Embed Size (px)
Citation preview

ORIGINAL ARTICLE
The earliest record of the genus Cola (Malvaceae sensu lato:Sterculioideae) from the Late Oligocene (28–27 Ma) of Ethiopiaand leaf characteristics within the genus
Aaron David Pan • Bonnie F. Jacobs
Received: 12 September 2007 / Accepted: 14 September 2009 / Published online: 10 October 2009
� Springer-Verlag 2009
Abstract A fossil leaf compression from the Late
Oligocene (28–27 Ma) of northwestern Ethiopia is the
earliest record of the African endemic moist tropical forest
genus Cola (Malvaceae sensu lato: Sterculioideae). Based
on leaf and epidermal morphology, the fossil is considered
to be very similar to two extant Guineo-Congolian species
but differences warrant designation of a new species. This
study also includes a review of the fossil record of Cola, a
comprehensive summary of leaf characteristics within
several extant species of Cola, Octolobus, and Pterygota,
and a brief discussion of the paleogeographic implications
of the fossil species affinity and occurrence in Ethiopia.
Keywords Cola � Cuticle � Octolobus � Pterygota �Sterculiaceae
Introduction
The Malvaceae sensu lato (which includes Bombacaceae,
Malvaceae sensu stricto, Sterculiaceae, and Tiliaceae) are
major woody components of present-day African tropical
moist forests (Judd and Manchester 1997; Alverson et al.
1999). For example, in most 0.1 ha African forest plots
examined by Gentry (1988), the Sterculiaceae or Tiliaceae
were almost always among the top ten most species-rich
angiosperm families (Phillips and Miller 2002). In these
plots, the most speciose group within Malvaceae s.l. is a
subclade of the Cola clade sensu Wilkie et al. (2006)
consisting of Cola Schott & Endl., Octolobus Welw., and
Pterygota Schott & Endl.
The African endemic Cola, the second largest genus
(approximately 100–125 species) in the subfamily Stercu-
lioideae, is almost entirely restricted to moist evergreen
and semideciduous forest formations (Cheek 2002; Bayer
and Kubitzki 2003; Judd et al. 2008). The genus, which
typically is a common component of the forest understorey,
includes shrubs, treelets, and trees that rarely reach more
than 30 m in height. While the majority of Cola species are
found in West and Central Africa, about 20 species occur
in eastern (Uganda, Tanzania, Kenya, and southern
Somalia), northern (Mali, Niger, and southern Sudan), and
southern Africa (Malawi, Mozambique, South Africa, and
Zambia) (Fig. 1; Keay and Brenan 1973; Verdoorn 1981;
White et al. 2001; Cheek 2002; Lebrun and Stork 2003;
Cheek personal communication).
Octolobus is closely related, and possibly sister to Cola
(Cheek and Frimodt-Møller 1998; Wilkie et al. 2006). This
African endemic tri- or tetratypic genus consists of small
forest trees found mainly in West and Central Africa,
although one species reaches Tanzania (Cheek and
Frimodt-Møller 1998; Bayer and Kubitzki 2003). Both
genera are similar in possessing indehiscent follicles and
exalbuminous seeds (Bayer and Kubitzki 2003; Wilkie
et al. 2006). Pterygota, a pantropical genus of about 15
species of tall forest trees, differs from both Cola and
Octolobus in possessing dehiscent follicle fruits with
winged seeds (Mabberley 1997; Bayer and Kubitzki 2003;
Lebrun and Stork 2003).
A. D. Pan (&)
Fort Worth Museum of Science and History,
1600 Gendy Street, Fort Worth, TX 76107-4062, USA
e-mail: [email protected]
B. F. Jacobs
Roy M. Huffington Department of Earth Sciences,
Southern Methodist University, 750395, Dallas,
TX 75275-0395, USA
e-mail: [email protected]
123
Plant Syst Evol (2009) 283:247–262
DOI 10.1007/s00606-009-0225-1

A recently discovered fossil leaf compression from the
Late Oligocene of Ethiopia (28–27 Ma) Guang River flora
can be identified confidently as Cola and is described
below. This fossil represents the oldest record of the genus.
The presence of a fossil Cola from Ethiopia is interesting
biogeographically because the genus is not present there
today (Friis 1992; Vollesen 1995) and indicates a different
distribution on the Afro-Arabian continent during the
Paleogene (Fig. 1). In addition, the genus is an indicator of
moist forest communities and thus provides insight into the
Late Oligocene vegetation on the northwestern Ethiopian
plateau.
Very little information on the epidermal anatomy of
living Cola has been published and no data are available
regarding the leaf epidermal anatomy of Octolobus or
Pterygota. The only sources of data about epidermal
micromorphology within Cola are Gehrig (1938), Metcalfe
and Chalk (1950), and Inamdar et al. (1983), none of which
are comprehensive. Consequently, leaf cuticle samples
from several extant species of Cola and some species of
Octolobus and Pterygota were prepared, examined, and
described to provide comparative epidermal micromor-
phological characters. These data have proven useful in
identification of the fossil and are provided here for future
systematic and phylogenetic investigations of the genus
and the Sterculioideae (Table 1 and Appendix).
Materials and methods
Cuticle was prepared from fossil leaf fragments (0.5–
1 cm2) by first rinsing the material in 10% hydrochloric
acid (HCl) for 10–30 min to remove carbonates. The leaf
material was then rinsed with distilled water and placed in
a solution of 48% hydrofluoric acid (HF) for 24–48 h to
dissolve the silicate matrix. Following this step, the cuticle
was placed in a 20% bleach (NaClO)–water solution for a
very brief amount of time (on the order of 20 s to 1 min) to
dissolve away excess organics. The cuticle was thoroughly
rinsed with distilled water and mounted on microscope
slides in glycerine.
Leaf cuticle samples from 38 extant species of Cola, 2
species of Octolobus, and 2 species of Pterygota were
sampled for comparative purposes from herbaria at the
Missouri Botanical Garden (MO) and the Royal Botanic
Gardens at Kew (K). Samples (0.5–1 cm2) were soaked in
distilled water for 24 h to resaturate. Subsequently, the
specimens were treated with a solution of 10% potassium
hydroxide (KOH) until they darkened fully, washed with
distilled water, and placed in baths of 30–60% bleach to
remove mesophyll. The cleared specimens were then
stained with a Safranine O solution, rinsed with 90%
ethanol (C2H5OH), and mounted on microscope slides in
glycerine jelly. Light microscope slides of both the fossil
and herbarium samples are housed in the Roy M.
Huffington Department of Earth Sciences at Southern
Methodist University in Dallas, TX.
Most of the terminology and definitions used in the text
and Appendix are taken from the Manual of Leaf Archi-
tecture (Ellis et al. 2009) and Dilcher (1974). Where terms
differ, they are defined in the text. Dominant abaxial
and adaxial trichome characters (characters 26 and 41 in
Table 1 and Appendix) were determined by randomly
observing 100 hairs among the prepared cuticle microscope
slides of each species. In cases where fewer than 100 hairs
were present on a slide, trichome dominance was based on
a percentage of fewer than 100 randomly observed hairs. If
the number of hairs observed was fewer than 20, and there
were more than two hair types present the character state
was scored as ‘‘no trichomes predominate.’’ Epidermal cell
sizes are recorded in measured micrometer ranges (lm;
characters 14 and 29 in Table 1, Appendix). For these
measurements only nonsubsidiary epidermal cells that
were not adjacent to veins or hairs were included.
The Guang River flora
The newly discovered Ethiopian Late Oligocene (28–
27 Ma) fossil is from a moderately diverse assemblage
(approximately 40 morphotypes, assumed to be species) of
well-preserved leaf, floral, and wood (logs and stumps)
Fig. 1 Distribution map of the genus Cola, modified from Lebrun
and Stork (2003) and Coates Palgrave (1977). Geometric shapes
correspond to the number of species that occur (or co-occur) within a
particular area: circles 1 or 2 species, triangles 3–5 species, squares6–9 species; pentagons, 10 or more species. Daggers denote
confirmed and purported fossil records of Cola
248 A. D. Pan, B. F. Jacobs
123

Ta
ble
1L
eaf
mo
rph
olo
gy
and
epid
erm
alm
icro
mo
rph
olo
gy
char
acte
rist
ics
wit
hin
Co
la,
Oct
olo
bu
s,an
dP
tery
go
ta
Sp
ecie
s1
23
45
67
89
10
11
12
13
14
15
Co
laa
cum
ina
ta(P
.B
eau
v.)
Sch
ott
&E
nd
l.0
00
10
00
,11
,21
10
20
?1
22
.5–
55
;1
9–
35
.50
Co
laa
ltis
sim
aE
ng
l.0
0,1
01
00
11
11
02
0?
11
3–
54
.5;
9.5
–2
2.5
0
Co
laa
mh
ara
ensi
ssp
.n
ov
.A
.D.
Pan
&B
.F.
Jaco
bs*
0?
01
00
0?
11
02
0?
11
6.5
–5
3;
9.5
–3
5.5
0
Co
laa
ttie
nsi
sA
ub
rev
.&
Pel
leg
r.0
0,1
,30
10
00
,11
,21
1,2
02
0?
11
9–
51
.5;
16
–2
60
Co
lab
all
ayi
Co
rnu
exH
eck
el0
0,2
,30
10
00
,11
,21
10
20
?1
16
–5
3;
8–
29
0
Co
lab
un
tin
gii
Bak
.f.
11
01
00
12
0?
02
0?
11
1–
64
.5;
9.5
–2
90
Co
laca
rica
efo
lia
(G.
Do
n)
K.
Sch
um
.0
0,1
00
12
31
,22
10
20
?1
19
–6
4.5
;1
3–
45
0
Co
lach
lam
yda
nth
aK
.S
chu
m.
10
,11
20
01
20
1,2
02
0?
11
4.5
–4
5;
16
–3
10
Co
lacl
ava
taM
ast.
01
01
00
01
10
,10
20
?1
12
.5–
32
.5;
9.5
–1
60
Co
laco
rdif
oli
a(C
av.)
R.
Br.
00
,20
10
,12
30
,1,2
1,2
0,1
02
0?
16
.5–
16
.5;
6.5
–1
30
Co
lad
igit
ata
Mas
t.1
00
11
00
11
0,1
02
0?
19
.5–
50
;6
.5–
32
.50
Co
lad
isco
gly
pre
mn
op
hyl
laB
ren
an&
A.
P.
D.
Jon
es0
00
20
0,2
1,3
?1
00
20
?1
16
–3
4;
8–
27
.50
Co
lafi
cifo
lia
Mas
t.0
0,1
01
10
,1,2
1,3
11
,21
,2,3
02
0?
12
5.5
–7
7.5
;9
.5–
42
0
Co
lafl
avo
-vel
uti
na
K.
Sch
um
.0
00
10
00
21
10
20
?1
16
–5
9.5
;9
.5–
35
.50
Co
lag
ab
on
ensi
sM
ast.
,n
on
sen
suA
ub
rev
.,n
ecse
nsu
M.
Bo
dar
d0
10
00
01
1,2
0,1
10
20
?1
35
.5–
61
;1
9–
42
0
Co
lag
reen
wa
yiB
ren
an0
0,1
01
00
11
,20
,11
,20
20
?1
16
–4
0;
9.5
–2
40
Co
lah
eter
op
hyl
la(P
.B
eau
v.)
Sch
ott
&E
nd
l.0
0,1
00
0,1
0,1
1,3
21
,20
,10
20
?1
24
–7
1;
22
.5–
35
0
Co
lah
isp
ida
Bre
nan
&K
eay
01
00
12
31
21
02
0?
13
2–
84
;2
9–
48
.50
Co
lala
teri
tia
K.
Sch
um
.0
0,1
,20
10
,12
30
,12
10
20
?1
24
–3
8.5
;9
.5–
32
.50
Co
lala
uri
foli
aM
ast.
00
01
00
10
,1
00
,10
20
?1
8–
32
.5;
6–
22
.50
Co
lale
pid
ota
K.
Sch
um
.1
0,2
11
00
0,1
10
10
20
?1
16
–6
5.5
;1
2.5
–3
90
Co
lale
tou
zeya
na
Nk
on
gm
enec
k0
11
10
1,2
31
,22
10
20
?1
29
–6
8;
19
.5–
29
0
Co
lali
ssa
chen
sis
Pel
leg
r.1
11
10
00
20
2,3
02
0?
11
3–
45
;1
3–
29
0
Co
lam
ah
ou
nd
ensi
sP
elle
gr.
00
,10
01
13
12
10
20
?1
22
.5–
77
.5;
14
.5–
32
.50
Co
lam
ars
up
ium
K.
Sch
um
.0
0,1
00
0,1
23
22
0,1
02
0?
12
9–
10
0;
16
–4
70
Co
lam
ille
nii
K.
Sch
um
.0
0,1
00
01
,21
,31
1,2
10
20
?1
12
.5–
58
;4
.5–
29
0
Co
lam
ino
rB
ren
an0
0,1
01
00
01
01
,20
20
?1
9.5
–3
9;
9.5
–1
9.5
0
Co
lam
oss
am
bic
ensi
sW
ild
00
,20
10
01
0,1
11
02
0?
19
.5–
32
.5;
6–
16
0
Co
lan
ata
len
sis
Oli
ver
00
,10
10
01
1,2
01
02
0?
19
.5–
39
;9
.5–
19
.50
Co
lan
iger
ica
Bre
nan
&K
eay
00
01
00
0,1
21
10
20
?1
20
.5–
58
;1
2.5
–3
90
Co
lan
itid
a(V
ent.
)S
cho
tt&
En
dl.
00
,10
10
01
1,2
10
,10
20
?1
22
.5–
58
;8
–3
10
Co
lap
ach
yca
rpa
K.
Sch
um
.1
0,1
02
0?
20
11
,20
20
20
?1
16
–6
1;
12
.5–
21
0
Co
lap
orp
hyr
an
tha
Bre
nan
00
,10
10
01
20
,10
,10
20
?1
9.5
–5
2;
8–
19
.50
Co
lasc
hef
fler
iK
.S
chu
m.
00
,20
00
,11
,21
,31
1,2
0,1
02
0?
12
2.5
–7
1;
12
.5–
53
.50
Co
lasi
mia
rum
Sp
rag
ue
exB
ren
an&
Kea
y0
0,3
00
20
12
10
,10
20
?1
9.5
–3
7;
6.5
–2
2.5
0
Co
last
elec
ha
nth
aB
ren
an0
0,1
01
00
11
,20
,11
02
0?
11
6–
68
;9
.5–
31
0
The earliest record of the genus Cola from the Late Oligocene 249
123

Ta
ble
1co
nti
nu
ed
Sp
ecie
s1
23
45
67
89
10
11
12
13
14
15
Co
lau
lolo
ma
Bre
nan
00
,11
10
01
1,2
00
,10
20
?1
15
.5–
42
;1
3–
23
0
Co
lau
sam
ba
ren
sis
En
gl.
00
01
00
11
,21
0,1
02
0?
11
6–
55
;9
.5–
22
.50
Oct
olo
bu
ssp
ecta
bil
isW
elw
.0
0,1
01
00
12
0,1
10
20
?1
19
–4
5;
12
.5–
29
0
Oct
olo
bu
sze
nke
riE
ng
l.0
00
10
01
21
10
20
?1
12
.5–
37
;9
.5–
19
.50
Pte
ryg
ota
beq
ua
erti
iD
eW
ild
.0
20
10
1,2
2,3
1,2
20
,10
20
?1
16
–4
8.5
;1
6–
22
.50
Pte
ryg
ota
ma
cro
carp
aK
.S
chu
m.
02
01
02
30
,12
0,1
02
0?
11
2.5
–5
5;
9.5
–3
5.5
0
Sp
ecie
s1
61
71
81
92
02
12
22
32
42
52
62
72
82
93
0
Co
laa
cum
ina
ta0
?1
–1
11
?2
?3
00
02
(?3
)(?
0?
)2?
30
10
?1
9.5
–4
3.5
;8
–2
7.5
0
Co
laa
ltis
sim
aE
ng
l.0
?1
–1
11
?2
?3
00
02
2?
30
00
?1
22
.5–
48
.5;
12
.5–
34
0
Co
laa
mh
ara
ensi
s*0
?1
–1
12
00
01
20
00
?1
24
–4
2;
14
.5–
31
0
Co
laa
ttie
nsi
s0
?1
–1
12
00
01
20
00
?1
16
–3
9;
14
.5–
21
0
Co
lab
all
ayi
0?
1–
11
1?
2?
30
00
12
00
0?
11
1–
39
;1
4.5
–2
60
Co
lab
un
tin
gii
1?
20
11
1?
30
00
22
?3
00
0?
11
6–
42
;1
3–
16
0
Co
laca
rici
foli
a1
?2
01
11
?2
?3
00
01
20
00
?1
16
–5
5;
12
.5–
35
.50
Co
lach
lam
yda
nth
a1
?2
01
10
?1
?2
01
,30
21
?3
00
0?
16
.5–
29
;4
.5–
18
0
Co
lacl
ava
ta1
–1
12
00
02
2?
31
00
?1
9.5
–2
4;
3–
19
.50
Co
laco
rdif
oli
a1
–1
12
00
02
2?
32
00
?1
6–
22
.5;
6–
16
0
Co
lad
igit
ata
0?
1?
20
11
10
00
22
?3
00
0?
11
1–
35
.5;
11
–2
2.5
0
Co
lad
isco
gly
pre
mn
op
hyl
la0
?1
–1
12
00
01
20
10
?1
16
–3
5.5
;1
1–
27
.50
Co
lafi
cifo
lia
1?
20
11
1?
20
30
22
?3
00
0?
11
9–
45
;1
2.5
–2
7.5
0
Co
lafl
avo
-vel
uti
na
0?
1–
11
20
00
22
?3
00
0?
11
9–
39
;1
6–
24
.50
Co
lag
ab
on
ensi
s2
01
12
00
02
2?
30
00
?1
25
.5–
64
.5;
22
.5–
42
0
Co
lag
reen
wa
yi1
?2
01
12
00
02
2?
32
10
?1
16
–3
9;
11
–2
90
Co
lah
eter
op
hyl
la1
?2
01
11
?2
00
02
2?
30
00
?1
39
–6
8;
35
–4
00
Co
lah
isp
ida
1?
20
11
20
00
22
?3
00
0?
13
8.5
–8
2;
25
.5–
51
.50
Co
lala
teri
tia
0?
1–
11
0?
1?
2?
30
00
12
00
0?
12
7–
38
.5;
8–
32
0
Co
lala
uri
foli
a0
?1
–1
10
00
02
2?
31
00
?1
11
–3
1;
6.5
–1
9.5
0
Co
lale
pid
ota
0?
1–
21
1?
20
1,3
02
2?
31
00
?1
13
–3
2;
13
–2
40
Co
lale
tou
zeya
na
1?
20
11
20
00
22
?3
01
0?
11
9–
40
;1
6–
29
0
Co
lali
ssa
chen
sis
0?
1–
11
0?
1?
2?
30
1,3
02
2?
31
00
?1
11
–2
5.5
;1
1–
22
.50
Co
lam
ah
ou
nd
ensi
s2
01
11
?2
00
03
0?
2?
31
00
?1
22
.5–
58
;2
2.5
–3
90
Co
lam
ars
up
ium
1?
20
11
20
00
12
00
0?
12
5.5
–5
5;
16
–2
90
Co
lam
ille
nii
0?
1–
11
1?
2?
30
00
30
?2
?3
00
0?
12
5.5
–5
1.5
;9
.5–
42
0
Co
lam
ino
r0
?1
–1
12
00
01
32
00
?1
9.5
–2
9;
6.5
–1
80
Co
lam
oss
am
bic
ensi
s0
?1
–1
12
00
02
2?
30
00
?1
12
.5–
26
;6
–1
9.5
0
250 A. D. Pan, B. F. Jacobs
123

Ta
ble
1co
nti
nu
ed
Sp
ecie
s1
61
71
81
92
02
12
22
32
42
52
62
72
82
93
0
Co
lan
ata
len
sis
1?
20
11
20
00
13
20
0?
19
.5–
32
.5;
9.5
–1
60
Co
lan
iger
ica
0–
11
1?
30
00
12
00
0?
11
6–
52
;1
2.5
–4
0.5
0
Co
lan
itid
a0
–1
11
?2
?3
00
02
2?
30
00
?1
12
.5–
42
;9
.5–
26
0
Co
lap
ach
yca
rpa
0?
1–
11
10
1,3
02
2?
30
00
?1
16
–4
5;
12
.5–
29
0
Co
lap
orp
hyr
an
tha
1?
20
11
1?
20
00
22
?3
00
0?
19
.5–
22
.5;
8–
21
0
Co
lasc
hef
fler
i2
01
11
?2
?3
00
01
20
00
?1
38
.5–
68
;1
9–
42
0
Co
lasi
mia
rum
0–
11
0?
1?
20
10
12
00
0?
19
.5–
51
.5;
6–
42
0
Co
last
elec
ha
nth
a0
?1
–1
11
?2
00
02
2?
30
00
?1
25
–5
8;
12
.5–
32
.50
Co
lau
lolo
ma
1–
11
20
00
22
?3
00
0?
11
6–
33
;1
2.5
–3
10
Co
lau
sam
ba
ren
sis
0?
1–
11
20
00
13
20
0?
11
6–
39
;1
2.5
–2
40
Oct
olo
bu
ssp
ecta
bil
is0
?1
–1
12
10
02
2?
31
00
?1
14
.5–
43
.5;
12
–3
70
Oct
olo
bu
sze
nke
ri0
?1
?2
01
12
00
02
2?
31
10
?1
14
.5–
32
.5;
9.5
–3
00
Pte
ryg
ota
beq
ua
erti
i2
02
11
?2
?3
00
02
2?
30
00
?1
16
–3
9;
9.5
–3
5.5
0
Pte
ryg
ota
ma
cro
carp
a1
?2
01
12
00
02
2?
30
00
?1
12
.5–
32
.5;
9.5
–1
9.5
0
Sp
ecie
s3
13
23
33
43
53
63
73
83
94
04
14
2
Co
laa
cum
ina
ta0
?1
–1
0–
––
–2
2?
30
1
Co
laa
ltis
sim
a0
?1
–1
0–
––
–2
2?
31
0
Co
laa
mh
ara
ensi
s*1
?2
01
0–
––
–?
??
0
Co
laa
ttie
nsi
s0
?1?
20
10
––
––
22
?3
00
Co
lab
all
ayi
0?
1–
10
––
––
13
20
Co
lab
un
tin
gii
1?
20
10
––
––
22
?3
20
Co
laca
rici
foli
a0
?1
–1
0–
––
–1
20
0
Co
lach
lam
yda
nth
a1
–1
10
,30
00
13
20
Co
lacl
ava
ta1
–1
0–
––
–1
32
0
Co
laco
rdif
oli
a0
?1
–1
0–
––
–2
2?
31
0
Co
lad
igit
ata
0?
1–
10
––
––
22
?3
10
Co
lad
isco
gly
pre
mn
op
hyl
la1
?2
01
10
00
01
32
0
Co
lafi
cifo
lia
0?
1–
10
––
––
22
?3
20
Co
lafl
avo
-vel
uti
na
1?
20
10
––
––
12
00
Co
lag
ab
on
ensi
s0
?1
–1
0–
––
–2
2?
30
0
Co
lag
reen
wa
yi1
?2
01
0–
––
–2
2?
32
0
Co
lah
eter
op
hyl
la1
–1
12
00
01
20
0
Co
lah
isp
ida
0?
1–
11
20
00
22
?3
00
Co
lala
teri
tia
0?
1–
10
––
––
22
?3
00
Co
lala
uri
foli
a0
?1
–1
0–
––
–2
2?
32
0
The earliest record of the genus Cola from the Late Oligocene 251
123
Ta
ble
1co
nti
nu
ed
Sp
ecie
s3
13
23
33
43
53
63
73
83
94
04
14
2
Co
lale
pid
ota
0–
10
––
––
13
20
Co
lale
tou
zeya
na
1?
20
10
––
––
22
?3
20
Co
lali
ssa
chen
sis
0?
1–
11
00
00
22
?3
20
Co
lam
ah
ou
nd
ensi
s1
?2
01
0–
––
–1
20
1
Co
lam
ars
up
ium
0?
1–
10
––
––
12
00
Co
lam
ille
nii
0?
1–
10
––
––
30
?2?
30
0
Co
lam
ino
r0
?1
–1
0–
––
–1
32
0
Co
lam
oss
am
bic
ensi
s0
?1
–1
0–
––
–1
32
0
Co
lan
ata
len
sis
0?
1–
10
––
––
13
20
Co
lan
iger
ica
0–
10
––
––
13
20
Co
lan
itid
a0
?1
–1
0–
––
–2
2?
30
0
Co
lap
ach
yca
rpa
0?
1–
10
––
––
11
00
Co
lap
orp
hyr
an
tha
1?
20
11
20
00
13
20
Co
lasc
hef
fler
i2
01
0–
––
–2
0?
20
0
Co
lasi
mia
rum
0?
1–
10
––
––
0?
??
Co
last
elec
ha
nth
a0
–1
0–
––
–2
2?
31
0
Co
lau
lolo
ma
1–
10
––
––
1?
20
Co
lau
sam
ba
ren
sis
1?
20
10
––
––
13
20
Oct
olo
bu
ssp
ecta
bil
is0
?1
–1
0–
––
–1
32
0
Oct
olo
bu
sze
nke
ri1
?2
01
0–
––
–2
2?
32
0
Pte
ryg
ota
beq
ua
erti
i1
20
––
––
22
?3
10
Pte
ryg
ota
ma
cro
carp
a1
?2
01
0–
––
–2
2?
30
0
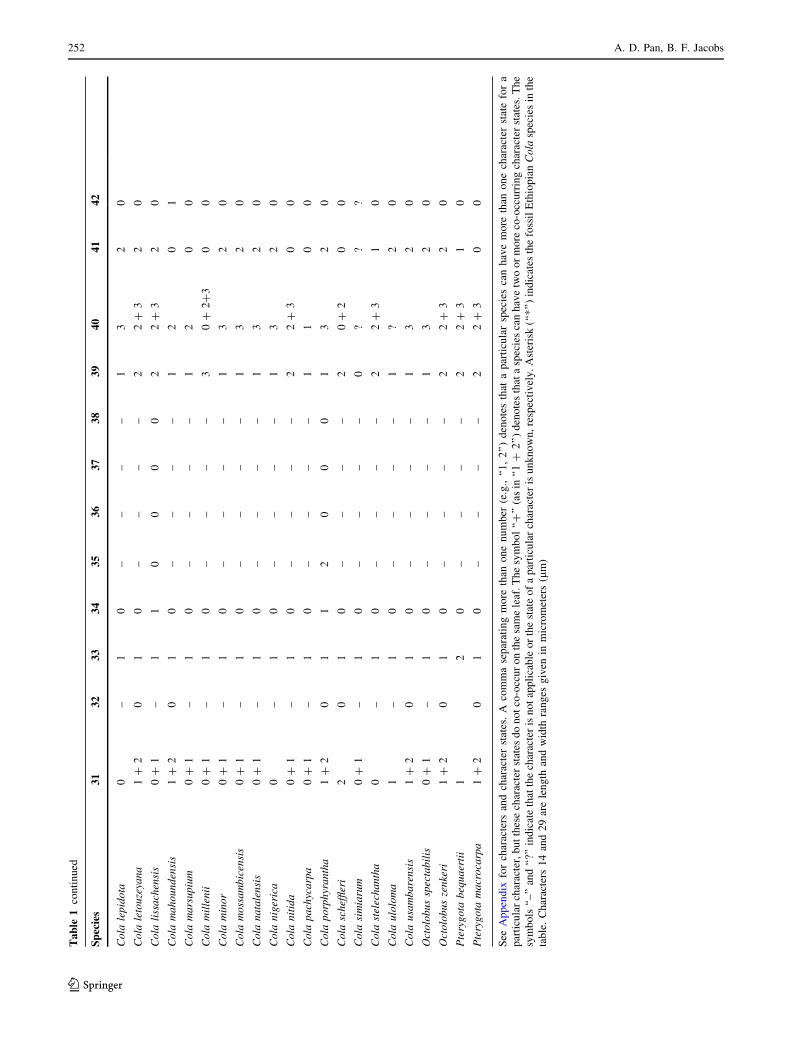
See
Ap
pen
dix
for
char
acte
rsan
dch
arac
ter
stat
es.
Aco
mm
ase
par
atin
gm
ore
than
on
en
um
ber
(e.g
.,‘‘
1,
2’’
)d
eno
tes
that
ap
arti
cula
rsp
ecie
sca
nh
ave
mo
reth
ano
ne
char
acte
rst
ate
for
a
par
ticu
lar
char
acte
r,b
ut
thes
ech
arac
ter
stat
esd
on
ot
co-o
ccu
ro
nth
esa
me
leaf
.T
he
sym
bo
l‘‘
?’’
(as
in‘‘
1?
2’’
)d
eno
tes
that
asp
ecie
sca
nh
ave
two
or
mo
reco
-occ
urr
ing
char
acte
rst
ates
.T
he
sym
bo
ls‘‘
–’’
and
‘‘?’
’in
dic
ate
that
the
char
acte
ris
no
tap
pli
cab
leo
rth
est
ate
of
ap
arti
cula
rch
arac
ter
isu
nk
no
wn
,re
spec
tiv
ely
.A
ster
isk
(‘‘*
’’)
ind
icat
esth
efo
ssil
Eth
iop
ian
Co
lasp
ecie
sin
the
tab
le.
Ch
arac
ters
14
and
29
are
len
gth
and
wid
thra
ng
esg
iven
inm
icro
met
ers
(lm
)
252 A. D. Pan, B. F. Jacobs
123

compressions, known as the Guang River flora (Pan et al.
2006; Pan 2007). The flora is located in the northwestern
plateau region of Ethiopia, about 60 km west of Gondar (in
the Chilga Woreda, Amhara region; Fig. 1). Rocks of this
region consist of massive Oligocene trap basalts inter-
spersed with tuffs, lignites, and fluvial volcaniclastic and
clastic sediments exposed along streams and gullies (Mohr
1971; Yemane et al. 1987; Kappelman et al. 2003; Jacobs
et al. 2005). Along a portion of the Guang River, a 100 m
sedimentary section has been dated to between 28 and
27 Ma (Chron C9n) based on K–Ar and 40Ar/39Ar radio-
metric dates and paleomagnetic reversal stratigraphy
(Kappelman et al. 2003; Cande and Kent 1995). Fossils are
prevalent in these sediments and include plants, mammals,
and invertebrates (Kappelman et al. 2003; Sanders et al.
2004; Pan et al. 2006; Garcıa Massini et al. 2006).
The autochthonous or parautochthonous Guang River
flora is preserved in an approximately 22-cm-thick massive
greenish-gray mudstone layer derived from a weathered
overbank (or pond) ash deposit, which provided the parent
material for an overlying Gleysol (Jacobs et al. 2005; Pan
et al. 2006; Pan 2007). The fossiliferous sediment is
underlain by a hard 1-cm-thick lignitic layer and overlain
by a 70-cm-thick yellowish-green silty claystone. Gleysols
are commonly formed in areas with high water tables that
are waterlogged for most of the year; lignites are derived
from organically rich, waterlogged soils characterized by
chemically reducing conditions (Mack et al. 1993). Based
on the sediments in which the fossils are preserved, the
large number of entire-margined notophyllous to meso-
phyllous leaf taxa present, species composition, and the
high species heterogeneity over short distances (based on
lateral sampling), the flora likely represents a moist tropical
forest community growing in swampy or water-saturated
(at least for a large portion of the year) conditions.
Results
Systematic Paleobotany
Malvaceae Juss. 1789
Sterculioideae Burnett 1835
Cola Schott & Endl. 1832
Cola amharaensis sp. nov. A. D. Pan and B. F. Jacobs
Diagnosis Entire-margined, mesophyllous leaf with
cuneate base, apical pulvinus, and triplinerved basal
venation (Fig. 2). Primary venation is pinnate, secondary
venation is brochidodromous, and tertiary venation is
alternate percurrent with admedially ramified veins
(Fig. 2). Abaxial leaf epidermal cells are rectangular and
isodiametric (Figs. 3, 4). Stomatal complexes are brac-
hyparacytic and restricted to the abaxial side of the leaf
(Fig. 3). Anticlinal cell walls are rounded to undulate and
have bead-like thickenings. Periclinal cell walls have a
stippled texture (Figs. 3, 4). Abaxial glandular hairs possess
globose or capitate heads and are uniseriate or biseriate and
multicellular (Fig. 4). Buttressed hair bases are present on
the adaxial surface.
Description Mesophyllous leaf compression fragment
(and counterpart) with length of 78 mm and maximum
width of 58.5 mm. The entire-margined lamina is either
elliptic or obovate and symmetrical. The base is cuneate
and has an acute angle. The petiole has an apical pulvinus
(4 mm long, 2.5 mm wide) which is attached at the top of a
13-mm-long petiole fragment (Fig. 2). The primary vena-
tion is pinnate, although the leaf is triplinerved at the base
with secondary veins (oriented at more acute angles than
subsequent secondaries) following parallel to the margin,
Fig. 2–7 Cola amharaensis Pan and Jacobs sp. nov. and Cola flavo-velutina. Fig. 2. Cola amharaensis Holotype CH41-P36/CH41-P41;
scale bar 15 mm. Fig. 3. Brachyparacytic stomata of C. amharaen-sis; scale bar 30 lm. Fig. 4. Multicellular glandular hairs of
C. amharaensis; scale bar 30 lm. Fig. 5. Leaf of Cola flavo-velutina;
scale bar 15 mm. Fig. 6. Brachyparacytic stomata of C. flavo-velutina; scale bar 30 lm. Fig. 7. Multicellular glandular hairs of
C. flavo-velutina; scale bar 30 lm
The earliest record of the genus Cola from the Late Oligocene 253
123

thinning toward a connection to the next pair of second-
aries (Fig. 2). Secondary venation is brochidodromous and
the spacing between pairs of secondary veins decreases
somewhat towards the base. Weak intersecondaries are
present and merge with the tertiary veins. Tertiary venation
is alternate percurrent with admedially ramified vein
course. Quaternary veins form a regular polygonal reticu-
late pattern. Cuticle: Abaxial cells are isodiametric to
rectangular in shape and are 16–53 lm long and 10–36 lm
wide (Fig. 3). Abaxial anticlinal cell walls are generally
straight to rounded and possess bead-like thickenings.
Abaxial epidermal periclinal cell surfaces are stippled. The
stomatal complex is brachyparacytic and stomata are
restricted to the abaxial leaf surface (Fig. 3). Glandular
trichomes are present and consist of a single foot cell and
uniseriate or biseriate multicellular, generally 3–4 layered,
globose or capitate head (Fig. 4). The glandular trichomes
range in size from 29 to 35.5 lm long and from 32 to
42 lm wide (Fig. 4), typically wider than long. Adaxial
cells are isodiametric, 24–42 lm long and 15–31 lm wide.
The adaxial epidermal cell surface ornamentation is similar
to that of the abaxial side. Anticlinal cell walls are rounded
to undulate. Trichome bases, which may have buttresses,
have been observed on the adaxial surface and, based on
morphology, likely represent glandular hairs (Fig. 17).
Holotype CH41-P36/CH41-P41 (Fig. 2); found at sublo-
cality CH41 in the Guang River flora. This specimen is
housed at the National Museum of Ethiopia in Addis Ababa.
Etymology The species is named for the Amhara Prov-
ince of Ethiopia where the fossil and the Guang River flora
were found.
Remarks The fossil can be placed in the genus Cola
based on the following characteristics: simple leaves with
entire margins, cuneate base with an acute angle and trip-
linerved veins, the presence of a pulvinus at the apex of the
petiole (Figs. 2, 5), weak brochidodromous secondary
venation, and the presence of multicellular glandular hairs
on the epidermal surfaces of the leaf (Figs. 4, 7). The fossil
differs from Octolobus in possessing a cuneate leaf base
and trichomes that are solely glandular. Also, Octolobus
spectabilis has stomatal complex chains and clusters
(Fig. 23), defined here as four or more adjacent stomatal
complexes lacking separating epidermal (nonsubsidiary)
cells, a characteristic that is absent in the fossil and almost
all extant species of Cola observed, excluding C. letouze-
yana. The fossil differs from Pterygota, which have sub-
cordate/cordate, rounded, or almost truncate bases.
Based on leaf and epidermal characters of the species,
the fossil most closely resembles Cola attiensis and
C. flavo-velutina (Fig. 5), two extant West and Central
African species. The fossil is similar to both in (1)
possessing a simple-entire margined leaf with a cuneate
base, (2) triplinerved basal veins which fade towards the
margin, (3) straight to rounded abaxial anticlinal cell walls
(Fig. 2), (4) brachyparacytic stomata (or rarely other para-
cytic stomatal complex types) restricted to the abaxial side
(Fig. 3), (5) multicellular glandular hairs present and
dominant (C70% of hairs encountered are glandular) on the
abaxial surface (Fig. 4), and (6) adaxial anticlinal cell walls
that are rounded to undulate. In addition, hair bases similar
to those of the multicellular glandular hairs of the abaxial
side are present on the adaxial surface. These bases may
indicate that glandular hairs are present and possibly pre-
dominant on the adaxial cell surface of the fossil, a
characteristic that also occurs within Cola attiensis and
C. flavo-velutina. Furthermore, the length and width ranges
of abaxial and adaxial epidermal cells in the Ethiopian Cola
are quite similar those in C. attiensis and C. flavo-velutina.
Cola attiensis is a small tree or shrub found along river
banks or moist ground in evergreen forest growing on
sandy clays in Cote d’Ivoire and has been reported from
Cameroon and Gabon (Halle 1961; Lebrun and Stork 2003;
Holmgren et al. 2004). Cola flavo-velutina is a shrub or
tree growing in the understorey of very moist rainforest in
Ghana, Nigeria, Cameroon, and Gabon (Halle 1961;
Lebrun and Stork 2003).
While the fossil differs little from Cola attiensis and
C. flavo-velutina based on the characters mentioned above,
the fossil is placed in a new species Cola amharaensis
because it differs by having buttresses on some adaxial hair
bases (Fig. 17; buttresses were absent from all hair bases
observed in C. attiensis and C. flavo-velutina), an absence
of stellate/peltate hairs or hair bases on the abaxial surface
(stellate hairs are present in Cola flavo-velutina), and a lack
of straight anticlinal cell walls on the adaxial surface (which
can occur within C. attiensis). The closest resemblance to
Cola amharaensis sp. nov. among the eastern African
species observed in this study is C. usambarensis, which
differs from the fossil in having a convex leaf base and
possessing only stellate/peltate hairs on its leaf surfaces.
Other eastern and southern Cola species not included in
this study are not similar to the fossil (Keay and Brenan 1973;
Brenan 1978; Verdoorn 1981; Coates Palgrave 1977; White
et al. 2001; Cheek 2002). Primary differences include
leaf base morphology (Cola chlorantha, C. congolana,
C. gigantea, C. lukei, and C. octoloboides), and the absence
of triplinerved or trinerved venation at the base of the leaf
(C. congolana).
Comparative leaf morphology in extant Cola,
Octolobus, and Pterygota
A variety of leaf types are present within the Cola ?
Octolobus ? Pterygota clade sensu Wilkie et al. (2006).
254 A. D. Pan, B. F. Jacobs
123

These include both simple and palmately compound leaved
species, which are entire-margined, and may be unlobed or
lobed. Simple, unlobed, entire-margined leaves are often
trinerved (or triplinerved). Cola, a relatively large genus,
has a diverse array of leaf morphologies, while Octolobus
consists solely of entire-margined, unlobed, simple-leaved
species. Octolobus spectabilis is usually not trinerved at the
base, but trinerved basal veins do occur consistently within
O. heteromerus and O. zenkeri. Pterygota species can have
either lobed or unlobed leaves, are usually palmately
veined, and often have a cordate or subcordate base,
although nearly truncate bases may also occur.
Micromorphology
Epidermal cells
Most epidermal cells within all three genera are isodia-
metric or rectangular in shape and 3–8 sided (Fig. 8).
Abaxial epidermal cells range in size from 8 to [100 lm
long and 6 to 48.5 lm wide. Adaxial cells range from 65 to
85 lm in length and 3 to 51.5 lm in width. Species typi-
cally have anticlinal cell walls that are either straight to
rounded, or rounded to undulate in shape. If undulate, the
sinuous walls are widely to narrowly U-shaped (Fig. 8).
Fine to prominent stippled ornamentation is present on the
abaxial and adaxial periclinal cell walls of almost all spe-
cies of Cola and Octolobus that we observed (Fig. 8).
Stippled ornamentation also occurs in Pterygota macro-
carpa. Striations are present on the abaxial and adaxial
(though slightly less defined) periclinal cell walls of
Pterygota bequaertii (Fig. 9) and abaxially in Cola lepi-
dota. Anticlinal cell wall thickenings are present on all
species observed, the most common types being ‘‘knob-
like’’ (Fig. 2) and ‘‘T-shaped,’’ which are defined and
illustrated in Dilcher (1974).
Trichomes
A variety of different trichome (hair) types occur within the
Malvaceae s.l. Although the family is often known for stellate,
peltate, and tufted trichomes (Figs. 10, 11; Metcalfe and
Chalk 1950; Bayer and Kubitzki 2003; Judd et al. 2008), other
types are also common. These include clavate or capitate
(globose) multicellular glandular (Figs. 12, 13), unicellular
glandular, and simple hairs (Fig. 14). Trichome types found
within Cola include glandular, simple unicellular, and stellate/
peltate hairs (Figs. 10, 12, 13, 14; in most cases a polygonal
base is usually the only indication of the presence of stellate/
peltate hairs; Fig. 11). Glandular hairs and stellate/peltate hair
bases were observed in all Octolobus and Pterygota species
examined; however, simple trichomes were absent among
observed species within these two genera.
Stellate/peltate hairs
The majority of species have deciduous stellate/peltate
hairs with only polygonal (usually pentagonal or hexago-
nal)-shaped hair bases with thickened margins and acute
points present (Fig. 11). However, whole stellate/peltate
trichomes are abundant on the abaxial surfaces of Cola
cordifolia and C. lepidota. Intact hairs also occur more
sporadically in Cola heterophylla (abaxial), C. lateritia
(adaxial), C. mahoudensis (abaxial; Fig. 10), C. millenii
(abaxial), and Pterygota bequaertii (adaxial).
Glandular hairs
Glandular hairs were observed in almost every species of
Cola, Octolobus, and Pterygota examined with the
exception of Cola minor, C. natalensis, and C. usambar-
ensis (Table 1; Appendix). Hair bases are often thickened
and can exhibit buttresses, as in Cola ballayi, C. discog-
lypremnophylla (Fig. 16), and C. simiarum. Glandular
hairs emerge from a unicellular rectangular base cell
(Fig. 12). Most glandular hairs occur singly, but twinned
glandular hairs can be found in Cola acuminate (Fig. 13),
C. discoglypremnophylla, C. letouzeyana, and Octolobus
zenkeri.
Simple hairs
Simple hairs are quite rare within Cola and were only
observed in C. mahoundensis (Fig. 14), C. millenii, and
C. scheffleri. While it should be noted that Inamdar et al.
(1983) reported occasional simple hairs in Cola acuminata,
none were observed in this study.
Silica bodies
Silica bodies were observed in several species of Cola,
particularly around the veins (Fig. 15). When silica bodies
were observed, the majority were found on the abaxial
surface of the leaf. Cola acuminata is unusual in having
silica bodies both abaxially and adaxially, but they are
noticeably more numerous on the abaxial surface.
Silica bodies resemble jagged spheres and are usually on
the order of 3–4 lm in diameter (Figs. 9, 15).
Stomata
Nearly all stomata found within Cola, Octolobus, and
Pterygota are restricted to the abaxial leaf surface, however
a few isolated stomata occur on the adaxial surface in Cola
chlamydantha, C. discoglypremnophylla, C. heterophylla,
C. hispida, and C. porphyrantha. The majority of species
have brachyparacytic and paracytic stomatal complexes
The earliest record of the genus Cola from the Late Oligocene 255
123

(Figs. 3, 16, 18; however, tetracytic (Fig. 20) and aniso-
cytic types are not uncommon and can be prevalent in some
species (Fig. 21; Table 1). Rarer stomatal complex types
including hemiparacytic, cyclocytic, hexacytic, and
anomocytic types can be found among a few species
(Figs. 19, 21; Table 1).
Discussion
Fossil record
The fossil record, besides the Guang River flora Cola sp.
leaf compression, includes a flower and possibly a leaf cast
Fig. 8–15 Leaf epidermal
micromorphology; scale bars30 lm. Fig. 8. Stippled
ornamentation on the adaxial
epidermal cells of Cola flavo-velutina. Fig. 9. Striated
epidermal abaxial surface and
silica bodies of Pterygotabequaertii. Fig. 10. Stellate hair
on the abaxial surface of Colamahoundensis. Fig. 11. Stellate
or peltate remnant hair base on
the abaxial of C. minor.
Fig. 12. Multicellular glandular
hair on the abaxial surface of
C. mahoundensis. Fig. 13.Twinned or coupled glandular
hairs on the abaxial surface of
C. acuminata. Fig. 14. Abaxial
unicellular simple trichome of
C. mahoundensis. Fig. 15.Adaxial stone cells of
C. buntingii
256 A. D. Pan, B. F. Jacobs
123

from the Early Miocene of Uganda, fossil wood of the form
genus Colaxylon from the Late Neogene of Ethiopia and
Chad, a fossil denoted as cf. Cola from the Early Miocene
of Kenya, and several species of Cola and Octolobus
reported from the Late Neogene of Cameroon (Fig. 1,
Menzel 1920; Hamilton 1968; Koeniguer 1973; Lemoigne
1978; Jacobs and Winkler 1992). A fossil seed attributed to
cf. Cola from the Cretaceous of Senegal (Monteillet and
Lappartient 1981) does not appear to be similar to the
extant genus.
Fossil wood of the form genus Colaxylon is reported by
Koeniguer (1973) and Lemoigne (1978) from the Late
Neogene of Chad and Ethiopia, respectively. While it is
possible that these fossil woods may have affinity to Cola,
Fig. 16–23 Leaf epidermal
micromorphology cont. Abaxial
surfaces; scale bars 30 lm.
Fig. 16. Buttressed hair base on
the abaxial surface of Coladiscoglypremnophylla.
Fig. 17. Buttressed hair base on
the adaxial surface of Colaamharaensis Pan sp. nov.
Holotype CH41-P36/CH41-P41.
Fig. 18. Brachyparacytic
stomata (a paracytic stomatal
complex type) of C. uloloma.
Fig. 19. Anomocytic stoma
(a polycytic stomatal complex
type) of C. ballayi.Fig. 20. Tetracytic stomata of
C. ballayi. Fig. 21. Hexacytic
stoma of C. nitida. Fig. 22.Thickened periclinal wall
surrounding the stomata of
C. lissachensis. Fig. 23.Stomatal complex cluster of
Octolobus spectabilis
The earliest record of the genus Cola from the Late Oligocene 257
123

many genera within the Sterculioideae have similar wood
anatomical structures (Metcalfe and Chalk 1950).
The fossil flower from the Bukwa locality in western
Uganda is Early Miocene (19.5–19.1 Ma based on40Ar/39Ar radiometric dating of lavas; Maclatchy et al.
2006). Hamilton (1968) considered the floral cast to have
affinities with either Cola or Pterygota based on the near-
sessile attachment of two whorls of bilobed anthers to the
staminal tube (androgynophore), and the presence of five
partially fused sepals. In the illustration provided by
Hamilton (1968: Fig. 1a), a structure at the apex of the
androgynophore appears to be 4–5 vestigial fused carpels
as occurs on male flowers in species of Cola and Pterygota.
Hamilton (1968), who compared the fossil to living
Ugandan Cola and Pterygota species, considered it to be
most similar to Cola gigantea A. Chev., which is found
only in Uganda and is often confused with Cola cordifolia,
a species restricted to western Africa (Lebrun and Stork
2003). However, in our opinion the fossil’s loose inter-
digitating whorls of stamens compares more closely to the
genus Pterygota, rather than Cola (Halle 1961 Bodard
1962; Germain 1963).
A lobed fossil leaf from the same bed as the flower was
noted by Hamilton (1968) as similar to either Euphorbia-
ceae or Sterculiaceae (e.g., Cola). Hamilton (1968) notes
in his description that each of the numerous main veins
(6–10?) leads to a leaf-lobe. However, Cola and Pterygota
species (that possess lobed, palmately veined, simple
leaves) generally have 3–5 lobes, and Cola gigantea leaves
are either unlobed or consistently possess three-lobes.
Thus, the affinity of this fossil leaf remains uncertain.
Cf. Cola fossil leaf impressions from the Early Miocene
Ngorora Formation (Fig. 24) have weak brochidodromous
secondary venation, alternate percurrent tertiary venation,
and possibly swollen pulvini, all features common to extant
Cola species. Moreover, the shallowly subcordate and
slightly asymmetrical base in association with both pinnate
basal secondary venation and obovate leaf shape is not
common within Cola. Only one species, Cola letouzeyana
from Cameroon, is similar in all these respects to the fossil
(Nkongmeneck 1985) and also compares favorably in
having a decrease in secondary vein angle and spacing
towards the leaf base. The fossil also looks superficially
similar to some species of Scaphopetalum Mast. (Malva-
ceae sensu lato: Byttnerioideae), for example S. thonneri
De Wild. & Th. Durand; however, it lacks the distinctive
trinerved venation arising from the base.
More than 700 specimens of leaf impressions found in
tuffs near Mount Cameroon were reported by Menzel
(1920), and among these he reports close affinities with 15
extant Cola and two Octolobus species. Menzel (1920)
considered all of these impressions to represent taxa that
still exist in the area today. Many of these fossils have
venation similar to that of Cola and Octolobus; however,
definitive identification cannot be established at this time.
While the age of these fossils is not known (considered
Late Neogene in age according to Menzel 1920), these
fossils are preserved in tuff sediments so radiometric dating
might be possible. It is interesting to note that today this
area of West Africa (the Lower Guinea center) has the
highest diversity and species richness of Cola in Africa
(Cheek et al. 1996; Cheek 2002) and is considered by
Hamilton (1976) to have been a major rainforest refugium
during the Pleistocene (Cheek et al. 1996; Linder 2001).
Leaf characters and relationships within Cola
Cola has often been split into a number of subgroups based
on leaf and reproductive characteristics by several authors,
most of which differ (at least somewhat) from one another
(see Bodard 1962 for a summary of taxonomic modifica-
tions within the genus). In this paper we have provided
leaf and epidermal characters for several species of Cola,
Octolobus, and Pterygota (summarized in Table 1 and
Appendix).
A number of purported subgenera were erected
by Halle (1961) to categorize Gabon Cola species. One of
these, Cola subgenus Neocourtenia (which includes
C. gabonensis, C. heterophylla, C. hispida, C. mahound-
ensis, C. marsupium, and C. urceolata), is characterized by
palmately veined, simple-leaved (often lobed) species with
inconspicuous tertiary and higher-order veins (only visible
in dried leaves), caducous stipules, flowers that are gen-
erally less than 2 cm in diameter possessing a uniseriate
whorl of stamens on the androecium, stamens numbering
one or two per carpel, panicle-like cyme inflorescences,
and seeds with a thick, very fibrous integument (Halle
Fig. 24 Leaf impressions of cf. Cola from the Early Miocene
Ngorora Formation of Kenya; scale bar 30 mm
258 A. D. Pan, B. F. Jacobs
123

1961). Within our study, we observed that charactaceous
leaf texture also supports close relationship amongst these
taxa (of the species listed above, C. urceolata was not
examined). Other species (Cola carcifolia, C. millenii, and
C. scheffleri) which share some morphological character-
istics, including palmately veined simple leaves with lobes
and chartaceous texture, with Halle’s (1961) subgenus
Neocourtenia may belong to (or closely related to) the
subgenus (Keay and Brenan 1973). Bodard (1962) con-
sidered Cola carcifolia and C. millenii to be closely related
and placed them in subgenus Haplocola along with
C. triloba (R. Br.) K. Schum., C. laurifolia, C. humilis
(which is considered a synonym of C. heterophylla;
Lebrun and Stork 2003), and C. reticulata (A. Chev.).
Cola chlamydantha, C. lepidota, C. lissachensis, and
C. pachycarpa appear to be closely related based on the
palmately compound leaves, the possession of anisocytic
stomatal complex types, thickened rims (?papillae) on the
periclinal wall of the subsidiary cells adjacent to the guard
cells (often encircling the stoma), and subsidiary cells with
striate ornamentation (Fig. 22). However, Cola chlamy-
dantha and C. lastoursvillensis (M. Bodard & Pellegr.)
N. Halle (not sampled) have often been placed in a separate
genus Chlamydocola K. Schum. based on a suite of char-
acters which is unique within the genus Cola including
androphore with anthers arranged in a subsinuous or digi-
tate wreath formation, bisexual flowers with 9–12 carpels
having numerous (22–26) ovules, albuminous seeds, and
large orbicular bracteoles surrounding the flower bud
(Halle 1961; Bodard 1962; Germain 1963; Keay and
Brenan 1973). Due to these unique features, Chlamydocola
may turn out to be a good genus. Cola ficifolia, a palmately
lobed simple-leaved species, also has stomata with thick-
ened to papillate rims on the periclinal wall of the sub-
sidiary cells and anisocytic stomata. Whether this species is
closely related to those mentioned above (and those species
which may or may not themselves be closely related) is
unknown, but is worth investigating.
Paleobiogeography of Cola
The presence of Cola amharaensis in Ethiopia is interest-
ing not only due to the absence of the genus in Ethiopia
today, but also due to the fossil’s close morphological
similarity with extant species found in the Guineo-Con-
golian forest region (West and Central Africa) as opposed
to living species that are found in closer geographic
proximity in Kenya, Somalia, and Sudan (Lebrun and Stork
2003; Fig. 1). At least two other fossil genera from the
Guang River flora share this pattern of absence from
Ethiopia today, and greater similarity to Guineo-Congolian
species than to extant East African taxa. These include a
fossil leaflet of the climbing calamoid palm Eremospatha
(G. Mann & H. Wendl.) H. Wendl. (Pan et al. 2006), a
genus restricted to West and Central Africa today, and
several fossil leaflet specimens of Cynometra (Legumino-
sae) that are most similar to two Central African species
(Pan 2007). In addition, fossil taxa with closer affinity to
extant Guineo-Congolian forest elements (than to East
African tropical forests) are also present in a Late Oligo-
cene palynoflora from northeastern Kenya (Mansonia
altissima type, Petersianthus type, and Triplochiton type;
Vincens et al. 2006). These ‘‘exotic’’ taxa, as they are
referred to in Vincens et al. (2006), and those from Ethi-
opia document a greater geographic distribution for some
tropical moist forest elements in the Paleogene than their
extant relatives, and indicate that extant species within
these genera in eastern Africa may be more distantly
related to these fossils than would otherwise be expected.
Acknowledgment We would like to thank the Authority for
Research and Conservation of Cultural Heritage, the Ministry of
Culture and Tourism, Ethiopia, and especially Ato Jara for permission
to conduct our continuing research in northwestern Ethiopia, and the
Director Mamitu Yilga and staff of the National Museum, Addis
Ababa, and the Gondar ARCCH and Chilga Ministry of Culture and
Sports Affairs for logistical support. We thank the Missouri Botanical
Garden, the Royal Botanic Gardens at Kew, and the collectors of the
herbaria specimens examined for assistance and access to their col-
lections. We are grateful to the National Museums of Kenya, the
Baringo Paleontological Research Project, the East African Herbar-
ium, and Christine Kabuye for their collaborative support of work in
Kenya. This project was funded by grants from the National Science
Foundation (EAR-0001259, EAR-0240251, and EAR-0617306), the
National Geographic Society, and the Dallas Paleontological Society.
Tillehun Selassie, Misege Birara, Habtewold Habtemichael, Mesfin
Mekonnen, and Drs. Ambachew Kebede and Aklilou Asfaw provided
valuable field assistance. We thank Dr. Martin Cheek for generously
sharing information about Cola and other sterculioids, for providing
advice, and for supplying cuticle specimens from the Royal Botanic
Gardens at Kew. We are grateful to Dr. Thomas Denk for images of
the Cameroon sterculioid fossils housed at the Swedish Museum of
Natural History. We also appreciatively acknowledge help from
Yohannes Desta, Yeshiwass Sitotaw, Gebremeskel Ayele, Elias
Addissu, and Teshome Yohannes at Chilga and laboratory assistance
from Kathryn Larson.
Appendix
Leaf and epidermal characteristics and states
1. Leaf type: (0) simple, (1) palmately compound,
(2) pinnately compound
2. Leaf(let) shape: (0) elliptic, (1) obovate, (2) ovate,
(3) oblong, (4) special (needles, awls, etc.)
3. Leaf(let) attachment: (0) petiolate [petiolulate for
compound-leaved species], (1) subsessile/sessile
4. Leaf(let) texture: (0) chartaceous (1) semicoriaceous,
(2) coriaceous
5. Leaf(let) margin type: (0) entire, (1) lobed, (2)
toothed
The earliest record of the genus Cola from the Late Oligocene 259
123

6. Leaf(let) base angle: (0) acute, (1) obtuse, (2) wide
obtuse
7. Leaf(let) base shape: (0) cuneate/concave/decurrent,
(1) convex/rounded, (2) truncate, (3) subcordate/
cordate/lobate
8. Leaf(let) apex morphology: (0) acute, (1) subacumi-
nate, (2) acuminate, (3) obtuse/rounded/slightly
emarginate, (4) deltate/cuspidate, (5) emarginate
9. Primary venation: (0) pinnate, (1) tri-nerved at the
base, (2) palmately veined
10. Number of secondary vein pairs: (0) 0–5; (1) 6–10;
(2) 11–15; (3) [16
11. Secondary vein categories: (0) brochidodromous,
(1) eucamptodromous, (2) festooned brochidodrom-
ous, (3) cladodromous, (4) reticulodromous, (5)
toothed-craspedodromous, (6) toothed-semicraspedo-
dromous, (7) toothed-festooned semicraspedodromous
12. Tertiary venation: (0) random reticulate, (1) opposite
percurrent, (2) alternate percurrent, (3) mixed oppo-
site/alternate percurrent
13. Abaxial epidermal cell shape: (0) isodiameteric,
(1) rectangular
14. Abaxial cell size: indicated by length (first set of
numbers) and width ranges, rounded to the nearest
0.5 lm
15. Abaxial epidermal cell arrangement: (0) random (a
combination of any of the following cellular arrange-
ments), (1) nonrandom–tetragonal, (2) nonrandom–
pentagonal, (3) nonrandom–hexagonal, (4) nonrandom–
polyagonal ([6-sided cells), (5) nonrandom–linear
16. Abaxial anticlinal cell wall pattern: (0) straight,
(1) rounded, (2) undulate
17. Abaxial anticlinal cell wall shape of undulation:
(0) U, (1) V, (2) X18. Abaxial epidermal cell surface ornamentation:
(0) absent, (1) stippled, (2) striate
19. Abaxial stoma: (0) absent, (1) present
20. Abaxial stomatal complex type: (0) polycytic
types (anomocyptic, cyclocytic), (1) anisocytic,
(2) paracytic types (paracytic, brachyparacytic,
amphibrachyparacytic, hemiparacytic), (3) tetracytic
types (staurocytic, anomotetracytic, paratetracytic,
brachyparatetracytic)
21. Abaxial stomatal complex chains or clusters:
(0) absent, (1) present
22. Abaxial stomatal ornamentation: (0) absent, (1) striate,
(2) papillate, (3) thickened areas on periclinal wall
23. Abaxial guard cells: (0) level, (1) sunken,
(2) raised
24. Number of abaxial trichomes types: (0) glabrous/absent
(no trichomes present), (1) one, (2) two, (3) three,
(4) four, (5) five or more
25. Abaxial hair types present: (0) unicellular simple,
(1) unicellular glandular, (2) multicellular glandular,
(3) stellate/peltate (usually only the polygonal base is
preserved),
26. Abaxial trichome dominance: (0) glandular trichomes
predominate [C70% glandular], (1) no trichome type
predominates (29–69%), (2) stellate/peltate trichomes
predominate [C70% stellate/peltate], (3) simple hairs
predominate (C70% simple)
27. Abaxial glandular hair pairs: (0) absent, (1) present
28. Adaxial epidermal cell shape: (0) isodiameteric,
(1) rectangular
29. Adaxial cell size: indicated by length (first set of
numbers) and width ranges, rounded to the nearest
0.5 lm
30. Adaxial epidermal cell arrangement: (0) random (a
combination of any of the following cellular
arrangements), (1) nonrandom–tetragonal, (2) non-
random–pentagonal, (3) nonrandom–hexagonal, (4)
nonrandom–polyagonal ([6-sided cells), (5) non-
random–linear
31. Adaxial anticlinal cell wall pattern: (0) straight,
(1) rounded, (2) undulate
32. Adaxial anticlinal cell wall shape of undulation:
(0) U, (1) V, (2) X33. Adaxial epidermal cell surface ornamentation:
(0) absent, (1) stippled, (2) striate
34. Adaxial stoma: (0) absent, (1) present
35. Adaxial stomatal complex type: (0) polycytic types
(anomocyptic, cyclocytic), (1) anisocytic, (2)
paracytic types (paracytic, brachyparacytic, amphi-
brachyparacytic, hemiparacytic), (3) tetracytic types
(staurocytic, anomotetracytic, paratetracytic, brachy-
paratetracytic)
36. Abaxial stomatal complex chains or clusters:
(0) absent, (1) present
37. Adaxial stomatal ornamentation: (0) absent, (1) striate,
(2) papillate, (3) thickened areas on periclinal wall
38. Adaxial guard cells: (0) level, (1) sunken, (2) raised
39. Number of adaxial trichomes types: (0) glabrous/
absent (no trichomes present), (1) one, (2) two, (3)
three, (4) four, (5) five or more
40. Adaxial hair types present: (0) unicellar simple,
(1) unicellular glandular, (2) multicellular glandular,
(3) stellate/peltate (usually only the polygonal base is
preserved)
41. Adaxial trichome dominance: (0) glandular trichomes
predominate (C70% glandular), (1) no trichome type
predominates (29–69%), (2) stellate/peltate trichomes
predominate (C70% stellate/peltate), (3) simple hairs
predominate (C70% simple)
42. Adaxial glandular hair pairs: (0) absent, (1) present
260 A. D. Pan, B. F. Jacobs
123

References
Alverson WS, Whitlock BA, Nyffeler R, Bayer C, Baum DA (1999)
Phylogeny of the core Malvales: evidence from ndhF sequence
data. Am J Bot 86:1474–1486
Bayer C, Kubitzki K (2003) Malvaceae. In: Kubitzki K, Bayer C (eds)
The families and genera of vascular plants. Springer, Berlin,
pp 225–310
Bodard M (1962) Contributions a l’etude systematique du genre Colaen Afrique Occidentale. Annales de Faculte des Sciences de
l’Universite de Dakar 7:1–187
Brenan JPM (1978) A new species of Cola (Sterculiaceae) from
East Africa. Kew Bull 33:283–286
Cande SC, Kent DV (1995) Revised calibration of the geomagnetic
polarity time scale for the Late Cretaceous and Cenozoic.
Journal of Geophysical Research 100:6093–6095
Cheek M (2002) A new species of Cola (Sterculiaceae) from the
Usambara Mountains, Tanzania. Kew Bull 57:417–422
Cheek M, Frimodt-Møller C (1998) The genus Octolobus (Sterculi-
aceae) new to East Africa. Kew Bull 53:682
Cheek M, Cable S, Hepper FN, Ndam N, Watts J (1996) Mapping
plant diversity on Mount Cameroon. In: van der Maesen LJG,
van der Burgt XM, de Rooy JM (eds) The Biodiversity of
African Plants: Proceedings XIV AETFAT Congress 22–27
August 1994. Kluwer Academic Publishers, Wageningen,
pp 110–120
Coates Palgrave K (1977) Trees of Southern Africa. C. Struik
Publishers, Cape Town
Dilcher DL (1974) Approaches to the identification of angiosperm
leaf remains. The Bot Rev 40:1–157
Ellis B, Daly DC, Hickey LJ, Johnson KR, Mitchell JD, Wilf P, Wing
SL (2009) Manual of leaf architecture. Cornell University Press,
Ithaca
Friis I (1992) Forests and forest trees of northeast tropical Africa.
Royal Botanic Gardens, Kew
Garcıa Massini JL, Jacobs BF, Pan AD, Tabor N, Kappelman J (2006)
The occurrence of the fern Acrostichum in Oligocene volcanic
strata of the northwestern Ethiopian plateau. International
Journal of Plant Sciences 167:909–918
Gehrig M (1938) Beitrage zur Pharmakognosie der Malvales.
Anatomie des Laubblattes. Thesis, Base
Gentry AH (1988) Changes in plant community diversity and floristic
composition on environmental and geographic gradients. Ann
Mo Bot Gard 75:1–34
Germain R (1963) Flora Du Congo & du Rwanda et du Burundi.
Spermatophytes 10(88): Sterculiaceae. Jardin botanique national
de Belgique, Brussels
Halle N (1961) Flore du Gabon, volume 2: Sterculiacees. Museum
National d’Histoire Naturelle, Paris
Hamilton A (1968) Some plant fossils from Bukwa. Ug J 32:157–164
Hamilton AC (1976) The significance of pattern of distribution.
Palaeoecol Afr Surround Isl 9:63–97
Holmgren M, Poorter L, Siepel A, Bongers F, Buitelaar M, Chatelain
C, Gautier L, Hawthorne WD, Helmink ATF, Jongkind CCH,
Os-Breijer HJ, Wieringa JJ, van Zoest AR (2004) Ecological
profiles of rare and endemic species. In: Poorter L, Bongers F,
Kouame FN, Hawthorne WD (eds) Biodiversity of West African
Forests: an ecological atlas of woody plant species. CABI
Publishing, Oxford, pp 101–389
Inamdar JA, Balakrishna Baht R, Ramana Rao TV (1983) Structure,
ontogeny, classification, and taxonomic significance of tric-
homes in Malvales. K J Bot 26:151–160
Jacobs BF, Winkler DA (1992) Taphonomy of a middle Miocene
autochthonous forest assemblage, Ngorora Formation, central
Kenya. Palaeogeogr Palaeoclimatol Palaeoecol 99:31–40
Jacobs BF, Tabor N, Feseha M, Pan A, Kappelman J, Rasmussen T,
Sanders W, Wiemann M, Crabaugh J, Garcia Massini JL (2005)
Oligocene-age (32.7–27.5 Ma) terrestrial strata of northwestern
Ethiopia: lithology, paleontology, and paleoclimate. Palaeonto-
logia Electronica 8:1–19. http://palaeoelectronica.org/2005_1/
jacobs25/issue1_05.htm
Judd WS, Manchester SR (1997) Circumscription of Malvaceae
(Malvales) as determined by a preliminary cladistic analysis of
morphological, anatomical, palynological, and chemical charac-
ters. Brittonia 49:384–405
Judd WS, Campbell CS, Kellogg EA, Stevens PF, Donoghue MJ
(2008) Plant systematics: a phylogenetic approach, 3rd edn.
Sinauer Associates Inc, Sunderland
Kappelman J, Rasmussen DT, Sanders WJ, Feseha M, Bown T,
Copeland P, Crabaugh J, Fleagle J, Glantz M, Gordon A, Jacobs
B, Maga M, Muldoon K, Pan A, Pyne L, Richmond B, Ryan T,
Seiffert ER, Sen S, Todd L, Wiemann MC, Winkler A (2003)
Oligocene mammals from Ethiopia and faunal exchange
between Afro-Arabia and Eurasia. Nature 426:549–552
Keay RWJ, Brenan JPM (1973) Cola. In: Keay RWJ (ed) Flora of
West Tropical Africa, volume 1, part 2, 2nd edn. The Whitefriare
Press Ltd., London, pp 321–332
Koeniguer JC (1973) Les bois heteroxyles de l’oasis de Kirdimi
(Tchad). C.R. 96e Congr. Nat. Soc. Sav., Toulouse, 1971. Paris
Sci 5:191–214
Lebrun J-P, Stork AL (2003) Tropical African flowering plants: ecology
and distributions, volume 1 Annonaceae-Balanitaceae. Conserva-
toire et Jardin botanique de la ville de Geneve, Switzerland
Lemoigne Y (1978) Flores tertiaires de la haute vallee de l’Omo
(Ethiopia) Palaeontogr. Abt B Palaeophytol 165:89–157
Linder HP (2001) Plant diversity and endemism in sub-Saharan
tropical Africa. J Biogeogr 28:169–182
Mabberley DJ (1997) The plant book: a portable dictionary of vascular
plants, 2nd edn. Cambridge University Press, Cambridge
Mack GH, James WC, Monger HC (1993) Classification of paleosols.
Geol Soc Am Bull 105:129–136
MacLatchy L, Deino A, Kingston J (2006) An updated chronology for
the Early Miocene of NE Uganda. J Vertebr Paleontol 26(Sup-
plement to 3):93A, abstract
Menzel P (1920) Uber Pflanzenreste aus Basalttuffen des Kam-
erungebietes. Beitrage zur Geologischen Erforschung der De-
utschen Schutzgebiete 18:17–32
Metcalfe CR, Chalk L (1950) Anatomy of the dicotyledons, volume I.
Oxford University Press, London
Mohr PA (1971) The geology of Ethiopia. Addis Ababa University
Press, Addis Ababa
Monteillet J, Lappartient J-R (1981) Fruits et graines du Cretace
superieur des carriers de Paki (Senegal). Rev Palaeobot Palynol
34:331–344
Nkongmeneck B-A (1985) Un nouveau Cola (Sterculiaceae) du
Cameroun. Bull. Mus. natn. Hist. nat. Paris, 4e ser., 7, section B.
Adansonia 3:337–339
Pan AD (2007) The Late Oligocene (28–27 Ma) Guang River flora
from the northwestern plateau of Ethiopia. Dissertation, South-
ern Methodist University, Dallas
Pan AD, Jacobs BF, Dransfield J, Baker WJ (2006) The fossil history
of palms (Arecaceae) in Africa and new records from the Late
Oligocene (28–27 Mya) of north-western Ethiopia. Bot J Linn
Soc 151:69–81
Phillips O, Miller JS (2002) Global patterns of plant diversity: Alwyn
H. Gentry’s forest transect data set. Missouri Botanical Garden,
St. Louis
Sanders WJ, Kappelman J, Rasmussen DT (2004) New large-bodied
mammals from the late Oligocene site of Chilga, Ethiopia. Acta
Palaeontologica Polonica 49:365–392
The earliest record of the genus Cola from the Late Oligocene 261
123

Verdoorn IC (1981) The genus Cola in southern Africa. Bothalia
13:277–279
Vincens A, Tiercelin J-J, Buchet G (2006) New Oligocene-early
Miocene microflora from the southwestern Turkana Basin,
palaeoenvironmental implications in the northern Kenya Rift.
Palaeogeography Palaeoclimatology Palaeoecology 239:470–486
Vollesen K (1995) Sterculiaceae. In: Edwards S, Tadesse M, Hedberg
I (eds) Flora of Ethiopia and Eritrea, volume 2, part 2:
Canellaceae to Euphorbiaceae. The National Herbarium and
Uppsala University, Addis Ababa and Uppsala, pp 165–185
White F, Dowsett-Lemaire F, Chapman JD (2001) Evergreen forest
flora of Malawi. Royal Botanic Gardens, Kew
Wilkie P, Clark A, Pennington RT, Cheek M, Bayer C, Wilcock CC
(2006) Phylogenetic relationships within the subfamily Stercu-
lioideae (Malvaceae/Sterculiaceae-Sterculieae) using the chlo-
roplast gene ndhF. Syst Bot 31:160–170
Yemane K, Robert C, Bonnefille R (1987) Pollen and clay mineral
assemblages of a late Miocene lacustrine sequence from the
northwestern Ethiopian highlands. Palaeogeogr Palaeoclimatol
Palaeoecol 60:123–141
262 A. D. Pan, B. F. Jacobs
123





























