Embed Size (px)
Citation preview

Psychopharmacology (2004) 171:306–321DOI 10.1007/s00213-003-1579-7
O R I G I N A L I N V E S T I G A T I O N
B. E. H. Sumner · L. A. Cruise · D. A. Slattery ·D. R. Hill · M. Shahid · B. Henry
Testing the validity of c-fos expression profiling to aidthe therapeutic classification of psychoactive drugs
Received: 25 March 2003 / Accepted: 21 June 2003 / Published online: 10 September 2003� Springer-Verlag 2003
Abstract Rationale: Different stimuli, including pharma-cological stimuli, induce different neuroanatomical pro-files of c-fos expression. Can these profiles be used inclassifying psychoactive drugs and predicting therapeuticutility? Objective: To test the validity of c-fos expressionprofiling to aid therapeutic classification. Methods: Anx-iolytics, antidepressants, antipsychotics and psychostim-ulants were compared. (i) A meta-analysis was performedand profiles compiled from literature reports of changes inc-fos expression in rat brain regions, measured by in situhybridisation histochemistry or immunohistochemistry,after acute injection of psychoactive drugs. (ii) Male ratbrains were profiled for changes in c-fos mRNA expres-sion induced by acute injection of psychoactive drugs.Results: (i) The meta-analysis showed that anxiolyticsactivate few (mostly stress-related) brain regions; antide-pressants activate more regions, including the centralamygdaloid nucleus; antipsychotics activate more regionsstill, including the nucleus accumbens and striatal areas;and psychostimulants activate the greatest number of all,including the most cortical regions (especially thepiriform cortex). Profiles also varied within drug classes.(ii) Our experimental profiles confirmed and extendedmeta-analysis profiles, showing more downregulation.(iii) Sites activated by mirtazapine (an antidepressant notpreviously profiled) matched those of the antidepressantimipramine. Conclusions: (i) Differences between drugclasses support their classification by means of c-fosprofiling. Differences within classes may reflect mecha-nistic variations. (ii) Greater downregulation in our
experiments might be because of inclusion of low,clinically relevant, drug doses and fuller coverage ofbrain regions. (iii) The agreement between mirtazapineand imipramine increases our confidence in the validity ofc-fos expression profiling to aid drug classification andpredict therapeutic utility.
Keywords c-fos expression profiling · Rat brain ·Anxiolytics · Antidepressants · Antipsychotics ·Psychostimulants
Introduction
Psychoactive drugs are usually classified first on theirclinical and therapeutic effects and, second, on theirmechanisms of action, if known (Cookson et al. 2002).Based on their chief therapeutic uses, most psychoactivedrugs can be classified as anxiolytics, antidepressants,mood stabilisers, antipsychotics, sedatives or hypnotics,antidementia drugs or psychostimulants. Subdivisions ofthese classes often describe pharmacological action, butmay also (or alternatively) describe chemical structure.For instance, selective serotonin re-uptake inhibitors(SSRIs) are a sub-class of antidepressants and benzodi-azepines are a sub-class of anxiolytics. Drugs of plantorigin may be grouped together, as in the case of theopiates. Other criteria for classification have also beeninvestigated, including the analysis of drug effects onelectroencephalogram (EEG) profiles recorded duringanimal sleep. This has now been developed as theautomated classification of sleep organisation (ACSO),which involves quantitative comparison, by computer, ofdrug-induced perturbations of EEG profiles (Ruigt et al.1989, 1993). Pharmacological magnetic resonance imag-ing may lead to a further method of classification (Leslieand James 2000; Preece et al. 2001). The method ofclassification evaluated in the following report relies oncomparisons between drug-induced patterns of expressionof an immediate early gene (IEG).
B. E. H. Sumner ()) · L. A. Cruise · D. A. Slattery · D. R. Hill ·M. Shahid · B. HenryDepartment of Pharmacology,Organon Laboratories Ltd.,Newhouse, Lanarkshire , ML1 5SH, Scotland, UKe-mail: [email protected]: +44-01698-736336
D. A. SlatteryPsychopharmacology Unit, School of Medical Sciences,University of Bristol,University Walk, Bristol, BS8 1TD, UK

In the mammalian nervous system, including the brain,induction of an IEG is one of the first signs of a genomicresponse to a stimulus (Sheng and Greenberg 1990). Thebest known of the IEGs are the proto-oncogene transcrip-tion factors, of which c-fos is a prime example (Beck-mann and Wilce 1997). IEG induction can bedemonstrated in cells or regions of tissue sections by insitu hybridisation (ISH) histochemistry or immunohisto-chemistry (IHC), which show mRNA or protein, respec-tively, in precise microscopic locations. With theseimaging techniques, c-fos expression has been extensivelystudied in the brain and elsewhere, and has becomewidely regarded as a mapping tool for sites of cellactivation (Herrera and Robertson 1996; Hoffman andLyo 2002; Kaczmarek and Robertson 2002).
Among the many stimuli that induce genomic re-sponses are pharmacological stimuli. Acute injection of adrug elicits responses which may include induction or,occasionally, downregulation of an IEG in the brain(Herrera and Robertson 1996). Do the patterns of IEGexpression betray the class of drug employed, and, if so,can this be used to aid the classification of novelcompounds and predict their therapeutic utility? Beck(1995) investigated antidepressants to find out whetherthey displayed a common c-fos induction profile anddiscovered that the central amygdaloid nucleus was theonly brain region activated by all the antidepressantstested. Robertson et al. (1994) investigated antipsychoticsto discover a means of recognising those that wereatypical. They developed a formula based on c-fosinduction profiles (see Discussion) and noted that all theantipsychotics tested activated the nucleus accumbens.However, there have been no comparative studiesbetween different classes of psychoactive drug to findout whether the different classes are easily recognisablevisually, at a glance, in a comparative map of their c-fosexpression profiles.
A drawback, for mapping purposes, is that there aresubpopulations of neurons and certain brain regions inwhich c-fos may not be either basally expressed orinducible, so any drug profile will be deficient in theseareas (Labiner et al. 1993; Chaudhuri 1997). For instance,c-fos expression is said to be not easily inducible in thesubstantia nigra (Dragunow and Faull 1989; Labiner et al.1993), although it has been activated in this region inmonkeys (Wirtshafter and Asin 1999) and after haloper-idol or caffeine in rats (Bennett and Semba 1998; Cochranet al. 2002). A further consideration is that c-fos is notequally inducible through all types of input or intracel-lular pathway (see Discussion). This might prove to be adrawback resulting in an inadequate map, or an advantageresulting in better discrimination between drug profiles. Athird consideration is the transience of c-fos expression.Activation of c-fos mRNA occurs within 30–60 min of anacute challenge with fluoxetine, or stimulation with light,and returns to baseline in most brain regions after 90 min(Torres et al. 1998; Zangenehpour and Chaudhuri 2002).The protein product, Fos, may peak between 2 h and 6 hafter an acute challenge, and be back to baseline well
before 24 h (Kaczmarek and Robertson 2002; Zangeneh-pour and Chaudhuri 2002). A comparative mappingproject, therefore, has to catch these windows of peakexpression.
Hypothesis
While being alert to the limitations of the c-fos mappingtechnique, our hypothesis was that different classes ofpsychoactive drug would elicit different c-fos expressionprofiles in the brain, by which the different classes couldbe recognised.
Aims
The first aim of this study was to carry out a meta-analysis of the c-fos literature to find out whether ourhypothesis was supported. The second aim was to makeour own experimental profiles of c-fos expression in adultrat brain using imaging technology for c-fos after acuteinjection of well-known psychoactive drugs. This willgrow into a reference database against which new drugscan be compared to establish and validate their class andcharacter, and predict their therapeutic utility. It wasdecided to start the latter process by profiling mirtazapine,an antidepressant not previously studied in this way, andcomparing its profile with that of other psychoactivedrugs to find out whether its character as an antidepres-sant would be confirmed.
Materials and methods
c-fos meta-analysis
The first step in the meta-analysis was to carry out full Medlinesearches (all years) for literature references in the combinedcategories of IEGs (or proto-oncogenes or c-fos) and drugs (namedindividually or in classes). We selected psychoactive drugs that arewell known, and for which there is a c-fos literature. Four majorclasses were chosen: anxiolytics, antidepressants, antipsychoticsand psychostimulants. The drugs in each class were as follows.
Anxiolytics
– Diazepam [benzodiazepine, increases gamma-amino-butyricacid (GABA)A receptor transmission, inhibits hypothalamic-pituitary-adrenal (HPA) axis; Salminen et al. 1996; Cookson etal. 2002]
– Chlordiazepoxide [CDP, benzodiazepine, increases GABAAtransmission, inhibits corticotrophin-releasing hormone (CRH)transcription; Imaki et al. 1995; Cookson et al. 2002]
– Buspirone [antagonist at dopamine (DA) D2 and D3 receptors,partial agonist at 5-hydroxytryptamine (5-HT)1A receptors;Palacios et al. 1996; Cookson et al. 2002].
Antidepressants
– Imipramine [tricyclic, inhibits noradrenaline (NA) and 5-HT re-uptake, increases available 5-HT and NA; Cookson et al. 2002]
307

– Desipramine (tricyclic, strongly inhibits NA re-uptake, slightlyinhibits 5-HT re-uptake, increases available NA and, to a lesserextent, 5-HT; Beck 1995; Cookson et al. 2002)
– Fluoxetine (SSRI, increases available 5-HT; also induces CRHtranscription; Torres et al. 1998; Cookson et al. 2002)
– Citalopram (SSRI, increases available 5-HT; Cookson et al.2002)
– Nortriptyline (tricyclic, inhibits NA re-uptake, increases avail-able NA; Beck 1995; Cookson et al. 2002)
– Tranylcypromine (monoamine oxidase inhibitor, MAOI, in-creases available NA, DA and 5-HT; Beck 1995; Cookson et al.2002)
– Buproprion (inhibits mostly DA, but also NA, re-uptake;increases available DA and NA; Beck 1995; Cookson et al.2002).
Antipsychotics (typical)
– Haloperidol (primarily a D2 receptor antagonist; Miyamoto etal. 2000; Cookson et al. 2002)
– Chlorpromazine (primarily a D2 receptor antagonist; Robertsonet al. 1994; Cookson et al. 2002)
– Raclopride (primarily a D2 receptor antagonist; Robertson et al.1994; Cookson et al. 2002).
Antipsychotics (atypical)
– Clozapine (broad spectrum antagonist acting at D1–5 receptors;also antagonistic at 5-HT2A and 2C, NA-alpha1 and 2, histamineH1, and acetylcholine M1 receptors; Miyamoto et al. 2000;Cookson et al. 2002)
– Risperidone (broad spectrum antagonist acting mainly at D2 and5-HT2A receptors, but also at 5-HT7, histamine H1 and NA-alpha1 and -alpha2 receptors; Miyamoto et al. 2000; Cookson etal. 2002)
– Olanzapine (broad spectrum antagonist acting at D1–4, 5-HT2A,2C, 3 and 6, NA alpha1, histamine H1 and acetylcholine M1–5receptors; Miyamoto et al. 2000; Cookson et al. 2002)
– Sulpiride (antagonist at D2 and especially D3 receptors; Preeceet al. 2001; Cookson et al. 2002).
Psychostimulants
– Amphetamine (inhibits DA and NA re-uptake, increases DAand NA release and transmission; Badiani et al. 1998; Cooksonet al. 2002)
– Methamphetamine (inhibits DA and NA re-uptake, increasesDA and NA release and transmission; Umino et al. 1995)
– Cocaine (inhibits DA re-uptake, increases available DA andenhances DA transmission; Johansson et al. 1994; Mach et al.1997)
– Phencyclidine [PCP, antagonist at N-methyl-d-aspartate(NMDA) receptors; also increases available 5-HT, and affectsACh receptors, DA, NA and GABA; Nakki et al. 1996; Martinet al. 1998]
– Ketamine (antagonist at NMDA receptors; Duncan et al. 1998)– Dizocilpine (MK-801, antagonist at NMDA receptors; Gao et
al. 1998)– Nicotine (agonist at nicotinic acetylcholine receptors, stimulates
DA neurons; Mathieu-Kia et al. 1998)– Caffeine (antagonist at adenosine receptors, also inhibits
phosphodiesterase, causing a rise in cyclic-AMP; Cookson etal. 2002; Singewald et al. 2003).
From these initial searches, every reference that used ISH tomeasure c-fos mRNA or IHC to measure the protein product, Fos,in drug-treated compared with vehicle-treated animals was scruti-nised further. Chronic studies and studies of all species other thanrats were rejected, and the final analysis was carried out on studies
of c-fos expression in sections of adult rat brain after a single acuteinjection of drug or vehicle. Eighty references met our criteria (seeAppendix for full list). From the data given in each quantitativestudy, the percentage change in c-fos mRNA content (ISH) or Fos-positive cell numbers (IHC) was calculated for each brain regionthought to be involved in mechanisms of stress, anxiety, depressionor psychosis. The list of brain regions was compiled from thoseimplicated in the relevant literature. Many authors had notsubdivided the nucleus accumbens and the striatum, but somehad, and had found differences between the different subdivisionsafter different antipsychotic treatments. In our list, these regionswere listed both whole and subdivided, so that data from paperswith different approaches could be adequately entered and assessed.A table of percentage changes for each drug (at each dose) wascompleted for each reference, and then entered into a combinedtable of results for each drug from all the references. An overallmean percentage change was calculated for each drug (at its mosteffective dose range) in each brain region and assembled into anoverall table so that expression profiles could be compared.However, the percentage changes obtained from different refer-ences were so diverse that it was decided this calculation of overallmeans was not useful. It was considered more useful to tabulateoverall increase, decrease or no change in c-fos expression in eachbrain region for each drug. This latter approach enabled us toinclude semi-quantitative studies, in which responses were gradedsubjectively, as well as strictly quantitative studies with detailedmeasurements. It also enabled us to make visual comparisonsbetween drug profiles at a glance.
Experimental c-fos profiles
Animals and drug treatments
In pursuit of our second aim, adult male Sprague-Dawley rats(Harlan Olac, UK; body weight 200–250 g) were calmed by dailyhandling and weighing for 5 min each day for 5 days before drugtreatment, to ensure a low basal expression of c-fos mRNA (Hess etal. 1995). All animals were injected i.p. with either a psychoactivedrug or vehicle (n=6 per group). Drugs and doses includedfluoxetine (5 mg/kg and 10 mg/kg), imipramine (15 mg/kg),haloperidol (0.05 mg/kg and 0.5 mg/kg), clozapine (2 mg/kg and30 mg/kg) and amphetamine (5 mg/kg). The higher doses for thesewell-known drugs were based on those used for c-fos expressionstudies in the literature. The lower doses were related to those usedclinically in humans (VanderSpek et al. 2002). Mirtazapine has notbeen profiled before. This drug, a NA and specific serotonergicantidepressant (NaSSA), was used at 2 mg/kg, based on previousexperiments in male rats (De Boer et al. 1994). The vehicle was 5%mulgofen EL-719 (GAF, Ltd., Manchester, UK) in sterile 0.9% w/vsaline. All the animal experiments were carried out according to theAnimals (Scientific Procedures) Act, 1986, Home Office, UK.
Tissue collection and processing
The animals were euthanased by excess CO2 45 min post-injection,and their brains were removed, frozen in iso-pentane cooled to�35�C by dry ice, and stored in sealed polythene bags in a freezer at�80�C. Serial coronal sections (20 mm) from bregma 4.7 mm to�10.3 mm (Paxinos and Watson 1997) were cut in a cryostat at�18�C, thaw-mounted three-per-slide on positively charged, nucle-ase-free slides (Superfrost Plus, VWR) and stored again briefly at�80�C. The slide-mounted sections were fixed in 4% paraformal-dehyde, acetylated, de-lipidised and stored at 4�C in 95% ethanoluntil required for in situ hybridisation (Wisden and Morris 1994).
Probes for c-fos mRNA
Two 48-base antisense oligonucleotide probes were designed totarget different, non-overlapping sequences of rat c-fos mRNA
308

(NCBI GenBank accession number: X06769). Probe 1 wascomplementary to bases 1001–1048 (probe sequence: 50-GGCTCC CAG TCT GCT GCA TAG AAG GAA CCA GAC AGGTCC ACA TCT GGC–30). Probe 2 was complementary to bases141–188 (probe sequence: 50-ACT GCA GCG GGA GGA TGACGC CTC GTA GTC CGC GTT GAA ACC CGA GAA–30). Usingthe NCBI Basic Local Alignment Search Tool (BLAST; Altschul etal. 1990), both the sense sequences were checked for homologywith other sequences and found to be specific for the c-fos gene.
ISH histochemistry
For ISH, the probes were 30-end-labelled with 33P-dATP (specificactivity: 92.5TBq/mmol; Amersham Biosciences, UK) and used ata concentration of 0.5 pmol/ml ’maximalist’ hybridisation buffer(Wisden and Morris 1994). Once specificity had been confirmed foreach labelled probe separately (see below), the labelled probes wereused together in the same hybridisation mixture thereafter, in orderto increase the hybridisation signal and reduce exposure time.Sections, which had been fixed, acetylated, de-lipidised (FADLd)and stored in 95% ethanol at 4�C, were air-dried at roomtemperature and then hybridised for 18 h at 42�C. After hybrid-isation, all the sections were taken through a series of washes ofincreasing stringency to remove unbound and non-specificallybound probe. These consisted of a brief rinse in 2� saline sodiumcitrate (SSC), two washes for 30 min each in 1� SSC, first at roomtemperature and then at 55�C, and, finally, a 10-min wash in 0.1�SSC at 55�C. Following the last wash, the sections were dehydratedin 70% and 95% ethanols for 2 min each and air-dried.
Specificity controls
Specificity of the probes for RNA was checked by hybridisation ofcontrol sections with 33P-labelled probes after RNase pre-treatment,which abolished all signal. To check the specificity of the probesfor the desired mRNAs, control sections were incubated inhybridisation buffer containing both 33P-labelled and unlabelledversions of each probe, the latter at 100-fold excess to compete withthe labelled probe for specific mRNA. Competition was successfuland no signal was found in the resulting autoradiographs. Probespecificity was also checked by comparing the distribution of signalin the autoradiographs with that found in the literature (seeResults).
Autoradiography and image analysis
The slides of dry sections were apposed to Kodak Biomax MR-1film together with a radioactive standard (14C-microscales, Amers-ham Biosciences, UK), and exposed for 2–3 weeks to obtainautoradiographic images. The films were developed for 3 min inKodak GBX developer/replenisher (20% aqueous), rinsed briefly inwater, fixed for 5 min in Kodak GBX fixer/replenisher (20%aqueous), and washed for 5 min in tap water. The slides removedfrom the film were stained with cresyl fast violet, haematoxylin andeosin, or pyronin to check histology. For each identified brainregion, the optical density of the autoradiographic image wasmeasured using an MCID image-analyser (InterFocus, Ltd., Hav-erhill, Suffolk, UK). The optical densities were converted toradioactivity values by means of the standard scale, in order tonormalise densities from film to film. Quantitative results fromdrug-treated and vehicle-treated rats were compared statistically byANOVA and Dunnett’s post-hoc test. This enabled us to identifybrain regions showing significant changes in c-fos mRNA expres-sion. Significant increases, decreases, and no changes in expressionwere tabulated in the same way as for the meta-analysis so thatinstant visual comparisons could be made between drug profiles.
Results
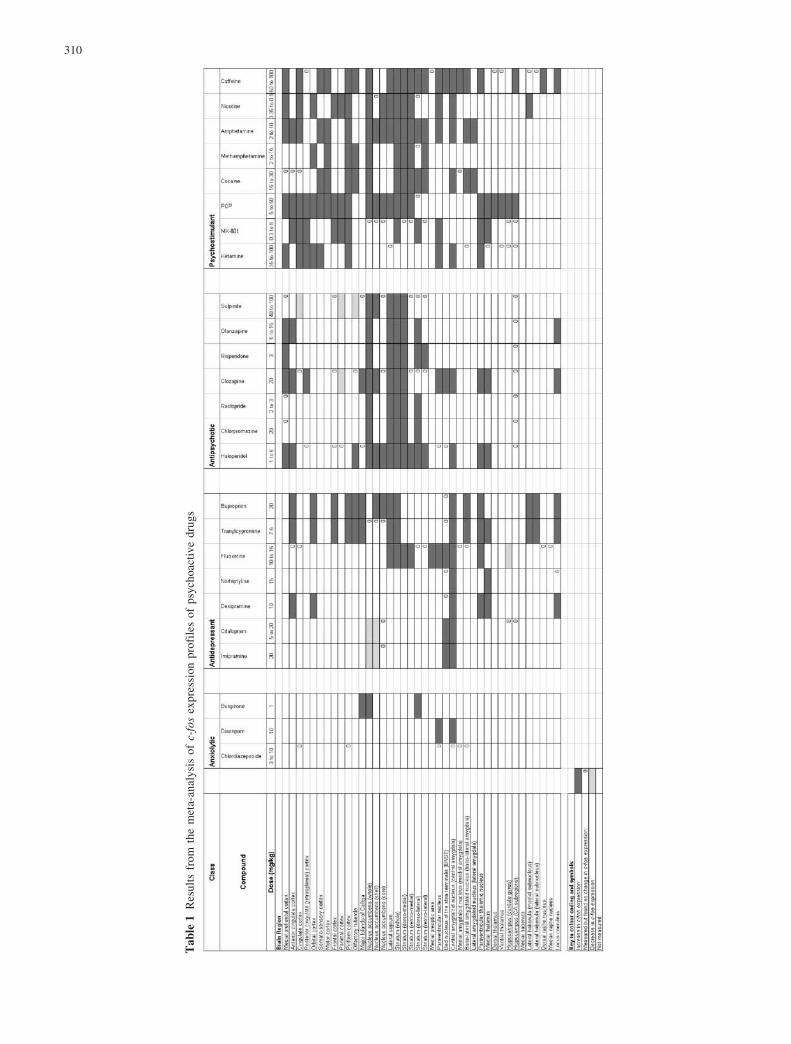
The results from the meta-analysis are shown in Table 1.Compared with other classes of drug, anxiolytics havehad the least investigation in the kind of study requiredfor inclusion in our survey. Analysis of three drugs in theliterature showed that they activated c-fos expression invery few brain regions, and displayed no cortical activa-tion at all. Buspirone activated the nucleus accumbens,lateral septum and dorso-lateral striatum; diazepamactivated the paraventricular nucleus and central amyg-daloid nucleus; and chlordiazepoxide failed to activateany of the regions measured in our survey.
Antidepressants have had more investigation and, onthe whole, they activated more brain regions thananxiolytics. These were mostly limbic regions, but threeof the seven drugs in our survey gave some corticalactivation also (desipramine, tranylcypromine and bupro-prion). Tranylcypromine and buproprion activated thelateral habenula, and these two drugs and desipramineactivated the locus coeruleus. All the antidepressantssurveyed induced c-fos expression in the central amyg-daloid nucleus. There were decreases of c-fos expressionin the nucleus accumbens (by imipramine and citalopram)and in the dentate gyrus of the hippocampus (byfluoxetine).
Antipsychotics have generated many more papers, butmost of these have been devoted to haloperidol andclozapine, as examples of typical and atypical antipsy-chotics, respectively. Our survey included three typicaland four atypical antipsychotics (see Materials andMethods), but the c-fos expression profiles compiledfrom all the papers surveyed showed no clear differencebetween these two categories unless the nucleus accum-bens and the striatum had been subdivided (Table 1). Inpapers that recognised subdivisions of the nucleusaccumbens, it was reported that clozapine and sulpirideactivated c-fos in the shell but not the core, whereashaloperidol activated both shell and core. Studies inwhich the striatum was subdivided found that haloperidolactivated the lateral, especially the dorso-lateral, striatum,whereas clozapine and sulpiride gave little or no activa-tion in this region. As a class, antipsychotics induced c-fosin more brain regions than either anxiolytics or antide-pressants. All the antipsychotics surveyed induced ex-pression in the nucleus accumbens, and striatal c-fosexpression was also activated to various degrees. Four ofthe seven antipsychotic drugs analysed induced activity inthe medial prefrontal cortex, three of these four alsoactivated the anterior cingulate cortex, but only one ofthem, clozapine, activated the posterior cingulate (retro-splenial) cortex. In the parietal cortex, c-fos expressionwas decreased by clozapine and sulpiride. Sulpiride alsodecreased expression in the cingulate cortex and olfactorytubercle.
Psychostimulants induced c-fos expression in morecortical regions than antipsychotics. All activated thepiriform cortex. There was a delayed activation of theposterior cingulate (retrosplenial) cortex by the NMDA
309
Tab
le1
Res
ults
from
the
met
a-an
alys
isof
c-fo
sex
pres
sion
prof
iles
ofps
ycho
acti
vedr
ugs
310

antagonists at high drug doses. The nucleus accumbensand striatum often showed activation also. Of all the fourclasses of psychoactive drug examined, psychostimulantsactivated the most brain regions. No decreases of c-fosexpression were found in the literature on psychostimu-lants.
Results from our own experimental c-fos expressionprofiles are shown in Table 2. We have not, as yet,investigated any anxiolytics. In agreement with the resultsfrom the meta-analysis, the antidepressants activated thefewest brain regions (in the absence of any anxiolytics),the antipsychotics were the next most effective (at thehigher doses), and the psychostimulant amphetamineactivated the most brain regions, including the mostcortical regions. Among antidepressants, the profileobtained from mirtazapine was similar in sites ofactivation to the profile from imipramine. The antide-pressants in our experiments all activated the centralamygdaloid nucleus (where measured) and both antipsy-chotics at the higher dose activated the nucleus accum-bens (although with clozapine only the shell wasactivated). Compared with the results from the meta-analysis, we found more downregulation of c-fos expres-
sion by antidepressants, especially in thalamic regions,but also in hippocampus, lateral septum and severalcortical regions. Clozapine also decreased c-fos expres-sion in more regions than reported in the majority ofpapers surveyed for the meta-analysis. At the lower,clinically relevant, dose used for clozapine (2 mg/kg), allthe responsive sites were sites of downregulation; therewas no significant activation. For cingulate cortex, lateralseptum and lateral striatum, this meant a switch fromactivation to downregulation between doses of 30 mg/kgand 2 mg/kg. Haloperidol and amphetamine agreed withthe literature in showing no sites of downregulation, andthe amount of c-fos activation was approximately similarto that in the literature for these two drugs.
Discussion
c-fos meta-analysis
For meaningful results, it is essential to compare like withlike. This is not easy in an analysis of the literature.Different studies have different aims and focus, and use
Table 2 Results from our ex-perimental c-fos expressionprofiles
311

different neuroanatomical terminology, groupings andsubdivisions, different drug doses and different tissue-processing and data-processing routines. To reduce thevariation between studies and make them more compa-rable, we only selected those papers that met certaincriteria (see Materials and methods). By this means, wewere able to carry out a comparative survey of psycho-active drugs according to their c-fos expression profilesand enter the consensus profiles into a table that could bevisually assessed at a glance. Nevertheless, dissimilaritiesbetween studies still remained. These were particularlyevident in the field of antipsychotics. Some authorsrecognised subdivisions of the nucleus accumbens and thestriatum, and analysed them separately, but others anal-ysed them as a whole. To overcome this problem, weincluded both whole and subdivided categories in our listof brain regions, and entered the data from the papers intothe appropriate category. In a survey of animal and humanstudies of antipsychotics, it has been shown that the drugdoses employed in animal studies are usually higher thanthose used clinically, and that the doses chosen fordifferent drugs may give different DA D2 receptoroccupancies, which make them difficult to compare(VanderSpek et al. 2002). For psychostimulants, authorswho used a range of drug doses in their animalexperiments found that low doses, which producedlocomotor effects, were ineffective at inducing c-fosexpression, and only the higher doses activated c-fos (forexample: Bennett and Semba 1998). In our survey, thetabulated results were drawn up from the most commonlyused and effective dose range reported for c-fos studies inrats for each drug. These dose ranges were high whencompared with dose ranges used in humans. In the usablereferences, IHC studies outnumbered ISH studies by 7 to3.
The consensus results suggest that there is variabilityin the c-fos expression profile between different drugs inthe same class. Nevertheless, there is an overall profile foreach class, which differs between classes. These differ-ences are not only in the nature of the brain regionsactivated, but also in their number. For instance, all theantidepressants in our literature survey activated thecentral amygdaloid nucleus and all the antipsychoticsactivated the nucleus accumbens, as had been found alsoby Beck (1995) for antidepressants and Robertson et al.(1994) for antipsychotics. Our survey, which is the first tomake comparisons between four major classes of psy-choactive drug, found that the number of brain regionsactivated tended to increase in the series from anxiolyticsto antidepressants to antipsychotics to psychostimulants.In addition, although we found strong striatal induction byboth antipsychotics and psychostimulants, these classesdiffered in that psychostimulants excited c-fos in morecortical regions than antipsychotics. Of course, to someextent these results reflect the focus of the papers in theliterature, which is usually directed towards the mostlikely brain regions to give positive results, based on theneuroanatomical distribution of the drug targets and thepathophysiology of related psychiatric illnesses. Unless a
brain region has been measured and definitely foundunresponsive, there is always the possibility that it mightbe responsive, but overlooked. Furthermore, negativeresults are not reported as often as positive, and mostpeople are seeking sites of activation. However, theliterature as it stands at present, incomplete though itmight be, supports our hypothesis that different classes ofdrug display different c-fos expression profiles whichenable the different classes to be recognised, after acutedrug administration.
What is the basis for these class differences? Evidencefrom experiments of drug action on stressed or anxiousanimals suggests that anxiolytics target stress mechanismsof the HPA axis, which are centred in the paraventricularnucleus (PVN) of the hypothalamus (Imaki et al. 1995).Even in unstressed animals, diazepam has been found toexcite c-fos expression in the PVN and amygdala(Table 1). CDP, however, failed to have any effect inunstressed animals. In stressed animals, CDP reducedboth the stress-induced c-fos expression and the stress-induced increase in transcription of CRH in the PVN,which would, in turn, reduce the stress-induced increasesof other hormones of the HPA axis (Imaki et al. 1995).Buspirone gave no evidence of HPA activation, butactivated restricted areas relating chiefly to DA. Buspi-rone was originally developed as a possible antipsychotic,but was found to be effective as an anxiolytic and is nowclassed as such (Baldessarini 1990; Cookson et al. 2002).It is also used in antidepressant treatment regimes (Menzaet al. 2003). The antidepressants surveyed all activatedthe central amygdaloid nucleus, which is a recognisedcentre for emotional processing, especially for negativeemotions such as fear and anxiety in rodents (Alheid andHeimer 1996; Davis and Shi 1999) and humans. Sustainedprocessing of negative emotions and even enlargement ofthe amygdala feature in human depressive illness (Siegleet al. 2002; Frodl et al. 2002). The chief neuroanatomicaltarget for antipsychotics, the nucleus accumbens, isassociated with dopaminergic psychomotor processingin rodents and humans, and with the positive symptoms ofschizophrenia in humans (Deutsch and Duman 1996).Schizophrenia may result from excess of DA andcorresponding DA hyperactivity (Palmer et al. 1997).Typical antipsychotics, which also activate the dorso-lateral (DL) striatum, are liable to cause motor sideeffects in humans, and the DL striatum is also a site of DAdepletion, with associated motor dysfunction, in Parkin-son’s disease (Deutsch and Duman 1996). The nucleusaccumbens and the striatum are particularly rich in DAreceptors. The D2 receptor is a common molecular targetfor antipsychotics, as has been confirmed in humanstriatum by positron emission tomography (PET) afterantipsychotic treatment (Seeman 2002). PET and mag-netic resonance imaging have also shown effects onregional blood flow in the striatum and the cingulatecortex after antipsychotic treatment of schizophrenicpatients (Miller et al. 2001). Activation of the medialprefrontal cortex by atypical antipsychotics may berelated to their successful treatment of the negative
312

aspects of schizophrenia in humans (Deutsch and Duman1996; Robertson and Fibiger 1996). Ohashi et al. (2000)have suggested that the action on the medial prefrontalcortex and on the locus coeruleus by atypical antipsy-chotics may involve the noradrenergic system and beta-adrenoceptor mediation. The cortex, often accompaniedby the striatum and nucleus accumbens, is a major targetfor c-fos activation by psychostimulants, and this agreeswith their capacity to heighten mental and psychomotoractivity and create euphoria in humans (Cookson et al.2002). The action of many psychostimulants results in anincrease in available DA and an enhancement of DAtransmission in humans, monkeys and rodents (Volkow etal. 1996; Mach et al. 1997; Ruskin and Marshall 1994),but chronic doses may cause a state resembling psychosis(Hamamura et al. 1997). The delayed activation of theposterior cingulate (retrosplenial) cortex by high doses ofNMDA antagonists is thought to be a toxic effect leadingto extensive cell death in this region (Fujimura et al.2000a).
Within classes the variability of c-fos profiles betweenindividual drugs probably reflects their different mecha-nisms and outcomes of action, and the different neuro-anatomical distributions of their molecular targets. Theremay also be differences of effectiveness by differentpathways. The literature suggests that, although neuronalc-fos is widely inducible by neurotransmitters, hormones,neuropeptides, growth factors and drugs, it is not equallyinducible through all types of input. For instance, amongthe antidepressants included in our meta-analysis, thosethat include increase of available DA in their action weremore effective at eliciting a c-fos response than those thatonly increase 5-HT or NA. Thus, buproprion andtranylcypromine (Beck 1995) activated more brain re-gions than the other antidepressants (Table 1). Not onlydoes the type of neurotransmitter affect the c-fosresponse, but also the receptor subtype that mediates theaction. Drugs that increase DA release and favourtransmission through D1 receptors, such as amphetamineand cocaine, are effective inducers of c-fos (Table 1), butagonists at D2 receptors are ineffective (Herdegen andLeah 1998). Antagonists at D2 receptors include the well-known antipsychotics of our study, which increased c-fosexpression in regions rich in this DA receptor subtype,especially the nucleus accumbens and striatum (Table 1).Several authors discovered differences between typicaland atypical antipsychotics in the profiles obtained fromdifferent subdivisions of either the nucleus accumbens orstriatum (Fink-Jensen and Kristensen 1994; Robertson etal. 1994; Fink-Jensen et al. 1995; Sebens et al. 1995;Hiroi and Graybiel 1996; Hurley et al. 1996; Semba et al.1996, 1999; Vahid-Ansari and Robertson 1996; Guitartand Farre 1998; Ishibashi et al. 1999; Fujimura et al.2000b; Werme et al. 2000). For instance, haloperidolinduced c-fos expression in both core and shell of thenucleus accumbens, whereas clozapine and sulpirideinduced it in the shell but not the core. Haloperidol wasa strong inducer of c-fos expression in the dorso-lateralstriatum, whereas clozapine and sulpiride were not.
Robertson et al. (1994) claimed that typical and atypicalantipsychotics could be distinguished on the basis of anindex obtained by subtracting the extent of induction inthe dorso-lateral striatum from that in the nucleusaccumbens. If the index was negative, the drug was atypical antipsychotic, and, if positive, an atypical anti-psychotic. These distinctions may reflect differencesbetween the action of haloperidol (which is predominant-ly a D2 antagonist) and clozapine (which is an antagonistat all DA receptors, D1–D5, and at 5-HT2A and 2Creceptors, and with action also at histamine H1, acetyl-choline M1 and NA-alpha1 and -alpha2 receptors; Cook-son et al. 2002; Miyamoto et al. 2000). Guo et al. (1995)have suggested that the action of clozapine on c-fosexpression in the nucleus accumbens may depend on D3receptors, but Hurley et al. (1996) disagree. Sulpiride ispredominantly a D2 and D3 antagonist, but again Hurleyet al. (1996) found no evidence that D3 receptors wereinvolved in the drug action on c-fos. Amino acids mayexcite electrical activity in neurons through AMPA,kainate and NMDA receptors, but IEG induction ismostly through NMDA inputs (Herdegen and Leah 1998).NMDA antagonists can also induce c-fos, as was foundfor PCP, ketamine, and MK-801 in our meta-analysis(Table 1). Not only are inputs important, but alsoHoffman and Lyo (2002) have pointed out that there aredominant intra-cellular pathways of c-fos regulation,which are more likely to induce a response than otherpathways. The most effective are those that involveincreases of calcium ions or cAMP and action through acalcium response/cAMP response element (CaRE/CRE)on the c-fos gene.
Experimental c-fos profiles
Compared with the c-fos expression profiles compiledfrom the literature, our own experimental profiles showedapproximately similar extents of upregulation but, inaddition, more extensive downregulation. Some of thisdownregulation was found at very low drug doses,especially in the case of clozapine, which showed noactivation and only downregulation at a low, clinicallyrelevant, dose (2 mg/kg). But the higher doses, whichresulted in downregulation, were similar to those used forc-fos studies in the literature. A possible explanationmight be that our experimental profiles were basedentirely on ISH results and the literature contains manymore IHC studies than ISH studies (70% in our meta-analysis). Methods for ISH using radioactively labelledprobes are more sensitive than IHC methods relying oncoloured reaction products. Downregulation might bemore easily spotted in an ISH preparation. Moreover, ISHmight show changes in mRNA which are not translatedinto changes in protein. However, neither of theseexplanations wholly fits the case. Over half the down-regulation reported in the literature is from IHC studies. Amore likely explanation is that downregulation is moreoften overlooked, whether ISH or IHC is used. Basal
313

expression of c-fos is often low, and downregulation maybe slight and easily dismissible in the search for sites ofactivation. In addition, most papers in the literature focuson a few brain regions rather than give a whole profile.The profiles in the meta-analysis were compiled frompapers of different focus. Few of these included thethalamus and hippocampus in their analyses, and much ofthe downregulation in our experimental profiles ofantidepressants is in these regions. Fuller coverage ofbrain regions in our experiments might also explain someof the downregulation found more anteriorly in our ISHpreparations.
Mirtazapine is an example of a newer type ofantidepressant, a noradrenergic and specific serotonergicantidepressant (NaSSA). This drug blocks NA-alpha2autoreceptors resulting in increased release of NA fromNA terminals, and also blocks NA-alpha2 heteroreceptorson 5-HT neurons, resulting in increased release of 5-HT(Cookson et al. 2002). The increased NA increases thefiring of 5-HT neurons by action through NA-alpha1receptors, thereby further enhancing both NA and 5-HTfunction. Mirtazapine also antagonises 5-HT2 and 5-HT3receptors, but not 5-HT1A. The net result is increased NAand 5-HT1 transmission. The older drug, imipramine, is atricyclic antidepressant, which increases available NAand 5-HT by blocking re-uptake (Cookson et al. 2002).Thus, both drugs increase available NA and 5-HTalthough their mechanisms of action are slightly different.It is interesting that the similarity of outcome results insimilar c-fos activation profiles for mirtazapine andimipramine (Table 2). Not only does this confirm thecharacter of mirtazapine as an antidepressant, but also itincreases our confidence in the validity of c-fos expres-sion profiling as a tool to aid the therapeutic classificationof new psychoactive agents.
Further developments
The expression-profiling basis of drug classification couldbe developed to include mapping of other IEGs. Forinstance, c-jun is an inducible transcription factor geneoften associated functionally with c-fos (Herdegen andLeah 1998). However, although its protein product, Jun,dimerises with Fos protein and induces transcription bybinding to AP-1 sites on DNA, c-jun can also actindependently of c-fos, and shows increased expression attimes of apoptosis or regeneration after injury, whensynaptic rearrangements are occurring (Dezhi et al. 1999;Herdegen and Leah 1998). Synaptic changes are thoughtto occur during the development of, and recovery from,depressive illness. Furthermore, there is prolonged in-crease of c-jun expression in the basolateral amygdalaafter olfactory bulbectomy, which is a recognised animalmodel of depression (Wrynne et al. 2000). Antipsychoticsare already known to increase c-jun (Herdegen and Leah1998). Krox-24 (zif268, NGFI-A, egr-1), although classedas an inducible transcription factor gene, has a high basalexpression in unstimulated animals (Herdegen and Leah
1998). This IEG has been found necessary in mechanismsof long-term potentiation and synaptic plasticity inmemory formation (Jones et al. 2001) and is inducibleby antidepressants (Morinobu et al. 1997). Krox-24 is alsosaid to have a lower induction threshold than c-fos inresponse to antipsychotics. Arc is an example of aneffector IEG, reported to be involved with synapticplasticity and regulated by 5-HT (Lyford et al. 1995; Peiet al. 2000). This IEG might be affected by depressiveillness and recovery, and might be sensitive to antide-pressants. Mapping of these IEGs in addition to c-foswould extend the basis of comparison with new com-pounds.
It could be argued that a better way of determining theclass of a supposedly therapeutic agent would be tomeasure its effect on IEG profiles induced by a conditionsuch as anxiety, depression or psychosis. This functionalapproach has been used to investigate the mechanism ofaction of anxiolytics, as cited above for CDP. It has alsobeen used to subclassify antidepressants. For instance, theforced swim test, which is a recognised model ofdepression and despair in rodents, induces an increaseof c-fos expression which can be reversed by somechronic antidepressant treatments (Duncan et al. 1996).The antidepressants that were effective were those thatmainly blocked NA uptake, namely imipramine, desipra-mine and nisoxetine, while fluoxetine, tranycylpromineand mianserin were ineffective. However, this approachto drug classification is limited by the difficulties ofmimicking psychiatric conditions in animal models.
The techniques of genomics and proteomics, in whichthe effects of drugs on multiple genes and proteins can bemapped simultaneously, will doubtless lead to newmethods of classification. For precise neuroanatomicallocation using these methods, precise microdissection ofbrain regions is necessary. Neuroanatomical resolutioncan be increased even to the cellular level by laser capturemicrodissection (Bonaventure et al. 2002; Wittliff andErlander 2002). The challenge will be to discern a patternamong the detail! Imaging methods, such as 2-deoxy-glucose for metabolic mapping (Sharp et al. 1993;Komisaruk et al. 2000) and BOLD fMRI for mappingblood flow (Leslie and James 2000; Preece et al. 2001),could be developed into methods for drug classification.In addition, behavioural and neurochemical paradigmscould be further explored to extend the criteria fortherapeutic classification.
Novel compounds
Although there are limitations to the c-fos mappingapproach, our analysis has shown that usable maps foraiding the classification of new psychoactive drugs can becompiled. These maps would also aid therapeutic predic-tion. If a novel compound were developed with a newmechanism of action, therapeutic classification andprediction would be assisted if some part of the actionaltered c-fos expression. The mechanism might involve
314

direct action on neurons expressing c-fos, or indirectaction, through other neurons connecting via circuitry toc-fos-responsive neurons. As shown by our meta-analysisof the literature, particular brain regions are characteris-tically involved in mechanisms of anxiety, depression,psychosis and psychostimulation, and the novel com-pound might ultimately target some of these regions. If,however, the novel compound were to act through amechanism with no effect on c-fos expression, anothermethod of therapeutic classification and prediction wouldhave to be used. It remains best to approach thetherapeutic classification of novel compounds by morethan one method, so that a balanced judgement can beachieved despite the limitations of the different methods.
Summary and conclusions
Our meta-analysis of the c-fos expression profiles ofpsychoactive drugs in four classes found that the profilesvaried between individual drugs, most probably accordingto their different mechanisms and targets. Nevertheless,there was an overall profile for each class, which differedbetween classes. Therefore, the meta-analysis supportsour hypothesis that different classes of drug displaydifferent c-fos expression profiles that characterise theirclasses.
Comparison of the c-fos profile induced by mirtaza-pine with those induced by well-known psychoactivedrugs confirmed the character of mirtazapine as anantidepressant. It follows that it is valid to use c-fosexpression profiles of well-known psychoactive drugs asa reference to aid the classification of novel psychoactiveagents, and to predict their therapeutic utility.
Appendix
References selected for the c-fos meta-analysis
– Badiani A, Oates MM, Day HE, Watson SJ, Akil H,Robinson TE (1998) Amphetamine-induced behavior,dopamine release, and c-fos mRNA expression: mod-ulation by environmental novelty. J Neurosci18:10579–10593
– Beck CHM (1995) Acute treatment with antidepres-sant drugs selectively increases the expression of c-fosin the rat brain. J Psychiatry Neurosci 20:25–32
– Bennett HJ, Semba K (1998) Immunohistochemicallocalization of caffeine-induced c-Fos protein expres-sion in the rat brain. J Comp Neurol 401:89–108
– Bujas-Bobanovic M, Robertson HA, Dursun SM(2000) Effects of nitric oxide synthase inhibitorN(G)-nitro-l-arginine methyl ester on phencyclidine-induced effects in rats. Eur J Pharmacol 409:57–65
– Carta AR, Gerfen CR (1999) Lack of a role for the D3receptor in clozapine induction of c-fos demonstratedin D3 dopamine receptor-deficient mice. Neuroscience90:1021–1029
– Castner SA, Becker JB (1996) Sex differences in theeffect of amphetamine on immediate early geneexpression in the rat dorsal striatum. Brain Res712:245–257
– Cenci MA, Kalen P, Mandel RJ, Wictorin K, Bjork-lund A (1992) Dopaminergic transplants normalizeamphetamine- and apomorphine-induced Fos expres-sion in the 6-hydroxydopamine-lesioned striatum.Neuroscience 46:943–957
– Cochran SM, McKerchar CE, Morris BJ, Pratt JA(2002) Induction of differential patterns of localcerebral glucose metabolism and immediate-earlygenes by acute clozapine and haloperidol. Neurophar-macology 43:394–407
– Cohen BM, Wan W (1996) The thalamus as a site ofaction of antipsychotic drugs. Am J Psychiatry153:104–106
– Dawe GS, Huff KD, Vandergriff JL, Sharp TL,O’Neill MJ, Rasmussen K (2001) Olanzapine activatesthe rat locus coeruleus: in vivo electrophysiology andc-fos immunoreactivity. Biol Psychiatry 50:510–520
– Day HE, Badiani A, Uslaner JM, Oates MM, VittozNM, Robinson TE, Watson SJ Jr, Akil H (2001)Environmental novelty differentially affects c-fosmRNA expression induced by amphetamine or cocainein subregions of the bed nucleus of the stria terminalisand amygdala. J Neurosci 21:732–740
– Deutsch AY, Ongur D, Duman RS (1995) Antipsy-chotic drugs induce Fos protein in the thalamicparaventricular nucleus: a novel locus of antipsychoticdrug action. Neuroscience 66:337–346
– Dragunow M, Faull RL (1990) MK801 induces c-fosprotein in thalamic and neocortical neurons of ratbrain. Neurosci Lett 113:144–150
– Duncan GE, Moym SS, Knapp DJ, Mueller RA,Breese GR (1998) Metabolic mapping of the rat brainafter subanesthetic doses of ketamine: potential rele-vance to schizophrenia. Brain Res 787:181–190
– Engber TM, Koury EJ, Dennis SA, Miller MS,Contreras PC, Bhat RV (1998) Differential patternsof regional c-Fos induction in the rat brain byamphetamine and the novel wakefulness-promotingagent modafinil. Neurosci Lett 241:95–98
– Fink-Jensen A, Kristensen P (1994) Effects of typicaland atypical neuroleptics on Fos protein expression inthe rat forebrain. Neurosci Lett 182:115–118
– Fink-Jensen A, Ludvigsen TS, Korsgaard N (1995)The effect of clozapine on Fos protein immunoreac-tivity in the rat forebrain is not mimicked by theaddition of alpha 1-adrenergic or 5HT2 receptorblockade to haloperidol. Neurosci Lett 194:77–80
– Fujimura M, Hashimoto K, Yamagami K (2000b) Theeffect of the antipsychotic drug mosapramine on theexpression of Fos protein in the rat brain: comparisonwith haloperidol, clozapine and risperidone. Life Sci67:2865–2872
– Gao XM, Hashimoto T, Tamminga CA (1998) Phen-cyclidine (PCP) and dizocilpine (MK801) exert time-
315

dependent effects on the expression of immediate earlygenes in rat brain. Synapse 29:14–28
– Gass P, Herdegen T, Bravo R, Kiessling M (1993)Induction and suppression of immediate early genes inspecific rat brain regions by the non-competitive N-methyl-D-aspartate receptor antagonist MK-801. Neu-roscience 53:749–758
– Guitart X, Farre AJ (1998) The effect of E-5842, asigma receptor ligand and potential atypical antipsy-chotic, on Fos expression in rat forebrain. Eur JPharmacol 363:127–130
– Guo N, Klitenick MA, Tham CS, Fibiger HC (1995)Receptor mechanisms mediating clozapine-induced c-fos expression in the forebrain. Neuroscience 65:747–756
– Habara T, Hamamura T, Miki M, Ohashi K, Kuroda S(2001) M100,907, a selective 5-HT(2A) receptorantagonist, attenuates phencyclidine-induced Fos ex-pression in discrete regions of rat brain. Eur JPharmacol 417:189–94
– Hinks GL, Brown P, Field M, Poat JA, Hughes J(1996) The anxiolytics CI-988 and chlordiazepoxidefail to reduce immediate early gene mRNA stimulationfollowing exposure to the rat elevated X-maze. Eur JPharmacol 312:153–161
– Hiroi N, Graybiel AM (1996) Atypical and typicalneuroleptic treatments induce distinct programs oftranscription factor expression in the striatum. J CompNeurol 374:70–83
– Hughes P, Dragunow M, Beilharz E, Lawlor P,Gluckman P (1993) MK801 induces immediate-earlygene proteins and BDNF mRNA in rat cerebrocorticalneurones. Neuroreport 4:183–186
– Hurley MJ, Stubbs CM, Jenner P, Marsden CD (1996)Dopamine D3 receptors are not involved in theinduction of c-fos mRNA by neuroleptic drugs:comparison of the dopamine D3 receptor antagonistGR103691 with typical and atypical neuroleptics. EurJ Pharmacol 318:283–293
– Hussain N, Flumerfelt BA, Rajakumar N (2001)Glutamatergic regulation of haloperidol-induced c-fosexpression in the rat striatum and nucleus accumbens.Neuroscience 102:391–399
– Imaki T, Wang XQ, Shibasaki T, Harada S, ChikadaN, Takahashi C, Naruse M, Demura H (1995) Chlor-diazepoxide attenuates stress-induced activation ofneurons, corticotropin-releasing factor (CRF) genetranscription and CRF biosynthesis in the paraventric-ular nucleus (PVN). Mol Brain Res 32:261–270
– Ishibashi T, Tagashira R, Nakamura M, Noguchi H,Ohno Y (1999) Effects of perospirone, a novel 5-HT2and D2 receptor antagonist, on Fos protein expressionin the rat forebrain. Pharmacol Biochem Behav63:535–541
– Jaber M, Cador M, Dumartin B, Normand E, Stinus L,Bloch B (1995) Acute and chronic amphetaminetreatments differently regulate neuropeptide messengerRNA levels and Fos immunoreactivity in rat striatalneurons. Neuroscience 65:1041–1050
– Javed A, Van De Kar LD, Gray TS (1997) p-Chlorophenylalanine and fluoxetine inhibit d-fenflur-amine-induced Fos expression in the paraventricularnucleus, cingulate cortex and frontal cortex but not inother forebrain and brainstem regions. Brain Res774:94–105
– Johansson B, Lindstrom K, Fredholm BB (1994)Differences in the regional and cellular localizationof c-fos messenger RNA induced by amphetamine,cocaine and caffeine in the rat. Neuroscience 59:837–849
– Kawashima N, Nakamura A, Okuyama S, Chaki S,Tomisawa K (1999) Effects of NRA0045, NRA0160,and NRA0215 on regional Fos-like immunoreactivityin the rat brain. Gen Pharmacol 32:637–646
– Kovacs KJ, Csejtei M, Laszlovszky I (2001) Doubleactivity imaging reveals distinct cellular targets ofhaloperidol, clozapine and dopamine D3 receptorselective RGH-1756. Neuropharmacology 40:383–393
– Lee S, Rivier C, Torres G (1994) Induction of c-fosand CRF mRNA by MK-801 in the parvocellularparaventricular nucleus of the rat hypothalamus. MolBrain Res 24:192–198
– Lillrank SM, Lipska BK, Bachus SE, Wood GK,Weinberger DR (1996) Amphetamine-induced c-fosmRNA expression is altered in rats with neonatalventral hippocampal damage. Synapse 23:292–301
– Lino-de-Oliveira C, Sales AJ, Del Bel EA, SilveiraMC, Guimaraes FS (2001) Effects of acute and chronicfluoxetine treatments on restraint stress-induced Fosexpression. Brain Res Bull 55:747–754
– MacGibbon GA, Lawlor PA, Bravo R, Dragunow M(1994) Clozapine and haloperidol produce a differen-tial pattern of immediate early gene expression in ratcaudate-putamen, nucleus accumbens, lateral septumand islands of Calleja. Mol Brain Res 23:21–32
– Mao L, Wang JQ (2002) Activation of metabotropicglutamate receptor mediates upregulation of transcrip-tion factor mRNA expression in rat striatum inducedby acute administration of amphetamine. Brain Res924:167–175
– Mathieu-Kia A-M, Pages C, Besson M-J (1998)Inducibility of c-fos protein in visuo-motor systemand limbic structures after acute and repeated admin-istration of nicotine in the rat. Synapse 29:343–354
– Merchant KM, Hanson GR, Dorsa DM (1994) Induc-tion of neurotensin and c-fos mRNA in distinctsubregions of rat neostriatum after acute methamphet-amine: comparison with acute haloperidol effects. JPharmacol Exp Ther 269:806–812
– Morelli M, Pinna A (1999) Antidepressants andatypical neuroleptics induce Fos-like immunoreactiv-ity in the central extended amygdala. Ann. N. Y. Acad.Sci. 877:703–706
– Morelli M, Pinna A, Ruiu S, Del Zompo M (1999)Induction of Fos-like immunoreactivity in the centralextended amygdala by antidepressant drugs. Synapse31:1–4
316

– Nakki R, Sharp FR, Sagar SM, Honkaniemi J (1996)Effects of phencyclidine on immediate early geneexpression in the brain. J Neuroscience Res 45:13–27
– Ohashi K, Hamamura T, Fujiwara Y, Suzuki H,Kuroda S (2000) Clozapine- and olanzapine-inducedFos expression in the rat medial prefrontal cortex ismediated by beta-adrenoceptors. Neuropsychopharma-cology 23:162–169
– Palacios G, Muro MA, Paz Marin A (1996) Differen-tial effects of haloperidol and two anxiolytic drugs,buspirone and lesopitron, on c-Fos expression in the ratstriatum and nucleus accumbens. Brain Res 742:141–148
– Palmer LC, Hess US, Larson J, Rogers GA, Gall CM,Lynch G (1997) Comparison of the effects of anampakine with those of methamphetamine on aggre-gate neuronal activity in cortex versus striatum. MolBrain Res 46:127–135
– Panegyres PK, Hughes J (1997) Activation of c-fosmRNA in the brain by the kappa-opioid receptoragonist enadoline and the NMDA receptor antagonistdizocilpine. Eur J Pharmacol 328:31–36
– Pinna A, Wardas J, Cozzolino A, Morelli M (1999)Involvement of adenosine A2A receptors in the induc-tion of c-fos expression by clozapine and haloperidol.Neuropsychopharmacology 20:44–51
– Robertson GS, Fibiger HC (1996) Effects of olanza-pine on regional c-fos expression in rat forebrain.Neuropsychopharmacology 14:105–110
– Robertson GS, Matsumura H, Fibiger HC (1994)Induction patterns of Fos-like immunoreactivity in theforebrain as predictors of atypical antipsychotic activ-ity. J Pharmacol Exp Ther 271:1058–1066
– Ruskin DN, Marshall JF (1994) Amphetamine- andcocaine-induced Fos in the rat striatum depends on D2receptor activation. Synapse 18:233–240
– Salminen O, Lahtinen S, Ahtee L (1996) Expression ofFos protein in various rat brain areas following acutenicotine and diazepam. Pharmacol Biochem Behav54:241–248
– Sato D, Umino A, Kaneda K, Takigawa M, NishikawaT (1997) Developmental changes in distribution pat-terns of phencyclidine-induced c-Fos in rat forebrain.Neurosci Lett 239:21–24
– Schilstrom B, de Villiers S, Malmerfelt A, SvenssonTH, Nomikos GG (2000) Nicotine-induced Fos ex-pression in the nucleus accumbens and the medialprefrontal cortex in the rat: role of nicotinic andNMDA receptors in the ventral tegmental area.Synapse 36:314–321
– Sebens JB, Koch T, Korf J (1996) Lack of cross-tolerance between haloperidol and clozapine towardsFos-protein induction in rat forebrain regions. Eur JPharmacol 315:269–275
– Semba J, Sakai M, Miyoshi R, Mataga N, FukamauchiF, Kito S (1996) Differential expression of c-fosmRNA in rat prefrontal cortex, striatum, N. accumbensand lateral septum after typical and atypical antipsy-
chotics: an in situ hybridization study. Neurochem Int29:435–442
– Semba J, Sakai MW, Suhara T, Akanuma N (1999)Differential effects of acute and chronic treatment withtypical and atypical neuroleptics on c-fos mRNAexpression in rat forebrain regions using non-radioac-tive in situ hybridization. Neurochem Int 34:269–277
– Semba J, Tanaka N, Wakuta M, Suhara T (2001)Neonatal phencyclidine treatment selectively attenu-ates mesolimbic dopamine function in adult rats asrevealed by methamphetamine-induced behavior andc-fos mRNA expression in the brain. Synapse 40:11–18
– Seppa T, Salminen O, Moed M, Ahtee L (2001)Induction of Fos-immunostaining by nicotine andnicotinic receptor antagonists in rat brain. Neurophar-macology 41:486–495
– Sharp JW (1997) Phencyclidine (PCP) acts at sigmasites to induce c-fos gene expression. Brain Res758:51–58
– Singewald N, Sharp T (2000) Neuroanatomical targetsof anxiogenic drugs in the hindbrain as revealed by Fosimmunocytochemistry. Neuroscience 98:759–770
– Singewald N, Salchner P, Sharp T (2003) Induction ofc-fos expression in specific areas of the fear circuitryin rat forebrain by anxiogenic drugs. Biol Psychiatry53:275–283
– Steiner H, Gerfen CR (1993) Cocaine-induced c-fosmessenger RNA is inversely related to dynorphinexpression in striatum. J Neurosci 13:5066–5081
– Sun YJ, Suzuki M, Kurachi T, Murata M, Kurachi M(1998) Expression of Fos protein in the limbic regionsof the rat following haloperidol decanoate. Brain Res791:125–136
– Suzuki M, Sun YJ, Murata M, Kurachi M (1998)Widespread expression of Fos protein induced byacute haloperidol administration in the rat brain.Psychiatry Clin Neurosci 52:353–359
– Torres G (1994) Acute administration of alcoholblocks cocaine-induced striatal c-fos immunoreactivityprotein in the rat. Synapse 18:161–167
– Torres G, Horowitz JM, Laflamme N, Rivest S (1998)Fluoxetine induces the transcription of genes encodingc-fos, corticotropin-releasing factor and its type 1receptor in rat brain. Neuroscience 87:463–477
– Turgeon SM, Case LC (2001) The effects of phency-clidine pretreatment on amphetamine-induced behav-ior and c-Fos expression in the rat. Brain Res 888:302–305
– Turgeon SM, Roche JK (1999) The delayed effects ofphencyclidine enhance amphetamine-induced behaviorand striatal c-Fos expression in the rat. Neuroscience91:1265–1275
– Umino A, Nishikawa T, Takahashi K (1995) Meth-amphetamine-induced nuclear c-Fos in rat brainregions. Neurochem Int 26:85–90
– Uslaner J, Badiani A, Norton CS, Day HE, Watson SJ,Akil H, Robinson TE (2001) Amphetamine andcocaine induce different patterns of c-fos mRNA
317

expression in the striatum and subthalamic nucleusdepending on environmental context. Eur J Neurosci13:1977–1983
– Vahid-Ansari F, Robertson GS (1996) 7-OH-DPATdifferentially reverses clozapine- and haloperidol-induced increases in Fos-like immunoreactivity in therodent forebrain. Eur J Neurosci 8:2605–2611
– Wan W, Ennulat DJ, Cohen BM (1995) Acuteadministration of typical and atypical antipsychoticdrugs induces distinctive patterns of Fos expression inthe rat forebrain. Brain Res 688:95–104
– Wang JQ, Smith AJ, McGinty JF (1995) A singleinjection of amphetamine or methamphetamine in-duces dynamic alterations in c-fos, zif/268 and prepro-dynorphin messenger RNA expression in rat forebrain.Neuroscience 68:83–95
– Wedzony K, Czyrak A (1996) Competitive and non-competitive NMDA receptor antagonists induce c-Fosexpression in the rat anterior cingulate cortex. JPhysiol Pharmacol 47:525–533
– Werme M, Ringholm A, Olson L, Brene S (2000)Differential patterns of induction of NGFI-B, Nor1 andc-fos mRNAs in striatal subregions by haloperidol andclozapine. Brain Res 863:112–119
– Wintrip N, Nance DM, Wilkinson M (1998) Sexuallydimorphic MK801-induced c-fos in the rat hypotha-lamic paraventricular nucleus. Neurosci Lett 242:151–154
– Young CD, Meltzer HY, Deutch AY (1998) Effects ofdesmethylclozapine on Fos protein expression in theforebrain: in vivo biological activity of the clozapinemetabolite. Neuropsychopharmacology 19:99–103
References
Alheid GF, Heimer L (1996) Theories of basal forebrain organi-zation and the ‘emotional motor system’. Prog Brain Res107:461–484
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990)Basic local alignment search tool. J Mol Biol 215:403–410
Badiani A, Oates MM, Day HE, Watson SJ, Akil H, Robinson TE(1998) Amphetamine-induced behavior, dopamine release, andc-fos mRNA expression: modulation by environmental novelty.J Neurosci 18:10579–10593
Baldessarini RJ (1990) Drugs and the treatment of psychiatricdisorders. In: Gilman AG, Rall TW, Nies AS, Taylor P (eds)Goodman and Gilman’s pharmacological basis of therapeutics,8th edn. Pergamon Press, New York, pp 383–435
Beck CHM (1995) Acute treatment with antidepressant drugsselectively increases the expression of c-fos in the rat brain. JPsychiatry Neurosci 20:25–32
Beckmann AM, Wilce PA (1997) EGR transcription factors in thenervous system. Neurochem Int 31:477–510
Bennett HJ, Semba K (1998) Immunohistochemical localization ofcaffeine-induced c-fos protein expression in the rat brain. JComp Neurol 401:89–108
Bonaventure P, Guo H, Tian B, Liu X, Bittner A, Roland B,Salunga R, Ma X-J, Kamme F, Meurers B, Bakker M, JurzakM, Leysen JE, Erlander MG (2002) Nuclei and subnuclei geneexpression profiling in mammalian brain. Brain Res 943:38–47
Bujas-Bobanovic M, Robertson HA, Dursun SM (2000) Effects ofnitric oxide synthase inhibitor N(G)-nitro-l-arginine methyl
ester on phencyclidine-induced effects in rats. Eur J Pharmacol409:57–65
Carta AR, Gerfen CR (1999) Lack of a role for the D3 receptor inclozapine induction of c-fos demonstrated in D3 dopaminereceptor-deficient mice. Neuroscience 90:1021–1029
Castner SA, Becker JB (1996) Sex differences in the effect ofamphetamine on immediate early gene expression in the ratdorsal striatum. Brain Res 712:245–257
Cenci MA, Kalen P, Mandel RJ, Wictorin K, Bjorklund A (1992)Dopaminergic transplants normalize amphetamine- and apo-morphine-induced Fos expression in the 6-hydroxydopamine-lesioned striatum. Neuroscience 46:943–957
Chaudhuri A (1997) Neural activity mapping with inducibletranscription factors Neuroreport 8:iii–vii
Cochran SM, McKerchar CE, Morris BJ, Pratt JA (2002) Inductionof differential patterns of local cerebral glucose metabolism andimmediate-early genes by acute clozapine and haloperidol.Neuropharmacology 43:394–407
Cohen BM, Wan W (1996) The thalamus as a site of action ofantipsychotic drugs. Am J Psychiatry 153:104–106
Cookson J, Taylor D, Katonia C (2002) Use of drugs in psychiatry,5th edn. Gaskell, Royal College of Psychiatrists, London
Davis M, Shi C (1999) The central extended amygdala: are thecentral nucleus of the amygdala and the bed nucleus of the striaterminalis differentially involved in fear versus anxiety? AnnNY Acad Sci 877:281–291
Dawe GS, Huff KD, Vandergriff JL, Sharp TL, O’Neill MJ,Rasmussen K (2001) Olanzapine activates the rat locuscoeruleus: in vivo electrophysiology and c-fos immunoreactiv-ity. Biol Psychiatry 50:510–520
Day HE, Badiani A, Uslaner JM, Oates MM, Vittoz NM, RobinsonTE, Watson SJ Jr, Akil H (2001) Environmental noveltydifferentially affects c-fos mRNA expression induced byamphetamine or cocaine in subregions of the bed nucleus ofthe stria terminalis and amygdala. J Neurosci 21:732–740
De Boer T, Nefkens F, Van Helvoirt A (1994) The alpha2-adrenoceptor antagonist Org 3770 enhances serotonin trans-mission in vivo. Eur J Pharmacol 253: R5–R6
Deutsch AY, Duman RS (1996) The effects of antipsychotic drugson Fos protein expression in the prefrontal cortex: cellularlocalization and pharmacological characterization. Neurosci-ence 70:377–389
Deutsch AY, Ongur D, Duman RS (1995) Antipsychotic drugsinduce Fos protein in the thalamic paraventricular nucleus: anovel locus of antipsychotic drug action. Neuroscience 66:337–346
Dezhi M, Liang W, Zhang G, Xiru W (1999) The relationshipbetween the c-jun mRNA expression and apoptosis of neuronsin rat brain following perinatal ischemic-hypoxia. Chin Med J(Engl) 112:40–43
Dragunow M, Faull R (1989) The use of c-fos as a metabolicmarker in neuronal pathway tracing. J Neurosci Methods29:261–265
Dragunow M, Faull RL (1990) MK801 induces c-fos protein inthalamic and neocortical neurons of rat brain. Neurosci Lett113:144–150
Duncan GE, Knapp DJ, Johnson KB, Breese GR (1996) Functionalclassification of antidepressants based on antagonism of swim-stress-induced fos-like immunoreactivity. J Pharmacol ExpTher 277:1076–1089
Duncan GE, Moym SS, Knapp DJ, Mueller RA, Breese GR (1998)Metabolic mapping of the rat brain after subanesthetic doses ofketamine: potential relevance to schizophrenia. Brain Res787:181–190
Engber TM, Koury EJ, Dennis SA, Miller MS, Contreras PC, BhatRV (1998) Differential patterns of regional c-Fos induction inthe rat brain by amphetamine and the novel wakefulness-promoting agent modafinil. Neurosci Lett 241:95–98
Fink-Jensen A, Kristensen P (1994) Effects of typical and atypicalneuroleptics on Fos protein expression in the rat forebrain.Neurosci Lett 182:115–118
318

Fink-Jensen A, Ludvigsen TS, Korsgaard N (1995) The effect ofclozapine on Fos protein immunoreactivity in the rat forebrainis not mimicked by the addition of alpha 1-adrenergic or 5HT2receptor blockade to haloperidol. Neurosci Lett 194: 77–80
Frodl T, Meisenzahl E, Zetzsche T, Bottlender R, Born C, Groll C,Jager M, Leinsinger G, Hahn K, Moller H-J (2002) Enlarge-ment of the amygdala in patients with a first episode of majordepression. Biol Psychiatry 51:708–714
Fujimura M, Hashimoto K, Yamagami K (2000a) Effects ofantipsychotic drugs on neurotoxicity: expression of Fos-likeprotein and c-fos mRNA in the retrosplenial cortex afteradministration of dizocilpine. Eur J Pharmacol 398:1–10
Fujimura M, Hashimoto K, Yamagami K (2000b) The effect of theantipsychotic drug mosapramine on the expression of Fosprotein in the rat brain: comparison with haloperidol, clozapineand risperidone. Life Sci 67:2865–2872
Gao XM, Hashimoto T, Tamminga CA (1998) Phencyclidine (PCP)and dizocilpine (MK801) exert time-dependent effects on theexpression of immediate early genes in rat brain. Synapse29:14–28
Gass P, Herdegen T, Bravo R, Kiessling M (1993) Induction andsuppression of immediate early genes in specific rat brainregions by the non-competitive N-methyl-d-aspartate receptorantagonist MK-801. Neuroscience 53:749–758
Guitart X, Farre AJ (1998) The effect of E-5842, a sigma receptorligand and potential atypical antipsychotic, on Fos expressionin rat forebrain. Eur J Pharmacol 363:127–130
Guo N, Klitenick MA, Tham CS, Fibiger HC (1995) Receptormechanisms mediating clozapine-induced c-fos expression inthe forebrain. Neuroscience 65:747–756
Habara T, Hamamura T, Miki M, Ohashi K, Kuroda S (2001)M100,907, a selective 5-HT(2A) receptor antagonist, attenuatesphencyclidine-induced Fos expression in discrete regions of ratbrain. Eur J Pharmacol 417:189–194
Hamamura T, Ichimaru Y, Fibiger HC (1997) Amphetaminesensitization enhances regional c-fos expression produced byconditioned fear. Neuroscience 76:1097–1103
Herdegen T, Leah JD (1998) Inducible and constitutive transcrip-tion factors in the mammalian nervous system: control of geneexpression by Jun, Fos and Krox, and CREB/ATF proteins.Brain Res Rev 28:370–490
Herrera DG, Robertson HA (1996) Activation of c-fos in the brain.Progr Neurobiol 50:83–107
Hess US, Lynch G, Gall CM (1995) Regional patterns of c-fosmRNA expression in rat hippocampus following exploration ofa novel environment versus performance of a well-learneddiscrimination. J Neurosci 15:7796–7809
Hinks GL, Brown P, Field M, Poat JA, Hughes J (1996) Theanxiolytics CI-988 and chlordiazepoxide fail to reduce imme-diate early gene mRNA stimulation following exposure to therat elevated X-maze. Eur J Pharmacol 312:153–161
Hiroi N, Graybiel AM (1996) Atypical and typical neuroleptictreatments induce distinct programs of transcription factorexpression in the striatum. J Comp Neurol 374:70–83
Hoffman GE, Lyo D (2002) Anatomical markers of activity inneuroendocrine systems. Are we all ‘Fos-ed’ out? J Neuroen-docrinol 14:259–268
Hughes P, Dragunow M, Beilharz E, Lawlor P, Gluckman P (1993)MK801 induces immediate-early gene proteins and BDNFmRNA in rat cerebrocortical neurones. Neuroreport 4:183–186
Hurley MJ, Stubbs CM, Jenner P, Marsden CD (1996) DopamineD3 receptors are not involved in the induction of c-fos mRNAby neuroleptic drugs: comparison of the dopamine D3 receptorantagonist GR103691 with typical and atypical neuroleptics.Eur J Pharmacol 318:283–293
Hussain N, Flumerfelt BA, Rajakumar N (2001) Glutamatergicregulation of haloperidol-induced c-fos expression in the ratstriatum and nucleus accumbens. Neuroscience 102:391–399
Imaki T, Wang XQ, Shibasaki T, Harada S, Chikada N, TakahashiC, Naruse M, Demura H (1995) Chlordiazepoxide attenuatesstress-induced activation of neurons, corticotropin-releasing
factor (CRF) gene transcription and CRF biosynthesis in theparaventricular nucleus (PVN). Mol Brain Res 32:261–270
Ishibashi T, Tagashira R, Nakamura M, Noguchi H, Ohno Y (1999)Effects of perospirone, a novel 5-HT2 and D2 receptorantagonist, on Fos protein expression in the rat forebrain.Pharmacol Biochem Behav 63:535–541
Jaber M, Cador M, Dumartin B, Normand E, Stinus L, Bloch B(1995) Acute and chronic amphetamine treatments differentlyregulate neuropeptide messenger RNA levels and Fos immu-noreactivity in rat striatal neurons. Neuroscience 65:1041–1050
Javed A, Van De Kar LD, Gray TS (1997) p-Chlorophenylalanineand fluoxetine inhibit d-fenfluramine-induced Fos expressionin the paraventricular nucleus, cingulate cortex and frontalcortex but not in other forebrain and brainstem regions. BrainRes 774:94–105
Johansson B, Lindstrom K, Fredholm BB (1994) Differences in theregional and cellular localization of c-fos messenger RNAinduced by amphetamine, cocaine and caffeine in the rat.Neuroscience 59:837–849
Jones MW, Errington ML, French PJ, Fine A, Bliss TVP, Garel S,Charnay P, Bozon B, Laroche S, Davis S (2001) A requirementfor the immediate early gene zif268 in the expression of lateLTP and long-term memories. Nat Neurosci 4:289–296
Kaczmarek L, Robertson HA (2002) Immediate early genes andinducible transcription factors in mapping of the centralnervous system function and dysfunction. In: Bjorklund A,Hokfelt T (eds) Handbook of chemical neuroanatomy, vol 19.Elsevier, Amsterdam
Kawashima N, Nakamura A, Okuyama S, Chaki S, Tomisawa K(1999) Effects of NRA0045, NRA0160, and NRA0215 onregional Fos-like immunoreactivity in the rat brain. GenPharmacol 32:637–646
Komisaruk BR, Rosenblatt JS, Barona ML, Chinapen S, NissanovJ, O’Brannon RT, Johnson BM, Del Cerro MCR (2000)Combined c-fos and C-14-2-deoxyglucose method to differen-tiate site-specific excitation from disinhibition: analysis ofmaternal behaviour in the rat. Brain Res 859:262–272
Kovacs KJ, Csejtei M, Laszlovszky I (2001) Double activityimaging reveals distinct cellular targets of haloperidol, cloza-pine and dopamine D3 receptor selective RGH-1756. Neuro-pharmacology 40:383–393
Labiner DM, Butler LS, Cao Z, Hosford DA, Shin C, McNamaraJO (1993) Induction of c-fos mRNA by kindled seizures:complex relationship with neuronal burst firing. J Neurosci13:744–751
Lee S, Rivier C, Torres G (1994) Induction of c-fos and CRFmRNA by MK-801 in the parvocellular paraventricular nucleusof the rat hypothalamus. Mol Brain Res 24:192–198
Leslie RA, James MF (2000) Pharmacological magnetic resonanceimaging: a new application for functional MRI. TrendsPharmacol Sci 21:314–318
Lillrank SM, Lipska BK, Bachus SE, Wood GK, Weinberger DR(1996) Amphetamine-induced c-fos mRNA expression isaltered in rats with neonatal ventral hippocampal damage.Synapse 23:292–301
Lino-de-Oliveira C, Sales AJ, Del Bel EA, Silveira MC, GuimaraesFS (2001) Effects of acute and chronic fluoxetine treatments onrestraint stress-induced Fos expression. Brain Res Bull 55:747–754
Lyford GL, Yamagata K, Kaufmann WE, Barnes CA, Sanders LK,Copeland NG, Gilbert DJ, Jenkins NA, Lanahan AA, WorleyPF (1995) Arc, a growth factor and activity-regulated gene,encodes a novel cytoskeleton-associated protein that is enrichedin neuronal dendrites. Neuron 14:433–445
MacGibbon GA, Lawlor PA, Bravo R, Dragunow M (1994)Clozapine and haloperidol produce a differential pattern ofimmediate early gene expression in rat caudate-putamen,nucleus accumbens, lateral septum and islands of Calleja.Mol Brain Res 23:21–32
Mach RH, Nader MA, Ehrenkaufer RLE, Line SW, Smith CR,Gage D, Morton TE (1997) Use of positron emission
319

tomography to study the dynamics of psychostimulant-induceddopamine release. Pharmacol Biochem Behav 57:477–486
Mao L, Wang JQ (2002) Activation of metabotropic glutamatereceptor mediates upregulation of transcription factor mRNAexpression in rat striatum induced by acute administration ofamphetamine. Brain Res 924:167–175
Mathieu-Kia A-M, Pages C, Besson M-J (1998) Inducibility of c-fos protein in visuo-motor system and limbic structures afteracute and repeated administration of nicotine in the rat. Synapse29:343–354
Martin P, Carlsson ML, Hjorth S (1998) Systemic PCP treatmentelevates brain extracellular 5-HT: a microdialysis study inawake rats. Neuroreport 9:2985–2988
Menza M, Marin H, Opper RS (2003) Residual symptoms indepression: can treatment be symptom specific? J Clin Psychi-atry 64:516–523
Merchant KM, Hanson GR, Dorsa DM (1994) Induction ofneurotensin and c-fos mRNA in distinct subregions of ratneostriatum after acute methamphetamine: comparison withacute haloperidol effects. J Pharmacol Exp Ther 269:806–812
Miller DD, Andreasen NC, O’Leary DS, Watkins GL, Ponto LLB,Hichwa RD (2001) Comparison of the effects of risperidoneand haloperidol on regional cerebral blood flow in schizophre-nia. Biol Psychiatry 49:704–715
Miyamoto S, Duncan GE, Mailman RB, Lieberman JA (2000)Developing novel antipsychotic drugs: strategies and goals.Curr Opin Investig Drugs 2:25–39
Morinobu S, Strausbaugh H, Terwilliger R, Duman RS (1997)Regulation of c-fos and NGFI-A by antidepressant treatments.Synapse 25:313–320
Morelli M, Pinna A (1999) Antidepressants and atypical neurolep-tics induce Fos-like immunoreactivity in the central extendedamygdala. Ann NY Acad Sci 877:703–706
Morelli M, Pinna A, Ruiu S, Del Zompo M (1999) Induction ofFos-like immunoreactivity in the central extended amygdala byantidepressant drugs. Synapse 31:1–4
Nakki R, Sharp FR, Sagar SM, Honkaniemi J (1996) Effects ofphencyclidine on immediate early gene expression in the brain.J Neurosci Res 45:13–27
Ohashi K, Hamamura T, Fujiwara Y, Suzuki H, Kuroda S (2000)Clozapine- and olanzapine-induced Fos expression in the ratmedial prefrontal cortex is mediated by beta-adrenoceptors.Neuropsychopharmacology 23:162–169
Palacios G, Muro MA, Paz Marin A (1996) Differential effects ofhaloperidol and two anxiolytic drugs, buspirone and lesopitron,on c-Fos expression in the rat striatum and nucleus accumbens.Brain Res 742:141–148
Palmer LC, Hess US, Larson J, Rogers GA, Gall CM, Lynch G(1997) Comparison of the effects of an ampakine with those ofmethamphetamine on aggregate neuronal activity in cortexversus striatum. Mol Brain Res 46:127–135
Panegyres PK, Hughes J (1997) Activation of c-fos mRNA in thebrain by the kappa-opioid receptor agonist enadoline and theNMDA receptor antagonist dizocilpine. Eur J Pharmacol328:31–36
Paxinos G, Watson C (1997) The rat brain in stereotaxiccoordinates, compact 3rd edn. Academic Press, San Diego
Pei Q, Lewis L, Sprakes ME, Jones EJ, Grahame-Smith DG,Zetterstrom TSC (2000) Serotonic regulation of mRNAexpression of Arc, an immediate early gene selectivelylocalized at neuronal dendrites. Neuropharmacology 39:463–470
Pinna A, Wardas J, Cozzolino A, Morelli M (1999) Involvement ofadenosine A2A receptors in the induction of c-fos expression byclozapine and haloperidol. Neuropsychopharmacology 20:44–51
Preece M, Mukherjee B, Huang CL-H, Hall LD, Leslie RA, JamesMF (2001) Detection of pharmacologically mediated changesin cerebral activity by functional magnetic resonance imaging:the effects of sulpiride in the brain of the anaesthetised rat.Brain Res 916:107–114
Robertson GS, Fibiger HC (1996) Effects of olanzapine on regionalc-fos expression in rat forebrain. Neuropsychopharmacology14:105–110
Robertson GS, Matsumura H, Fibiger HC (1994) Induction patternsof Fos-like immunoreactivity in the forebrain as predictors ofatypical antipsychotic activity. J Pharmacol Exp Ther271:1058–1066
Ruigt GSF, van Proosdij JN, van Wezenbeek LACM (1989) Alarge-scale, high-resolution, automated system for rat sleepstaging. I. Methodology and technical aspects. II. Validationand application. Electroencephalogr Clin Neurophysiol 73:52–71
Ruigt GSF, Engelen S, Gerrits A, Verbon F (1993) Computer-basedprediction of psychotropic drug classes based on a discriminantanalysis of drug effects on rat sleep. Neuropsychobiology28:138–153
Ruskin DN, Marshall JF (1994) Amphetamine- and cocaine-induced Fos in the rat striatum depends on D2 receptoractivation. Synapse 18:233–240
Salminen O, Lahtinen S, Ahtee L (1996) Expression of Fos proteinin various rat brain areas following acute nicotine anddiazepam. Pharmacol Biochem Behav 54:241–248
Sato D, Umino A, Kaneda K, Takigawa M, Nishikawa T (1997)Developmental changes in distribution patterns of phencycli-dine-induced c-Fos in rat forebrain. Neurosci Lett 239:21–24
Schilstrom B, de Villiers S, Malmerfelt A, Svensson TH, NomikosGG (2000) Nicotine-induced Fos expression in the nucleusaccumbens and the medial prefrontal cortex in the rat: role ofnicotinic and NMDA receptors in the ventral tegmental area.Synapse 36:314–321
Sebens JB, Koch T, Korf J (1996) Lack of cross-tolerance betweenhaloperidol and clozapine towards Fos-protein induction in ratforebrain regions. Eur J Pharmacol 315:269–275
Seeman P (2002) Atypical antipsychotics: mechanism of action.Can J Psychiatry 47:27–38
Semba J, Sakai M, Miyoshi R, Mataga N, Fukamauchi F, Kito S(1996) Differential expression of c-fos mRNA in rat prefrontalcortex, striatum, N. accumbens and lateral septum after typicaland atypical antipsychotics: an in situ hybridization study.Neurochem Int 29:435–442
Semba J, Sakai MW, Suhara T, Akanuma N (1999) Differentialeffects of acute and chronic treatment with typical and atypicalneuroleptics on c-fos mRNA expression in rat forebrain regionsusing non-radioactive in situ hybridization. Neurochem Int34:269–277
Semba J, Tanaka N, Wakuta M, Suhara T (2001) Neonatalphencyclidine treatment selectively attenuates mesolimbicdopamine function in adult rats as revealed by methamphet-amine-induced behavior and c-fos mRNA expression in thebrain. Synapse 40:11–18
Seppa T, Salminen O, Moed M, Ahtee L (2001) Induction of Fos-immunostaining by nicotine and nicotinic receptor antagonistsin rat brain. Neuropharmacology 41:486–495
Sharp FR, Sagar SM, Swanson RA (1993) Metabolic mapping withcellular resolution: c-fos vs 2-deoxyglucose. Crit Rev Neuro-biol 7:205–228
Sharp JW (1997) Phencyclidine (PCP) acts at sigma sites to inducec-fos gene expression. Brain Res 758:51–58
Sheng M, Greenberg ME (1990) The regulation and function of c-fos and other immediate early genes in the nervous system.Neuron 4:477–485
Siegle GJ, Steinhauer SR, Thase ME, Stenger VA, Carter CS(2002) Can’t shake that feeling: event-related fMRI assessmentof sustained amygdala activity in response to emotionalinformation in depressed individuals. Biol Psychiatry 51:693–707
Singewald N, Sharp T (2000) Neuroanatomical targets of anxio-genic drugs in the hindbrain as revealed by Fos immunocyto-chemistry. Neuroscience 98:759–770
Singewald N, Salchner P, Sharp T (2003) Induction of c-fosexpression in specific areas of the fear circuitry in rat forebrainby anxiogenic drugs. Biol Psychiatry 53:275–283
320

Steiner H, Gerfen CR (1993) Cocaine-induced c-fos messengerRNA is inversely related to dynorphin expression in striatum. JNeurosci 13:5066–5081
Sun YJ, Suzuki M, Kurachi T, Murata M, Kurachi M (1998)Expression of Fos protein in the limbic regions of the ratfollowing haloperidol decanoate. Brain Res 791:125–136
Suzuki M, Sun YJ, Murata M, Kurachi M (1998) Widespreadexpression of Fos protein induced by acute haloperidoladministration in the rat brain. Psychiatry Clin Neurosci52:353–359
Torres G (1994) Acute administration of alcohol blocks cocaine-induced striatal c-fos immunoreactivity protein in the rat.Synapse 18:161–167
Torres G, Horowitz JM, Laflamme N, Rivest S (1998) Fluoxetineinduces the transcription of genes encoding c-fos, corticotropin-releasing factor and its type 1 receptor in rat brain. Neurosci-ence 87:463–477
Turgeon SM, Case LC (2001) The effects of phencyclidinepretreatment on amphetamine-induced behavior and c-Fosexpression in the rat. Brain Res 888:302–305
Turgeon SM, Roche JK (1999) The delayed effects of phencycli-dine enhance amphetamine-induced behavior and striatal c-Fosexpression in the rat. Neuroscience 91:1265–1275
Umino A, Nishikawa T, Takahashi K (1995) Methamphetamine-induced nuclear c-Fos in rat brain regions. Neurochem Int26:85–90
Uslaner J, Badiani A, Norton CS, Day HE, Watson SJ, Akil H,Robinson TE (2001) Amphetamine and cocaine induce differ-ent patterns of c-fos mRNA expression in the striatum andsubthalamic nucleus depending on environmental context. Eur JNeurosci 13:1977–1983
Vahid-Ansari F, Robertson GS (1996) 7-OH-DPAT differentiallyreverses clozapine- and haloperidol-induced increases in Fos-like immunoreactivity in the rodent forebrain. Eur J Neurosci8:2605–2611
VanderSpek SC, Nobrega J, Brownlee BA, Seeman P, Kapur S(2002) Animal studies of antipsychotics use doses which arenot representative of the clinical condition—potential implica-tions and suggested solutions. Biol Psychiatry 51:121S
Volkow ND, Wang G-J, Fowler JS, Gatley SJ, Ding Y-S, Logan J,Dewey SL, Hitzeman R, Lieberman J (1996) Relationship
between psychostimulant-induced “high” and dopamine trans-porter occupancy. Proc Natl Acad Sci USA 93:10388–10392
Wan W, Ennulat DJ, Cohen BM (1995) Acute administration oftypical and atypical antipsychotic drugs induces distinctivepatterns of Fos expression in the rat forebrain. Brain Res688:95–104
Wang JQ, Smith AJ, McGinty JF (1995) A single injection ofamphetamine or methamphetamine induces dynamic alterationsin c-fos, zif/268 and preprodynorphin messenger RNA expres-sion in rat forebrain. Neuroscience 68:83–95
Wedzony K, Czyrak A (1996) Competitive and non-competitiveNMDA receptor antagonists induce c-Fos expression in the ratanterior cingulate cortex. J Physiol Pharmacol 47:525–533
Werme M, Ringholm A, Olson L, Brene S (2000) Differentialpatterns of induction of NGFI-B, Nor1 and c-fos mRNAs instriatal subregions by haloperidol and clozapine. Brain Res863:112–119
Wintrip N, Nance DM, Wilkinson M (1998) Sexually dimorphicMK801-induced c-fos in the rat hypothalamic paraventricularnucleus. Neurosci Lett 242:151–154
Wirtshafter D, Asin KE (1999) Haloperidol induces c-fos expres-sion in the globus pallidus and substantia nigra of cynomolgusmonkeys. Brain Res 835:154–161
Wisden W, Morris BJ (eds) (1994) In situ hybridization protocolsfor the brain. Academic Press, London
Wittliff JL, Erlander MG (2002) Laser capture microdissection andits applications in genomics and proteomics. Methods Enzymol356:12–25
Wrynne AS, Sebens JB, Koch T, Leonard BE, Korf J (2000)Prolonged c-jun expression in the basolateral amygdalafollowing bulbectomy: possible implications for antidepressantactivity and time of onset. Mol Brain Res 76:7–17
Young CD, Meltzer HY, Deutch AY (1998) Effects of desmethyl-clozapine on Fos protein expression in the forebrain: in vivobiological activity of the clozapine metabolite. Neuropsycho-pharmacology 19:99–103
Zangenehpour S, Chaudhuri A (2002) Differential induction anddecay curves of c-fos and zif268 revealed through dual activitymaps. Mol Brain Res 109:221–225
321





























