Embed Size (px)
Citation preview

Plant Molecular Biology 26: 1775-1783, 1994. © 1994 Kluwer Academic Publishers. Printed in Belgium. 1775
The C 3 plant Flaveria pringlei contains a plastidic NADP-malic enzyme which is orthologous to the C 4 isoform of the C 4 plant F. trinervia
B~bel Lipka, Klaus Steinm/aller, Elke Rosche, Dagmar Btrsch and Peter Westhoff* Institut fiir Entwicklungs- und Molekularbiologie der Pflanzen, Heinrich-Heine-Universitiit, D-40225 Diisseldorf, Germany (* author for correspondence)
Received 11 March 1994; accepted in revised form 7 September 1994
Key words: C 4 photosynthesis, molecular evolution, nadp-malic enzyme, flaveria
Abstract
To study the molecular evolution of NADP-dependent malic enzyme (NADP-ME) in the genus Flaveria a leaf-specific cDNA library of the C3 plant F. pringlei was screened for the presence of sequences ho- mologous to the C4 isoform gene (named modA) of the C4 plant F. trinervia. The cDNAs isolated contained varying numbers of identical restriction fragments suggesting that they were derived from a single gene. This was supported by Southern hybridisation experiments with genomic DNA from F. trinervia and F. pringlei. Nucleotide sequence analysis of a full-size clone identified the presence of a typical plastidic transit peptide and revealed that the mature modA proteins of F. trinervia (C4) and F. pringlei (C3) are 90~o similar. These findings indicate that C3 plants, like C4 species, possess a plastidic isoform of NADP-ME and that the modA genes of the two species represent orthologous genes. Northern analyses showed that modA transcripts accumulate to similar levels in leaves, stems and roots of F. pringlei. The expression of this gene in F. pringlei thus appears to be rather constitutive. In contrast, the modA gene of F. trinervia is abundantly expressed in leaves, but maintains its expression in stems and roots. It has to be concluded from these data that the leaf-specific increase in the expression level was a key step which was taken during the evolution of the C4 isoform modA gene starting from a C3 ancestral gene.
Introduction
NADP-dependent malic enzyme (NADP-ME; oxaloacetate-decarboxylating, EC 1.1.1.40) is an abundant protein in malate-transporting C4 plants and serves as a diagnostic criterion for the classification of C4 plants [ 10, 12]. The enzyme is located in bundle-sheath chloroplasts and ca-
talyses the oxidative decarboxylation of malate [13]. NADP-malic enzyme plays also a central role in crassulacean acid metabolism [35] and, moreover, NADP-ME activity has been detected in non-photosynthetic tissues of Ca and C4 plants. It has been proposed that this non-photosynthetic NADP-ME is involved in an alternative branch of the glycolytic pathway and that it functions in
The nucleotide sequence data reported will appear in the EMBL, GenBank and DDBJ Nucleotide Sequence Databases under the accession number X78069.

1776
pH regulation (reviewed in [6, 32]). However, the exact metabolic role of the non-photosynthetic isoform still needs to be clarified. There is also confusion in the literature about the intracellular location of the C 3 isoform of NADP-ME. It has been suggested, but never definitely proven, that the C3 isoforms of NADP-ME are cytosolic pro- teins (reviewed in [6, 32]). In accordance with this view, two NADP-ME cDNA sequences have been published whose encoded proteins lack a plastidic transit peptide [29, 31]. On the other hand, recent cell-fractionation experiments with photosynthetic as well as non-photosynthetic tis- sues of various C3 plants clearly indicate that NADP-ME activity should be confined to plas- rids [8].
The involvement of NADP-ME in different metabolic pathways and, hence, the existence of various isoforms raises the question of how they originated during evolution. This is partictilarly interesting with respect to the formation of the C4 isoform, since plants with the C4 pathway of pho- tosynthesis have evolved several times indepen- dently from C3 ancestral species during the evo- lution of angiosperms [21]. To analyse the evolutionary origin of the C4 isoform of NADP- ME, our laboratory is engaged in studying the molecular genetics of this enzyme in Flaveria. The genus Flaveria has been selected, because it con- tains C3 and C4 species and, in addition, a large number of C3-C4 intermediates suggesting that evolution towards C4 photosynthesis is still in progress within this genus [7]. Sequence analysis of a cDNA clone encoding the C4 isoform of NADP-ME of the C4 dicot F. trinervia revealed that the transit peptides of the Flaveria and maize enzymes lack any significant sequence similarity, whereas the mature proteins of the two species are about 75 70 identical at the amino acid level [2]. This finding suggested an independent origin of the two transit peptides and provided further evidence for the polyphyletic origin of C4 photo- synthesis.
To clarify the molecular evolution of the C4 isoform of NADP-ME (gene designation modA) in Flaveria we have searched for the presence of modA-homologous sequences in the C3 plant F.
pringlei by cDNA cloning and genomic Southern hybridisation. Sequence analysis of a full-size cDNA clone isolated from a leaf-specific cDNA library showed that the encoded protein contains a typical plastidic transit sequence and, hence, must be located in the chloroplast. Northern blot experiments revealed that the modA gene is not only expressed in leaves, but also in stems and roots. A basal expression in stems and roots was also observed for the C4 isoform modA gene ofF. trinervia. This indicates that the modA gene main- tained its ancient C3-1ike expression pattern dur- ing evolution of C4 photosynthesis, but in addi- tion acquired a new feature, i.e. a high level of expression in the leaves.
Materials and methods
Plant material
Seeds of F. trinervia (C4), F. brownii (C4-1ike), F. floridana (C3-C 4 intermediate), F. linearis (C3-C 4 intermediate), F. chloraefolia (C3-C4 intermediate) and F. pringlei. (C3) were obtained from H. B auwe (Institut ftir Genetik und Kulturpflanzenfors- chung, Gatersleben, Germany) or S. Holaday (Texas Tech University, Lubbock), respectively. Plants were grown in the greenhouse as described [14].
Construction and screening of the F. pringlei leaf- specific cDNA library
Poly(A) + RNA isolated from leaves ofF. pringlei was converted to double-stranded cDNA using the cDNA cloning kit from Stratagene Cloning Systems (San Diego, USA). The double-stranded cDNAs were size-fractionated on a short column (about 3 cm in length) of Sephacryl S-400 and fragments larger than 600 bp in size were ligated into Xho I/Eco RI-digested ZAP II. The DNA was packaged into phage using Gigapack II ex- tracts (Stratagene Cloning Systems) resulting in about 250 000 independent recombinant phage. About 180 000 phage of the amplified library were screened by plaque hybridisation essentially as

outlined before [25]. Hybridisation was carried out at 57 °C in the sodium phosphate/dodecyl sulphate medium of Church and Gilbert [ 3 ] using the radio-labelled insert of lcFtrma152 [2] as a probe. Washings were at the same temperature once each in 5 x , 2 x and l x SSC. Teninde- pendent phage were finally recovered and con- verted to double-stranded plasmids as described in the manual of the cloning kit.
RNA and DNA blot analysis
Total or poly(A + ) RNA was size-fractionated on 1.5~ agarose gels and blotted onto Biodyne A membranes (Fa. Pall, Dreieich, Germany) as de- scribed [33], The RNA filters were hybridized overnight at 64 °C in sodium phosphate/dodecyl sulphate medium [ 3 ]. Washings were at the same temperature once each in 5 x , 2 x , 1 x and 0.5 x SSC. The blots were fluorographed for varying times using Kodak XAR-4 films.
Hybridisation of Southern-blotted DNA was as described above for RNA. After hybridisation, the filters were washed five times for 15 min each in 0.1 x SSC, 0.15 ~o SDS at 62 °C and fluoro- graphed for 2 to 4 days.
Nucleotide and protein sequences
The nucleotide sequence of 1cFprma133 was de- termined on both strands by the dideoxy-chain termination method modified for double-stranded plasmid DNA [25]. Nucleic acid and protein alignments as well as the calculation of similarity scores were performed with CLUSTAL V [15] on Apple Macintosh computers. The parsimony analysis was conducted with PAUP version 3.0i [28] using the branch and bound option.
Results and discussion
Isolation and sequencing of a NADP-ME cDNA clone from a leaf-specific library of F. pringlei
A leaf-specific cDNA library of F. pringlei was screened with lcFtrma152 encoding the
1777
NADP-ME C4-isoform of F. trinervia [2]. Re- striction analyses and Southern hybridisations using lcFtrma152 as a probe showed that all of the ten independent clones isolated contained a cen- trally located 330 Eco RI/Xho I fragment (see Fig. 3), but varying sizes of 5'- and 3'-extended sequences. This suggested that all clones were derived from the same mRNA. Three clones car- ried inserts of about 2.2 kb indicating that they were of full size. One of them, lcFprma133, was selected for nucleotide sequencing.
The predicted protein
The sequence of 2213 bp obtained contains a long open reading frame of 1941 bp which can be translated into a polypeptide of 647 amino acid residues. The protein encoded is 71 kDa and hence of the same size as the F. trinervia NADP-ME [2]. Figure 1 illustrates that the amino terminal part of the F. pringlei NADP-ME is rich in hydroxylated amino acids (15 out of 61) and has a positive net charge. It shows the typi- cal features of a plastidic transit peptide function- ing in targeting a protein into the stroma [30]. The F. pringlei and F. trinervia transit peptides are about 70 ~o similar. Remarkably, the positions of twelve serine/threonine residues are conserved as is the region surrounding the putative cleavage site (see Fig. 1). We infer from these finding that the NADP-ME isoform encoded by lcFprma133 is a plastidic protein like its C4 isoform counter- part of F. trinervia [2].
The two mature proteins possess 90~o identi- cal amino acid residues. A multiple sequence alignment of the F. pringlei protein with all known plant NADP-ME sequences allowed identifica- tion of blocks of similarities and hence putative functional significance. A strong sequence con- servation is found in three regions (I to III in Fig. 1) which are indicative of dinucleotide bind- ing folds of NAD- (box II) or NADP-linked oxi- doreductases (boxes I and III)[ 11, 24, 26, 34]. In addition, five cysteine residues (marked C in Fig. 1) are conserved in all plant NADP-MEs. Two of them are adjacent to putative dinucleotide

1778
F.p. MMSLNS S SVVK S S I SGVSWTQ S Q SVRL SVRRPMVVAMI2NSNGRPERSVGVSVDGAVKDVNAPVAVEVADS E SKKP 75 F.t. NI SLNS S FLERS SVTGG SRTQ S QS LRL SARRPVVTSMLNSNS L P ERIqVSVSVDSAVRDVNAPVAVEVDRSVGEKP 75 M.C. MGGSNALNEMTNG 13 P.V. MSSISLKENGGEVSVKK 17 P.t. MESTLKEMRDGASVLDMDP 19 Z.m. ML S TRTAAVAASAS PAS PWKLGGRS EGGAS C DGCRTYRNTLRRRAAPAKVRALP PRRVDAVAMV 64
F.p. TAVVGGGVEDVYGEDSATEDHFITPWSVSVASGYSLLRDPHHNKGLAFTEKERDAHYLRGLLPPVIrVNHDLQVKK 150 F.t. FAAVGGGVEDMYGEDTATEDHYITPWSVSVASGYSLLRDPHHNKGLAFTEKERDAHFLRGLLPPVVVNHDLQVKK 150 M.c. S DG I TGGVADVYGEE FATQDQLVTPWSF SVACGHSLLRDPQHNKGLAFTEKERDAHFLRGLLP PWL SQELQEKK 88 P.v. DYSNGGGVRDLYGEDSATEDHL I TPWTF SVASGC SLLRDPRYITKGLAFTEGE RDAHYLRGLL p p SVFNQE LQEKR 92 P.t. KS TVGGGVEDVYGEDRATEDQLVT PWT I SVASGYTLLRDPHHNKGLAFTEKERDAHYLRGLL p PTT I SQ QLQEKK 94 Z.m. SNAETETEKEQEEAAAASEELPVMPWATSVASGYTLLRDPHHNKGLAFTEEERDGHYLRGLLPPAVLSQELQIKK 139
C
* * * * * * * * * * * * * * * * * * * * * * * * * * * * e * * * * * * * * * * * * * *
C4 II C C4 C
m . p .
F . t .
M.C. P.V. P.t. Z.m.
YIGLRQRRASGKEYAELMNEFMSAVKQNYGEKVL I QFEDFANHNAFDL LEKYRTTHLVFNDD I QGTASVVLAGL I 375 YIGLKQKRAAGQEYAELMNEFMSAVKQNYGENLL I QFEDFANHNAFDL LEKYRTTHLVFNDD I QGTASVVLGGL I 375 YI GLKQKRATGEEYAEFVQEFMSAVKQNYGEKI LVQFEDFANHNAFEL LEKYRTTHLVFNDD I QGTASVVLAGL I 313
319
C4 III 0 4
C
C4 C4
C

binding sites. Six amino acid positions (marked C4 in Fig. 1) can be detected which are identical in the C4 isoform of the dicot F. trinervia and of the monocot Zea mays, but differ in the C3 and CAM isoforms. Whether these residues are at least partly responsible for the reported differ- ences in the kinetic properties of C3 and C4 NADP-ME isozymes (cf. [6]), is not known. The availability of cloned C3 and C4 NADP-ME iso- forms from the two closely related plants F. tri- nervia and F. pringlei will allow this to be tested by producing active enzymes in Escherichia coIi and searching for structure-function relation- ships.
Phylogenetic comparisons
To investigate the phylogenetic relationship of the various plant and animal NADP-MEs a parsi- rnony analysis was performed (Fig. 2). A single minimum length tree of 1112 steps was obtained using NAD-malic enzyme of Bacillus stearother- mophilus as an outgroup. The ctadogram obtained shows that the mono- and dicotyledonous NADP-malic enzymes are placed on different branches of the tree. This supports the previous conclusion that the transit pepfides of the F. tri- nervia and Z. mays C4 isoforms are not homolo- gous and, hence, that the mono- and dicotyledon- ous C4 isoforms evolved independently from each other [2]. The cladogram shows also that the plastidic and cytosolic isoforms of NADP-ME group separately among the dicotyledonous spe- cies suggesting that these two isoforms evolved after the monocot-dicot divergence. However, at the current state of knowledge it is too early to speculate, because no monocotyledonous se- quences of cytosolic NADP-MEs have been re-
1779
Bac i l l us st. (NAD-ME)
........... M . m u s c u l u s
F. ~rirterv]a (04; chloroplast)
E p r i ng le i (03; chloroplast)
, fR vu lgar is (C3; cytosol)
P. t r i c h o c a r p a (03; cytosol)
M. c rys ta t#nurn (CAM; cytosol)
Z, m a y s (C4; chloroplast)
Fig. 2. Parsimony analysis of NADP-malic enzymes from F. pringlei, F. trinervia [2], Z. rnays [24], Phaseolus vulgaris [29], Populus trichocarpa [ 31 ], M. crystalIinum [ 5 ], Mus muscuIus [ 1 ] and Rattus rattus [20]. The NAD-malic enzyme of Bacillus stearothermophilus [ 16] was included as an outgroup. The fol- lowing amino acid residues of the malic enzymes were used for the alignment with CLUSTAL V [ 15]: M. musculus: 11 to 553; R. rattus: 11 to 553; F. pringlei and F. trinervia: 104 to 647; Z. mays: 93 to 636; M. crystallinum: 41 to 585; P. vulgaris: 13 to 589; P. trichocarpa: 9 to 589. Parsimony analysis with PAUP 3.0 [28] was performed using the branch and bound option.
ported yet. It is also not known whether plastidic NADP-MEs are an acquisition of the an- giosperms or whether they are common to gym- nosperms, pteridophytes and bryophytes. Thus one has to await further analyses, before a con- clusive statement can be made.
Southern analysis
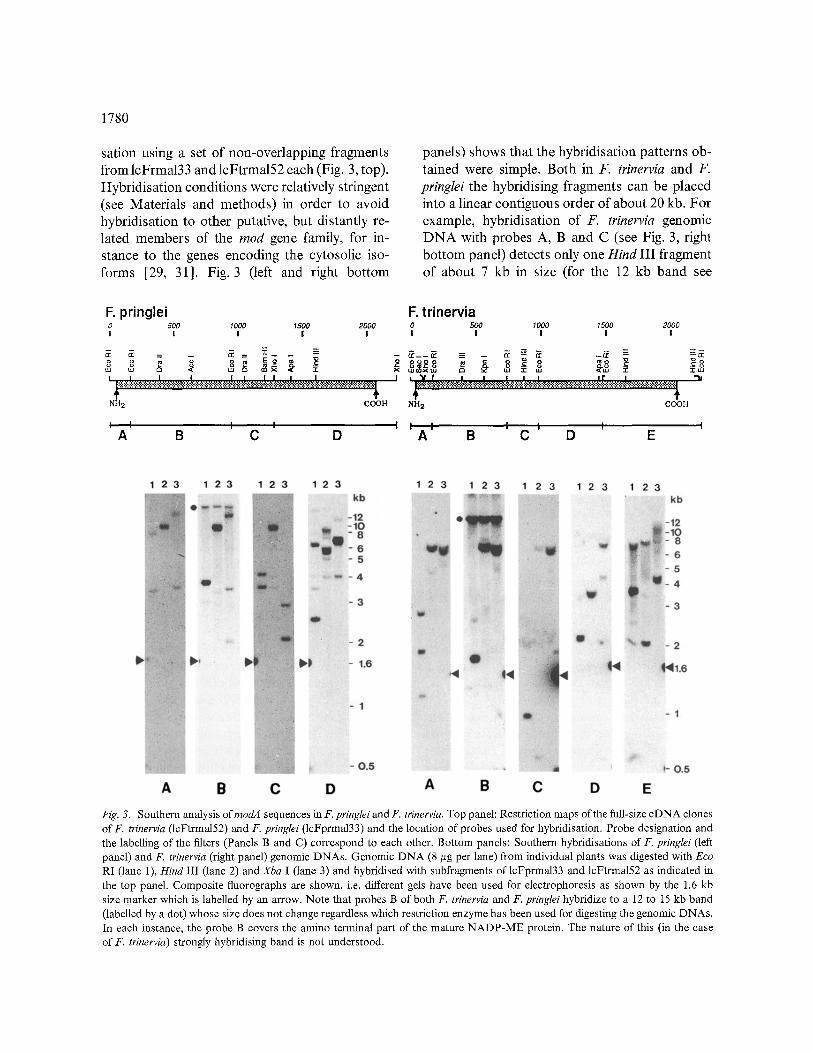
The close sequence similarity of the NADP-MEs of F. pringlei and F. trinervia and the fact that apparently only one class of cDNAs was isolated from the F. pringlei leaf cDNA library suggested that the two NADP-MEs were orthologous pro- teins [4], i.e. the least common ancestor of F. pringtei and F. trinervia already contained a modA gene. To challenge this conclusion the genomic representation of modA sequences was analysed in F. pringlei and F. trinervia by Southern hybridi-
Fig. 1. Amino acid sequence of the F. pringlei modA isoform of NADP-ME and alignment with NADP-MEs from F. trinervia (C4), Zea mays (C4), Phaseolus vulgaris (C3), PopuIus trichocarpa (C3) and Mesemb~yanthemum crystallinurn (CAM). The amino acid sequence of NADP-ME of the C3-C 4 intermediate F. lineatis was not included in the comparison, because no complete sequence is available yet [22]. The putative cleavage sites of the transit peptides of the NADP-ME precursors from F. trinervia, F. pringlei and Z. mays are indicated by arrows; they have been tentatively assigned by applying the rules of yon Heijne [9, 30]. Identical amino acids of the F. trinervia and F. pringlei precursor polypeptides are marked by dots, while residues identical in all six pro- teins are underlined by an asterisk. Grey boxes indicate amino acid residues of putative functional importance (see text).
1780
sation using a set of non-overlapping fragments from lcFrma133 and lcFtrma152 each (Fig. 3, top). Hybridisation conditions were relatively stringent (see Materials and methods) in order to avoid hybridisation to other putative, but distantly re- lated members of the rood gene family, for in- stance to the genes encoding the cytosolic iso- forms [29, 31]. Fig. 3 (left and right bottom
panels) shows that the hybridisation patterns ob- tained were simple. Both in F. trinervia and F. pringlei the hybridising fragments can be placed into a linear contiguous order of about 20 kb. For example, hybridisation of F. trinervia genomic DNA with probes A, B and C (see Fig. 3, right bottom panel) detects only one Hind III fragment of about 7 kb in size (for the 12 kb band see
F. pringlei F, trinervia 0 500 1000 1500 2 0 0 0 0 5 0 0 11200 1500 2 0 0 0 I I I | ! I I ! ! I
v,J uJ l × ~ [ ) × | × ,~ ,~ x a I , "4 t , , , ~ , , t , " ~
q ~ q NH 2 COOH NH 2 COOH
1 I I I I I I l I I I A B C D A B C D E
Fig. 3. Southern analysis ofmodA sequences in F. pringlei and F. trinervia. Top panel: Restriction maps of the full-size cDNA clones of F. trinervia (lcFtrma152) and F. pringlei (lcFprma133) and the location of probes used for hybridisation. Probe designation and the labelling of the filters (Panels B and C) correspond to each other. Bottom panels: Southern hybridisations of F. pringlei (left panel) and F. trinervia (right panel) genomic DNAs. Genomic DNA (8 #g per lane) from individual plants was digested with Eco RI (lane 1), Hind III (lane 2) and Xba I (lane 3) and hybridised with subfragments of lcFprma133 and lcFtrma152 as indicated in the top panel. Composite fluorographs are shown, i.e. different gels have been used for electrophoresis as shown by the 1.6 kb size marker which is labelled by an arrow. Note that probes B of both F. trinervia and F. pringlei hybridize to a 12 to 15 kb band (labelled by a dot) whose size does not change regardless which restriction enzyme has been used for digesting the genomic DNAs. In each instance, the probe B covers the amino terminal part of the mature NADP-ME protein. The nature of this (in the case of F. trinervia) strongly hybridising band is not understood.

legend to Fig. 3) The two remaining, i.e. carboxy- terminal fragments of the cDNA hybridise to three further fragments of 3.5 (probe D), 3.6 and 7 kb (probe E) resulting in a total of 20 kb hybridising sequences. Taking into account that the maize C4 NADP-ME gene has been reported to contain 18 introns (cited after [6]) a gene size of more than 10 kb can be envisaged. The data suggest, there- fore, that both in F. trinervia and F. pringlei modA is a single-copy gene and that the two modA genes are orthologous.
Expression analysis
The high conservation of modA sequences be- tween F. pringlei and F. trinervia suggested that it should be possible to reliably compare the abun- dance of modA transcripts in the various Flaveria species by using one common hybridisation probe, i.e. the cDNA of F. pringlei. Therefore, total RNA was isolated from young, expanding leaves of the C4 species F. trinervia, the C4-1ike species F. brownii, the C3-C4 intermediates F. floridana, F. linearis and F. chloraefolia as well as the C3 species F. pringlei and analysed by north- ern hybridisation. Comparable loadings of RNA were tested by hybridising the northern filters with a cDNA probe encoding hsp70 m R N A sequences (data not shown; see Fig. 5 of [23]). Figure4 (Panel A) shows that - as to be expected from the cDNA cloning experiments - modA sequences are expressed in the leaves ofF. pringlei, but that the modA RNA levels increase gradually when comparing the C3, C3-C4 intermediate and C4 species. Thus the amount of modA transcript in the leaves is linked with the degree of C4 charac- teristics expressed [7, 17] as has also been found for pyruvate orthophosphate dikinase transcript levels [23].
To study the organ specificity of modA expres- sion in F. pringlei a northern analysis was carried out with RNA from leaves, stems and roots. Figure 4 (Panel B) depicts that in F. pringlei the modA gene is more or less constitutively ex- pressed, i.e. comparable levels of its m R N A are detectable in all three organs. Surprisingly, modA
1781
Fig. 4. Species- and organ-specific accumulation patterns of modA transcripts in Flaveria. Panel A: transcript abundance in the leaves ofF. trinervia (C4), F. brownii (C4-1ike), F. flori- dana (C3-C4), F. linearis (C3-C4), F. ehloraefolia (C3-C4) and F. pringlei (C3). Twenty #g of total RNA was applied per lane. The northern blot was hybridised with probe D of the the F. pringlei cDNA (see map in Fig. 3). Exposure time 12 h. Panel B: modA transcript levels in leaves, stems and roots of F. pringlei. Two/*g poly(A) + RNA was analysed in each case using the F. pringlei cDNA (probe D) as a probe. Exposure time 2 days. Panel C: modA transcript levels in leaves, stems and roots of F. trinervia. One #g poly(A) + RNA from leaves and 10/zg poly(A) + RNA from stem and root, respectively, were applied per lane. The blot was hybridised with probe E of the F. trinervia cDNA (see Fig. 3). Exposure time 2 days. All northern hybridisations were carried out at least in dupli- cate using independent RNA preparations. Representative blots are shown.
transcripts can also be found in stems and roots of the C4 plant F. trinervia (Fig. 4, Panel C) indi- cating that the expression of the C4 isoform gene is not confined to the leaves (cf. [2]). However, the visual comparison of the fluorographs clearly demonstrates that in contrast to F. pringlei modA expression in F. trinervia is by far more abundant in the leaves than in stems and roots (cf. [2]). Therefore, the leaf-specific increase in expression levels must have been one crucial step during the evolution of the C4 modA gene. It appears that during this evolutionary change the constitutive expression pattern of the C3 ancestral gene as evidenced by the F. pringlei modA has been main- tained in the C4 plant F. trinervia, but in addition a novel feature, i.e. a high level of expression in the leaves has been acquired.
A similar evolutionary scenario has also been reported for phosphoenolpyruvate carboxylase. A

1782
transgenic analysis showed that essentially only one novel cis-acting domain or element distin- guishes the C4 phosphoenolpyruvate carboxylase gene ofF. trinervia from its orthologous C3 coun- terpart in F. pringlei. This cis-acting domain is located in the 5' upstream region of the F. tri- nervia ppcA gene and is responsible for its high level of expression in the leaves [27].
With respect to the evolution of C4-specific ex- pression patterns it will be very interesting to compare whether similar strategies, i.e. combina- tions of cis-acting elements and their correspond- ing trans-regulatory factors, have been exploited by the various C4 cycle genes. It is also not known yet, how the strict compartmentation of modA expression in the bundle-sheath cells of the leaves [18, 19] has been achieved. Two orthologous modA genes are available now to tackle this prob- lem and to identify the changes in regulatory cis and trans elements which had to occur in order to shape a C4 isozyme gene starting from a C3 an- cestral gene. The fact that the modA gene of F. pringlei encodes a plastidic protein may also en- courage plant biochemists to re-address the func- tion of a plastid-located NADP-ME in C 3 plants.
Acknowledgements
This work was supported by grants from the Deutsche Forschungsgemeinschaft (SFB 189), the Bundesminister f~lr Forschung und Technolo- gie and the Fonds der Chemischen Industrie.
References
1. Bagchi S, Wise LS, Brown ML, Bregman D, Sul HS, Rubin CS: Structure and expression of murine malic en- zyme mRNA. J Biol Chem 262:1558-1565 (1987).
2. BOrsch D, Westhoff P: Primary structure of NADP- dependent malic enzyme in the dicotyledonous C 4 plant Flaveria trinervia. FEBS Lett 273:111-115 (1990).
3. Church GM, Gilbert W: Genomic sequencing. Proc Natl Acad Sci USA 81:1991-1995 (1984).
4. Craig M, Hillis DM: Molecular systematics: context and controversies. In: Hillis DM, Craig M (eds) Molecular Systematics, pp 1-10. Sinauer Associates, Sunderland (1990).
5. Cushman JC: Characterization and expression of a NADP-malic enzyme cDNA induced by salt stress from the facultative crassulacean acid metabolism plant, Mesembryanthemum crystallinum. Eur J Biochem 208: 259-266 (1992).
6. Edwards GE, Andreo CS: NADP-malic enzyme from plants. Phytochemistry 31:1845-1857 (1992).
7. Edwards GE, Ku MSB: Biochemistry of C3-C 4 interme- diates. In: Hatch MD, Boardman NK (eds) The Bio- chemistry of Plants, vol. 10, pp 275-325. Academic Press, New York (1987).
8. E1-Shora HM, ap Rees T: Intracellular location of NADP + -linked malic enzyme in C 3 plants. Planta 185: 362-367 (1991).
9. Gavel Y, von Heijne G: A conserved cleavage-site motif in chloroplast transit peptides. FEBS Lett 261:455-458 (1990).
10. Gutierrez M, Gracen VE, Edwards GE: Biochemical and cytological relationship in C 4 plants. Planta 119:279-300 (1974).
11. Hanukoglou I, Gutfinger T: cDNA sequence of adreno- doxin reductase. Identification of NADP-binding sites in oxidoreductases. Eur J Biochem 180:479-484 (1989).
12. Hatch MD, Kagawa T, Craig S: Subdivision of C 4 path- way species based on differing C 4 acid decarboxylating systems and ultrastructural features. Aust J Plant Physiol 2:111-128 (1975).
13. Hatch MD, Osmond CB: Compartmentation and trans- port in C 4 photosynthesis. In: Stocking CR, Heber U (eds) Transport in Plants. III. Intracellular Interactions and Transport Processes. Encyclopedia of Plant Physi- ology, vol. 3, pp 144-184. Springer-Verlag, Berlin (1976).
14. Hermans J, Westhoff P: Analysis of expression and evo- lutionary relationships of phosphoenolpyruvate carboxy- lase genes in Flaveria trinervia (C4) and F. pringlei (C3). Mol Gen Genet 224:459-468 (1990).
15. Higgins DG, Bleasby AJ, Fuchs R: CLUSTAL V: Im- proved software for multiple sequence alignment. Corn- put Appl Biosci 8:189-191 (1992).
16. Kobayashi K, Doi S, Negoro S, Urabe I, Okada H: Structure and properties of malic enzyme from Bacillus stearothermophilus. J Biol Chem 264:3200-3205 (1989).
17. Ku MSB, Wu J, Dai Z, Scott RA, Chu C, Edwards GE: Photosynthetic and photorespiratory characteristics of Flaveria species. Plant Physiol 96:518-528 (1991).
18. Langdale JA, Rothermel BA, Nelson T: Cellular pattern of photosynthetic gene expression in developing maize leaves. Genes Devel 2:106-115 (1988).
19. Langdale JA, Zelitch I, Miller E, Nelson T: Cell position and light influence C 4 versus C 3 patterns of photosyn- thetic gene expression in maize. EMBO J 7:3643-3651 (1988).
20. Magnuson MA, Morioka H, Tecce MF, Nikodem VM: Coding nucleotide sequence of rat liver malic enzyme mRNA. J Biol Chem 261:1183-1186 (1986).
21. Monson RK, Moore Bd: On the significance of C3-C 4

intermediate photosynthesis to the evolution of C 4 pho- tosynthesis. Plant Cell Environ 12:689-699 (1989).
22. Rajeevan MS, Bassett CL, Hughes DW: Isolation and characterization of cDNA clones for NADP-malic en- zyme from leaves of Flaveria: transcript abundance dis- tinguishes C3, C3-C 4 and C4 photosynthetic types. Plant Mol Biol 17:371-383 (1991).
23. Rosche E, Streubel M, Westhoff P: Primary structure of the photosynthetic pyruvate orthophosphate dikinase of the C 3 plant Flaveria pringlei and expression analysis of pyruvate orthophosphate dikinase sequences in C3, C3-C 4 and C4 Flaveria species. Plant Mol Biol, in press (1994).
24. Rothermel BA, Nelson T: Primary structure of the maize NADP-dependent malic enzyme. J Biol Chem 264: 19587-19592 (1989).
25. Sambrook J, Fritsch EF, Maniatis T: Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1989).
26. Scrutton NS, Berry A, Perham RN: Redesign of the co- enzyme specificity of a dehydrogenase by protein engi- neering. Nature 343:38-43 (1990).
27. Stockhaus J, Poetsch W, Steinm~iller K, Westhoff P: Evolution of the C 4 phosphoenolpyruvate carboxylase promoter of the C 4 dicot Flaveria trinervia: an expression analysis in the C 3 plant tobacco. Mol Gen Genet, in press (1994).
28. Swofford DL: PAUP: Phylogenetic Analysis Using Par- simony, version 3.0. Illinois Natural History Survey, Champaign, Illinois (1989).
1783
29. van Doorsselaere J, Villarroel R, van Montagu M, Inz6 D: Nucleotide sequence of a cDNA encoding malic en- zyme from poplar. Plant Physiol 96:1385-1386 (1991).
30. von Heijne G, Steppuhn J, Herrmann RG: Domain structure of mitochondrial and chloroplast targeting pep- tides. Eur J Biochem 180:535-545 (1989).
31. Waiter MH, Grima-Pettenati J, Grand C, Boudet AM, Lamb CJ: Extensive sequence similarity of the bean CAD4 (cinnamyl-alcohol dehydrogenase) to a maize malic enzyme. Plant Mol Biol 15:525-526 (1990).
32. Wedding RT: Malic enzymes of higher plants. Charac- terization, regulation, and physiological function. Plant Physiol 90:367-371 (1989).
33. WesthoffP, Offermann-Steinhard K, H6fer M, Eskins K, Oswald A, Streubel M: Differential accumulation of plas- tid transcripts encoding photosystem II components in the mesophyll and bundle-sheath cells of monocotyledon- ous NADP-maiic enzyme-type C 4 plants. Planta 184: 377-388 (1991).
34. Wierenga RK, Terpstra P, Hol WGJ: Prediction of the occurence of the ADP-binding beta-aipha-beta-fold in proteins, using an amino acid sequence fingerprint. J Mol Biol 187:101-107 (1986).
35. Winter K: Crassulacean acid metabolism. In: Barber J, Baker NR (eds) Photosynthetic Mechanisms and the En- vironment, pp 329-387. Elsevier Science Publishers (Bio- medical Division), Amsterdam, New York/Oxford (1985).





























