Embed Size (px)
Citation preview

www.elsevier.com/locate/hal
Harmful Algae 6 (2007) 29–47
The first description of the potentially toxic dinoflagellate,
Alexandrium minutum in Hunts Bay,
Kingston Harbour, Jamaica
Emma R. Ranston a,*, Dale F. Webber a, Jacob Larsen b
a Department of Life Sciences, University of The West Indies, Mona Campus, Mona, Kingston 7, Jamaica, West Indiesb IOC-DANIDA, Science and Communication Centre on Harmful Algae,
University of Copenhagen, Øster Farimagsgade 2D, DK-1353 Copenhagen K., Denmark
Received 2 February 2006; received in revised form 20 April 2006; accepted 30 May 2006
Abstract
The occurrence and morphology of the potentially toxic dinoflagellate species Alexandrium minutum found for the first time in
Jamaica, were examined and described by light and scanning electron microscopy. Classical morphological examinations of whole
cells, the thecal plate pattern of intact cells and more importantly the structure of individual thecal plates of squashed cells, were
conducted in an attempt to positively identify the species. Characteristics such as a tear-drop shaped apical pore plate with a comma-
shaped apical pore and no anterior attachment pore; a narrow sixth precingular plate; a narrow anterior sulcal plate longer than or
approximately as long as it is wide; and a posterior sulcal plate wider than long, confirmed the Jamaican species as A. minutum. This
dinoflagellate which produces potent neurotoxins responsible for paralytic shellfish poisoning (PSP) in humans in many parts of the
World, as well as mass mortality of various marine flora and fauna, was identified in water samples collected during an extensive
bloom of the species in the brackish to saline water body of Hunts Bay, an estuarine arm of Kingston Harbour, Jamaica in August
1994. The highest cell concentration was 4.6 � 105 cells l�1, a concentration which far exceeds acceptable concentrations
(<103 cells l�1) of PSP-toxin producing A. minutum in several countries including: Spain and Denmark. No PSP human symptoms
were reported during the bloom; however it was accompanied by a large kill of small pelagic fish extending across a third of the bay.
Since then, smaller blooms of A. minutum have occurred with the most recent in February and April 2004. Hunts Bay is an important
fishing, shrimping and to some extent oyster/mussel collection area and provides an important source of livelihood and food for
many fishermen in nearby fishing communities as well as an important source of food for members of other communities. Although
there are no known records of human illness due to PSP in Jamaica, the occurrence and blooming in Jamaican waters of this
potentially toxic dinoflagellate, is great cause for concern.
# 2006 Elsevier B.V. All rights reserved.
Keywords: Alexandrium minutum; First description; Hunts Bay; Kingston Harbour; Jamaica
* Corresponding author at: P.O. Box 34, Red Hills P.O., St. Andrew,
Jamaica, West Indies. Tel.: +876 944 4624; fax: +876 944 4324.
E-mail address: [email protected] (E.R. Ranston).
1568-9883/$ – see front matter # 2006 Elsevier B.V. All rights reserved.
doi:10.1016/j.hal.2006.05.006
1. Introduction
Alexandrium minutum was first described in Alex-
andria, Egypt (Halim, 1960). It has also been found in
Europe (Montresor et al., 1990), Asia (Chang et al.,
1997), Australia and North America (Hallegraeff et al.,
1988, 1991) where it can form blooms in estuarine
E.R. Ranston et al. / Harmful Algae 6 (2007) 29–4730
waters (Hallegraeff et al., 1988), eutrophic brackish
lagoons (Giacobbe et al., 1996) and aquaculture ponds
(Yoshida et al., 2000).
A. minutum is one of the at least nine toxic species of
the Alexandrium genus, known to be responsible for
PSP in many parts of the world including South
Australia (Hallegraeff et al., 1988) and France (Erard-
Le Denn, 1991; Belin, 1993). Dinoflagellates such as A.
minutum are therefore receiving increasing attention
due to the public health risk they pose and the economic
impacts they have on fisheries and aquaculture
developments (Hallegraeff et al., 1991).
The coastal waters of Jamaica have been the site of
several confirmed and unconfirmed red tides over the
years, sometimes accompanied by fish kills (Steven,
1966; Goodbody, 1970; Wade, 1971; Simmonds, 1997).
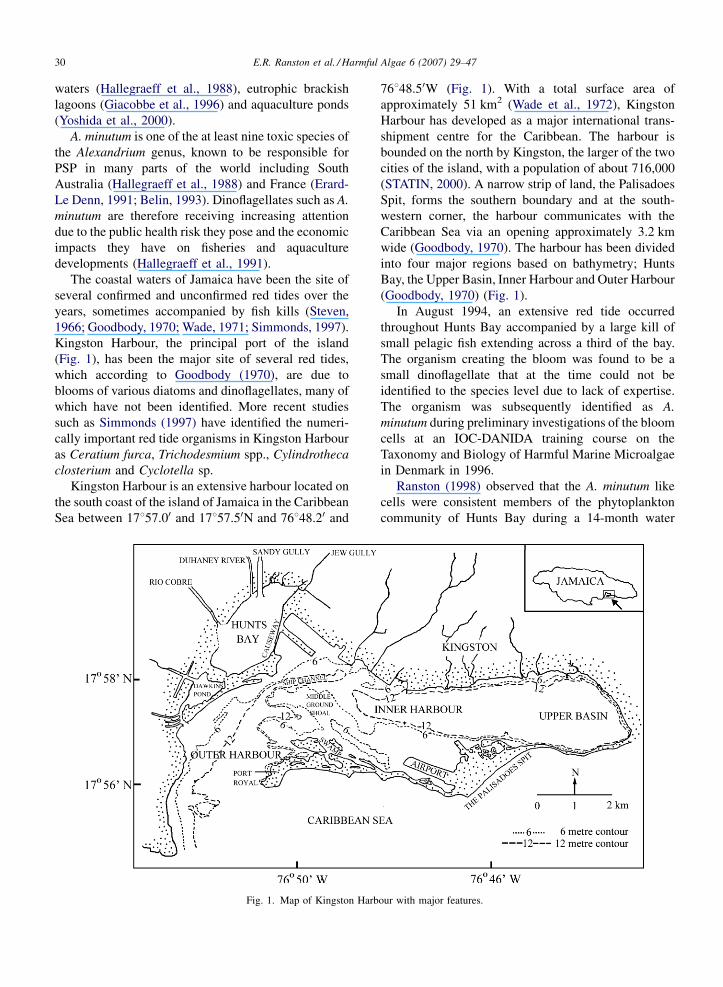
Kingston Harbour, the principal port of the island
(Fig. 1), has been the major site of several red tides,
which according to Goodbody (1970), are due to
blooms of various diatoms and dinoflagellates, many of
which have not been identified. More recent studies
such as Simmonds (1997) have identified the numeri-
cally important red tide organisms in Kingston Harbour
as Ceratium furca, Trichodesmium spp., Cylindrotheca
closterium and Cyclotella sp.
Kingston Harbour is an extensive harbour located on
the south coast of the island of Jamaica in the Caribbean
Sea between 17857.00 and 17857.50N and 76848.20 and
Fig. 1. Map of Kingston Harbour with major features.
76848.50W (Fig. 1). With a total surface area of
approximately 51 km2 (Wade et al., 1972), Kingston
Harbour has developed as a major international trans-
shipment centre for the Caribbean. The harbour is
bounded on the north by Kingston, the larger of the two
cities of the island, with a population of about 716,000
(STATIN, 2000). A narrow strip of land, the Palisadoes
Spit, forms the southern boundary and at the south-
western corner, the harbour communicates with the
Caribbean Sea via an opening approximately 3.2 km
wide (Goodbody, 1970). The harbour has been divided
into four major regions based on bathymetry; Hunts
Bay, the Upper Basin, Inner Harbour and Outer Harbour
(Goodbody, 1970) (Fig. 1).
In August 1994, an extensive red tide occurred
throughout Hunts Bay accompanied by a large kill of
small pelagic fish extending across a third of the bay.
The organism creating the bloom was found to be a
small dinoflagellate that at the time could not be
identified to the species level due to lack of expertise.
The organism was subsequently identified as A.
minutum during preliminary investigations of the bloom
cells at an IOC-DANIDA training course on the
Taxonomy and Biology of Harmful Marine Microalgae
in Denmark in 1996.
Ranston (1998) observed that the A. minutum like
cells were consistent members of the phytoplankton
community of Hunts Bay during a 14-month water

E.R. Ranston et al. / Harmful Algae 6 (2007) 29–47 31
quality study of the area extending from December
1993 to February 1995. In February 2004, another
extensive red tide was observed throughout the bay with
previously observed A. minutum like cells dominating.
A smaller bloom of the species was reported in April
2004; however this occurred outside of Hunts Bay along
a section of the northern shoreline of Kingston Harbour.
It is not known whether any illnesses were associated
with the previous and more recent blooms of this
species.
Countries such as Jamaica which experience blooms
of potentially toxic species and lack historical records of
these species can protect and prepare themselves from
potential problems through adequate species identifica-
tion, thus allowing scientists and officials to project
potential impacts to public health, aquaculture and
coastal community economy (Steidinger et al., 1989).
The main aim of this research was thus to confirm the
specific identity of the Alexandrium species in Hunts
Bay by examination and description of the morpholo-
gical characteristics of the cells and comparison with
classical taxonomical descriptions of Alexandrium,
such as Balech (1989, 1995). This paper therefore
describes the morphology of A. minutum discovered for
the first time in Jamaican coastal waters.
2. Materials and methods
2.1. Study site description
Hunts Bay is a relatively shallow, semi-enclosed arm
of the Kingston Harbour (Fig. 1), with an average depth
of about 2.4 m and an area of approximately 6.5 km2
(Ranston and Webber, 2003). The bay has a soft, black,
highly anoxic mud bottom and receives fresh water
discharges from two rivers: the Rio Cobre and the
Duhaney Rivers, and from two major gullies – the Jew
and Sandy Gullies (Fig. 1).
The Rio Cobre, one of the largest rivers of Jamaica,
drains major sugar cane fields and other agricultural
lands to the west of Hunts Bay and receives a number of
industrial and municipal discharges (Fig. 2). The
Duhaney River and Jew Gully pass through major
industrial sectors to the north and north-east of the bay,
respectively, and the Sandy Gully drains a very large
section of the residential area of the city of Kingston
which lies to the north of the harbour (Fig. 1). Near the
west-south-western shore of the bay, a small sewage
disposal culvert, the Portsmouth Sewage outfall, directs
semi-treated sewage into the bay (Fig. 2).
With all this nutrient input, the area has been
described as one under severe ecological stress,
exhibiting all the typical characteristics of a eutrophic
body of water (Wade, 1976), which are known to
encourage the development of red tides (Lam and Yip,
1990). Ranston (1998) confirmed increasing eutrophic
conditions in Hunts Bay and reported observing
obnoxious red tides occasionally accompanied by fish
kills, throughout the bay, during routine sampling of the
area over a 14-month period.
Hunts Bay is characterized by a water column
stratified into a fresh to brackish water surface layer, due
to freshwater input from the rivers and gullies, and a
saline deeper layer. The surface waters are often
supersaturated, high in nutrient concentrations, phyto-
plankton biomass and abundance, while deeper water is
characterized by oxygen deficiency, lower nutrient
concentrations and lower phytoplankton biomass and
abundance (Ranston and Webber, 2003).
Hunts Bay used to be a very productive and
important fishing and shrimping ground and also served
as a nursery area for young commercial species that are
caught in the open sea (Goodbody, 1970). The
decreasing water quality of the area over the years
has contributed to a drastic reduction of the fish and
shrimp populations of the bay. At present all that
remains is a small shrimping industry and an even
smaller fishery for bait and other small fish throughout
the bay, resulting in general hardship for fishermen of
the surrounding communities. Despite this decline,
Hunts Bay is still an important fishing, shrimping and to
a lesser extent, oyster/mussel collection area and still
functions to some extent as a nursery area for fish and
shrimp, as well as a feeding area for dolphins during the
beginning of each year. Everyday up to 30 fishing
vessels can be seen in the bay engaged in line and net
fishing, as well as the setting of fish and crab pots. The
small pelagic fish caught in the area are used as bait to
catch larger fish out in the open ocean. Some fishermen
collect and sell the various species of oysters and
mussels that grow on the mangrove roots surrounding
the bay. Hunts Bay is thus an important source of
livelihood for the many fishermen in the area, an
important feeding and nursery area for fish, shrimp and
marine mammals, and an important source of food for
the surrounding communities.
2.2. Sampling station locations
The A. minutum blooms coincided with routine water
monitoring exercises being conducted in Hunts Bay in
August 1994 and February 2004. Quantitative and
qualitative bloom samples were therefore collected at
the same stations established for the monitoring
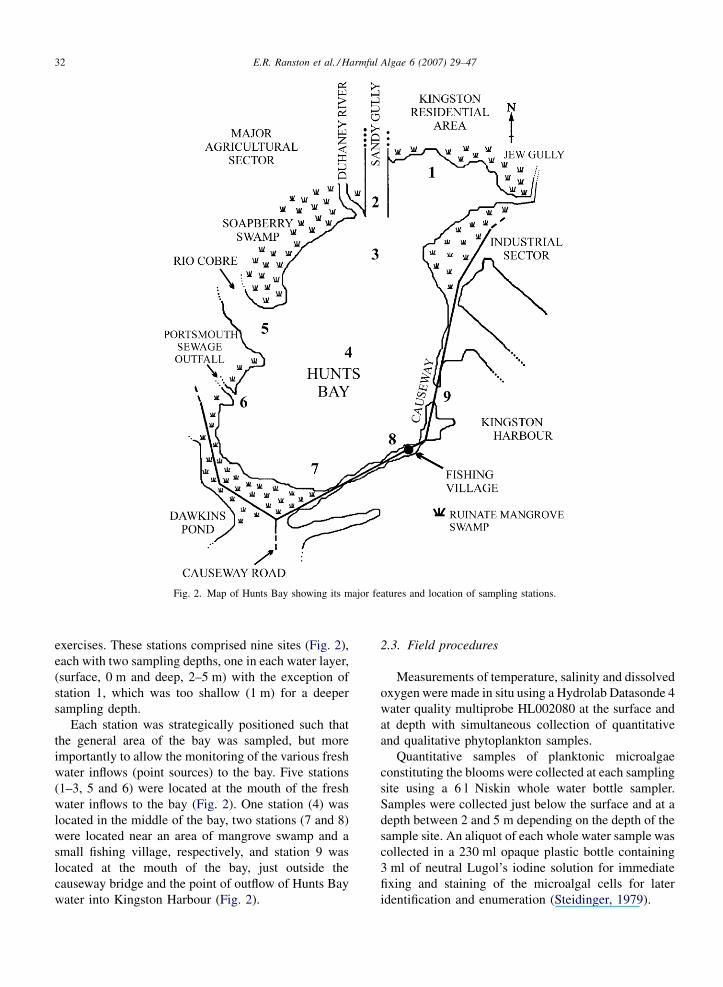
E.R. Ranston et al. / Harmful Algae 6 (2007) 29–4732
Fig. 2. Map of Hunts Bay showing its major features and location of sampling stations.
exercises. These stations comprised nine sites (Fig. 2),
each with two sampling depths, one in each water layer,
(surface, 0 m and deep, 2–5 m) with the exception of
station 1, which was too shallow (1 m) for a deeper
sampling depth.
Each station was strategically positioned such that
the general area of the bay was sampled, but more
importantly to allow the monitoring of the various fresh
water inflows (point sources) to the bay. Five stations
(1–3, 5 and 6) were located at the mouth of the fresh
water inflows to the bay (Fig. 2). One station (4) was
located in the middle of the bay, two stations (7 and 8)
were located near an area of mangrove swamp and a
small fishing village, respectively, and station 9 was
located at the mouth of the bay, just outside the
causeway bridge and the point of outflow of Hunts Bay
water into Kingston Harbour (Fig. 2).
2.3. Field procedures
Measurements of temperature, salinity and dissolved
oxygen were made in situ using a Hydrolab Datasonde 4
water quality multiprobe HL002080 at the surface and
at depth with simultaneous collection of quantitative
and qualitative phytoplankton samples.
Quantitative samples of planktonic microalgae
constituting the blooms were collected at each sampling
site using a 6 l Niskin whole water bottle sampler.
Samples were collected just below the surface and at a
depth between 2 and 5 m depending on the depth of the
sample site. An aliquot of each whole water sample was
collected in a 230 ml opaque plastic bottle containing
3 ml of neutral Lugol’s iodine solution for immediate
fixing and staining of the microalgal cells for later
identification and enumeration (Steidinger, 1979).

E.R. Ranston et al. / Harmful Algae 6 (2007) 29–47 33
Qualitative concentrated samples (net samples) of
phytoplankton were collected by hauling and towing a
20 mm plankton net through the water at each sampling
site. A vertical haul was conducted by gently lowering
the net to a point just above the seafloor and drawing the
net several times up through the water column until the
water in the net became unclear or coloured by the
concentrated algae. A horizontal surface tow was
conducted by allowing the net to sink below the water
surface followed by slow towing in a circular path
behind the boat for about 2 min. About 25 ml of one of
these samples was collected into a bottle and stored in a
cool, dark igloo as a live sample until analysis. Each net
sample was washed into a 230 ml opaque plastic bottle
containing 3 ml of neutral Lugol’s iodine solution,
using water in a spray bottle collected from each site.
Preserved qualitative and quantitative plankton samples
were stored in a cool dark area until analysis was
possible.
2.4. Laboratory procedures
2.4.1. Preliminary identification and enumeration
procedures
On return to the lab sub-samples of the live and
preserved planktonic net samples were observed on a
microscope slide using a Leitz Wetzlar Dialux 20 EB
compound microscope (model no. 020-452.008), in an
attempt to identify the dominant species of the bloom.
Quantitative determination of the species constitut-
ing the bloom was conducted using the Utermohl
method (Utermohl, 1958). Lugol’s preserved quantita-
tive samples were gently homogenised by inversion and
a 5 ml aliquot of each sample was made up to 10 ml
with filtered seawater in a 10 ml settling chamber. The
chambers were left to stand overnight to allow settling
of the phytoplankton cells before examination. Settled
samples were examined using a Leitz Labovert (model
no. 020-435.025) inverted microscope. Thirty random
fields of view of each settled sample were examined to
remove the edge effect in the settling of phytoplankton
cells (Sandgren and Robinson, 1984). The phytoplank-
ton in these thirty fields were identified and enumerated
at 320� and then each settled sample was fully scanned
at 100� to ensure that no phytoplankton species was
overlooked.
2.4.2. Confirmatory identification and microscopy
procedures
2.4.2.1. Scanning electron microscopy. Cells for elec-
tron microscopy were processed at the Electron
Microscopy Laboratory of the Botanical Institute,
Copenhagen University, using the following proce-
dures. Lugol’s preserved net samples of the bloom were
filtered directly on to a Nuclepore polycarbonate filter
by gravity filtration using a Swinnex filterholder.
Gravity filtration was used to thoroughly wash the
filtered samples with distilled water in order to get rid of
salt in the samples. After washing, samples were
chemically dehydrated using acidified 2,2-dimethox-
ypropane (DMP) (three drops 1N HCl to 25 ml DMP)
for a 10 min period. This dehydration process was
repeated and followed by critical point drying in which
the dehydration liquid in the samples was replaced by
liquid carbon dioxide, which was then vaporized. The
dried material on the filters was transferred on to a
double-sided adhesive carbon disc mounted on an SEM
stub. The material was coated with a thin layer of gold–
palladium applied by a sputtering process before
viewing using a scanning electron microscope. Electron
micrographs of the bloom cells were taken using a
camera attached to the microscope.
2.4.2.2. The squash technique. The squashing method
(Steidinger, 1979) was used to separate the thecal plates
of the bloom cells, allowing each plate to be observed
and used to aid in confirming the identity of the species.
A drop of a net sample from the bloom was placed on
a microscope slide and covered with a coverslip, gently
removing any excess seawater with a small piece of
blotting paper. The sample was observed using a Leitz
Wetzlar Dialux 20 EB compound microscope and a
Canon Power Shot G6 digital camera was used to
photograph the intact cells at 100� and 400� in an
attempt to determine the identity of the cells based on
size, shape and general morphological features.
A drop of 5% sodium hypochlorite (commercial
bleach) solution (1:1 mixture of sodium hypochlorite
and distilled water) was placed along a margin of the
coverslip and allowed to run under and across by
placing a small piece of blotting paper under the
opposite coverslip margin. A drop of neutral Lugol’s
solution was added in a similar manner in order to stain
the thecal plates. The cells were located under the
microscope and squashed by applying firm but gentle
pressure on the coverslip using a dissecting needle.
Individual thecal plates were identified with the aid
of descriptive and taxonomic references, with the
principal references being Balech (1989, 1995). Thecal
plates were viewed at 400� and 1000� with the aid of
immersion oil and photographs of these plates were
taken to show their morphological characteristics and
aid in the confirmation of the identity of the
dinoflagellate cells.

E.R. Ranston et al. / Harmful Algae 6 (2007) 29–4734
3. Results and discussion
3.1. Qualitative and quantitative analyses
The phytoplankton bloom that occurred in August
1994 in Hunts Bay was bright orange in colour and
concentrated in the fresh to brackish water surface layer
of the bay. The bloom was accompanied by a large kill
of small pelagic fish belonging to the clupeid and
carangid groups, which extended across approximately
2 km2 of the bay and average salinity, temperature and
dissolved oxygen concentrations of 11 ppt, 26.5 8C, and
6.50 mg l�1, respectively.
Qualitative analysis of the phytoplankton in the
bloom samples revealed that the samples were
dominated by a dinoflagellate belonging to the genus
Alexandrium, which occurred along with a few other
dinoflagellate and flagellate species. Preliminary exam-
inations of the Alexandrium cells led to the subsequent
identification of the species as A. minutum.
Quantitative analyses found that the highest cell
concentration of this species in the bloom was
4.6 � 105 cells l�1, a concentration which far exceeds
acceptable concentrations of PSP-toxin producing A.
minutum in several countries. In the Balearic Islands of
Spain, cell concentrations reaching 103 cells l�1 result
in intensified monitoring and closure of shellfishery
areas (Anderson, 1996) while in Denmark, a concen-
tration as low as 500 cells l�1 is considered to be the
maximum concentration for closing or imposing special
restrictions on shellfisheries (Anderson, 1996).
3.2. Species description and identification
The genus Alexandrium is notably homogenous with
the exception of a few species and lacks conspicuous
elements, frequent in other genera such as apical horns
and spines, which help to distinguish among species
(Balech, 1995). It is therefore necessary to conduct
extremely detailed observations in order to identify
Alexandrium cells to the species level, a process not
necessary for most dinoflagellates. Confirmation of the
identity of A. minutum cells in the red tide samples was
therefore based on examination of the general cell
morphology including characteristics such as size and
shape and more importantly, thecal plate pattern of
intact cells and the morphology of individual thecal
plates of squashed cells.
3.2.1. General morphology
Cells of this species were solitary and small in size, a
little longer than wide (Fig. 3A), with lengths ranging
from 18 to 31.5 mm (mean length � S.D. = 23.6 �2.11 mm) and widths ranging from 15.3 to 23.2 mm
(mean width � S.D. = 18.55 � 1.52 mm). These dimen-
sions are similar to those recorded by Balech (1989), who
reported thecae of the species ranging in length from 17
to 29 mm, with most ranging from 21 to 26 mm. Balech
(1995) reported that the width of A. minutum equals the
length, sometimes being larger but, more often, some-
what smaller, as observed for the Jamaican specimens.
Size is a secondary characteristic, as evidenced by the
large variations within certain species, however some
species such as A. minutum are differentiated by their
normally small size (Balech, 1995), as observed for the
Jamaican specimens.
Shape is an important taxonomic character although
it can be considerably altered by environmental
conditions, sexual reproductive stages such as zygotes,
small changes in position of the specimen, coverslip
pressure and adherence of the specimen to the glass
(Balech, 1995). The shape of the cells matched descrip-
tions put forward by Balech (1989, 1995) and varied
from irregularly oval to elliptical in dorsal view,
with roughly equal epitheca and hypotheca in length
(Fig. 3B). The epitheca varied in shape from hemi-
elliptical to almost hemispherical in both ventral
and dorsal views (Fig. 3B and C), while the shape of
the hypotheca ranged from hemielliptical in dorsal
view (Fig. 3B) to hemielliptical, with oblique antapical
flattening created by the sulcus, in ventral view
(Fig. 3C).
No spines, or horns were present and plate
ornamentation in the form of fairly strong, coarse,
irregular reticulations were observed on the hypo-
theca (Fig. 3D and E). The epitheca lacked any
obvious form of ornamentation and was therefore
smooth in comparison to the hypotheca (Fig. 3D).
Balech (1995) described A. minutum as typically
having the beginning of irregular sculpture rather than
the distinct, coarse reticulations observed in the
Jamaican specimens, but the degree of reticulation
was variable, possibly due to external factors such as
environmental conditions. He reported strong and true
reticulation in the hypotheca and barely perceptible
reticulation in the epitheca of Italian specimens of A.
minutum from the Gulf of Naples, which was similar
to that observed in the Jamaican specimens. Mon-
tresor et al., 1990 also described similar heavy
reticulation of hypothecal plates in specimens from
the Tyrrhenian Sea.
Each cell had a median cingulum that was deeply
excavated, left-handed, without lists and descending
one cingular width (c in Fig. 3F). The cingulum met a
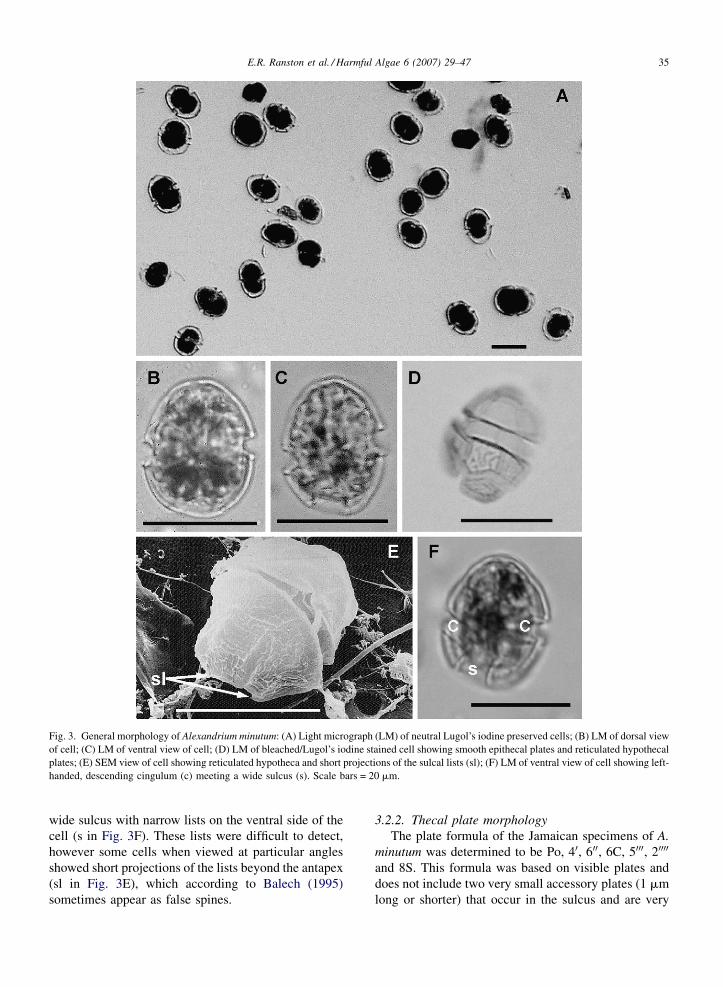
E.R. Ranston et al. / Harmful Algae 6 (2007) 29–47 35
Fig. 3. General morphology of Alexandrium minutum: (A) Light micrograph (LM) of neutral Lugol’s iodine preserved cells; (B) LM of dorsal view
of cell; (C) LM of ventral view of cell; (D) LM of bleached/Lugol’s iodine stained cell showing smooth epithecal plates and reticulated hypothecal
plates; (E) SEM view of cell showing reticulated hypotheca and short projections of the sulcal lists (sl); (F) LM of ventral view of cell showing left-
handed, descending cingulum (c) meeting a wide sulcus (s). Scale bars = 20 mm.
wide sulcus with narrow lists on the ventral side of the
cell (s in Fig. 3F). These lists were difficult to detect,
however some cells when viewed at particular angles
showed short projections of the lists beyond the antapex
(sl in Fig. 3E), which according to Balech (1995)
sometimes appear as false spines.
3.2.2. Thecal plate morphology
The plate formula of the Jamaican specimens of A.
minutum was determined to be Po, 40, 600, 6C, 5000, 20000
and 8S. This formula was based on visible plates and
does not include two very small accessory plates (1 mm
long or shorter) that occur in the sulcus and are very
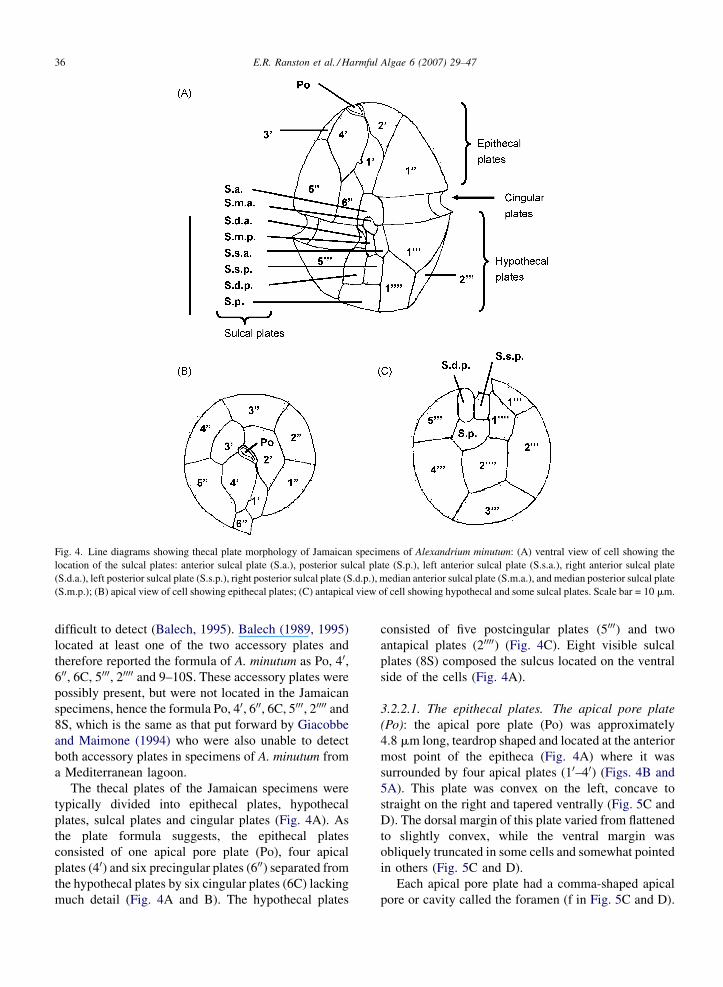
E.R. Ranston et al. / Harmful Algae 6 (2007) 29–4736
Fig. 4. Line diagrams showing thecal plate morphology of Jamaican specimens of Alexandrium minutum: (A) ventral view of cell showing the
location of the sulcal plates: anterior sulcal plate (S.a.), posterior sulcal plate (S.p.), left anterior sulcal plate (S.s.a.), right anterior sulcal plate
(S.d.a.), left posterior sulcal plate (S.s.p.), right posterior sulcal plate (S.d.p.), median anterior sulcal plate (S.m.a.), and median posterior sulcal plate
(S.m.p.); (B) apical view of cell showing epithecal plates; (C) antapical view of cell showing hypothecal and some sulcal plates. Scale bar = 10 mm.
difficult to detect (Balech, 1995). Balech (1989, 1995)
located at least one of the two accessory plates and
therefore reported the formula of A. minutum as Po, 40,600, 6C, 5000, 20000 and 9–10S. These accessory plates were
possibly present, but were not located in the Jamaican
specimens, hence the formula Po, 40, 600, 6C, 5000, 20000 and
8S, which is the same as that put forward by Giacobbe
and Maimone (1994) who were also unable to detect
both accessory plates in specimens of A. minutum from
a Mediterranean lagoon.
The thecal plates of the Jamaican specimens were
typically divided into epithecal plates, hypothecal
plates, sulcal plates and cingular plates (Fig. 4A). As
the plate formula suggests, the epithecal plates
consisted of one apical pore plate (Po), four apical
plates (40) and six precingular plates (600) separated from
the hypothecal plates by six cingular plates (6C) lacking
much detail (Fig. 4A and B). The hypothecal plates
consisted of five postcingular plates (5000) and two
antapical plates (20000) (Fig. 4C). Eight visible sulcal
plates (8S) composed the sulcus located on the ventral
side of the cells (Fig. 4A).
3.2.2.1. The epithecal plates. The apical pore plate
(Po): the apical pore plate (Po) was approximately
4.8 mm long, teardrop shaped and located at the anterior
most point of the epitheca (Fig. 4A) where it was
surrounded by four apical plates (10–40) (Figs. 4B and
5A). This plate was convex on the left, concave to
straight on the right and tapered ventrally (Fig. 5C and
D). The dorsal margin of this plate varied from flattened
to slightly convex, while the ventral margin was
obliquely truncated in some cells and somewhat pointed
in others (Fig. 5C and D).
Each apical pore plate had a comma-shaped apical
pore or cavity called the foramen (f in Fig. 5C and D).
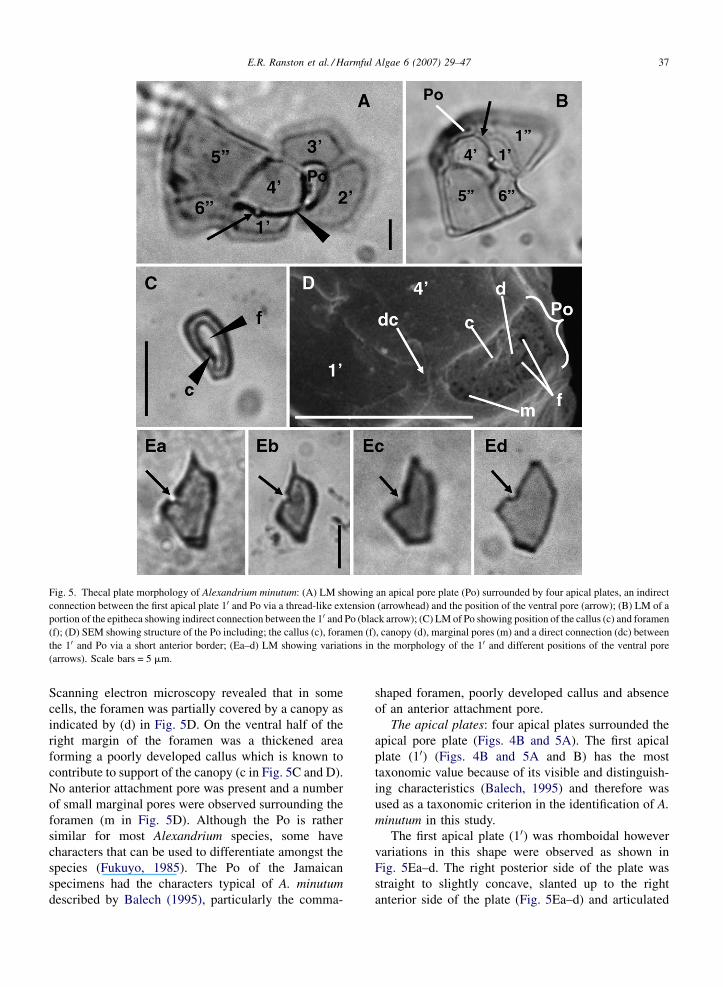
E.R. Ranston et al. / Harmful Algae 6 (2007) 29–47 37
Fig. 5. Thecal plate morphology of Alexandrium minutum: (A) LM showing an apical pore plate (Po) surrounded by four apical plates, an indirect
connection between the first apical plate 10 and Po via a thread-like extension (arrowhead) and the position of the ventral pore (arrow); (B) LM of a
portion of the epitheca showing indirect connection between the 10 and Po (black arrow); (C) LM of Po showing position of the callus (c) and foramen
(f); (D) SEM showing structure of the Po including; the callus (c), foramen (f), canopy (d), marginal pores (m) and a direct connection (dc) between
the 10 and Po via a short anterior border; (Ea–d) LM showing variations in the morphology of the 10 and different positions of the ventral pore
(arrows). Scale bars = 5 mm.
Scanning electron microscopy revealed that in some
cells, the foramen was partially covered by a canopy as
indicated by (d) in Fig. 5D. On the ventral half of the
right margin of the foramen was a thickened area
forming a poorly developed callus which is known to
contribute to support of the canopy (c in Fig. 5C and D).
No anterior attachment pore was present and a number
of small marginal pores were observed surrounding the
foramen (m in Fig. 5D). Although the Po is rather
similar for most Alexandrium species, some have
characters that can be used to differentiate amongst the
species (Fukuyo, 1985). The Po of the Jamaican
specimens had the characters typical of A. minutum
described by Balech (1995), particularly the comma-
shaped foramen, poorly developed callus and absence
of an anterior attachment pore.
The apical plates: four apical plates surrounded the
apical pore plate (Figs. 4B and 5A). The first apical
plate (10) (Figs. 4B and 5A and B) has the most
taxonomic value because of its visible and distinguish-
ing characteristics (Balech, 1995) and therefore was
used as a taxonomic criterion in the identification of A.
minutum in this study.
The first apical plate (10) was rhomboidal however
variations in this shape were observed as shown in
Fig. 5Ea–d. The right posterior side of the plate was
straight to slightly concave, slanted up to the right
anterior side of the plate (Fig. 5Ea–d) and articulated
E.R. Ranston et al. / Harmful Algae 6 (2007) 29–4738
with the sixth precingular plate (Figs. 4B and 5A and B).
The right anterior side of the 10 was in contact with the
fourth apical plate (40) (Figs. 4B and 5A and B) and was
slightly concave in some cases (Fig. 5Ea), straight in
others (Fig. 5Eb) and angular in a few cells (Fig. 5Ec).
All 10 had a small ventral pore which in most cells was
situated close to the posterior extreme of the anterior
right margin, on the suture between the first and fourth
apical plates as reported by Balech (1995) (black arrow
in Fig. 5A and black arrow in Fig. 5Ea). The presence of
the ventral pore is considered to be another important
taxonomic characteristic in the identification of A.
minutum (Balech, 1995). A small number of cells had
the ventral pore located close to the middle of the suture
as shown in Fig. 5Eb. A similar observation was made in
Vietnamese specimens of A. minutum (Yoshida et al.,
2000).
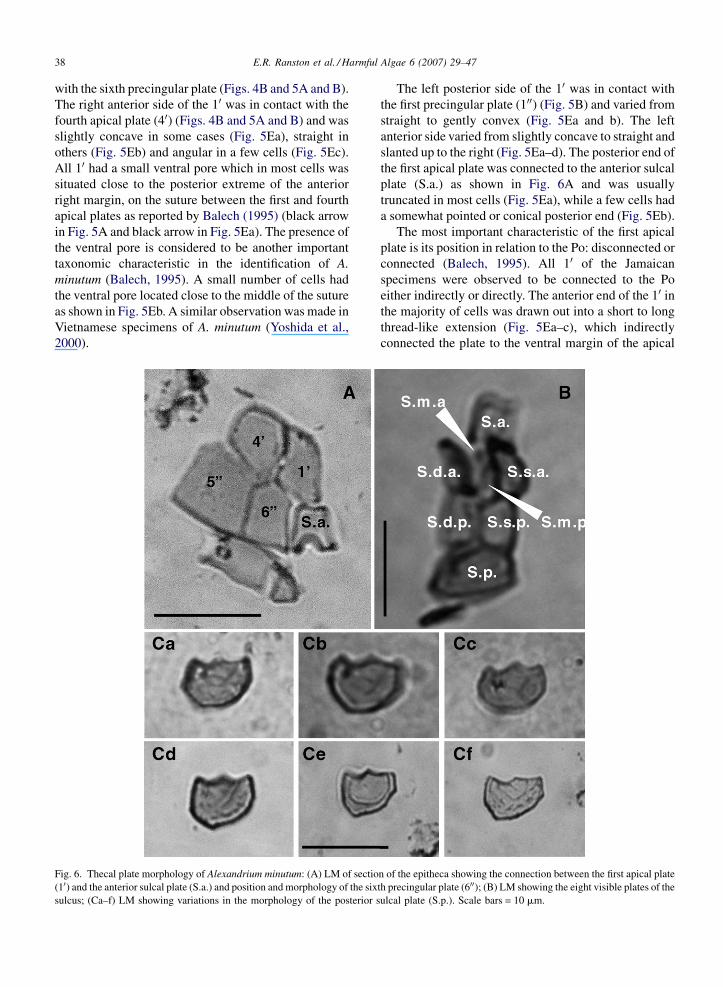
Fig. 6. Thecal plate morphology of Alexandrium minutum: (A) LM of sectio
(10) and the anterior sulcal plate (S.a.) and position and morphology of the six
sulcus; (Ca–f) LM showing variations in the morphology of the posterior s
The left posterior side of the 10 was in contact with
the first precingular plate (100) (Fig. 5B) and varied from
straight to gently convex (Fig. 5Ea and b). The left
anterior side varied from slightly concave to straight and
slanted up to the right (Fig. 5Ea–d). The posterior end of
the first apical plate was connected to the anterior sulcal
plate (S.a.) as shown in Fig. 6A and was usually
truncated in most cells (Fig. 5Ea), while a few cells had
a somewhat pointed or conical posterior end (Fig. 5Eb).
The most important characteristic of the first apical
plate is its position in relation to the Po: disconnected or
connected (Balech, 1995). All 10 of the Jamaican
specimens were observed to be connected to the Po
either indirectly or directly. The anterior end of the 10 inthe majority of cells was drawn out into a short to long
thread-like extension (Fig. 5Ea–c), which indirectly
connected the plate to the ventral margin of the apical
n of the epitheca showing the connection between the first apical plate
th precingular plate (600); (B) LM showing the eight visible plates of the
ulcal plate (S.p.). Scale bars = 10 mm.

E.R. Ranston et al. / Harmful Algae 6 (2007) 29–47 39
pore plate as indicated by the black arrowhead in
Fig. 5A and black arrow in Fig. 5B. Some 10 lacked the
extension altogether (Fig. 5Ed) and were directly
connected to the apical pore plate by a very short
anterior border as indicated by the arrow (dc) in Fig. 5D.
Balech (1989) similarly reported both direct and more
often indirect connection between the Po and 10 of his A.
minutum specimens.
Characteristics of the remaining apical plates were of
little taxonomic value with the exception of the third
apical plate (30) for which symmetry or asymmetry has
some differential value (Balech, 1995). The 30 of the
Jamaican specimens was six sided and appeared to be
almost symmetrical as shown in Figs. 4B and 5A. These
characters were typical of the 30 of A. minutum cells
described by Balech (1989).
The precingular plates: surrounding the apical plates
and immediately above the cingulum were six
precingular plates known as the first, second, third,
fourth, fifth and sixth precingulars (100–600, respectively)
(Fig. 4A and B). The most characteristic and
taxonomically valuable of these plates is the sixth
precingular (600) (Fig. 6A) which can be used to some
extent to differentiate between Alexandrium species
(Balech, 1995). In the Jamaican specimens, the 600 was
small and narrow—longer than wide in comparison to
the other precingular plates, with a length/width ratio of
about 2:1, typical of A. minutum species, as described
by Balech (1989). The right posterior margin of the 600
was straight to slightly convex and articulated with the
500 (Fig. 6A). The right anterior margin was straight to
concave and articulated with the fourth apical plate
(Fig. 6A). The left posterior margin was arched or
slightly concave, to allow for attachment of the convex
right margin of the anterior sulcal plate (S.a.) (Fig. 6A),
while the left anterior margin was straight or slightly
convex to allow for articulation of the 10.
3.2.2.2. The sulcal plates. All sulcal plates have
taxonomic value (Balech, 1995). Each cell had a large
ventrally located sulcus which widened slightly poster-
iorly (Fig. 3F) and has been reported as being composed
of ten plates, eight of which were relatively visible
(Figs. 4A and 6B) in the Jamaican specimens and two
which are very small and were not observed during this
study.
The eight visible sulcal plates are shown in Fig. 6B
and were composed of one anterior sulcal plate (S.a.)
and one posterior sulcal plate (S.p.). Two lateral pairs
of plates were present with one pair being posterior
and composed of the left and right posterior sulcal
plates (S.s.p. and S.d.p.). The other pair was anterior
and composed of the left and right anterior sulcal
plates (S.s.a and S.d.a.). Two small median plates
known as the median posterior and anterior sulcal
plates (S.m.p. and S.m.a.) occurred between the two
anterior lateral sulcal plates, one above the other. Of
these sulcal plates the most distinctive characters are
found in the posterior and anterior sulcal plates
(Fukuyo, 1985).
The posterior sulcal plate (S.p.): the posterior sulcal
plate was somewhat variable in the length/width ratio,
usually being wider than long (Fig. 6Ca), however some
cells had a S.p. with a length approximately equal to the
width as shown in Fig. 6Cb. Most S.p. were somewhat
symmetrical with an irregularly thickened anterior
margin composed of gently concave right and left
regions, which met in the middle of the margin
(Fig. 6Ca). In some cells, the left region of the anterior
margin was almost straight (Fig. 6Cf) and the right
region of the margin in some cases extended down into
the plate creating a small indentation as shown in
Fig. 6Ca and b. Both the left and right sides of the S.p.
varied from straight to slanting to gently convex, with
the left side usually shorter than the right side
(Fig. 6Ca–f). The posterior margin was generally
convex, slanting down toward the left and then curving
up towards the anterior margin (Fig. 6Ca–f).
The observed variations in the morphology of the
S.p. are considered to be important characteristics in the
identification of A. minutum and were similar to
variations reported by Balech (1989), who particularly
noted variation in the length/width ratio and shape of the
anterior edge.
The surface of the S.p. was irregularly reticulated as
was typical of the hypothecal plates and this reticulation
varied in pattern from cell to cell (Fig. 6Ca–d and f).
Some S.p. had less reticulation and a stria running near
the periphery of the left, right and posterior margins as
shown in Fig. 6Ce. No attachment pore was present on
the S.p. as also reported by both Yuki (1994) and
Yoshida et al. (2000) for A. minutum cells from Japan
and Vietnam, respectively. The anterior margin of the
S.p. articulated with two smaller lateral sulcal plates
known as the right and left posterior sulcal plates (S.d.p.
and S.s.p.) (Figs. 6B and 7A).
The right posterior sulcal plate (S.d.p.): the right
posterior sulcal plate (S.d.p.) was longer than wide and
had a generally rectangular shape which varied from
cell to cell (Fig. 7Ba–e), but fit previous descriptions of
this plate by Balech (1989, 1995). The posterior margin
of the S.d.p. articulated with the right region of the
anterior margin of the posterior sulcal plate (Fig. 7A)
and varied from straight to irregularly convex but most
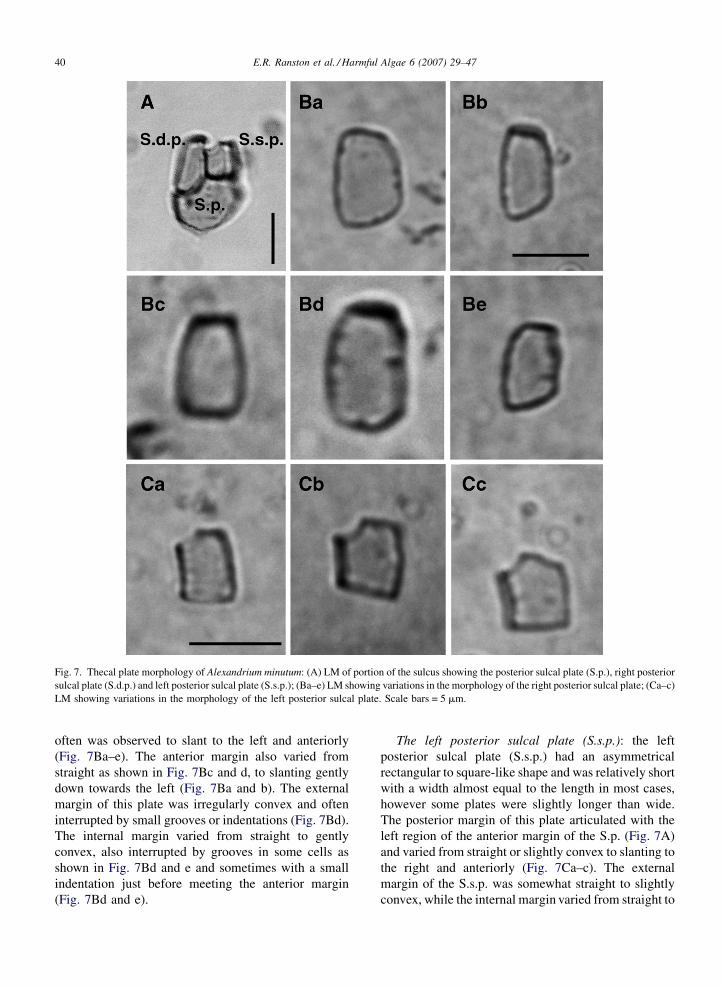
E.R. Ranston et al. / Harmful Algae 6 (2007) 29–4740
Fig. 7. Thecal plate morphology of Alexandrium minutum: (A) LM of portion of the sulcus showing the posterior sulcal plate (S.p.), right posterior
sulcal plate (S.d.p.) and left posterior sulcal plate (S.s.p.); (Ba–e) LM showing variations in the morphology of the right posterior sulcal plate; (Ca–c)
LM showing variations in the morphology of the left posterior sulcal plate. Scale bars = 5 mm.
often was observed to slant to the left and anteriorly
(Fig. 7Ba–e). The anterior margin also varied from
straight as shown in Fig. 7Bc and d, to slanting gently
down towards the left (Fig. 7Ba and b). The external
margin of this plate was irregularly convex and often
interrupted by small grooves or indentations (Fig. 7Bd).
The internal margin varied from straight to gently
convex, also interrupted by grooves in some cells as
shown in Fig. 7Bd and e and sometimes with a small
indentation just before meeting the anterior margin
(Fig. 7Bd and e).
The left posterior sulcal plate (S.s.p.): the left
posterior sulcal plate (S.s.p.) had an asymmetrical
rectangular to square-like shape and was relatively short
with a width almost equal to the length in most cases,
however some plates were slightly longer than wide.
The posterior margin of this plate articulated with the
left region of the anterior margin of the S.p. (Fig. 7A)
and varied from straight or slightly convex to slanting to
the right and anteriorly (Fig. 7Ca–c). The external
margin of the S.s.p. was somewhat straight to slightly
convex, while the internal margin varied from straight to
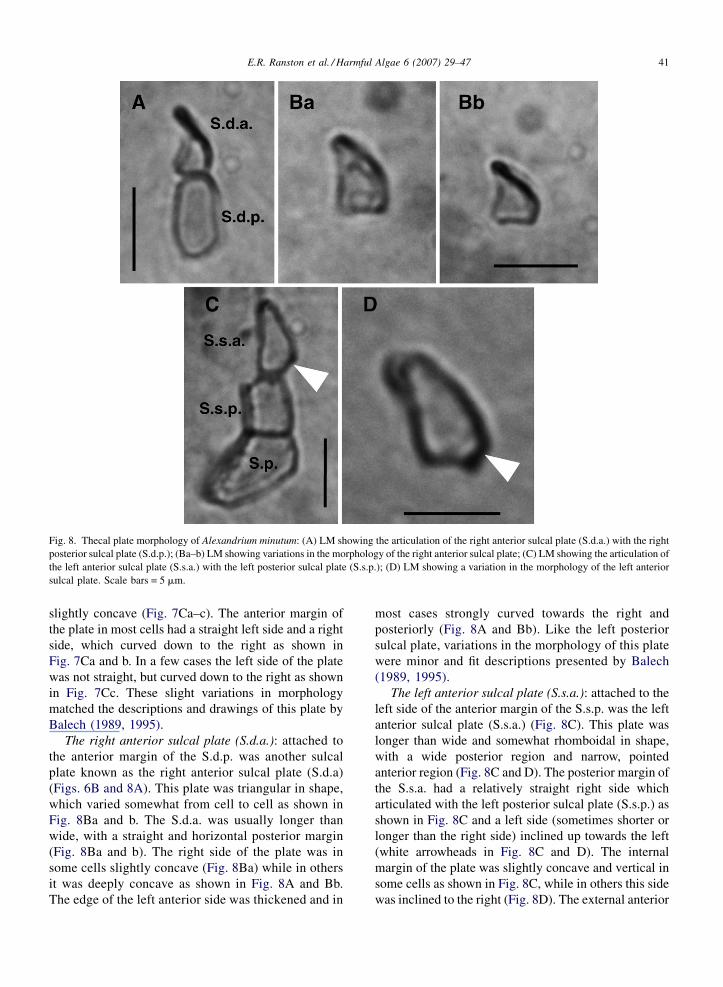
E.R. Ranston et al. / Harmful Algae 6 (2007) 29–47 41
Fig. 8. Thecal plate morphology of Alexandrium minutum: (A) LM showing the articulation of the right anterior sulcal plate (S.d.a.) with the right
posterior sulcal plate (S.d.p.); (Ba–b) LM showing variations in the morphology of the right anterior sulcal plate; (C) LM showing the articulation of
the left anterior sulcal plate (S.s.a.) with the left posterior sulcal plate (S.s.p.); (D) LM showing a variation in the morphology of the left anterior
sulcal plate. Scale bars = 5 mm.
slightly concave (Fig. 7Ca–c). The anterior margin of
the plate in most cells had a straight left side and a right
side, which curved down to the right as shown in
Fig. 7Ca and b. In a few cases the left side of the plate
was not straight, but curved down to the right as shown
in Fig. 7Cc. These slight variations in morphology
matched the descriptions and drawings of this plate by
Balech (1989, 1995).
The right anterior sulcal plate (S.d.a.): attached to
the anterior margin of the S.d.p. was another sulcal
plate known as the right anterior sulcal plate (S.d.a)
(Figs. 6B and 8A). This plate was triangular in shape,
which varied somewhat from cell to cell as shown in
Fig. 8Ba and b. The S.d.a. was usually longer than
wide, with a straight and horizontal posterior margin
(Fig. 8Ba and b). The right side of the plate was in
some cells slightly concave (Fig. 8Ba) while in others
it was deeply concave as shown in Fig. 8A and Bb.
The edge of the left anterior side was thickened and in
most cases strongly curved towards the right and
posteriorly (Fig. 8A and Bb). Like the left posterior
sulcal plate, variations in the morphology of this plate
were minor and fit descriptions presented by Balech
(1989, 1995).
The left anterior sulcal plate (S.s.a.): attached to the
left side of the anterior margin of the S.s.p. was the left
anterior sulcal plate (S.s.a.) (Fig. 8C). This plate was
longer than wide and somewhat rhomboidal in shape,
with a wide posterior region and narrow, pointed
anterior region (Fig. 8C and D). The posterior margin of
the S.s.a. had a relatively straight right side which
articulated with the left posterior sulcal plate (S.s.p.) as
shown in Fig. 8C and a left side (sometimes shorter or
longer than the right side) inclined up towards the left
(white arrowheads in Fig. 8C and D). The internal
margin of the plate was slightly concave and vertical in
some cells as shown in Fig. 8C, while in others this side
was inclined to the right (Fig. 8D). The external anterior
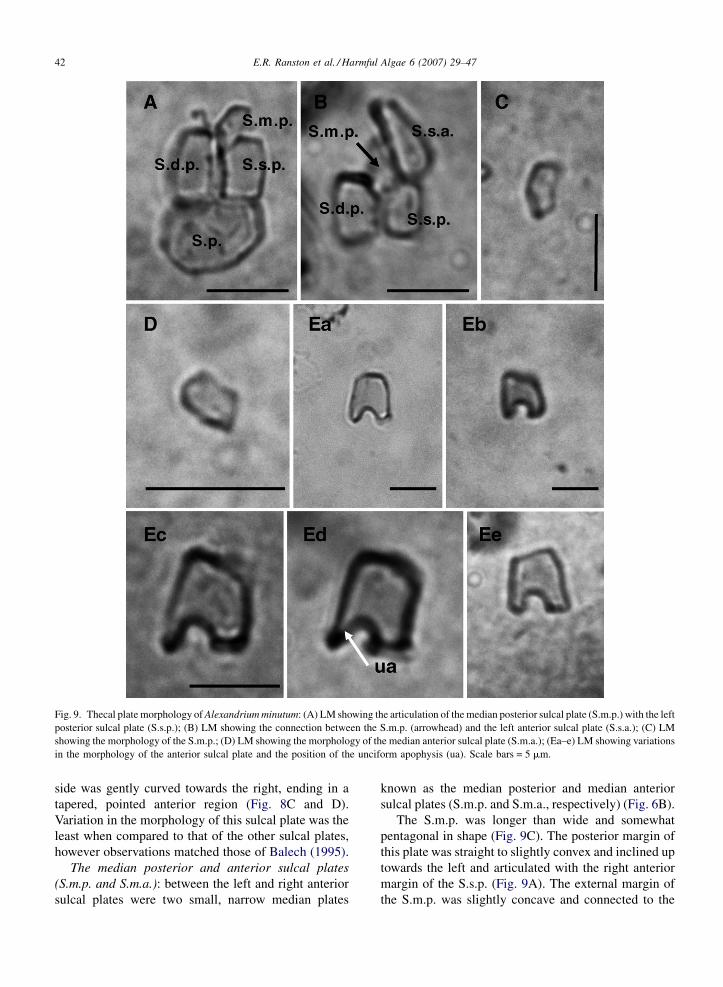
E.R. Ranston et al. / Harmful Algae 6 (2007) 29–4742
Fig. 9. Thecal plate morphology of Alexandrium minutum: (A) LM showing the articulation of the median posterior sulcal plate (S.m.p.) with the left
posterior sulcal plate (S.s.p.); (B) LM showing the connection between the S.m.p. (arrowhead) and the left anterior sulcal plate (S.s.a.); (C) LM
showing the morphology of the S.m.p.; (D) LM showing the morphology of the median anterior sulcal plate (S.m.a.); (Ea–e) LM showing variations
in the morphology of the anterior sulcal plate and the position of the unciform apophysis (ua). Scale bars = 5 mm.
side was gently curved towards the right, ending in a
tapered, pointed anterior region (Fig. 8C and D).
Variation in the morphology of this sulcal plate was the
least when compared to that of the other sulcal plates,
however observations matched those of Balech (1995).
The median posterior and anterior sulcal plates
(S.m.p. and S.m.a.): between the left and right anterior
sulcal plates were two small, narrow median plates
known as the median posterior and median anterior
sulcal plates (S.m.p. and S.m.a., respectively) (Fig. 6B).
The S.m.p. was longer than wide and somewhat
pentagonal in shape (Fig. 9C). The posterior margin of
this plate was straight to slightly convex and inclined up
towards the left and articulated with the right anterior
margin of the S.s.p. (Fig. 9A). The external margin of
the S.m.p. was slightly concave and connected to the
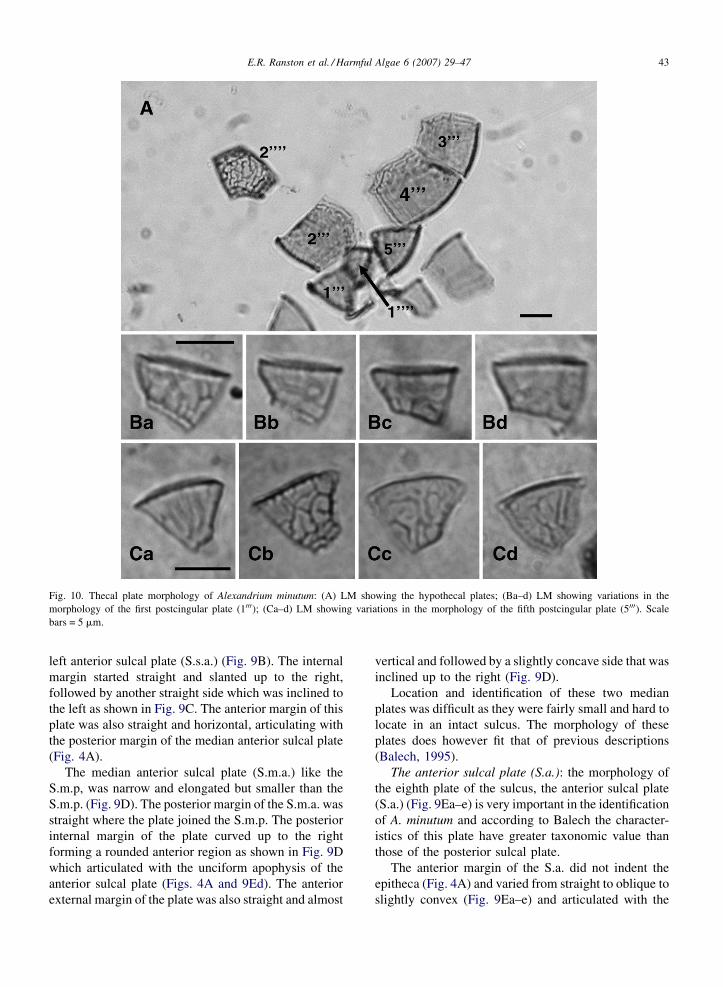
E.R. Ranston et al. / Harmful Algae 6 (2007) 29–47 43
Fig. 10. Thecal plate morphology of Alexandrium minutum: (A) LM showing the hypothecal plates; (Ba–d) LM showing variations in the
morphology of the first postcingular plate (1000); (Ca–d) LM showing variations in the morphology of the fifth postcingular plate (5000). Scale
bars = 5 mm.
left anterior sulcal plate (S.s.a.) (Fig. 9B). The internal
margin started straight and slanted up to the right,
followed by another straight side which was inclined to
the left as shown in Fig. 9C. The anterior margin of this
plate was also straight and horizontal, articulating with
the posterior margin of the median anterior sulcal plate
(Fig. 4A).
The median anterior sulcal plate (S.m.a.) like the
S.m.p, was narrow and elongated but smaller than the
S.m.p. (Fig. 9D). The posterior margin of the S.m.a. was
straight where the plate joined the S.m.p. The posterior
internal margin of the plate curved up to the right
forming a rounded anterior region as shown in Fig. 9D
which articulated with the unciform apophysis of the
anterior sulcal plate (Figs. 4A and 9Ed). The anterior
external margin of the plate was also straight and almost
vertical and followed by a slightly concave side that was
inclined up to the right (Fig. 9D).
Location and identification of these two median
plates was difficult as they were fairly small and hard to
locate in an intact sulcus. The morphology of these
plates does however fit that of previous descriptions
(Balech, 1995).
The anterior sulcal plate (S.a.): the morphology of
the eighth plate of the sulcus, the anterior sulcal plate
(S.a.) (Fig. 9Ea–e) is very important in the identification
of A. minutum and according to Balech the character-
istics of this plate have greater taxonomic value than
those of the posterior sulcal plate.
The anterior margin of the S.a. did not indent the
epitheca (Fig. 4A) and varied from straight to oblique to
slightly convex (Fig. 9Ea–e) and articulated with the
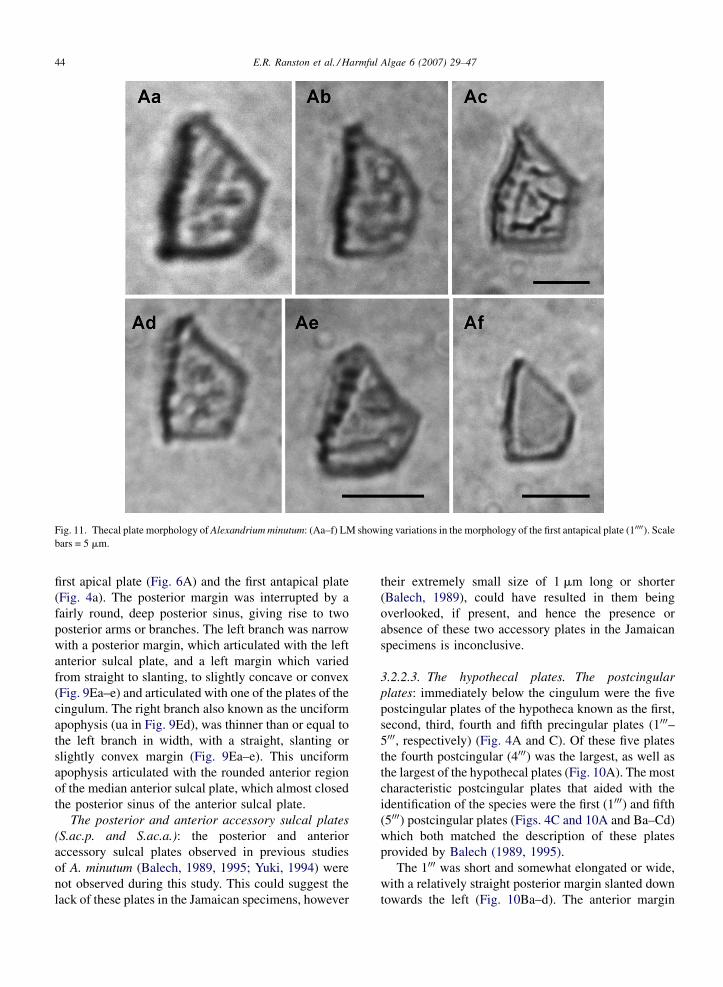
E.R. Ranston et al. / Harmful Algae 6 (2007) 29–4744
Fig. 11. Thecal plate morphology of Alexandrium minutum: (Aa–f) LM showing variations in the morphology of the first antapical plate (10000). Scale
bars = 5 mm.
first apical plate (Fig. 6A) and the first antapical plate
(Fig. 4a). The posterior margin was interrupted by a
fairly round, deep posterior sinus, giving rise to two
posterior arms or branches. The left branch was narrow
with a posterior margin, which articulated with the left
anterior sulcal plate, and a left margin which varied
from straight to slanting, to slightly concave or convex
(Fig. 9Ea–e) and articulated with one of the plates of the
cingulum. The right branch also known as the unciform
apophysis (ua in Fig. 9Ed), was thinner than or equal to
the left branch in width, with a straight, slanting or
slightly convex margin (Fig. 9Ea–e). This unciform
apophysis articulated with the rounded anterior region
of the median anterior sulcal plate, which almost closed
the posterior sinus of the anterior sulcal plate.
The posterior and anterior accessory sulcal plates
(S.ac.p. and S.ac.a.): the posterior and anterior
accessory sulcal plates observed in previous studies
of A. minutum (Balech, 1989, 1995; Yuki, 1994) were
not observed during this study. This could suggest the
lack of these plates in the Jamaican specimens, however
their extremely small size of 1 mm long or shorter
(Balech, 1989), could have resulted in them being
overlooked, if present, and hence the presence or
absence of these two accessory plates in the Jamaican
specimens is inconclusive.
3.2.2.3. The hypothecal plates. The postcingular
plates: immediately below the cingulum were the five
postcingular plates of the hypotheca known as the first,
second, third, fourth and fifth precingular plates (1000–5000, respectively) (Fig. 4A and C). Of these five plates
the fourth postcingular (4000) was the largest, as well as
the largest of the hypothecal plates (Fig. 10A). The most
characteristic postcingular plates that aided with the
identification of the species were the first (1000) and fifth
(5000) postcingular plates (Figs. 4C and 10A and Ba–Cd)
which both matched the description of these plates
provided by Balech (1989, 1995).
The 1000 was short and somewhat elongated or wide,
with a relatively straight posterior margin slanted down
towards the left (Fig. 10Ba–d). The anterior margin
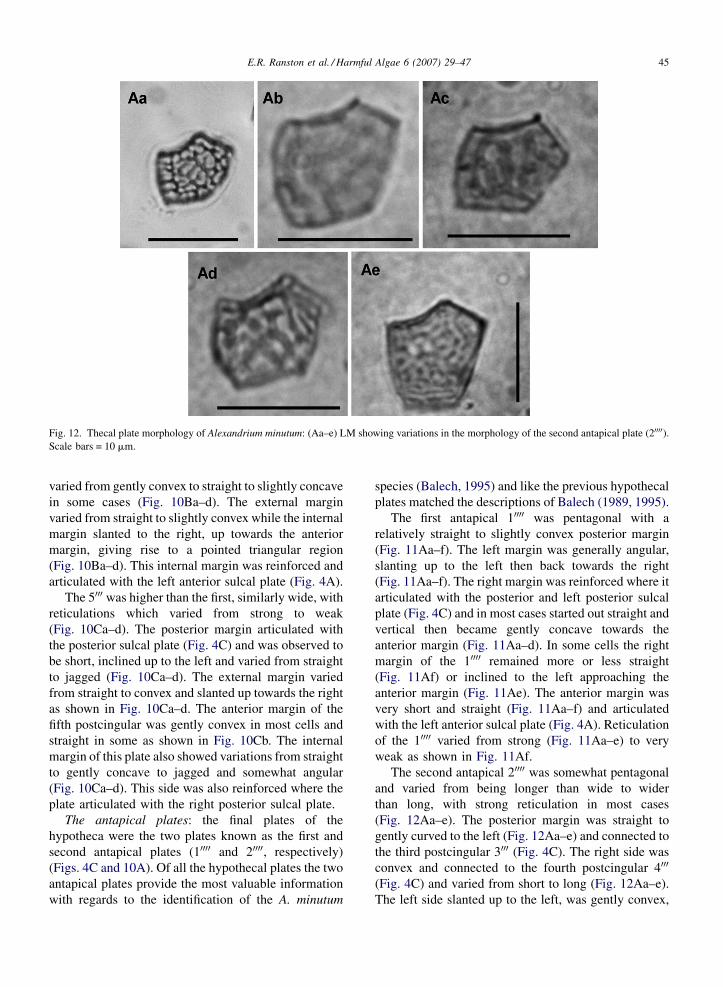
E.R. Ranston et al. / Harmful Algae 6 (2007) 29–47 45
Fig. 12. Thecal plate morphology of Alexandrium minutum: (Aa–e) LM showing variations in the morphology of the second antapical plate (20000).Scale bars = 10 mm.
varied from gently convex to straight to slightly concave
in some cases (Fig. 10Ba–d). The external margin
varied from straight to slightly convex while the internal
margin slanted to the right, up towards the anterior
margin, giving rise to a pointed triangular region
(Fig. 10Ba–d). This internal margin was reinforced and
articulated with the left anterior sulcal plate (Fig. 4A).
The 5000 was higher than the first, similarly wide, with
reticulations which varied from strong to weak
(Fig. 10Ca–d). The posterior margin articulated with
the posterior sulcal plate (Fig. 4C) and was observed to
be short, inclined up to the left and varied from straight
to jagged (Fig. 10Ca–d). The external margin varied
from straight to convex and slanted up towards the right
as shown in Fig. 10Ca–d. The anterior margin of the
fifth postcingular was gently convex in most cells and
straight in some as shown in Fig. 10Cb. The internal
margin of this plate also showed variations from straight
to gently concave to jagged and somewhat angular
(Fig. 10Ca–d). This side was also reinforced where the
plate articulated with the right posterior sulcal plate.
The antapical plates: the final plates of the
hypotheca were the two plates known as the first and
second antapical plates (10000 and 20000, respectively)
(Figs. 4C and 10A). Of all the hypothecal plates the two
antapical plates provide the most valuable information
with regards to the identification of the A. minutum
species (Balech, 1995) and like the previous hypothecal
plates matched the descriptions of Balech (1989, 1995).
The first antapical 10000 was pentagonal with a
relatively straight to slightly convex posterior margin
(Fig. 11Aa–f). The left margin was generally angular,
slanting up to the left then back towards the right
(Fig. 11Aa–f). The right margin was reinforced where it
articulated with the posterior and left posterior sulcal
plate (Fig. 4C) and in most cases started out straight and
vertical then became gently concave towards the
anterior margin (Fig. 11Aa–d). In some cells the right
margin of the 10000 remained more or less straight
(Fig. 11Af) or inclined to the left approaching the
anterior margin (Fig. 11Ae). The anterior margin was
very short and straight (Fig. 11Aa–f) and articulated
with the left anterior sulcal plate (Fig. 4A). Reticulation
of the 10000 varied from strong (Fig. 11Aa–e) to very
weak as shown in Fig. 11Af.
The second antapical 20000 was somewhat pentagonal
and varied from being longer than wide to wider
than long, with strong reticulation in most cases
(Fig. 12Aa–e). The posterior margin was straight to
gently curved to the left (Fig. 12Aa–e) and connected to
the third postcingular 3000 (Fig. 4C). The right side was
convex and connected to the fourth postcingular 4000
(Fig. 4C) and varied from short to long (Fig. 12Aa–e).
The left side slanted up to the left, was gently convex,

E.R. Ranston et al. / Harmful Algae 6 (2007) 29–4746
connected to the second postcingular 2000 (Fig. 4C) and
also varied from short to long (Fig. 12Aa–e). The left
side of the anterior margin was straight, slanting to the
right and connected to the first antapical 10000 (Figs. 4C
and 12Aa–e). This side was followed by a concave right
side, which connected, to the posterior sulcal plate
(Figs. 4C and 12Aa–e).
4. Conclusion
The reported observations on the morphology and
specifically the plate patterns of the Jamaican speci-
mens of A. minutum were in good agreement with
classical descriptions of A. minutum presented by
Balech (1989, 1995) and do not present substantial
differences in plate tabulation and morphology.
A. minutum is characterized by its small size, the
rhomboidal first apical 10 which is directly or, more
often, indirectly connected to the comma shaped Po, a
ventral pore which is situated in the posterior half of the
right upper side of 10, the narrow S.a. as long as wide or
longer than wide, the short, wider than long S.p., the
narrow precingular 600 and the constant lack of anterior
and posterior attachment pores (Balech, 1995). These
are all characters, which are defined as typical of A.
minutum and are used to distinguish this species from
other species of Alexandrium, and were characters
possessed by the Jamaican specimens.
These specimens showed some slight variations from
the typical description of the species, however these are
not considered to be significant. A distinctly reticulated
hypotheca as seen in the Jamaican specimens is not
considered to be typical (Balech, 1995), but A. minutum
cells from Italy also had strong reticulations suggesting
high variability in the appearance of the hypothecal
surface possibly due to environmental conditions
(Balech, 1995). The ventral pore of the Jamaican
specimens was not always near the posterior margin of
the 10, but sometimes in the middle. Such a variation
was also described by Yoshida et al. (2000) for A.
minutum specimens from Vietnam.
To date nostudiesof the toxic potentialof the Jamaican
specimens of A. minutum have been conducted, however
all Alexandrium species should be regarded as potentially
toxic until proven otherwise. The fact that a massive fish
kill accompanied the bloom suggests the possibility that
the Jamaican specimens of A. minutum are toxic. The fish
kill may have been as a result of oxygen depletion during
the bloom, however the possibility exists that death could
have occurred due to close contact with, or direct
ingestion of paralytic shellfish toxin producing A.
minutum cells, or ingestion of zooplankton which had
fed on the cells (White, 1981). The relationships between
fish kills, paralytic shellfish toxins and the marine food
web have been reviewed by White (1984). Marine fish are
in general very sensitive to paralytic shellfish toxins
although some species can accumulate and retain the
toxins without being affected (Geraci et al., 1989).
With potentially toxic A. minutum being a consistent
and sometimes dominant member of the phytoplankton
community of Hunts Bay, and with concentrations far
exceeding the acceptable limits in other countries, the
continual use of this bay as a source of food is cause for
great concern. Fishermen of surrounding communities
fish for finfish, crabs, shrimp and shellfish in Hunts Bay
everyday, including during red tide occurrences. Bait fish
caught every morning in the bay are used to catch larger
fish outat sea, compounding the potential for toxic events.
There are no known cases of PSP in Jamaica, but the
potential for its occurrence is ever present as long as A.
minutum continues to occur and bloom in Hunts Bay
and possibly other Jamaican coastal waters that are used
as food sources by humans and other animals. PSP is
appearing in regions where it has never been known
(Kao, 1993) and Jamaica is certainly one of these
regions. With no historical data on this species and lack
of expertise on PSP events, such an occurrence could
have detrimental effects on commercial fisheries, public
health and the economies of local areas in the country.
Acknowledgements
We gratefully acknowledge the IOC-DANIDA
(Intergovernmental Oceanographic Commission, Dan-
ish Agency for International Development Aid) Science
and Communication Centre on Harmful Algae for
funding this study and providing literature and training
at workshops on the identification of harmful marine
microalgae at the University of Copenhagen. Thanks to
staff of the Botanical Institute, University of Copenha-
gen for their assistance with the electron microscope
observations and photography. Thanks also to The
Department of Life Sciences, University of the West
Indies, Mona Campus in Jamaica, for use of inverted
and compound microscopes and the Port Royal Marine
Laboratory in Jamaica for providing boat transport and
accommodation during the field work phase of both the
1994 and 2004 exercises.[SS]
References
Anderson, P., 1996. Design and implementation of some harmful algal
monitoring systems. In: IOC Technical Series No. 44. UNESCO,
Paris 102pp.

E.R. Ranston et al. / Harmful Algae 6 (2007) 29–47 47
Balech, E., 1989. Redescription of Alexandrium minutum Halim
(dinophyceae) type species of the genus Alexandrium. Phycologia
28, 206–211.
Balech, E., 1995. The Genus Alexandrium Halim (Dinoflagellata).
Sherkin Island Marine Station, Sherkin Island Co., Cork, Ireland,
151 pp.
Belin, C., 1993. Distribution of Dinophysis spp. and Alexandrium
minutum along French coasts since 1984 and their DSP and PSP
toxicity levels. In: Smayda, T.J., Shimizu, Y. (Eds.), Toxic Phy-
toplankton Blooms in the Sea. Elsevier, Amsterdam, pp. 469–474.
Chang, F.H., Anderson, D.M., Kulis, D.M., Till, D.G., 1997. Toxin
production of Alexandrium minutum (Dinophyceae) from the Bay
of Plenty, New Zealand. Toxicon 35, 393–409.
Erard-Le Denn, E., 1991. Recent occurrence of red tide dinoflagellate
Alexandrium minutum Halim from the north western coasts of
France. In: Park, J.S., Kim, H.J. (Eds.), Recent Approaches on
Red Tides. Department of Oceanography and Marine Resources,
National Fisheries Research & Development Agency, Republic of
Korea, pp. 85–98.
Fukuyo, Y., 1985. Morphology of Protogonyaulax tamarensis
(Lebour) Taylor and Protogonyaulax catenella (Whedon and
Kofoid) Taylor from Japanese waters. Bull. Mar. Sci. 37, 529–537.
Geraci, J.A., Anderson, D.M., Timperi, R.J., St. Aubin, D.J., Early,
G.A., Prescott, J.A., Mayo, C.A., 1989. Humpback whales (Mega-
ptera novaeangliae) fatally poisoned by dinoflagellate toxin. Can.
J. Fish. Aquat. Sci. 46, 1895–1898.
Giacobbe, M.G., Maimone, G., 1994. First report of Alexandrium
minutum Halim in a Mediterranean lagoon. Cryptogamie, Algol
15, 47–52.
Giacobbe, M.G., Oliva, F.D., Maimone, G., 1996. Environmental
factors and seasonal occurrence of the dinoflagellate Alexandrium
minutum, a PSP potential producer, in a Mediterranean lagoon.
Est. Coast. Shelf Sci. 42, 539–549.
Goodbody, I.M., 1970. The biology of Kingston harbour. J. Sci. Res.
Counc., Jamaica 1, 10–34.
Halim, Y., 1960. Alexandrium minutum nov. g. nov. sp. dinoflagellate
provocant des ‘‘eaux rouges’’ Vie et Milieu 11, 102–105.
Hallegraeff, G.M., Steffensen, D.A., Wetherbee, R., 1988. Three
estuarine Australian dinoflagellates that can produce paralytic
shellfish toxins. J. Plankton Res. 10, 533–541.
Hallegraeff, G.M., Bolch, S.I., Blackburn, S.I., Oshima, Y., 1991.
Species of the toxigenic dinoflagellate genus Alexandrium in
South-eastern Australian waters. Botanica Marina 34, 575–587.
Kao, C.Y., 1993. Paralytic shellfish poisoning. In: Falconer, I.R.
(Ed.), Algal Toxins in Seafood and Drinking Water. Academic
Press, London, pp. 75–86.
Lam, C.W.Y., Yip, S.S.Y., 1990. A three-month red tide event in Hong
Kong. In: Graneli, E., Sundstrom, B., Edler, L., Anderson, D.M.
(Eds.), Toxic Marine Phytoplankton: Proceedings of the Fourth
International Conference on Toxic Marine Phytoplankton, Lund,
Sweden, June 26–30. Elsevier, New York, pp. 481–486.
Montresor, M., Marino, D., Zingone, A., Dafnis, G., 1990. Three
Alexandrium species from coastal Tyrrhenian waters (Mediterra-
nean Sea). In: Graneli, E., Sundstrom, B., Edler, L., Anderson,
D.M. (Eds.), Toxic Marine Phytoplankton: Proceedings of the
Fourth International Conference on Toxic Marine Phytoplankton,
Lund, Sweden, June 26–30. Elsevier, New York, pp. 82–87.
Ranston, E.R., 1998. The Phytoplankton Community and Water
Quality of a Highly Eutrophic Bay: Hunts Bay, Kingston Harbour,
Jamaica. MPhil. Thesis. University of the West Indies, Mona,
Jamaica, 249 pp.
Ranston, E.R., Webber, D.F., 2003. Phytoplankton distribution in a
highly eutrophic estuarine bay, Hunts Bay, Kingston Harbour,
Jamaica. Bull. Mar. Sci. 73, 307–324.
Sandgren, C.D., Robinson, J.V., 1984. A stratified sampling approach
to compensating for non-random sedimentation of phytoplankton
cells in inverted microscope settling chambers. Br. Phycol. J. 19,
67–72.
Simmonds, R.A., 1997. The Phytoplankton Community and Water
Quality of a Eutrophic Embayment; Kingston Harbor, Jamaica.
MPhil. Thesis. University of the West Indies, Mona, Jamaica, 214
pp.
STATIN, 2000. Demographic Statistics 2000. Statistical Institute of
Jamaica, STATIN Press, Kingston, 63 pp.
Steidinger, K.A., 1979. Collection, enumeration and identification, of
free-living dinoflagellates. In: Taylor, D.L., Selinger, H.H. (Eds.),
Toxic Dinoflagellate Blooms. Elsevier, North Holland, Inc., pp.
435–442.
Steidinger, K., Babcock, C., Mahmoudi, B., Tomas, C., Truby, E.,
1989. Conservative taxonomic characters in toxic dinoflagellate
species identification. In: Okaichi, T., Anderson, D.M., Nemoto,
T. (Eds.), Red Tides: Biology, Environmental Science and Tox-
icology. Elsevier, Amsterdam, pp. 285–288.
Steven, D.M., 1966. Characteristics of a red water bloom in Kingston
Harbour, Jamaica, West Indies. J. Mar. Res. 24, 113–123.
Utermohl, H., 1958. Zur vervollkommung der qualitativen phyto-
plankton methodisk. Mitteilungen Internationale Veroingung fur
Theoretische und Sgeurandt Limnologie 9, 1–38.
Wade, B.A., 1971. Marine pollution problems in Jamaica. Mar. Poll.
Bull. 2, 1.
Wade, B.A., 1976. The Pollution Ecology of Kingston Harbour,
Jamaica. Volume 1. Scientific Report of the U.W.I./Kingston
Harbour Research Project, 142 pp.
Wade, B.A., Antonio, L., Mahon, R., 1972. Increasing organic pollu-
tion in Kingston Harbour. Mar. Poll. Bull. 3, 106–110.
White, A.W., 1981. Marine zooplankton can accumulate and retain
dinoflagellate toxins and cause fish kills. Limnol. Oceanogr. 26,
103–109.
White, A.W., 1984. Paralytic shellfish toxins and finfish. In: Ragelis,
E.P. (Ed.), Seafood Toxins, ACS Symposium Series No. 262, ACS
Washington, DC, pp. 171–180.
Yoshida, M., Ogata, T., Van, T.C., Matsuoka, K., Fukuyo, Y., Hoi,
N.C., Kodama, M., 2000. The first finding of toxic dinoflagellate
Alexandrium minutum in Vietnam. Fish. Sci. 66, 177–179.
Yuki, K., 1994. First report of Alexandrium minutum Halim (Dino-
phyceae) from Japan. Jpn. J. Phycol. 42, 425–430.





























