Embed Size (px)
Citation preview

Eur. J. Biochem. 211,259-266 (1993) 0 FEBS 1993
The cooperative binding of phenylalanine to phenylalanine 4-monooxygenase studied by H-NMR paramagnetic relaxation Changes in water accessibility to the iron at the active site upon substrate binding
Aurora MARTiNEZ, Sigridur OLAFSDOTTIR and Torgeir FLATMARK Department of Biochemistry, University of Bergen, Norway
(Received July 17/September 23,1992) - EJB 92 1025
The effect of the paramagnetic high-spin Fe(II1) ion in phenylalanine 4-monooxygenase (phenyl- alanine hydroxylase, EC 1.14.16.1) on the water proton longitudinal relaxation rate has been used to study the environment of the iron center. The relaxation rate was measured as a function of the concentration of enzyme, substrate (phenylalanine), inhibitor (noradrenaline) and activator (lysolecithin), asrwell as of the temperature (1 8 - 40 "C) and the external magnetic field strength (100 - 600 MHz). From the frequency dependence of the relaxation rate, an effective correlation time (7,) of 4.2( f0.5) x 10- lo s was calculated for the enzyme-substrate complex, which most likely represents the electron spin relaxation rate (zs) for Fe(II1) ( S = 5/2) in this complex. The relaxation rate was proportional to the concentration of enzyme (0.04 - 1 mM) both in the absence and presence of phenylalanine, but the paramagnetic molar relaxivity at 400 MHz and 22 "C decreased from 2.2(+0.05) x lo3 s - l . M-' i n the enzyme as isolated to 1.2(+0.06) x lo3 s-l . M-' in the presence of saturating concentrations of the substrate. The activation energy of the relaxation rate also decreased from 11.3 k 0.8 kJ/mol to - 1.5 k 0.2 kJ/mol upon incubation of the enzyme with 5 mM phenylalanine. The results obtained can be interpreted in terms of a slowly exchanging water molecule coordinated to the catalytic paramagnetic Fe(II1) in the native and resting enzyme, and that this water molecule seems to be displaced from coordination on the binding of substrate or inhibitor. Moreover, the effect of increasing concentrations of phenylalanine and noradrenaline on the water proton relaxation rate and on the hydrophobic surface properties of the enzyme indicate that substrate and inhibitor induce a similar cooperative conformational change upon binding at the active site. By contrast, the activator lysolecithin does not seem to affect the interaction of water with the catalytic Fe(II1).
Phenylalanine 4-monooxygenase (phenylalanine hy- droxylase) catalyzes the hydroxylation of L-phenylalanine to L-tyrosine in the liver, using (6R)-l-erythro-tetrahydro- biopterin (BH,) as the electron donor (Kaufman and Fisher, 1974). The rat enzyme contains approximately 1 mol iron/mol
Correspondence to A. Martinez, Department of Biochemistry,
Fax: + 47-5-20 6400. Abbreviutions. C, protein concentration; E,, activation energy;
BH4, (6R)-l-erythro-tetrahydrobiopterin; q, number of water ligands coordinated to the paramagnetic ion; T;;, diamagnetic contribution to the longitudinal relaxation rate; T;d, longitudinal paramagnetic relaxation rate of the bound water protons; T;:, paramagnetic contri- bution to the longitudinal relaxation rate; T;d, water proton longi- tudinal relaxation rate in protein-free buffer solutions; Tl,tbsr ob- served water proton longitudinal relaxation rate; T&, outer-sphere contribution to the relaxation rate; z., dipolar correlation time; tM, exchange lifetime; zR, rotational correlation time; zs, electron-spin relaxation time; wI and ws, proton and electron Larmor frequencies, respectively.
Enzymes. Phenylalanine 3-monooxygenase or phenylalanine hy- droxylase (EC 1.14.16.1).
University of Bergen, N-5009 Bergen, Norway
subunit, which is mainly in the ferric form in the enzyme as isolated (Martinez et al., 1991); in the catalytic cycle, this iron is reduced to Fe(I1) by the pterin cofactor (Wallick et al., 1984; Bloom et al., 1986). The holoenzyme (A4 x 200000) is composed of four identical subunits and is subjected to activation (and other types of regulation) by several chemical agents, among which the substrate (phenylalanine) is of par- ticular physiological significance. Cooperativity is manifested both in its rate/concentration curve and its equilibrium bind- ing (Shiman and Gray, 1980; Parniak and Kaufman, 1981), but the underlying mechanism of the activation by the sub- strate is, however, poorly understood. Evidence has recently been presented supporting the proposal that activation occurs by a cooperative binding of the substrate to the active site (Martinez et al., 1990). In the present study we focus on the mechanism of the cooperative binding of phenylalanine, using noradrenaline as a reference compound shown to specifically bind at the active site, with a cooperativity similar to that of phenylalanine (Martinez et al., 1990). Phenylalanine hy- droxylase, as well as the homologous enzyme tyrosine hy- droxylase, is inhibited by catecholamines (dopamine,

260
noradrenaline and adrenaline), which bind by a direct coordi- nation to the Fe(1II) at the active site (Cox et al., 1988; Anderson et al., 1988; Martinez et al., 1991). Thus, catechol- amines can be regarded as probes for the catalytic (iron- binding) site in these enzymes (Martinez et al., 1990; Haavik et al., 1991).
The iron center in rat phenylalanine hydroxylase is in the ferric high-spin state ( S = 5/2) (Wallick et al., 1984; Martinez et al., 3 991), and should induce a large enhancement of water proton relaxation rates if the water in the first and/or second coordination sphere of the iron exchanges with bulk water at a sufficiently fast rate on the NMR time scale (for reviews see Burton et al., 1979; Mildvan et al., 1980, and Banci et al., 1991). Previous measurements of the paramagnetic effect of the iron on the longitudinal relaxation times ( T1) of the aro- matic protons of phenylalanine and noradrenaline have shown that these ligands bind close to the metal, and that the cate- cholamine displaces the substrate from the active site (Martinez et al., 1991). Using this method, we show in the present study that the water accessibility to the iron center is modulated by the binding of phenylalanine, compatible with a cooperative binding of the substrate to the active site.
EXPERIMENTAL PROCEDURES
Materials
L-Noradrenaline, L-phenylalanine, lysolecithin (lysophos- phatidylcholine), catalase and dithiothreitol were purchased from the Sigma Chemical Co. (St. Louis, MO). Phenyl- Sepharose CL4B was obtained from Pharmacia LKB A. B. (Uppsala, Sweden). 6-Methyl-5,6,7,8-tetrahydropterin (6- MPH4) was from Dr. B. Schircks Laboratories (Jona, Switzerland) and 'H,O (99.8%) from Hydro (Norway).
Purification and assay of phenylalanine hydroxylase
Phenylalanine hydroxylase was isolated from rat liver by the method (procedure I1 D) of Shiman et al. (1979), yielding homogeneous enzyme preparations with typical specific activi- ties of 7.2 -+_ 0.3 pmol L-tyrosine . min-l . mg protein-l when assayed at pH 6.8 and 20°C. The enzyme activity was measured spectrophotometrically as described (Shiman et al., 1979). The concentration of purified protein was estimated using an absorption coefficient at 280 nm of 1.0 cm-' for 1 mg/ml (Shiman et al., 1979).
Metal analyses
The metal content of the enzyme preparations was deter- mined by a Perkin-Elmer model 402 atomic absorption spectrophotometer equipped with a graphite furnace (type HGA-76B from Perkin Elmer). The enzyme preparations utilized in these experiments contained 1.1 _+ 0.1 atom iron/ enzyme subunit.
Adsorption of phenylalanine hydroxylase to Phenyl-Sepharose
In order to study the effect of phenylalanine and noradrenaline on the surface hydrophobicity of the enzyme, purified rat phenylalanine hydroxylase (1 30 pg) was incubated for 15 min at 25 "C in 20 mM potassium phosphate pH 7.25, containing 0.2 M KC1 and L-phenylalanine (2 and 20 mM) or L-noradrenaline (0.1 -5 mM), in a final volume of 100 pl.
Controls were incubated in the same buffer, without substrate or inhibitor. Then, 100 p1 (approximately 50 mg dry gel) Phe- nyl-Sepharose CL-4B, equilibrated in the incubation medium, was added and the incubation continued for 10 min at 25°C with gentle stirring. Samples were then centrifuged for 5 min (1000 x g), and the protein concentration in the supernatants was determined as described by Bradford (1976).
Measurements of water proton relaxation rates
Longitudinal relaxation rates of the residual water signal (HDO) were measured on enzyme samples prepared in 'H,O, containing 20 mM potassium phosphate of pH* 7.2 - 7.4 (un- corrected value in 'HzO) and 0.2 M KC1. Deuteration of exchangeable protons of the samples was performed as de- scribed (Martinez et al., 1991). In order to measure the relax- ation rates in non-deuterated samples, the enzyme was pre- pared in 20 mM potassium phosphate pH 7.25, 0.2 M KC1, or, when indicated, in 15 mM Tris/HCl pH 7.25,0.2 M KC1; both buffers were prepared from double-distilled water. The spectra were recorded on a Bruker AM-400 WB spectrometer using internal deuterium lock and standard Fourier-transfor- mation techniques. In experiments with variable field, mea- surements at 100 MHz were made on a Jeol JNM-FX100 spectrometer and at 250, 500 and 600 MHz on Bruker AM- 250, AM-500 and AM-600 spectrometers, respectively. The longitudinal relaxation times (Tl,obs) were measured at the indicated probe temperatures by using a standard inversion- recovery sequence (180 '-z-9Oo), recycle delay 5 x T1. In the titration experiments, the enzyme solution was allowed to equilibrate with the indicated concentrations of L-phenylala- nine, L-noradrenaline or lysolecithin in the NMR tubes for 5 min at room temperature, prior to measurements. Titrations of the enzyme with substrate, inhibitor or activator were performed by adding microliter amounts of concentrated solu- tions; final dilution of the enzyme samples was 12.5%.
Theoretical details
The longitudinal paramagnetic relaxation time of the bound water proton resonance (TIM) is described by the di- polar term of the Solomon-Bloembergen equation (Solomon, 1955; Bloembergen, 1957), i.e.
where yI is the nuclear gyromagnetic ratio, g is the electronic g factor (isotropic splitting factor), p is the Bohr magneton, S is the electronic spin at the ground state of the paramagnetic ion (Wuthrich, 1976), r is the metal-proton internuclear dis- tance, wI and cos are the nuclear and electron Larmor pre- cession frequencies, respectively, and 7, the effective dipolar correlation time, which describes the molecular events which modulate the electron-nuclear dipolar coupling, and is defined as
( 2 ) where zs is the electron-spin relaxation time, T~ is the lifetime of the water protons in the bound state of the paramagnetic complex (exchange lifetime) and 7 R is the rotational corre- lation time of the paramagnetic complex. The value of z, is determined by the fastest rate process and in the present work it was calculated from the frequency dependence of the longi- tudinal paramagnetic relaxation (Peacocke et al., 1969 ; Dwek, 1975).
Tc-l = 7;' + 7 i 1 + 7,'

The observed longitudinal relaxation rate of water protons
(3)
in a protein solution (T;,OIbs), is given by the equation,
TT,ibs = TTp' f T;d where T;p' is the longitudinal relaxation rate due to the para- magnetic ion and T;& is the diamagnetic contribution of the protein, due to the effects of protein residues-solvent interac- tions. The water proton longitudinal relaxation time in pro- tein-free buffer solutions was designated T;;, and its contri- bution is included in T I D . T;; was estimated using sodium- dithionite-treated phenylalanine hydroxylase, in which the iron is fully reduced (Martinez et al., 1991). TTp' values for water protons, normalised to a protein concentration (Cj of 1 M subunit, are expressed as paramagnetic molar relaxivity
The paramagnetic molar relaxivity is related to the longi- tudinal paramagnetic relaxation time of the bound water pro- tons, T I M , by the following expression (Mildvan et al., 1980):
(T;; . C').
where q is the number of water ligands which are coordinated to the Paramagnetic ion (i.e. in the first coordination sphere) and T& is the outer-sphere contribution to the relaxation rate. T& is usually small in paramagnetic systems in which the water is directly coordinated to the iron. However, the paramagnetic contribution to the relaxation can be dominated by outer-sphere effects in systems where water or solvent pro- tons diffuse sufficiently close to the paramagnetic ion, but the ion does not coordinate water at all or only with severe exchange limitations (Koenig et al., 1981). In these cases, T& is dependent on the distance between the water protons and the paramagnetic ion (Eqn 1). The effect of varying tem- perature and frequency on T;: was used to determine the predominant contributions (TIM, zM or To.s.) to the observed relaxation rate (Dwek, 1975).
RESULTS
Effect of rat phenylalanine hydroxylase on the water proton relaxation rates
Proton spin-lattice relaxation rates (T&s) were measured at 400 MHz on the bulk residual water (HDO) signal (4.8 ppm) at 295 K in deuterated samples of phenylalanine hydroxylase at various concentrations (0.04 - 1 mM enzyme subunit), in the absence and presence of substrate (Fig. 1). The T;d values (Eqn 3), obtained with the dithionite-reduced enzyme preparations, were similar to the longitudinal relax- ation rates measured in the protein-free deuterated buffer (T;,&,S = 0.071 s-'; Fig. I), indicating that, when measured at 400 MHz, the diamagnetic contribution due to protein- solvent interactions, has little effect on The large effect of the enzyme concentration on Tl,obs indicates that the high- spin Fe(II1) site is accessible to exchangeable water molecules. The paramagnetic molar relaxivity (T;: . C - l) was found to decrease from 2.2( k0.05) x lo3 M-' . s-l in the native en- zyme to 1.2(f0.06) x lo3 M-' . s-' in the presence of 5 mM phenylalanine. The paramagnetic molar relaxivity of the native enzyme also decreased to a value of 1.3( k0.06) x lo3 M-' . s-' upon binding of the active site inhibitor noradre- naline (Martinez et al., 1991).
Longitudinal relaxation rates were also measured in water- based, non-deuterated solutions of native phenylalanine hy- droxylase and the paramagnetic molar relaxivity was found to be dependent on the buffer utilized to prepare the enzyme
2.5
2.0
1.5 - 1 (D - * - 0
c; 1.0
0.5
0.0 0.0 0.2 0.4 0.6 0.8 1.0 1.2
261
1
[Phonylalanine hydroxylaso subunit] (mM)
Fig. 1. Longitudinal relaxation rate of the residual water signal (HDO) in solutions of phenylalanine hydroxylase. Measurements were made at 295 K and 400 MHz, in solutions of rat phenylalanine hydroxylase as a function of subunit concentration. Native enzyme in 20mM deuterated (99.8% 'HzO) potassium phosphate, pH* 7.2 and 0.2 M KCl (0), with 5 mM phenylalanine ( 0 ) and reduced with 1 mM sodium dithionite (7). The effect of the enzyme concentration on the relaxation rate of the water protons can be expressed by the equation: T& = [2.2(*0.05) . C + 0.0471 s-' and that of the enzyme- substrate complex by T , & = [1.2(f0.06) . c + 0.0451 s-'; c = subunit concentration (mM).
solution. Thus, when phenylalanine hydroxylase was prepared in 20 mM potassium phosphate pH 7.25,0.2 M KCl (T;: = 0.303 s-'), the paramagnetic molar relaxivity (T;: . C- ') obtained at 400 MHz and 295 K in the absence of ligands, i. e. 2 .2(+0.1)~ lo3 M-' . S-' , was found to be the same as that obtained in enzyme samples prepared in deuterated potassium phosphate buffer (see above), which indicates that there is no isotope effect on the TTp' . C - ' values. However, the paramag- netic molar relaxivity measured on enzyme samples prepared in 15 mM Tris/HCl pH 7.25, 0.2 M KC1 (T;: = 0.300 s-') was only l.l(+O.l) x lo3 M - l . s-' , and this value did not decrease in the presence of saturating concentrations of phe- nylalanine. Tris is an inhibitor of the enzyme, competitive to the pteridine cofactor (Marota and Shiman, 1984), and we have recently found that the EPR spectrum of phenylalanine hydroxylase at 3.6 K taken in Tris buffer (axial-type of sig- nals), is different from that obtained in phosphate buffer (rhombic-type signals; Martinez et al., 1991). Moreover, the EPR spectra- taken in potassium phosphate buffer, when noradrenaline is added as an active-site inhibitor, resemble those taken in Tris buffer (Martinez et al., 1991). These results indicate an interaction of Tris with the catalytic Fe(II1) in phenylalanine hydroxylase. The low paramagnetic molar re- laxivity value obtained in Tris buffer, comparable to that obtained with noradrenaline, is in agreement with our pre- vious results and supports the conclusion that Tris binds at the active site. By contrast, phosphate does not seem to bind to the enzyme and has no inhibitory effect on enzyme activity (Marota and Shiman, 1984; Martinez et al., 1991) and it was the buffer used in these studies.

262
I I
3.2 3.3 3.4 3.5
103/T ( k-')
Fig. 2. Temperature dependence of the paramagnetic molar relaxivity. Measurements made at 400 MHz in solutions of rat phenylalanine hydroxylase (60 FM subunit), prepared in 20 mM deuterated potassi- um phosphate pH* 7.2 and 0.2 M KCI, in the absence (0) and in the presence of 5 mM phenylalanine (0). The slopes of linear regression lines yield activation energies of 11.3 0.8 and - 1.5 -t 0.2 kJ/mol, respectively.
Temperature dependence of water proton relaxation rates
The effect of temperature (18 -40°C) on the paramagnetic molar relaxivity of water protons is shown in Fig. 2. The Arrhenius activation energy (E,) for the paramagnetic contri- bution to the relaxation (T;:) decreased from 11.3 f 0.8 kJ/ mol in the native enzyme to - 1.5 f 0.2 kJ/mol upon incu- bation with 5 mM phenylalanine (enzyme-substrate complex). In general, the exchange-lifetime, zM, has positive temperature coefficients, with E, values 2 9 kJ/mol (for review, see Dwek, 1975 and Burton et al., 1979) and typical E, for TC: in slow exchange processes are 2 13 kJ/mol (Peacocke et al., 1969), as observed for methemoglobin (Gupta and Mildvan, 1975; Kaivarainen et al., 1984) and metmyoglobin (Kaivarainen et al., 1984). Thus, our results indicate that the relaxation of water protons in native phenylalanine hydroxylase, in the absence of substrate, is partially exchange-limited, i.e. Z~ is of similar magnitude as TIM or larger, and affects T;: . C-'. However, for the enzyme-substrate complex, the paramagnetic molar relaxivity seems to be independent of the temperature in the range studied (Fig. 2), consistent with a fast exchange of water protons in the paramagnetic environment or with an outer-sphere effect (Dwek, 1975). In this case, the paramag- netic relaxation rate of the water proton resonance is depen- dent on the distance between the exchangeable water mol- ecules and the ferric iron (Dwek, 1975; Koenig et al., 1981).
Field dependence of water proton relaxation rates In solutions of phenylalanine hydroxylase without sub-
strate present, the paramagnetic contribution to the relaxation time (Tlp . C) was found to be nearly frequency-independent, when measured at three different Lannor frequencies, i. e. 250, 400 and 600 MHz and 295 K (Fig. 3), which is consistent with an exchange limited relaxation (see above). For the enzyme- substrate complex the paramagnetic relaxation rate of water protons is frequency-dependent (Fig. 3), which further sup- ports a fast exchange condition of water protons in the para-
1.4
1.2
1 .o 1
u)
0.8 E 0
0.4
0.2
1 I I I I I I
I I I I I I I I 0 2 4 6 8 1 0 1 2 1 4
W,' (x lo-'*) (rad/s>'
Fig. 3. Effect of magnetic field on the water proton paramagnetic relax- ation times. Measurements were performed for the enzyme at 250,400 and 600 MHz (0 ) and at 100, 250, 400, 500 and 600 MHz on the enzyme-substrate complex (0), in samples prepared in 20 mM deuterated potassium phosphate pH* 7.2, containing 0.2 M KCI and 85 pM enzyme subunit (0 ) or 50 pM enzyme subunit and 5 mM phenylalanine (0). The temperature was 295 K. wI is the nuclear Larmor precession frequency. The calculated value o f t , in the en- zyme-substrate complex is 4.20(+0.5) x lo-" s.
magnetic environment (Dwek, 1975). A linear fit of the Tlp . C values at all of the frequencies (two additional measurements were done at 100 and 500 MHz) was used to calculate the effective dipolar correlation time (z,, Eqn l), under the as- sumption that it is constant in this frequency range (Peacocke et al., 1969; Dwek, 1975; Jarori et al., 1989). The value for z, thus obtained is 4.2(f0.5) x lo-'' s. This value should be considered as an approximation since it was calculated from measurements at high field (2 100 MHz), where dispersion of magnetization may occur, especially in high-spin (5/2) Fe(II1) systems (Bertini and Luchinat, 1986).
Effect of the substrate (phenylalanine) and the active-site inhibitor (noradrenaline) on the paramagnetic molar relaxivity and the hydrophobic surface properties of the enzyme
The paramagnetic relaxation rate of the residual water proton resonance is affected by the binding of both substrate and inhibitor to the enzyme, and the effects of ligand concen- tration (Fig. 4) reflect their different affinities [Kd x 0.25 pM for noradrenaline (Martinez et al., 1990) and x 0.2 mM for phenylalanine (Parniak and Kaufman, 1981)l. However, bind- ing parameters (stoichiometry and Kd values) cannot be calcu- lated from these titration curves since several processes seem to contribute to the paramagnetic relaxation rates obtained at each ligand concentration (see below). Both titration curves are very similar in the sense that they are three-phasic and non-hyperbolic. This is in contrast to previous observations on non-cooperative enzymes where this type of titration curves are found to be essentially hyperbolic (Mildvan et al., 1980; Sellin et al., 1982; Bhargava and Chatterji, 1988). When the titration was carried out in non-deuterated buffer and the paramagnetic molar relaxivity values were measured in the
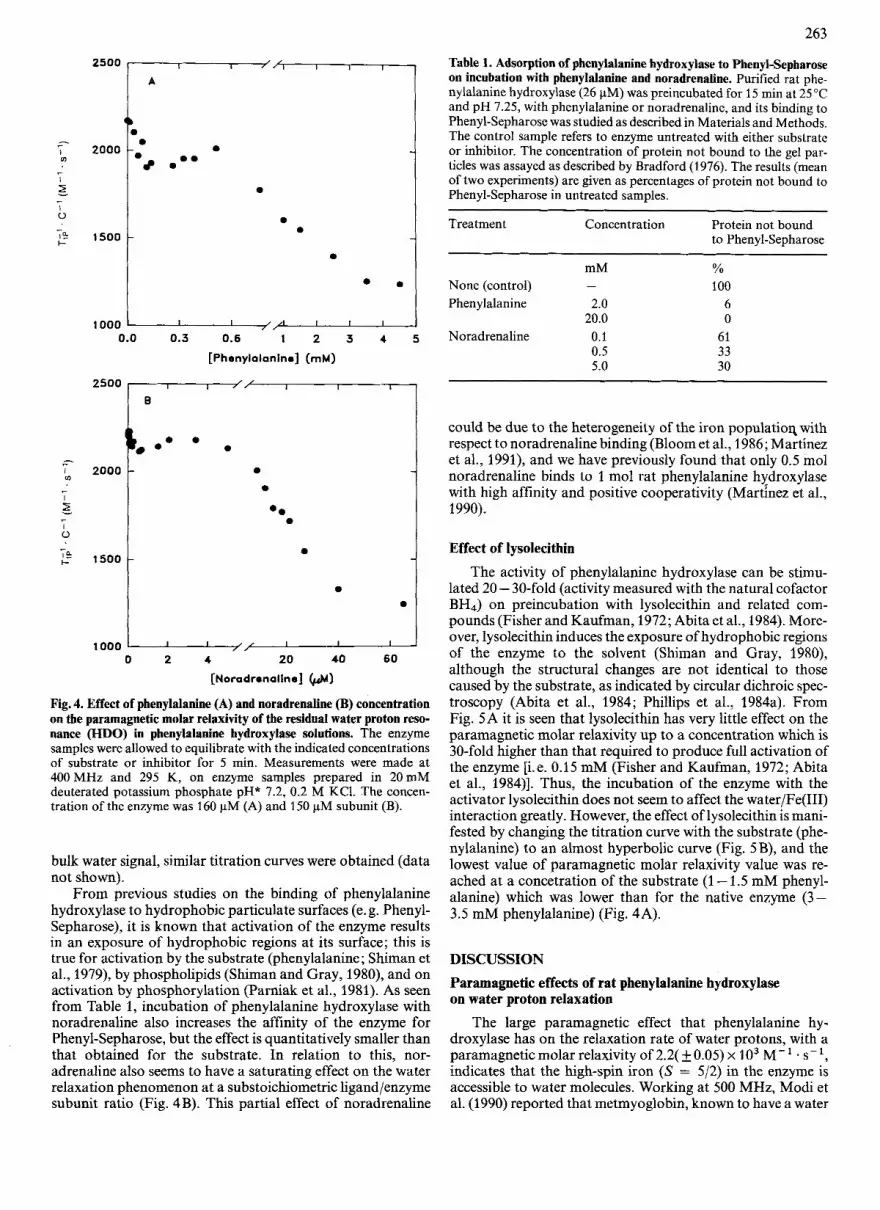
263
c
1000 I,~,-l 0.0 0.3 0.6 1 2 3 4 5
[Phanylalanina] (mM)
9 1500
1000 0 2 4 20 40 60
[Noradrenalina] (@)
Fig. 4. Effect of phenylalanine (A) and noradrenaline (B) concentration on the paramagnetic molar relaxivity of the residual water proton reso- nance (HDO) in phenylalanine hydroxylase solutions. The enzyme samples were allowed to equilibrate with the indicated concentrations of substrate or inhibitor for 5 min. Measurements were made at 400 MHz and 295 K, on enzyme samples prepared in 20 mM deuterated potassium phosphate pH* 7.2, 0.2 M KC1. The concen- tration of the enzyme was 160 pM (A) and 150 pM subunit (B).
bulk water signal, similar titration curves were obtained (data not shown).
From previous studies on the binding of phenylalanine hydroxylase to hydrophobic particulate surfaces (e. g. Phenyl- Sepharose), it is known that activation of the enzyme results in an exposure of hydrophobic regions at its surface; this is true for activation by the substrate (phenylalanine; Shiman et al., 1979), by phospholipids (Shiman and Gray, 1980), and on activation by phosphorylation (Parniak et al., 1981). As seen from Table 1, incubation of phenylalanine hydroxylase with noradrenaline also increases the affinity of the enzyme for Phenyl-Sepharose, but the effect is quantitatively smaller than that obtained for the substrate. In relation to this, nor- adrenaline also seems to have a saturating effect on the water relaxation phenomenon at a substoichometric ligand/enzyme subunit ratio (Fig. 4 B). This partial effect of noradrenaline
Table 1. Adsorption of phenylalanine hydroxylase to Phenyl-Sepharose on incubation with phenylalanine and noradrenaline. Purified rat phe- nylalanine hydroxylase (26 pM) was preincubated for 15 min at 25 "C and pH 7.25, with phenylalanine or noradrenaline, and its binding to Phenyl-Sepharose was studied as described in Materials and Methods. The control sample refers to enzyme untreated with either substrate or inhibitor. The concentration of protein not bound to the gel par- ticles was assayed as described by Bradford (1976). The results (mean of two experiments) are given as percentages of protein not bound to Phenyl-Sepharose in untreated samples.
Treatment Concentration Protein not bound to Phenyl-Sepharose
mM None (control) -
Phenylalanine 2.0 20.0
Noradrenaline 0.1 0.5 5.0
% 100
6 0
61 33 30
could be due to the heterogeneity of the iron populatioq with respect to noradrenaline binding (Bloom et al., 1986; Martinez et al., 1991), and we have previously found that only 0.5 mol noradrenaline binds to 1 mol rat phenylalanine hydroxylase with high affinity and positive cooperativity (Martinez et al., 1990).
Effect of lysolecithin
The activity of phenylalanine hydroxylase can be stimu- lated 20 - 30-fold (activity measured with the natural cofactor BH4) on preincubation with lysolecithin and related com- pounds (Fisher and Kaufman, 1972; Abita et al., 1984). More- over, lysolecithin induces the exposure of hydrophobic regions of the enzyme to the solvent (Shiman and Gray, 1980), although the structural changes are not identical to those caused by the substrate, as indicated by circular dichroic spec- troscopy (Abita et al., 1984; Phillips et al., 1984a). From Fig. 5A it is seen that lysolecithin has very little effect on the paramagnetic molar relaxivity up to a concentration which is 30-fold higher than that required to produce full activation of the enzyme [i.e. 0.15 mM (Fisher and Kaufman, 1972; Abita et al., 1984)l. Thus, the incubation of the enzyme with the activator lysolecithin does not seem to affect the water/Fe(III) interaction greatly. However, the effect of lysolecithin is mani- fested by changing the titration curve with the substrate @he- nylalanine) to an almost hyperbolic curve (Fig. 5B), and the lowest value of paramagnetic molar relaxivity value was re- ached at a concetration of the substrate (1 - 1.5 mM phenyl- alanine) which was lower than for the native enzyme (3- 3.5 mM phenylalanine) (Fig. 4A).
DISCUSSION Paramagnetic effects of rat phenylalanine hydroxylase on water proton relaxation
The large paramagnetic effect that phenylalanine hy- droxylase has on the relaxation rate of water protons, with a paramagnetic molar relaxivity of 2.2( kO.05) x lo3 M-' . s-', indicates that the high-spin iron ( S = 5/2) in the enzyme is accessible to water molecules. Working at 500 MHz, Modi et al. (1990) reported that metmyoglobin, known to have a water

264
2500
1
7* 2000
- I
E .- I 0
2 1500 -
1000
-- A
-',-
0 .
0
1000 0.0 0.5 1 .o 1.5 2.0 2.5
[ P heny la la nine] (m M)
Fig. 5. Variations in the paramagnetic molar relaxivity of the residual water proton resonance (HDO) with increasing concentrations of lysolecithin (A) and with increasing concentrations of phenylalanine in the presence of 5 mM lysolecithin 9). The enzyme samples were al- lowed to equilibrate with the indicated concentrations of lysolecithin or phenylalanine for 5 min. Measurements were made at 400 MHz and 295 K in 20mM deuterated potassium phosphate pH* 7.2, 0.2 M KC1. The concentration of the enzyme was 120 pM subunit.
molecule in the first coordination sphere of the ferric ion, had a paramagnetic molar relaxivity of about 3 x lo3 M-' . s-' at neutral pH. Other high-spin iron paramagnetic proteins such as horseradish peroxidase, lactoperoxidase, and cytochrome c at neutral pH, in which water is proposed to be in the outer sphere of the Fe(III), i.e. not coordinated, had paramagnetic molar relaxivities around 0.5 - 1.5 x lo3 M- ' . s - l (Lanir and Schejter, 1975; Emptage et al., 1981; Modi et al., 1990), when measured at high field. When measured at lower magnetic fields (0 - 100 MHz), somewhat higher values of molar relaxivity have been found for paramagnetic proteins containing coordinated water in the native state (Lanir and Aviram, 1975; Que et al., 1977; Koenig and Brown, 1983). A 1.5 - 3-fold reduction in paramagnetic molar relaxivity has been reported upon binding of ligands known to displace the bound water (Hershberg and Chance, 1975; Que et al., 1977;
Mildvan et al., 1980), except for the special cases of fluorornethemoglobin and other proteins where an anion re- places the water which is liganded to the Fe(II1) (Gupta and Mildvan, 1975; Koenig and Brown, 1983). In the present study we have also found a reduction of the paramagnetic molar relaxivity from 2 . 2 ~ lo3 M-' . s-' to 1 . 2 ~ 1 0 ~ M- ' . s- l upon incubation of the enzyme with the substrate phenylala- nine and the inhibitor noradrenaline. Since the binding of phenylalanine or noradrenaline to the enzyme does not affect the spin state of the Fe(II1) (Martinez et al., 1991), the decrease in paramagnetic molar relaxivity upon binding of these ligands cannot be explained by a high-spin to low-spin transition of the iron.
Estimation of the Fe(II1) - water proton internuclear distances in phenylalanine hydroxylase and the enzyme-substrate complex
The paramagnetic molar relaxivity in the enzyme-substrate complex seems to be essentially independent of temperature (Fig. 2), which is consistent with a fast exchange of water molecules close to the paramagnetic center with those of bulk water. This is further confirmed by the observed frequency dependence of the relaxation rates (Fig. 3), which yields a dipolar correlation time (7,) = 4.2(f0.5) x lo-'' s . As seen from Eqn (2), the effective z, is determined by the fastest of the individual components zs, zM and zR. The value of zR may be estimated from Stokes' law for the rotation of a rigid sphere in a viscous medium as x2.7 x 10- l3 x the relative molecular mass (Mildvan et al., 1980), giving a value of 5.4 x lo-' s for phenylalanine hydroxylase (MR x 200000), and z M has been shown to be > lov9 s (Mildvan and Cohn, 1970; Burton et al., 1979; Mildvan et al., 1980). Thus, the correlation time in the enzyme-substrate complex seems to be dominated by zs, the electron-spin relaxation time for the high-spin Fe(II1) center (Wiithrich, 1976; Mildvan et al., 1980), in agreement with the temperature independence and the strong frequency dependence of TIP for this complex (Gupta and Mildvan, 1975; Dwek, 1975; Jarori et al., 1989). This zs value has been estimated under the assumption that it is frequency- independent in the frequency range studied (Peacocke et al., 1969; Dwek, 1975). This is probably a valid assumption since there is no evidence for a dependence of zs on frequency for Fe(II1) proteins (Bertini and Luchinat, 1986; Lanir and Schejter, 1975), in contrast to the magnetic field dependence found for zs of Fe(II1) in the hexaaqua complex (Levanon et al., 1970; Poupko and Luz, 1972).
Both the temperature dependence (Fig. 2) and the fre- quency independence (Fig. 3) of the paramagnetic contri- bution to the relaxivity indicate that the relaxation rates are limited by the exchange of water in native phenylalanine hy- droxylase. This implies that only upper limits can be estimated for the metal - water proton internuclear distance in the native resting enzyme. By using the paramagnetic molar relaxivity obtained (2.2 x lo3 M-' . s-' ) and assuming that this value is due to the coordination of one molecule to the paramagnetic ion (i.e. q = 1, Eqn 4), TIM is calculated to be 58.17 x s and the estimated distance between the iron and the water protons (averaged distance for the two protons) is 20.35 nm (Eqn 1). However, the interaction with the substrate could affect zs, and consequently z, (Gupta and Mildvan, 1975; Burton et al., 1979), and a different z, value than that in the enzyme-substrate complex may determine the paramagnetic relaxation in the enzyme. A z, value of lO-'Os has been obtained for several high-spin iron proteins containing heme (Gupta and Mildvan, 1975; Wuthich, 1976) and non-heme

265
iron, such as transferrin (Bertini and Luchinat, 1986). By using z, = lo-'' s, the iron-proton distance was calculated to be 10.27 nm. The crystallographic distance from Fe(II1) to the proton of a coordinated water ligand in hemoglobin is 0.26
0.01 nm (Perutz, 1970; Evans and Brayer, 1990). Assuming that more than one water molecule are coordinated to Fe(II1) (i. e. q > l), the calculated distances are too long to be due to direct water-metal coordination. Therefore, our results suggest that in phenylalanine hydroxylase as isolated, the enhance- ment of water proton magnetic relaxation rates is due to one water molecule being directly coordinated to the Fe(II1) center (inner sphere effects).
In the case of the phenylalanine-hydroxylase - phenylala- nine complex, in which water is in a fast exchange condition in the paramagnetic environment (see above), the distance between the water protons and Fe(II1) is estimated to be about 0.39 nm (Eqn 1). Thus, the binding of substrate to the enzyme seems to increase the water - iron distance by 2 11 YO (2 31 YO if a z, value of lo-'' s is used for the enzyme without phenyl- alanine), indicating a displacement of the coordinated water, the remaining paramagnetic enhancement of solvent proton relaxation rates being due to outer sphere effects (Dwek, 1975; Goldberg et al., 1980). Accordingly, Kohzuma et al. (1989) have shown by comparison of the EPR spectrum of phenylala- nine hydroxylase from Chromobacterium violaceum and those of model compounds, that the catalytic Cu(I1) is coordinated to a displaceable water molecule. We have also considered the possibility that the decrease in paramagnetic molar relaxivity, observed upon binding of the substrate to the enzyme, could be due to an entrapment or occlusion of the coordinated water molecule and not to its displacement, i. e. the exchange-lifetime for water protons is increased upon substrate binding and the exchange is so slow on the NMR time scale that the bound water molecule does not contribute to the observed paramag- netic molar relaxivity (Burton et al., 1979). In this case, the paramagnetic enhancement of solvent proton relaxation rates should also be due to outer sphere effects (Koenig et al., 1981). This possibility, however, seems to be less likely in view of the EPR (Wallick et al., 1984) and 57Fe Mossbauer spectroscopic studies (Davis and Kaufman, 1986), showing that phenylala- nine alters the coordination geometry of the catalytic iron in phenylalanine hydroxylase. Thus, displacement of water could be due to the rearrangement of the coordination environment of the metal upon substrate binding which could be ac- companied by enzyme activation (i. e. a change from six-coor- dinated iron to five-coordinated iron). Although a direct chelation or coordination to the iron does not seem to be a likely possibility for the binding of phenylalanine, the results obtained with the active-site inhibitor noradrenaline are interesting. Thus, at saturating concentrations of noradrenaline, the paramagnetic molar relaxivity of the en- zyme solution is also reduced to a similar value to that obtained with the substrate (Fig. 4). Catecholamines have been shown to coordinate to the Fe(1II) of the aromatic amino acid hydroxylases by a catechol to iron charge-transfer inter- action (Cox et al., 1988; Andersson et al., 1988), and displace- ment of coordinated water would be expected upon their binding (Whittaker and Lipscomb, 1984; Arciero et al., 1985). However, noradrenaline could not displace water from all the iron centers, since it binds to the enzyme with a ligandlenzyme ratio lower than one (Fig. 4; Martinez et al., 1990), whereas phenylalanine seems to bind to all the subunits (Parniak and Kaufman, 1981; Phillips et al., 1984b). Therefore, the results rather suggest that, when correcting for differences in the number of subunits occupied by ligand in each case, a higher
outer-sphere contribution occurs in the enzyme-substrate complex than in the tight inhlbitor-enzyme complex, in- dicating that water can diffuse closer to the metal in the former complex.
Effect of cooperative binding of phenylalanine and noradrenaline on the water proton relaxation rate
The phenylalanine hydroxylase holoenzyme is composed of four identical subunits and is subject to allosteric regulation and activation by several chemical agents, among which phe- nylalanine is of particular physiological interest (for review, see Martinez et al., 1990). In steady-state kinetics phenylala- nine hydroxylase exhibits a nonhyperbolic (sigmoidal) depen- dence of activity on substrate concentration, which reflects a slow transition (minutes) of the enzyme from a low-activity to a high-activity state, characteristic of a hysteretic enzyme. Experimental evidence has recently been presented in support of a model in which the cooperative binding of substrate/ inhibitor occurs at the active site (Martinez et al., 1990) and not to a putative regulatory site (Shiman, 1980; Kaufman and Mason, 1982; Shiman et al., 1990). The 'H-NMR data presented here (Fig. 4) further support this proposal. Acti- vation of the enzyme by its substrate is manifested as a trans- formation from a system in which water is coordinated to Fe(II1) at the active site and slowly exchanging with the bulk water, to a system in which water is rapidly exchanging at a site close to the iron, but not coordinated. At concentrations of phenylalanine <0.1 mM, which is the concentration of substrate that activates the enzyme by 50% at pH 6.8 and 25 "C (Shiman and Gray, 1980), phenylalanine decreases the paramagnetic molar relaxivity in a similar way to low concen- trations of the active-site inhibitor noradrenaline (< 1 pM), indicating that the substrate also binds at the catalytic site. At higher concentrations of phenylalanine (0.1 -0.6 mM) and noradrenaline (1 - 4 pM), the paramagnetic molar relaxivity increases, which is most likely related to the cooperative bind- ing of the two ligands, resulting in a conformational change in which more subunits are in an open conformation than those occupied by ligand. An open conformation of the sub- units eliminates the exchange limitations characteristic of the resting non-liganded enzyme (see discussion above) and, therefore, the paramagnetic molar relaxivity of the enzyme increases. At even higher concentrations of phenylalanine (> 0.6 mM) and noradrenaline (> 4 KM), the relaxivity pro- gressively decreases due to an increasing number of subunits with water displaced from coordination to Fe(II1) at the active site. It is also interesting to note that the conformational change of the enzyme induced by noradrenaline seems to be qualitatively similar to that obtained by phenylalanine, as indicated by the increase in its hydrophobic surface properties on ligand binding (Table 1). By contrast, binding of lysolecithin, which is also a very potent activator of phenylala- nine hydroxylase (Fisher and Kaufman, 1972; Abita et al., 1984), does not seem to displace water coordinated to Fe(II1) in the resting enzyme (Fig. 5A), although it induces an ex- posure of hydrophobic regions of the enzyme (Shiman and Gray, 1980). Furthermore, the conformational changes in- duced by lysolecithn results in an essentially hyperbolic ti- tration curve for phenylalanine (Fig. 5 B), characteristic of enzymes with non-cooperative binding of ligands (Mildvan et al., 1980; Sellin et al., 1982; Bhargava and Chatterji, 1988).
We are very grateful to Professors Hans Grasdalen, Steffen B. Petersen and Albert S. Mildvan for their contribution to the NMR

measurements at variable field and their valuable comments and discussions. We also wish to thank to Dr. Jan Haavik and Prof. Einar Sletten for helpful discussions. This work was supported by grants from the Norwegian Research Council for Science and the Humanities and the Royal Norwegian Council for Scientific and Industrial Re- search.
qEFERENCES Abita, J.-P., Parniak, M. & Kaufman, S. (1984) J . Biol. Chem. 259,
Andersson, K. K., Cox, D. D., Que, L., Jr., Flatmark, T. & Haavik, J. (1988) J . Biol. Chem. 263, 18621 -18626.
Arciero, D. M., Orville, A. M. & Lipscomb, J. D. (1985) J . Biol. Chem. 260, 14035- 14044.
Banci, L., Bertini, I. & Luchinat, C. (1991) in Nuclear and electron relaxation: the magnetic nucleus-unpaired electron coupling in solu- tion, VCH Verlagsgesellschaft mbH, Weinheim.
Bertini, I. & Luchinat, C. (1986) in NMR of paramagnetic molecules in biological systems, pp 47 - 83, The Benjamin/Cummings Pub- lishing Company, California.
14560- 14566,
Bhargava, P. & Chatterji, D. (1988) FEBS Lett. 241, 33-37. Bloembergen, N. (1957) J . Chem. Phys. 27, 572-573. Bloom, L. M., Benkovic, S. J. & Gaffney, B. J. (1986) Biochemistry
Bradford, M. (1976) Anal. Biochem. 72,248-254. Burton, D. R., Forsen, S., Karlstrom, G. & Dwek, R. A. (1979) Prog.
Nuclear Magn. Reson. Spectrosc. 13, 1-45. Cox., D. D., Benkovic, S. J., Bloom, L. M., Bradley, F. C., Nelson,
M. J., Que, L. Jr & Wallick, D. E. (1988) J. Am. Chem. SOC. 106,
25,4204-4210.
2026- 2032. Davis, M. D. & Kaufman, S. (1986) Biophys. J. 49, 532a. Dwek, R. A. (1975) in Nuclear magnetic resonance in biochemistry:
applications to enzyme systems, ch. 9 - 11, Oxford University Press, Oxford.
Emptage, M. H., Xavier, A. V., Wood, J. M., Alsaadi, B. M., Moore, G. R., Pitt, R. C., Williams, R. J. P., Ambler, R. P. & Bartsch, R. G. (1981) Biochemistry 20, 58-64.
Evans, S. V. & Brayer, G. D. (1990) J . Mot. Biof. 213, 885-897. Fisher, D. B. & Kaufman S. (1972) J . Biol. Chem. 247,2250-2252. Goldberg, M., Vuk-Pavlovic, S. & Pecht, I. (1980) Biochemistry 19,
Gupta, R. K. & Mildvan, A. S. (1975) J. Biol. Chem. 250,246-253. Haavik, J., Le Bourdelles, B., Martinez, A., Flatmark, T. & Mallet,
Hershberg, R. D. & Chance, B. (1975) Biochemistry 14, 3885-3891. Jarori, G. K., Ray, B. D. & Rao, B. D. N. (1989) Biochemistry 28,
Kaivarainen, A. S., Goryunov, A. S. & Sukhanova, G. (1984) Folia Biol. (Prague) 30,396-403.
Kaufman, S. & Fisher, D. (1974) in Molecular mechanisms of oxygen activation (Hayaishi, O., ed.) pp. 285-369, Academic Press, New York.
Kaufman, S. & Mason, K. (1982) J. Biol. Chem. 257,14667-14678. Koenig, S . & Brown, R. D. I11 (1983) in The coordination chemistry
of metalloenzymes (Bertini, I., Drago, R. S. & Luchinat, C., eds.) pp. 19-33, D. Reidel Publishing Company, Dordrecht.
5181-5189.
J. (1991) Eur. J. Biochem. 199, 371 -378.
9343 - 9350.
Koenig, S. H., Brown, R. D. I11 & Lindstrom, T. R. (1981) Biophys.
Kohzuma, T., Masuda, H. & Yamauchi, 0. (1989) J . Am. Chem. SOC.
Lanir, A. & Aviram, I. (1975) Arch. Biochem. Biophys. 166,439-445. Lanir, A. & Schejter, A. (1975) Biochem. Biophys. Res. Commun. 62,
Levanon, H., Stein, G. & Luz, Z. (1970) J. Chem. Phys. 53, 876-
Marota, J. J. A. & Shiman, R. (1984) Biochemistry 23, 1303 - 131 1. Martinez, A,, Haavik, J. & Flatmark, T. (1990) Eur. J . Biochem. 193,
Martinez, A., Andersson, K. K., Haavik, J. & Flatmark, T. (1991)
Mildvan, A. S . & Cohn, M. (1970) Adv. Enzymol. 33, I -70. Mildvan, A. S., Granot, J., Smith, G. M. & Liebman, M. N. (1980)
in Methods for determining metal ion environments in proteins: structure and function of metalloproteins (Darnall, D. W. & Wilkins, R. G., eds) pp. 211 -236, Elsevier North Holland, New York.
Modi, S., Behere, D. V. & Mitra, S. (1990) J . Inorg. Biochem. 38,17 - 25.
Parniak, M. A. & Kaufman, S. (1981) J . Biol. Chem. 256, 6876- 6882.
Parniak, M. A., Hasegawa, H., Wilgus, H. & Kaufman, S. (1981) Biochem. Biophys. Res. Commun. 99,707 - 714.
Peacocke, A. R., Richards, R. E. & Sheard, B. (1969) Mol. Phys. 16,
Perutz, M. F. (1970) Nature 228, 726-739. Phillips, R. S., Parniak, M. A. & Kaufman, S. (1984a) Biochemistry
Phillips, R. S., Parniak, M. A. & Kaufman, S. (1984b) J . Biol. Chem.
Poupko,R.&Luz,Z.(1972)J. Chem. Phys.57,3311-3318. Que, L. Jr, Lipscomb, J. D., Munck, E. & Wood, J. M. (1977) Biochim.
Sellin, S . , Goran Eriksson, L. E. & Mannervik, B. (1982) Biochemistry
Shiman, R. (1980) J . Biol. Chem. 255,10029- 10032. Shiman, R. & Gray, D. W. (1 980) J . Biof. Chem. 255,4793 - 4800. Shiman, R., Gray, D. W. & Pater, A. (1979) J . Biol. Chem. 254,
Shiman, R., Jones, S. H. & Gray, D. W. (1990) J . Biol. Chem. 265,
Solomon, I. (1955) Phys. Rev. 99, 559-565. Wallick, D. E., Bloom, L. M., Gaffney, B. J. & Benkovic, S. J. (1984)
Biochemistry 23, 1295 - 1302. Whittaker, J. W. & Lipscomb, J. D. (1984) J. Biol. Chem. 259,4487-
4495. Wuthrich, K. (1976) in NMR in biological research: peptides and
proteins, pp. 221 - 292. North-Holland Publishing Company, Amsterdam.
J. 34,397-408.
I l l , 3431-3433.
199 - 203.
887.
211-219.
Eur. J . Biochem. 198,675 - 682.
177 - 189.
23,3836- 3842.
259, 271 - 277.
Biophys. Acta 485, 60 - 74.
21,4850-4857.
11 300- 11 306.
11 633 - 11 642.





























