Embed Size (px)
Citation preview

Author's personal copy
The cytoplasmic domain of neuropilin-1 regulates focal adhesionturnover
Himabindu Reddy Seerapu a, Susmita Borthakur b, Nathan Kong a, Sudesh Agrawal a, Judy Drazba c,Amit Vasanji d, Alessandro Fantin e, Christiana Ruhrberg e, Matthias Buck b, Arie Horowitz a,b,⇑a Department of Molecular Cardiology, Lerner Research Institute, the Cleveland Clinic, Cleveland, OH 44195, United Statesb The Department of Physiology and Biophysics, Case Western Reserve University, Cleveland, OH 44106, United Statesc Imaging Core Facility, Lerner Research Institute, the Cleveland Clinic, Cleveland, OH 44195, United Statesd Image-IQ, Cleveland, OH 44106, United Statese University College London, Institute of Ophthalmology, London, UK
a r t i c l e i n f o
Article history:Received 2 August 2013Accepted 26 August 2013Available online 8 September 2013
Edited by Veli-Pekka Lehto
Keywords:Neuropilin-1Cytoplasmic domainFilamin AFocal adhesion
a b s t r a c t
Though the vascular endothelial growth factor coreceptor neuropilin-1 (Nrp1) plays a critical role invascular development, its precise function is not fully understood. We identified a group of novelbinding partners of the cytoplasmic domain of Nrp1 that includes the focal adhesion regulator, Filam-in A (FlnA). Endothelial cells (ECs) expressing a Nrp1 mutant devoid of the cytoplasmic domain (nrp1cyto-D/D) migrated significantly slower in response to VEGF relative to the cells expressing wild-type Nrp1(nrp1+/+ cells). The rate of FA turnover in VEGF-treated nrp1cytoD/D ECs was an order of magnitude lowerin comparison to nrp1+/+ ECs, thus accounting for the slower migration rate of the nrp1cytoD/D ECs.
Structured summary of protein interactions:NRP1 physically interacts with alpha enolase, Myh10, Myh9, EEF1alpha1 and FlnA by anti bait coimmu-noprecipitation (View interaction)FlnA and NRP1 colocalize by fluorescence microscopy (View interaction)NRP1 and rab11 colocalize by fluorescence microscopy (View interaction)NRP1 physically interacts with Myh10, Dync1h1, Myh9 and EEF1alpha1 by anti bait coimmunoprecipi-tation (View interaction)NRP1 and FlnA bind by isothermal titration calorimetry (View interaction)NRP1 physically interacts with p130Cas by anti bait coimmunoprecipitation (View interaction)NRP1 and p130Cas colocalize by fluorescence microscopy (View interaction)NRP1 binds to FlnA by surface plasmon resonance (View interaction)
� 2013 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
1. Introduction
Nrp is a single-pass transmembrane protein, which has a highlyconserved short cytoplasmic domain that lacks catalytic activity[1,2]. While the downstream signaling of the tyrosine kinase recep-tor of vascular endothelial growth factor (VEGF)-A, VEGF receptor 2(VEGFR2), has been the subject of numerous studies and its majorelements have been established [3], the contribution of Nrp1 toVEGF signaling is less well understood. Several studies suggestedthat Nrp1 might have a VEGFR2-independent function in mediat-ing VEGF signaling [4–7]. Indeed, Nrp1 has binding partners other
than VEGFR2, such as receptors of the plexin family [8], but it re-mains unclear how the cytoplasmic domain of Nrp1 mediates sig-naling. Presumably, the cytoplasmic domain could recruit otherproteins that may be able to transduce VEGF signaling. The beststudied Nrp1 cytoplasmic binding protein is synectin, also namedNrp1-interacting protein (NIP) [9]. Synectin contains a single PDZ(postsynaptic density 95, disk large, zona occludens-1) domain,through which it binds to the C-terminus of Nrp1. Synectin recruitsthe molecular motor myosin VI to uncoated endocytic vesicles,thus facilitating the trafficking of endocytosed membrane recep-tors [10], such as VEGFR2 in complex with Nrp1, to promote arte-riogenesis [11].
Focal adhesions are large aggregates of proteins that anchor thecell to the extracellular matrix and that undergo cycles of assemblyand disassembly during cell movement [12,13]. Among other sig-nals, the dynamics of FAs are regulated by growth factors [14],
0014-5793/$36.00 � 2013 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.http://dx.doi.org/10.1016/j.febslet.2013.08.040
⇑ Corresponding author at: Department of Molecular Cardiology, Lerner ResearchInstitute, the Cleveland Clinic, Cleveland, OH 44195, United States. Fax: +1 216 4458204.
E-mail addresses: [email protected], [email protected] (A. Horowitz).
FEBS Letters 587 (2013) 3392–3399
journal homepage: www.FEBSLetters .org

Author's personal copy
including VEGF-A [15], a major stimulant of vasculogenesis andangiogenesis during development and in the adult organism[16,17]. Treatment with VEGF increased FA density in ECs [18].The signaling of VEGF to FAs appeared to follow the canonical VEG-FR2-dependent pathway, and to be mediated by the non-receptorFA kinase (FAK), and by a closely related kinase, Pyk2 [19] TheVEGF-induced increase in FA assembly was accompanied by an in-crease in EC migration. Though Nrp1 had not been observed di-rectly in FAs [19], proteomics studies identified the VEGF co-receptor Nrp1 as a FA component [20].
The regulation of FA turnover is not fully understood, and it islikely to involve multiple molecular mechanisms [13]. One ofthese invoked FlnA [18], a large scaffold protein that binds fila-mentous actin (F-actin) and numerous other proteins, includingtransmembrane receptors [21,22]. FlnA is composed of 24 IgG re-peats including a C-terminal dimerization domain [21]. Most ofits ligands, excluding F-actin, bind to the last three C-terminalIgG repeats.
Here, we investigated if the Nrp1-mediated intracellular signal-ing of VEGF originates from a protein complex bound to the cyto-plasmic domain of Nrp1. We first combined immunoprecipitationand mass spectroscopy to identify several novel Nrp1-associatedproteins, including FlnA. Separately, we found by immunoprecipi-tation and immunoblotting that the scaffold protein p130Cas alsoassociates with Nrp1. Both proteins are known to be involved inthe dynamics of FAs [23]. We tested, therefore, if Nrp1 is involvedin FA turnover. Our results support this premise.
2. Materials and methods
See Supplemental materials.
3. Results
3.1. Identification of novel ligands to the cytoplasmic domain of Nrp1
We used a proteomics approach to identify cytoplasmic pro-teins that associate with Nrp1. Immunoprecipitates of Nrp1 from
quiescent and from VEGF-A164-treated (20 ng/ml, 5 min) primarymouse heart ECs were resolved by SDS–PAGE. The band patternsof the two samples were partially different (Fig. 1ote, Table 1).The proteins were identified by liquid chromatography–tandemmass spectroscopy (LC–MS). In the quiescent ECs, these proteinswere cytoplasmic dynein heavy chain 1 (Dync1h1), myosin heavychains 9 and 10 (Mhy9, Mhy10), and eukaryotic translation elonga-tion factor 1 a1 (EEF1a1). Mhy9, Mhy10, and EEF1a1 were presentin the immunoprecipitate from both VEGF-treated and quiescentECs. Mhy9 and Myh10 are subunits of non-muscle myosin II,known also as heavy chains IIa and IIb. They are involved in endo-somal trafficking [24], cytokinesis [25], and cell shape remodeling[26], and are localized to early endosomes, stress-fibers, and FAs[27]. The proteins unique to the VEGF-treated sample were a-eno-lase and Filamin-A (FlnA). Due to its connection to the actin cyto-skeleton and protein transport, the involvement of FlnA wasinvestigated further.
3.2. FlnA binds Nrp1 directly
Given that FlnA binds directly several classes of transmembranereceptors [21,22], we sought to determine if the association of FlnAto Nrp1 is similarly direct. As shown by the MS analysis, we iden-tified Nrp1 as an FlnA-associated protein by co-immunoprecipita-tion from the lysate of nrp1+/+ ECs (Fig. 1). We used twoindependent in vitro approaches to test if FlnA and Nrp1 are capa-ble to bind directly to each other. First, we used surface plasmonresonance (SPR) to determine the equilibrium binding kinetics be-tween FlnA and the Nrp1 cytoplasmic domain. FlnA is a modularprotein consisting of 24 immunoglobulin-like (Ig) repeats, whichbind several proteins including F-actin. We tested the interactionof Nrp1 cytoplasmic region with the most common binding sitesof FlnA, the repeats 10–11, 18, 19, 20, 21, 22 and 23–24 of FlnAand found that only repeats 23–24 of FlnA associates with theNrp1 cytoplasmic region with an appreciable affinity (KD of9.6 ± 1.2 lM) (Fig. 2A–C). This was confirmed by isothermal titra-tion calorimetry, which showed a closely similar dissociation con-stant of 12.1 ± 2.0 lM (Fig. 2D).
3.3. Nrp1 colocalizes with FlnA in vesicles in response to VEGF-A
To gain insight into the functional significance of the interactionbetween Nrp1 and FlnA, we tracked their localization in VEGF-A-treated nrp1+/+ ECs. Whereas there was little or no colocalizationin quiescent ECs (Fig. 3A), the two proteins colocalized extensivelyin vesicular punctae in the cytoplasm 5 min after the introductionof VEGF-A (Fig. 3B). The extent of colocalization decreased at thelater time points of 10 and 30 min (Fig. 3C–E).
To test for the dependence of the colocalization of Nrp1 andFlnA on the cytoplasmic interaction of the two proteins, we probed
Fig. 1. Identification of novel binding partners for the cytoplasmic domain of Nrp1.Coomassie-stained tris-glycine acrylamide (10%) showing bands of proteinsimmunoprecipitated by Nrp1 from primary mouse heart ECs incubated withoutor with 20 ng/ml VEGF-A165 for 10 min. (1) Dynein heavy chain; (2) myosin heavychains 9 and 10; (3) eukaryotic translation elongation factor 1 a1; (4) filamin A; (5and 6) myosin heavy chains 9 and 10; (7) eukaryotic translation elongation factor 1a1; (8) a enolase. None of these bands were present in a sample that wasimmunoprecipitated by non-immune goat IgG (not shown). Table 1 listing theproteins that were coimmunoprecipitated together with Nrp1, and whose corre-sponding bands were identified by LC–MS.
Table 1Protein ligands of the Nrp1 cytoplasmic domain that were identified by LC–MS. Theprotein name, gene name, peptide coverage, and Mascot scores are listed.
Protein Bandnumber
Genename
Peptides Mascotscore
Non-muscle myosin IIa heavychain
2, 5, 6 myh9 83 5871
Non-muscle myosin IIb heavychain
2, 5, 6 myh10 40 2479
Cytoplasmic dynein heavychain 1
1 dyhc1 4 148
Filamin A 4 flnA 2 85Eukaryotic translation
elongation factor 1 a13, 7 ef1a1 5 275
a enolase 8 enoa 3 175
H.R. Seerapu et al. / FEBS Letters 587 (2013) 3392–3399 3393

Author's personal copy
their localization in nrp1cytoD/D ECs. The Nrp1 population did notundergo noticeable redistribution after the application of VEGF-A,and it did not colocalize with FlnA (Fig. 3A0 and B0). Rather, Nrp1remained concentrated along the cell edges, as in quiescentnrp1cytoD/D cells.
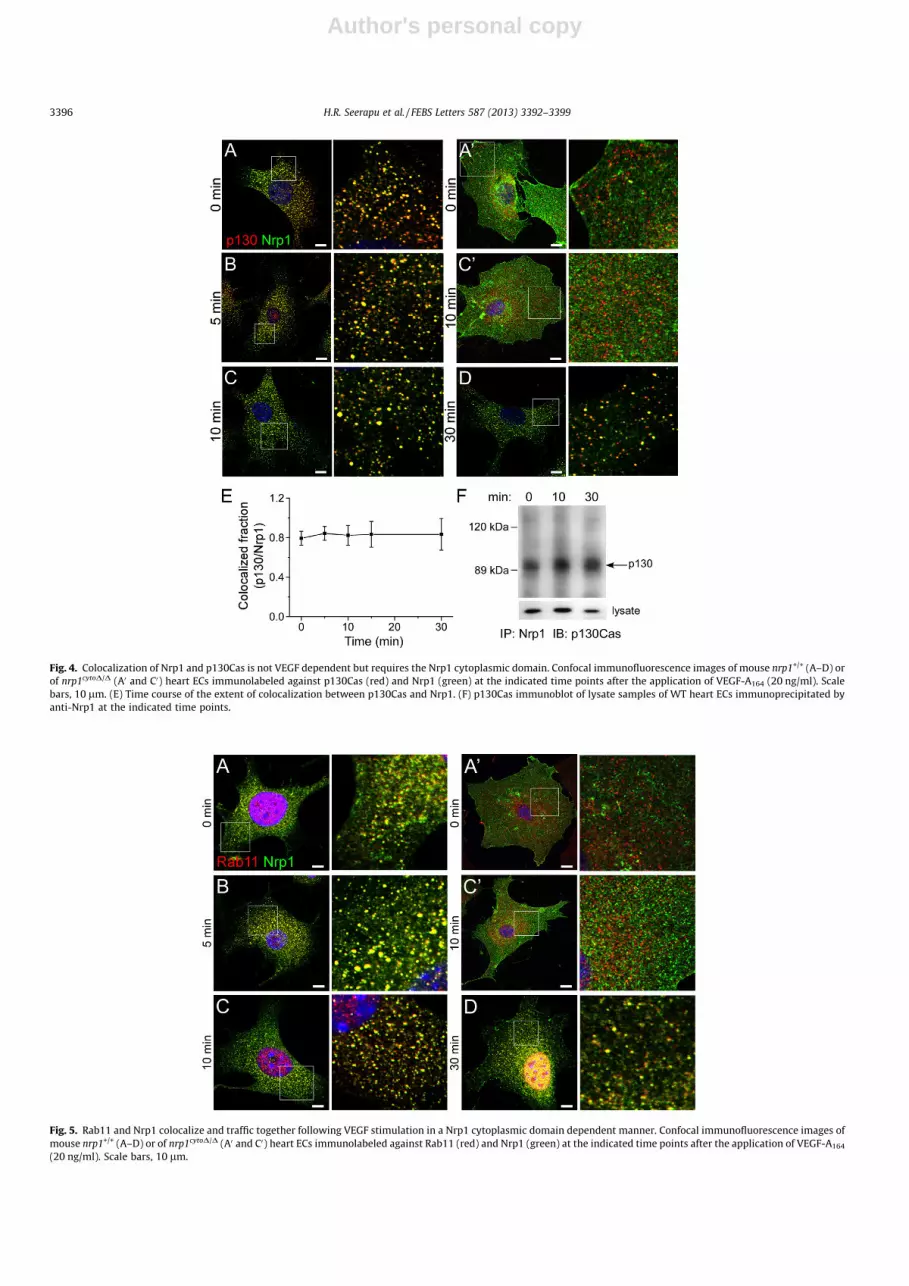
3.4. Nrp1 associates with FA proteins
Because of the known association of FlnA with FAs, we sought,to determine if Nrp1 binds other FA proteins. Several studies hadshown that Nrp1-dependent cell migration is mediated byp130Cas and its tyrosine phosphorylation [19,28,29]. We sought,therefore, to test if the two proteins associate with each otherin a VEGF-dependent manner. Nrp1 and p130Cas colocalizedextensively in quiescent ECs (Fig. 4A–D), and remained colocal-ized to a similar degree 30 min after the administration of VEGF(Fig. 4E). This is vastly different from the colocalization of Nrp1with FlnA, which was much more transient. Further, Nrp1 andp130Cas coimmunoprecipitated in both quiescent EC and VEGF-
treated ECs, similar to their colocalization (Fig. 4F). Despite theobserved presence of p130Cas in FAs [30], we detected p130Casonly in vesicular punctae. Apparently, the p130Cas that we visu-alized was a dynamic fraction that may have been recycling toand/or from FAs, whereas the association with FAs may have beentoo transient to be detected at the time points we sampled. Sim-ilar to FlnA, Nrp1 and p130Cas did not colocalize in nrp1cytoD/D
ECs (Fig. 4A0 and C0).
3.5. Nrp1 recycles in a Rab11-dependent manner
While it is known that Nrp1 undergoes clathrin and Rab5-dependent endocytosis [31], the mechanisms that confer c be char-acterized. We sought to identify which Rab GTPase colocalizes withit. Nrp1 colocalized extensively with Rab11 (Fig. 5A–D), but notwith Rab4 (data not shown), suggesting that it underwent slowrecycling via perinuclear endosomes [32]. Indeed, by 30 min themajority of the cytoplasmic Nrp1 population was localized to theperinuclear region (Fig. 5D). In nrp1cytoD/D ECs, Nrp1 did not colo-
Fig. 2. Binding between FlnA and the Nrp1 cytoplasmic domain. (A) SPR sensorgrams of FlnA repeats 23-24 binding to immobilized Nrp1 cytoplasmic region. A range ofanalyte concentrations were tested, which showed concentration dependence of binding. (B) The fitted SPR binding data gives a dissociation constant, KD, of 9.6 (±1.2) lM. (C)SPR data for the interaction between Nrp1 cytoplasmic region and FlnA repeats 10–11; there was no observable binding between the two domains. (D) ITC binding data forthe titration of Nrp1 cytoplasmic domain (0.4 mM) with FlnA repeats 23–24 (0.04 mM). The Nrp1 cytoplasmic region bound these repeats with a KD of 12.9 (±2.0) lM.
3394 H.R. Seerapu et al. / FEBS Letters 587 (2013) 3392–3399

Author's personal copy
calize with Rab11 at all, indicating that its cytoplasmic domain isrequired for the recruitment of Nrp1 to Rab11-carrying vesicles(Fig. 5A0 and C0).
3.6. VEGF-induced migration and rate of FA turnover are slower innrp1cytoD/D ECs
The association of Nrp1 with FA proteins both in vesicles and inFAs of VEGF-treated ECs raised the possibility that Nrp1 may be in-volved in FA turnover, and, consequently, in regulating VEGF-in-duced cell migration. To test this premise, we compared the ratesof gap closure by nrp1+/+ and nrp1cytoD/D ECs in the presence ofVEGF-A. The rate of VEGF-induced gap closure by nrp1cytoD/D ECswas significantly lower than the gap closure rate by the same celltype in the absence of VEGF (Fig. 6).
We chose kindlin-2 as a marker for tracking FAs in live cells,because it associates with FlnA [33], like Nrp1. To test directlythe rate of FA turnover in nrp1+/+ and nrp1cytoD/D ECs, we transfec-ted each cell type by kindlin-2 fused to the fluorescent proteinmCherry (mCherry-K2). We then imaged the dynamics of themCherry-K2 in basal FAs using total internal reflection fluores-cence (TIRF). By tracking the rate of the appearance or removalof kindlin-2 from individual FAs, we measured the rate constantsof FA assembly and disassembly, respectively, in wild type and in
nrp1cytoD/D ECs, after the administration of VEGF-A164. We foundthat the rates of both the assembly and disassembly of FAs wereapproximately an order of a magnitude lower in the nrp1cytoD/D
ECs (Fig. 7, Supplemental videos 1, 2), thus suggesting that thecytoplasmic domain of Nrp1 is required for the turnover of FAsin VEGF-treated ECs.
4. Discussion
The precise contribution of Nrp1 to VEGF signaling and in par-ticular, the role of the Nrp1 cytoplasmic domain have been debatedsince the discovery that Nrp1 is a VEGF co-receptor [34]. Recent re-port suggests that the Nrp1 cytoplasmic tail is dispensable fordevelopmental and pathological angiogenesis [35], but essentialfor VEGF-induced arteriogenesis in both physiological and ische-mic settings [11]. As recently discussed [36], by default, the puta-tive contribution of Nrp1 to VEGF signaling would involve thecytoplasmic domain of Nrp1. Since the cytoplasmic domain is only44 residues long and appears to be devoid of catalytic activity, themost likely manner in which this domain acts in VEGF signaling isby the binding and assembly of other proteins, such as synectin[11], or FlnA.
The picture that emerges from this study is that Nrp1 localizesmainly along the trafficking pathway of several FA components,
Fig. 3. Confocal immunofluorescence images showing that the colocalization of Nrp1 and FlnA depends on the presence of the Nrp1 cytoplasmic domain and on VEGF. Mousenrp1+/+ (A–D) or nrp1cytoD/D (A0 and C0) heart ECs immunolabeled against FlnA (red) and Nrp1 (green) are seen at the indicated time points after the application of VEGF-A164
(20 ng/ml). The magnified subfields correspond to the area in the white squares. Arrowheads denote colocalized punctae of FlnA and Nrp1. Scale bars, 10 lm.
H.R. Seerapu et al. / FEBS Letters 587 (2013) 3392–3399 3395
Author's personal copy
Fig. 4. Colocalization of Nrp1 and p130Cas is not VEGF dependent but requires the Nrp1 cytoplasmic domain. Confocal immunofluorescence images of mouse nrp1+/+ (A–D) orof nrp1cytoD/D (A0 and C0) heart ECs immunolabeled against p130Cas (red) and Nrp1 (green) at the indicated time points after the application of VEGF-A164 (20 ng/ml). Scalebars, 10 lm. (E) Time course of the extent of colocalization between p130Cas and Nrp1. (F) p130Cas immunoblot of lysate samples of WT heart ECs immunoprecipitated byanti-Nrp1 at the indicated time points.
Fig. 5. Rab11 and Nrp1 colocalize and traffic together following VEGF stimulation in a Nrp1 cytoplasmic domain dependent manner. Confocal immunofluorescence images ofmouse nrp1+/+ (A–D) or of nrp1cytoD/D (A0 and C0) heart ECs immunolabeled against Rab11 (red) and Nrp1 (green) at the indicated time points after the application of VEGF-A164
(20 ng/ml). Scale bars, 10 lm.
3396 H.R. Seerapu et al. / FEBS Letters 587 (2013) 3392–3399

Author's personal copy
Fig. 7. Deletion of the Nrp1 cytoplasmic domain slows down FA turnover. (A and A0) TIRF mages of FAs (encircled in red line) in nrp1+/+ and nrp1cytoD/D ECs, respectively. (Band C, B0 and C0) Time course of the relative fluorescence intensities of assembling (B and B0) or disassembling (C and C0) individual FAs. (D) The mean rates of assembly anddisassembly of nrp1+/+ (black columns) and nrp1cytoD/D (grey columns) ECs. The rates from four cells and 15 FAs were measured for each cell type (p = 1.3 � 10�7; 4.9 � 10�9).
Fig. 6. Cell migration is impeded by deletion of the Nrp1 cytoplasmic domain. (A) Images of gaps between monolayers of nrp1+/+ or of nrp1cytoD/D (KI) cells before and 6 h aftertreatment with 50 ng/ml VEGF-A164. The edges of the monolayers are delineated by a black line. (B) Time course of gap closure of nrp1+/+ (triangles) and nrp1cytoD/D ECs(squares) ECs (n = 3, p = 0.036, 0.009).
H.R. Seerapu et al. / FEBS Letters 587 (2013) 3392–3399 3397

Author's personal copy
primarily the scaffold proteins p130Cas and FlnA, whereas it re-sides in FAs only transiently. Since the distribution of Nrp1 in qui-escent ECs is along the cell borders, it appears that the role of Nrp1in FA dynamics triggered by the application of VEGF is in drivingtheir disassembly (Fig. 8). The membrane traffic of Nrp1 appearsto be dependent on Rab11, in agreement with previous studies[11,37]. Thus, it is likely that Nrp1 mediates FA turnover in re-sponse to VEGF by recycling several FA components, includingFlnA. While these components can be thought of as cargo, Nrp1probably provides the means to mobilize the vesicles to which theyare bound by cross-linking them to myosin VI via synectin. Thevastly slower FA turnover in the nrp1cytoD/D ECs in response toVEGF, where the truncated Nrp1 mutant is unable to bind synectin,is the likely cause of the reduced rate of migration of these cells.This impaired migration could account at least in part for the vas-cular defect observed in vivo in the nrp1cytoD/D mouse model [11].
Other studies proposed that Nrp1 is involved in VEGF-inducedtyrosine phosphorylation of FA kinase and, consequently, in theregulation of FA assembly [38]. The possible involvement of Nrp1in the regulation of FA turnover was deduced also from its rolein stimulating the phosphorylation of the scaffold proteinp130Cas, a known component of FAs [30]. Functional associationsbetween Nrp1 and FAs have been reported previously. Severalstudies found links between Nrp1 and the FA protein p130Cas,wherein Nrp1 promoted the migration of several cell types viathe tyrosine phosphorylation of p130Cas [19,28,29]. Here we ex-pand those studies by observing the association between Nrp1and p130Cas, and their widespread colocalization. Nrp2 was alsoreported to be involved in the formation of FAs through the regu-lation of the activity of integrin a6b1, and was shown to be abun-dant in FAs [39].
Interestingly, FlnA had been reported to regulate the recyclingof the calcitonin receptor (CTR), a G-protein coupled receptor thatis involved in the maintenance of calcium homeostasis [40]. Simi-lar to Nrp1, the C-terminus of CTR binds FlnA. While our study didnot test directly if FlnA regulates the recycling of Nrp1, it should bepointed out that the recycling mechanisms and interaction withFlnA differ between the two receptors. Whereas CTR undergoes to-nic endocytosis and binds FlnA constitutively, Nrp1 undergoesVEGF-triggered endocytosis and, given the immunofluorescencedata, binds FlnA transiently. It is unlikely therefore that FlnA regu-lates the recycling of Nrp1. In a separate study, FlnA was found toregulate the rate of FA disassembly in a calpain-dependent manner[18]. We observed that once cells were activated by VEGF, FlnA wasdisplaced from the cell borders, where it would have been in closeproximity to peripheral FAs. It is possible, therefore, that the VEGF/Nrp1-dependent dynamics of FlnA contributes to FA disassembly.
In conclusion, Nrp1 interacts with several focal adhesion proteins.Since Nrp1 is also a VEGF coreceptor, it is in a position to act as aninterface between the VEGF signaling pathway and FA dynamics.
Acknowledgments
We thank Dr. E. Plow, Cleveland Clinic, for sharing with us themCherry-kindlin-2 construct, and Soon Jeung Kim, Case WesternReserve, for help with molecular biology. This study was supportedin part by National Institutes of Health Grants GM073071 andGM092851 (to M.B.) and by an American Heart Association post-doctoral fellowship (to S.B.).
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.febslet.2013.08.040.
References
[1] Schwarz, Q. and Ruhrberg, C. (2010) Neuropilin, you gotta let me know: shouldI stay or should I go? Cell Adhes. Migr. 4, 61–66.
[2] Zachary, I.C. (2011) How neuropilin-1 regulates receptor tyrosine kinasesignalling: the knowns and known unknowns. Biochem. Soc. Trans. 39, 1583–1591.
[3] Koch, S., Tugues, S., Li, X., Gualandi, L. and Claesson-Welsh, L. (2011) Signaltransduction by vascular endothelial growth factor receptors. Biochem. J. 437,169–183.
[4] Pan, Q. et al. (2007) Blocking neuropilin-1 function has an additive effect withanti-VEGF to inhibit tumor growth. Cancer Cell 11, 53–67.
[5] Pan, Q. et al. (2007) Neuropilin-1 binds to VEGF121 and regulates endothelialcell migration and sprouting. J. Biol. Chem. 282, 24049–24056.
[6] Wang, L., Mukhopadhyay, D. and Xu, X. (2006) C terminus of RGS-GAIP-interacting protein conveys neuropilin-1-mediated signaling duringangiogenesis. FASEB J. 20, 1513–1515.
[7] Wang, L., Zeng, H., Wang, P., Soker, S. and Mukhopadhyay, D. (2003)Neuropilin-1-mediated vascular permeability factor/vascular endothelialgrowth factor-dependent endothelial cell migration. J. Biol. Chem. 278,48848–48860.
[8] Hota, P.K. and Buck, M. (2012) Plexin structures are coming: opportunities formultilevel investigations of semaphorin guidance receptors, their cellsignaling mechanisms, and functions. Cell. Mol. Life Sci. 69, 3765–3805.
[9] Cai, H. and Reed, R.R. (1999) Cloning and characterization of neuropilin-1-interacting protein: a PSD-95/Dlg/ZO-1 domain-containing protein thatinteracts with the cytoplasmic domain of neuropilin-1. J. Neurosci. 19,6519–6527.
[10] Naccache, S.N., Hasson, T. and Horowitz, A. (2006) Binding of internalizedreceptors to the PDZ domain of GIPC/synectin recruits myosin VI to endocyticvesicles. Proc. Natl. Acad. Sci. USA 103, 12735–12740.
[11] Lanahan, A. et al. (2013) The neuropilin 1 cytoplasmic domain is required forVEGF-A-dependent arteriogenesis. Dev. Cell 25, 156–168.
[12] Hanein, D. and Horwitz, A.R. (2012) The structure of cell-matrix adhesions:the new frontier. Curr. Opin. Cell Biol. 24, 134–140.
[13] Wehrle-Haller, B. (2012) Structure and function of focal adhesions. Curr. Opin.Cell Biol. 24, 116–124.
[14] Sieg, D.J., Hauck, C.R., Ilic, D., Klingbeil, C.K., Schaefer, E., Damsky, C.H. andSchlaepfer, D.D. (2000) FAK integrates growth-factor and integrin signals topromote cell migration. Nat. Cell Biol. 2, 249–256.
[15] Avraham, H.K., Lee, T.H., Koh, Y., Kim, T.A., Jiang, S., Sussman, M., Samarel, A.M.and Avraham, S. (2003) Vascular endothelial growth factor regulates focaladhesion assembly in human brain microvascular endothelial cells throughactivation of the focal adhesion kinase and related adhesion focal tyrosinekinase. J. Biol. Chem. 278, 36661–36668.
[16] Ruhrberg, C. (2003) Growing and shaping the vascular tree: multiple roles forVEGF. BioEssays 25, 1052–1060.
[17] Vieira, J.M., Ruhrberg, C. and Schwarz, Q. (2010) VEGF receptor signaling invertebrate development. Organogenesis 6, 97–106.
[18] Xu, Y. et al. (2010) Filamin A regulates focal adhesion disassembly andsuppresses breast cancer cell migration and invasion. J. Exp. Med. 207, 2421–2437.
[19] Evans, I.M., Yamaji, M., Britton, G., Pellet-Many, C., Lockie, C., Zachary, I.C. andFrankel, P. (2011) Neuropilin-1 signaling through p130Cas tyrosinephosphorylation is essential for growth factor-dependent migration ofglioma and endothelial cells. Mol. Cell Biol. 31, 1174–1185.
[20] Kuo, J.C., Han, X., Hsiao, C.T., Yates 3rd, J.R. and Waterman, C.M. (2011)Analysis of the myosin-II-responsive focal adhesion proteome reveals a rolefor beta-Pix in negative regulation of focal adhesion maturation. Nat. Cell Biol.13, 383–393.
Fig. 8. Schematic representation of the function of Nrp1 in focal adhesion turnover.According to this scenario, Nrp1 is endocytosed in response to VEGF, targeted tofocal adhesions, where it resides transiently, and then departs with FlnA bound toits cytoplasmic domain. The Nrp1–FlnA complex recycles back in a Rab11-dependent manner. Speculatively, p130Cas remains associated with Nrp1 through-out this process.
3398 H.R. Seerapu et al. / FEBS Letters 587 (2013) 3392–3399

Author's personal copy
[21] Nakamura, F., Stossel, T.P. and Hartwig, J.H. (2011) The filamins: organizers ofcell structure and function. Cell. Adhes. Migr. 5, 160–169.
[22] Zhou, A.X., Hartwig, J.H. and Akyurek, L.M. (2010) Filamins in cell signaling,transcription and organ development. Trends Cell Biol. 20, 113–123.
[23] Zamir, E. and Geiger, B. (2001) Components of cell-matrix adhesions. J. Cell Sci.114, 3577–3579.
[24] Hara, T., Honda, K., Shitashige, M., Ono, M., Matsuyama, H., Naito, K.,Hirohashi, S. and Yamada, T. (2007) Mass spectrometry analysis of thenative protein complex containing actinin-4 in prostate cancer cells. Mol. Cell.Proteomics 6, 479–491.
[25] Maciver, S.K. (1996) Myosin II function in non-muscle cells. BioEssays 18,179–182.
[26] Shelden, E. and Knecht, D.A. (1996) Dictyostelium cell shape generationrequires myosin II. Cell Motil. Cytoskeleton 35, 59–67.
[27] Schiller, H.B., Friedel, C.C., Boulegue, C. and Fassler, R. (2011) Quantitativeproteomics of the integrin adhesome show a myosin II-dependent recruitmentof LIM domain proteins. EMBO Rep. 12, 259–266.
[28] Frankel, P., Pellet-Many, C., Lehtolainen, P., D’Abaco, G.M., Tickner, M.L., Cheng,L. and Zachary, I.C. (2008) Chondroitin sulphate-modified neuropilin 1 isexpressed in human tumour cells and modulates 3D invasion in the U87MGhuman glioblastoma cell line through a p130Cas-mediated pathway. EMBORep. 9, 983–989.
[29] Pellet-Many, C., Frankel, P., Evans, I.M., Herzog, B., Junemann-Ramirez, M. andZachary, I.C. (2011) Neuropilin-1 mediates PDGF stimulation of vascularsmooth muscle cell migration and signalling via p130Cas. Biochem. J. 435,609–618.
[30] Harte, M.T., Hildebrand, J.D., Burnham, M.R., Bouton, A.H. and Parsons, J.T.(1996) P130Cas, a substrate associated with v-Src and v-Crk, localizes to focaladhesions and binds to focal adhesion kinase. J. Biol. Chem. 271, 13649–13655.
[31] Salikhova, A., Wang, L., Lanahan, A.A., Liu, M., Simons, M., Leenders, W.P.,Mukhopadhyay, D. and Horowitz, A. (2008) Vascular endothelial growth factor
and semaphorin induce neuropilin-1 endocytosis via separate pathways. Circ.Res. 103, e71–9.
[32] Ullrich, O., Reinsch, S., Urbe, S., Zerial, M. and Parton, R.G. (1996) Rab11regulates recycling through the pericentriolar recycling endosome. J. Cell Biol.135, 913–924.
[33] Tu, Y., Wu, S., Shi, X., Chen, K. and Wu, C. (2003) Migfilin and Mig-2 link focaladhesions to filamin and the actin cytoskeleton and function in cell shapemodulation. Cell 113, 37–47.
[34] Soker, S., Takashima, S., Miao, H.Q., Neufeld, G. and Klagsbrun, M. (1998)Neuropilin-1 is expressed by endothelial and tumor cells as an isoform-specific receptor for vascular endothelial growth factor. Cell 92, 735–745.
[35] Fantin, A., Schwarz, Q., Davidson, K., Normando, E.M., Denti, L. and Ruhrberg,C. (2011) The cytoplasmic domain of neuropilin 1 is dispensable forangiogenesis, but promotes the spatial separation of retinal arteries andveins. Development 138, 4185–4191.
[36] Horowitz, A. and Seerapu, H.R. (2012) Regulation of VEGF signaling bymembrane traffic. Cell. Signal. 24, 1810–1820.
[37] Ballmer-Hofer, K., Andersson, A.E., Ratcliffe, L.E. and Berger, P. (2011)Neuropilin-1 promotes VEGFR-2 trafficking through Rab11 vesicles therebyspecifying signal output. Blood 118, 816–826.
[38] Herzog, B., Pellet-Many, C., Britton, G., Hartzoulakis, B. and Zachary, I.C. (2011)VEGF binding to NRP1 is essential for VEGF stimulation of endothelial cellmigration, complex formation between NRP1 and VEGFR2, and signaling viaFAK Tyr407 phosphorylation. Mol. Biol. Cell 22, 2766–2776.
[39] Goel, H.L., Pursell, B., Standley, C., Fogarty, K. and Mercurio, A.M. (2012)Neuropilin-2 regulates alpha6beta1 integrin in the formation of focaladhesions and signaling. J. Cell Sci. 125, 497–506.
[40] Seck, T., Baron, R. and Horne, W.C. (2003) Binding of filamin to the C-terminaltail of the calcitonin receptor controls recycling. J. Biol. Chem. 278, 10408–10416.
H.R. Seerapu et al. / FEBS Letters 587 (2013) 3392–3399 3399





























