomised between 5
and 7 days of larval development.
Functional deficits in Tx Xenopus
Previous analyses bave shown that early-Tx Xenopus develop acrippled tbymus-dependent immune system, as judged byimpaired allo reactivity in vivo (S, 46) and in vitro (47), loss ofhelper activity for antibody responses to thymus-dependentantigens (48), and inability of lymphocytes to respond toT-cell mitogen stimulation by enhanced proliferation (47, 49,50) and release of lympbokines (51). Nevertheless, tbe possi-bihty that certain residual "T-cell-like" functions (e,g, chronicallograft rejection (52) and low level T-cell mitogen reactivity(53)) can develop in some early-Tx froglets bas also been noted
(7).Recent functional studies of 1 -year-old control and Tx X.
loevis fail to provide evidence for extra tbymic T-cell maturation.Tbese experiments (54) revealed that MF€C-disparate skingrafts from X, luevis/X. mudleri (LM3) clonal Xenopus were rejected
250 Immunologica! Reviews 166/1998

Horton et al • T<ell and NK development in Tx Xenopus
-^•^•'•'if i " w i i * ^* ' •*-• A - ; • ? : ' • ^
Fig. 5. Imraunohistochemical analysis ofspleen and gut sections from control andthymectomised Xenopus adults. Four to six-micromelrc cryostat sections from spleen andintestine of 1-year-old X. loevis were fixed inacetotie and incubated with anti-CDS (A-C) oranti-CD8 (AM22) mAbs (D-E).ImmunosUining employed a biotin-avidin-peroxida.se detection technique (36). A. Spleenfrom control showing CD.S+ cells in red pulp(RP) margitial zone, jusl outside white pulp(WP), which also conuins a few scatteredT cells. B. Thymectomised spleen showingabsence of CDS*" cells. Arrows in A and B pointto bntmdary layer which separates white 2ndred pulp, and whicb frequently displaysperoxidase reaction product even in controlCT3-sUined sections. C . D. Intestine fromcontrol Xenopus stained for CDS* (C) and CD8*(D) intraepithelial lymphocytes (IEL). Nodularcollections (arrowed) of lymphocytes in laminapropria also express these antigens. E. Intestinefrom thymectomised animal showing absenceof identifiable CDS* lymphocytes. Ceils of theintestine are stained heavily with methyl greencounterstain, but no specific peroxidasereaction product is found in the thymectomisedanimal. L = ltnnen of gut. Data representativefrom five separate experiments. Magnification:X200.
by control recipients in <3 weeks, whereas grafts persisted in
perfect condition for >100 days on all Tx recipients. Immuno-histochemical analysis illustrated that LM3 grafts on controlsbecame heavily infiltrated by CD5"̂ and CD8+ lymphocytes,whereas MHC-disparate grafts on Tx animals remained devoidof such cells. In vitro-cultured splenocytes from Tx siblings ofthe grafted animals proved unreactive to the T-cell mitogenconcanavalin A (ConA), whereas in keeping with the higherproportion of B cells, splenocytes from Tx frogs responded bet-ter (higher incorporation of ^H-thymidine (TdR)) to Escherichiacoli lipopolysaccharide (LPS) than did splenocytes from con-trols. Mitogen reactivity of lymphocytes from 1 -year-old Xenopusliver and intestine have also been analysed. CeUs were culturedfor 3 days with ConA, or che pan-lymphocyte stimulator, phor-bol myristate acetate (PMA). and examined for mitogen-induced 'H-TdR uptake (Fig. 8). ConA was highly stimulatoryfor lymphocytes extracted from spleen, liver and gut from con-
trols, whereas cells from all lymphoid tissues from Tx frogs
failed to respond. The viability of cultured lymphocytes from
spleen, hver and intestine of Tx frogs is demonstrated by their
ability to proliferate in response to PMA stimtilation.
Effect of thymectomy; summary
This analysis of the Immune system in Xenopus thymectomizedas 5-7 day larvae indicates the virtual ahsence of identifiableT cells, greatly reduced expression of TCRp transcripts, and lossof detectable T-cell function. Our study thus provides evidencethat T-cell development is minimal in the absence of the thy-mus in this amphibian representative. On the other hand, theappearance of CD5'". AM22'" cells (which may representCD8aK+ cells) in the liver and gut, together with the low levelsof TCR|3 transcripts in lymphoid cells of Tx frogs, indicates avery limited developmeni of cells with "T-like" feattires inthese athymic animals. It seems likely that these latter cells have
Immunologitfll Reviews i 66/1998 251

Horton et al • T-cell and NK development in Tx Xenopus
Spleen Intestine
normal Tx normal Tx
Fij. 6. Analysis of TCRp, IgM and elongation factor la transcripts insplenic and intestinal lymphocytes from control and thymectomizedXenopus adults. Total RNA was obtained from splenic and intestinallymphocytes from one pair of thymectomised (Tx) X. loevis (I -year-old)and one pair of normal (control) siblings. First strand cDNA template wassynthesised using oligo dT. PCR amplification followed witholigonucleotide primers corresponding to TCRp chain framework region2, position 79-99, 5' AAC ATG TTC TGG TAC CAA CAG 3' and a reverseprimer complementing constant region position 742—762, S' AGC CGTGAC AAT GAG CCC ATA 3', Primers amphfying portions of CM at position569-589, 5' TAT AAA CCT GTG ATG CAA TCT 3' in exon 1 and reverseposition 740-760, 5' GGATGG AGG GTG AAT AGA CAC 3' in exon 2, Thesequence of a 5' primer in exon 4, position 675—695 of elongation factor1 a is CCA TGG TTT AAG GGA TGG AAA and its 3' partner in exon 5,position 890-91 0 is AGT TGT TAC ATT AAC AGG GGC. Cyclingparameters were as follows: 1 minat 94°C, 1 minat S8°C, 30 minat 72°Cfor 30 cycles followed by a 10 mine extension at 72°, For Southern blotof the PCR products, I Ofil of a SOpl PCR volume were electrophoresed ona 4% Nusieve GTG agarose gel. The resultant gel was Southern blotted tonylon aiid UV cross-linked. Oligonucleotide probes corresponding toposition 670-690 of TCR Cp, 690-710 of C|j, and 795-815 of EF la wereS' end labelled with ^'P 7ATP and hybridized to the membrane at 42° in50% formamide. The membrane was washed to a stringency of 45° inO.IXSSC, 0,1% SDS,
emerged extra tbymicalty, but it is conceivable that they devel-oped from cells influenced by the tbymus for the very briefperiod prior to larval thymectomy The question of whethersome yS T-hneage cells can develop in Tx Xenopus also remainsunanswered. The Y§ T-cell receptor genes also arose early in ver-tebrate evolution, baving been described togetber with aP TCRgenes in cartilaginous fish (55), birds and mammals, althoughnot yet defined in Xenoptjs, Tbe severe impairment of T-celldevelopment demonstrated in Tx Xenopus nevertheless mitigatesagainst a substantial extrathymic pathway of T-cell develop-ment eariy in vertebrate evolution.
Search for candidate NK cells in thymectomized Xenopus
Baci<ground
Mammahan NK cells are large granular lymphocytes distrib-uted in diverse lympboid tissues. Tbese cells, wbich can be dis-tinguished from T cells by their lack of rearranging TCRs,exhibit innate, non-antigen-specific defence against certaintumour targets and virally infected cells, especially those tbatare deficient in MHC class I protein expression (5 6). Such aber-rant targets cannot be dealt with by cytotoxic T lymphocytes(CTL), whose clonally distributed antigen receptors are MHICrestricted. Tbe reahsation that NK cells provide crucial immunedefence, together witb increasing awareness that these cells areimportant regulators of adaptive immunity (57), has lead to awealth of research attempting to defme the molecular basis ofantigen receptors and target ligands involved in the activationand inhibition of mamtnahan NK cells (58-62), NK-cell inhib-itory receptors in mouse and man are beginning to be charac-terised in considerable deptb and it is now realised that the twodifferent structural types of inhibitory receptor existed beforethe divergence of primates and rodents (61).
Since NK cells are believed to be the evolutionary forerun-ners of CTL (63), studies on lower vertebrates may reveal novelanimal models to explore crucial issues concerning NK-cellbiology For example, the identification of cytoplasmic CD3proteins and botb surface CDS and interleukin (IL)-2 receptorson cbicken NK cells (64) support the notion (65) that T cellsand NK cells derive from a common progenitor. As mentionedin the introduction, studies on catfish indicate the conservationof NK-cell antigen receptors (4). Heterogeneous populationsof non-specific cytotoxic cells (NCC) bave recently beendescribed in catfisb, where separable peripberal blood leuco-cyte populations from unimmunised animals are spontane-ously cytotoxic to eitber allogeneic or virally infected targets(66), Cytotoxicity mediated by these two populations is notinhibited by an anti-NCC mAb (4, 66), vv̂ hereas tbe xenoreac-tive NCC cells extracted from catfisb kidney are inhibited bytbis mAb, Distinct populations of alloreactive and xenoreactivenon-specific cytotoxic cells have also been observed in tumour-bearing damselfisb (67), Altbougb kiUing by fisb cytotoxiccells frequently displays lack of antigen specificity, the unavail-ability of either T-cell-depleted fish or appropriate anti-fishT-cel! antibodies (68) precludes documentation as to whethersuch killing represents diverse NK-like populations rather thanT cells.
Amongst amphibians, NK activity against xenogeneictumour targets and viruses is suspected in several anuran spe-cies and its role in the very effective tumour immunity at thisevolutionary level has been discussed (69). Our ongoing
252 Immunological Reviews 166/1998

Horton et al • T-cell and NK development in Tx Xenopus
10-'
10
Thymectomised
Normal
Fig. 7. Approximate mean (± SD) numbers ofcells needed to detect TCRp transcripts indiverse lymphoid tissues of control andthymectomized adult Xenopus. Numbers oflymphocytes from spleen, liver and intesdne(from 1 -year-old X. lams) used lo extract RNAwere titrated [o allow a semi-quantitacive RT-PCR, based on methodology given in Fig. 6.Data from eight separate experiments, each onindividual normal (control) and 5-6-day-chymectomised Xenopos.
Spleen Liver
50,000
•
•
Medium
ConA
PMA
Spleen Liver Gut Spleen Liver Gut
fig. 8. Mitogen responsiveness of lymphocytesfrom control and thymectomised adnltXenopus. Graph shows mean DPM ^H-TdRincorporation for lymphocytes from spleen, liverand gut of individual control and chymectomizedX. laais aged 12 months, stimulated in vitro with2.5 fig ml-1 concanavalin A (ConA) or lOngmh'phorbol myristace acetate (PMA). or cultured inmedium alone. Lymphocytes were cultured at27°C for 48 h, before being pulsed with ^H-TdR18-20 h prior to harvesting and liquidscintillation counting. Data are representative ofthree separate experiments.
Control Thymectomised
research is attempting to identify candidate NK cells in Tx Xeno-
pus.
Xenopus splenocytes exhibit NK-like activity
The remarkable capacity of early-Tx Xenopus to survive in the
absence of bona fide T cells (see above) attests to the importance
of otber elements of host defence in these frogs. Altbough
powerful antibiotic magainins in Xenopus skin (70) may be
essential in this respect, it seems likely that NK cells play an
important role in tbe survival of Tx Xenopus. Our initial experi-
ments (71, 71) revealed that splenoeyte effectors taken directly
from Tx (1-year-old) X. laevis, but not from control sibhngs.
showed natural lytic activity towards allogeneic thymus
tumour-derived lymphoid cells (6-h ^'Cr release assay). (Thy-
mus tumour-derived lymphoid cell lines, such as the B3B7 cells
from homozygous Xeiiopus laevis family ff, are appropriate targets
for candidate NK cells, since they are deficient in class I
(class Ia) and II MHC antigen expression (73),) In contrast,
some Tx batches of Xenopus tested directly ex vivo fail to lyse allo-
tumour targets (71). Such variation in natural cytotoxicity may
well relate to differences in natural antigenic challenge in vivo
and associated cytokine modulation. In this respect lytic capac-
ity towards allogeneic tumour cells can be readily induced in
control Xenopus and elevated in Tx frogs by a single injection of
Immunological Reviews 166/1998 253

Horton et al • T-cell and NK development in Tx Xenopus
tumour cells, with maximal splenocyte cytotoxicity occurring3 days post-injection (71). These investigations also revealedthat splenocytes, even those from tumour-injected frogs, wereunable to lyse allogeneic splenic lymphohlasts or erythrocytetargets, even when the latter are coated with IgY (the XenopusIgG equivalent); moreover, we were unable to demonstrate anysplenocyte-induced lysis of the hmnan NK-cell target K562.
Lymphokine-activated natural c/totoxicity
Lymphokine-activated killing (LAK) in Xenopus is suggested (71)by the ohservation that when splenocytes from 1-year-old Txfrogs were cultured for 48 h in growth factor-rich medium(GFM) from ConA-stimulated cells {probably containing IL-2-like material (74)), their capacity to kill allogeneic tumour tar-get cells was significantly enhanced (Fig, 9). Although thiscytokine-indnced cytotoxicity is reminiscent of LAK activity,the question of whether culture in GFM induces splenocytesfrom Tx frogs to kill a wide array of target types was not exam-ined. It would be interesting to observe whether prolongedincubation of Xenopus splenocytes in GFM (>48 hours) results inthe increased promiscuity of target cell killing that is typical ofmammalian LAK cells (75). LAK activity was first suspected inXenopus splenocytes from frogs injected with the iridovirus FV3,which, after restimulation in vilro in the presence of T-cellgrowth factor-rich supernatant, exhibited non-specific cyto-toxicity (76). The potential of GFM to stimulate NK-like activ-ity in Tx splenocytes is consistent with previous studies, wheresupernatants from activated T cells were shown to induceenhanced tritiated TdR incorporation by Ig" splenocytes fromTx Xenopus (51). In hnmans, a NK-cell prohferation-inducingfactor distinct from IL-2 has recently heen identified (7 7),
Splenocytes from control Xenopus are also induced to displayanti-tumour cytotoxicity following culture in tumour medium,but this was not dependent on addition of GFM (Fig. 9). Itseems likely (see below) that T lymphocytes in the controlspleen, hy responding to elements in the culture medium,achieve effective activation of NK-like cells, without the needfor addition of exogenous T-cell-derived cytokines. In theabsence of splenic T cells, such exogenous cytokines are crucialfor in vitro activation of NK-like cells.
Cellular basis of NK-like activityMagnetic cell sorting of mAb-labelled T and B cells has recentlyheen used to examine the cellular basis of Xenopus splenocytecytotoxicity towards allogeneic thymns tumour-derived lym-phoid cells, flow cytometry being employed to confirm thepurity of spleen subpopulations (78). Splenocytes from controlfrogs, depleted of T cells and then cultured for 48 hours.
behaved hke unsorted Tx splenocytes, in that they became allo-cytotoxic effectors, but only after culture in GFM. Sorting withanti-Ig mAbs revealed that purified B cells from hoth controland Tx Xenopus, even after culture in GFM, failed to kill allotu-mour targets, whereas B-cell-depleted splenocytes from thesesame frogs hecame effective killers (Fig, 10). These experimentstherefore indicate that non-T/non-B splenic leucocytes inunimmunised control and Tx Xenopus can display natural lyticactivity towards allogeneic tumour cell targets and representNK candidates, (The viahility of purified B cells produced hyimmunomagnetic sorting was shown hy their ability torespond to the B-cell mitogen, LPS (72).)
Experiments employing magnetic cell sorting have alsoindicated that cytotoxicity against tumour cells (but not alloge-neic lymphoblast targets) is mediated by CD5+ T cells fromXenopus spleen following culture in GPM (Fig. 10), This findingthat T cells from non-immunised, outbred X. laevis can lyse allo-geneic B3B7 tumour cells is of interest, since in vitro cytotoxicityof allogeneic splenic lymphoblasts by CTL requires prior Jn vivoimmunisation with allogeneic lymphocytes or skin grafts, andrestimulation of splenocytes in vitro for 5 days (79). Onr obser-vation that cultured T cells can kill allogeneic tumour cells isconsistent with recent findings from early thymectomy exper-iments (80), indicating that T cells play an important role inthe rejection of ff strain thymus tumour cells implanted intominor histocompatibility antigen-disparate ff Xenopus hosts.
The question of which T-cell subsets in unimmunisedcontrol Xenopus are capable of lysing allothymus tumour targetsfollowing cell cnlture has been investigated through sequentialmagnetic sorting (78); this procedure obtained cytotoxicT cells (CD8+CD5+), putative helper T cells (CD8-CD5+), anda non-T-cell subset (CD8- CD5-). The CD8- lymphocytesbecame effective killers after a 48-h culture in GFM (Fig, 11), afinding that was not entirely unexpected in view of the knownabihty of mammahan GTL to become NK-like after IL-2 treat-ment. More surprising was the allocytotoxicity exhibited bythe CD8- CD5+ T-cell population cultured in GFM. (There isno anti-CD4 reagent available to identify the Xenopus CD4+T cells,) This population achieved higher levels of tumour celllysis than the enriched CD8+T-cell (and T-cell-depleted) sub-populations. In mammals, CD4-̂ CTL clones exist that can killtargets relatively quickly, probably through binding to thetumour necrosis factor receptor family memher Fas (81, 82).Interestingly, the presence of Fas has recently been demon-strated on apoptosing Xenopus lymphocytes (L, N. Ruben, per-sonal communication).
254 Immimologica! Reviews 166/1998

Horton et al • T-cell and NK development in Tx Xenopus
Control
10:1 20:1 40:1
E:T ratio
10:1 20:1
E:T ratio
40:1
Fig. 9. Elevated cytotoxicity towards BjB?lymphoid tumour targets by splenocytes fromcontrol and thymectomised adult XenopusfoUowing in vitro culture. Splenocytes from one-year-old control and thymectomised (Tx) X. laeviswere tested directly ex vivo (0 hrs) and then againafter 48 h in tumour culture medium alone (med)or in culture medium supplemented with 25%growth factor-rich medium (GFM) generatedfrom ConA-stimulated cells. Data show meanpercentage specific '•'Cr release (±SE) from 5x10*B3B7 targets at various effector: target (E: T) ratiosfollowing 6-h assay. (All cell culture andcytotoxicily assays carried oul at 27°C.) Nineseparate experiments were carried out withspleens from individual control frogs, five withindividual Tx frogs.
u
40
30
20
10
0
-10
40
30 -
20
10
0 +
-10
CD5-ve
5:1 10:1
Ig-ve
20:1
5:1 10:1 20:1E:T ratio
40
30
20 4
10
0
-10
40 -r
30 -
20 -
10
0
-10
CD5+ve
5:1 10:1
Ig+ve
20:1
5:1 10:1
E:T ratio20:1
Fig. 10. Cytotoxicity towards allogeneic B3B7lymphoid tumour targets mediated bysplenocyte suhpopulations from control andthymectomised adult Xenopus. The subsets wereobtained by magnetic bead separation (78) usinganti-CD5 mAb 2B1 to deplete/enrich for T cellsor a 50:50 mix of anti-IgM mAb 8E4 (89) and acocktail of three anti-Xenopus Ig light chain mAbs(90) to deplete/enrich for B cells. The mAb-coated splenocytes were then incubated withMACS anti-mouse Ig paramagnetic microbeadsand populations enriched (>95%) or depleted(<2%) for T or B cells were collected by magneticcell sorting. The sorted cells were cultured for 48hours in medium alone or in GFM and thenincubated with BjB, targets at various effector:target (E:T) ratios for a 6-h ^'Cr release assay. Topgraphs sbow mean data for T-cell-depletedsplenocytes (CD5-) and purified T cells (CD5+)from four control animals. Mean data for B-cell-depleted splenocytes (Ig-) and purified B cells(Ig+) from three 1 -year-old control frogs areshown in tbe middle grapbs, and from threesibhng Tx frogs in the bottom graphs. Meanspecific 5'Cr release ±SE from 5x10* targets isshown.
U
_ Tx Ig-ve
50 -
40
30
20
10 +
0
-10
5:1 10:1
E:T ratio20:1
60
50
40
30
20
10
0
-10
Tx Ig+ve
Medium
GFM
5:1 10:1 20:1
E:T ratio
Immunological Reviews 166/1998 255

Horton et al • T-cell and NK development in Tx Xenopus
CD8 +ve (CD5+ve) CDS -ve (CD5+ve) CD8 -ve (CD5-ve)
20:1 20:1 20:1
Fig. 11, Cytotoxicity towards allogeneic BjB, lymphoid tumour cellsmediated by two T-cell subsets and a non-T-splenocyte popnlationfrom adult Xenopus. Splenocytes from a control, 1 -year-old frog wereinitially incubated with mAb AM2 2 to label CD8+cells and a subset highlyenriched (98%) for CD8 collected by magnetic separation (78). The CD8-cells were then incubated with mAb 2B1 (anti-CD5) and again
magnetically .sorted to produce highly enriched (99%) CD5+ (CD8") andalso CD5" CDS populations. Each of these three populations was culturedfor 48 h in medium alone or in growth factor-rich medium (GFM — seetext) and then tested for tytotoxicity against B1B7 targets al variouseffector: target (E:T) ratios in 6-h ^'Cr release assay. Mean data from threeanimals show specific '̂ Cr release ±SE from 5 x i 0̂ targets.
Ontogeny of NK-like activity
Our initial ontogenetic studies of NK-like activity revealed that
splenocytes from 6—7-week-old (stage 56/57) X. laevis larvae
were unable to display cytotoxicity towards B3B7 cells, even
after 2 days in vitro culture in GFM (78), These in vitro findings
are consistent with in vivo experiments in which Xenopus larvae,
in contrast to young adults, were unable to destroy minor H
antigen-disparate tymphoid tumour cells (73, 80, 83). Even
though in vivo reactivity towards MHC-disparate tumour cells by
tbe larval immune system has been described (73), our in vitro
studies indicate chac components of such larval aiiocytotoxicity
remain defective.
The lack of NK-like activity in Xenopus larvae is not unex-
pected, given tbat MHC class I antigen expression is essential
for inhibition of NK ceils (59, 60) and tbat MHG class Ia anti-
gens are not detectable on Xenopus cells prior to metamorphosis
(84). If NK cells were functional in the MHC class I-negative
Stage, larvae might risk auto immtrne destruction!
Xenopus NK cells: summary and future directions
The above experiments employing adult Xenopus splenocyte
populations indicate tbat a non-T/non-B population and two
T-cell subsets all display natural cytotoxicity towards allogeneic
lymphoid tumour cells, following a 48-h culture in growth fac-
tor-rich medium; such natural cytotoxicity is not evident when
larval splenocytes were similarly tested. Early-Tx Xenopus, with
their dearth of T cells, may provide a useful phylogenetic
model for generating and screening of mAbs against the non-
T-cytotoxic population, that is likely to include candidate NK
cells. Tbe generation of mAbs that identify Xmopus candidate NK
cells, using both hybridoma and phage display (85. 86) tech-
nologies, is a major current focus. Such mAbs will allow us to
explore lymphoid tissue sites where NK-cell development is
initiated, wbich will include the gnt, since this site is a major
reservoir of NK-Hke cells in birds (64). If novel receptors are
identified on Xenopus NK candidates witb the anti-Xenoptis mAbs
generated, conservation of tbese receptors on mammalian NK
cells will be sought.
References
Du Pasquier L, Schwager J, Flajnik ME Theimmune system of Xenopus.Annu Rev Immunol 1989;7:2Sl-275.Chretien I. Robert J, Marcuz A.Garcia-Sanz JA. Courtet M, Du Pasquier LCTX, a novel molecule specifically expressedon the surface of cortical thymocytes inXmopus.Eur J Immunol 199fa;26:780-791.
3, Katevuo K, Gobel TF, Dunon D, Koskinen R,Imhof BA. Vaino O. ChT 1, a thymocyiemolecule ofthe igSF involved in colonizationof peripheral organs during development.ScanJImmunol 1996;43:694,
4. Harris D, ct al. A species-cunserved NK celiantigen receptor is a novel vimentin-likemolecule.Dev Comp Immimol I992;16:39S-4O3.
5. Horton ]D, Manning MJ. Response lo skinallografts in Xenopus laevis followingthymectomy at early stages of lymphoidorgan maturation.Transplantation 1972;14:141-154.
6. Florton JD. Thymectomy and transplantationin Xenopus. In: Lefkovits I, ed. Immunologymethods manual. London: Academic Press;1997. p. 2395-2406.
256 Immunoioflical Reviews 166/1998

Horton et al • T-cell and NK development in Tx Xenopus
7. Horton JD, Horton TL, Ritchie P hnmune
system of Xenopus: T cell biology. In: Kobel
HR, Tinsley RC. eds. The Biology of Xmopus.
Oxford: Oxford University Press; 1996.
p. 279-299.
8. Guy-Grand D, Cerf-Bensussan N, Mahssen B,
Malassis-Seris M, Briottei C, Vassalli R Two
gut intraepithelial CD8+ lymphocyte
populations with different T cell receptors: a
role for the gut epithelium in T cell
differentiation.
JExpMed 1 9 9 l ; i 7 3 : 4 7 1 ^ 8 l .
9. Guy-Grand D, Vassalli P. Gut intraepitlieiial
T lymphocytes.
Curr Opin Immunol 1993;5:247-252.
i 0, Rocha B, Guy-Grand D, Vassalli P Extrathymic
T cell differentiation,
Curr Opin Immunol 199S;7:235-242.
11. Poussier P, Julius M. Thymus independent
T cell development and selection in the
intestinal epithelium.
Annu Rev lmmnnol 1994;12:52 1-553.
12. MosleyRl, Klein JR. Peripheral engraftment of
fetal intestine into athymic mice sponsors
T cell development: direct evidence for
tiiymopoietic function of murine small
intestine.
JExpMed i992;i76:1365-1373.
13. Makino Y, et al, Extra-thymic development of
V-a-14-positive T-cells.
JExpMed 1993;177:1399-1408.
14. Wang J, Klein JR. Tliymus-neuroendocrine
interactions in extrathymic T cell
development.
Science l994;26S:i860-1862.
i 5, Lin T, Matsuzaki G, Kenai H, Nomoto K,
Extrathymic and thymic origin of murine IEL:
are most IEL in euthymic mice derived from
the thymus?
Immunol Cell Biol 1995;73:469-3-73,
! 6. Ilai T, et al. Ontogeny and development of
extrathymic T cells in mouse liver.
Immunology 1992;77:5S6-S63.
i 7. Ohteki T, et al. Age-dependent increase of
extratliymic T cells in the liver and their
appearance in the periphery of older mice.
Jlmmunol 1992;149:1562-1570.
18. Olusuka K, et al. Similaiities and differences
between extrathymic T ceils residing in mouse
liver and intestine.
Cell Immunol !994;153:52-66.
19. Poggi A, el al. Extratliymic differentiation of
T lymphocytes and natural killer cells from
human embryonic liver precursors.
Proc Natl Acad Sd USA 1993;90:4465-l-469.
20. MakinoY, Kanno R, Koseki H, Taniguclii M.
DeveiopmeniofV-a-14(+) NKT-celhin the
early stages ol embryo genesis.
Proc Natl Acad Sci USA ]996;93:6516-6520.
21. Garcia Ojeda ME, Dejbaklish Jones S,
Weissman IL, Strober S. An alternative pathway
for T cell development supported by the bone
marrow microenvironment: recapitulation of
thymic maturation.
JExpMed 1998;187:1813-1823.
22. Lake JP Pierce CW, Kennedy JD. T cell receptor
expression by T Cells that mature
extrathymically in nude mice.
Celi Immunol 199l;US;259-265
23. Lin T, Matsuzaki, G, Kenai H, Nakamiira T,
Nomoto K. Thymus influences the
development of extrathymically derived
intestinal intraepithelial lymphocytes.
EurJImmunoI 1993:23:1968-1974.
24. Ikehara S, et al. Thymic rudiments are
responsible for induction of functional T cells
in Du/nu mice.
Thymus 1987;10:193-20S.
2 5. Hein WR, Dudler L, Morris B. Differential
peripheral expansion and in vivo antigen
reactivity of ap and 78 T ceOs emigrating from
the early fetal lamb thymus.
EurJ Immunol 1990:20:1805-1813.
26. KasaharaY, Chen CH, Gobel TWF, Bucy RP,
Cooper MD. Intraepithelial lymphocytes in
birds. In: Kiyono H, McGhee JR, eds, Mucosal
immunology: intraepithelial lymphocyes.
New York: Raven Press: 1994. p, 163-174.
27. Chen CH, Bucy RP, Cooper MD.T cell
differentiation in birds.
Semin Immunol 1990;2:79-86.
28. Coltey M, et al. Analysis of the first two waves
of thymus homing stem cells and their T cell
progeny in chick-quail chimeras.
JExpMed 1989;I70:S43-S57,
29. Dunon D, Cooper MD, Imhof BA. Thymic
origin of embryonic intestinal 76 T cells.
JExpMed !993:177:2S7-263.
30. Abo T, et al, Extrathymic T ceils stand at an
intermediate phylogenetic position between
natural kiUer cells and diymus-derived T cells.
Natlmmun 1995:14:173-187,
31. Lefrancois L, Extratliymic differentiation of
ijitraepithelial lymphocytes: generation of
separate and unequal T-cell repertoires.
Immunol Today 1991:12:436-438,
32. Horton JD, Horton TL, Development of
transplantation immtmity and restoration
experiments in the thymectomized
amphibian,
AmZool 1975:15:73-84.
33. Turpen JB, Smith PB. Precursor immigration
and thymocyce succession during laii-al
development and metamorphosis in Xenopus,
Jlmmunol 1989:142:41^7.
34. Nagata S, T cell-specific antigen in Xenopus
identified with a mouse monoclonal antibody:
biochemical characterization and species
distribution.
ZoolSci 1988:5:77-83.
35, Nagata S, Development of T lymphocytes in
Xenopus laevis: appearance of tJie antigen
recognized by an anti-thymocyte mouse
monoclonal antibody
DevBiol 1986:114:389-394,
36. Gravenor I, Horton TL, Ritchie P Flint E,
Horton JD, Ontogeny and thymus-dependence
of T cell stixface antigens in Xenoptis: flow
cytometric studies on monodonal antibody-
stained thymus and spleen,
Dev Comp Immunol 1995:19:507-523.
3 7, Jturgens JB, Gartland LA, Du Pasquier L,
Horton JD, Gobel TWE, Cooper MD,
identification of a candidate CD5 homologue
in the amphibian Xenopus laevis,
Jlinmimol 1995:155:42 18--1-223,
38. IbrahimB,etal, Analysis of T cell development
in Xaiopus.
Fed Proc 1991:5:7651.
39. Flajnik MF, EerroneS, Cohen N, DuPasquierL.
Evolution of the MHC: antigenicity and
unusual tissue distribution of Xenopus (frog)
class 11 molecules,
Mol Immunol 1990:27:451-462.
40. Chretien I, Marcuz A, Fellah J, Charlemagne J,
Du Pasquier L. The T cell receptor p genes of
Xenopus,
EurJImmunoI 1997:27:763-771,
41. Cohen N, Effrige NJ, Parsons SCy Rollins-
Smith LA, Nagata S, Albright D, Identification
and treatment of a lethal nematode (Copillana
.fenopodis) infestation in the South African frog,
Xenopus laevis,
DevelComp Immunol 1984:8:739-741.
42. Weiss N, Horton JD, Dn Pasquier I. The effect
of thymectomy on cell surface associated and
serum immunoglobtilin in the toad Xenopu.s
laevis (Daudin): a possible inliibitory role of the
thymus on the expression of
immunoglobulins. In: Liacopouios P. Panijel J,
eds. CoUoque Inserm, Paris: Ministry of Public
Health: 1973, p, 165-174,
43. Gravenor I, Ontogeny, thymus dependence
and in vitro stimulation of lymphocyte subsets
in Xoiopus [PliD Thesis], Durham: Durliam
University: 1996,
44. Bleicher PA, Cohen N. Monoclonal anti-IgM
can separate T cell from B cell proiiferative
responses in the frog, Xenopus laevis.
Jlmmunol 198 1;127:1549-1 555,
45. Horton JD Amphibians. In: Turner RJ, ed.
Immunology: a comparative approach,
Cliichester: Wiley: 1994, p, 101-135.
46. Katagiri C, Tochinai S. Ontogeny of thymus-
dependent immtuie responses and lymphoid
cell differentiation in Xenopus laevis.
Dev Grow[h Differ 1987;29:297-305,
Immunological Reviews 166/1998 257

Horton et al • T-cell and NK development in Tx Xenopus
47, Du Pasquier L, Horton JD, The effect of
thymectomy on the mixed leucocyte reaction
andphytohemaggiutinin reponsiveness in the
clawed toad Xenopus laevis.
Immtinogenetics 1976:3:105-112.
48, Du Pasquier L, Horton JD. Restoration of
antibody responsiveness in early
thymectomized Xaiopus by implantation of
major liistocompatibility comp 1 ex-
mismatched larval thymus.
EurJImmunoI 1982:12:546-551,
49, Horton JD, Sherif NEHS. Sequential
ih)inectomy in the clawed toad: effect on
mixed leucocyte reactivity and
phytoliaemagglutinin responsiveness. In:
Solomon JB, Horton JD, eds. Developmental
immunology Amsterdam: Elsevier: 1977, p,
283-290,
50, Manning MJ, Collie MH, The ontogeny of
thymic dependence in the amphibian Xenopus
iaevi.?. In: Solomon JB, Horlon JD, eds.
Developmental immunology. Amsterdam:
Elsevier; 1977, p, 291-298.
51, Turner SL, Horton TL, Ritchie R Horton ]D.
Splenocyte response to T cell-derived
cytokines in thymectomized Xenopus,
Dev Comp Immunol l991;I5:319-328.
52, Nagata S, Cohen N, Specific in vivo and non
specific in viiro alloreactivities of adult frogs
(Xenopus Icevis) that were thymectomized during
early larval life.
EurJImmunoI 1983:13:541-545,
53, Manning MJ, Donnelly N, Cohen N,
Thymus-dependent and thymus-independent
components of the amphibian immune
system. In: Wright RK, Cooper EL, eds.
Phylogeny of thymus and bone marrow- bursa
cells. Amsterdam: Elsevier; 1976. p. 123—132.
54, Horton p , Horton TL, Ritchie P. Gravenor I,
Horsham K. Immune system development in
Xenopus. In: Stolen JS, et al. eds. Modulators of
immune responses: the evolutionary trail. Fair
Haven: SOS Publications USA: 1996. p.
131-147.
55, Rast JP, Anderson MK, Strong SJ, Luer C,
Litman RT, Liiman GW a, p, v. and 5 T cell
antigen receptor genes arose early in vertebrate
evolution.
Immunity 1997:6:1-11.
56, Valiante NM, Parham P NK cells and CTL:
opposite sides of the same coin. In: MorettaL,
ed. Molecular basis of NK cell recognition and
function, Basel: Karger: 1996, p. 146-163,
57, Kos FJ, Engleman EC, Immune regulation: a
critical link between NK cells and CTLs,
Immunol Today 1996;17:174-176,
58, Yokoyama WM. Natural killer cell receptors
Curr Opin Immunol 1998:10:298-305,
59, Raulet DH, Recognition events that inhibit and
activate natural killer cells,
Curr Opin Immunol 1996;8:372-377.
60, Lanier LL, Natural killer cell receptors and
MHC class I interactions,
Curr Opin Imnnunol !997;9:126-131,
61. LongEO, WagtmannN. Natural killer ceil
receptors,
Curr Opin Immunol ]997;9:344-350,
62. Parham P, ed. NK cells, MHC class I antigens
and missing self,
hnmunolRev 1997:155:1-221,
63. Janeway CA. Natural kiUer cells: a primitive
immtme system.
Nature 1989:341:108.
64. Gobel TWF, et al. Characterisation of avian
natural kilter cells and their intracellular CD3
protein complex,
Eiir J Immunol 1994;24:1685-1691,
65. Brooks CG, Georgiou A, Jordan RK, The
majority of immature fetal thymocyces can be
induced to proliferate to lL-2 and differentiate
into cells indistinguishable from mature
natural kiUer cells,
Jlmmtino! !993;]2:664.S-6656,
66. Hogan RJ, Stuge TB, Clem LW, Miller NW
Cbinchar VG, Anti-viral cytotoxic cells in the
channel catfish {Ictalurus punctatus),
Dev Comp Immunol 1996:20:115-1 27.
67. McKinney EC, Schmaie MC. Damselfish with
neurofibromatosis exliibit cytotoxicity
towards retrovirus infected cells,
Dev Comp Immunol 1997;21:287-298.
68. Trede NS, Zon LI. Development of T cells
during fish embryogenesis.
Devel Comp Immunol 1998:22:253-263.
69. GhoneumM,CooperEL, Sadekl. Variability of
natural killer cell activity in anuran
amphibians.
Dev Comp Immunol 1990;14:3S9-36S.
70. Marsh J, Goode JA, eds. Antimicrobial
peptides, Ciba foundation symposium: 186,
Chichester: Wiley; 1995.
71. Horton TL, Ritchie P Watson MD, Horton JD.
NK-like activity against allogeneic tumour
cells demonstrated in the spleen of control and
thymectomized Xenopus.
Immunol Cell Biol 1996:74:365-373,
72. Horton TL, Ritchie P Watson MD, Horton JD.
NK cell evolution: studies on Xenopus.
Biochem Soc Trans 1997;2S:263S.
73. Robert J, Guiet C, Du Pasquier L. Lymphoid
tumours of Xenopus iaevis with different
capacities for growth in larvae and adults.
Dev Immunol 1994:3:297-307.
74. Haynes L, Cohen N, Further characterization
of an interleukin-2 like cytokine produced by
Xenopu.s laevis T lymphocytes.
Dev Immunol 1993;3:23 1-238,
75. Brooks CG, Urdal DL, Henney CS,
Lymphokine-driven "differentiation" of
cytotoxic T-cell dones into cells with NK-like
specificity: correlations with display of
membrane macromolecules,
Immunol Rev 1983;72:43-72,
76. Waikins, D, T ceU function in )toopus: studies
on T cell ontogeny and cytotoxicity using an
IL-2 like growth factor [PhD. Tliesis]. New
York: University of Rochester, 1985.
7 7. Saxena QB, Saxena RK, Goldfarb RH,
Herberman RB, Whiteside, TL. Identification
and partial puriflcation of a human natural
kiiler cell prohferation-inducing factor
JBiosci 1996:21:455-469.
78. Horton TL, Ritchie P Watson MD, Horton JD.
Natural cytotoxicity towards allogeneic
[umour targets in Xenopus mediated by diverse
splenocyte populations.
Dev Comp Immimol 1998:22:217-230.
79. Bernard CCA, Bordmann G, Blomberg B,
Du Pasquier L. Immunogenetic studies on the
cell-mediated cytotoxicity in the clawed toad
Xenopus laevis.
Immunogenetics 1979:9:443^54.
80. Robert J, Guiet C, Cohen N. Du Pasquier L.
Effects of thymectomy and tolerance induction
on tumor immunity in adult Xenopus laecis.
IntJCancer 1997:70:330-334,
81. Aposov SD, Redegeld F, Sitkovsky M,
Cell-mediated cytotoxicity: contact and
secreted factors,
Curr Opin Immunol ]993;S:404-410,
82. Squier MKT, Cohen JJ, Cell-mediated cytotoxic
mecbanisms,
Curr Opin Immunol 1994;6:447^52,
83. Robert J, Guiet C, Du Pasqtiier L. Ontogeny of
the alloimmune response against a
transplanted tumor in Xenopus laevis.
Differentiation l995;59:135-]44.
84. Flajnik M, Kaufman JF, Hsu H, Manes M,
Parisot R, Du Pasquier L, Major
histocompatibihty complex encoded class II,
but not class I, molecules are present in
immunologically competent Xenopus before
metamorpbosis.
] Immunol 1986:137:3891-3899.
85. Winter G, Griffiths AD, Hawkins RE,
Hoogenboom HR, Making antibodies by
phage display technology
Annu Rev Immunol 1994;12:433-4-55.
86. Hayden MS. Gilliland LK, Ledbetter JA.
Antibody engineering.
Curr Opin Immunol 1997:9:201-212.
87. Chen CH, Ager LL, Gardand GL, Cooper MD,
Identification of a T3/T cell receptor complex
in chickens.
JExpMed 1986:164:37.5-380.
88. Lefrancois L Isolation of mouse small
intestinal intraepithelial lymphocytes.
Current Protocols Immimol
1993;1:19.]-19.7.
89. Langeberg L, Ruben LN, Clothier RH, Shiigi S.
The characterization of the toad splenocytes
which bind mouse anti-human IL-2 receptor
antibody
Immunol Lett 1987:16:43-l-8.
90. Hsu E, Lefkovits J, Flajnik M, Du Pasquier L,
Light chain heterogeneity in the ampliibian
Xenopus,
Mol Immunol 1991:28:985-994.
258 Immunological Reviews 166/1998






























