Embed Size (px)
Citation preview

VITAMINS AND HORMONES, VOL. 61
The Biosynthesis of Coenzyme A in Bacteria
TADHG P. BEGLEY, CYNTHIA KINSLAND, AND CRICK STRAUSS
Department of Chemistry and Chemical Biology, Cornell University, Ithaca, New York
I. Introduction H. The Coenzyme A Biosynthetic Pathway in Bacteria
A. Aspartate 1-Decarboxylase B. Ketopantoate Hydroxymethyltransferase C. Ketopantoate Reductase D. Pantothenate Synthase E. Pantothenate Kinase F. 4'-Phosphopantothenoylcysteine Synthase G. 4'-Phosphopautothenoylcysteine Decarboxylase H. 4'-Phosphopantetheine Adenylyltransferase I. Dephospho-Coenzyme A Kinase J. 4'-Phosphopantetheinyltransferase
III. Regulation of Coenzyme A Biosynthesis in Escherichia coli IV. Coenzyme A Biosynthesis in Saccharomyces cerevisiae V. Coenzyme A in Human Nutrition
VI. Summary and Conclusions References
Coenzyme A (I) and enzyme-bound phosphopantetheine (II) func- tion as acyl carriers and as carbonyl activating groups for Claisen reactions as well as for amide-, ester-, and thioester-forming reac- tions in the cell. In so doing, these cofactors play a key role in the biosynthesis and breakdown of fatty acids and in the biosynthesis of polyketides and nonribosomal peptides. Coenzyme A is biosyn- thesized in bacteria in nine steps. The biosynthesis begins with the decarboxylation of aspartate to give ~-alanine. Pantoic acid is formed by the hydroxymethylation of ~-ketoisovalerate followed by reduction. These intermediates are then condensed to give pan- tothenic acid. Phosphorylation of pantothenic acid followed by condensation with cysteine and decarboxylation gives 4'-phospho- pantetheine. Adenylation and phosphorylation of 4'-phosphopan- tetheine completes the biosynthesis ofcoenzyme A. This review will focus on the mechanistic enzymology of coenzyme A biosynthesis in bacteria. © 2001 Academic Press.
157 Copyright © 2001 by Academic Press. All rights of reproduction in any form reserved.
0083-6729/01 $35.00

158 TADHG P. BEGLEY et al.
o SH
.b dP I
II
FIG. 1. The structure of eoenzyme A (I) and enzyme-bound phosphopantetheine (II).
I. INTRODUCTION
Coenzyme A (I) and enzyme-bound phosphopantetheine (II) (Fig. 1) function as acyl carriers and as carbonyl activating groups for Claisen reactions, as well as for amide-, ester-, and thioester-forming reactions in the cell. In so doing, these cofactors play a key role in the biosynthe- sis and breakdown of fatty acids and in the biosynthesis of nonriboso- mal peptides and polyketides. It has been estimated that 4% of the known enzymes use coenzyme A as a cofactor (Lee and Chen, 1982). This review will focus on the mechanistic enzymology of coenzyme A biosynthesis in bacteria.
I I . THE COENZYME A BIOSYNTHETIC PATHWAY IN BACTERIA
Bacteria synthesize coenzyme A from aspartate, ~-ketovalerate, cys- teine, and ATP. The biosynthesis (Fig. 2) begins with the decarboxyla- tion of aspartate (III) to give ~-alanine (IV). Pantoic acid (VII) is formed by the hydroxymethylation of a-ketoisovalerate (V) followed by reduc- tion. These intermediates are then condensed to give pantothenic acid (VIII). Phosphorylation of pantothenic acid followed by condensation with cysteine and decarboxylation gives 4'-phosphopantetheine (XI). Adenylation and phosphorylation of 4'-phosphopantetheine completes the biosynthesis of coenzyme A. The acyl carrier protein is phospho- pantetheinylated by the displacement of the AMP moiety of coenzyme A by an active site serine.
The biosynthesis of coenzyme A requires nine enzymes: aspartate l-decarboxylase, ketopantoate hydroxymethyltransferase, ketopantoate

BIOSYNTHESIS OF COENZYME A IN BACTERIA 159
Aspartate l-decarboxylase H N,~ ~COOH (PanD) -
2 H~,/COOH ,. H 2 N ~ C O O Aspartic acid ~3-Alanine
III IV Ketopantoate
reductase ~ . COOH /],~,.COO-hydroxymethyltransferase ( /~COO-Ketopantoa te
" ~ (PanB) (ApbA or PanE)" HO 0 NADPH H~) H" ~)H
ct-Ketoisovalerate c~-Ketopantoate Pantoic acid V Vl VII
Pantothenate ~ Pantothenatc O synthetase kinase "X~ ~. ~/L2OOH (PanC) H~ H4~'oHN/~"~OOH (CoaA) " ( H ~ I H N
PO ATP + IV Pantothenie acid 4'-Phosphopantothenie acid
VIII IX
4'-Phosphopantothenoyl- 4'-Phosphopantothenoylcysteine cysteine synthetase decarboxylase
(CoaB) (CoaC) , , .o. 9 H gOOH
Cysteine + CTP r /~I I" . - N "%"J~Y ~Q'/S H P6 ffb." H
4'-Phosphopantothenoylcysteine X
O 4'-Phosphopantetheine adenylyltransferase N / , . . ~ N ~ S H (CoaD)
-I H~bH H H ATP t'u 4'-Phosphopantetheine
XI
N
(~p.O O ] ' ' [ (CoaE) . _
"O"x~/ H6 (~H ATP ,~;\ , O O
H ~3H Coenzyme A
Dephospho Coenzyme A I XII
Phosphopantetheinyl transferase ~ S H ACP-Se
Acyl carrier protein-Ser-OH I1
FIG. 2. The coenzyme A biosynthetic pathway in bacteria.

160 TADHG P. BEGLEY et al.
reductase, pantothenate synthetase, pantothenate kinase, 4'-phospho- pantothenoylcysteine synthetase, 4'-phosphopantothenoylcysteine de- carboxylase, 4'-phosphopantetheine adenylyltransferase, and dephos- pho-coenzyme A kinase. The following sections review the progress made on the mechanistic enzymology of these enzymes during the 1990s (Jackowski, 1996).
A. ASPARTATE 1-DECARBOXYLASE
Aspartate 1-decarboxylase has been overexpressed from Escherichia coli (PanD, tetramer of 13.8-kDa subunits, K m = 151 txM, kca t = 0.6 s - 1 ) (Ramjee et al., 1997). This enzyme contains a pyruvamide cofactor that is formed by an autocatalytic cleavage between glycine 24 and serine 25 (Fig. 3). This cleavage reaction occurs by an N-O acyl shift to give XV followed by elimination and enamine hydrolysis. The autoprocess- ing reaction is quite slow (half-life of 16 h at 50°C), and the overex- pressed enzyme after purification consists primarily of the unprocessed protein (90%). This raises the interesting possibility that an additional protein may be involved in the in vivo formation of the pyruvoyl cofac- tor because the time spent by E. coli between divisions (20-60 min) is much shorter than the in vitro cofactor processing time (Ramjee et al., 1997).
The crystal structure of aspartate 1-decarboxylase has been solved at 2.2 A resolution (Albert et al., 1998). In this structure, only three of the subunits contain the pyruvoyl cofactor, and the fourth subunit has been trapped as the ester intermediate (XV). The structure suggests that the
Gly 24 H H IOI O O N ~ N ~ LI'~ NHCyS26 n ~+r ~ " ~ NHCys26 H3~'~ ~U'~ NHCy s26 %-x,,, ) o1,%
O O
H 2 N ' ~ " NHCys26 ~ _ 0 ~ NHCys26
G I Y % o H [ [ XVII + NH3 I I O
XVl
FIG. 3. Mechanism for the formation of the pyruvoyl cofactor.

BIOSYNTHESIS OF COENZYME A IN BACTERIA 161
A subunit serine 25 is deprotonated by a proton relay system consist- ing oftyrosine 58 from the A subunit and lysine 9 and histidine 11 from the D subunit. The collapse of the resulting tetrahedral intermediate XIV is facilitated by the transfer of this proton from the relay to the amine via tyrosine 58. Either tyrosine 58 or threonine 57 may also func- tion as the base catalyzing the elimination reaction (XV to XVI). In ad- dition, the structure also suggests that the N - O acyl shift (XIII to XV) and the elimination reaction (XV to XVI) are driven by relief of strain in a short peptide loop connecting [3 sheet 1' and [3 sheet 2 and by pro- tonation of the amine XIV. The processing of the fourth subunit does not occur because of an unfavorable conformation for the elimination in this subunit and because there is no base close to the C proton of ser- ine 25 to carry out the deprotonation reaction essential for the conver- sion of XV to XVI.
A mechanism for the decarboxylation reaction, based on mechanistic studies carried out on histidine decarboxylase, is outlined in Fig. 4 (Van Poelje and Snell, 1990). Formation of a protonated imine (XVIII) be- tween aspar ta te and the pyruvoyl cofactor activates aspar ta te for decarboxylation because the resulting anion (XIX) is stabilized by an electrostatic interaction with the positively charged nitrogen and by de- localization (Bach and Canepa, 1997). Protonation of this anion fol- lowed by hydrolysis of the Schiffbase completes the reaction. The Schiff base of aspar ta te has been modeled into the active site of one of the fully processed subunits. This model suggests that aspar ta te is bound via a salt bridge to arginine A54 and that a water molecule hydrogen bonded to lysine A9 protonates the carbanion intermediate. Other in- teractions involved in the formation and hydrolysis of the Schiff base and[ in the decarboxylation have not yet been identified.
0 H
III / 0 XVII
0 O-
XVIII 0
O
H O O C ~ N H 2 , ~ H'Enz IV ~ 0 XVlI /
+ CO2
noocv-~n ~ H + CO 2
~" N. En z
XIX 0
FIG. 4. The mechanism of aspartate 1-decarboxylase.
162 TADHG R BEGLEYet al.
.R
H2 H
XX
-OOC~O _
v
H'J~H R
H XXIV
2 H2 H
XXI! XXIII
H ~H -00 H N +
~H2N H ~ O H XXI VI
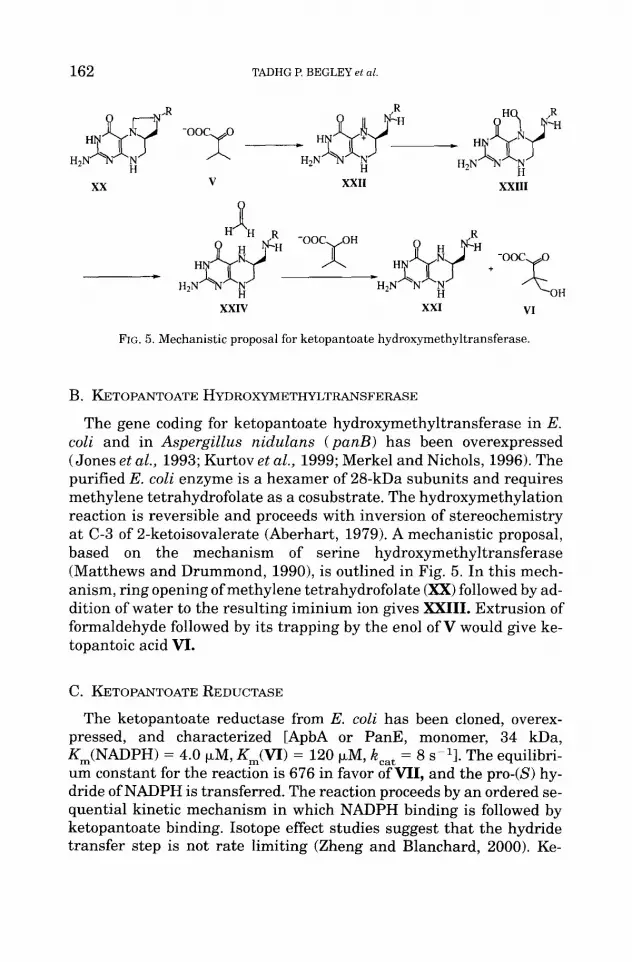
FIG. 5. Mechanistic proposal for ketopantoate hydroxymethyltransferase.
B. KETOPANTOATE HYDROXYMETHYLTRANSFERASE
The gene coding for ketopantoate hydroxymethyltransferase in E. coli and in Aspergillus nidulans (panB) has been overexpressed (Jones et al., 1993; Kurtov et al., 1999; Merkel and Nichols, 1996). The purified E. coli enzyme is a hexamer of 28-kDa subunits and requires methylene tetrahydrofolate as a cosubstrate. The hydroxymethylation reaction is reversible and proceeds with inversion of stereochemistry at C-3 of 2-ketoisovalerate (Aberhart, 1979). A mechanistic proposal, based on the mechanism of serine hydroxymethyltransferase (Matthews and Drummond, 1990), is outlined in Fig. 5. In this mech- anism, ring opening of methylene tetrahydrofolate (XX) followed by ad- dition of water to the resulting iminium ion gives XXIII. Extrusion of formaldehyde followed by its trapping by the enol of V would give ke- topantoic acid VI.
C. KETOPANTOATE REDUCTASE
The ketopantoate reductase from E. coli has been cloned, overex- pressed, and characterized [ApbA or PanE, monomer, 34 kDa, Km(NADPH) = 4.0 ~M, Km(VI) = 120 ~M, kca t = 8 s - l ] . The equilibri- um constant for the reaction is 676 in favor of VII, and the pro-(S) hy- dride of NADPH is transferred. The reaction proceeds by an ordered se- quential kinetic mechanism in which NADPH binding is followed by ketopantoate binding. Isotope effect studies suggest that the hydride transfer step is not rate limiting (Zheng and Blanchard, 2000). Ke-

BIOSYNTHESIS OF COENZYME A IN BACTERIA 163
topantoic acid reductases from Salmonella typhimurium (Frodyma and Downs, 1998) and from Pseudomonas maltophilia (Shimizu et el., 1988) have also been characterized.
D. PANTOTHENATE SYNTHASE
Although the gene for pantothenate synthase (panC) has been iden- tiffed, it has not been overexpressed from any bacterial source. The E. coli enzyme has been purified 500-fold and is a homotetramer of 18-kDa subunits. The K m values for ~-alanine, ATP, and pantoic acid are 150, 100, and 63 txM, respectively. The enzyme follows a Ping-Pong Bi Uni Bi type mechanism (Miyatake et el., 1979).
Pantothenate is the most advanced coenzyme A precursor that is tak- en up by E. coli. The transport protein, pantothenate permease (PanF), has been cloned. Uptake occurs by a sodium ion-dependent cotransport mechanism (Jackowski and Alix, 1990).
The pantothenate synthetase genes from Lotus japonicus and from Oryza sat ivum (rice) have also been cloned (Genschel et el., 1999). The Lotusjaponicus enzyme has been overexpressed and characterized [ho- modimer of 34-kDa subunits, KIn(VII) = 44 ~M, Kin(IV) = 44 ~M, hea t = 0.62 s-l) .
E. PANTOTHENATE KINASE
Pantothenate kinase has been cloned from E. coli and characterized [CoaA homodimer, Km(ATP) = 136 txM, KIn(VIII) = 36 IxM, k c a t = 17 rain 1) (Song and Jackowski, 1992). The enzyme is expressed as two closely related translation products (35.4 and 36.4 kDa). The signifi- cance of this observation is unknown. The phosphorylation reaction proceeds by an ordered sequential mechanism with ATP binding first. The enzyme is inhibited by coenzyme A and its esters, shows highly co- operative ATP binding, and is a key regulatory enzyme on the coenzyme A biosynthesis pathway (Song and Jackowski, 1994). Pantothenate ki- nase has been overexpressed and characterized from AspergiUus nidu- lens (Calder et el., 1999), and the enzyme has also been purified from wild-type Brevibacterium ammoniagenes (Shimizu et el., 1973).
F. 4%PHosPHOPANTOTHENOYLCYSTEINE SYNTHASE
4'-Phosphopantothenoylcysteine synthase activity has been de- tected in a cell-free system, but the enzyme has not been purified from

164 TADHG P. BEGLEY et al.
any bacterial source (Brown, 1959). The gene coding for this enzyme has not been identified.
G. 4 ' -PHosPHOPANTOTHENOYLCYSTEINE DECARBOXYLASE
4'-Phosphopantothenoylcysteine decarboxylase has been purified from E. coli (homotetramer of 35-kDa subunits, K m = 0.9 mM, k c a t =
21 min -1 assuming one active site per subunit) (Yang and Abeles, 1987). This enzyme utilizes a pyruvoyl group and is mechanistically in- teresting because all other pyruvoyl-dependent enzymes require a free amino group a to the carboxylate in order to catalyze the decarboxyla- tion (see aspartate 1-decarboxylase, Fig. 4). No mechanistic studies have been carried out on this enzyme other than the demonstration that the decarboxylation catalyzed by the rat liver enzyme occurs with re- tention of stereochemistry (Aberhart et al., 1985). The gene has not yet been identified in any microorganism.
A mechanistic proposal for this interesting decarboxylase is outlined in Fig. 6. In this mechanism an N-S acyl shift unmasks the amino group to give XXVI. The free amine could then participate in a pyru- voyl-assisted decarboxylation as previously described in Fig. 4 for as- partate decarboxylase. Reversal of the N-S acyl shift would complete the reaction. Enzyme-catalyzed acyl shift reactions have been previ- ously identified in the intein processing reaction (Paulus, 1998; Perler, 1998; Perler et al., 1997; Shao and Paulus, 1997), in the formation of
0 OH
y "-00- N - - ' ( • y "OH2N % 0 N
( R _/S | S ENZYME ENZYME ] ] ENZYME R ~
XXV X XXV 0 XXV] XXV[I 0
| R ~S / HS ENZYME - '~ ENZYME " ~ ENZYME
0 0 XXVIII XXV XXIX XXV XI
FIG. 6. Proposed mechanism for 4'-phosphopantothenoylcysteine decarboxylase (R = 4'-phosphopantothenoyl).

BIOSYNTHESIS OF COENZYME A IN BACTERIA 165
the pyruvoyl cofactor (Van Poelje and Snell, 1990), and in the serine and cysteine proteases.
H. 4'-PHosPHOPANTETHEINE ADENYLYLTRANSFERASE
Tihe 4'-phosphopantetheine adenylyltransferase gene (kdtB or coaD) has been overexpressed and purified from E. coli (Geerlofet al., 1999). It is a hexamer of 17.8-kDa subunits. The adenylation reaction is re- versible [Km(XI) = 7 ~M, Km(PP i) = 0.2 raM, kca t =- 3.3 s-l]. The crys- tal structure of this enzyme complexed with dephospho-coenzyme A has been described (Izard and Geerlof, 1999).
I. DEPHOSPHO-COENZYME A KINASE
The E. coli dephospho-coenzyme A kinase gene has recently been identified, and the enzyme has been overexpressed and characterized (D. Drueckhammer, unpublished results, 2000).
4'-Phosphopantetheine adenylyltransferase and dephospho-coen- zyrae A kinase have been used for the enzymatic synthesis of coenzyme A analogs (Bibart et al., 1999; Martin et al., 1994). These analogs have been elegantly used as mechanistic probes of coenzyme A-utilizing en- zymes (Gu et al., 1999; Kurz et al., 1997; Schwartz and Drueckhammer, 1996; Schwartz et al., 1995; Usher et al., 1994).
J. 4'-PHosPHOPANTETHEINYLTRANSFERASE
The phosphopantetheinyl moiety of the acyl carrier protein is formed by displacement of AMP from coenzyme Aby an active site ser- ine in a reaction catalyzed by a phosphopantetheinyltransferase. The enzyme has been overexpressed and characterized from several sources. These include the acyl carrier protein synthase from E. coli involved in fatty acid biosynthesis (Flugel et al., 2000), Lys5 from Sac- charomyces cerevisiae involved in lysine biosynthesis (Ehmann et al., 1999), EntD from E. coli involved in enterobactin biosynthesis (Gehring et al., 1998), and Spf from B. subt i l is involved in surfactin biosynthesis (Quadri et al., 1998). The Spf phosphopantetheinyl- transferase shows relaxed substrate specificity and has been used to transfer the phosphopantetheinyl moiety to apo-6-methylsalicylic acid synthase in S. cerevisiae and in E. coli, giving rise to active syn- thase and the overproduction of 6-methylsalicylic acid (Carreras et al., 1997; Kealey et al., 1998).

166 TADHG P. BEGLEY et al.
III. REGULATION OF COENZYME A BIOSYNTHESIS IN Escherichia coli
Pantothenic acid is not a rate-limiting intermediate because E. coli produces 15 times more pantothenate than is required for coenzyme A biosynthesis. Furthermore, the overexpression of pantothenate perme- ase in the presence ofpantothenate does not result in an increase in the concentration of coenzyme A in the cell (Jackowski and Rock, 1981; Jackowski and Alix, 1990).
Pantothenate kinase is a key control point on the coenzyme A biosyn- thesis pathway. It is subject to feedback inhibition by coenzyme A and to a lesser extent by coenzyme A thioesters (Vallari et al., 1987; Song and Jackowski, 1994). As a consequence, 76-fold overexpression of pan- tothenate kinase resulted in only a 2.7-fold increase in the concentra- tion of cellular coenzyme A (Song and Jackowski, 1992). The concen- tration of coenzyme A in the cell is also influenced by coenzyme A degradation (Vallari and Jackowski, 1988). This degradation reaction is not catalyzed by phosphopantetheine adenylyltransferase (Geerlofet al., 1999), and the enzyme involved has not yet been identified.
IV. COENZYME A BIOSYNTHESIS IN Saccharomyces cerevisiae
In Saccharomyces cerevisiae pantothenate is t ransported into the cell by the Fen2p transporter (Stolz and Sauer, 1999) and converted to coen- zyme A by a 375- to 400-kDa multienzyme complex (Bucovaz et al., 1997). The biosynthetic pathway (Fig. 7) uses essentially the same chemistry as that found in the bacterial pathway although the steps oc- cur in a different order. Pantothenate (VIII) is first adenylylated and then condensed with cysteine to give XXXI. Decarboxylation followed by a final phosphorylation completes the biosynthesis. Our under- standing of the mechanistic enzymology of coenzyme A biosynthesis in yeast is still at an early stage.
V. COENZYME A IN HUMAN NUTRITION
Pantothenic acid (vitamin B 5) is the most advanced coenzyme A pre- cursor taken up by cells. It is not biosynthesized by humans and is, therefore, an essential vitamin. Annual production of pantothenic acid is 5000 tons, and the average daily intake, primarily in the form ofcoen- zyme A and coenzyme A esters, is 5 -10 mg per person. A recommended

VIII
BIOSYNTHESIS OF COENZYME A IN BACTERIA
~.O_P/=O HO 0tl
167
N
-o-pL = o o
XXXI
O•P 0 HO OH
XI1 H ~'OH 1
F I a . 7. C o e n z y m e A b i o s y n t h e s i s i n Saccharomyces cerevisiae.
dietary allowance has not been established because this vitamin is widely distributed in plants and animals and because spontaneous pan- tothenic acid deficiency in humans has not been recorded. Dietary coen- zyme A is hydrolyzed in the lumen of the intestine to pantothenic acid, which is then taken up and reconverted to coenzyme A (Brody, 1998; Tahiliani and Beinlich, 1991). Our current understanding of the mech- anistic enzymology of coenzyme A biosynthesis in mammals is still at an early stage.
VI. SUMMARY AND CONCLUSIONS
Bacteria biosynthesize coenzyme A from aspartate, ~-ketovalerate, cysteine, and ATP. The biosynthesis begins with the decarboxylation of aspartate (III) to give ~-alanine (IV) and the formation ofpantoic acid (VII) by the hydroxymethylation of ~-ketoisovalerate (V) followed by reduction. These intermediates are then condensed to give pantothenic acid (VIII). Phosphorylation of pantothenic acid followed by conden- sation with cysteine and decarboxylation gives phosphopantetheine (XI). Adenylation and phosphorylation of this complete the biosynthe- sis of coenzyme A. Overall, the coenzyme A biosynthesis pathway in-

168 TADHG P. BEGLEY et al.
volves well-understood chemistry except for the decarboxylation of phosphopantothenoylcysteine. This reaction is a novel pyruvoyl- dependent decarboxylation reaction in which the substrate does not have a free amino group. All of the biosynthetic genes except for the phosphopantothenoylcysteine synthetase gene and the phosphopan- tothenoylcysteine decarboxylase gene have been identified. Structural studies on the pathway are at an early stage, and only two structures have been solved (aspartate decarboxylase and 4'-phosphopantetheine adenylyltransferase). The mechanistic enzymology of coenzyme A biosynthesis in eukaryotes is also at an early stage.
REFERENCES
Aberhart, D. J. (1979). Stereochemistry of pantoate biosynthesis from 2-ketoisovalerate. J. Am. Chem. Soc. 101, 1354-1355.
Aberhart, D. J., Ghoshal, P. K., Cotting, J.-A., and Russell, D. J. (1985). Coenzyme A biosynthesis: Steric course of4'-phosphopantothenoyl-L-cysteine decarboxylase. Bio- chemistry 24, 7178-7182.
Albert, A., Dhanaraj, V., Genschel, U., Khan, G., Ramjee, M. K., Pulido, R., Sibanda, B. L., von Delft, F., Witty, M., Blundell, T. L., Smith, A. G., and Abell, C. (1998). Crystal structure of aspartate decarboxylase at 2.2 A resolution provides evidence for an es- ter in protein self-processing. Nat. Struct. Biol. 5, 289-293.
Bach, R. D., and Canepa, C. (1997). Theoretical model for pyruvoyl-dependent enzymic decarboxylation of a-amino acids. J. Am. Chem. Soc. 119, 11725-11733.
Bibart, R. T., Vogel, K. W., and Drueckhammer, D. G. (1999). Development of a second generation coenzyme A analog synthon. J. Org. Chem. 64, 2903-2909.
Brody, T. (1998). "Nutritional Biochemistry," 2nd ed. Academic Press, San Diego, CA. Brown, G. M. (1959). The metabolism of pantothenic acid. J. Biol. Chem. 234, 370-378. Bucovaz, E. T., Macleod, R. M., Morrison, J. C., and Whybrew, W. D. (1997). The coen-
zyme A synthesizing protein complex and its proposed role in CoA biosynthesis in bakers' yeast. Biochimie 79, 787-798.
Calder, R. B., Williams, R. S., Ramaswamy, G., Rock, C. O., Campbell, E., Unkles, S. E., Kinghorn, J. R., and Jackowski, S. (1999). Cloning and characterization of a eu- karyotic pantothenate kinase gene (panK) from Aspergillus nidulans. J. Biol. Chem. 274, 2014-2020.
Carreras, C. W., Gehring, A. M., Walsh, C. T., and Khosla, C. (1997). Utilization of en- zymically phosphopantetheinylated acyl carrier proteins and acetyl-acyl carrier pro- teins by the actinorhodin polyketide synthase. Biochemistry 36, 11757-11761.
Ehmann, D. E., Gehring, A. M., and Walsh, C. T. (1999). Lysine biosynthesis in Saccha- romyces cerevisiae: Mechanism of ~-aminoadipate reductase (Lys2) involves post- translational phosphopantetheinylation by Lys5. Biochemistry 38, 6171-6177.
Flugel, R. S., Hwangbo, Y., Lambalot, R. H., Cronan, J. E., and Walsh, C. T. (2000). Holo- (acyl carrier protein) synthase and phosphopantetheinyl transfer in Escherichia coli. J. Biol. Chem. 275, 959-968.
Frodyma, M. E., and Downs, D. (1998). ApbA, the ketopantoate reductase enzyme of Sal- monella typhimurium, is required for the synthesis of thiamine via the alternative pyrimidine biosynthetic pathway. J. Biol. Chem. 273, 5572-5576.

BIOSYNTHESIS OF COENZYME A IN BACTERIA 169
Geerlof, A., Lewendon, A., and Shaw, W. V. (1999). Purification and characterization of phosphopantetheine adenylyltransferase from Escherichia coll. J. Biol. Chem. 274, 2'7105-27111.
Gehring, A. M., Mori, I., and Walsh, C. T. (1998). Reconstitution and characterization of tile Escherichia coli enterobactin synthetase from EntB, EntE, and EntF. Biochem- istry 37, 2648-2659.
Genschel, U., Powell, C. A., Abell, C., and Smith, A. G. (1999). The final step of pan- tothenate biosynthesis in higher plants: Cloning and characterization of pantothen- ate synthetase from Lotusjaponicus and Oryza sativum (rice). Biochem. J. 341,669- 678.
Gu, Z, Drueckhammer, D. G., Kurz, L., Liu, K., Martin, D. P., and McDermott, A. (1999). Solid state NMR studies of hydrogen bonding in a citrate synthase inhibitor complex. Biochemistry 38, 8022-8031.
Izard: T., and Geerlof, A. (1999). The crystal structure of a novel bacterial adenylyl- transferase reveals half of sites reactivity. EMBO J. 18, 2021-2030.
Jackowski, S. (1996). Biosynthesis of pantothenic acid and coenzyme A. In "Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology" (F. C. Neidhardt et al., eds.), 2nd ed., pp. 687-694. American Society for Microbiology, Washington, DC.
Jackowski, S., and Alix, J. H. (1990). Cloning, sequence and expression of the pan- tothenate permease (PanF) gene of Escherichia coli. J. Bacteriol. 172, 3842-3848.
Jackowski, S., and Rock, C. O. (1981). Regulation of coenzyme A biosynthesis. J. Bacte- rliol. 148, 926-932.
Jones;, C. E., Brook, J. M., Buck, D., Abell, C., and Smith, A. G. (1993). Cloning and se- quencing of the Escherichia coli panB gene, which encodes ketopantoate hydroxy- methyltransferase, and overexpression of the enzyme. J. Bacteriol. 175, 2125- 2130.
Kealey, J. T., Liu, L., Santi, D. V., Betlach, M. C., and Barr, P. J. (1998). Production of a polyketide natural product in nonpolyketide-producing prokaryotic and eukaryotic hosts. Proc. Natl. Acad. Sci. U.S.A. 95, 505-509.
Kurtov, D., Kinghorn, J. R., and Unkles, S. E. (1999). The Aspergillus nidulans panB gene encodes ketopantoate hydroxymethyltransferase, required for biosynthesis of pan- tothenate and coenzyme A. Mol. Gen. Genet. 262, 115-120.
Kurz, L. C., Roble, J. H., Nakra, T., Drysdale, G. R., Buzan, J. M., Schwartz, B., and Drueckhammer, D. G. (1997). Ability of single-site mutants of citrate syuthase to cat- alyze proton transfer from the methyl group of dethiaacetyl-coenzyme A, a non- thioester substrate analog. Biochemistry 36, 3981-3990.
Lee, C. H., and Chen, A. F. (1982). Immobilized coenzymes and derivatives. In "The Pyri- dine Nucleotide Coenzymes" (J. Everse, B. Anderson, and K. You, eds.), pp. 189. Aca- demic Press, New York.
Martin, D. P., Bibart, R. T., and Drueckhammer, D. G. (1994). Synthesis of novel analogs of acetyl coenzyme A: Mimics of enzyme reaction intermediates. J. Am. Chem. Soc. 116, 4660-4668.
Matthews, R. G., and Drummond, J. T. (1990). Providing one-carbon units for biological methylations: Mechanistic studies on serine hydroxymethyltransferase, methyl- enetetrahydrofolate reductase, and methyltetrahydrofolate-homocysteine methyl- transferase. Chem. Rev. 90, 1275-1290.
Merkel, W. K., and Nichols, B. P. (1996). Characterization and sequence of the Escherichia coli panBCD gene cluster. FEMB Microbiol. Lett. 143, 247-252.

170 TADHG P. BEGLEY et al.
Miyatake, K., Nakano, Y., and Kitaoka, S. (1979). Pantothenate synthetase from Es- cherichia coli. In "Methods in Enzymology" (D. B. McCormick and L. D. Wright, eds.), Vol. 62, Part D, pp. 215-219. Academic Press, New York.
Paulus, H. (1998). The chemical basis of protein splicing. Chem. Soc. Rev. 27, 375-386. Perler, F. B. (1998). Breaking up is easy with esters. Nat. Struct. Biol. 5, 249-252. Perler, F. B., Xu, M.-Q., and Paulus, H. (1997). Protein splicing and autoproteolysis mech-
anisms. Curr. Opin. Chem. Biol. 1, 292-299. Quadri, L. E. N., Weinreb, P. H., Lei, M., Nakano, M. M., Zuber, P., and Walsh, C. T.
(1998). Characterization of Sfp, a Bacillus subtilis phosphopantetheinyl transferase for peptidyl carrier protein domains in peptide synthetases. Biochemistry 37, 1585- 1595.
Ramjee, M. K., Genschel, U., Abell, C., and Smith, A. G. (1997). Escherichia coli L-aspar- tate-(~-decarboxylase: Preprotein processing and observation of reaction intermedi- ates by electrospray mass spectrometry. Biochem. J. 323, 661-669.
Schwartz, B., and Drueckhammer, D. G. (1996), A stereochemical probe of the tetrahe- dral intermediate in the reactions of acetyl-coenzyme A dependent acetyltransferas- es. J. Am. Chem. Soc. 118, 9826-9830.
Schwartz, B., Drueckhammer, D. G., Usher, K. C., and Remington, S. J. (1995). ~-Fluoro acid and ~-fluoro amide analogs of acetyl-CoA as inhibitors of citrate synthase: Ef- fect o fpK matching on binding affinity and hydrogen bond length. Biochemistry 34, 15459-15466.
Shao, Y., and Paulus, H. (1997). Protein splicing: Estimation of the rate of O-N and S- N acyl rearrangements, the last step of the splicing process. J. Pept. Res. 50, 193- 198.
Shimizu, S., Kubo, K., Tani, Y., and Ogata, K. (1973). Metabolism of pantothenic acid in microorganisms. VII. Purification and properties of pantothenate kinase from Bre- vibacterium ammoniagenes IFO 12071. Agric. Biol. Chem. 37, 2863-2870.
Shimizu, S., Kataoka, M., Chung, M., and Yamada, H. (1988). Ketopantoic acid reductase of Pseudomonas maltophilia 845. Purification, characterization, and role in pan- tothenate biosynthesis. J. Biol. Chem. 263, 12077-12084.
Song, W.-J., and Jackowski, S. (1992). Cloning, sequencing, and expression of the pan- tothenate kinase (coaA ) gene of Escherichia coli. J. Bacteriol. 174, 6411-6417.
Song, W.-J., and Jackowski, S. (1994). Kinetics and regulation of pantothenate kinase from Escherichia coli. J. Biol. Chem, 269, 27051-27058.
Stolz, J., and Sauer, N. (1999). The fenpropimorph resistance gene FEN2 from Saccha- romyces cerevisiae encodes a plasma membrane H÷-pantothenate symporter. J. Biol. Chem. 274, 18747-18752.
Tahiliani, A. G., and Beinlich, C. J. (1991). Pantothenic acid in health and disease. Vitam. Horm. (N.Y,)46, 165-228.
Usher, K. C., Remington, S. J., Martin, D. P., and Drueckhammer, D. G. (1994). A very short hydrogen bond provides only moderate stabilization of an enzyme-inhibitor complex of citrate synthase. Biochemistry 33, 7753-7759.
Vallari, D. S., and Jackowski, S. (1988). Biosynthesis and degradation both contribute to the regulation of coenzyme A content in Escherichia coli. J. Bacteriol. 170, 3961.
Vallari, D. S., Jackowski, S., and Rock, C. O. (1987). Regulation of pantothenate kinase by coenzyme A and its thioesters. J. Biol. Chem. 262, 2468-2471.
Van Poelje, P. D., and Snell, E. E. (1990). Pyruvoyl-dependent enzymes. Annu, Rev. Biochem. 59, 29-59.
Yang, H., and Abeles, R. H. (1987). Purification and properties ofEscherichia coli 4'-phos-

BIOSYNTHESIS OF COENZYME A IN BACTERIA 171
phopantothenoylcysteine decarboxylase: Presence of covalently bound pyruvate. Bio- chemistry 26, 4076-4081.
Zheng, R., and Blanchard, J. S. (2000). Kinetic and mechanistic analysis of the E. coli pan E-.encoded ketopantoate reductase. Biochemistry 39, 3708-3717.





























