Embed Size (px)
Citation preview

UNIVERSIDAD NACIONAL AUTÓNOMA DE MÉXICO
POSGRADO EN CIENCIAS BIOLÓGICAS
FACULTAD DE MEDICINA BIOMEDICINA
POSIBLE EMPLEO DE PÉPTIDOS MIMOTOPOS OBTENIDOS POR PHAGE DISPLAY DEL
LPS DE E. coli O157 EN EL DISEÑO DE VACUNAS CONTRA LA BACTERIA
TESIS
QUE PARA OPTAR POR EL GRADO DE:
DOCTOR EN CIENCIAS
PRESENTA:
NAVARRO OCAÑA ARMANDO
TUTOR PRINCIPAL DE TESIS: DR. CARLOS ALBERTO ESLAVA CAMPOS. FACULTAD DE MEDICINA, UNAM.
COMITÉ TUTOR: DR. EDGAR ARTURO ZENTENO GALINDO. FACULTAD DE MEDICINA, UNAM.
DRA. BERTHA GONZÁLEZ PEDRAJO. INSTITUTO DE FISIOLOGÍA CELULAR, UNAM.
MÉXICO, CD. MX., DICIEMBRE, 2016.

Uo. 1""" .. Rom;,., W. n<o 0;'0<1010 (¡""o ral do Adminft.tr.oi<>n EOCOlar, UN.OM Pr ... nl.
Me p«fr.'" nformar ~ "tC.O qo.;o '" Subcornt<' jo Ik>log i. Expcri".ont.>l 1 B"""-'<l~ ", ~ P'Oo¡¡raDO "" C. """ .. Biol<:~~"" .n su S,.,," oroino,", <lrl dio 15:. loe,.ro do 2016, oprobó 01 jlrodo ,.,,~ L> "' .... 0'*' 00 do .", 'r on p.ae;¡ ""' ....... 01 9"'00 ,~ DOCTOR EN CIENCIAS ,101
. Jre"" NAVARRO OCAflA ARMANDO """ nirle!O do o""ota U206U9. 00' l. " ... "",,1,,,. ·POSI8LE EMPLEO OE PEpnoos MIMOTOPOli 08TENlDOS POR PHA(jE DISPLAY DEL LP5 DE E, ~o'i O'~1 EN EL D'SEÑO DE VA.CUNA5 CONTRI, LA. BACTERIA. , "'. '_ . boJO la dI'oceiOo 0.4 DR. CARLOS AL8E!110 ESLAVA Ci\MPOS:
P",sidon' o:
"~ Stcretor",
Sv,ole"""
S"" lente
DRA YO!.ANOA LÓP EZ IIICA.L
DRA ARIAOtI'A DEl- CARMEN CRUZ C6RDQ'/A
DRA LOUROES . ..,.00,,/\ ARRLI\(l/\ PtZNIO
DR C=SAA RA.Ul GC~lAlEl60"" ' "
DR EDGAR /lRTtJRO lENTENO GALI\oo
ATENTAMENTE · PO~ MI R.lZA HA6LA"A EL EspíA'TU"
Cd \J"i, .,oitW, C~, ~',,, , 13 00 X tl. .,.. eo 2,)1~
~ Mc.,~ DRA. MARiA OEL CORO ARIZMENOI ARRIAGA
COORDINADORA OH PROGRAMA

AGRADECIMIENTOS
Agradezco muy especialmente al Programa Académico del Posgrado en Ciencias Biológicas de la Universidad Nacional Autónoma de México (UNAM) por la formación académica recibida durante el desarrollo del Posgrado.
Al Fondo Sectorial de Investigación para la Educación SEP-CONACYT, Ciencia Básica, Proyecto CB-2012-01/179000.
Con mucha gratitud y afecto a mi Comité tutor: Dr. Carlos Alberto Eslava Campos, Dr. Edgar Arturo Zenteno Galindo y Dra. Bertha González Pedrajo, por su apoyo y orientación en el desarrollo del Proyecto tesis.
A los miembros del Jurado: Dra. Yolanda López Vidal, Dra. Ariadnna del Carmen Cruz Córdova, Dra. Lourdes Andrea Arriaga Pizano, Dr. Cesar Raúl González Bonilla y Dr. Edgar Arturo Zenteno Galindo, por su valiosa revisión y recomendaciones al manuscrito de tesis.

DEDICATORIA
A mis hijos, Armando y Rodrigo, muy especialmente a Irma por su comprensión y apoyo incondicional durante el tiempo en que desarrollé el Posgrado en Ciencias Biológicas.
Agradezco a Delia Licona Moreno, Gabriel Pérez y Luis León Alamilla por su apoyo técnico en el desarrollo de mi Proyecto de tesis.

ÍNDICE
ÍNDICE DE CUADROS Y FIGURAS.......................................................................................... i
RESUMEN ................................................................................................................................ 1
ABSTRACT ............................................................................................................................... 2
1. INTRODUCCCIÓN ............................................................................................................... 3
1.1 Importancia de las enfermedades diarreicas en México. .................................................... 3 1.2 Agentes etiológicos de la diarrea: Escherichia coli. ............................................................ 4 1.3 E. coli diarreogénica (DEC). ............................................................................................... 5 1.4 E. coli productora de la toxina Shiga (STEC). ..................................................................... 6 1.5 Prevalencia de E. coli O157:H7 en México. ........................................................................ 8 1.6 Estructura del LPS de E. coli O157 y epitopos comunes con otras enterobacterias. ......... 9 1.7 Respuesta inmune contra el LPS O157 en población expuesta y no expuesta. ............... 10 1.8 E. coli O157:H7 en población mexicana y respuesta inmune. .......................................... 11 1.9 Vacunas contra E. coli O157. ........................................................................................... 12 1.10 Phage display: usos del método. .................................................................................... 16 2. JUSTIFICACIÓN ................................................................................................................. 17
3. OBJETIVOS ........................................................................................................................ 18
Objetivo general ...................................................................................................................... 18 Objetivos particulares ............................................................................................................. 18 4. MATERIAL Y MÉTODOS ................................................................................................... 19
4.1 Extracción de lipopolisacáridos ......................................................................................... 19 4.2 Sueros anti-LPS O157 ...................................................................................................... 19 4.3 Obtención de inmunoglobulinas (IgG) ............................................................................... 20 4.4 Selección de mimotopos ................................................................................................... 20 4.5 Secuenciación de DNA ..................................................................................................... 21 4.6 Péptidos sintéticos ............................................................................................................ 22 4.7 Suero anti-fagotopos y anti-péptidos sintéticos ................................................................ 22 4.8 IgG anti-fagotopo .............................................................................................................. 23 4.9 Respuesta de sueros anti-péptido .................................................................................... 24 4.10 Ensayos de competencia ................................................................................................ 24 4.11 Sitio que mimetiza SP12 en el LPS O157 ....................................................................... 25 4.12 Reactividad de sueros humanos contra SP12 ................................................................ 26 4.13 Análisis estadístico ......................................................................................................... 26 5. RESULTADOS ................................................................................................................... 27
5.1 Anticuerpos anti-LPS O157 .............................................................................................. 27 5.2 Selección de mimotopos ................................................................................................... 28 5.3 Análisis de péptidos .......................................................................................................... 28 5.4 Reactividad de IgG anti-fagotopos .................................................................................... 30 5.5 Inmunogenicidad de péptidos sintéticos ........................................................................... 31 5.6 Antigenicidad del péptido SP12 ........................................................................................ 32 5.7 Sitio en el LPS O157 en el que SP12 es mimotopo .......................................................... 33

5.8 Inhibición competitiva ........................................................................................................ 34 5.9 Respuesta de sueros humanos contra el péptido SP12 ................................................... 36 6. DISCUSIÓN ........................................................................................................................ 37
7. CONCLUSIONES ............................................................................................................... 44
8. PERSPECTIVAS ................................................................................................................ 45
9. LITERATURA CITADA ....................................................................................................... 46
10. ANEXO. ARTÍCULO REQUISITO..................................................................................... 54

i
ÍNDICE DE CUADROS Y FIGURAS
Figura 1 Reactividad del suero de conejo anti-LPS O157 contra el LPS
O157 27
Cuadro1 Clonas de fagotopos seleccionados con su secuencia de aminoácidos y reconocimiento por IgG anti-LPS O157
29
Cuadro 2 Las IgGs anti-fagotopos reconocieron las clonas de fagos homólogos y péptidos sintéticos
30
Figura 2 Reconocimiento de péptidos mimotopos y LPS O157 por el suero anti-LPS O157
31
Figura 3 SP12 es un mimotopo inmunogénico de una fracción del LPS de E. coli O157
32
Figura 4 Inmunoelectrotransferencia (Western blot) del LPS O157 purificado
33
Figura 5 SP12 bloquea los sitios de unión de los anticuerpos anti-LPS O157 34 Figura 6 El LPS O157 bloquea los sitios de unión de los anticuerpos anti-
SP12 35
Figura 7 SP12 fue reconocido por muestras de sueros humanos sanos de población abierta
36

1
RESUMEN
INTRODUCCIÓN. Escherichia coli O157:H7 es un patógeno de importancia clínica y epidemiológica en varios países. El lipopolisacárido (LPS) es un componente que participa en la patogenicidad de la bacteria y activa la respuesta inmune, sin embargo, debido a su efecto tóxico no es un buen candidato para el desarrollo de vacunas. OBJETIVO. Identificar por el procedimiento de Phage display mimotopos de epitopos inmunodominantes del LPS O157 y evaluar su capacidad inmunogénica para considerar su empleo como vacunas. MATERIAL Y MÉTODOS. Con IgGs anti-LPS O157 se seleccionaron fagotopos (fago M13) que expresan péptidos mimotopos. Las secuencias de DNA de los fagotopos se tradujeron a aminoácidos para generar péptidos sintéticos (SP), con ambos fagotopos y péptidos sintéticos por separado se inmunizaron conejos. La respuesta de anticuerpos se evaluó por ELISA contra el LPS O157 y los péptidos. Para conocer la especificidad del péptido mimotopo, se realizó inhibición competitiva y se analizó la respuesta de los sueros anti-SP12 y anti-LPS O157. El sitio de ubicación del mimotopo se definió por la interacción del suero anti-péptido con el LPS O157 en un ensayo de Western-blot. Además, se evaluó la respuesta de muestras de suero humano contra el péptido mimotopo identificado. RESULTADOS. Se obtuvieron 63 fagotopos, 15 reaccionaron con el suero anti-LPS O157, en cinco de ellos (Fgtp12, Fgtp13, Fgtp5, Fgtp14 y Fgtp16) se identificó el motivo consenso LxP/YP/SxL. Se obtuvo la síntesis de los péptidos SP5, SP12, SP16, SP11 y SP3, los dos últimos con motivo diferente a los previos. El suero anti-SP12 (SLSPPLSGLGRA) reconoció tanto al péptido homólogo (1:800) como al LPS O157 (1:200). Así mismo el suero anti-LPS O157 reaccionó con SP12 (1:200) pero no con SP3 (control de especificidad). El ensayo de competencia, mostró disminución significativa de los valores de D.O. en la respuesta del suero anti-LPS O157 (previa incubación con SP12) contra el antígeno homólogo. En el análisis de la ubicación del mimotopo evaluado por inmunotransferencia, los sueros anti-SP12 y anti-LPS O157 reconocieron una región localizada entre 35 y 50 kDa del LPS O157 que corresponde a las cadenas laterales del LPS O157. El ensayo con los sueros humanos que reaccionan con el LPS O157, mostró que 18 (82 %) de 22 reconocieron el SP12. Conclusión. Se seleccionó un péptido mimotopo (SP12) que corresponde a un epitopo inmunodominante del LPS O157. El péptido SP12 mostró capacidad inmunogénica y antigénica por lo que pudiera ser un buen candidato para el posible desarrollo de una vacuna contra E. coli O157.

2
ABSTRACT
POSSIBLE USE OF MIMOTOPE PEPTIDES OBTAINED BY PHAGE DISPLAY FROM LPS OF E. coli O157 IN THE DESIGN OF VACCINES AGAINST BACTERIA INTRODUCTION. Escherichia coli O157:H7 is a pathogen of clinical and epidemiological importance in various countries. Lipopolysaccharide (LPS) is one of the components involved in the bacterial pathogenicity. Despite being an immunogenic molecule due to its toxic activity, its use in the vaccine development is not recommended. OBJECTIVE. To use Phage display method to identify immunodominant epitopes (mimotopes) from O157 LPS and to evaluate their immunogenicity to propose their possible use in the development of vaccines. MATERIAL AND METHODS. Using IgG anti-O157 LPS and biopanning M13 filamentous phage clones were selected. With phagotopes (phage clones) and synthetic peptides (conjugated to KLH) were immunized rabbits. To obtain anti-phage IgG, rabbits were immunized with the selected phagotopes and by ELISA test, the IgG were evaluated to know their reactivity against the phage clones and synthetic peptides (SP). A competition assay was performed to confirm if the synthetic peptides were mimotopes of O157 LPS. To know the possible location of the peptide mimotope on the LPS, anti-SP12 serum reactivity was analyzed by immunoblotting, and as positive control, anti-O157 LPS serum was used. Finally, the response against the SP12 of human sera samples with reactivity against O157 LPS was evaluated by ELISA assay. RESULTS. Fifteen phagotopes out of 63 selected with anti-O157 LPS sera showed a high positive response against the anti-O157 LPS serum. In five of these (Fgtp12, Fgtp13, Fgtp5, Fgtp14 and Fgtp16), the consensus motive LxP/YP/SxL was identified. With the derived amino acid sequences obtained from the phagotopes DNA nucleotides, the synthetic peptides SP5, SP12 and SP16 were obtained. Additionally, two more peptides (SP3, SP11) that correspond to phagotopes with a different motif were synthesized. The rabbit serum obtained against SP12 with the amino acid sequence SLSPPLSGLGRA showed a positive response against the homologous peptide (1:800) and O157 LPS (1:200). Similar results were obtained with the anti-O157 LPS serum (1:200) that reacted with the SP12 but not with the SP3 peptide with a different sequence. The competition assay, showed a significant decrease in the ELISA OD values of the anti-O157 LPS serum response against the O157 LPS previously incubated with SP12. The immunoblotting assay on the O157 LPS, showed that anti-SP12 serum as well as anti-O157 LPS serum reacted with a region located between 35 and 50 kDa. The carbohydrate repeating units of O157 LPS are located in this molecular weight range, suggesting that this region corresponds to mimotope. The response of human sera samples with positive reaction against O157 LPS showed that 18 (82 %) out of 22 samples reacted with SP12. CONCLUSION. Phage display method allowed the selection of a mimotope peptide (SP12) that corresponds to an immunodominant epitope of O157 LPS located on carbohydrate repeating units. Although further studies are required, SP12 could be considered an immunogenic peptide possible candidate to vaccine development against E. coli O157.

3
1. INTRODUCCCIÓN
1.1 Importancia de las enfermedades diarreicas en México.
La diarrea se define como el incremento en el número de evacuaciones y la disminución
de la consistencia de las mismas. Se puede presentar en tres tipos clínicos; diarrea aguda
con evacuaciones líquidas (varias horas o días), aguda con sangre (también llamada
disentería) y persistente cuya duración es de 14 o más días [1]. Una estimación a nivel
mundial en 1990 sobre la diarrea en niños menores de cinco años que viven en países
en vías de desarrollo reportó, 1,900 millones de episodios y en 2010 aproximadamente
1700 millones [2], causando anualmente entre 1.6-2.5 millones de muertes en estas
regiones [3]. Aunque se ha observado que la media estimada de muertes a consecuencia
de este padecimiento ha descendido de 4.6 millones en 1982 a 3.0 millones en 1992 y a
2.5 millones en 2000, la diarrea es causa de alrededor del 21 % de todas las muertes
registradas en este grupo de edad [3].
En México, de acuerdo a datos publicados por la Dirección General de Epidemiología [4],
en el 2001 registraron 6.9 millones de casos de diarrea en niños menores de cinco años,
en 2007, 5.45 millones y en 2011, 6.03 millones de casos, lo que significó una incidencia
de 5521 casos por 100, 000 habitantes y un aumento de 10.6 % respecto a 2007. Datos
de 2014 en niños menores de cuatro años, reportan una incidencia de 10356.3 por 100
000 habitantes (SUIVE/DGR/Secretaria de Salud), siendo niños de familias con bajo nivel
de ingresos económicos de áreas rurales los más afectados por este padecimiento.
Aunque la reducción de las tasas de mortalidad y morbilidad de las enfermedades
diarreicas en niños continúa siendo un pendiente para este importante problema de salud
pública.

4
1.2 Agentes etiológicos de la diarrea: Escherichia coli.
Se conoce que entre los agentes etiológicos relacionados con las infecciones intestinales
se encuentran virus, parásitos y bacterias. Las bacterias Vibrio cholerae y algunos
géneros de la familia Enterobacteriaceae son los principales microorganismos
involucrados en estas infecciones. La familia Enterobacteriaceae está integrada por
varios géneros entre los que se incluyen Salmonella, Escherichia, Shigella, Proteus,
Enterobacter, entre otros [5]. Desde el punto de vista microbiológico las enterobacterias
se caracterizan por no formar esporas, son anaerobios facultativos, fermentan la glucosa,
no producen oxidasa y tienen movilidad variable, lo que depende de la presencia o no de
flagelos.
El género Escherichia es uno de los principales agentes causantes de diarrea, este lo
constituyen las especies: E. coli, E. albertii, E. blattae, E. fergusonii, E. hermannii y E.
vulneris [5]. La descripción microbiológica de E. coli tiene sus antecedentes desde 1885,
con el pediatra alemán Theodor Escherich quién trabajando en Austria con muestras de
heces de niños aisló a la bacteria que describió como bacilos delgados y ligeramente
curvos los cuales tenían la capacidad para crecer en medios artificiales. Escherich llamó
a este microorganismo Bacterium coli commune, posteriormente Castelani y Chalmer la
llamaron Escherichia coli en honor a Escherich [6]. En 1889, Laurelle sugirió una
asociación entre Bacterium coli commune con la presencia de diarrea y vómito, once años
después Le Sage demostró que muestras de suero de pacientes con diarrea aguda
aglutinaban a los microorganismos aislados de pacientes de brotes epidémicos, sin que
la reacción fuera observada con las bacterias aisladas de niños sanos. A partir de esa
época se empezaron a realizar esfuerzos para diferenciar cepas de E. coli patógenas y
no patógenas. Uno de los primeros intentos para caracterizar a E. coli aislada de caso de

5
gastroenteritis fue el realizado por Adam en Alemania en 1923 [6] quien comenzó a utilizar
métodos serológicos, además mediante pruebas bioquímicas logró diferenciar grupos de
E. coli. En la década siguiente, en 1933, Goldschmidt continuó con el trabajo de Adam y
desarrollo la tipificación serológica por aglutinación en laminilla del grupo “dyspepsiekoli”.
Mediante esta técnica, Goldschmidt realizó estudios de epidemiología de gastroenteritis
infantil y señaló la importancia del portador sano de cepas con propiedades epidémicas.
En la década siguiente, Kauffmann en 1944, propuso por primera vez un esquema de
tipificación por métodos serológicos [7]. En el primer esquema antigénico publicado por
Kauffmann se describieron 20 variedades del antígeno O (serogrupos), para tal propósito
utilizó suspensiones preparadas con bacterias hervidas. Un año después Knipschildt [8]
adicionó al esquema cinco nuevos grupos O y en 1947 Kauffmann publicó la primera
ampliación al esquema, el cual comprendía 25 antígenos O, 55 antígenos K (capsular) y
19 antígenos H [9]. En la actualidad el Centro Internacional de Escherichia coli y
Klebsiella, en Copehnague, Dinamarca reconoce 187 variedades del antígeno O [10].
1.3 E. coli diarreogénica (DEC).
Los primeros estudios que relacionaron a E. coli con un evento diarreico, utilizaron como
elementos de diagnóstico el suero de los pacientes y las cepas de E. coli aislada de las
heces de los mismos pacientes, los resultados obtenidos mostraron que las cepas
reaccionaban con el suero de los pacientes, lo que demostró que estas cepas eran las
responsables de la enfermedad. Diferentes estudios mostraron que la mayoría de los
casos de diarrea en niños, sobre todo en los brotes eran causados por un grupo reducido
de serogrupos de E. coli. Estos grupos fueron denominados por Neter [11], como
enteropatógenos y propuso el termino de E. coli enteropatógena (EPEC) para referirse a
varios serogrupos y serotipos de E. coli asociadas epidemiológicamente con infecciones

6
intestinales. Posteriormente Levine [12] propuso la clasificación de E. coli enteropátogena
en diferentes grupos diarreogénicos (DEC) de acuerdo a sus propiedades de virulencia y
cuadro clínico que producían, inicialmente se propusieron cinco grupos diarreogénicos
de E. coli, enteropatógena (EPEC), E. coli enteroinvasiva (EIEC), E. coli enterotoxigénica
(ETEC), E. coli enterohemorrágica (EHEC) y E. coli enteroagregativa (EAggEC). Más
adelante se reconocieron seis grupos diarreogénicos denominados como patotipos, en
estos grupos, EHEC se propuso como un subtipo de E. coli productora de la toxina Shiga
(STEC), además de los patotipos mencionados se agregó a E. coli con adherencia difusa
(DAEC) [13, 14]. Sin embargo, recientemente nuevas formas híbridas de la bacteria han
emergido, por lo que dos nuevos grupos se incluyeron en los grupos DEC; E. coli
adherente invasiva (AIEC) asociada a la enfermedad de Crohn y E. coli
enteroagregativa/productora de la toxina Shiga (STEAEC) responsable de un brote de
colitis hemorrágica (CH) y del síndrome urémico hemolítico (SUH) en Alemania durante
2011 [15].
1.4 E. coli productora de la toxina Shiga (STEC).
A principios de la década ochenta del siglo pasado, en los Estados Unidos y Canadá se
describieron brotes de CH y SUH asociada a E. coli del serotipo O157:H7 [16, 17]. Riley,
[16] reportó la participación de la mencionada bacteria en brotes de enfermedad
gastrointestinal caracterizada por dolor abdominal severo sin fiebre acompañado de
diarrea acuosa con sangre, a la cual llamaron colitis hemorrágica. Simultáneamente
Karmali [17], reportó la participación del mismo microorganismo en casos aislados de
SUH, padecimiento que se caracteriza por daño renal agudo, trombocitopenia y anemia
hemolítica microangiopática.

7
Posteriormente a partir los brotes que se presentaron de CH y SHU en otros países como
Alemania, Francia, Austria y Japón, entre otros, además de los Estados Unidos y Canadá
fue que se le reconoció como un patógeno emergente de importancia clínica y
epidemiológica en países industrializados [18, 19, 20]. A través de diferentes estudios
epidemiológicos ha quedado establecido que los bovinos, ovinos, venados, cerdos,
caprinos, entre otros son los reservorios naturales de E. coli O157:H7 [21, 22, 23, 24]. Su
transmisión a humanos se realiza a través del consumo de carne mal cocida, de leche
sin pasteurizar y de la ingesta de germinados de alfalfa, y vegetales regados con agua
contaminada con heces de estos animales [25, 26, 27, 28, 29].
E. coli O157 posee diferentes factores de virulencia, uno de los cuales consiste en la
producción de diferentes variedades de la toxina conocida como toxina Shiga (Stx). A E.
coli O157:H7 además se le conoce también como E. coli productora de la toxina Vero
(VTEC) por el efecto citopático en células de riñón de mono verde [30]. La familia de
toxinas tipo Shiga contiene principalmente las toxinas Stx1 y Stx2, estas dos toxinas
difieren en su secuencia de aminoácidos por lo que son antigénicamente diferentes.
Dichas toxinas son codificadas por bacteriófagos lisogénicos insertados en el cromosoma
de la bacteria. La toxina Stx1 es similar a la toxina Shiga de Shigella dysenteriae tipo 1,
en la actualidad se conocen las variantes stx/vtx1a, vtx1c, vtx1d de ésta toxina. Con
relación a Stx2 se conocen las variantes Stx2a, Stx2b, Stx2c, Stx2d, Stx2e, Stx2f y Stx2g,
aunque, la nomenclatura es intercambiable cuando se utiliza el término de Verotoxina
vtx2: vtx2a, vtx2b, vtx2c, vtx2d, vtx2e, vtx2f and vtx2g [31]. De estas toxinas el tipo Stx2
es el que se ha asociado con la producción del SUH.

8
1.5 Prevalencia de E. coli O157:H7 en México.
En México en un estudio realizado por Valdivia G, [3rd International Symposium and
Workshop on Shiga toxin VTEC 1997 abstr. V167/IV] reportó un brote de diarrea con
sangre en becerros. Las cepas obtenidas en dicho brote fueron caracterizadas por
tipificación con sueros en el laboratorio de Salud Pública de la Facultad de Medicina de
la UNAM, el resultado mostró que eran cepas de E. coli del serotipo O157:NM (no móvil)
y que una de las cepas era portadora de los genes que se han relacionado con la
virulencia de esta bacteria. Callaway [32] por su parte, en colaboración con la USDA
(College Station, Texas, USA), estudiaron asilamientos de cepas de E. coli de muestras
fecales de 240 bovinos y 240 de cerdos de cuatro diferentes ranchos situados en la región
central de México. La frecuencia de aislamiento de E. coli O157:NM fue de 1.25% en
bovinos y en cerdos de 2.1%. En un estudio realizado por Amezquita-López [33] en
animales domésticos entre los que se incluyeron bovinos, carneros y pollos del Valle de
Culiacán, México, se reportó que E. coli O157:H7 fue aislada en el 5.4 % de 240 muestras
de heces de los animales, las cepas se caracterizaron por presentar los genes stx2, eae
y ehxA. En otros estudios realizados en canales de reses en rastros de Guadalajara Jal.,
y Toluca, Estado de México [34, 35], realizaron el aislamiento de cepas de E. coli
O157:H7. En el estudio de Varela-Hernández [34] fueron aisladas cepas de E. coli
O157:NM y O157:H7 en una frecuencia de 5 % y 2.7 % respectivamente; Con respecto
al estudio realizado por Reyes-Rodríguez [35] de 37 cepas analizadas obtenidas de
canales de reses encontraron 6 (16 %) cepas de E. coli O157:H7. Por otro lado, en un
estudio de colonización intestinal por E. coli en un hato de vacas lecheras del estado de
Jalisco, se identificó la presencia de E. coli O157:H7 en dos (4 %) de un hato de 46 vacas
[36]. Aunque, estos estudios muestran la presencia de E. coli O157:H7 en bovinos de

9
diferentes zonas geográficas del país, hasta el momento no se ha identificado que esta
bacteria afecte la población mexicana.
1.6 Estructura del LPS de E. coli O157 y epitopos comunes con otras
enterobacterias.
El lipopolisacárido (LPS) de E. coli está formado por tres regiones, la primera con
características hidrofóbicas incluye el lípido A, la región central o core está compuesta
por oligosacáridos (OS) y la tercera que se conoce como antígeno O (somático). El lípido
A es una región conservada entre las enterobacterias, está integrado por un disacárido
de N-acetil-glucosamina unido por un enlace 1-6 y fosforilado en las posiciones 1 y 4´.
Este disacárido contiene seis cadenas de ácidos grasos, de los cuales 4 están acilados
directamente a los residuos de glucosamina en las posiciones 2, 3, 2´ y 3´, y los otros son
cadenas cortas de laurato y miristato esterificadas en las cadenas 2´ y 3´ [37]. El lípido A
es una molécula de aproximadamente 1200 Da, se menciona que es un potente activador
de la inflamación y relacionado con el choque endotóxico [38]. En cuanto a la región del
core se divide en dos subregiones estructurales, el interno compuesto por heptosas y por
el ácido 3-deoxy-D-manno-octulosónico (KDO), también conocido como el ácido 2-ceto-
3-deoxyoctónico y la región externa compuesta principalmente de hexosas y
carbohidratos amino-acetilados. En el caso de E. coli se reconocen cinco tipos de core
externo que se han designado como R1, R2, R3, R4 y K12, cada uno de ellos con una
composición diferente de carbohidratos [39, 40]. La tercera región se conoce como
antígeno somático (O) también referida como cadenas laterales, su composición es
principalmente de carbohidratos y está relacionada con la diversidad del LPS de E. coli,
además de que es una región que proporciona una superficie hidrofílica a la pared celular
de los microorganismos [41, 42]. En su estructura ésta molécula presenta más de un

10
carbohidrato en forma repetida, por lo que se les denomina unidades repetidas de
carbohidratos.
Las unidades repetidas del LPS de E. coli O157 es un tretrasacárido no ramificado
compuesto de D-glucosa, L-fucosa, 2-acetamido-2-deoxy-D-galactosa, y 4-acetamido-
4,6-dideoxy-D-mannosa [43], y la fracción común que E. coli O157 comparte con los otros
grupos bacterianos es el carbohidrato 4-acetamido-4,6-dideoxy-D-mannosa. Como es el
caso de los serogrupos de E. coli O7 y O116 [5] y de otros grupos bacterianos como
Brucella abortus, Yersinia enterocolitica, Citrobacter freundii, Escherichia hermannii y V.
cholerae O1 Inaba y Salmonella Urbana y S. Arizonae [44, 45, 46, 47, 48] la presencia
de este carbohidrato puede estar relacionado con la reactividad antigénica cruzada que
se presenta en sueros de conejos inmunizados contra estas bacterias.
1.7 Respuesta inmune contra el LPS O157 en población expuesta y no expuesta.
Estudios realizados en pacientes infectados con E. coli O157:H7 han mostrado que las
infecciones por estas bacterias dan lugar al desarrolló de anticuerpos contra el
lipopolisacárido (LPS) de la bacteria [49], así como contra la toxina Stx [50]. En el caso
de individuos que desarrollan SUH se pueden detectar anticuerpos IgM e IgG en suero e
IgA en heces y suero [51, 52, 53, 54]. En trabajos realizados por diferentes investigadores
[55, 56, 57, 58], se ha identificado la presencia de anticuerpos anti-LPS O157 en
manejadores de ganado y en un 13 a 20 % de la población que vive en áreas rurales en
comparación con individuos de áreas urbanas en la misma región, en los que menos del
4 % mostraban anticuerpos contra el LPS O157. En los mismos estudios se ha
identificado que la población de estas áreas rurales presenta un riesgo menor a infección
por E. coli O157 y de manera consecuente al desarrollo de SUH. Niños que viven en

11
zonas rurales que muestran anticuerpos contra el LPS O157 y presentan menos visitas
al médico por cuadros diarreicos [59].
1.8 E. coli O157:H7 en población mexicana y respuesta inmune.
Estudios relacionados con el aislamiento de E. coli O157 en México, muestran que la
bacteria se ha aislado de niños con y sin diarrea y de muestras de agua, sin embargo,
ninguno de los aislados presentó el antígeno H7 e incluso una gran mayoría fueron no
móviles. Por otro lado, al realizar el análisis genético de estas bacterias, los resultados
mostraron que no eran portadoras de genes relacionados con la virulencia del
microorganismo como stx1-2, ehxA y eae (datos no publicados).
En México no existen reportes de SUH asociado a infección por E. coli O157. Con el
propósito de explicar que factores pudieran estar participando en la ausencia de
infecciones por E. coli O157:H7, Navarro A, et al [60] reportaron la evaluación de la
respuesta inmune contra el LPS de E. coli O157 en población mexicana abierta. En este
trabajo se analizó la respuesta inmune humoral de 605 muestras de suero de niños,
adolescentes y adultos que pudiera estar implicada con la baja frecuencia de aislamientos
de E. coli O157:H7 observada en México. Se encontró que 121 (20 %) de los sueros
presentaron anticuerpos contra el LPS O157. Además, se determinó que al inmunizar
conejos con las cepas de E. coli O7, O116 y O157 hervidas con vapor fluente, sus sueros
presentaban actividad bactericida contra los antígenos heterólogos de las bacterias a
títulos más altos que los observados contra los antígenos homólogos. Por lo que, los
resultados obtenidos en dicho estudio sugieren que la respuesta inmune de la población
mexicana contra el LPS O157 pudiera ser una respuesta inmune heteróloga inespecífica,
pero protectora contra la colonización intestinal por la bacteria. La respuesta inmune

12
contra LPS O157 pudiera tener su origen en la exposición de la población a bacterias que
comparten fracciones antigénicas comunes con el LPS O157.
1.9 Vacunas contra E. coli O157.
Uno de los primeros estudios del uso de proteínas de la bacteria como vacunas fue el
realizado por Potter [61] quién reportó el uso de proteínas del sistema III de secreción de
la bacteria como inmunógenos para prevenir la colonización por E. coli O157:H7 en
ganado lechero (vacas lecheras). Una de éstas era la proteína translocadora del receptor
de intimina (Tir) y la segunda era una proteína efectora denominadas proteínas
secretadas de E. coli enteropatógena (EspA) las cuales tienen una función importante en
la adherencia de la bacteria al enterocito del epitelio intestinal. La vacuna desarrollada
por Potter [61] consistía de las dos proteínas mencionadas, las cuales fueron aplicadas
por vía subcutánea en dos dosis de 2 mL cada una en intervalos de 21 días. Después de
dos semanas de aplicar la última dosis se desafió a los animales inmunizados con una
dosis de 108 unidades formadoras de colonias (UFC)/mL de E. coli O157:H7 por vía
intragástrica observándose una disminución en el número y tiempo de excreción de la
bacteria comparada con los controles a los cuales sólo se les administró un placebo.
Otra estrategia para inhibir la colonización por E. coli O157 ha sido la vacunación con
antígenos del LPS de E. coli O157 conjugado con proteínas [62, 63]. Para utilizar esta
estrategia los autores se basaron en la hipótesis que proponía que la respuesta por
anticuerpos (IgG) contra las cadenas laterales del polisacárido del LPS de E. coli O157
pueden conferir protección mediante la inducción de una actividad bactericida contra los
microorganismos. La propuesta anterior surgió de la observación de que niños vacunados
con polisacáridos conjugados con proteínas producen IgG con capacidad protectoras. En

13
un primer estudio Konadu [62], utilizaron los carbohidratos de las cadenas laterales del
LPS de E. coli O157 conjugados de forma separada a tres proteínas, una de ellas era
albumina bovina (BSA), la segunda, la exotoxina C de Clostridium welchii y la tercera la
exoproteina A recombinante de Pseudomonas aeruginosa. Las cadenas laterales
conjugadas a dichas proteínas fueron aplicadas a ratones BALB/c por vía subcutánea en
tres dosis en intervalos de catorce días. Después de la inmunización los anticuerpos (IgG)
de los ratones presentaron actividad bactericida contra una cepa de E. coli O157.
En un estudio similar reportado por el mismo autor [63] se utilizó una vacuna utilizando
los polisacáridos del LPS O157 conjugada a la exoproteína A de P. aeruginosa. La
vacuna mencionada se aplicó a 87 voluntarios adultos por vía intramuscular en una dosis
de 25 g de la vacuna. En estos voluntarios no se registraron reacciones adversas a la
vacuna, observándose un incremento en los niveles de anticuerpos IgG e IgM, además
dichos anticuerpos presentaron actividad bactericida contra una cepa de E. coli O157.
Sin embargo, estudios reportados por Conlan [64], en el cual se inmunizaron ratones
BALB/c y C57BL/6 por vía subcutánea con el antígeno O de E. coli O157 conjugado con
albumina de caballo (HSA). Los autores observaron que en los ratones BALB/c la
respuesta inmune estaba dirigida principalmente contra la albumina de caballo y una
respuesta pobre contra los carbohidratos del antígeno O de E. coli O157. Así mismo se
observó que la inmunización no protegía a los ratones contra la colonización intestinal
por E. coli O157 después del reto de los animales con la bacteria. Concluyendo que una
vacunación por vía subcutánea no era la vía más adecuada a utilizarse contra un
microorganismo que infecta al intestino.

14
Otros esfuerzos de los mismos autores para desarrollar vacunas contra la colonización
intestinal contra E. coli O157 [65], consistieron en analizar la capacidad de Salmonella
Landau, para conferir protección contra la colonización intestinal en un modelo animal
(ratones BALB/c) contra E. coli O157. Los autores utilizaron la cepa de S. Landau por el
hecho de que esta bacteria comparte epitopos en el antígeno somático de E. coli O157.
En el estudio los autores administraron en ratones BALB/c por vía oral dos dosis de
2X1010 UFC (S. Landau) en un volumen de 0.2 ml en intervalos de catorce días. Los
ratones desarrollaron títulos altos de IgG en suero e IgA en heces contra E. coli O157.
Posteriormente los ratones inmunizados fueron retados con un inóculo de E. coli O157
los cuales mostraron una reducción significativa en el número y tiempos de excreción de
la bacteria comparada con la de los ratones controles que no fueron inmunizados con S.
Landau, sugiriendo que una respuesta inmune en la mucosa intestinal dirigida contra E.
coli O157 puede incrementar la resistencia a la colonización contra E. coli O157.
Otras alternativas para el desarrollo de vacunas contra E. coli O157 consisten en utilizar
LPS O157 conjugado a la subunidad B de la toxina Shiga [66]. En estos estudios se
planteó una hipótesis similar a la propuesta previamente por Konadu [62], de que las
vacunas basadas en el LPS tienen la propiedad de inducir anticuerpos IgG anti-LPS con
actividad bactericida. Considerando la hipótesis anterior los autores sintetizaron el
polisacárido O-específico (O-SP) de E. coli O157 y lo acoplaron a la subunidad B no
toxigénica de la toxina Stx1, con lo cual se pudiera inducir anticuerpos en suero IgG anti-
LPS O157 y anticuerpos neutralizantes para Stx1. En el estudio mencionado los autores
utilizaron dos conjugados, uno denominado O-SP conjugado directamente con la
subunidad de Stx1B (OSP-StxB1), y el segundo el O-SP unido a ADH (adipic acid
dihydrazide) y acoplado a la subunidad Stx1B (OSP-AH-StxB1), ambos conjugados

15
administrados subcutáneamente en ratones en tres dosis, cada dosis en intervalos de
catorce días con una concentración de 2.5 g del O-SP. En este estudio los autores
encontraron que después de aplicar las tres dosis, los ratones desarrollaron anticuerpos
IgG e IgM los cuales mostraron diferencias significativas comparadas con los ratones a
los que no se les aplicó ninguno de los conjugados. El suero de los ratones inoculados
con ambos tipos de conjugados, OSP-Stx1B y OSP-AH-Stx1B presentaban la propiedad
de actividad bactericida contra la bacteria. La actividad bactericida presentada por los
sueros de los ratones fue eliminada mediante la absorción de los sueros de los ratones
con el LPS de E. coli O157, lo que significó que los anticuerpos dirigidos contra el O-SP
eran los responsables de la actividad bactericida. Además, los sueros de los ratones
inmunizados con los conjugados de O-PS presentaron la propiedad de neutralizar la
toxina Stx1. La conclusión del estudio fue que este tipo de conjugados pudieran utilizarse
en el control de brotes epidémicos de infecciones por E. coli O157.
Los diferentes estudios previamente referidos indican que la respuesta de anticuerpos
anti-O157 está dirigida contra las cadenas laterales del LPS. Lo anterior provee
elementos importantes para proponer que dichos compuestos sean considerado en el
desarrollo de posibles vacunas contra E. coli O157 [63, 67, 68]. Existen, sin embargo,
limitantes para utilizar el LPS como inmunógeno, entre ellas el efecto tóxico del lípido A,
el cual en caso de ser eliminado hace al LPS un compuesto escasamente inmunogénico.
Con relación a las cadenas laterales de carbohidratos, la respuesta que inducen es timo-
independiente y por lo mismo de corta duración [69, 70].

16
1.10 Phage display: usos del método.
Dada la importancia clínica y epidemiológica de E. coli O157, es que se han planteado
alternativas para la búsqueda de un inmunógeno con capacidad protectora, pero que no
muestre los efectos tóxicos colaterales del LPS. Una de dichas alternativas es el
procedimiento de Phage display [71], este sistema utiliza una biblioteca de péptidos (1012)
de 7 y 12 residuos de aminoácidos cuyas secuencias se integraron al gene que codifica
para la Proteína III (pIII) del fago filamentoso M13, el sistema requiere anticuerpos
dirigidos contra el epitopo del inmunógeno en estudio para seleccionar el o los péptidos
(mimotopos) expresados en la superficie del fago M13 [72, 73]. Con las secuencias de
estos se diseñan péptidos sintéticos los cuales después de ser evaluados y en caso de
tener capacidad protectora, pudieran ser utilizados para el desarrollo de vacunas seguras
con propiedades inmunogénicas.
Utilizando anticuerpos preparados contra componentes de una bacteria, tal como el LPS,
se seleccionan los fagos que expresan péptidos que mimetizan un epítopo (mimotopos)
del LPS u de otros compuestos de la pared celular de la bacteria. Con este procedimiento
se han obtenido mimotopos del LPS de diferentes microorganismos como Shigella
flexneri, Brucella abortus y Neisseria meningitidis [74, 75, 76]. En algunos casos se ha
observado que la inmunización de animales con los péptidos mimotopos se induce una
respuesta protectora contra los mismos microorganismos [77, 78]. El procedimiento
también se ha empleado para la localización de motivos funcionales en proteasas de E.
coli y Shigella flexneri [79], así como para obtener péptidos con actividad bactericida [80].
En este trabajo se presentan resultados sobre la identificación de péptidos mimotopos
del LPS O157 obtenidos por Phage display y su capacidad para inducir una respuesta
inmune contra los péptidos sintéticos y el LPS O157.

17
2. JUSTIFICACIÓN
Los polisacáridos y glicoconjugados de la pared celular de E. coli son complejos y
muestran una diversidad estructural dentro de los serogrupos de esta bacteria. En la
última década, las bibliotecas de péptidos que se han construido en fagos han contribuido
al desarrollo de una técnica útil para buscar péptidos imitadores (mimotopos) de azucares
[81, 82]. En el presente estudio se empleó el método de Phage display para identificar
mimotopos de carbohidratos de la pared celular de E. coli O157 usando una librería de
péptidos presentes en el fago M13 y seleccionados con anticuerpos de conejo producidos
contra el LPS (lipopolisacárido) de E. coli O157. En caso de seleccionar fagotopos que
expresen péptidos con secuencias consenso, se podrá analizar su capacidad
inmunogénica y evaluar si los anticuerpos inducidos en conejo por estos fagotopos tienen
la capacidad de reconocer el LPS de E. coli O157, lo que dará elementos para obtener
péptidos sintéticos que pudieran inducir la producción de anticuerpos con capacidad
protectora contra la infección producida por este grupo de E. coli.

18
3. OBJETIVOS
Objetivo general:
Identificar por el procedimiento de Phage display mimotopos del LPS O157 y evaluar su
capacidad inmunogénica para proponer su posible empleo en el desarrollo de vacunas
con capacidad protectora.
Objetivos particulares:
1. Obtener fagotopos que expresen péptidos mimotopos del LPS O157 utilizando IgG
de conejos inmunizados con el LPS O157.
2. Obtener la síntesis de péptidos con las secuencias de los diferentes fagotopos
previamente seleccionados.
3. Generar en conejos anticuerpos contra los péptidos mimotopos y evaluar su
capacidad para reaccionar con el LPS de E. coli O157.
4. Evaluar si antisueros obtenidos contra el LPS de E. coli O157 reaccionan contra
péptidos sintéticos.
5. Determinar sí los péptidos mimotopos sintéticos compiten por los sitios de unión
de los anticuerpos anti-LPS O157.
6. Identificar la región del LPS O157 que es reconocida por anticuerpos anti-péptidos
mimotopos, para definir a que región del LPS corresponde.
7. Evaluar si sueros de origen humanos con respuesta contra el LPS O157
reaccionan con el o los péptidos mimotopos del LPS O157.

19
4. MATERIAL Y MÉTODOS
4.1 Extracción de lipopolisacáridos. Se obtuvo el LPS de E. coli O157:H7 (ATCC
700927, FMU 108287) mediante el método de extracción fenol-agua [83]. El LPS se trató
con 20 g/ mL de DNasa, RNasa y se incubó a 37 ° C durante 3 h, a la suspensión anterior
se adicionaron 200 g/mL de proteinasa K y se incubó nuevamente a 50 °C durante 3
horas, después de la incubación la suspensión del LPS fue centrifugado a 35 000g
durante 3 h, finalmente se conservó liofilizado a 4 °C hasta su uso.
4.2 Sueros anti-LPS O157. Con el LPS de E. coli O157 previamente obtenido se
inmunizaron conejos blancos Nueva Zelanda [5], a cada uno se le administraron cinco
dosis por vía subcutánea cada una con 100 g/mL del LPS diluido en PBS (pH 7.2), la
primera contenía adyuvante completo de Freund (1:1) y las cuatro restantes el mismo
adyuvante de Freund pero sin Mycobacterium [84], las dosis se aplicaron en intervalos
de siete días. Además, se obtuvieron anticuerpos contra la bacteria completa, para esto
conejos blancos Nueva Zelanda fueron inmunizados por vía intravenosa con una cepa
de E. coli O157:H7 (ATCC 700927, FMU 108287) previamente hervida con vapor fluente
(100 °C) durante 1 hora, siguiendo el protocolo de inmunización previamente reportado
por Ewing, [5]. En ambos casos, antes de la inmunización se tomó una muestra de sangre
para obtener los sueros preinmune y utilizarlos como control negativo en los diferentes
ensayos. Una semana después de aplicar la última dosis, previa aplicación por vía
intravenosa de pentobarbital (Anestesal, Pfizer) a la concentración de 6 mg en 1.0 mL
por Kg de peso del conejo se realizó una sangría de los animales. Los experimentos con
animales fueron desarrollados de acuerdo a la Norma Oficial Mexicana NOM-062-ZOO-

20
1999 (Técnicas específicas para la producción, cuidado y uso de animales de laboratorio)
[85].
4.3 Obtención de inmunoglobulinas (IgG). A partir de muestras de sueros preinmune
e inmunes de los conejos se obtuvieron las IgGs por precipitación con sulfato de amonio
4.1 M (Sigma) primero al 50 % (v/v) de saturación y posteriormente al 33 % (v/v) [86]. La
purificación de las mismas se realizó por cromatografía de afinidad con Protein G Agarose
(Invitrogen), de acuerdo a las instrucciones del productor. La concentración de IgGs se
determinó con el reactivo de Bradford de acuerdo a las instrucciones del fabricante
(Coomassie Plus, Kit. Thermo Scientific, 23236).
4.4 Selección de mimotopos. Para este propósito se utilizó el procedimiento
previamente descrito por Smith y Dower [72, 73], con una biblioteca de péptidos (1012) de
12 residuos de aminoácidos (New England, BioLabs). Para la captura de los fagotopos
(Fgtp), clona de fagos, se emplearon las IgGs purificadas obtenidas de los sueros de
conejos antes y después de ser inmunizados. Brevemente, en una microplaca de 96
pozos (Maxisorp F96) se fijaron 15 g de IgG disueltas en 100 L de PBS pH 7.2, la placa
se incubó toda la noche a 4 ºC y se lavó 6 veces con PBS-Tween 0.1%(v/v),
posteriormente en cada pozo se colocaron 100 L con 4 x 1011unidades formadoras de
placa (ufp) del fago M13 disueltos en PBS (pH 7.2). La placa se incubó una hora a
temperatura ambiente con agitación suave, los fagos que no reaccionaron con las IgGs
se eliminaron lavando 10 veces con 200 L de PBS-Tween, los fagos capturados por las
IgG se liberaron con 100 L de buffer de Glicina-HCI 0.2 M y pH 2.2. Las suspensiones
de fagos obtenidas de cada pozo se transfirieron a microtubos de 1.6 mL y el pH se
neutralizó con 15 L de buffer Tris-base pH 9.0. En total se realizaron tres rondas de

21
bioselección, utilizando la misma concentración de fagos y una concentración de 7.5
g/mL y 3.75 g/mL de IgG para la segunda y tercera ronda respectivamente. En cada
ronda de biopanning los fagos se propagaron en la cepa E. coli ER 2738 (New England
BioLabs) utilizando 300 L de caldo Luria Bertani (LB, Difco), para la infección se
utilizaron 70 L de los fagos diluidos en PBS, la mezcla se inoculó en 20 mL de caldo
2YT (Extracto de levadura, Bacto triptona y NaCl) y se incubó a 37 ºC con agitación (250
rpm) por 4:30 h. Los fagos se recuperaron del sobrenadante por centrifugación a 8000g
(Sorvall RC5) durante 10 min a 4 °C y posteriormente se precipitaron con polyethylene
glycol (PEG) 8000-NaCl (1/4 de volumen total) durante toda la noche a 4 ºC, finalmente
la muestra se centrifugó a 8000g durante 10 min y los fagos se recuperaron del
precipitado. En la tercera ronda, la cepa de E. coli ER 2738 se infectó de manera similar
y se inoculó en una placa de agar LB con 250 L (0.1 M) de isopropyl -D-
thiogalactopyranoside (IPTG), la placa de LB/IPTG se incubó a 37 °C durante 18 h. A
partir del crecimiento obtenido en la placa del cultivo anterior, fueron seleccionadas al
azar 15 clonas de cada fagotopo (Fgtp) y se propagaron de manera individual como se
describió anteriormente.
4.5 Secuenciación de DNA. El DNA de los fagotopos (Fgtps) seleccionados se obtuvo
por el procedimiento descrito por Wilson [87]. Brevemente, a 1 mL de la suspensión de
fagos previamente propagados se les adicionaron 400 L de buffer PEG-8000/NaCl
(20%/40%), la mezcla se dejó a -20 °C (10 min). La suspensión de fagos se centrifugo a
10000g (10 min), el pellet se suspendió en 100 L de buffer de Ioduro de Sodio y se
adicionaron 250 L de etanol absoluto para precipitar el DNA. Para obtener el DNA la
muestra se centrifugó (10000 g/ 10 min) y el “pellet” se suspendió en etanol absoluto (250

22
L), finalmente se centrifugó, se eliminó el sobrenadante y el DNA se secó con vacío
(Automatic Environmental Speed VacAES-1010, Thermo Fisher Scientific). A cada una
de las muestras de DNA se le realizó secuenciación automatizada (Genetic analyzer
3100) utilizando el iniciador -96 gIII (5´-CCC TCA TAG TTA GCG TAA CG-3´) incluido en
el kit de BioLabs. Las secuencias se editaron con el programa Chromas y se realizó su
traducción con el programa Translate tool ubicada en ExPASy Proteomics Server
(http://www.expasy.ch/tools/dna.html). Para identificar la secuencia consenso de los
péptidos seleccionados se utilizó el programa MIMOX (http://i.uestc.edu.cn/mimox/cgi-
bin/mimosa.pl).
4.6 Péptidos sintéticos. Para la síntesis química de los péptidos (Bio-Synthesis) se
utilizaron como molde las secuencias de aminoácidos identificadas en los fagos
seleccionados, los péptidos (SP) fueron sintetizados sin conjugar y conjugados a
hemocianina de lapa Californiana (Keyhole Limpet Hemocyanine, KLH). La síntesis de
los péptidos conjugados con KLH se solicitó con la adición de una extensión en el grupo
amino-terminal de 5 aminoácidos que consistió de Cys-Ser-Gly-Gly-Gly (CSGGG).
4.7 Suero anti-fagotopos y anti-péptidos sintéticos. Anticuerpos contra los mimotopos
del LPS O157 se obtuvieron al inmunizar conejos blancos Nueva Zelanda (1.5 Kg) con
las clonas de los fagotopos seleccionados previamente (1013 ufp) o con el péptido
sintético conjugado con KLH (500 g la primera y 250 g las restantes), de acuerdo al
protocolo reportado por Galfré [88] y Gevorkian [84] respectivamente. Una semana
después de la quinta inmunización se realizó una sangría siguiendo el procedimiento
referido previamente y el suero obtenido se conservó en alícuotas a -20 ºC hasta su uso.

23
Previo a las inmunizaciones se tomaron muestras de sangre de los conejos, para obtener
el suero pre-inmune y utilizarlo como control negativo en los diferentes ensayos.
4.8 IgG anti-fagotopo. Para evaluar la respuesta de las IgG anti-fagotopo contra los
fagotopos homólogos se realizó un ensayo de ELISA [79], en microplacas de 96 pozos
(Maxisorp F96) se fijaron 10 g/100 L de IgG anti-fagotopos (Fgtp5, Fgtp11, Fgtp12 y
Fgtp16) disueltas en 200 L en PBS pH 7.4. Las placas se incubaron a 4 °C durante 18
h y se lavaron tres veces con PBS/Tween20 (0.05%), con 200 L de PBS/BSA se
bloquearon los espacios libres (2 h, temperatura ambiente), se lavó (5 veces) con
PBS/Tween y se adicionaron 1011 fagos de Fgtp5, Fgtp11, Fgtp12 y Fgtp16 disueltos en
200 L de PBS pH 7.4, se incubó a temperatura ambiente por 2 horas. La reacción se
reveló utilizando un anticuerpo monoclonal anti M13-pIII (1:500) conjugado con
peroxidasa de rábano (New England Biolabs, USA), y se visualizó a los 30 minutos
después de adicionar el sustrato 2’,2’-azinobis-3-ethylbenzothiazoline-6-sulphonico
(ABTS, Sigma-Aldrich). Se realizaron lecturas de la reacción en un lector de ELISA
(Dynatech) a 405 nm de densidad óptica (DO).
Con las mismas IgG anti-fagotopos y con un ensayo de ELISA similar al descrito
anteriormente se analizó su reactividad contra los péptidos sintéticos (SP) no conjugados
(SP5, SP11, SP12 y SP16. Se describe brevemente, en microplacas de 96 pozos
(Maxisorp F96) se pegaron los péptidos SP disueltos en buffer de carbonatos pH 9.6 la
concentración de 10 g/mL, las placas se incubaron a 37 °C (2 h) y a 4 °C (18-24 h),
posteriormente las placas se bloquearon con 200 L de PBS/leche descremada al 1.0 %
(Svelty) a temperatura ambiente (2 h). Las placas se lavaron (3 veces) con PBS/Tween
0.05 % y se les adicionaron las concentraciones IgGs anti-fagotopos (10 g/mL) disueltas

24
en PBS pH 7.4, nuevamente se incubó a 37 °C durante 2 h. Las placas se lavaron tres
veces (PBS/Tween20) y a cada uno de pozo se adicionaron 100 L de anti-IgG de conejo
obtenido en cabra (1:1000) conjugado con fosfatasa alcalina (Invitrogen). Se incubó a 37
°C durante 2 h y la reacción se visualizó al adicionar 200 µL del substrato p-nitrofenil
fosfato (Sigma) en buffer de diethanolamina (pH 9.8, Sigma), la reacción se detuvo
adicionando 25 µL de NaOH 3 M. Para las lecturas de DO de la reacción se utilizó un
lector de ELISA (MR 580, Dynatech) a 405 nm. Todos los ensayos se realizaron por
duplicado y en dos ensayos independientes, como control negativo se utilizaron las IgGs
de los sueros preinmunes.
4.9 Respuesta de sueros anti-péptido. Con la prueba de ELISA descrita previamente
con modificaciones menores, se evaluó la respuesta de los sueros obtenidos contra cada
uno de los péptidos sintéticos y el LPS O157. En microplacas de 96 pozos (Maxisorp F96)
se pegaron los péptidos sintéticos (10g/mL) disueltos en buffer de carbonatos pH 9.6,
las placas se incubaron a 37 °C (2 h) y a 4 °C (18-24 h), posteriormente las placas se
bloquearon (200 L) con PBS/leche descremada al 1.0 % (Svelty) a temperatura
ambiente durante 2 h. La reacción se reveló utilizando anti-IgG de conejo obtenida en
cabra (1:1000) y se realizaron lecturas de DO a 405 nm.
4.10 Ensayos de competencia. Para confirmar que SP12 era mimotopo de un sitio
inmunogénico del LPS de E. coli O157, se realizó un ensayo de competencia de acuerdo
al procedimiento previamente descrito por Phalipon [89] con modificaciones menores. En
microplacas de 96 pozos (Maxisorp F96) se pegaron 10 g/mL del LPS O157,
previamente muestras de suero anti-LPS O157 (1:100) se incubaron con las
concentraciones de 10 g/100 L hasta 100 g/100 L del péptido SP12 a 37 °C durante

25
2 h. Para evaluar la respuesta a cada pozo se le adicionó 100 L de la suspensión del
suero anti-LPS O157/SP12 y se incubó a 37 °C durante 2 h, posteriormente la reacción
se reveló y realizaron lecturas de DO como el ensayo anterior. Todos los ensayos se
realizaron por duplicado y en dos ensayos independientes, como control de la
especificidad de la reacción muestras de sueros anti-LPS O157 fueron incubadas con el
péptido SP3 y el ensayo se realizó como se describió arriba.
Se realizó un ensayo similar al descrito anteriormente, en este ensayo en una placa
placas de 96 pozos de ELISA (Maxisorp F96) el péptido SP12 se pegó a la concentración
de 10 g/mL, previamente muestras de suero anti-péptido SP12 (1:100) se incubaron con
las concentraciones de 10 g/100 L hasta 200 g/100 L del LPS O157 a 37 °C durante
2 h. Para evaluar la respuesta a cada pozo se le adicionó 100 L de la suspensión del
suero anti-SP12/LPS O157 y se incubó a 37 °C durante 2 h. Todos los ensayos se
realizaron por duplicado y en dos ensayos independientes, como control de la
especificidad de la reacción muestras de sueros anti-SP12 fueron incubadas con un LPS
de E. coli O179 (E43478) sin relación antigénica con el LPS O157 y el ensayo se reveló
como se describió arriba.
4.11 Sitio que mimetiza SP12 en el LPS O157. Para identificar el posible sitio del cual
fue seleccionado SP12 en el LPS O157 se realizó un ensayo de Western-blot. A muestras
de 10 g (1 mg/mL) del LPS O157, se les realizó SDS-PAGE [90] en un gel de
poliacrilamida (15 %) con 4M de urea. La electroforesis se realizó en una cámara mini-
protean III (BioRad, Laboratories) aplicando una corriente de 25 mA durante 30 min y 45
mA durante 1 h. La transferencia [91] a membranas de nitrocelulosa de 0.45 m (BioRad,
Laboratories) se realizó con una cámara (Mini Protean BioRad laboratories) aplicando

26
una corriente de 200 mA constantes durante 2 h. Posteriormente las membranas fueron
cortadas en tiras y se bloquearon con Buffer Salino Tris-Base (TBS)/leche descremada
(1.0 %), se incubaron a temperatura ambiente durante 30 min, se lavaron tres veces con
TBS/Tween 0.05 %. Las tiras de nitrocelulosa se incubaron con una dilución 1:100 del
suero anti-SP12 en TBS/ 1.0 % de leche descremada y se incubaron a 4°C durante 12 h.
Después de la incubación las tiras fueron lavadas tres veces. Para visualizar la reacción
las tiras se incubaron con anti-IgG de conejo marcada con fosfatasa alcalina a una
dilución 1:1000 (Millipore) durante 1 h a temperatura ambiente, al final de la incubación
las tiras fueron lavadas nuevamente tres veces. Para visualizar la reacción antígeno-
anticuerpo se adicionaron 250 L de 5-bromo-4-cloro-3-indolylphosphate (BCIP)-
nitroblue tetrazolium (Sigma Fast).
4.12 Reactividad de sueros humanos contra SP12. Con el ensayo de ELISA que
previamente se describió, se analizó la respuesta contra el péptido SP12 de muestras de
suero de humanos que presentaban reactividad contra el LPS O157 [60]. Placas de 96
pozos (Maxisorp F96) se recubrieron con una concentración de 10 g/mL de SP12, los
sueros con reactividad positiva contra el LPS O157 y los controles negativos se utilizaron
en una dilución 1:100.
4.13 Análisis estadístico. En todos los ensayos de ELISA se realizó un análisis de
varianza de una cola del promedio de las DO de dos ensayos realizados de manera
independiente y por duplicado con un nivel de significancia de <0.05.
27
5. RESULTADOS
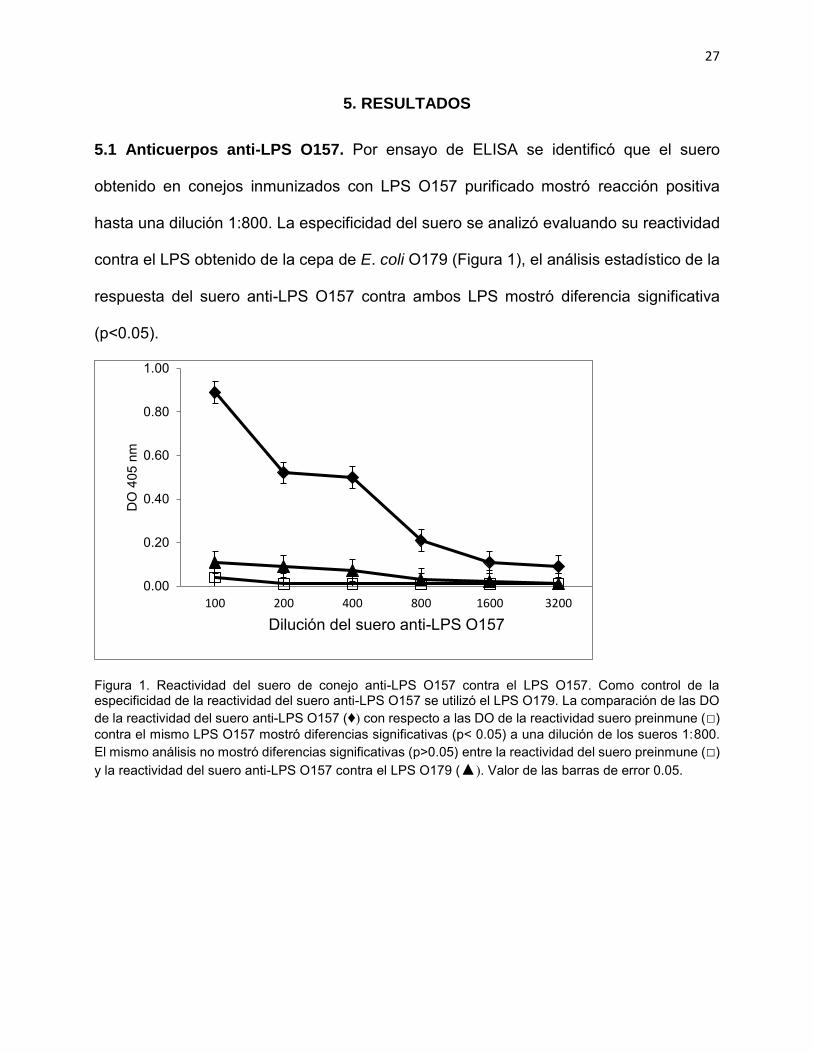
5.1 Anticuerpos anti-LPS O157. Por ensayo de ELISA se identificó que el suero
obtenido en conejos inmunizados con LPS O157 purificado mostró reacción positiva
hasta una dilución 1:800. La especificidad del suero se analizó evaluando su reactividad
contra el LPS obtenido de la cepa de E. coli O179 (Figura 1), el análisis estadístico de la
respuesta del suero anti-LPS O157 contra ambos LPS mostró diferencia significativa
(p<0.05).
Figura 1. Reactividad del suero de conejo anti-LPS O157 contra el LPS O157. Como control de la especificidad de la reactividad del suero anti-LPS O157 se utilizó el LPS O179. La comparación de las DO de la reactividad del suero anti-LPS O157 (♦) con respecto a las DO de la reactividad suero preinmune (□) contra el mismo LPS O157 mostró diferencias significativas (p< 0.05) a una dilución de los sueros 1:800. El mismo análisis no mostró diferencias significativas (p>0.05) entre la reactividad del suero preinmune (□) y la reactividad del suero anti-LPS O157 contra el LPS O179 (▲). Valor de las barras de error 0.05.
0.00
0.20
0.40
0.60
0.80
1.00
100 200 400 800 1600 3200
DO
405
nm
Dilución del suero anti-LPS O157

28
5.2 Selección de mimotopos. Se obtuvieron 63 clonas de fagos, de éstas 33 fueron
seleccionadas con IgGs de dos sueros de conejos inmunizados con la bacteria completa
(O157/2001 y O157/2004), 15 que se obtuvieron con IgG del suero preinmune y 15 más
fueron seleccionadas con las IgG del suero de conejos inmunizados con el LPS O157
purificado.
5.3 Análisis de péptidos. A partir de las secuencias de nucleótidos de los 63 fagotopos
seleccionados, se realizó su traducción para conocer la conformación de aminoácidos de
los péptidos mimotopos (http://www.expasy.ch/tools/dna.html). Las secuencias obtenidas
con las IgG del suero preinmune se caracterizaron por presentar, His (con carga positiva),
Trp y Phe (aminoácidos aromáticos), Pro (cíclico no polar), Ile, Leu (no polares) y Thr
(polar sin carga con grupo hidroxilo) sin una secuencia en común (datos no mostrados).
Con relación a los péptidos obtenidos con IgGs del suero anti-E. coli O157/2004, estos
se distinguieron por contener His, Thr, Phe, Pro, y Leu (aminoácidos: básico, polar sin
carga con grupo hidroxilo, aromáticos, cíclico no polar y no polar respectivamente). Los
aminoácidos de los péptidos obtenidos con las IgG del suero anti-E. coli O157/2001,
presentaron His, Pro, Ser Leu, Ile y Thr (aminoácidos: básico, cíclico no polar, polar sin
carga con grupo hidroxilo, no polares, polar sin carga con grupo hidroxilo
respectivamente).
Con respecto a las quince clonas seleccionadas con las IgG anti-LPS O157, éstas se
incluyeron en tres grupos (Cuadro 1), en el grupo I, se colocaron 5 (33 %) clonas en las
que se identificó el motivo LxP/YP/SxL como secuencia consenso; en el grupo II, se
ubicaron diez clonas en las que no se identificó una secuencia consenso. Estos péptidos
se caracterizaron por presentar los aminoácidos, Pro, Thr, Leu, Ala y Ser (aminoácidos:
cíclico no polar, polar sin carga con grupo hidroxilo, no polar o alifático y polar sin carga,
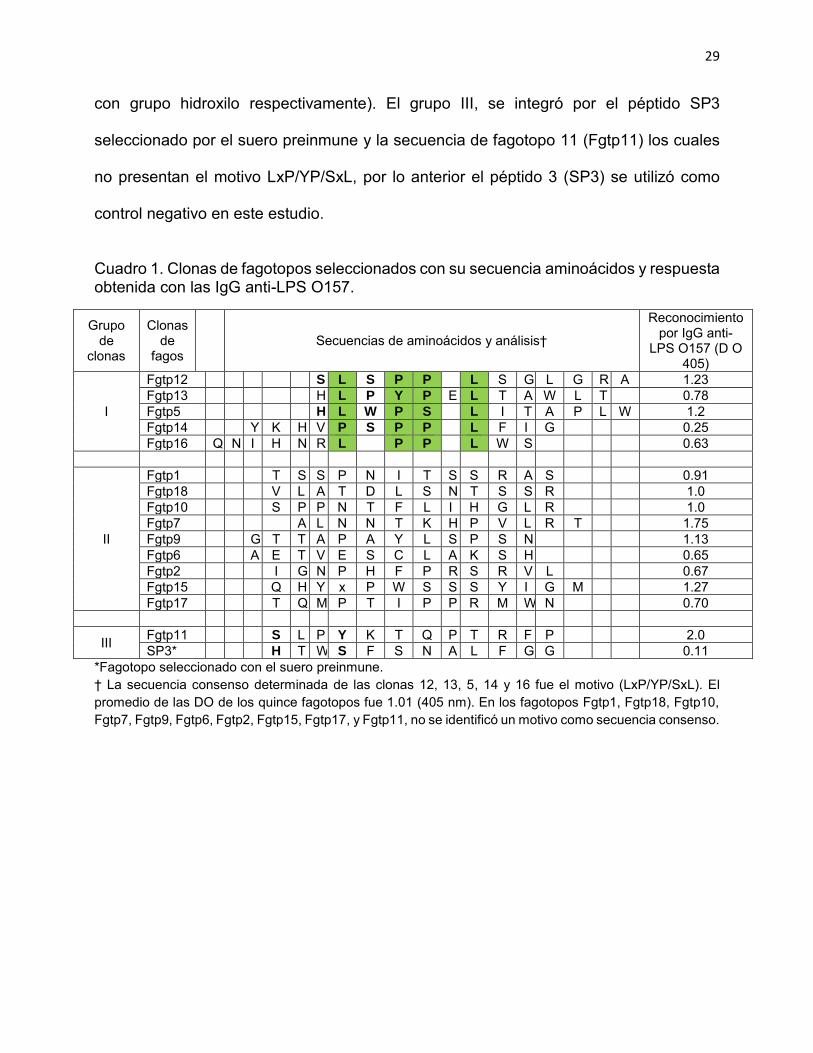
29
con grupo hidroxilo respectivamente). El grupo III, se integró por el péptido SP3
seleccionado por el suero preinmune y la secuencia de fagotopo 11 (Fgtp11) los cuales
no presentan el motivo LxP/YP/SxL, por lo anterior el péptido 3 (SP3) se utilizó como
control negativo en este estudio.
Cuadro 1. Clonas de fagotopos seleccionados con su secuencia aminoácidos y respuesta obtenida con las IgG anti-LPS O157.
Grupo de
clonas
Clonas de
fagos Secuencias de aminoácidos y análisis†
Reconocimiento por IgG anti-
LPS O157 (D O 405)
I
Fgtp12 S L S P P L S G L G R A 1.23 Fgtp13 H L P Y P E L T A W L T 0.78 Fgtp5 H L W P S L I T A P L W 1.2 Fgtp14 Y K H V P S P P L F I G 0.25 Fgtp16 Q N I H N R L P P L W S 0.63
II
Fgtp1 T S S P N I T S S R A S 0.91 Fgtp18 V L A T D L S N T S S R 1.0 Fgtp10 S P P N T F L I H G L R 1.0 Fgtp7 A L N N T K H P V L R T 1.75 Fgtp9 G T T A P A Y L S P S N 1.13 Fgtp6 A E T V E S C L A K S H 0.65 Fgtp2 I G N P H F P R S R V L 0.67 Fgtp15 Q H Y x P W S S S Y I G M 1.27 Fgtp17 T Q M P T I P P R M W N 0.70
III Fgtp11 S L P Y K T Q P T R F P 2.0 SP3* H T W S F S N A L F G G 0.11
*Fagotopo seleccionado con el suero preinmune. † La secuencia consenso determinada de las clonas 12, 13, 5, 14 y 16 fue el motivo (LxP/YP/SxL). El promedio de las DO de los quince fagotopos fue 1.01 (405 nm). En los fagotopos Fgtp1, Fgtp18, Fgtp10, Fgtp7, Fgtp9, Fgtp6, Fgtp2, Fgtp15, Fgtp17, y Fgtp11, no se identificó un motivo como secuencia consenso.

30
5.4 Reactividad de IgG anti-fagotopos. Se obtuvieron anticuerpos contra los fagotopos
Fgtp5, Fgtp12 y Fgtp16 que presentaron la secuencia consenso LxP/YP/SxL y Fgtp11
con la secuencia SLPYKTQPTRFP que, aunque, no presentaba secuencia consenso,
mostró reactividad intensa con las IgG anti-LPS O157. La reactividad de las IgG obtenidas
de los sueros anti-fagotopos (SFgtp) contra los fagotopos y péptidos sintéticos
homólogos, los resultados para ambos fueron positivos mostrando diferencias
significativas al comparar las DO obtenidas cuando se utilizaron las IgG de los sueros
pre-inmunes (Cuadro 2).
Cuadro 2. Las IgG anti-fagotopos reconocieron las clonas de fagos homólogos y péptidos sintéticos.
IgG anti-fagotpos Fagotopos homólogos*
Reconocimiento del fagotopo (DO, 405 nm)
Péptidos Sintéticos**
Reconocimiento del SP (DO, 405 nm)
SFgtp-5 Fgtp5 0.87 SP5 1.23 SFgtp-11 Fgtp11 1.19 SP11 1.17 SFgtp-12 Fgtp12 0.81 SP12 0.82 SFgtp-16 Fgtp16 0.87 SP16 1.34
*Reactividad de IgG obtenidas de los sueros anti-fagotopos contra las clonas de fagos. La comparación de las DO de la reactividad de IgG anti-fagotopos mostraron diferencias significativas (p<0.05) con respecto a las DO de las IgG del suero preinmune (DO, 0.05). **Reactividad de las IgG obtenidas de sueros anti-fagotopos contra los péptidos sintéticos (SP). La comparación de las DO de las IgG anti-fagotopos mostraron diferencias significativas (p<0.05) con respecto a las DO de las IgG del suero preinmune (DO, 0.03).
31
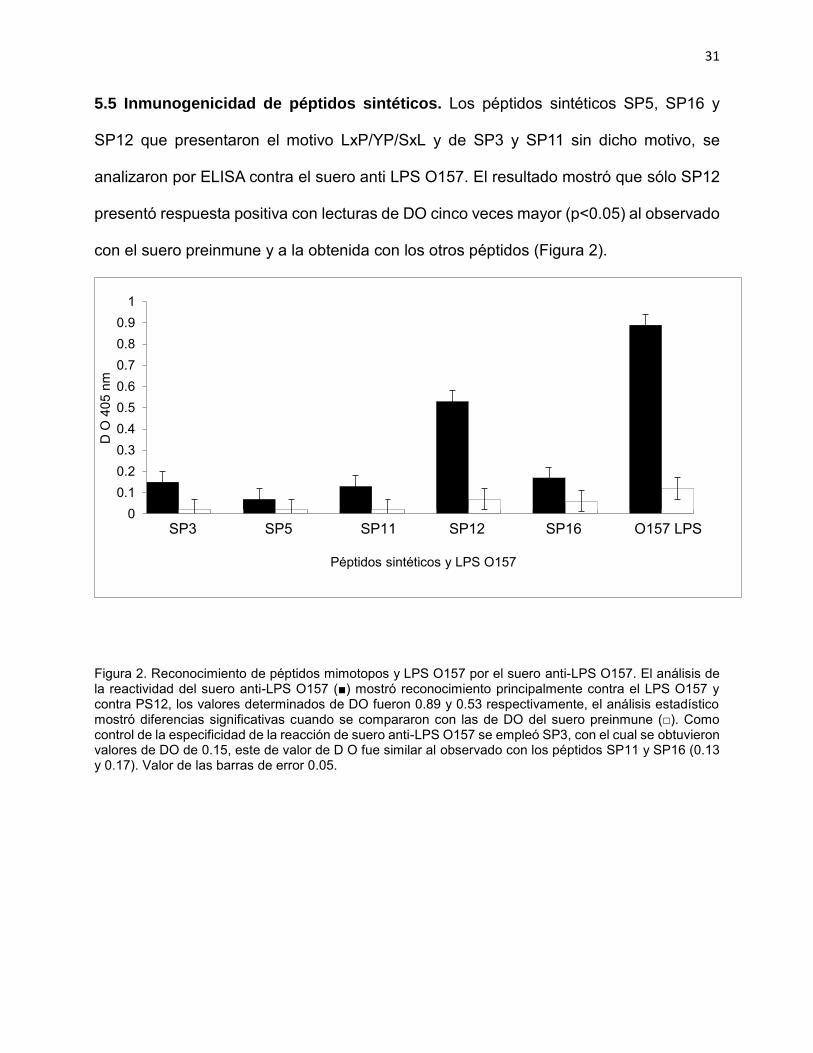
5.5 Inmunogenicidad de péptidos sintéticos. Los péptidos sintéticos SP5, SP16 y
SP12 que presentaron el motivo LxP/YP/SxL y de SP3 y SP11 sin dicho motivo, se
analizaron por ELISA contra el suero anti LPS O157. El resultado mostró que sólo SP12
presentó respuesta positiva con lecturas de DO cinco veces mayor (p<0.05) al observado
con el suero preinmune y a la obtenida con los otros péptidos (Figura 2).
Figura 2. Reconocimiento de péptidos mimotopos y LPS O157 por el suero anti-LPS O157. El análisis de la reactividad del suero anti-LPS O157 (■) mostró reconocimiento principalmente contra el LPS O157 y contra PS12, los valores determinados de DO fueron 0.89 y 0.53 respectivamente, el análisis estadístico mostró diferencias significativas cuando se compararon con las de DO del suero preinmune (□). Como control de la especificidad de la reacción de suero anti-LPS O157 se empleó SP3, con el cual se obtuvieron valores de DO de 0.15, este de valor de D O fue similar al observado con los péptidos SP11 y SP16 (0.13 y 0.17). Valor de las barras de error 0.05.
00.10.20.30.40.50.60.70.80.9
1
D O
405
nm
Péptidos sintéticos y LPS O157
SP3 SP5 SP11 SP12 SP16 O157 LPS
32
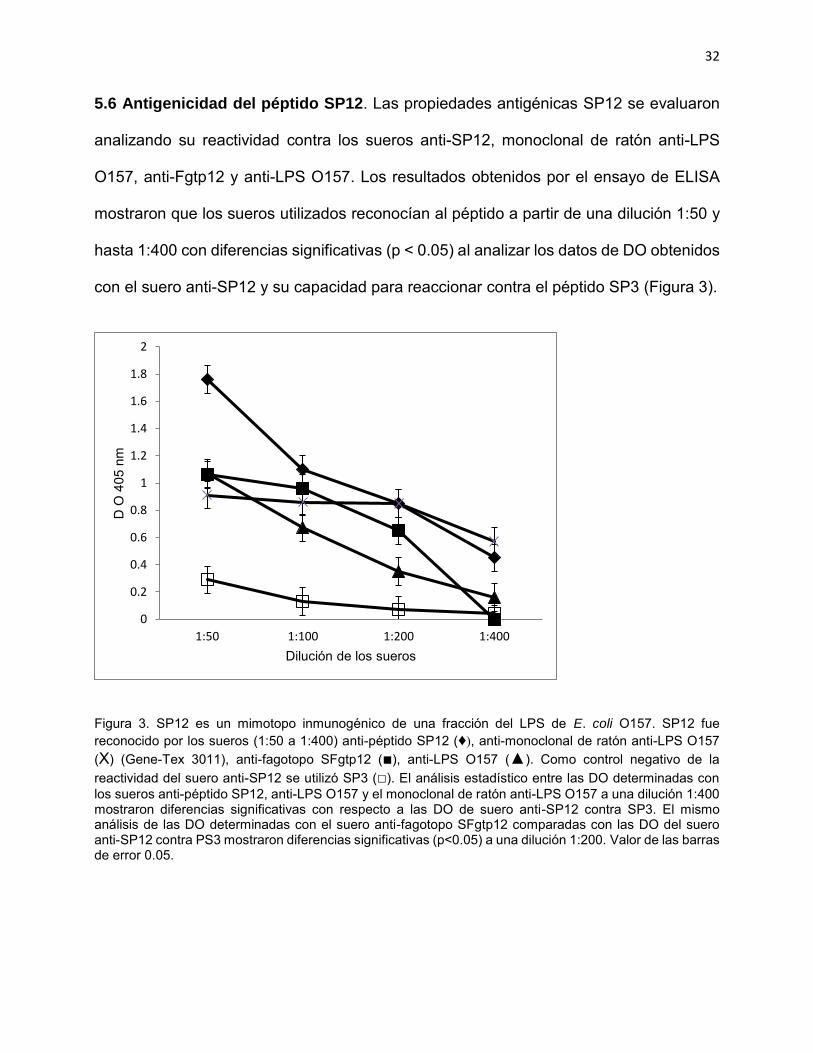
5.6 Antigenicidad del péptido SP12. Las propiedades antigénicas SP12 se evaluaron
analizando su reactividad contra los sueros anti-SP12, monoclonal de ratón anti-LPS
O157, anti-Fgtp12 y anti-LPS O157. Los resultados obtenidos por el ensayo de ELISA
mostraron que los sueros utilizados reconocían al péptido a partir de una dilución 1:50 y
hasta 1:400 con diferencias significativas (p < 0.05) al analizar los datos de DO obtenidos
con el suero anti-SP12 y su capacidad para reaccionar contra el péptido SP3 (Figura 3).
Figura 3. SP12 es un mimotopo inmunogénico de una fracción del LPS de E. coli O157. SP12 fue reconocido por los sueros (1:50 a 1:400) anti-péptido SP12 (♦), anti-monoclonal de ratón anti-LPS O157 (Χ) (Gene-Tex 3011), anti-fagotopo SFgtp12 (■), anti-LPS O157 (▲). Como control negativo de la reactividad del suero anti-SP12 se utilizó SP3 (□). El análisis estadístico entre las DO determinadas con los sueros anti-péptido SP12, anti-LPS O157 y el monoclonal de ratón anti-LPS O157 a una dilución 1:400 mostraron diferencias significativas con respecto a las DO de suero anti-SP12 contra SP3. El mismo análisis de las DO determinadas con el suero anti-fagotopo SFgtp12 comparadas con las DO del suero anti-SP12 contra PS3 mostraron diferencias significativas (p<0.05) a una dilución 1:200. Valor de las barras de error 0.05.
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
2
1:50 1:100 1:200 1:400
D O
405
nm
Dilución de los sueros
33
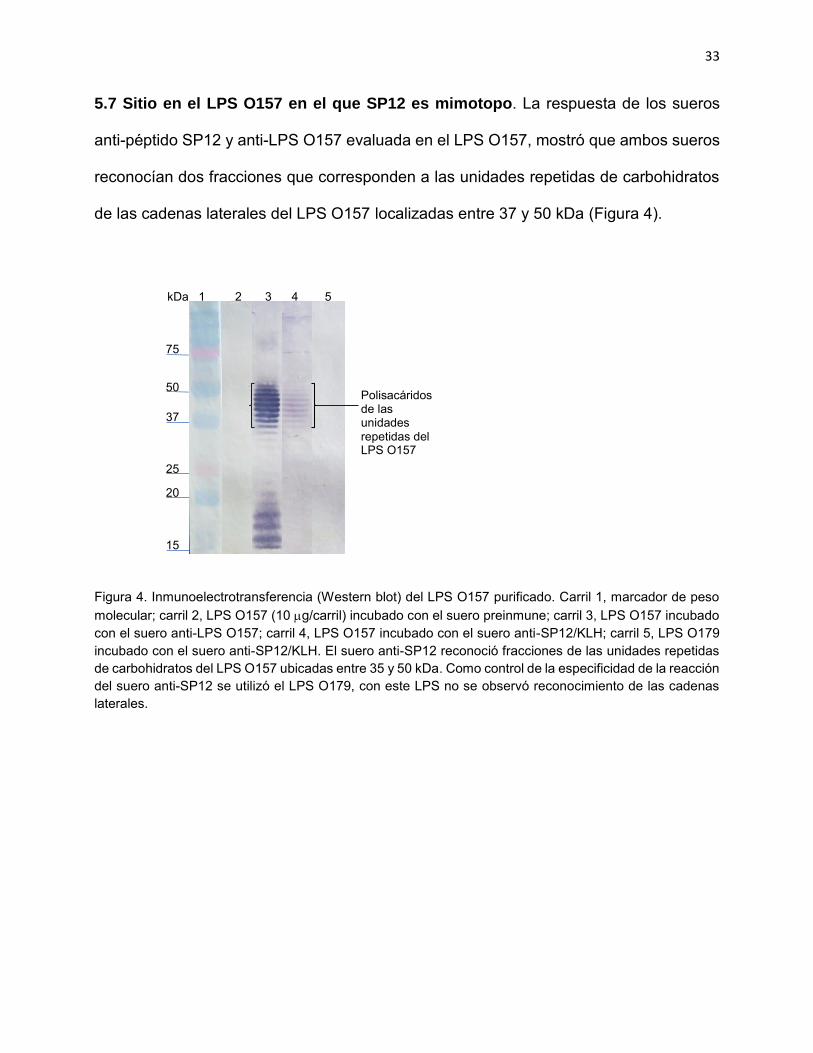
5.7 Sitio en el LPS O157 en el que SP12 es mimotopo. La respuesta de los sueros
anti-péptido SP12 y anti-LPS O157 evaluada en el LPS O157, mostró que ambos sueros
reconocían dos fracciones que corresponden a las unidades repetidas de carbohidratos
de las cadenas laterales del LPS O157 localizadas entre 37 y 50 kDa (Figura 4).
Figura 4. Inmunoelectrotransferencia (Western blot) del LPS O157 purificado. Carril 1, marcador de peso molecular; carril 2, LPS O157 (10 g/carril) incubado con el suero preinmune; carril 3, LPS O157 incubado con el suero anti-LPS O157; carril 4, LPS O157 incubado con el suero anti-SP12/KLH; carril 5, LPS O179 incubado con el suero anti-SP12/KLH. El suero anti-SP12 reconoció fracciones de las unidades repetidas de carbohidratos del LPS O157 ubicadas entre 35 y 50 kDa. Como control de la especificidad de la reacción del suero anti-SP12 se utilizó el LPS O179, con este LPS no se observó reconocimiento de las cadenas laterales.
75
50
37
25 20
15
kDa 1 2 3 4 5
Polisacáridos de las unidades repetidas del LPS O157
34
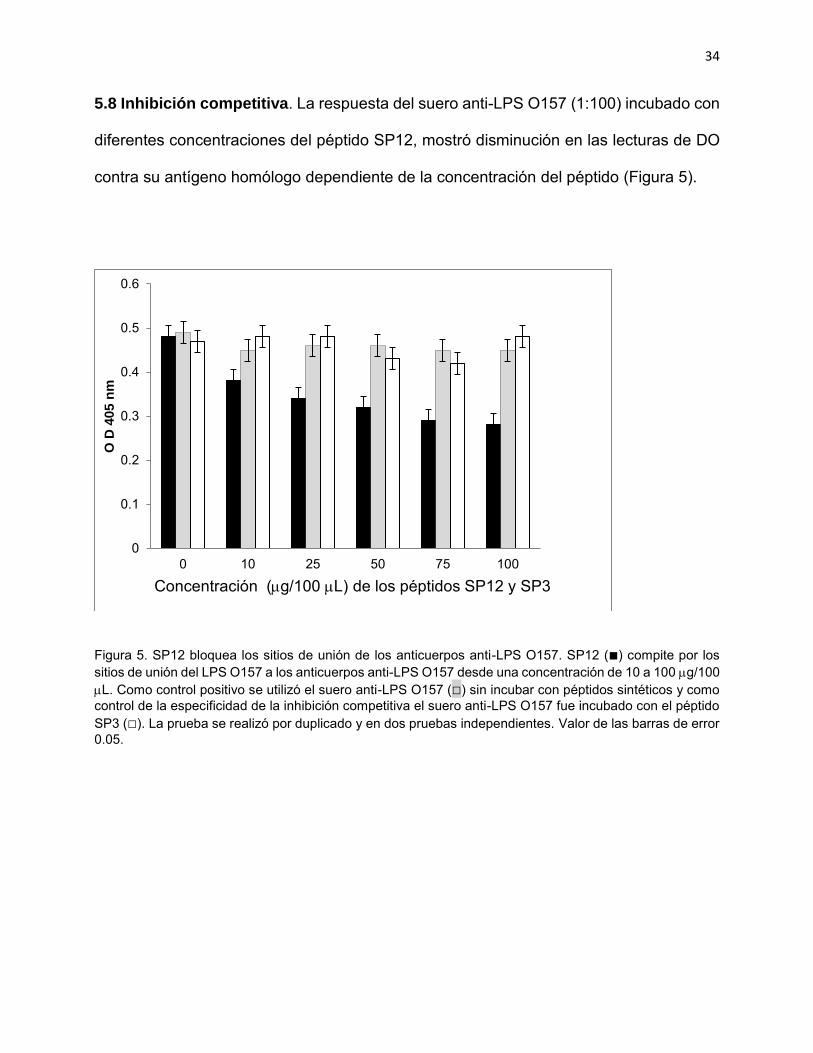
5.8 Inhibición competitiva. La respuesta del suero anti-LPS O157 (1:100) incubado con
diferentes concentraciones del péptido SP12, mostró disminución en las lecturas de DO
contra su antígeno homólogo dependiente de la concentración del péptido (Figura 5).
Figura 5. SP12 bloquea los sitios de unión de los anticuerpos anti-LPS O157. SP12 (■) compite por los sitios de unión del LPS O157 a los anticuerpos anti-LPS O157 desde una concentración de 10 a 100 g/100 L. Como control positivo se utilizó el suero anti-LPS O157 (□) sin incubar con péptidos sintéticos y como control de la especificidad de la inhibición competitiva el suero anti-LPS O157 fue incubado con el péptido SP3 (□). La prueba se realizó por duplicado y en dos pruebas independientes. Valor de las barras de error 0.05.
0
0.1
0.2
0.3
0.4
0.5
0.6
0 10 25 50 75 100
O D
405 n
m
Concentración (g/100 L) de los péptidos SP12 y SP3
35
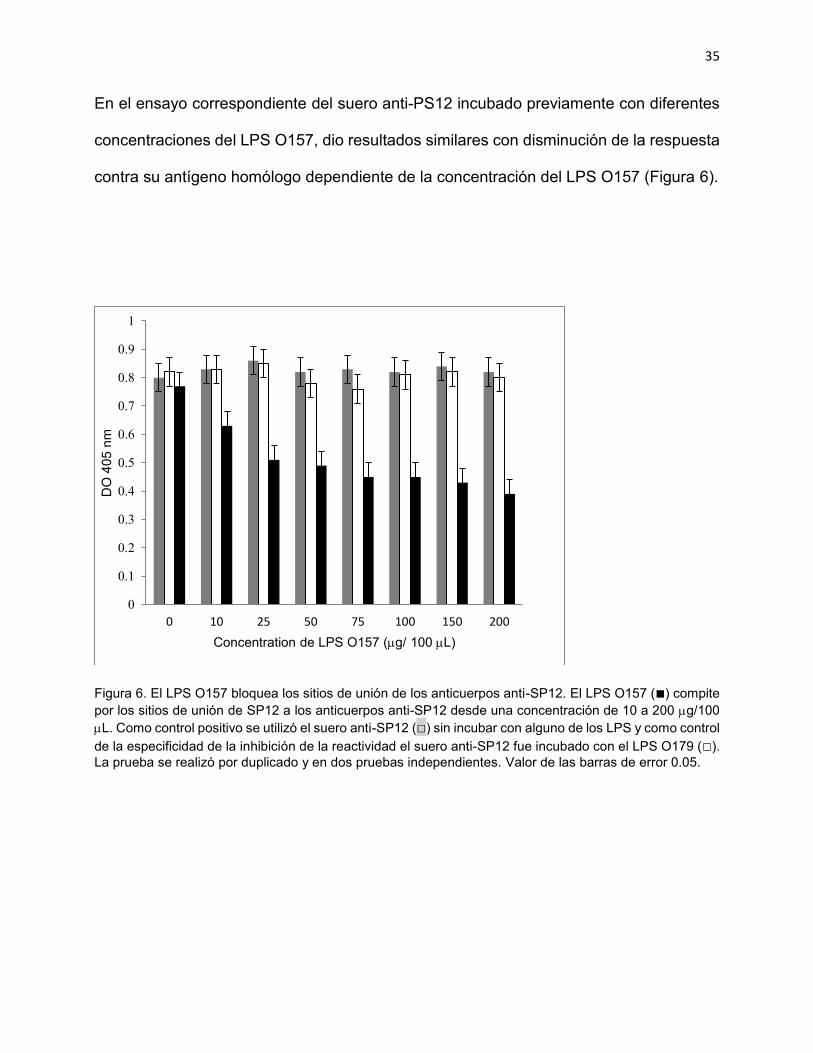
En el ensayo correspondiente del suero anti-PS12 incubado previamente con diferentes
concentraciones del LPS O157, dio resultados similares con disminución de la respuesta
contra su antígeno homólogo dependiente de la concentración del LPS O157 (Figura 6).
Figura 6. El LPS O157 bloquea los sitios de unión de los anticuerpos anti-SP12. El LPS O157 (■) compite por los sitios de unión de SP12 a los anticuerpos anti-SP12 desde una concentración de 10 a 200 g/100 L. Como control positivo se utilizó el suero anti-SP12 (□) sin incubar con alguno de los LPS y como control de la especificidad de la inhibición de la reactividad el suero anti-SP12 fue incubado con el LPS O179 (□). La prueba se realizó por duplicado y en dos pruebas independientes. Valor de las barras de error 0.05.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0 10 25 50 75 100 150 200
DO
405
nm
Concentration de LPS O157 (g/ 100 L)
36
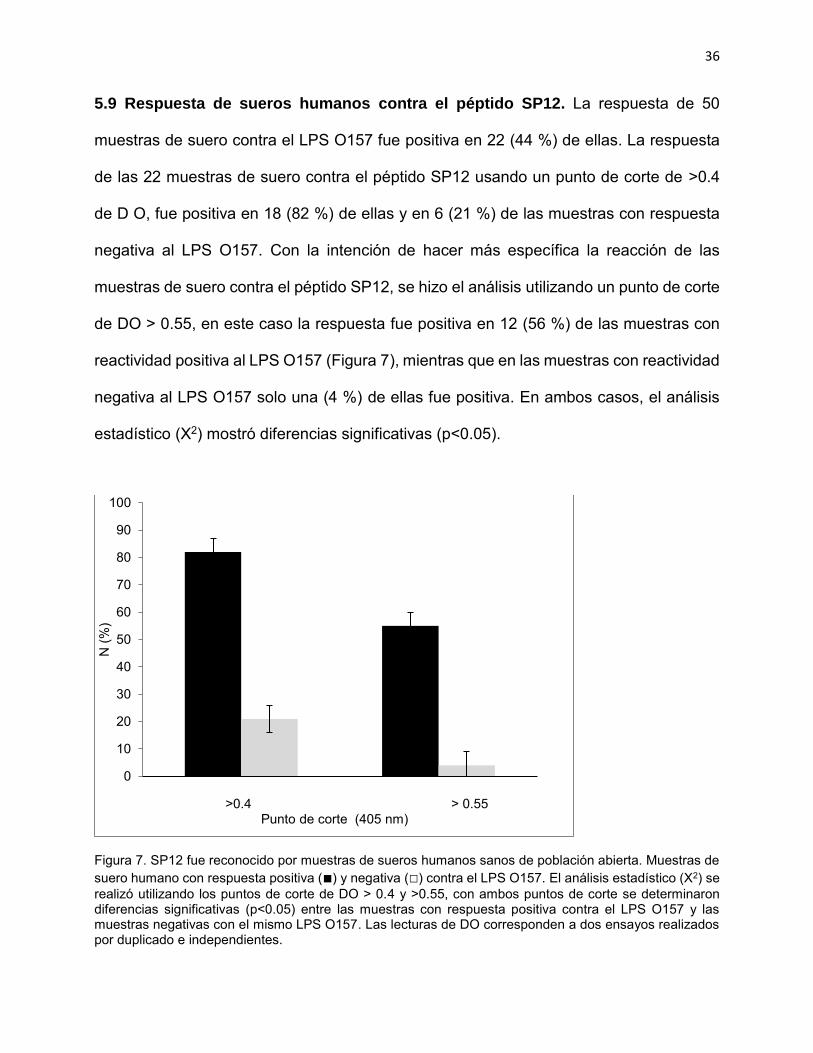
5.9 Respuesta de sueros humanos contra el péptido SP12. La respuesta de 50
muestras de suero contra el LPS O157 fue positiva en 22 (44 %) de ellas. La respuesta
de las 22 muestras de suero contra el péptido SP12 usando un punto de corte de >0.4
de D O, fue positiva en 18 (82 %) de ellas y en 6 (21 %) de las muestras con respuesta
negativa al LPS O157. Con la intención de hacer más específica la reacción de las
muestras de suero contra el péptido SP12, se hizo el análisis utilizando un punto de corte
de DO > 0.55, en este caso la respuesta fue positiva en 12 (56 %) de las muestras con
reactividad positiva al LPS O157 (Figura 7), mientras que en las muestras con reactividad
negativa al LPS O157 solo una (4 %) de ellas fue positiva. En ambos casos, el análisis
estadístico (X2) mostró diferencias significativas (p<0.05).
Figura 7. SP12 fue reconocido por muestras de sueros humanos sanos de población abierta. Muestras de suero humano con respuesta positiva (■) y negativa (□) contra el LPS O157. El análisis estadístico (X2) se realizó utilizando los puntos de corte de DO > 0.4 y >0.55, con ambos puntos de corte se determinaron diferencias significativas (p<0.05) entre las muestras con respuesta positiva contra el LPS O157 y las muestras negativas con el mismo LPS O157. Las lecturas de DO corresponden a dos ensayos realizados por duplicado e independientes.
0
10
20
30
40
50
60
70
80
90
100
N (%
)
>0.4 > 0.55 Punto de corte (405 nm)

37
6. DISCUSIÓN
La importancia clínica y epidemiológica de E. coli O157:H7 como patógeno responsable
de la colitis hemorrágica y el síndrome urémico hemolítico ha sido ampliamente
documentada [92, 93]. La aparición de brotes de ambos cuadros clínicos asociados con
ésta bacteria ha dado lugar a la propuesta de medidas que ayuden a controlar la
transmisión e infección por E. coli O157:H7. Una de estas propuestas para controlar las
enfermedades diarreicas consiste en la elaboración de vacunas. En el caso de E. coli
O157 se han desarrollado algunas vacunas [94, 95, 96, 97], ninguna de las cuales ha
dado los resultados esperados. En diferentes trabajos [55, 56, 57], se ha identificado que
la respuesta inmune por anticuerpos contra el LPS de la bacteria adquirida de manera
natural participa en la protección contra la infección por E. coli O157. Sin embargo, para
el caso de humanos, el LPS O157 no se puede utilizar como inmunógeno por el efecto
tóxico del lípido A que forma parte de la molécula [38]. Por otra parte, Konadu, Ahmed y
Szu [63, 67, 68], refieren que la respuesta de anticuerpos contra el LPS O157 se realiza
principalmente contra las cadenas laterales de carbohidratos. Sin embargo, el uso de
estas cadenas como inmunógenos, está limitado por la corta duración que tiene este tipo
de respuesta inmune de tipo timo-independiente [98]. Ambas situaciones han conducido
a buscar alternativas que permitan contar con compuestos con capacidad inmunogénica
capaces de inducir una respuesta protectora contra la bacteria sin los efectos tóxicos del
LPS.
El procedimiento de Phage display se ha utilizado con buenos resultados para la
identificación de péptidos mimotopos de epitopos inmunodominantes presentes en el LPS
de diferentes bacterias [74, 75, 76]. En el presente trabajo se logró identificar un péptido

38
mimotopo de un epitopo del LPS O157 de E. coli utilizando éste procedimiento. Este
péptido tiene capacidad inmunogénica y es capaz de inducir la producción de anticuerpos
que reaccionaron contra el LPS O157. La selección del fagotopo que expresa este
péptido mimotopo sólo se realizó con IgG de conejos inmunizados con el LPS O157
purificado pero no con IgG de conejos inmunizados con la bacteria completa. Esto sugiere
que el uso del LPS O157 purificado probablemente favorece la expresión de algunos
epitopos que indujeron la producción de anticuerpos específicos capaces de reaccionar
con fagotopos portadores de los péptidos mimotopos. La traducción de la secuencia del
DNA de 15 fagotopos seleccionados con las IgG anti-LPS O157 mostró la presencia de
Ser, Tre, Leu, Ile, Val y Pro como aminoácidos más frecuentes en los péptidos. El análisis
de las secuencias de los péptidos referidos mostró en cinco (33 %) la secuencia consenso
LxP/YP/SxL, la cual contiene Leu, Ser y Tyr, con dos Prolinas (imino ácido cíclico) en el
centro de la secuencia. La presencia de las prolinas es relevante ya que es un aminoácido
que confiere un limitado grado de libertad que influye en la rigidez de la estructura
secundaria de las proteínas [99]. Dharmasena [77] ha descrito también que la prolina es
importante para la reactividad de péptidos contra anticuerpos monoclonales anti-LPS de
V. cholerae O1 Ogawa. Por otro lado, la presencia de Pro en la secuencia
SLSPPLSGLGRA del péptido mimotopo SP12 pudiera contribuir en forma importante a
la respuesta de los anticuerpos contra éste péptido. La prolina es un aminoácido
hidrofóbico cíclico, Angostino [100] señala que péptidos con aminoácidos que poseen
cadenas laterales hidroxiladas (Tyr, Thr y Ser) y no polares como Leu y Trp, además de
Pro mimetizan mejor la estructura de carbohidratos. Así mismo, se ha identificado que
péptidos con motivos Tyr-Pro-Tyr [82] actúan como mimotopos de carbohidratos de la
pared celular de bacterias. Gevorkian [84] con el procedimiento de Phage display realizó

39
la selección de péptidos mimotopos de la pared celular de Mycobacterium tuberculosis
constituida por arabinomanana y lipoaribinomanana. Las secuencias de los péptidos
seleccionados identificadas en dicho estudio (Pro-Leu-X-Gly-[Thr/Val]-Leu-Pro)
presentaban Pro y Leu, aminoácidos que se identificaron en la secuencia consenso
(LxP/YP/SxL) encontradas en el presente estudio. Se ha reportado también que
aminoácidos con residuos no polares y aromáticos pueden mimetizar carbohidratos [82,
101], como sería el caso de la secuencia consenso (LxP/YP/SxL) seleccionada por las
IgG del suero anti-LPS O157 que contiene la presencia de Pro y Tyr, (aminoácidos, cíclico
y con un anillo aromático).
Con el fin de corroborar que algunos aminoácidos en la secuencia de SP12 eran
importantes en la reactividad contra los anticuerpos se comparó con la secuencia de
aminoácidos de SP3 (HTWSFSNALFGG) que no presentó el motivo consenso. Aunque
SP3 contiene los aminoácidos (Trp y Phe) que flanquean a una Ser, en este caso estudios
previos se ha mostrado que dichos aminoácidos están presentes en motivos que
mimetizan carbohidratos [100], al analizar por ensayo de ELISA la reactividad de
anticuerpos anti-LPS O157 y anti-SP12 contra SP3, la respuesta contra este fue pobre.
La limitada antigenicidad de SP3 puede deberse a que no contiene un dipéptido de
Prolina. Un estudio con un péptido mimotopo de perosamina (4-P8) de las cadenas
laterales del LPS de V. cholerae Ogawa con la secuencia Pro-Pro-Leu-Ser-Leu-Leu [77],
mostró que el dipéptido de Pro-Pro-, en las posiciones 5 y 6, y Leu en la posición 9 del
péptido eran importantes en la reactividad del péptido mimotopo contra anticuerpos
monoclonales obtenidos contra el antígeno O del LPS de V. cholerea Ogawa. En dicho
estudio al realizar la substitución de las Pro y Leu por Ala se observó disminución

40
importante en la reactividad de los anticuerpos monoclonales contra el péptido mimotopo.
En nuestro estudio tanto el mimetismo observado con las cadenas laterales del LPS
O157, como la reactividad con el suero anti-LPS O157, pudiera estar relacionado con el
dipéptido Pro-Pro y Leu en las posiciones 4, 5 y 9 de SP12, aminoácidos que como se
hizo referencia resultan importantes para la reactividad de anticuerpos monoclonales
contra el LPS de V. cholerea. La importancia de prolinas también ha sido reportada en el
caso de péptidos mimotopos del LPS de Shigella flexneri [74]. En este último estudio se
refiere que la presencia de Prolina en el péptido mimotopo, participa en la unión de
anticuerpos monoclonales obtenidos contra el LPS de S. flexneri. Finalmente, la baja
respuesta del suero anti-LPS O157 confirma que la presencia de ciertos aminoácidos son
necesarios para que el péptido mimetice la fracción inmunodominante del LPS O157.
La capacidad inmunogénica de algunos de los péptidos sintéticos utilizados en este
estudio quedo demostrada cuando se logró la producción de anticuerpos específicos en
conejos inmunizados con estos antígenos. Para definir el posible sitio del cual SP12 es
mimotopo en el LPS O157 se realizó un ensayo de Western-blot utilizando el LPS O157
completo, en este ensayo se estableció respuesta con ambos sueros, (anti-LPS O157 y
anti-SP12), el sitio de interacción se localizó en la región que corresponde a las unidades
repetidas de los carbohidratos de las cadenas laterales del LPS. Observando que la
respuesta del suero anti-SP12 no dio el mismo patrón de reconocimiento que el suero
anti-LPS O157 lo anterior puede deberse a que el motivo LxP/YP/SxL presente en SP12
sólo mimetiza una fracción del LPS O157. Esta situación ha sido referida en otros trabajos
en los que se han identificado péptidos mimotopos de carbohidratos [102]. Van
Regenmortel y Deroo [103, 104] y sugieren que para considerar un péptido como
mimotopo se requiere que este reaccione con su anticuerpo homólogo y que los

41
anticuerpos obtenidos contra el péptido reaccionen con el antígeno natural. Los
resultados obtenidos con SP12 cumplen a satisfacción con lo propuesto por estos grupos
de investigadores [103, 104]. Los resultados anteriores se confirmaron con los ensayos
de competencia con anticuerpos anti-SP12 y anti-LPS O157 saturados con el LPS y el
péptido respectivamente. Los ensayos muestran que la reactividad del suero anti-LPS
O157 contra su antígeno homólogo no se eliminó completamente al incubarlo con SP12,
aunque la reactividad disminuyo sugiriendo que el péptido compite por los sitios de unión
del anticuerpo anti-LPS O157. Un resultado similar se observó al incubar el suero anti-
SP12 con el LPS O157 para evaluar su reactividad contra el péptido sintético. La
propiedad de inmunogenicidad y la capacidad de péptidos mimotopos para inducir
protección ha sido analizada para diferentes microorganismos. Hai-Bo [105] mostró que
un mimotopo del LPS de una enterobacteria conjugado con KLH (Blue Carrier, BC) fue
capaz de inducir una respuesta protectora mediada por anticuerpos en ratones BALB/c
desafiados con Salmonella Typhi y con E. coli. Dharmasena [77] por su parte, reportó que
ratones inmunizados con el LPS de Vibrio cholerae Ogawa precedida por la aplicación
de un refuerzo con el péptido 4P-8-KLH (mimotopo del LPS) favorece la inducción
prolongada de IgG e IgM contra el LPS Ogawa. En el estudio anterior se observó además
que los anticuerpos presentaban actividad bactericida, por lo que los autores hicieron la
propuesta de utilizarlo como inmunógeno alterno al LPS de V. cholerae O1 Ogawa.
Además de la respuesta en conejos, en el presente trabajo se analizó la respuesta de un
lote de sueros de humanos con reactividad contra el LPS O157 contra el péptido sintético
SP12. Los resultados en estos estudios mostraron que más de la mitad de los sueros
fueron capaces de reaccionar también contra SP12 en comparación con la escasa

42
respuesta al SP12 encontrada en sueros humanos sin respuesta al LPS O157. Lo anterior
corrobora que el péptido SP12 es un mimotopo de un epitopo inmunodominante del LPS
O157, que presenta propiedades antigénicas e inmunogénicas.
La respuesta de los sueros humanos contra el péptido SP12 tienen correlación con dos
estudios independientes realizados por Tang y Thong [106, 107], quienes reportaron la
respuesta inmune de suero de pacientes infectados con S. Typhi contra fagotopos que
expresaban péptidos mimotopos del polisacárido capsular del antígeno Vi de S. Typhi.
En estos estudios se propone que los mimotopos que reaccionan con los sueros anti-
antígeno Vi forman parte de determinantes antigénicos de un epitopo inmunodominante.
En nuestro estudio los resultados observados con la respuesta de sueros humanos contra
SP12 son similares, lo que en cierta forma confirma que SP12 es mimotopo de un epitopo
inmunodominante del LPS O157. La composición estructural del LPS O157 es ([→4)-β-
D-Glc-(1→3)-α-D-PerNAc-(1→4)-α-D-GalNAc-(1→3)-α-l-Fuc-(1→]n), en este se
determinó la presencia de α-D-PerNAc (4-acetamido-4,6-dideoxy-D-mannose)
carbohidrato que se ha reportado como posible determinante antigénico relacionado con
la respuesta inmune y la reactividad antigénica cruzada entre el LPS de diferentes
bacterias Gram-negativas (Brucella abortus, E. hermannii, Salmonella spp. del grupo N)
[108], por ensayos previos con diferentes LPS (datos no mostrados) se observó que suero
anti SP12 reacciona con algunos de ellos, por lo que consideramos que el péptido
mimotopo SP12 pudiera corresponder a α-D-PerNAc.
Los resultados de nuestro estudio muestran que mediante el procedimiento de Phage
display, fue posible seleccionar péptidos mimotopos del LPS O157 con propiedades
inmunogénicas, cuyos anticuerpos reconocen al LPS O157 y compiten por los sitios de

43
unión del anticuerpo con el LPS. Sin embargo, se requieren más estudios para proponer
a estos péptidos mimotopos como inmunógenos alternativos para inducir una respuesta
protectora contra la infección por E. coli O157:H7 evitando el riesgo de usar el LPS O157
como inmunógeno.

44
7. CONCLUSIONES
1. Se identificó un péptido que es un mimotopo de la región de las cadenas laterales del
LPS de E. coli O157.
2. El péptido sintético (SP12) diseñado con la secuencia del mimotopo acoplado a KLH,
indujo la producción de anticuerpos que reaccionaron con el LPS de E. coli O157.
2. Se confirmó que el péptido sintético 12 (SP12) es un mimotopo del LPS de E. coli O157
ya que compite por los sitios de unión de anticuerpos anti LSP O157.
3. Anticuerpos de sueros humanos con reactividad positiva con el LPS de E. coli O157,
reaccionan con el péptido sintético (SP12), lo que reafirma que el péptido es mimotopo
de un sitio inmunodominante del LPS O157.
4. Los resultados generados en el presente trabajo dan elementos para proponer el
empleo de péptidos sintéticos como SP12, como inmunógenos protectores contra las
infecciones por E. coli O157.

45
8. PERSPECTIVAS
1. Estudios previos en el laboratorio hemos determinado que los anticuerpos anti-LPS
O157 reconocen a los LPS de Salmonella Urbana, S. Arizonae y S. Typhi por lo que
proponemos conocer sí los anticuerpos anti-SP12 reconocen a los LPS de Salmonella
spp.
2. El LPS de E. coli O157 lo componen los carbohidratos [→4)-β-D-Glc-(1→3)-α-D-
PerNAc-(1→4)-α-D-GalNAc-(1→3)-α-l-Fuc-(1→]n, en donde α-D-PerNAc es 4-
acetamido-4,6-dideoxy-D-mannosa, este último carbohidrato se reporta que pudiera ser
el epitopos antigénico relacionado con la reactividad antigénica cruzada con los LPS de
Salmonella spp., por lo que se propone establecer sí este carbohidrato es la estructura
que SP12 mimetiza.
3. Realizar el modelado de la estructura de SP12 con su interacción con los anticuerpos
anti-LPS O157 para establecer los grupos funcionales de los aminoácidos que están
relacionados el mimetismo de SP12.
4. Diseñar un modelo animal (ratones CD1) para mostrar que las propiedades
inmunogénicas de SP12 protegen a los animales contra la infección de E. coli O157.
5. Realizar biopanning con IgG anti-Salmonella spp. para obtener péptidos mimotopos
del LPS de Salmonella que comparte epitopos inmunodominantes con E. coli O157, con
lo anterior se pudiera diseñar la formulación de un inmunógeno polivalente contra
enterobacterias que comparten epitopos inmunodominantes comunes.

46
9. LITERATURA CITADA
1. Organización Mundial de la Salud (OMS). Organización Mundial de la Salud: OMS. [Online]. [cited 2015 Marzo 17. Available from: http://www.who.int/mediacentre/factsheets/fs330/es/.
2. Walker C.L.F., Perin J., Aryee M.J., Boschi-Pinto C., and Black R.E. (2012) Diarrhea incidence in low- and middle-income countries in 1990 and 2010: a systematic review. BMC Public Health 12, 220-.
3. Kosek M., Bern C., and Guerrant R.L. (2003) The global burden of diarrhoeal disease, as estimated from studies published between 1992 and 2000. Bull. World Health Organ. 81, 197-204.
4. Dirección General de Epidemiología (DGE). [Online].; 2012 [cited 2014 Noviembre 25. Disponible en: http://www.epidemiologia.salud.gob.mx/doctos/infoepid/vig_epid_manuales/1_2012_Manual_NuTraVe_vFinal_6nov12.pdf.
5. Ewing W. H. (1986) Edwards and Ewing´s identification of Enterobacteriaceae. Elsevier Science Publishing Inc., New York.
6. Lior H. 1994. (1994) Classification of Escherichia coli. In: Escherichia coli in domestic animals and humans, Gyles, C.L. (ed). Wallingford (United Kingdom): CAB International.- ISBN 0-85198-921-7. p. 31-72.
7. Kauffmann F. (1944) Zur serology der coli-gruppe. Acta Path. Et. Microb. Sacand. 21, 20-45.
8. Knipschildt H. E. (1945) Undersogelser over coligruppens serology med saerligt Henblik paa Kapselformerne. Nyt Nordisk Forlag; Arnold Busck, Copenhagen.
9. Kauffmann F. (1947) The serology of the coli group. J. Immunol. 57, 71-100. 10. Scheutz F., Cheasty T., Woodward D., and Smith H. R. (2004) Designation of O174
and O175 to temporary O groups OX3 and OX7, and six new E. coli O groups that include Verocytotoxin-producing E. coli (VTEC): O176, O177, O178, O179, O180 and O181. APMIS 112, 569–584.
11. Neter E., Westphal O., Lüderitz O., Gino R.M., and Gorzynski E.A. (1955) Demonstration of antibodies against enteropathogenic Escherichia coli in sera of children of various ages. Pediatrics 16, 801-808.
12. Levine M.M. (1987) Escherichia coli that cause diarrhea: enterotoxigenic, enteropathogenic, enteroinvasive, enterohemorrhagic, and enteroadherent. J. Infect. Dis. 155, 377-389.
13. Nataro J., and Kaper J. (1998) Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 11, 142-201.
14. Kaper J., Nataro J., Mobley H. (2004) Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2, 123-140.
15. Clements A., Young J.C., Constantinou N., and Frankel G. (2012) Infection strategies of enteric pathogenic Escherichia coli. Gut Microbes. 3, 71-87.
16. Riley L.W., Remis R.S., Helgerson S.D., McGee H.B., Wells J.G., David B.R., Hebert R.J., Olcott E.S., Johnson L.M., Hargrett N.T., et al. (1983) Hemorrhagic colitis associated with a rare Escherichia coli serotype. N. Engl. J. Med. 308, 681-85.

47
17. Karmali M. A., Steele B.T., Petric M., and Lim C. (1983) Sporadic cases of haemolytic-uraemic syndrome associated with faecal cytotoxin and cytotoxin-producing Escherichia coli in stools. Lancet I, 619-20.
18. Barrett T.J., Lior H., Green J.H., Khakhria R., Wells J.G., Bell B.P., Greene K.D., Lewis J., and Griffin P.M. (1994) Laboratory investigation of a multistate food-borne outbreak of Escherichia coli O157:H7 by using pulsed-field gel electrophoresis and phage typing. J. Clin. Microbiol. 32, 3013-17.
19. Bitzan M., Ludwig K., Klemt M., König H., Büren J., and Müller-Wiefel D.E. (1993) The role of Escherichia coli O 157 infections in the classical (enteropathic) haemolytic uraemic syndrome: results of a Central European, multicentre study. Epidemiol. Infect. 110, 183-196.
20. Watanabe H., Wada A., Inagak Y., and Tamura K. (1996) Outbreaks of enterohaemorrhagic Escherichia coli O157:H7 infection by two different genotype strains in Japan. Lancet 348, 831-32.
21. Hancok D.D., Rice D.H., Thomas L.A., Dargatz D.A., and Besser T.E. (1997) Epidemiology of Escherichia coli O157 in feedlot cattle. J. Food Prot. 60, 462-5.
22. Chapman P.A., Siddons C.A, Gerdan Malo A.T., and Harkin M.A. (1997) A 1-year study of Escherichia coli O157 in cattle, sheep, pigs and poultry. Epidemiol. Infect. 119, 245–250.
23. Rabatsky-Ehr T., Dingman D., Marcus R., Howard R., Kinney A., and Mshar P. (2002) Deer meat as the source for a sporadic case of Escherichia coli O157:H7 infection. Emerg. Infect. Dis. 8, 525-527.
24. Nakazawa M., and Akiba M. (1999) Swine as a potential reservoir of Shiga toxin-producing Escherichia coli O157: H7 in Japan. Emerg. Infect. Dis. 5, 833.
25. Center for Disease Control and Prevention (CDC). (1993) Update: multistate outbreak of Escherichia coli O157:H7 infections from hamburgers - Westerns United States, 1992-1993. MMWR Morb. Mortal. Wkly. Rep. 42, 15-38.
26. Keene W.E., Hedberg K., Herriott D.E, Hancock D.D., McKay R.W., Barrett T.J., and Fleming D.W. (1997) A prolonged outbreak of Escherichia coli O157:H7 infections caused by commercially distributed raw milk. J. Infect. Dis. 176, 815-818.
27. Breuer T., Benkel D.H., Shapiro, R.L., Hall W.N., Winnett M.M., Linn M.J., Neimann J., Barrett T.J., Dietrich S., Downes F.P., et al. (2001) A multistate outbreak of Escherichia coli O157:H7 infections linked to alfalfa sprouts grown from contaminated seeds. Emerg. Infect. Dis. 7, 977-982.
28. Chekabab S.M., Paquin-Veillette J., Dozois C.M., and Harel J. (2013) The ecological habitat and transmission of Escherichia coli O157:H7. FEMS Microbiol. Lett. 341, 1-12.
29. Xicohtencatl-Cortes J., Sánchez Chacón E., Saldaña Z., Freer E., and Girón J.A. (2009) Interaction of Escherichia coli O157: H7 with leafy green produce. J. Food Prot. 72, 1531-1537.
30. Konowalchuk J., Speirs J.I., and Stavric S. (1977) Vero response to a cytotoxin of Escherichia coli. Infect. Immun. 18, 775–779.
31. Scheutz F., Teel L.D., Beutin L., Piérard D., Buvens G., Karch H., Mellmann A., Caprioli A., Tozzoli R., Morabito S., et al. (2012) Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 50, 2951-.

48
32. Callaway T.R., Anderson R.C., Tellez G., Rosario C., Nava G.M., Eslava C., Blanco M.A., Quiroz M.A., Olguín A., Herradora M., et al. (2004) Prevalence of Escherichia coli O157 in cattle and swine in Central Mexico. J. Food Protect. 67, 2274-2276.
33. Amézquita-López B.A., Quiñones B., Cooley M.B., León-Félix J., Castro-del Campo, N., Mandrell R.E, Jimenez M., and Chaidez C. (2012) Genotypic analyses of Shiga toxin-producing Escherichia coli O157 and non-O157 recovered from feces of domestic animals on rural farms in Mexico. PloS ONE 7, e51565 10.1371/journal.pone.0051565.
34. Varela-Hernández J.J., Cabrera-Diaz E., Cardona-López M.A., Ibarra-Velázquez L.M., Rangel-Villalobos H., Castillo A., Torres-Vitela M.R., and Ramírez-Alvarez A. (2007) Isolation and characterization of Shiga toxin-producing Escherichia coli O157:H7 and non-O157 from beef carcasses at a slaughter plant in Mexico. Int. J. Food Microbiol. 113, 237-241.
35. Reyes-Rodríguez N., Soriano-Vargas E., Barba-León J., Navarro A., Talavera-Rojas M., Sanso A., and Bustamante A.V. (2015) Genetic characterization of Escherichia coli isolated from cattle carcasses and feces in Mexico State. J. Food Protect. 78, 796-801.
36. Navarro A., Licona D., León L.A., Pérez G., Ariza M.C., Gutiérrez A., Trejo A., Díaz M., Díaz S.P., Meza J.D., Cravioto A., and Eslava C. (2012) Escherichia coli O104 isolation frequency and immune response against O104 LPS in dairy cattle, from different states of Mexico. Special Issue: Abstracts from 8th International Shiga Toxin (Verocytotoxin)-producing Escherichia coli Infection. Zoonoses Public Health 59, 19–90.
37. Raetz C.R.H. (1987) Structure a biosynthesis of lipid A in Escherichia coli. In Neidhardt CF. Ed. Escherichia coli and Salmonella typhimurium. ASM Press Washington D C; 498-503.
38. Beutler B., and Rietschel E. Th. (2003) Innate immune sensing and its roots: the story of endotoxin. Nature Rev. Immunol. 3, 169-176.
39. Appelmelk B.J., An Y-Q., Hekker A.M., Thijs L.G., MacLaren D.M., and Graaf J. (1994) Frequencies of lipopolysaccharide core types in Escherichia coli strains from bacteraemic patients. Microbiology 140, 1119-24.
40. Heinrichs D.E., Jeremy A., Yethon J.A., and Whitfield C. (1998) Molecular basis for structural diversity in the core regions of the lipopolysaccharides of Escherichia coli and Salmonella enterica. Mol. Microbiol. 30, 221–32.
41. Heine H., Rietschel E.T. and Ulmer A.J. (2001) The biology of endotoxin. Mol. Biotechnol. 19, 279-296.
42. Raetz C.R, and Whitfield C. (2002) Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 71, 635–700.
43. Perry M.B., MacLean L., and Griffith D.W. (1986) Structure of O-chain polysaccharide of the phenol-phase soluble lipopolysaccharide of Escherichia coli O157: H7. Biochem. Cell Biol. 64, 21-28.
44. Chart H., Cheasty T., Georgiou T., and Rowe B. (1993) Antigenic cross-reactions between Escherichia coli O157, Vibrio cholera O1 (Inaba) and group N Salmonella. Serodiagnosis and Immunotherapy in Infectious Disease 5, 81-84.
45. Park C., Martin E., and White E. 1998. Isolation of a nonpathogenic strain of Citrobacter sedlakii which expresses Escherichia coli O157 antigen. J. Clin. Microbiol. 36, 1408-1409.

49
46. Nishiuchi Y., Doe M., Hotta H., and Kobayashi K. (2000) Structure and serologic properties of O-specific polysaccharide from Citrobacter freundii possessing cross-reactivity with Escherichia coli O157:H7. FEMS Immunol. Med. Microbiol. 28, 163–171.
47. Rice E., Sowers E., Johnson C., Dunnigan M., Strockbine N., and Edberg S. (1992) Serological cross-reactions between Escherichia coli O157 and other species of the genus Escherichia. J. Clin. Microbiol. 30, 1315-1316.
48. Navarro A., Eslava C., García de la Torre G., León L.A., Licona D., León L., Zarco L. A., and Cravioto A. (2007) Common Epitopes in lipopolysaccharides of different Enterobacteriaceae are associated with an immune response against Escherichia coli O157 in bovine serum samples. J. Med. Microbiol. 56, 1447-1454.
49. Notenboom R.H., Borczyk A., Karmali M.A., and Duncan L.M. (1987) Clinical relevance of a serological cross-reaction between Escherichia coli O157 and Brucella abortus. The Lancet ii, 744.
50. Ludwig K., Sarkim V., Bitzan M., Karmali M.A., Bobrowski C., Ruder H., Laufs R., Sobottka I., Petric M., Karch H., et al. (2002) Shiga toxin-producing Escherichia coli infection and antibodies against Stx2 and Stx1 in household contacts of children with enteropathic hemolytic-uremic syndrome. J. Clin. Microbiol. 40, 1773-1782.
51. Chart H., Scotland S., and Rowe B. (1989) Serum antibodies to Escherichia coli serotype O157:H7 in patients with hemolytic uremic syndrome. J. Clin. Microbiol. 27, 285-290.
52. Greatorex J., and Thorne G. (1994) Humoral immune response to Shiga-Like Toxin and Escherichia coli O157 lipopolysaccharide in hemolytic-uremic syndrome patients and healthy subjects. J. Clin. Microbiol. 32, 1172-1178.
53. Banatvala N, Griffin PM, Greene KD, Barrett TJ, Bibb WF, Green JH, and Wells J G. (2001) The United States national prospective hemolytic uremic syndrome study: microbiologic, serologic, clinical, and epidemiologic findings. J. Infect. Dis. 183, 1063-1070.
54. Siddons C.A., and Chapman P.A. (1993) Detection of serum and faecal antibodies in haemorrhagic colitis caused by Escherichia coli O157. J. Med. Microbiol. 39, 408-415.
55. Reymond D., Johnson R.P., Karmali M.A., Petric M., Winkler M., Johnson S., Rahn K., Renwick S., Wilson J., Clarke R.C., and Spika J. (1996) Neutralizing antibodies to Escherichia coli Vero cytotoxin 1 and antibodies to O157 lipopolysaccharide in healthy farm family members and urban residents. J. Clin. Microbiol. 34, 2053-2057.
56. Evans J., Chalmers R.M., Chart H., Salmon R.L., Kench S.M., Coleman T.J., Meadows D., Morgan-Capner P., Softley P., Sillis M., et al. (2000) Evidence of persisting serum antibodies to Escherichia coli O157 lipopolysaccharide and Verocytotoxin in members of rural communities in England. Eur. J. Epidemiol. 16, 885-9.
57. Haack J.P., Jelacic S., Besser T.E., Weinberger E., Kirk D.J., McKee G.L, Harrison S.M., Musgrave K.J., Miller G., Price T.H., et al. (2003) Escherichia coli O157 exposure in Wyoming and Seattle: serologic evidence of rural risk. Emerg. Infect. Dis. 9, 1226-31.
58. Quilliam R.S., Chalmers R.M., Williams A.P., Chart H., Willshaw G.A., Kench S.M., Edwards-Jones G., Evans J., Thomas D.Rh., Salmon R.L., et al. (2012) Seroprevalence and risk factors associated with Escherichia coli O157 in a farming population. Zoonoses Public Health 59, 83-88.
59. Belongia E.A., Chyou P-H., Greenlee R.T., Perez-Perez G., Bibb W.F., and DeVries E.O. (2003) Diarrhea incidence and farm-related risk factor for Escherichia coli O157:H7 and Campylobacter jejuni antibodies among rural children. J. Infect. Dis. 187,

50
1460–1468. 60. Navarro A., Eslava C., Hernández U., Navarro-Henze., J.L., Avilés M., García de
la Torre G., and Cravioto A. (2003) Antibody response to Escherichia coli O157 and other lipopolysaccharides in healthy children and adults. Clin. Diag. Lab. Immunol. 10, 797-801.
61. Potter A.A., Klashinsky S., Li Y., Frey E., Townsed H., Rogan D., Erickson G., Hinkley S., Klopfenstein T., Moxley R.A., et al. (2004) Decreased shedding of Escherichia coli O157:H7 by cattle following vaccination with type III secreted proteins. Vaccine 22, 362-369.
62. Konadu E., Robbins J.B., Schiloach J., Bryla D.A., and Szu S.C. (1994) Preparation, characterization, and immunological properties in mice of Escherichia coli O157 O-specific polysaccharide-protein conjugate vaccine. Infect. Immun. 62, 5048-5054.
63. Konadu E.Y., Parke J.C., Tran H.T., Bryla D.A., Robin J.B., and Szu S.C. (1998) Investigational vaccine for Escherichia coli O157: phase 1 study of O157 O-specific polysaccharide-Pseudomonas aeruginosa recombinant exoprotein A (rEPA) conjugate in adults. J. Infect. Dis. 177, 383-387.
64. Conlan J.W., Cox A.D., KuoLee R., Webb A., and Perry M.B. (1999) Parenteral immunization with a glycoconjugate vaccine containing the O157 antigen of Escherichia coli O157: H7 elicits a systemic humoral immune response in mice, but fails to prevent colonization by the pathogen. Can. J. Microbiol. 45, 279-286.
65. Conlan J.W., KuoLee R., Webb A., and Perry M.B. (1999) Salmonella landau as a live vaccine against Escherichia coli O157: H7 investigated in a mouse model of intestinal colonization. Can. J. Microbiol. 45, 723-731.
66. Konadu E., Donohue-Rolfe A., Calderwood S.B., Pozsgay V., Shiloach J., Robbins J.B., and Szu S.C. (1999) Syntheses and immunologic properties of Escherichia coli O157 O-specific polysaccharide and Shiga toxin 1 B subunit conjugates in mice. Infect. Immun. 67, 6191-6193.
67. Ahmed A., Li J., Shiloach Y., Robbins J.B., and Szu S.C. (2006) Safety and immunogenicity of Escherichia coli O157 O-specific polysaccharide conjugate vaccine in 2–5-year-old children. J. Infect. Dis. 193, 515-521.
68. Szu S., and Ahmed A. (2014) Clinical Studies of Escherichia coli O157:H7 conjugate vaccines in adults and young children. Microbiol Spectrum 2, EHEC-0016-2013.
69. Nagy G., and Pál T. (2008) Lipopolysaccharide: a tool and target in enterobacterial vaccine development. Biol. Chem. 389, 513-520.
70. Ludwig K., Bitzan M., Bobrowski C., and Müller-Wiefel D.E. (2002) Escherichia coli O157 fails to induce a long-lasting lipopolysaccharide-specific, measurable humoral immune response in children with hemolytic-uremic syndrome. J. Clin. Microbiol. 186, 566-569.
71. Smith G.P. (1985) Filamentous fusion phage: novel expression vectors that display cloned antigens on the virion surface. Science 228, 1315-1317.
72. Smith G.P., and Scott J.K. (1993) Libraries of peptides and proteins displayed on filamentous phage. Methods Enzymol. 217, 228–57.
73. Dower W.J., and Cwirla S.E. (1994) Epitope mapping using libraries of random peptides displayed on phage. In: Wisdom GB, ed. Peptide antigens. A practical approach. New York, Oxford University Press, pp. 219–43.

51
74. Clément M-J., Fortuné A., Phalipon A., Marcel-Peyre V., Simenel C., Imberty A., Delepierre M., and Mulard L.A. (2006) Toward a better understanding of the basis of the molecular mimicry of polysaccharide antigens by peptides: The example of Shigella flexneri 5a. J. Biol. Chem. 281, 2317–32.
75. De Bolle X., Laurent T., Tibor A., Godfoid F., Weynants-Jacques L.V., and Mertens P. (1999) Antigenic properties of peptidic mimics for epitopes of the lipopolysaccharide from Brucella. J. Mol. Biol. 294, 181-91.
76. Grothaus M.C., Srivastava N., Smithson S.L., Kieber-Emmons T., Williams D.B., Carlone G.M., and Westerink M.A.J. (2000) Selection of an immunogenic peptide mimic of the capsular polysaccharide of Neisseria meningitidis serogroup A using a peptide display library. Vaccine 18, 1253-63.
77. Dharmasena M.N., Jewell D.A., and Taylor R.K. (2007) Development of peptide mimics of a protective epitope of Vibrio cholerae Ogawa O-antigen and investigation of the structural basis of peptide mimicry. J. Biol. Chem. 282, 33805–33816.
78. Falklind-Jerkérus S., Felici F., Cavalieri C., Lo Passo C., Garufi G., Pernice I., Islam M.M., Qadri F., and Weintraub A. (2005) Peptides mimicking Vibrio cholerae O139 capsular polysaccharide elicit protective antibody response. Microbes Infect. 7, 1453-60.
79. Ulises H.C., Gazarian T., Gazarian K., Mendoza-Hernández G., Xicohtencatl-Cortes J., and Carlos E. (2009) Peptide sequences identified by phage display are immunodominant functional motifs of Pet and Pic serine proteases secreted by Escherichia coli and Shigella flexneri. Peptides 30, 2127-2135.
80. Rao S.S., Mohan K.V., and Atreya C.D. (2013) A Peptide derived from phage display library exhibits antibacterial activity against E. coli and Pseudomonas aeruginosa. PloS One 8, e56081.
81. Scott J.K., and Smith G.P. (1990) Searching for peptide ligands with an epitope library. Science 27, 386-390.
82. Oldenburg K. R., Loganathan D., Goldstein I.J., Schultz P.G., and Gallop M.A. (1992) Peptide ligands for a sugar-binding protein isolated from a random peptide library. Proc. Natl. Acad. Sci. USA. 89, 5393–5397.
83. Westphal O., and Jann K. (1965) Bacterial lipopolysaccharide: extraction with phenol-water and further applications of the procedure. Meth. Carbohydr. Chem. 5, 83-91.
84. Gevorkian G., Segura E., Acero G., Palma J.P., Espitia C., Manoutcharian K., and López-Marín L.M. (2005) Peptide mimotopes of Mycobacterium tuberculosis carbohydrate immunodeterminants. Biochem. J. 387, 411-17.
85. Norma Oficial Mexicana -NOM-062-ZOO-1999. Especificaciones técnicas para la producción, cuidado y uso de los animales de laboratorio.
86. Green A. A., and Walter L.H. (1955) Protein fractionation on the basis of solubility in aqueous solutions of salts and organic solvents. Methods Enzymol. 1, 67-90.
87. Wilson R.K. (1993) High-throughput purification of M13 templates for DNA sequencing. Biotechniques 15, 414-6.
88. Galfré G., Monaci A., Nicosia A., Luzzago F., Felici F., and Cortese R. (1996) Immunization with phage-displayed mimotopes. Methods Enzymol, 267, 109-115.
89. Phalipon A., Costachel C., Grandjean C., Thuizat A., Guerreiro C., Tanguy M., Nato F., Vulliez-Le Normand B., Bélot F., Wright K., et al. (2006) Characterization of functional oligosaccharide mimics of the Shigella flexneri serotype 2a O-antigen:

52
implications for the development of a chemically defined glycoconjugate vaccine. J. Immunol. 176,1686-1694.
90. Laemmli U.K. (1970) Cleavage of structural protein during the assembly of the head of bacteriophage T4. Nature 227, 680-685.
91. Towbin H., Staehelin T., and Gordon J. (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc. Natl. Acad. Sci. USA. 76, 4350-4354.
92. Tarr P.I., Gordon C.A., and Chandler W.L. (2005) Shiga-toxin-producing Escherichia coli and haemolytic uraemic syndrome. Lancet 365, 1073–1086.
93. Scallan E., Hoekstra R.M., Angulo F.J., Tauxe R.V., Widdowson M.A., Roy S.L., Jones J.L., and Griffin P.M. (2011) Foodborne illness acquired in the United States—major pathogens. Emerg. Infect. Dis. 17, 7-15.
94. Vilte D.A., Larzábal M., Garbaccio S., Gammella M., Rabinovitz B.C., Elizondo A.M., Cantet R.J., Delgado F., Meikle V., Cataldi A., and Mercado E.C. (2011) Reduced faecal shedding of Escherichia coli O157:H7 in cattle following systemic vaccination with γ-intimin C₂₈₀ and EspB proteins. Vaccine 29, 3962-8.
95. Yekta M.A., Goddeeris B.M., Vanrompay D., and Cox E. (2011) Immunization of sheep with a combination of intiminγ, EspA and EspB decreases Escherichia coli O157:H7 shedding. Vet. Immunol. Immunopathol. 140, 42-46.
96. McNeilly T.N., Naylor S.W., Mahajan A., Mitchell M.C., McAteer S., Deane D., Smith D.G., Low J.C., Gally D.L., and Huntley J.F. (2008) Escherichia coli O157:H7 colonization in cattle following systemic and mucosal immunization with purified H7 flagellin. Infect. Immun. 76, 2594-2602.
97. McNeilly T.N., Mitchell M.C., Rosser T., McAteer S., Low J.C., Smith D.G., Huntley J.F., Mahajan A., and Gally D.L. (2010) Immunization of cattle with a combination of purified intimin-531, EspA and Tir significantly reduces shedding of Escherichia coli O157:H7 following oral challenge. Vaccine 28, 1422-8.
98. Kieber-Emmons T., Luo P., Qiu J., Chang T.Y., Insug O., Blaszczyk-Thurin M., and Steplewski Z. (1999) Vaccination with carbohydrate peptide mimotopes promotes anti-tumor responses. Nature Biotech. 17, 660 – 665.
99. MacArthur M.W., and Thornton J.M. (1991) Influence of proline residues on protein conformation. J. Mol. Biol. 218, 397-412.
100. Agostino M., Sandrin M.S., Thompson P.E., Farrugia W., Ramsland P.A., and Yuriev E. (2011) Carbohydrate-mimetic peptides: structural aspects of mimicry and therapeutic implications. Expert. Opin. Biol. Ther. 11, 211-224.
101. Hoess R., Brinkmann U., Handel T., and Pastan I. (1993) Identification of a peptide which binds to the carbohydrate-specific monoclonal antibody B3. Gene 128, 43-9.
102. Phalipon A., Folgori A., Arondel J., Sgaramella G., Fortugno P., Cortese R., Sansonetti P. J., and Felici F. (1997) Induction of anti-carbohydrate antibodies by phage library-selected peptide mimics. Eur. J. Immunol. 27, 2620-2625.
103. Van Regenmortel M.H. (2009) What is a B-cell epitope. Methods Mol. Biol. 524, 3-20.
104. Deroo S., and Muller C.P. (2001) Antigenic and immunogenic phage displayed mimotopes as substitute antigens: applications and limitations. Chem. High Throughput Screen 4, 75-110.

53
105. Hai-Bo L., Jian Z., Ping Z., Lai-Qiang H., and Ning F. (2008) Protective immunity induced by peptide mimics to LPS in mice with endotoxic shock. Prog. Biochem. Biophys. 35, 1312-19.
106. Tang S.S., Tan W.S., Dev S., Wang L.F., Pang T., and Thong K.L. (2003) Mimotopes of the Vi antigen of Salmonella enterica serovar Typhi identified from phage display peptide library. Clin. Diagn. Lab. Immunol. 10, 1078-1084.
107. Thong K.L., Tang S.S., Tan W.S., and Devi S. (2007) Peptide mimotopes of complex carbohydrates in Salmonella enterica serovar Typhi which react with both carbohydrate-specific monoclonal antibody and polyclonal sera from typhoid patients. Microbiol. Immunol. 51, 1045-1052.
108. Nishiuchi Y., Doe M., Hotta H., and Kobayashi K. (2000) Structure and serologic properties of O-specific polysaccharide from Citrobacter freundii possessing cross-reactivity with Escherichia coli O157:H7. FEMS Immunol. Med. Microbiol. 28, 163–171.

54
10. ANEXO. ARTÍCULO REQUISITO

55

56
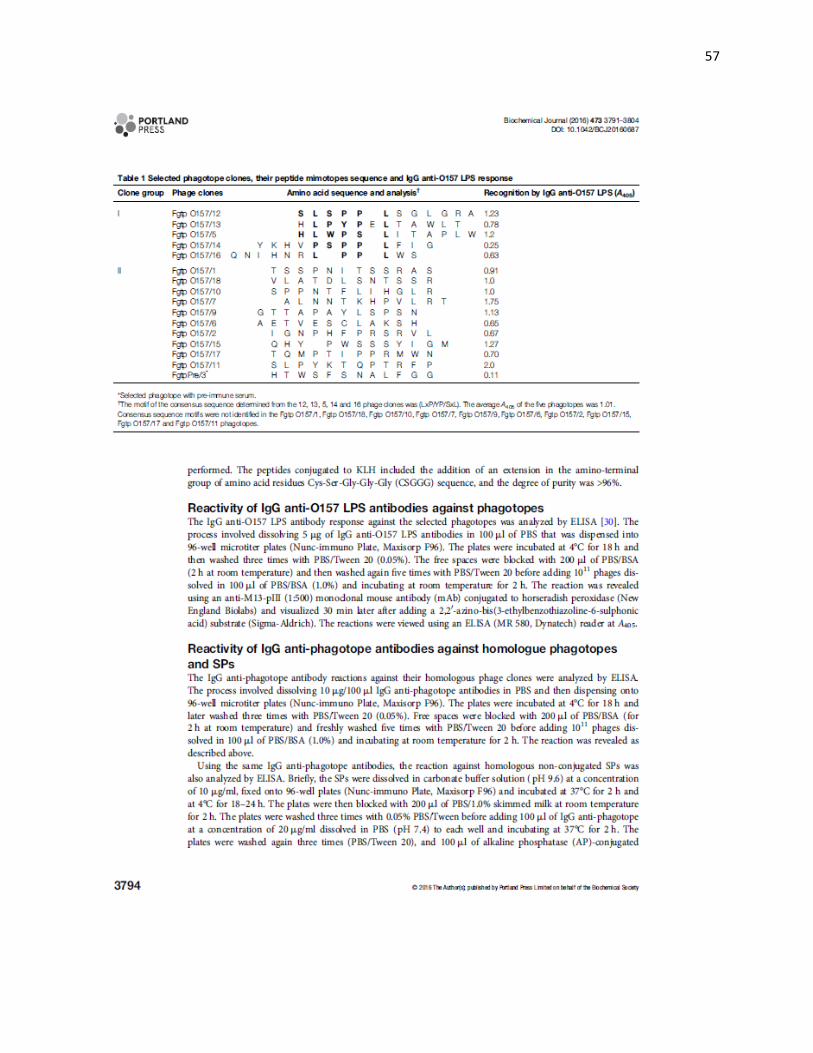
57

58

59

60
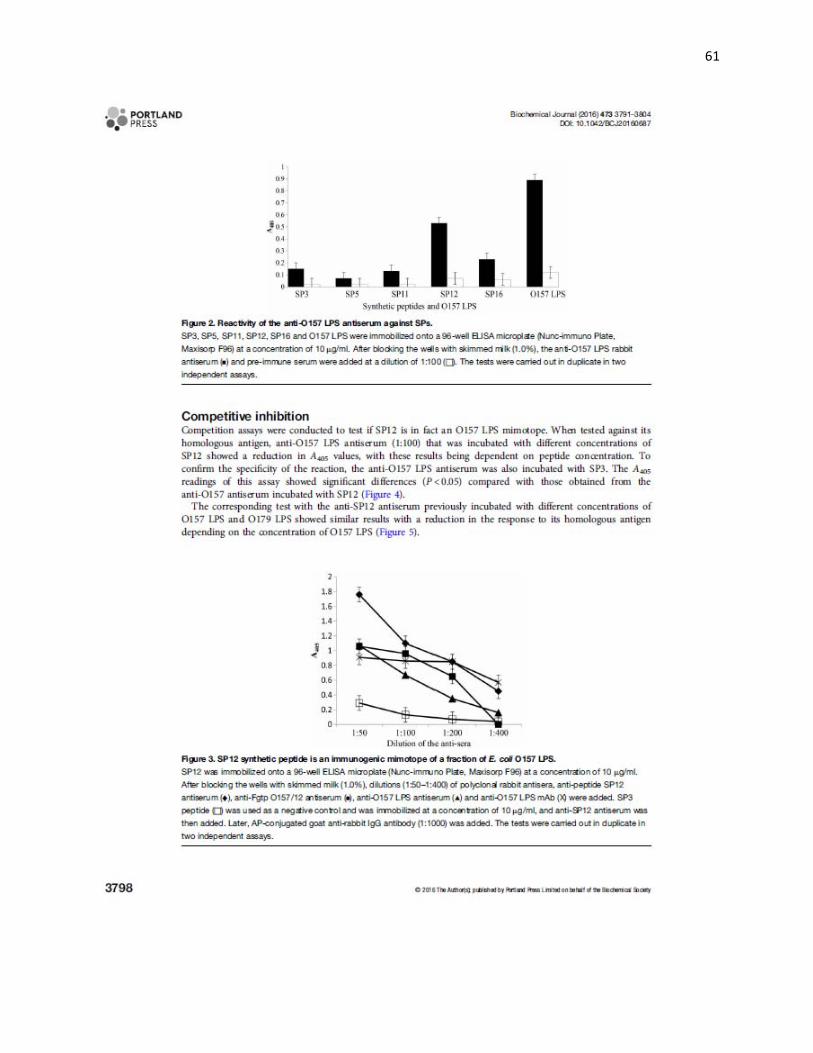
61
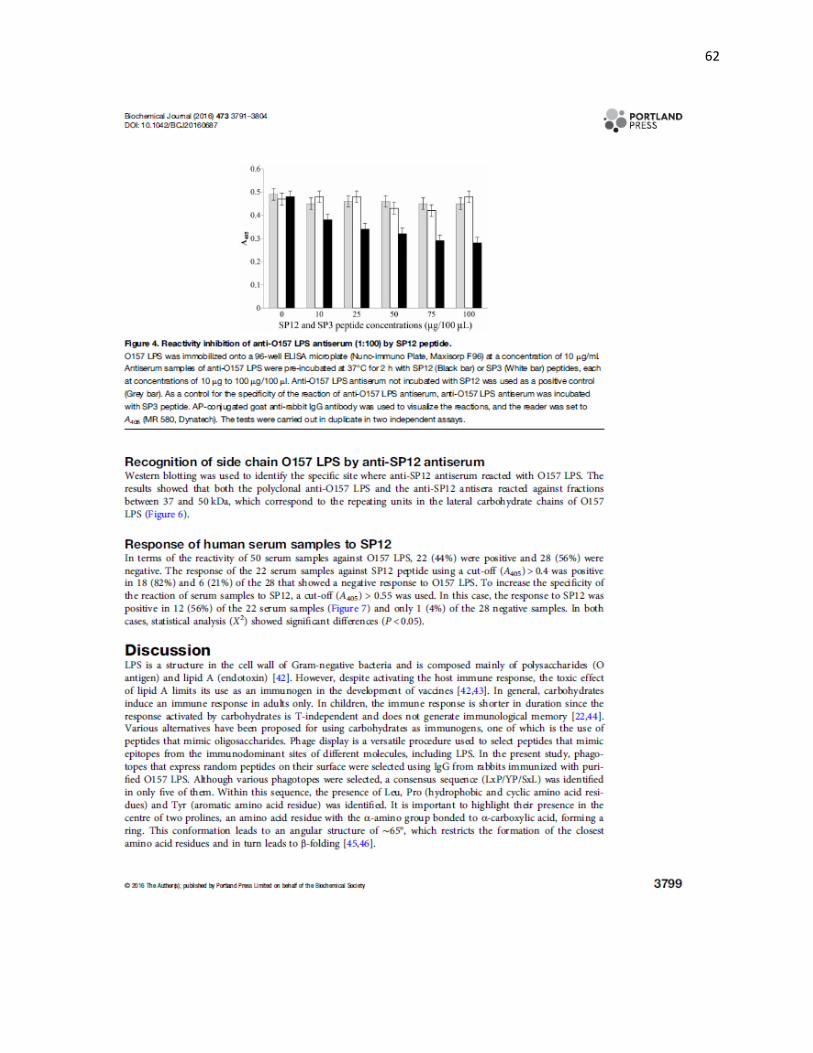
62
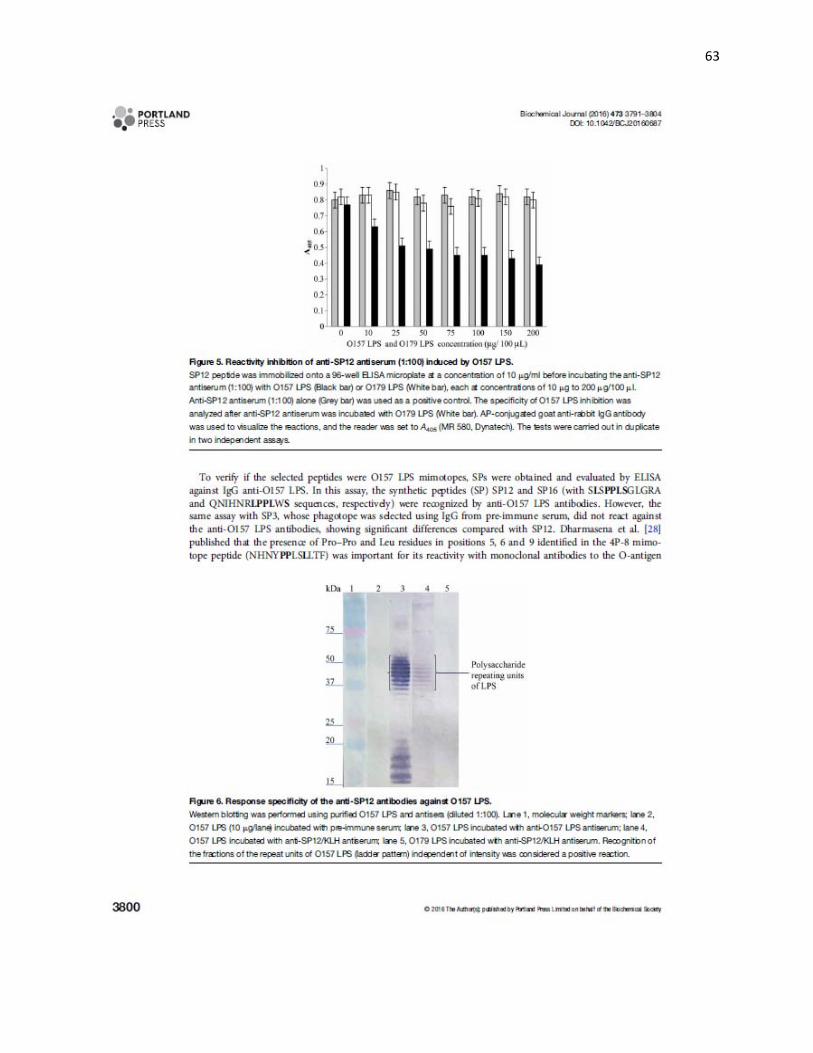
63
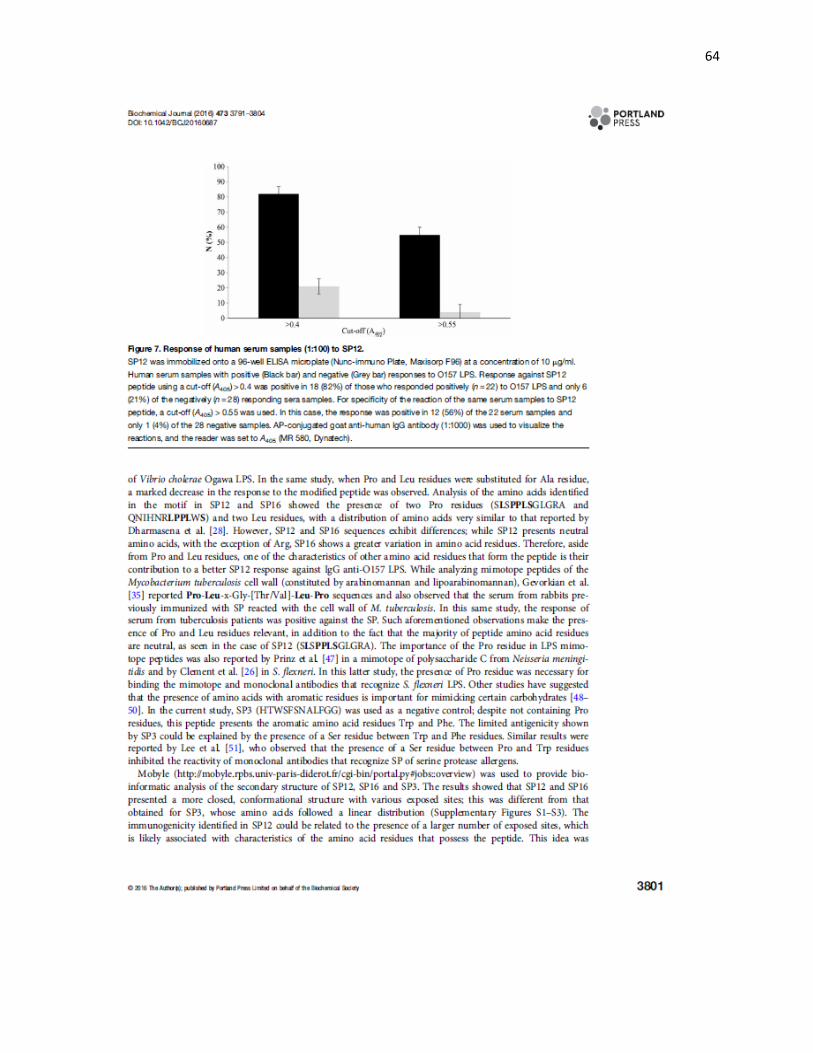
64

65

66

67












![Otra Universidad es posible [2004]](https://img.pdfslide.net/doc/110x75/6313c32bc32ab5e46f0c907a/otra-universidad-es-posible-2004.jpg)
















