Embed Size (px)
Citation preview

R
Ts
MG
a
AA
KHEHPF
C
S
0d
Journal of Chromatography A, 1218 (2011) 8791– 8798
Contents lists available at ScienceDirect
Journal of Chromatography A
jou rn al h om epage: www.elsev ier .com/ locat e/chroma
eview
he development and characterization of protein-based stationary phases fortudying drug–protein and protein–protein interactions
itesh Sanghvi, Ruin Moaddel ∗, Irving W. Wainererontology Research Center, National institute on Aging, National Institutes of Health, 5600 Nathan Shock Drive, Baltimore, MD 21224, USA
r t i c l e i n f o
rticle history:vailable online 27 May 2011
a b s t r a c t
Protein-based liquid chromatography stationary phases are used in bioaffinity chromatography forstudying drug–protein interactions, the determination of binding affinities, competitive and allosteric
eywords:sp90RRSArotein fishingrontal affinity chromatography
interactions, as well as for studying protein–protein interactions. This review addresses the develop-ment and characterization of protein-based stationary phase, and the application of these phases usingfrontal and zonal chromatography techniques. The approach will be illustrated using immobilized heatshock protein 90� and the immobilized estrogen related receptor stationary phases. In addition, thereview discusses the use of the protein-coated magnetic beads for ligand and protein fishing as well asfor the identification of unknown ligands from cellular or botanical extracts.
Published by Elsevier B.V.
ontents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87922. Experimental approach . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8793
2.1. Synthesis of protein-immobilized LC phases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87932.2. Chromatographic techniques . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8793
2.2.1. Frontal affinity chromatography . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87932.2.2. Non linear chromatography . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8793
3. Application of biochromatography . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87943.1. Estrogen related receptor (ERR� and ERR�) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87943.2. Human serum albumin (HSA) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87953.3. Heat shock protein � (Hsp90�) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8795
3.3.1. ATPase activity of Hsp90�-CT column . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87953.3.2. Thiol oxidation of Hsp90� . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8796
3.4. Ligand fishing using Hsp90� coated magnetic beads (Hsp90� – MB) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87963.4.1. Preparation of Hsp90�-MB . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87963.4.2. Application of Hsp90�(CT)-MB and Hsp90�(NT)-MB . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8797
3.4.3. Protein fishing with protein coated magnetic beads . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 87974. Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8797Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8798References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8798
∗ Corresponding author at: Bioanalytical and Drug Discovery Unit, National Institute on Aging, National Institutes of Health, Gerontology Research Center, 5600 Nathanhock Drive, Baltimore, MD 21224-6825, USA. Tel.: +1 410 558 8294; fax: +1 410 558 8409.
E-mail address: [email protected] (R. Moaddel).
021-9673/$ – see front matter. Published by Elsevier B.V.oi:10.1016/j.chroma.2011.05.067

8 atogr.
1
afltstcAttDoddwAtilaattdbrTdmamofipf
emsoatitaobpbmwatc�rivhTatm
792 M. Sanghvi et al. / J. Chrom
. Introduction
The measurement of protein–protein interactions has become key factor in drug discovery. The methods currently usedor these measurements include but are not limited to staticight scattering (SLS) [1,2], ultracentrifugation [3], X-ray scat-ering [4], self-interaction chromatography (SIC) [5,6] andize-exclusion chromatography (SEC) [7,8]. The characterization ofhe protein–protein interactions using these methods is typicallyarried out by determining the second virial coefficient parameter.
positive coefficient implies that repulsion interactions dominatehe protein–protein interaction, while a negative number is indica-ive of a net attraction for the protein–protein interactions [9].umetz et al., for example, carried out SIC to measure the effectf salt concentration on the protein–protein interactions, [6] andetermined that the protein interactions show very low salt depen-ence for sodium chloride solutions, with a very pronounced effecthen ammonium sulfate is employed, for their proteins tested.
limitation of SIC is that it requires prior immobilization of pro-ein, which can affect protein structure and thus protein–proteinnteraction. In addition, SIC uses the same protein as ligand andigate and thus is not useful to study multiprotein complexes,nd several other methods are limited to protein–protein inter-ctions between two different proteins. Bloustine et al. used SECechnique to determine the solute distribution coefficient fromhe retention times measured by refractive index detector andiode array detector [7]. They also compared the result obtainedy SEC technique to the results obtained by frontal chromatog-aphy [10,11] and from light-scattering measurements [12–14].herefore, SIC and SEC techniques have the advantage of a shorteruration time relative to the other techniques. Although, theseethods are very useful, they still use larger amounts of protein,
lbeit, Garica et al. have recently shown that SIC technique can beiniaturized to a microchip, thus significantly reducing the amount
f protein required for the determination of the second virial coef-cient parameter [9]. SLS technique also requires large amounts ofrotein in order to determine the coefficient parameter and there-ore is seldom used [1,2].
Protein–protein and protein-ligand interactions have also beenxplored through protein immobilization on solid supports of chro-atographic and non-chromatographic experimental techniques
uch as microarrays [15,16], biosensors [17] and nanotechnol-gy [18]. In protein microarrays, functionally active proteins wererrayed using microfabricated polyacrylamide gel pads to cap-ure their samples [19], while more recently, Macbeath et al.mmobilized the protein onto the surface of the plates in ordero probe thousands of protein–protein and ligand–protein inter-ctions [15,16]. This method was developed to take advantagef existing instrumentation. This was accomplished by immo-ilizing the protein onto a solid support while preserving itsroper conformation. More specifically, the protein was immo-ilized covalently onto the surface of smooth flat surfaces oficroscope slides. Although, several chemically derivatized slidesere used the majority of the studies were carried out using the
ldehyde-containing slides. This takes advantage of immobilizinghe protein through primary amine containing residues, specifi-ally the N-terminus lysine residue as they are a more reactive-amine. For details on the fabrication of protein microarrays c.f.
ef Macbeth et al. Briefly, nanoliter volumes of protein sampless delivered on the slides in PBS with 40% glycerol, which pre-ents evaporation of the nanodroplets, thus keeping the proteinsydrated. The unreactive functional groups are capped using BSA.
hese slides have been used to study protein–protein interactionss well as small molecule protein interactions. Protein interac-ion at specific sites can also be studied using protein-domainicroarrays, Espejo et al., used a microarray approach to study
A 1218 (2011) 8791– 8798
signal transduction issues and interactions that are sensitive toarginine methylation. This was accomplished using a glutathione S-transferase (GST)-fusion protein that contained a peptide-specificbinding motif, allowing for the immobilization onto a supportto study the specific protein interaction at a specific site [20].Several other methods have been reported for protein microar-rays and have been previously reviewed [21]. For example, Zhuet al. constructed yeast proteome microarrays containing 5800yeast proteins and screened it for various biochemical activi-ties [16]. Many known kinases and calcineurins were identifiedalong with 33 new calmodulin binding partners. Knezevic et al.showed the alteration in specific levels of more than ten cancerrelated proteins expression due to ionizing radiation treatment[22]. Miller et al. also successfully showed the antibody microarrayscontaining 184 antibodies to profile the serum of patient diag-nosed with prostate cancer to identify potential biomarkers [23].The further development and characterization of this technologycan lead to its application in a personalized medicine, where atreatment can be tailored to specific individual for an increasedefficacy.
An alternative approach is to use the immobilized protein asa stationary phase in bioaffinity chromatography. This techniqueis based on specific reversible interaction between the ligand andthe immobilized protein. A widely used method for the synthe-sis of protein-based stationary phases is the immobilization ofthe protein on the solid support using adsorption or covalentimmobilization. The resulting protein-based LC phases (SPs) canbe used to determine and characterize ligand–protein interac-tions [24,25]. The theory and applicability of using immobilizedproteins to explore the interactions between ligands/substratesand an immobilized cytosolic protein or enzyme were initiallydescribed by Chaiken [26] and Carr [27] and expanded to thestudy of transmembrane proteins by Lundahl et al. [24,25]. It wasfurther expanded to the immobilization of cellular membrane frag-ments and their use in cellular membrane affinity chromatography(CMAC) [28]. Recent data using frontal and zonal chromatographictechniques have demonstrated that ligand binding affinities (Kdvalues) obtained using protein based stationary phases are com-parable to affinities obtained using standard membrane bindingtechniques [29].
In addition, Belanger immobilized Mex67-Nep1 onto sepharosebeads and determined protein–protein interactions by LC/MS andWestern blotting techniques [30]. Magnetic beads have gained asignificant amount of interest as an alternative method for ‘fish-ing’ experiments for both ligand and protein binders. It has beendemonstrated and will be discussed in greater detail below thatthe formation of protein–protein complexes remain intact on thesurface of the protein coated magnetic beads [31–34]. The proteincoated magnetic beads, were successfully used to fish out bindersfrom a mixture of binders and non-binders for HSA [35], and theidentification of ligands that modulate protein–protein interac-tions [31]. Further, Jonker et al., have shown that the magneticnano-particles can be used as a novel high throughput screeningmethodology, to determine whether a compound has an affinityfor an immobilized target in a ‘yes’ or ‘no’ method [32]. The forma-tion of a multiprotein complex was also carried out on the surfaceof the protein coated magnetic beads, and it has been demonstratedthat the resulting protein coated magnetic bead was able to fish outa binding partner in a complex matrix, the KU-812 cellular matrix[34].
The development of protein-based stationary phase and theircharacterization using frontal and zonal chromatographic tech-niques will be demonstrated using Hsp90� and the estrogen related
receptors. The use of HSA and Hsp90�-coated magnetic beads forligand and protein fishing will also be reviewed.
atogr.
2
mami
2
ritsod
fiftpe
isctvsHie
tHtcaett([im
wiiwsdal[boowaTtpa
M. Sanghvi et al. / J. Chrom
. Experimental approach
The protein immobilized stationary phases utilized in biochro-atography can be created using a variety of experimental
pproaches and the resulting columns used in a variety of chro-atographic techniques. These various approaches are described
n Section 2.1.
.1. Synthesis of protein-immobilized LC phases
Protein immobilization onto a stationary phases can be car-ied out using adsorption, covalent immobilization and directionalmmobilization using his-tag or GST fusion proteins. Apart fromhese widely used approaches, it can also be carried out on theurface of an open tubular capillary or magnetic beads. The vari-us advantages and disadvantages of these approaches have beeniscussed below.
The major advantage of an adsorption technique is that modi-cation of the protein is not required. Conversely, this technique
orms a weak and mainly reversible interaction between the sta-ionary phase and protein. As a result, a slow leakage of immobilizedrotein from the stationary phase can be observed, which wouldventually result in loss of activity [36].
The most frequently used method of protein immobilizations covalent immobilization, although commonly used, this occa-ionally prevents the protein from immobilizing in its properonformation. The immobilizations are typically carried out usinghe N-terminal or C-terminal of the protein with the intent of pre-enting steric interaction between the solid support and the activeites of the protein, which are not usually present at the terminals.owever, this method has the disadvantage of potentially disrupt-
ng the proper protein conformation. These methods have beenxtensively reported and will not be discussed here [34,37–44].
Directional immobilization can be carried out using variousechniques, including the use of fusion proteins with either ais-tag [45] or glutathione S-transferase (GST) tagged fusion pro-
eins [46]. In addition, the proteins can be immobilized with theirorresponding binder, nickel and glutathione, respectively. Thispproach (His-tag) has been used to immobilize the His-taggedstrogen receptor ligand binding domain, ER-LBD [47] and the His-agged DNA unwinding element binding (DUE-B) [45] using Ni+2 ashe coordinating metal ion. In addition Jonker et al., used a CobaltII) coated magnetic bead to immobilize the His-tagged protein32], while McFadden et al. used a Nickel coated magnetic bead tosolate a His-tagged protein from a mixture of proteins and small
olecules [31].Open tubular capillaries [28,37,48], are a promising format for
hich to immobilize proteins as a significant reduction in run times seen, thus increasing throughput. However, due to the decreasen surface area, a significant reduction in the active binding site
ill also be observed. The immobilization of proteins onto theurface of magnetic beads in order to analyze protein–protein orrug–protein interaction by Western-blot or by LC/MS system haslso been carried out. Several articles have been recently pub-ished on the use of magnetic beads for ligand and protein fishing31,32,34,35]. The major advantage of protein-coated magneticeads is the ease in isolating an active compound from a mixturef compounds, since it does not require any additional columnsr fraction collectors. Using this approach, human serum albuminas immobilized onto the surface of silica coated magnetic beads
nd was able to differentiate between ligands and nonligands.
his screening process was also automated [25]. This applica-ion can be extended to immobilize other proteins to determinerotein–protein and ligand–protein interactions and potential lig-nd fishing from plant extracts.A 1218 (2011) 8791– 8798 8793
2.2. Chromatographic techniques
2.2.1. Frontal affinity chromatographyFrontal affinity chromatography (FAC) is used in bioaffinity
chromatography to study ligand–protein interactions. Schriemeret al. and Calleri et al. have reviewed the application of FAC byimmobilizing various proteins onto the columns and using it todetermine binding affinities of potential therapeutic compounds[49,50]. Briefly, a marker ligand is placed in the mobile phase andpassed through the column in the presence or absence of serialconcentrations of a displacing ligand under dynamic equilibriumconditions in FAC [51]. The marker ligand can be monitored using aradioflow detector (radiolabeled ligand), mass spectrometer, UVor fluorescence (for a fluorescence ligand). The resulting chro-matographic traces contains initially a relatively flat initial portion,which represents the nonspecific and specific binding of the markerligand to the stationary phase, followed by a vertical breakthrough,which reflects the saturation of the specific binding sites on theimmobilized protein, and ending in a plateau, which correspondsto the complete saturation of these sites, Fig. 1.
Frontal chromatography allows the determination of the bind-ing affinity of the competing ligand (Kd) and the number of activebinding sites (Bmax) for the immobilized protein. This is determinedusing Eq. (1)
[D](V − Vmin) = P[D](Kd + [D])−1 (1)
where [D] is the concentration of displacer ligand, V is the reten-tion volume corresponding to 50% maximal response; Vmin is theretention volume of displacer ligand when the specific interaction iscompletely suppressed and P is the product of the Bmax (the numberof active binding sites) and (Kd/KdM). From the plot of [D] (V − Vmin)versus [D], dissociation constant values (Kd), for displacer ligandcan be obtained. For the results to be quantitative, Eq. (1) assumesthat the concentration of the Marker ligand is at least one order ofmagnitude less than its Kd for the immobilized protein.
FAC can be used to screen mixture of compounds, for example,a library of 356 �-galactopyranoside compounds were divided in agroup of 25–40 compounds which were screened using an immo-bilized �-galactosidase enzyme on a LC stationary phase column[52].
2.2.2. Non linear chromatographyNon-linear chromatograph (NLC) is also used in bioaffinity
chromatography to study ligand–protein interactions. The zonalinjections of ligands result in asymmetric peak profiles with tail-ing that is proportional to the injected concentration The shapeof this chromatographic peak represents specific and nonspe-cific interactions between the solute and the stationary phase.When the stationary phase contains an immobilized protein, themass transfer process defined by the association and dissocia-tion of a ligand–protein complex usually is slow, producing broad,non-Gaussian chromatographic peaks with significant tailing. Thedegree of deviation from a Gaussian distribution is a function ofapplied ligand concentration, and the concentration-dependentasymmetry can be used with NLC techniques to characterizethe separation processes occurring on the column, includingthe kinetics involved in the formation and dissociation of thesolute–stationary phase complex, the association (ka) and dis-sociation (kd) rate constants, and the equilibrium constant (Ka,calculated as ka/kd) [37,53].
The observed peak asymmetries were analyzed using ImpulseInput Solution (see Eq. (2) below as described previously [53–55]).
y = a0
a3
[1 − exp
(−a3
a2
)][√(a1/x)I1(2
√a3x/a2) exp((−x − a1)/a2)
1 − T[(a′1/a2)(x/a2)][1 − exp(−a3/a2)]
](2)

8794 M. Sanghvi et al. / J. Chromatogr. A 1218 (2011) 8791– 8798
princR
watftKaw
3
bamatraupc
3
ppto
dato(i[v
iptd
Ect3wia
maximum binding capacity value was 40-fold lower relative to theERR�-silica column (4 nmol) [37]. In contrast to ERR�-silica col-umn, the maximum binding capacity of ERR� on the silica support
Fig. 2. A. The effect of increasing concentration of diethylstilbesterol on its chro-
Fig. 1. Cartoon showing the basiceprinted from Ref. [51].
here y is intensity of signal, x is reduced retention time, I0 and I1re modified Bessel functions, a0 is area parameter and a1 is cen-er parameter (which determines the true thermodynamic capacityactor k′), a2 is width parameter, and a3 is distortion parame-er. Kinetic parameters can be calculated as follows: kd = 1/a2/t0,a = a3/C0, and ka = Ka/kd, where t0 is the dead time of the columnnd C0 is the concentration of ligand injected multiplied by theidth of the injection pulse [55,56].
. Application of biochromatography
Various membranes and cytosolic proteins have been immo-ilized onto silica-based stationary phases by several groups,llowing them to study the binding interactions of single orultiple compounds using biochromatography techniques. These
pplications are exemplified here by the synthesis and applica-ion of stationary phases containing immobilized estrogen relatedeceptors � and � (ERR�, ERR�), heat shock protein 90� (HSP90�)nd human serum albumin (HSA) protein. These phases have beensed in high-throughput screening methods including ligand androtein fishing using protein coated magnetic beads, microaffinityolumns, etc. [34,35].
.1. Estrogen related receptor (ERR ̨ and ERR�)
ERRs are a subfamily of nuclear receptors that is currently com-osed of three isoform (ERR�, ERR�, and ERR�) [57,58]. All threeroteins possess the typical functional domains of nuclear recep-ors and were orphan receptors identified based on their high levelf sequence identity with estrogen receptors (ER) � and � [36].
There are a number of characterized ERR agonists which includeaidzein, biochanin A, genistein and 6,3′,4′-trihydroxyflavone [59]nd DY131. DY131, in particular, has been shown to increase ERR�ranscriptional activity by ∼3–4 fold in CV-1 cells [60]. ERR antag-nists include diethylstilbestrol (DES) and 4-hydroxytamoxifen4-OHT) [59]. 4-OHT is selective for the ERR� and EPP� while DESs an antagonists for all three isoforms, ERR �, ERR � and ERR�59,61–63]. Greschik et al. reported the mechanism of ERR deacti-ation upon binding of antagonists like DES and 4-OHT [63].
The ERR proteins (Genscript, Piscataway, NJ) were initiallymmobilized onto the surface of an amine-coated silica stationaryhase. The ERR � and ERR� columns were characterized by FAC andhe obtained binding affinities correlated with previously reportedata [36].
Although the results of the initial studies with the ERR� andRR� LC column demonstrated that the phases could be used toharacterize binding to the immobilized receptor, the application ofhese columns to compound screening was limited by the required
–6 h wash time. In order to increase throughput, ERR� and ERR�as immobilized on the surface of open tubular capillaries resultingn ERR�-OT and ERR�-OT columns. DES was used as the marker lig-nd to characterize the ERR�-OT and ERR�-OT columns using FAC
ipal of Frontal Chromatography.
techniques and obtained chromatograph using various concentra-tions of DES is shown in Fig. 2. The calculated Kd value for ERR�-OT,237 nM [37], was the same as the value obtained on the ERR�-silica column, which clearly indicates that the protein interactionswith DES were similar on both columns. However, the calculated
matographic retention on the ERR�-OT column from right to left (0.05, 0.1, 0.25 and0.5 �M). B. The effect of increasing concentration of diethylstilbesterol on its chro-matographic retention on the ERR�-column from right to left (0.05, 0.1, 0.25, 0.5, 1,2 �M).Reprinted from Ref. [37].

M. Sanghvi et al. / J. Chromatogr. A 1218 (2011) 8791– 8798 8795
0
10
20
30
40
50
60
70
80
90
LabetololFenoterol NicotineNaproxenAZTWarfarin
A1B1A4B4
Fig. 3. HSA coated magnetic beads were incubated with warfarin, Azidothymidine, naproxen, nicotine, fenoterol and labetolol. Nonabsorbed and eluted material werea bed) V(R
dt
3
bmotaaTliAtalsssTne
3
efkmspitgaaeA
tt
nalyzed. A direct comparison of the manual ligand fishing results [A1 (Non-absorEluted)].eprinted from Ref. [35].
id not differ significantly between the open tubular format andhe silica column and the calculated Kd value was 929 nM [37].
.2. Human serum albumin (HSA)
Ligand fishing using protein coated magnetic beads (MB) haseen reported for the first time by our laboratory [35]. HSA coatedagnetic beads were used to fish out known binders from a mixture
f nonbinders. The protocol was optimized by studying incubationime and elution conditions. A mixture of known binders (warfarin,zidothymidine and naproxen) and nonbinders (nicotine, fenoterolnd labetolol) were incubated with magnetic beads for 15 min.he supernatant was collected using Dynal magnetic separator fol-owed by wash and elution. Load, wash and elution containingndividual or mixture of binders were detected and quantified bygilent LC-MSD. As shown in Fig. 3, HSA ligands were retained until
he final elution (A4), whereas the nonbinders were not retainednd were present in the loading solution (A1) (nicotine, fenoterol,abetolol). This method was then automated using the Magtrationystem 12GC PSS Bio Instruments Inc., which can run up to 12amples in parallel. The known binder ligands were successfullyeparated from nonbinders using the magtration system (Fig. 3).his demonstrates the versatility of the use of protein coated mag-etic beads to isolate ligands/proteins from a complex matrix, forxample cellular or botanical extracts.
.3. Heat shock protein ̨ (Hsp90˛)
Hsp90� is the only inducible form in a family of molecular chap-rones (Hsp90�, Hsp90�, Grp94, Trap1), which are involved in theolding, intracellular disposition and proteolytic turnover of manyey regulators for cell growth and survival [64]. Hsp90� exists inultiple conformations including, an ATP-free state, ATP-bound
tate or in a protein complex with other co-chaperones and clientroteins. It has been recently demonstrated that Hsp90� is signif-
cantly elevated in glioma cells compare to normal astrocytes andhis disparity indicates that Hsp90� is a potential therapeutic tar-et for the treatment of astrocytomas [65]. Hsp90� is also vieweds a promising target for the treatment of many forms of cancer and
number of drug development programs are aimed at the discov-ry of compounds that will selectively inhibit the proteins intrinsic
TPase activity [66].HSP90� usually exists as a homodimer made up of monomershat consist of three main domains, each of which have impor-ant functional interactions, Fig. 4 [64]. Atomic resolution of crystal
s A4 (Eluted)] and the automated ligand fishing results [B1 (Non-absorbed) Vs B4
structures have only been solved for individual monomers due tointrinsic conformational mobility of the intact protein. The proteincontains highly conserved N-, middle-, and C-terminal domains[64]. The N-terminal domain contains an adeninenucleotide-binding pocket, which has been associated with the intrinsic ATPaseactivity. There are some reports on extensive structural alterationsdriven by the hydrolysis of ATP to ADP in the adenine nucleotide-binding pocket, which have an essential role in the chaperoningactivity of the HSP90 dimer [67,68]. The chaperone function ofHsp90� is sensitive to oxidative stress and it has been confirmedby using chromatographic approach (See Section 3.3.2).
Hsp90� was immobilized at N-terminus (Fig. 5a) and C-terminus (Fig. 5b) onto the silica stationary phase and theresulting columns were characterized using C-terminal lig-ands and N-terminal ligands respectively [38]. Coumermycin A1(CA1) and novobiocin (NOVO), C-terminal ligands, were usedto characterize binding to the exposed C terminus on theHsp90�-NT column, while geldanamycin (GM), 17-allylamino-17-demethoxygeldanamycine (17-AAG) and radicicol (RAD) were usedto characterize the exposed N-terminus on the HSP90�-CT column.The addition of GM, an N-terminus ligand, to the mobile phase didnot displace NOVO on the HSP90�-NT, indicating that they are notcompeting for the same site. These results indicate that GM didnot specifically bind to the Hsp90�-NT column, nor did it com-petitively or allosterically displace the C-terminus ligands. Similarresults were obtained with the Hsp90�-CT column and C-terminusligand binders.
The relative affinities were determined using the Hsp90�-CTcolumn, RAD > GM > 17-AAG and are consistent with the previouslyreported relative IC50 values for the inhibition of Hsp90 ATPaseactivity [69].
3.3.1. ATPase activity of Hsp90˛-CT columnSince the inhibition of the ATPase activity of Hsp90� is one of
the properties used to screen for new drug candidates, the effectof immobilization of Hsp90� on ATPase activity and the sensitivityto inhibition of this activity were determined. The ATPase activityof Hsp90� has been associated with an adenine nucleotide bindingpocket in the N-terminus [65,70], and, therefore, the Hsp90�-CT
column was used in these studies. The initial studies demonstratedthat the ATPase mediated conversion of ATP to ADP occurred on theHsp90� CT-column [38], and that this hydrolysis could be inhibitedby GM.
8796 M. Sanghvi et al. / J. Chromatogr. A 1218 (2011) 8791– 8798
Fig. 4. The numbering 1–732 indicates the approximate positions in the amino acid sequence of the human protein that define its functional domains. ‘CR’ refers to a chargedr . The
a are alR
3
swiSp[dNfdtHotmi
FaR
egion which serves as a flexible linker between the N-terminal and middle domainsnd modulate its function are indicated. The biochemical functions of each domaineprinted from Ref. [64].
.3.2. Thiol oxidation of Hsp90˛The chaperone function of Hsp90� is sensitive to oxidative
tress [71], therefore the Hsp90� CT column was used to ascertainhether a reversible S-thiolation reaction plays a role in regulat-
ng the intrinsic ATPase function of Hsp90� by carrying out the-glutathionylation of the immobilized Hsp90� This reaction waserformed according to the protocol described in Caplan et al.72]. Briefly, Hsp90� CT was incubated with 2 mM GSH and 1 mMiamide in degassed buffer A [40 mM Tris-HCl (pH 8.0), 140 mMaCl, and 0.2 mM EDTA] for 1 h at room temperature in the dark,
ollowed by extensive washing to remove excess reagents. 3 mMithiothreitol was included in buffer A as a control to monitorhe reversibility of diamide-induced S-thiolation of immobilizedsp90�. ATPase activity was carried out in the presence or absence
f 10 �M GM in the mobile phase as discussed previously [38]. Fur-her, the thiol oxidation of Hsp90�-column by diamide plus GSHediated functional impairment in the chaperone’s ATPase activ-ty, which was reversed by subsequent incubation of the column
ig. 5. The synthetic approaches used in the covalent immobilization of Hsp90� on an amnd (b) carboxyl terminus.eprinted from Ref [38].
locations where various small molecules bind HSP90 (heat-shock protein of 90 kDa)so shown. 17AAG, 17-allylaminogeldanamycin; GA, geldanamycin.
with the reducing agent dithiothreitol, as shown in Fig. 6. Thisdata demonstrated that the immobilized Hsp90� could undergo S-thiolation, which interfered in the intrinsic ATPase activity and thusresulted in a disruption of the chaperone function toward clientproteins.
3.4. Ligand fishing using Hsp90˛ coated magnetic beads(Hsp90 ̨ – MB)
3.4.1. Preparation of Hsp90˛-MBHsp90� was also immobilized through the N-terminus and
C-terminus onto the surface of magnetic beads resulting inthe Hsp90�(CT)-MB and Hsp90�(NT)-MB, respectively [34]. Thedetailed procedure for the immobilization of Hsp90� at N-terminus
and C-terminus onto magnetic beads has been previously reported[34]. Briefly, N-terminus of Hsp90� protein was immobilized ontoan amine coated magnetic beads by reductive amination usinggluteraldehyde and sodium cyanoborohydride. In order to immo-inopropyl silica liquid chromatography stationary phase via the (a) amino terminus
M. Sanghvi et al. / J. Chromatogr. A 1218 (2011) 8791– 8798 8797
Fig. 6. Effect on the ATPase activity of an Hsp90�(CT) column of the addition ofeither the Hsp90-binding drug, geldanamycin, to the mobile phase or from diamide-mediated S-glutathionylation of the immobilized Hsp90. Calculation of the ADP/ATPrsd
bit
3
iNbucttlHtl
3
wabnttp1rwdcbrtHmawnecccpH
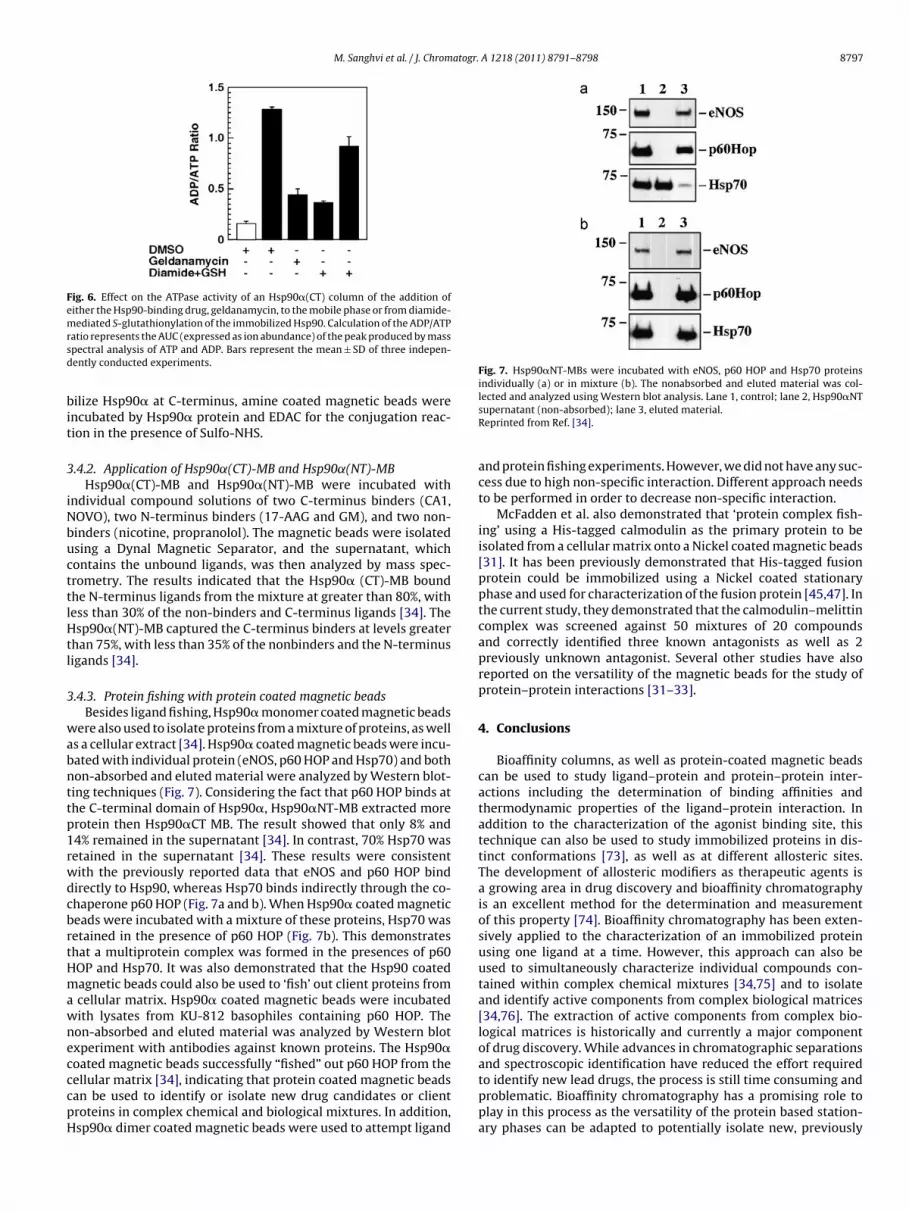
Fig. 7. Hsp90�NT-MBs were incubated with eNOS, p60 HOP and Hsp70 proteinsindividually (a) or in mixture (b). The nonabsorbed and eluted material was col-lected and analyzed using Western blot analysis. Lane 1, control; lane 2, Hsp90�NT
atio represents the AUC (expressed as ion abundance) of the peak produced by masspectral analysis of ATP and ADP. Bars represent the mean ± SD of three indepen-ently conducted experiments.
ilize Hsp90� at C-terminus, amine coated magnetic beads werencubated by Hsp90� protein and EDAC for the conjugation reac-ion in the presence of Sulfo-NHS.
.4.2. Application of Hsp90˛(CT)-MB and Hsp90˛(NT)-MBHsp90�(CT)-MB and Hsp90�(NT)-MB were incubated with
ndividual compound solutions of two C-terminus binders (CA1,OVO), two N-terminus binders (17-AAG and GM), and two non-inders (nicotine, propranolol). The magnetic beads were isolatedsing a Dynal Magnetic Separator, and the supernatant, whichontains the unbound ligands, was then analyzed by mass spec-rometry. The results indicated that the Hsp90� (CT)-MB boundhe N-terminus ligands from the mixture at greater than 80%, withess than 30% of the non-binders and C-terminus ligands [34]. Thesp90�(NT)-MB captured the C-terminus binders at levels greater
han 75%, with less than 35% of the nonbinders and the N-terminusigands [34].
.4.3. Protein fishing with protein coated magnetic beadsBesides ligand fishing, Hsp90� monomer coated magnetic beads
ere also used to isolate proteins from a mixture of proteins, as wells a cellular extract [34]. Hsp90� coated magnetic beads were incu-ated with individual protein (eNOS, p60 HOP and Hsp70) and bothon-absorbed and eluted material were analyzed by Western blot-ing techniques (Fig. 7). Considering the fact that p60 HOP binds athe C-terminal domain of Hsp90�, Hsp90�NT-MB extracted morerotein then Hsp90�CT MB. The result showed that only 8% and4% remained in the supernatant [34]. In contrast, 70% Hsp70 wasetained in the supernatant [34]. These results were consistentith the previously reported data that eNOS and p60 HOP bindirectly to Hsp90, whereas Hsp70 binds indirectly through the co-haperone p60 HOP (Fig. 7a and b). When Hsp90� coated magneticeads were incubated with a mixture of these proteins, Hsp70 wasetained in the presence of p60 HOP (Fig. 7b). This demonstrateshat a multiprotein complex was formed in the presences of p60OP and Hsp70. It was also demonstrated that the Hsp90 coatedagnetic beads could also be used to ‘fish’ out client proteins from
cellular matrix. Hsp90� coated magnetic beads were incubatedith lysates from KU-812 basophiles containing p60 HOP. Theon-absorbed and eluted material was analyzed by Western blotxperiment with antibodies against known proteins. The Hsp90�oated magnetic beads successfully “fished” out p60 HOP from the
ellular matrix [34], indicating that protein coated magnetic beadsan be used to identify or isolate new drug candidates or clientroteins in complex chemical and biological mixtures. In addition,sp90� dimer coated magnetic beads were used to attempt ligandsupernatant (non-absorbed); lane 3, eluted material.Reprinted from Ref. [34].
and protein fishing experiments. However, we did not have any suc-cess due to high non-specific interaction. Different approach needsto be performed in order to decrease non-specific interaction.
McFadden et al. also demonstrated that ‘protein complex fish-ing’ using a His-tagged calmodulin as the primary protein to beisolated from a cellular matrix onto a Nickel coated magnetic beads[31]. It has been previously demonstrated that His-tagged fusionprotein could be immobilized using a Nickel coated stationaryphase and used for characterization of the fusion protein [45,47]. Inthe current study, they demonstrated that the calmodulin–melittincomplex was screened against 50 mixtures of 20 compoundsand correctly identified three known antagonists as well as 2previously unknown antagonist. Several other studies have alsoreported on the versatility of the magnetic beads for the study ofprotein–protein interactions [31–33].
4. Conclusions
Bioaffinity columns, as well as protein-coated magnetic beadscan be used to study ligand–protein and protein–protein inter-actions including the determination of binding affinities andthermodynamic properties of the ligand–protein interaction. Inaddition to the characterization of the agonist binding site, thistechnique can also be used to study immobilized proteins in dis-tinct conformations [73], as well as at different allosteric sites.The development of allosteric modifiers as therapeutic agents isa growing area in drug discovery and bioaffinity chromatographyis an excellent method for the determination and measurementof this property [74]. Bioaffinity chromatography has been exten-sively applied to the characterization of an immobilized proteinusing one ligand at a time. However, this approach can also beused to simultaneously characterize individual compounds con-tained within complex chemical mixtures [34,75] and to isolateand identify active components from complex biological matrices[34,76]. The extraction of active components from complex bio-logical matrices is historically and currently a major componentof drug discovery. While advances in chromatographic separationsand spectroscopic identification have reduced the effort required
to identify new lead drugs, the process is still time consuming andproblematic. Bioaffinity chromatography has a promising role toplay in this process as the versatility of the protein based station-ary phases can be adapted to potentially isolate new, previously
8 atogr.
uttb
A
gc
R
[[[
[[[[
[
[[
[
[
[
[
[[[[[[[[[
[[
[
[[
[
[[[[[[[
[[[[[
[
[
[[[[[[
[
[
[
[[[[
[
[[
[
[
[
[
[[[
S. Daniele, G. Caccialanza, D. Lecca, C. Lambertucci, M.L. Trincavelli, G. Marucci,
798 M. Sanghvi et al. / J. Chrom
nknown, therapeutic drugs. In addition, to the columns, our ini-ial work with the protein-coated magnetic beads demonstrateshat this technique may have a greater advantage than the protein-ased stationary phase in this regard.
cknowledgements
This research was supported by the Intramural Research Pro-ram of the NIH, National Institute on Aging. There are no financialonflicts.
eferences
[1] R. Curtis, A.J. Ulrich, A. Montaser, J.M. Prausnitz, H.W. Blanch, Biotechnol. Bio-eng. 79 (2002) 367.
[2] R. Piazza, J. Cryst. Growth 196 (1999) 415.[3] J. Behlke, O. Ristau, Biophys. Chem. 76 (1999) 13.[4] D. Vivares, F. Bonnete, Acta Crystallogr. D 58 (2002) 472.[5] P.M. Tessier, A.M. Lenhoff, S.I. Sandler, Biophys. J. 82 (2002) 1620.[6] A.C. Dumetz, A.M. Snellinger-O’Brien, E.W. Kaler, A.M. Lenhoff, Protein Sci. 16
(2007) 1867.[7] J. Bloustine, V. Berejnov, S. Fraden, Biophys. J. 85 (2003) 2619.[8] H. Bajaj, V.K. Sharma, D.S. Kalonia, Biophys. J. 87 (2004) 4048.[9] C.D. Garica, D.J. Hadley, W.W. Wilson, C.S. Henry, Biotechnol. Prog. 19 (2003)
1006.10] L.W. Nichol, R.J. Siezen, D.J. Winzor, Biophys. Chem. 9 (1978) 47.11] P.R. Willis, L.W. Nichol, R.J. Siezen, Biophys. Chem. 11 (1980) 71.12] C. Gripon, L. Legrand, I. Rosenman, O. Vidal, M.C. Robert, F. Boue, J. Cryst. Growth
178 (1997) 575.13] A.M. Kulkarni, Master’s Thesis, Uni of Illinois, Urbana-Champaign, IL.14] O.D. Veleve, E.W. Kaler, A.M. Lenhoff, Biophys. J. 75 (1998) 2682.15] G. MacBeath, S.L. Schreiber, Science 289 (2000) 1760.16] H. Zhu, M. Bilgin, R. Bangham, D. Hall, A. Casamayor, P. Bertone, N. Lan, R.
Jansen, S. Bidlingmaier, T. Houfek, T. Mitchell, P. Miller, R.A. Dean, M. Gerstein,M. Snyder, Science 293 (2001) 2101.
17] G. Wu, R.H. Datar, K.M. Hansen, T. Thundat, R.J. Cote, A. Majumdar, Nat. Biotech-nol. 19 (2001) 856.
18] V.K. Yadavalli, J.G. Forbes, K. Wang, Langmuir 22 (2006) 6969.19] P. Arenkov, A. Kukhtin, A. Gemmell, S. Voloshchuk, V. Chupeeva, A. Mirzbekov,
Anal. Biochem. 278 (2000) 123.20] A. Espejo, J. Cote, A. Bednarek, S. Richard, M.T. Bedford, Biochem. J. 367 (2002)
697.21] D. Stoll, M.F. Templin, J. Bachmann, T.O. Joos, Curr. Opin. Drug Discov. Dev. 8
(2005) 239.22] V. Knezevic, C. Leethanakul, V.E. Bichsel, J.M. Worth, V.V. Prabhu, J.S. Gutkind,
L.A. Liotta, P.J. Munson, E.F. Petricoin, Proteomics 1 (2001) 1271.23] J.C. Miller, H. Zhou, J. Kwekel, R. Cavallo, J. Burke, E.B. Butler, B.S. Teh, B.B. Haab,
Proteomics 3 (2003) 56.24] E. Brekkan, A. Lundqvist, P. Lundahl, Biochemistry 35 (1996) 12141.25] Q. Yang, P. Lundahl, Biochemistry 34 (1995) 7289.26] I.M. Chaiken, J. Chromatogr. 376 (1986) 11.27] J.L. Wade, A.F. Bergold, P.W. Carr, Anal. Chem. 59 (1987) 1286.28] R. Moaddel, I.W. Wainer, Nat. Protoc. 4 (2009) 197.29] R. Moaddel, K. Jozwiak, I.W. Wainer, Med. Res. Rev. 27 (2007) 723.30] K.D. Belanger, Life Sci. Edu. 8 (2009) 214.31] M.J. McFadden, M.S. Junop, J.D. Brennan, Anal. Chem. 82 (2010) 9850.
32] N. Jonker, A. Krestchmer, J. Kool, A. Fernandez, D. Kloos, J.G. Krabbe, H. Linge-man, H. Irth, Anal. Chem. 81 (2009) 4263.33] J.S. Li, J.P. Ge, Y.D. Yin, W.W. Zhong, Anal. Chem. 80 (2008) 7068.34] M.P. Marszall, R. Moaddel, S. Kole, M. Gandhari, M. Bernier, I.W. Wainer, Anal.
Chem. 80 (2008) 7571.
[
A 1218 (2011) 8791– 8798
35] R. Moaddel, M.P. Marszall, F. Bighi, Q. Yang, X. Duan, I.W. Wainer, Anal. Chem.79 (2007) 5414.
36] L.S. Wong, F. Khan, J. Micklefield, Chem. Rev. 109 (2009) 4025.37] M. Sanghvi, R. Moaddel, C. Frazier, I.W. Wainer, J. Pharm. Biomed. Anal. 53
(2010) 777.38] M.P. Marszall, R. Moaddel, K. Jozwiak, M. Bernier, I.W. Wainer, Anal. Biochem.
373 (2008) 313.39] P.O. Larsson, Methods Enzymol. 104 (1984) 212.40] U. Erler, G. Heublein, J. Chromatogr. 588 (1991) 340.41] Q. Zhang, R. Hunag, L. Guo, Chin. Sci. Bull. 54 (2009) 2620.42] R. Axen, J. Porath, S. Ernback, Nature 214 (1967) 1302.43] J. Kohn, M. Wilchek, Biochem. Biophys. Res. Commun. 197 (1982) 878.44] R. Paul, G.W. Anderson, J. Chem. Soc. 82 (1960) 4596.45] R. Moaddel, G.B. Price, J.M. Juteau, M. Leffak, I.W. Wainer, J. Chromatogr. B 820
(2005) 197.46] D.B. Smith, K.S. Johnson, Gene 67 (1988) 31.47] R. Moaddel, L. Lu, M. Baynham, I.W. Wainer, J. Chromatogr. B 768 (2002) 41.48] R. Moaddel, P.I. Bullock, I.W. Wainer, J. Chromatogr. B 799 (2004) 255.49] E. Calleri, C. Temporini, G. Massolini, J. Pharm. Biomed. Anal. 54 (2011) 911.50] E.S.M. Ng, N.W.C. Chora, D.F. Lewis, O. Hindsgaul, D. Schriemer, Nat. Protoc. 2
(2007) 1907.51] E. Calleri, C. Temporini, G. Caccialanza, G. Massolini, Chem. Med. Chem. 4 (2009)
905.52] N.W. Chan, D.F. Lewis, S. Hewko, O. Hindsgaul, D.C. Schriemer, Comb. Chem.
High Throughput Screen 5 (2002) 395.53] J.E. Schiel, K.S. Joseph, D.S. Hage, Adv. Chromatogr. 48 (2010) 145.54] K. Jozwiak, J. Haginaka, R. Moaddel, I.W. Wainer, Anal. Chem. 74 (2002) 4618.55] J. Wade, A.F. Bergold, P.W. Carr, Anal. Chem. 59 (1987) 1286.56] A. Jaulmes, C. Vidal-Madjar, Adv. Chromatogr. 28 (1989) 1.57] V. Gigue‘re, N. Yang, P. Segui, R.M. Evans, Nature 331 (1988) 91.58] J.D. Eudy, S. Yao, M.D. Weston, M. Ma-Edmonds, C.B. Talmage, J.J. Cheng, W.J.
Kimberling, J. Sumegi, Genomics 50 (1998) 382.59] M. Suetsugi, L. Su, K. Karlsberg, Y.C. Yuan, S. Chen, Mol. Cancer Res. 1 (2003)
981.60] P. Coward, D. Lee, M.V. Hull, J. Lehmann, Proc. Natl. Acad. Sci. U. S. A 98 (2001)
8880.61] G.B. Tremblay, T. Kunath, D. Bergeron, L. Lapointe, C. Champigny, J.A. Bader, J.
Rossant, V. Giguere, Genes Dev. 15 (2001) 833.62] G.B. Tremblay, D. Bergeron, V. Giguere, Endocrinology 142 (2001) 4572.63] H. Greschik, R. Flaig, J. Renaud, D. Moras, J. Biol. Chem. 279 (2004) 33639.64] L. Whitesell, S.L. Lindquist, Nat. Rev. Cancer 5 (2005) 671.65] A. Shervington, N. Cruickshanks, R. Lea, G. Roberts, T. Dawson, L. Shervington,
Cancer Invest. 26 (2008) 900.66] A. Kamal, L. Thao, J. Sensintaffar, L. Zhang, M.F. Boehm, L.C. Fritz, F.J. Burrows,
Nature 425 (2003) 407.67] P. Meyer, Mol. Cell 1 (2003) 647.68] S.H. McLaughlin, L.A. Ventouras, B. Lobbezoo, S.E. Jackson, J. Mol. Biol. 344
(2004) 813.69] M.R. Rowlands, Y.M. Newbatt, C. Prodromou, L.H. Pearl, P. Workman, W. Aherne,
Anal. Biochem. 327 (2004) 176.70] S.K. Calderwood, M.D. Khaleque, D.B. Sawyer, D.R. Ciocca, Trends Biochem. Sci.
31 (2006) 164.71] S. Song, S. Kole, P. Precht, M.J. Pazin, M. Bernier, Int. J. Biochem. Cell Biol. 42
(2010) 1856.72] J.F. Caplan, N.R. Filipenko, S.L. Fitzpatrick, D.M. Waisman, J. Biol. Chem. 279
(2004) 7740.73] R. Moaddel, I.W. Wainer, Anal. Chim. Acta 564 (2006) 97.74] J. Chen, D.S. Hage, Nat. Biotechnol. 22 (2004) 1445.75] E. Calleri, S. Ceruti, G. Cristalli, C. Martini, C. Temporini, C. Parravicini, R. Volpini,
I.W. Wainer, G. Ranghino, P. Fantucci, M.P. Abbracchio, G. Massolini, J. Med.Chem. 53 (2010) 3489.
76] A. Maciuk, R. Moaddel, J. Haginaka, I.W. Wainer, J. Pharm. Biomed. Anal. 48(2008) 238.





























