Embed Size (px)
Citation preview

216 Brain Research, 537 (1990) 210-226 Elsevier
BRES 16193
The circadian activity rhythms of rats with mid- and parasagittal 'split-SCN' knife cuts and pinealectomy
J.A. Yanovski, A.M. Rosenwasser*, J.D. Levine and N.T. Adler
Department of Psychology, University of Pennsylvania, Philadelphia, PA 19104 (U.S.A.)
(Accepted 31 July 1990)
Key words: Circadian activity; Suprachiasmatic nucleus; Pinealectomy
To explore the multioscillator nature of the neurohumoral circuitry controlling the expression of circadian rhyttunicity, rats' wheel running circadian activity rhythms were compared following sagittal knife cuts in the region of the supraehiasmatic nucleus (SSCN), following pinealectomy (PX) and following the combination of SSCN and PX. 25% of animals with knife cuts that passed through one SCN had disturbed running activity under constant illumination; rhythmic disturbances were seen neither in animals with sham knife cuts nor in rats with knife cuts on the midline or lateral to the SCN. Animals with both SSCN and PX were twice as likely to show severe rhythmic disruptions under free-running conditions as rats with SSCN and sham PX. Rats with PX and sham SSCN did not display disrupted activity rhythms. When animals with PX alone or SSCN alone were first observed under free-running conditions and then subjected to a second surgical procedure so that all animals underwent both PX and SSCN, all PX and most SSCN animals demonstrated coherent activity rhythms after the first operation, but 35% showed disruptions in circadian activity patterns only following the second surgery. The activity rhythms of rats with knife cuts placed either on the midline or lateral to the SCN did not deteriorate when combined with PX. Rats with coherent rhythms following knife cuts damaging one SCN had rhythm disruptions after the addition of PX. The effects of pinealectomy may indicate that the pineal gland plays a role in maintaining the coupling relationships in the multioseillator system controlling circadian activity rhythms. The results of this study also suggest that neither the direct commissural connection of the SCNs nor the humoral output of the pineal gland is indispensable for the expression of coherent circadian activity rhythms in the rat.
INTRODUCTION
Vertebrate circadian rhythms appear to be the product of the interaction of multiple, coupled oscillators 27' 30,36,44,53. The bilateral suprachiasmatic nucleus (SCN) of
the anterior hypothalamus, considered of great impor- tance in the control of mammalian circadian rhythms, may itself comprise a functionally complex multi-oscil- lator system 28'29'44 with specialization of function within
the SCN 3'12'42. As each SCN appears to have the capacity to function as a competent circadian oscillator 3'14'34'42'51,
it has been suggested that the two SCNs are a pair of mutually coupled circadian pacemakers. Thus, the effects of surgical separation of the SCN's two, reciprocally innervated 4'33'37'48 substituent nuclei have been exam-
ined. In hamsters, a preliminary report suggests that midsagittal knife cuts may produce changes in the patterning of already split free-running activity rhythms 46. Such knife cuts alter the rate of reentrainment after phase shifts of the l ight-dark cycle in ratsa9"s°; however, the effects of midline knife cuts on free-running rat circadian rhythms have not been investigated.
It has also been postulated that there are other neurohumoral mechanisms by which both SCNs maintain coupling 9'2°. A candidate hormone for such a role is
melatonin, produced primarily by the pineal gland. Both
pinealectomy and the administration of melatonin have profound effects on the expression of circadian rhythms of many species of birds, reptiles and fish 17'18'21"23'54"55'57. However , rodents who are either pinealectomized or
exposed to continuous administration of melatonin can express coherent entrained and free-running circadian rhythms 2A1'43'52, even though their rates of reentrain-
ment to phase shifts are often more rapid than those of sham-operated animals 16"24'39'4°. Intact rats kept either in
constant light or in constant darkness can phase lock with daily injections of the pineal hormone melatonin 1'5a°'41. Furthermore, melatonin has a phase-dependent phase- shifting capability for animals kept in constant conditions that depends upon the presence of the SCN 6. Other
experiments have shown that melatonin binds to high affinity receptors within the SCN s6 and can affect the activity of SCN neurons 7'8'26'45.
The present experiment was designed to explore the
* Present address: Department of Psychology, University of Maine, Orono, ME 04469, U.S.A. Correspondence: J.A. Yanovski, The National Institutes of Health, Building 10, Room 10N262, Bethesda, MD 20892, U.S.A.
0006-8993/90/$03.50 © 1990 Elsevier Science Publishers B.V. (Biomedical Division)

functional implications of the anatomic organization of
the SCN and to examine the role of the pineal gland as
a signal coordinating the expression of circadian rhyth-
micity in the mammal ian circadian system. Free running
circadian activity rhythms were examined in rats with
sham, mid- and parasagittal SCN knife cuts, with sham
and actual pinealectomy, and with combinations of knife
cuts plus pinealectomy.
MATERIALS AND METHODS
Animals and apparatus Adult female Sprague-Dawley (Charles River) rats were placed
in Wahmann running wheel cages with attached home cage compartments within light-tight cabinets to record their running activity.
Surgery Operative procedures were carried out during the animals'
subjective night. Surgical anesthesia was induced with intraperito- heal injections of ketamine hydrochioride (100 mg/kg) plus acepro- mazine (2.4 mg/kg).
(.4) Surgical separation of the SCNs (SSCN). Mid- and parasa- gittal knife cuts were made with a retracting wire knife TM. The cuts were aimed to sever the tissue between the third ventricle and the optic chiasm throughout its rostrocaudal extent. Sham-operated animals (sham SSCN), were prepared by lowering the microliter syringe into the animal's brain to the coordinates used to split the SCNs and extending the plunger wire; however, the knife assembly was not lowered to the base of the brain.
(B) Pinealectomy (PX). Pinealectomy was accomplished by modified rapid pinealectomy 22. The venous confluence of sinuses was torn, and the pineal gland visualized with the aid of a dissecting microscope. The gland was grasped by a thin pair of forceps and removed. Sham PX was performed by interrupting the venous confluence of sinuses, just as in actual PX. The pineal gland was visualized in these animals, but not removed. The degree of damage to surrounding brain structures was comparable to that induced by actual PX.
Experimental protocols Experiment 1. Rats were randomly assigned to undergo either
SSCN or Sham SSCN. Long-term activity was recorded under LD 12:12, constant light (LL), constant darkness (DD) and then LD 12:12.
Experiment 2. Rats were randomly assigned to one of 3 surgical groups: simultaneous knife cut and sham pinealectomy (SSCN/sham PX), simultaneous sham knife cut and pinealectomy (sham SSCN/ PX), or simultaneous knife cut and pinealectomy (SSCN/PX). Long term running activity records were obtained under LD 12:12, LL, and DD.
Experiment 3. Rats were randomly assigned to one of two surgical groups. Half of the animals were pinealectomized while the other half underwent SSCN. Running activity was recorded under LD 12:12, LL, DD, and again under LD 12:12. Following stable reentrainment to LD 12:12, each PX animal received a knife cut (PX+SSCN). Similarly, the SSCN animals were pinealectomized (SSCN+PX). Running activity rhythms were again recorded under conditions of LD 12:12, LL, DD, and LD 12:12.
Histological evaluation Following the conclusion of behavioral testing, brain sections
were obtained from the region of the SCN and pineal. Every third 20/~m section was saved in the region of the SCN and stained with 1% Thionine solution. Differences in the sizes of the two SCNs after knife cuts were quantified by use of a computerized planimetric system: The areas of each of the two SCNs were measured in all
217
sections containing this structure, and these areas cumulated to form a statistic proportional to the volume of each SCN, the 'total area' (TA). In order to determine the most conservative estimate of SCN size, all clusters of cell bodies in the suprachiasmatic nucleus region that could be distinguished from ghotic scars were considered to be SCN. The areas of all such SCN fragments were added together to determine the cross sectional area of the SCN in any one section. From the TAs, the percent disparity between the two SCNs was determined by dividing the estimated TA of the smaller SCN by the TA of the larger SCN within each animal, multiplying by 100, and subtracting this number from 100; thus percent disparity increases to indicate increased unilateral SCN damage. Estimates of SCN TAs of 5 unoperated animals were also determined for comparison with those of the undamaged SCNs of knife-cut animals. The TAs from the SCNs of unoperated animals always had percent disparities of less that 13% and did not differ significantly from the TAs of knife-cut animals' undamaged SCNs (P -- 1.26). Therefore, the TA of the undamaged SCN was a reasonable reference for evaluation of knife cut-induced SCN damage. The TA SCN volumetric estimate was employed, rather than individual section areas, in order to render unimportant any possible asymmetries that resulted from brain sectioning technique.
All sections in which the optic chiasm or SCN could be recognized were examined. Either clear separation of the left and fight parts of the chiasm and SCN, or evidence of a gliotic scar emending from the third ventricle to the optic chiasm throughout the entire length of the chiasm was required for classification of a knife cut as having caused complete splitting of the chiasm and SCNs. Efficacy of PX was determined by examination of gross and microscopic brain morphology in the region of the pineal.
Quantitative and statistical evaluation Free-running period (tan) was estimated by eye-fitting a straight
fine to the approximate midpoints of each daffy activity bout and graphically determining the slope of that line. The presence of statistically significant circadian rhythmicity was evaluated by Chi-square periodogram analysis 4T.
RESULTS
Experiment 1
Behavioral and histological observations were obtained
from 34 rats, including 29 SSCN and 5 S H A M animals
(Table I). SSCN knife cuts could be divided on the basis
of histological appearance into 3 categories (Fig. 1): knife
cuts were on the midline, sparing both SCNs in 8 animals
(28%), and displaced laterally so as to pass through one
SCN in 14 animals (48%) and produce damage lateral to
the SCN in 7 animals (24%). S H A M knife cuts produced
damage solely to brain regions dorsal to the SCN.
The 8 animals with SSCN knife cuts that were found to
be accurately placed on the midline had SCN disparities
of less than 14%. In addit ion, these cuts usually (5/8)
severed the optic chiasm completely. All 8 animals with
midline knife cuts showed clear evidence of coherent
circadian rhythmicity under both en t ra inment and free-
running conditions (Fig. 2). Mean f ree-running LL tan
for animals with midline SSCN knife cuts was 25.09 +
0.37 h.
Ten of the 14 SSCN knife cuts that damaged one SCN
severed the optic chiasm completely. Of these 14 rats, 3
did not run sufficiently for an evaluat ion of rhythmicity

218
and seven showed coherent activity rhythms with a mean free-running period of 24.90 + 0.38 h under LL. of the
14
Four rats with unilateral SCN damage displayed dis-
rupted free-running rhythmicity under LL. One of these animals appeared to demonstrate 'splitting' of its activity
rhythms into two distinct components with different
circadian periods. An additional 3 rats with unilateral
SCN damage showed more severe rhythmic disturbances under LL. For example, an animal that had a 64%
difference in SCN size showed multiple activity compo-
nents with no significant circadian peak, but with a
significant 6-h period, as detected by periodogram anal- ysis (Fig. 3). Arrhythmicity was also displayed by two animals with 80% and 62% SCN disparities.
Five of the 7 SSCN knife cuts that were sufficiently
lateral so as to leave both SCNs intact severed the optic
chiasm. Five animals with lateral knife cuts had coherent
rhythms under both LL and DD, while two did not
generate enough wheel-running behavior for evaluation of rhythmicity. The mean free-running period under LL
for these 5 animals was 25.0 + 0.50 h. After SHAM knife cuts there were differences of less
than 11% in the estimated TAs of the two SCNs.
Coherent circadian rhythmicity was observed under LD, LL and DD in all 5 SHAM animals, with mean LL tau
25.15 + 0.39 h.
For those animals with detectable free running rhythms, no significant differences in free-running period
TABLE I
Behavioral and histological categorization of animals from Expt. 1
The location of the knife cut is divided into midline (knife cut passed between the two SCN without damaging either), lateral (knife cut passed lateral to the SCN without damaging it), SCN (knife cut produced unilateral SCN damage) and sham (knife cut sparing the SCN and other structures on the ventral part of the brain). The group of animals with knife cuts passing through one SCN is subdivided into large SCN disparities (more than 50%) and small SEN disparities (less than 50%), Behavior under LL is categorized into coherent (not distinguishable from intact animals), sprit (showing evidence of 'splitting' of the running activity rhythm into two components with differing periods), disorganized (more than two free-running compo- nents or no evident circadian periodicity), and insufficient (not enough running activity to evaluate circadian rhythms). Percentages are given in parentheses; n is total number of animals in each histological group.
Knife cut Coherent Split or dis- Insuff4cient n organized
Midline 8(100%) 0 (0%) 0 (0%) 8 Lateral 5 (71%) 0 ( 0 % ) 2(29%) 7 SCN (large) 3 (33%) 4 (45%) 2 (22%) 9 SCN (small) 4 (80%) 0 (0%) 1 (20%) 5 SHAM 5(100%) 0 (0%) 0 (0%) 5 Allanimals 25 (73%) 4 (12%) 5 (15%) 34
under LL or DD were found (t-tests, n.s.). Disruptions in rhythmicity occurred only in animals with extensive
unilateral SCN damage (Table I): only rats with greater
than 50% disparity between their SCNs displayed severe
rhythm disruptions such as splitting and rhythmic disor- ganization (Fig. 3). Circadian rhythms were disrupted
significantly more frequently in rats with more than
50% unilateral SCN damage (P = 0.0002, Fisher exact test).
Experiment 2
The results from the twenty-six rats who completed this
experiment are summarized in Table II. Phase of entrain- ment to light-dark cycles was not distinct for any group.
All PX/sham-SSCN rats showed coherent circadian ac-
tivity patterns under LL and DD. Mean free running
period (tau) for these animals under LL was 24.88 + 0.67
h. Five of the 7 SSCN/sham PX animals also had coherent circadian rhythms under both LL and DD; mean LL tau
for these animals was 24.91 + 0.54 h. Only two rats with SSCN/sham PX showed disruptions in their free,running
activity rhythms. An activity pattern similar to 'splitting' and one that appeared to be arrhythmic under constant conditions were seen.
Of the 12 rats that received SSCN/PX, two rats did not
display enough activity to determine the presence of circadian rhythmicity. Of the remaining 10 rats, 4
displayed coherent circadian rhythms under LL and DD,
with average LL tau of 24.90 + 0.72 h. Six had
disruptions in their circadian rhythmicity, with two rats
demonstrating split-like running activity and 4 appearing to have arrhythmic activity patterns. Four of the six
SSCN/PX rats with rhythmic disruptions under LL demonstrated disturbances of rhythmicity under DD.
1 2 3
Fig. 1. Schematic representation of the SSCN knife cuts. Coronal brain section based on Paxinos and Watson ~2. Midline lesions, sparing both SCNs, 1; through one SCN sparing the other, 2; lateral to the SeN sparing both, 3. Fornix, F; third ventricle, 3 r° V; supraoptic nucleus, SON; optic chiasm, OC; suprachiasmatic nucleus, SCN.

219
A
B SSCN, LD
LL
i i
D D - - _ " - - ~ -
mid " k I i
I
- . ~ m I
h
I I
Im
i .
- - h . 11 ~ i
m - • - -
• i i i - ~
. f - - L
i
m
J l
. . . . . Z 2 - - - " -
• _ u .
I h I
I b
I l l i i
~L
i
- - _ m a " L
m m
i • . L
i l | i i
, n
. . . . d l lU l
I
Fig. 2. A: photomicrograph from a rat with a midiine SSCN knife cut that severed the optic chiasm completely, sparing both SCNs. There was a 9% disparity in the two SCNs. B: circadian actogram from this animal. Successive days are represented from top to bottom along the vertical axis and time of day is represented along the horizontal axis. The record is double-plotted. Light-dark 12:12 schedule, LD; first full day of constant light, LL; first full day of constant darkness, DD.

sluouodtuoo £l!^!loe ue!peoa!ouou old.qlnm ql!~ tuolled ,(l!A!lOe 8u!uuna e ~u!l~11suotu~p [etu!ue s!ql tuoaj tue.=~oloe ue!peaa!o :[I "lno ~J!U~l N~)SS aolJ~ ,(l.ulcds!p %t,9 e ql!A~ lea e tuoaj qd~.li]oJa.Kuoloqd' :V "£ "~!et
#'lt'l
I,
T~ .. r-- .... --~T.: " --
_ Z__ _ ..$ .T. if__ .~
Ill 'NOSS 8
V

TABLE II
The quality of the running activity rhythms under LL of animals from Expt. 2
Behavior is characterized as in Table I. Animals simultaneously undergoing both PX and SSCN (SSCN/PX) are twice as likely to have rhythmic disturbances as those with SSCN and sham PX (SSCN/ Sham PX). Animals with PX but sham SSCN (Sham SSCN/PX) always show coherent circadian activity rhythms.
Group Coherent Split or dis- Insufficient n organized
Sham SSCN/PX 7 (100%) 0 (0%) 0 (0%) 7 SSCN/Sham PX 5 (72%) 2 (28%) 0 (0%) 7 SSCN/PX 4 (33%) 6 (50%) 2 (17%) 12 Total 16 (61%) 8(31%) 2 (8%) 26
Exper iment 3
Results ob ta ined f rom 40 rats in this exper iment are
summar ized in Table III . Act ivi ty records and coronal
sections of the anter ior hypotha lamus at the level of the
SCN from representa t ive animals are shown in Figs. 4 -7 .
Behavior o f rats with P X + S S C N
Sixteen rats survived initial p inealec tomy; none
showed any disrupt ions in circadian rhythmici ty pr ior to
SSCN. Average free running tau under L L was 25.05 +
0.71 h.
Af t e r the second opera t ion (SSCN) 5 animals contin-
ued to display coherent circadian rhythms, (average L L
tau 25.10 + 0.77 h). Five o ther animals had insufficient
activity to de te rmine the presence or absence of free-
running circadian rhythmicity. Six rats showed disrup-
A
B SSCN, LD
LL
DD
LD, PX
221
TABLE III
Behavior and histological categorization under LL for animais in Expt. 3 foUowing both PX and SSCN
Those animals who received SSCN first and PX second are designated SSCN+PX; those with the reverse order are designated PX+SSCN.
Knife cut Coherent Split or dis- Insufficient n organized
SSCN + PX Midfine 4 (67%) 0 (0%) 2 (33%) 6 Lateral 3 (60%) 0 (0%) 2 (40%) 5 SCN (large) 1 (14%) 4 (57%) 2 (29%) 7 SCN (small) 1 (17%) 4 (66%) 1 (17%) 6 Total 9 (38%) 8 (33%) 7 (29%) 24
PX + SSCN Midline 2 (67%) 0 (0%) 1 (33%) 3 Lateral 2 (67%) 0 (0%) 1 (33%) 3 SCN (large) 0 (0%) 1 (33%) 2 (67%) 3 SCN (small) 1 (14%) 5 (72%) 1 (14%) 7 Total 5 (31%) 6 (38%) 5 (31%) 16
LL
DD
LD
Fig. 4. A: photomicrograph of a coronal brain section of the anterior hypothalamus from a rat that underwent pinealectomy (PX) and then a knife cut (SSCN). The knife cut passed through the midline, sparing each SCN. SCN disparity is 11%. B: circadian actogram from this animal. All animals with midline knife cuts had coherent circadian rhythms.

222
tions in their circadian activity patterns under constant conditions; 4 of these animals appeared to be arrhythmic under both LL and DD while the remaining two displayed disruptions of running activity rhythms under LL and regrouping of activity rhythms under DD. Periodogram analysis showed splitting in one animal and failed to detect a significant circadian peak in activity for the remaining animals under LL. Under DD, circadian peaks were detected only for the two animals that appeared to regroup under DD.
Behavior of rats with SSCN+ PX Twenty-four rats received SSCN as their first opera-
tion. Twenty SSCN rats displayed normally coherent circadian rhythms under LD, LL and DD (average LL tau 25.0 + 0.85 h). Three SSCN rats displayed disrup- tions in circadian rhythmicity under constant conditions. One rat appeared arrhythmic under LL while two showed evidence of activity splitting under LL. All 3 had coherent circadian rhythms under DD.
Of the twenty-two SSCN rats surviving subsequent PX, 5 did not produce enough behavior for analysis of their free-running rhythms. Nine animals displayed coherent circadian rhythms (average LL tan 24.93 + 0.79 h), 7 displayed free-running activity patterns that were markedly less coherent than their activity rhythms prior to PX, and the one animal who was arrhythmic only in LL following SSCN expressed a disrupted running activ- ity pattern under both LL and DD after PX. Periodo- gram analysis revealed no dominant circadian activity peak after PX under either LL or DD for 6 of the 8 animals with rhythmic disturbances; the two remaining rats demonstrated disrupted running activity rhythms only under- LL.
Correlating anatomy with behavior Pinealectomy was microscopically complete in all
animals. The brain damage produced by SSCN surgery was classified as in Fig. 1.
All animals with SSCN knife cuts lateral to the SCN had SCN size disparities of less than 11%. All displayed coherent circadian rhythms under LD, LL and DD, regardless of whether PX preceded or followed SSCN (Table III). All animals with SSCN knife cuts on the midline had SCN size disparities of less than 12% and displayed coherent activity rhythms under LD, LL and DD regardless of whether PX preceded or followed SSCN (Fig. 4, Table III). Six of the 9 midline knife cuts appeared to have severed the tissue between the two SCNs and the underlying optic chiasm completely. There were no differences in tau between animals with com- plete or incomplete knife cuts (t-tests).
All rats that demonstrated disturbances in free-running
rhythmicity had SSCN knife cuts which passed through one SCN (Table III). The addition of PX to SSCN made the appearance of disrupted rhythmicity significantly
B PX, LD
LL
DD
SSCN, LD
LL
DD
Fig. 5. A: photomicrograph from a rat with a knife cut through one SCN and PX. SCN disparity is 37%. B: circadian actogram shows disordered rhythm under LL and DD following SSCN.
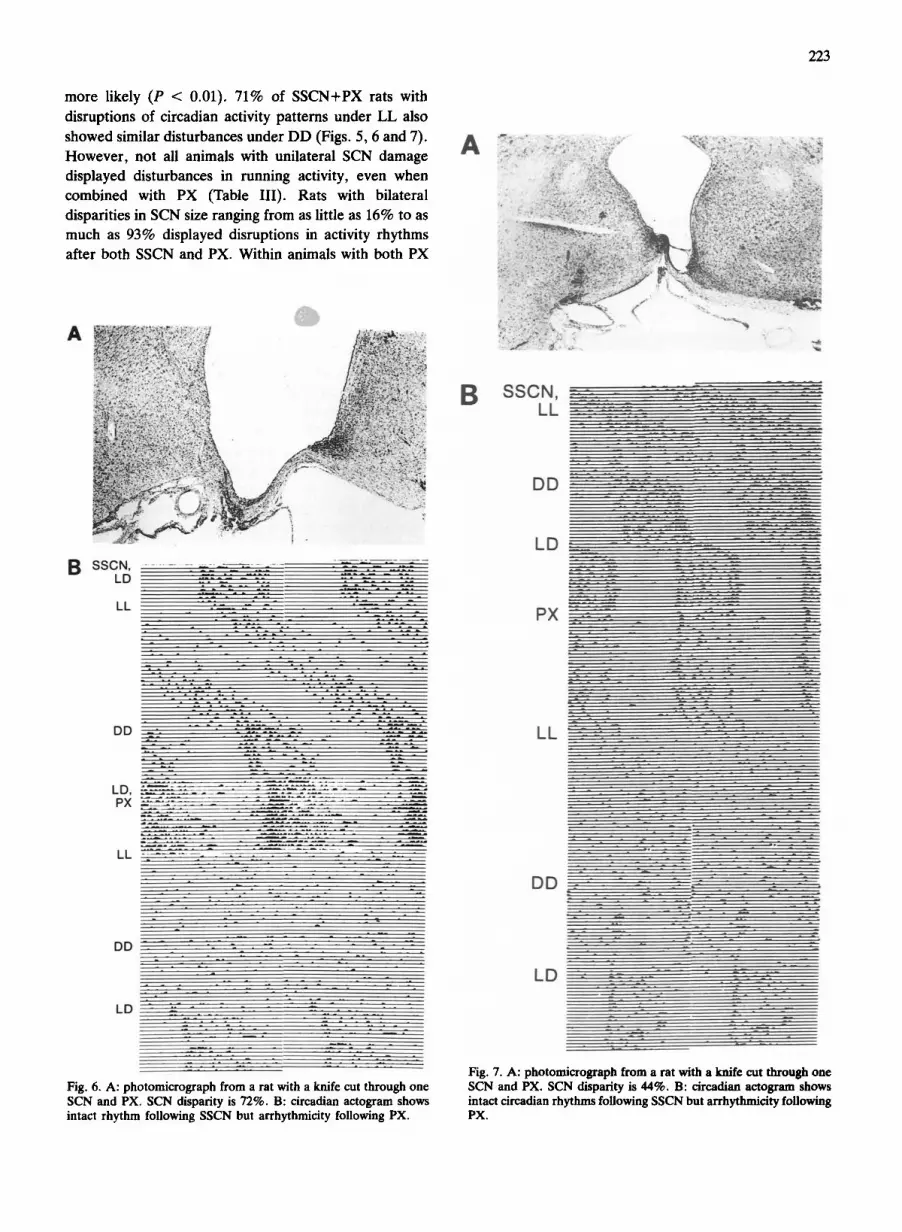
more likely (P < 0.01). 71% of S S C N + P X rats with
disruptions of circadian activity pat terns under LL also
showed similar disturbances under D D (Figs. 5, 6 and 7). However , not all animals with unilateral SCN damage displayed disturbances in running activity, even when combined with PX (Table I I I ) . Rats with bilateral disparities in SCN size ranging from as little as 16% to as much as 93% displayed disruptions in activity rhythms after both SSCN and PX. Within animals with both PX
/t
B SSCN, LD .~'~ _ -:-'- _~ ~1-- ~-~ ~ - Z - -:~'~
LL
== - - - x . . . . -~ : =
A
B SSCN, LL
DD
LD
PX
223
DD = - - - -
w
¢,
LD, PX
LL
D D
LD
LL
DD
LD
Fig. 6. A: photomicrograph from a rat with a knife cut through one SCN and PX. SCN disparity is 72%. B: circadian actogram shows intact rhythm following SSCN hut arrhythmicity following PX.
Fig. 7. A: photomicrograph from a rat with a knife cut through one SeN and PX. SCN disparity is 44%. B: circadian actogram shows intact circadian rhythms following SSCN hut arrhythmicity following PX.

224
and partial unilateral SCN damage, no correlation was found between the likelihood of disruptions of circadian rhythmicity and the area or amount of SCN damage.
DISCUSSION
Effects of subtotal, unilateral SCN damage alone Knife cuts passing through the SCNs have now been
examined in the sagittal, horizontal 3 and coronal planes 12. All 3 types of knife cuts produce subtotal SCN damage and can disrupt the expression of circadian activity rhythms.
In these experiments, several animals with sagittal knife cuts leading to a disparity in SCN TA of more than 50% had significant disruptions in the expression of circadian rhythmicity under LL. Interestingly, complete unilateral SCN destruction does not alter the expression of rat free-running circadian rhythms 14'3s'42'51. However,
following incomplete bilateral SCN lesions, the degree of rhythmic disruption does depend on the extent of SCN damage 3A2,a3,lS'31,3s'42. Arrhythmicity under LL may
occur in intact rats, but because of the relatively low illumination employed in these experiments (200 lux), is infrequent in our experimental apparatus. As arrhythmi- city was seen only in animals with knife cuts through the SCN, it appears likely that the observed effects are related to SCN damage and not solely to the effects of LL.
It is possible that SSCN knife cuts affecting circadian rhythms did so by interfering with communication be- tween the SCNs and other neural areas participating in the expression of circadian rhythmicity 3. It is also possible that the selective destruction of particular SCN subdivisions or cell types may underlie the observed behavioral disruptions 29'3s. Furthermore, knife-cut in-
duced differences in the innervation of the SCN by the retinohypothalamic tract might disrupt oscillator function or render the animal more susceptible to the effects of LEa9, 50.
The pineal gland in the expression of circadian rhythmicity The results of Expts. 2 and 3 indicate that the
combination of PX and SSCN produces more frequent and more profound disturbances in free-running circa- dian activity rhythms than either operation alone and suggest a role for the pineal gland in the expression of rat circadian rhythms.
Expt. 2 found that animals with both SSCN and PX were twice as likely to manifest disruptions in activity patterns as were animals with SSCN and sham PX. Because Expt. 1 found that some animals with SSCN knife cuts causing unilateral SCN damage may have decreases in the coherence of their circadian rhythms
even without the addition of PX, one could not conclude definitively from Expt. 2 alone that PX changed the likelihood of rhythmic disruptions in animals with SCN damage. It was possible that those SSCN/PX animals that expressed disordered circadian rhythms were destined to show them solely as a result of their knife cut-induced SCN damage rather than from the combination of the knife cut with PX.
In Expt. 3, SSCN animals with partial SCN damage but coherent circadian rhythms under LL and DD were shown to have disruptions in their free-running circadian running patterns after the addition of PX. Thus, PX did increase the likelihood of rhythmic disruptions, suggest- ing a role for the pineal gland in the rat circadian rhythm generating apparatus that is uncovered only after the SCN is damaged by a knife cut.
Animals with both SSCN and PX are quantitatively different from those with SSCN alone in two major respects. First, PX renders unimportant the extent of SCN damage induced by a knife cut. Animals with SSCN alone showed rhythmic disturbances only when they had large SCN size disparities; however, some animals with small differences in SCN size were arrhythmic following PX (Table III). Second, most animals that have disrup- tions in their running activity patterns after partial SCN lesions show regrouping of their circadian rhythms under DD (Expt. 1). This was not the case for most animals with both PX and SSCN-induced SCN damage; such rats frequently showed disruptions in their free-running cir- cadian rhythms under both LL and DD. Thus the addition of PX to SSCN produced both more frequent and more profound disturbances in circadian rhythmicity than SSCN alone.
The effect of the combination of SSCN and PX may be a consequence of the response of the pineal gland to constant conditions. Under LL, the pineal gland of the intact rat produces little melatonin as light inhibits pineal production of melatonin by means of a polysynaptic system that is gated through the SCN 25, Under DD, however, the activity of the pineal gland of the intact rat has a high amplitude circadian rhythm. If the pineal gland's humoral output acts to increase the coordination of the multiple oscillators involved in the determination of the running activity rhythm, then under DD, when that output normally expresses circadian rhythmicity, coherence may be reestablished. Thus, if a rat sustains SCN damage that uncouples one or more oscillating SCN subregions from the rest, such damage might produce an arrhythmic activity pattern under LL. However, once the rat is placed under DD, periodic output from the pineal gland driven by the intact portion of the SCN might help to coordinate the neural activity of regions previously oscillating out of phase with one another. In the

pinealectomized rat, no pineal-derived melatonin cou-
pling signal could exist; if incoherent circadian rhythms
were expressed under LL, there could be no rhythmic pineal signal under D D that could act to aid in the coordination of such uncoupled SCN subregions.
A role for the pineal-derived hormone melatonin as an
agent facilitating the coordination of the oscillators
controlling circadian activity rhythms is consistent with findings that functional melatonin receptors are located on SCN neurons 7'8'26'45'56. It is presumably through
interaction with these receptors that melatonin injections entrain circadian rhythms 1'5'6'41 and that PX affects the rate of reentrainment following phase shifts 16'24"39'4°.
The two SCNs as two physically separable, coupled
pacemakers
The coherent free-running activity patterns obtained
from rats with midline SSCN knife cuts despite PX are taken as evidence that destruction of the direct fibers
interconnecting the two SCNs does not preclude the
expression of coherent circadian rhythmicity, even in the absence of pineal melatonin. Such results do not support
ABBREVIATIONS
LD LL DD
light-dark cycle constant light constant darkness
REFERENCES
1 Armstrong, S.M., Cassone, V.M., Chesworth, M.J., Redman, J.R. and Short, R.V., Synchronization of mammalian circadian rhythms by melatonin, Z Neural Transm. SuppL, 21 (1986) 375-394.
2 Asehoff, J., Gerecke, U., von Goetz, C., Groos, G. and Turek, EW., Phase responses and characteristics of free running activity rhythms in the golden hamster: independence of the pineal gland. In J. Aschoff, S. Daan and G.A. Groos (Eds.), Vertebrate Circadian Systems, Springer, Berlin, 1982.
3 Brown, M.H. and Nunez, A.A., Hypothalamic circuits and circadian rhythms: effects of knife cuts vary with their placement within the suprachiasmatic area, Brain Res. Bull., 16 (1986) 705-711.
4 Card, J.E, Brecha, N., Karten, H.J. and Moore R.Y., Immu- nocytochemicai localization of vasoactive intestinal polypeptide- containing cells and processes in the suprachiasmatic nucleus of the rat, J. Neurosci., 1 (1981) 1289-1303.
5 Cassone, V.M., Chesworth, M.J. and Armstrong, S.M., Dose- dependent entrainment of rat circadian rhythms by daily injection of melatonin, J. Biol. Rhythms, 1 (1986) 219-229.
6 Cassone, V.M., Chesworth, M.J. and Armstrong, S.M., En- trainment of rat circadian rhythms by daily injection of mela- tonin depends on the hypothaiamic suprachiasmatic nuclei, Physiol. Behav., 36 (1986) 1111-1121.
7 Cassone, V.M., Roberts, M.H. and Moore, R.Y., Melatonin inhibits metabolic activity in the rat suprachiasmatic nuclei, Neurosci. Lea., 81 (1987) 29-34.
8 Cassone, V.M., Roberts, M.H. and Moore, R.Y., Effects of melatonin on 2-deoxy-[1-14C]glucose uptake within rat suprachi- asmatic nucleus, Am. J. Physiol., 255 (1988) R332-337.
9 Cassone V.M. and Menaker, M., Is the avian circadian system
225
the hypotheses that the two SCNs represent two distinct
circadian oscillators whose coupling is dependent upon
their neural interconnections or upon the humoral output of the pineal gland.
Continued coherent rhythmicity after surgical disrup-
tion of the direct SCN-SCN neural pathway does not necessarily imply that communicat ion between the paired
SCNs is of no importance in the control of circadian rhythmicity. After the knife cut, a few neurons connect-
ing the SCNs may have remained that could support
coherent circadian rhythmicity. There also may have
been regrowth of the connections between each SCN,
Finally, the SCNs may be coupled by other neural pathways and endocrine feedback loops 9'2° that must first
be disrupted to demonstrate the role of the direct
SCN-SCN connection. Alternatively, the bilateral struc- ture of the SCN may be of no particular functional
significance.
Acknowledgement. This research was supported by NSF Grant BNS 82-17281 to N.T.A. and A.M.R. and Grant NIH 5-T32- 6M07170 to J.A.Y.
SCN SSCN
PX
suprachiasmatic nucleus of the hypothalamus parasagittal knife cut intended to split the bilateral SCN and optic chiasm pinealectomy
a neuroendocrine loop? J. Exp. Zool., 232 (1984) 539-549. 10 Chesworth, M.J., Cassone, V.M. and Armstrong, S.M., Effects
of daily melatonin injections on activity rhythms of rats in constant light, Am. J. Physiol., 253 (1987) R101-107.
11 Cheung, P.W. and McCormack, C., Failure of pinealectomy or melatonin to alter circadian activity rhythm of the rat, Am. J. Physiol., 242 (1982) R261-R264.
12 Dark, J., Partial isolation of the suprachiasmatic nuclei: effects on circadian rhythms of rat drinking behavior, Physiol. Behav., 25 (1980) 863-873.
13 Davis, EC. and Gorski, R.A., Unilateral lesions of the hamster suprachiasmatic nuclei, J. Comp. Physiol., 154 (1984) 221-232.
14 Donaldson, J.A. and Stephan, EK., Entrainment of circadian rhythms: retinofugai pathways and unilateral suprachiasmatic nucleus lesions, Physiol. Behav., 29 (1982) 1161-1169.
15 Eastman, C.I., Mistlberger, R.E. and Rechtshaffen, A., Supra- chiasmatic nuclei lesions eliminate circadian temperature and sleep rhythms in the rat, Physiol. Behav., 32 (1983) 357-368.
16 Finkelstein, J., Baum, E and Campbell, C., Entrainment of the female hamster to reversed photoperiod: role of the pineal, Physiol. Behav., 21 (1978) 105-111.
17 Fuchs, J.L., Effects of pinealectomy and subsequent melatonin implants on activity rhythms in the house finch, J. Comp. Physiol., 153 (1983) 413-419.
18 Gaston, S., The influence of the pineal organ on the circadian activity rhythm in birds. In M. Menaker (Ed.), Biochronometry, Natl. Acad. Sci., Washington, DC, 1971, pp. 541-548.
19 Gold, R.W., Kapatos, G. and Carey, R., A retracting wire knife for stereotaxic brain surgery made from a microliter syringe, Physiol. Behav., 10 (1973) 813-815.
20 Groos, G.A., The neurophysiology of the mammalian suprachi- asmatic nucleus and its visual afferents. In J. Aschoff, S. Daan and G.A. Groos (Eds.), Vertebrate Circadian Systems, Springer,

226
Berlin, 1982. 21 Gwinner, E. and Benzinger, I., Synchronization of a circadian
rhythm in pinealectomized European starlings by daily injections of melatonin, J. Comp. Physiol., 127 (1978) 209-213.
22 Hoffman, R.A. and Reiter, R.J., Rapid pinealectomy in hamsters and other small rodents, Anat. Rec., 153 (1965) 19-22.
23 Kavaliers, M., The pineal organ and circadian organization of teleost fish, Rev. Can. Biol., 38 (1979) 281-292.
24 Kincl, F.A., Chang, C.C. and Buzkova, V., Observations on the influence of changing photoperiod on spontaneous wheel- running activity of neonatally pinealectomized rats, Endocrinol- ogy, 87 (1970) 38-42.
25 Klein, D.C., Smoot, R., Weller, J.L., Niya, S., Markey, S.P., Creed, G.J. and Jacobowitz, D.M., Lesions of the paraventric- ular nucleus area of the hypothalamus disrupt the suprachias- matic-spinal cord circuit in the melatonin rhythm generating system, Brain Res. Bull., 10 (1983) 647-652.
26 Mason, R. and Brooks, A., The electrophysiological effects of melatonin and a putative melatonin antagonist (N-acetyltrypta- mine) on rat suprachiasmatic neurones in vitro, Neurosci. Len., 95 (1988) 296-301.
27 Menaker, M., The search for principles of physiological orga- nization in vertebrate circadian systems. In J. Aschoff, S. Daan and G.A. Groos (Eds.), Vertebrate Circadian Systems, Springer, Berlin, 1982.
28 Moore, R.Y., The anatomy of central neural mechanisms regulating endocrine rhythms. In D.T. Krieger (Ed.), Endocrine Rhythms, Raven Press, New York, 1979.
29 Moore, R.Y., Organization and function of a central nervous system circadian oscillator: the suprachiasmatic hypothalamic nucleus, Fed. Proc., 42 (1983) 2783-2789.
30 Moore-Ede, M.C., The circadian timing system in mammals: two pacemakers preside over many secondary oscillators, Fed. Proc., 42 (1983) 2802-2808.
31 Mosko, S.S. and Moore, R.Y., Neonatal suprachiasmatic nu- cleus lesions, Brain Research, 164 (1979) 17-38.
32 Paxinos, G. and Watson, C., The Rat Brain in Stereotaxic Coordinates, 2nd edn., Academic Press, New York, 1986, 22-24.
33 Pickard, G.E., The afferent connections of the supraehiasmatic nucleus of the golden hamster with emphasis on the retino- hypothalamic projection, J. Comp. NeuroL, 211 (1982) 65-83.
34 Pickard, G.E. and Turek, EW., The suprachiasmatic nuclei: two circadian clocks? Brain Research, 268 (1983) 201-210.
35 Pickard, G.E. and Turek, F.W., Effects of partial destruction of the suprachiasmatic nuclei on two circadian parameters, J. Comp. Physiol. A, 156 (1985) 803-815.
36 Pittendrigh, C.S., Circadian systems: general perspective. In J. Aschoff (Ed.) Biological Rhythms, Plenum, New York, 1981, pp. 57-80.
37 van den Pol, A.N., The hypothalamic suprachiasmatic nucleus of the rat: intrinsic anatomy, J. Comp. Neurol., 191 (1980) 661-702.
38 van den Pol, A.N. and Powley, T., A fine-grained anatomical analysis of the role of the rat suprachiasmatic nucleus in circadian rhythms of feeding and drinking, Brain Research, 160 (1979) 307-326.
39 Quay, W.B., Physiological significance of the pineal during adaptation to shifts in the photoperiod, Physiol. Behav., 5 (1970) 353-360.
40 Quay, W.B., Precocious entrainment and associated character- istics of activity patterns following pinealectomy and reversal of photoperiod, Physiol. Behav., 5 (1970) 1281-1290.
41 Redman, J., Armstrong, S. and Ng, K.T., Free-running activity rhythms in the rat: entrainment by melatonin, Science, 219 (1983) 1089-1091.
42 Rietveld, W.J., The effect of partial lesions of the hypothatamic suprachiasmatic nucleus on the circadian control of behavior, Annu. Rev. Chronopharmacol., 1 (1984) t-4.
43 Richter, C.P., Sleep and activity: their relation to the 24-h clock, Proc. Assoc. Res. Nerv. Ment. Dis., 45 (1967) 8-27.
44 Rosenwasser, A.N, and Adler, N.T., Structure and function in circadian timing systems: evidence for multiple coupled circadian oscillators, Neurosci. Biobehav. Rev., 10 (1986) 431-448.
45 Shibata, S., Cassone, V.M. and Moore, R.Y., Effects of melatonin on neuronal activity in the rat suprachiasmatic nucleus in vitro, Neurosci. Lett., 97 (1989) 140-144.
46 Sisk, C.L. and Turek, EW., Role of the inter-connections of the suprachiasmatic nuclei in the hamster circadian system, Soc. Neurosci. Abstr., 8 (1982) 6.
47 Sokolove, P.G. and Bushell, W.N., The chi square periodogram: its utility for analysis of circadian rhythms, J. Theor. Biol., 72 (1978) 131.
48 Stephan, F.K., Berkeley, K.J. and Moss, R.L., Efferent con- nections of the rat suprachiasmatic nucleus, Neuroscience, 6 (1981) 2625-2641.
49 Stephan, F.K. and Nunez, A.A., Role of retino-hypothalamic pathway in the entrainment of drinking rhythms, Brain Res. Bull., 1 (1976) 495-497.
50 Stephan, F.K. and Nunez, A.A., Developmental plasticity in retinohypothalamic connections and the entrainment of circa- dian rhythms, Behav. Biol., 22 (1978) 77-84.
51 Stephan, EK. and Zucker, I., Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalamic lesions, Proc. Natl. Acad. Sci. U.S.A., 69 (1972) 1583-1586.
52 Takahashi, K., Inoue, K. and Takahashi, Y., No effect of pinealectomy on the parallel shift in circadian rhythms of adrenocortical activity and food intake in blinded rats, Endo- crinol. Japan, 23 (1976) 417-421.
53 Turek, F.K., Circadian neural rhythms in mammals, Annu. Rev. Physiol., 47 (1985) 49-64.
54 Turek, EW., McMillan, J.P. and Menaker, M., Melatonin: effects on the circadian locomotor rhythm of sparrows, Science, 194 (1976) 1441-1443.
55 Underwood, H. and Harless, M., Entrainment of the circadian activity rhythm of a lizard to melatonin injections, Physiol. Behav., 35 (1985) 267-270.
56 Vanecek, J., Melatonin binding sites, J. Neurochem,, 51 (1988) 1436-1440.
57 Zimmerman, N.H. and Menaker, M., The pineal: a pacemaker within the circadian system of the house sparrow, Proc. Natl. Acad. Sci. U.S.A., 76 (1979) 999-1003.





























