Embed Size (px)
Citation preview

Ibis
(2005),
147
, 772–782
© 2005 British Ornithologists’ Union
Blackwell Publishing, Ltd.
The incidence of the parasitic disease trichomoniasis and its treatment in reintroduced and wild Pink Pigeons
Columba mayeri
K. J. SWINNERTON,
1,2,3,4
* A. G. GREENWOOD,
5
R. E. CHAPMAN
3
& C. G. JONES
1,3
1
Durrell Wildlife Conservation Trust, Les Augrès Manor, Trinity, Jersey JE3 5BP, Channel Islands, UK
2
Wildlife Preservation Trust Canada, 120 King Street, Guelph, Ontario, N1E 4PS, Canada
3
Mauritian Wildlife Foundation, Grannum Road, Vacoas, Mauritius
4
Durrell Institute of Conservation and Ecology, University of Kent, Canterbury, Kent CT2 7NS, UK
5
International Zoo Veterinary Group, Keighley Business Centre, South Street, Keighley, W. Yorkshire BD21 1AG, UK
The recovery programme for the endangered Pink Pigeon
Columba mayeri
Prévost, 1843of Mauritius has involved intensive management to promote rapid population growth. Asthe Pink Pigeon population has increased in size and distribution, disease problems haveemerged that have affected wild and reintroduced birds. Trichomoniasis first appeared inthe remnant wild population in 1992, and became established in a reintroduced subpopu-lation on the offshore island of Ile aux Aigrettes. We aimed to control the disease by treatingindividuals and, on Ile aux Aigrettes, by routinely treating the population with medicateddrinking water. On Ile aux Aigrettes, 49% of squabs were clinically diseased and only 27%of all squabs survived without treatment. Treatment of squabs that were not clinically dis-eased improved survival from 36 to 82%, suggesting that subclinical infections also affectedsurvival. Subclinical infections were demonstrated by microscopy in 22% of Pink Pigeonsand 17% of exotic doves on Ile aux Aigrettes. Clinical disease was observed in 4.5% of adultsand 3.1% of juveniles, but more juveniles than adults died from the disease. It is likely thatTrichomoniasis and other diseases will continue to affect the Pink Pigeon’s recovery and thespecies may require long-term management to counteract the effects of disease and otherlimiting factors.
Pathogens are increasingly being implicated in thedecline or extinction of wild populations (Cooper1989, Daszak
et al
. 2000, Deem
et al
. 2001, Dobson& Foufopoulos 2001, Friend
et al
. 2001). Recentexamples include the spread of West Nile virus in wildand captive avian populations and the decline intropical frogs associated with cutaneous chytridio-mycosis (Berger
et al
. 1998, Steele
et al
. 2000). Theincreasing disease risk to wildlife may be associatedwith changes in human activity patterns causingchanges in the dynamics of host and vector popula-tions. These changes include increasing proximity ofwildlife to humans and domestic animals, habitatand environmental degradation, and the transfer ofspecies and vectors around the globe. Isolated island
populations are particularly vulnerable, in particularfrom generalist pathogens spread by common intro-duced hosts (Begon & Bowers 1995), and this hasbeen well documented in the Hawaiian avifauna(Warner 1968, van Riper
et al
. 1986). Endangeredspecies may be susceptible to disease as they areprobably also suffering from other environmentaland physiological stresses, and local epidemics canthreaten to wipe out small populations (Thorne &Williams 1988).
There is growing concern that disease may impedeefforts to conserve endangered species (Scott 1988,Cohn 1991, Viggers
et al
. 1993, Wolff & Seal 1993,Ballou 1999). Self-sustaining wild populations maybe difficult to achieve when reservoirs of pathogensare maintained by large populations of common hostspecies. In the Black-footed Ferret
Mustela nigripes
,plague and distemper, which nearly caused thespecies’ extinction, are prevalent in Prairie Dogs
*Corresponding author. Present address: Maui Forest BirdRecovery Project, 2465 Olinda Road, Makawao, HI 96768, USA.Email: [email protected]

© 2005 British Ornithologists’ Union,
Ibis
,
147
, 772–782
Trichomoniasis and Pink Pigeons
773
Cynomys
spp. and Coyotes
Canis latrans
occupyingmuch of the remaining Ferret habitat, and threatenthe successful establishment of reintroducedpopulations (Thorne & Williams 1988). Similarly, anincrease in rabies outbreaks associated with increas-ing domestic dog populations threatens many Africancarnivores, including the endangered Ethiopian Wolf
Canis simiensis
(Laurenson
et al
. 1998).In endangered-species recovery programmes, the
risk of introducing new pathogens to wild populations,through reintroduction and translocation, is a majorconcern (Woodford & Rossiter 1994, Cunningham1996). Of equal concern is the risk to reintroducedanimals from pathogens maintained in wild popula-tions, to which they may have little resistance. Inmost cases, captive species are maintained in adifferent epidemiological environment and maylack, or have lost, immunity to pathogens encounteredat the reintroduction site. Even translocations overshort distances can result in disease problems owingto often highly localized requirements of pathogensand vectors (Woodford & Rossiter 1994). Bothendemic and introduced diseases, which may evenhave been involved in the species’ original decline,will continue to operate on reintroduced populationsand may affect the success of a recovery programme.
In the conservation programmes for the endemicbirds of Mauritius, disease research and managementhave been integral to the successful recovery of thePink Pigeon
Columba mayeri
and Echo Parakeet
Psittacula eques
(Greenwood 1996). During the PinkPigeon reintroduction programme, three diseasesbecame apparent as the free-living population increased:trichomoniasis, leucocytozoonosis and avian pox(Wheler
et al
. 1996, Peirce
et al
. 1997, Swinnerton2001). In this paper we investigate the incidence ofone disease, trichomoniasis, observed in reintroducedand wild Pink Pigeons. We study cases of the diseasein Pink Pigeon adults, juveniles and squabs anddemonstrate the effects of treatment for the diseaseon squab survival. We compare juvenile and adultsurvival at mainland subpopulations with birds onthe offshore island of Ile aux Aigrettes, where thedisease is more established. We demonstrate, by micro-scopy, subclinical infections in Pink Pigeons andexotic columbids, and discuss the implications of thedisease for the species’ recovery and long-term survival.
Trichomoniasis
Trichomoniasis is a common protozoan parasiticdisease primarily affecting pigeons and turkeys,
although infections have been reported in otheravian groups including raptors and parrots, and theparasite
Trichomonas gallinae
has been recorded froma range of pigeon species (Stabler 1954, Murton 1965,Rowan 1983, Tudor 1991). The parasite primarilyinfects the upper digestive tract, causing necroticulceration, and may invade the circulatory systemand other tissues. Mouth lesions prevent feeding,and death occurs from emaciation and tissue necrosis(Stabler 1954, Charlton
et al
. 1991, Narcisi
et al
.1991). Transmission is either directly via crop milkand regurgitated food, or indirectly through contam-inated water and food. The parasite was probablyintroduced to Mauritius with exotic doves, theMadagascar Turtle-Dove
Streptopelia picturata
, ZebraDove
Geopelia striata
, Spotted Dove
Streptopeliachinensis
and the Feral Pigeon
Columba livia
, in the18th and 19th centuries.
The Pink Pigeon
The Pink Pigeon is an endangered endemic bird fromthe island of Mauritius in the Indian Ocean. It hasbeen rare for more than a century and numberedfewer than 20 birds in 1975 (see review in Jones1987). A conservation programme for the speciescomprises captive breeding, reintroduction, trans-location and management of wild populations. Since1987, 305 birds have been reintroduced and trans-located into native forest (Jones
et al
. 1988, 1992,1999), and in 2003, the free-living population wasestimated at 350 birds in five subpopulations. Therehave been several investigations of infectious disease,developmental deformities and causes of mortalityin captive adult Pink Pigeons and squabs (Flach1984, Cooper
et al
. 1987, Jones
et al
. 1989, Flach &Cooper 1991, Spencer 1993). Trichomoniasis wasfirst recorded in Pink Pigeons in a captive squabbeing foster-reared by Madagascar Turtle-Doves(McKelvey 1976), and later in a population introducedto Pamplemousses Botanical Gardens in Mauritius(Jones
et al
. 1989).
METHODS
Between 1987 and 1998, we recorded cases oftrichomoniasis in Pink Pigeon adults, juveniles andsquabs. Birds were monitored in four discretesubpopulations; the remnant wild subpopulation atPigeon Wood and two reintroduced subpopulationson the mainland, and one reintroduced subpopulationon the predator-free offshore island of Ile aux

774
K. J. Swinnerton
et al.
© 2005 British Ornithologists’ Union,
Ibis
,
147
, 772–782
Aigrettes (Fig. 1). The disease was studied mainly inbirds on Ile aux Aigrettes because (1) there weremany more cases, (2) squabs could be monitoredclosely as nests were easily accessible, (3) squabsand fledged birds could be treated routinely for thedisease, and (4) nest failure from mammalian pred-ators, which might mask the presence of disease, waseliminated. All subpopulations used supplementaryfeeding stations and, on Ile aux Aigrettes, birds usedartificial watering stations due to the lack of freshwater on the island. Milton™ Sterilizing Fluid wasadded to the drinking water (10 p.p.m., whichdissipates to 1 p.p.m), which we had demonstratedreduced the persistence of
T. gallinae
in the water(Howes unpubl. data).
Detection of Trichomoniasis in Pink Pigeons
Trichomoniasis is the manifestation of the diseaseand is not synonymous with infection by
T. gallinae
in birds. Disease here means a clinically abnormalstate resulting from infection that affects physiologicalfunction or performance. In Pink Pigeons, clinical signswere observed and recorded as evidence of disease.The first sign was usually stringy mucus in the mouthand a putrid smell from the gape. As the disease
progressed, yellowish cheesy nodules appeared onthe tongue and mucous membranes in the mouth,which extended into the pharynx and oesophagus.Tissues around the throat, eyes, nares and sinusesbecame swollen, and the bird lost weight. In fledgedbirds, lethargy and wet stained feathers around thegape were recorded as an early indication of sickness.Twisted or over-extended mandibles often indicatedrecovery from previous disease, but could alsoindicate a previous avian pox infection.
Subclinical infections in birds not showing signs ofdisease were detected by direct wet-mount micro-scopic examination for the parasite from crop swabs.Crop swabs were taken from 41 randomly selectedPink Pigeons and 121 exotic doves that did not showclinical signs of the disease. Crop swabs from fivePink Pigeon squabs were also examined microscopi-cally to investigate parasite load. Four individualswere sampled once, and the fifth individual wassampled three times. A McMaster chamber wasused to count individual flagellates in a wet-mountpreparation using lactated Ringer’s solution as amedium with a drop of glycerine added to reduceflagellate movement. The mean number of flagel-lates in a 0.25-mL aliquot was determined from sixsamples of each aliquot, and recorded as a meannumber per millilitre.
Treatment
Any Pink Pigeon with a suspected or confirmedinfection with
T. gallinae
was treated with the anti-protozoal carnidazole (Spartrix™). Adult birds andjuveniles were administered a single dose of 10 mg(one tablet). On Ile aux Aigrettes, young squabs(
≤
18 days old) were administered half of one 10-mgtablet (approximately 5 mg) on alternate days for aslong as signs of infection persisted. Older squabs(> 18 days old) were administered 10 mg with arepeat treatment every 2–4 days until fledged.Twenty-one squabs from Ile aux Aigrettes wereremoved to captivity because of severe disease and/or parental neglect. At mainland sites, few squabswere individually treated because of difficult nestaccess.
In July 1994, all reintroduced Pink Pigeons (22birds) on Ile aux Aigrettes were re-trapped and indi-vidually treated with carnidazole, following the para-site’s detection in a crop swab from one bird. The Ileaux Aigrettes subpopulation was further treatedbetween once and four times a year from November1994 to January 1999, with ronidazole (Ronitrol™)
Figure 1. The island of Mauritius showing the four sub-populations of free-living Pink Pigeons sampled for Trichomonasgallinae. The shaded area represents the Black River GorgesNational Park. Most reintroduced Pink Pigeons were bred at theGovernment captive-breeding centre in Black River village.

© 2005 British Ornithologists’ Union,
Ibis
,
147
, 772–782
Trichomoniasis and Pink Pigeons
775
or dimetridazole (Harkanker™) administered viathe drinking water. However, only a proportion ofbirds drank from the treated water source andattendance was higher during hot, dry weather thanwhen it rained. During the October 1997 treatment,33/49 (67%) Pink Pigeons were observed drinkingfrom the treated water source over the 5-day treat-ment during 9 h of observation per day. In October1998, a total of 39/55 (66%) birds drank from thetreated water source during 9 h of observation perday over 2 days of treatment. Daily attendance ofbirds that used the stations also varied. In October1998, 38 birds were observed drinking on day 1(11 h of observation) and only 19 birds on day 2 (9 hof observation) including one bird that was notobserved drinking on day 1. Variation in attendancewas also observed in a control experiment usinguntreated water during the 2 days prior to the Octo-ber 1998 treatment; a total of 26 birds drank on day1 and only six on day 2 including one bird that wasnot observed drinking on day 1. Effective treatmentrequires that the birds drink from the medicatedwater at least once a day.
Survival analyses
Studbook data for all reintroduced captive-bredbirds have been maintained since 1987, as they havefor the remaining wild population and for the wild-fledged progeny of reintroduced birds since 1989.Fledging age was standardized for wild-hatched birdsto 23 days old (mean age at fledging, Swinnerton2001) and for captive-bred reintroduced birds to30 days old (the age at which they were removedfrom the parents and were thus largely independent).A juvenile was defined as fledged but
≤
150 days old(Jones 1995) and an adult was defined as > 150 daysold. Birds were ringed routinely as part of the recov-ery programme, and all fledged birds were identifiedby a unique number and colour-ring combination. Asquab was a young Pink Pigeon in the nest untilfledged. When two squabs were present, the featherson the forehead or on one wing were marked with acoloured pen until ringed. At about 6–8 days old,each squab was ringed with a unique numbered ringand colour-ring combination. Numbered rings wereassigned sequentially in accordance with the studbook.On Ile aux Aigrettes, dates of squab hatch, squabdeath, squab removal and fledging were recorded.
On Ile aux Aigrettes, survival analyses wereperformed for squabs from hatch to fledge underdifferent disease treatment regimes. A chi-squared
contingency table was used to test for seasonaldifferences in disease incidence and for the effect oftreatment on the number of squabs that fledged.Survival analyses were also performed for fledgedbirds, and comparisons made between the survival offledged birds on Ile aux Aigrettes and at mainlandsites. Survival of individuals was monitored fromfledging to death, and birds alive on 31 December1998 were treated as censored data (Collett 1994).
We investigated survival using Kaplan–Meiernon-parametric estimates of the survivor function(Kaplan & Meier 1958, Collett 1994). We used Cox’sproportional hazards regression model (Cox 1972)with disease status (diseased/not diseased) enteredas a covariate. The significance of terms entered inthe model was tested by Wald tests, which yield anapproximate chi-squared statistic (Collett 1994).Covariates are assumed to induce proportional shiftsof the arbitrary hazard function. In the Cox model,the data were stratified owing to the different survivalprobabilities between subpopulations. Stratificationallowed the survival pattern to vary for cases withdifferent values while keeping the coefficientsgoverning the hazard shifts common across all strata.
RESULTS
Trichomoniasis was first detected in the wild in July1992, in a squab in the remnant wild population atPigeon Wood. At mainland sites, further cases werenot recorded until 1994, in an adult, and then in1996, in a squab, in the Brise Fer reintroducedsubpopulation. In 1997 and 1998, 16 cases wererecorded in adults and juveniles at mainland sites, ofwhich eight survived. The incidence of trichomo-niasis was highest on Ile aux Aigrettes, where it wasfirst confirmed in October 1994 in the recentlyreintroduced population. These first cases were in afledgling and in a brood of two squabs, both rearedby reintroduced parents. The disease subsequentlybecame established in this subpopulation.
Trichomoniasis in squabs on Ile aux Aigrettes
Trichomoniasis was recorded in 49% of squabs(Table 1). There were no seasonal differences in theincidence of trichomoniasis cases (January–April:hot, wet; May–August: cool, dry; September–December: hot, dry) (
χ
2
= 0.89,
df
= 2,
P
> 0.05,
n
= 222). Disease incidence in squabs in singlebroods and in broods of two was similar. Squabs first
776
K. J. Swinnerton
et al.
© 2005 British Ornithologists’ Union,
Ibis
,
147
, 772–782
developed signs of disease between 1 and 24 days old(mean
±
sd = 8
±
5 days old,
n
= 108). Some squabsthat died were probably infected but died before anyobvious clinical signs developed. Of the 48 squabs inwhich disease status was not recorded, 21 (44%)died in the first 2 days. Diseased squabs that diedbefore fledging died between 2 and 25 days old(mean
±
sd = 11
±
6 days,
n
= 61).On Ile aux Aigrettes, disease, inanition and injury,
and interactions between them, were the main causesof squab mortality and comprised 39% of squab deaths(
n
= 159) (Swinnerton 2001). Injuries includedpuncture wounds from unsuitable nest materialor were incurred through poor parental care (e.g.excessive feather plucking by a parent, or parentsfighting at the nest), or by falling from the nest. Dis-placement from the nest usually led to the death ofyoung squabs (either because of injuries or becausethe parents abandoned them), but older squabs thatsurvived the fall continued to be fed by the parentson the ground. Nest displacement was caused bypoor nest structure, poor parental behaviour, poorsquab health, bad weather, fighting at the nest withinor between pairs, and displacement by a secondsquab. In some cases a combination of factors (e.g.poor parental care, inanition and bad weather) led tomortality. Unknown causes of mortality comprised13% of squab deaths and an additional 30% ofsquabs that died disappeared from the nest, 55%(26/47) of which were between hatch and 3 daysold.
On Ile aux Aigrettes, trichomoniasis was widespreadin the offspring of breeding pairs: 30/35 males and26/33 females reared squabs which became diseased.In the remaining birds, few squabs were reared andinfection was not confirmed. Of the parent birds that
reared diseased squabs, the proportion of squabs thatbecame diseased differed between pairs. For exam-ple, only two out of 19 squabs monitored fromone pair became diseased. By contrast, 16 out of 18squabs from one female and all of six squabs rearedby one male became diseased.
The effect of treatment on squab survival
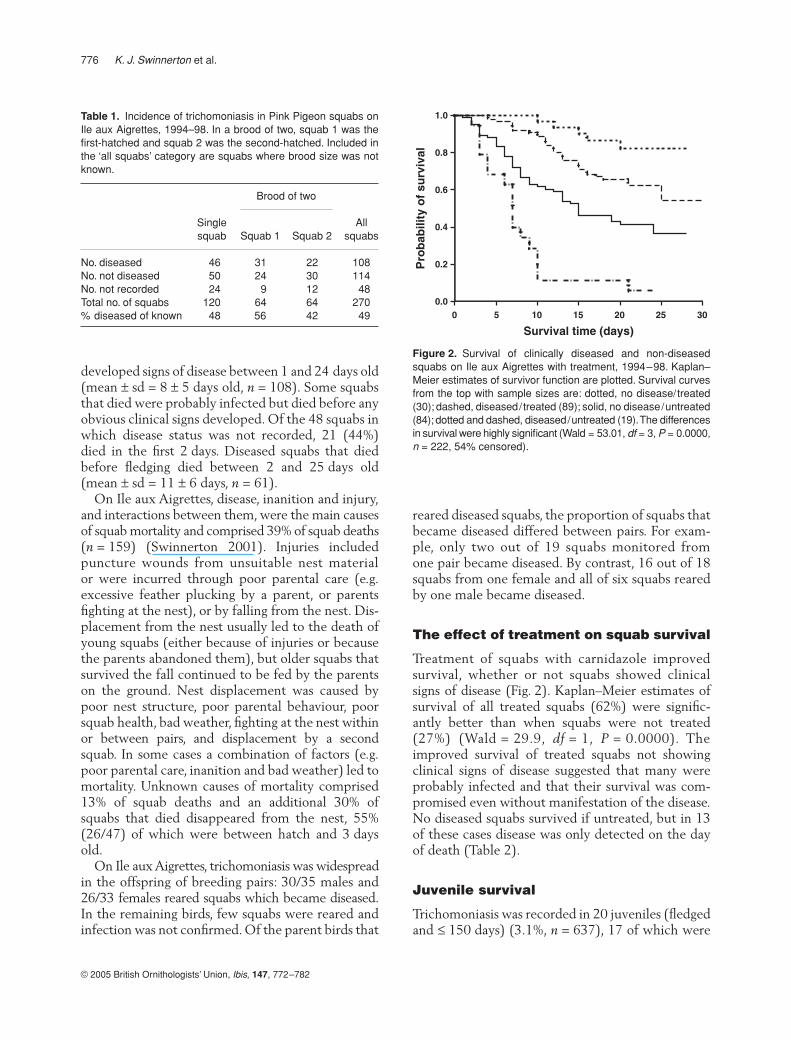
Treatment of squabs with carnidazole improvedsurvival, whether or not squabs showed clinicalsigns of disease (Fig. 2). Kaplan–Meier estimates ofsurvival of all treated squabs (62%) were signific-antly better than when squabs were not treated(27%) (Wald = 29.9,
df
= 1,
P
= 0.0000). Theimproved survival of treated squabs not showingclinical signs of disease suggested that many wereprobably infected and that their survival was com-promised even without manifestation of the disease.No diseased squabs survived if untreated, but in 13of these cases disease was only detected on the dayof death (Table 2).
Juvenile survival
Trichomoniasis was recorded in 20 juveniles (fledgedand
≤
150 days) (3.1%,
n
= 637), 17 of which were
Table 1. Incidence of trichomoniasis in Pink Pigeon squabs onIle aux Aigrettes, 1994–98. In a brood of two, squab 1 was thefirst-hatched and squab 2 was the second-hatched. Included inthe ‘all squabs’ category are squabs where brood size was notknown.
Brood of two
Single squab Squab 1 Squab 2
All squabs
No. diseased 46 31 22 108No. not diseased 50 24 30 114No. not recorded 24 9 12 48Total no. of squabs 120 64 64 270% diseased of known 48 56 42 49
Figure 2. Survival of clinically diseased and non-diseasedsquabs on Ile aux Aigrettes with treatment, 1994–98. Kaplan–Meier estimates of survivor function are plotted. Survival curvesfrom the top with sample sizes are: dotted, no disease/treated(30); dashed, diseased/treated (89); solid, no disease/untreated(84); dotted and dashed, diseased/untreated (19). The differencesin survival were highly significant (Wald = 53.01, df = 3, P = 0.0000,n = 222, 54% censored).
© 2005 British Ornithologists’ Union,
Ibis
,
147
, 772–782
Trichomoniasis and Pink Pigeons
777
fledglings (mean age
±
sd = 32
±
7 days old). Thedisease probably caused death in 15 juveniles, one ofwhich died 72 days after clinical diagnosis of thedisease (Table 3). The one diseased juvenile thatsurvived without treatment was the oldest juvenileto develop the disease, at 147 days old. On Ile auxAigrettes, undetected cases and subclinical infectionwith the parasite probably contributed to the gener-ally poor survival observed in juveniles at this site.Survival of wild-fledged juveniles was significantlypoorer on Ile aux Aigrettes (46%,
n
= 126) than atmainland sites (85%,
n
= 315) where the disease wasless established (Wald = 24.5,
df
= 1,
P
= 0.0000).Treatment of squabs did not significantly affect
their survival as juveniles from fledging to 150 daysold (Wald = 1.55,
df
= 1,
P
= 0.21). Of 95 squabsmonitored post-fledging on Ile aux Aigrettes, 46(48%) died at less than 70 days old, 35 of which weretreated and 11 of which were not treated as squabs.
Adult survival
Trichomoniasis was recorded in 23 adults(> 150 days old) (4.5%,
n
= 508), 14 of which werein their first year (151–365 days old). Ten out of 12treated adults survived (but one of the two that died
survived for 109 days post-treatment) but only 5/11untreated adults survived (Table 3). Adults that diedof the disease died between 0.5 and 2.6 years old(mean
±
sd = 1.4
±
0.7 years), and birds that survivedthe disease probably became carriers. On Ile auxAigrettes, survival of wild-fledged adults in their firstyear (82%,
n
= 52) was similar to survival in theirsecond and third years (83%,
n
= 37), and wassimilar to survival at mainland sites (first year = 88%,
n
= 236; second and third years = 74%,
n = 200).On Ile aux Aigrettes, both adults and juvenilesprobably had similar exposure to the pathogen, butjuveniles were more compromised by the disease.
Group treatment of juveniles and adults
On Ile aux Aigrettes, group treatments of juvenilesand adults with anti-protozoals in the drinking waterhad no long-term effect of reducing disease in thissubpopulation, and birds quickly became re-infectedonce treatment stopped. Even during treatments,the effect was variable. For example, between Juneand December 1996 (three group treatments),diseased squabs were recorded in 10/23 nests, andthree diseased juveniles were reported. BetweenOctober and December 1998 (monthly treatments),
Table 2. Disease status of Pink Pigeon squabs on Ile aux Aigrettes and treatment regime, 1994–98. Kaplan–Meier estimates of squabsurvival to fledge are shown. The number of squabs that fledged when treated was greater than expected (χ2 = 37.3, df = 3, P < 0.001).
No. of squabs
No. fledged
No.died
No. removed
% survival to fledge
No disease/treated 30 24 5 1 82.4Diseased/treated 89 47 34 8 54.2No disease/untreated 84 28 45 11 36.7Diseased/untreated 19 0 18 1 0.0All squabs 222 99 102 21 –
Table 3. Number of juvenile (fledged and ≤ 150 days old) and adult Pink Pigeons treated for trichomoniasis and the number of birds thatsurvived, 1992–98.
Juveniles Adults All birds
SiteTreated/Survived
Untreated/Survived
Treated/Survived
Untreated/Survived
Treated/Survived
Untreated/Survived
Brise Fer 0/0 0/0 2/2 3/2 2/2 3/2Pigeon Wood 1/1 3/0 0/0 1/0 1/1 4/0Bel Ombre 0/0 2/1 1/0 4/3 1/0 6/4Ile aux Aigrettes 6/3 8/0 9/8 3/0 15/11 11/0Total 7/4 13/1 12/10 11/5 19/14 24/6
778 K. J. Swinnerton et al.
© 2005 British Ornithologists’ Union, Ibis, 147, 772–782
diseased squabs were recorded in 5/12 nests, andthree diseased juveniles were reported.
Microscopic detection of the parasite
The parasite was detected microscopically in wet-mountpreparations of crop swabs taken from free-livingPink Pigeons and exotic doves that did not showclinical signs of disease (Table 4). The prevalence ofinfection in Pink Pigeons (22%) was similar to theoverall prevalence in exotic doves (17%).
Examination of crop swabs from five Pink Pigeonsquabs also demonstrated that squabs showing noclinical signs of disease were infected with the para-site to a varied degree. A worsening of clinical signs,and of the disease, was associated with an increase inthe number of flagellates. In squab A285, flagellatenumbers increased rapidly over several days despitetreatment (Table 5).
DISCUSSION
The first case of trichomoniasis in wild Pink Pigeonswas in 1992 in the remnant wild population at Pigeon
Wood. Between 1992 and 1998, 153 cases wererecorded in free-living Pink Pigeons, 108 of whichwere in squabs on Ile aux Aigrettes. On Ile auxAigrettes, the disease was confirmed only 7 monthsafter the first birds were reintroduced and only2 months after the first squabs hatched. Individualtreatment of squabs with carnidazole improvedsurvival to fledging, whether or not squabs showedsigns of the disease. On Ile aux Aigrettes, survival ofjuvenile Pink Pigeons (46%) was very poor comparedwith that of juveniles at mainland sites (85%); thiswas probably in part due to undetected or subclinicalinfections with the parasite. Juveniles were susceptibleto the disease whereas some adults could withstandinfection and some probably became carriers.Reintroduced Pink Pigeons must have initiallyacquired their infections from exotic doves, whichwere demonstrated to be infected without signs ofdisease.
The incidence of clinical disease given here isprobably underestimated. Many cases in fledgedbirds were probably missed owing to the difficulty ofobserving clinical signs in the field. The prevalence ofclinical disease in squabs at mainland sites is poorlyknown owing to problems of nest access, but periodicoutbreaks have been recorded, such as in July 2000when 10 of 24 squabs that died in the Brise Fersubpopulation died from trichomoniasis. Subclinicalinfections were detected microscopically andprobably affect survival. In 2001, trial screeningsfor T. gallinae using InPouch™ cultures (Cover et al.1994) of crop swabs from free-living Pink Pigeonsin the Brise Fer mainland site showed that 10%(n = 30) of clinically non-diseased adults were, infact, infected. Subsequent research has shown thatcultures are a significantly more sensitive test forT. gallinae infection than wet-mount preparations
Table 4. Detection of Trichomonas gallinae in crop swabs fromPink Pigeons and exotic doves on Ile aux Aigrettes, 1995–98.The Pink Pigeon sample included 4/6 infected squabs.
No. sampled
No.+ve
% +ve
Pink Pigeon 41 9 22Madagascar Turtle-dove 109 17 16Spotted Dove 3 1 33Zebra Dove 9 3 33Total 162 30 19
Table 5. Number of Trichomonas gallinae flagellates counted in 0.25-mL wet-mount preparations of crop swabs taken from five PinkPigeon squabs from Ile aux Aigrettes in 1995.
Ring no.
Age(days)
Counts in 0.25 mLMean (mL) Clinical signsC1 C2 C3 C4 C5 C6
A281 22 46 29 37 38 28 31 139 NoneA282 16 168 170 179 152 121 135 617 Cheesy caseous matter in pharynxA283 11 1 0 0 0 0 0 1 NoneA284 20 37 33 44 46 59 55 183 None, poor growthA285 17 0 0 0 0 0 0 0 None, poor growth, treatedA285 19 15 108 90 60 72 91 291 Cheesy caseous matter in pharynxA285 20 95 171 160 158 166 143 595 Caseous matter increased

© 2005 British Ornithologists’ Union, Ibis, 147, 772–782
Trichomoniasis and Pink Pigeons 779
and indicate a much higher incidence of subclinicalinfections (N. Bunbury pers. comm.).
Clinical disease is usually only found in youngbirds, and adults probably do not die from the diseasealone. Incidence rates of between 14 and 65% havebeen reported in wild and feral columbids (Stabler1951, 1954, Johnston & Janiga 1995, Ostrand et al.1995, McKeon et al. 1997) and squabs may sufferup to 75% mortality in an epidemic (Stabler 1954).Virulent strains can cause high mortality; in California,a lethal strain of T. gallinae was responsible for thedeaths of at least 16 000 native Band-tailed PigeonsColumba fasciata in 1988 (Conti 1993). An avirulentinfection, or the survival of a virulent one, may provideprotective immunity against further virulent strains(Stabler 1954).
Mortality may depend on the immunocompet-ence of the bird, in turn affected by other factorssuch as age, concurrent disease, food shortages, bodycondition and genetic variation: all factors whichalso affect Pink Pigeon survival. Infection with theparasite may lower resistance to other disease andthere may be interactions between pathogens(Richie 1988). Infection with Leucocytozoonmarchouxi and avian pox also cause mortality inPink Pigeons (Wheler et al. 1996, Peirce et al. 1997,Swinnerton 2001 and unpubl. data, Swinnertonet al. 2005). Eight Pink Pigeons suffering fromtrichomoniasis had a combined infection withL. marchouxi, nine birds suffered from trichomonia-sis and avian pox, and one bird suffered from allthree pathogens.
Squab survival on Ile aux Aigrettes
On Ile aux Aigrettes, trichomoniasis in squabsseverely reduced fledging success, which would havebeen even lower without treatment. Squab survivalwas probably influenced by the age of the squabwhen infected and treated, the severity of the dis-ease, the virulence of the strain and differences inparental care. About half of the diseased squabsdeveloped a severe infection. Squabs typicallyshowed either mouth lesions or inflammation of thesinuses, tissue around the eyes and head, with mouthlesions scarce or absent, which may have reflecteddifferent parasite strains. Stabler (1954) reportedthat certain strains have a predilection for certaintissues.
First-hatched squabs are more likely to survivethan second-hatched squabs (broods vary betweenone and two squabs, Swinnerton 2001) which, if
diseased, were further compromised by competitionfor food with the first-hatched squab. In eight broodsof two in which both squabs were treated, one diedof trichomoniasis while the other fledged. Overallsquab condition was improved by treatment, whichmay have reduced mortality from other factors. Parentbirds that produced diseased squabs may themselvesnot always retain infections. Infected parents re-infected the squab post-fledging until its completeindependence (up to 90 days old). Re-infection ratesand the number of parasites transmitted could partlyaccount for differences in the severity of infectionand survival between individual squabs.
Implications of disease in Pink Pigeon decline and recovery
It is probable that disease was a contributing factorto the historical decline of the Pink Pigeon, parti-cularly in lowland habitats where exotic doves aremore common, but at the beginning of the recoveryprogramme its significance was not understoodbecause of the very small population size and othermore obvious limiting factors, such as predation,masking disease effects. Reintroduced Pink Pigeons,free of infection, must have initially acquired theirinfections from exotic doves but as the populationhas increased in size and distribution, Pink Pigeonsthemselves have increasingly become an importanthost. The Pink Pigeon’s range expansion and carryingcapacity on Mauritius may be influenced by the pop-ulation dynamics and distribution of reservoir hostsfor T. gallinae and other pathogens, and Pink Pigeonpopulations may be less successful in some habitats.The severity of the disease on Ile aux Aigrettes, com-pared with mainland sites, is probably due to higherdensities of exotic doves on the island (and hencea higher parasite load) and because fresh water(through which parasites are probably transferred) isscarce and all birds on the island drink from communalsources. Studies on the role of exotic doves as reservoirhosts are under way (N. Bunbury pers. comm.). Theintroduction to Mauritius of new hosts and patho-gens could be a serious threat to the Pink Pigeon,such as the recently introduced Laughing DoveStreptopelia senegalensis, which is now established.Preventing new introductions of hosts and vectors isimportant for the species’ recovery.
The Pink Pigeon recovery programme involvesintensive management to encourage population growth,including predator control and eradication, supple-mental feeding and habitat restoration. Supplemental

780 K. J. Swinnerton et al.
© 2005 British Ornithologists’ Union, Ibis, 147, 772–782
feeding of Pink Pigeons has contributed most toraising survival rates and improving productivity.However, feeding stations can also be sources ofinfection if they are poorly maintained (Wilson &Macdonald 1967, Conti 1993, Greenwood 1996,Hartup et al. 2001), and artificial watering stationswere partly responsible for higher transmission ratesof T. gallinae on Ile aux Aigrettes.
Treatment of birds for trichomoniasis was limitedin its success. Treatment of individual squabs onlyreduced mortality in the short term and was labourintensive. Group treatments with medicated waterhad little effect on controlling the disease becauseattendance at the watering stations and acceptabilityof the medication varied. Dimetridazole (Harkanker™)appeared less acceptable than ronidazole (Ronitrol™),and attendance was higher during hot, dry days thanduring days of showers or rain when birds drankfrom other water sources. For example, in October1998 severe rainstorms during the last 3 days oftreatment probably reduced the effectiveness of anytreatment received in the first 2 days because birdsdid not receive a full course of medication. Variationin daily attendance also occurs at feeding stationsand is influenced by age and breeding status(Mauritian Wildlife Foundation unpubl. data). Inaddition, post-treatment re-infection occurred quicklybecause not all birds received the full course of treat-ment. Future management of trichomoniasis shouldfocus on reducing the sources of infection (communaldrinking water, exotic birds) and targeting factorsthat reduce resistance to disease (e.g. other pathogens,inanition from food shortages). Disease managementmay be needed in perpetuity for some subpopulationsin high-risk habitats, such as Ile aux Aigrettes.
The impacts of disease on a population areunlikely to be wholly additive to other causes ofmortality and contributing causes are often apparent,for example individuals weakened by starvation maybe vulnerable to disease. The effects of disease mayalso be compensatory to other causes of mortality sothat, in the absence of premature deaths from disease,a similar number of premature deaths would occurfrom other causes (Newton 1998). In Pink Pigeons,the effects of disease on predator-free Ile aux Aigretteshas to some degree replaced the effects of predationat mainland sites, and reproductive success is similarat both sites (Swinnerton 2001). Conclusive infor-mation about the effects of disease at the populationlevel is therefore difficult to obtain. In small popula-tions the effect of pathogens may be masked byother factors. Disease problems in the Pink Pigeon
became clearer once other limiting factors (food andpredators) were controlled and a larger populationsize was achieved.
The factors that caused the Pink Pigeon’s declinecontinue to operate on the free-living population,including habitat loss and degradation, food shortagesand introduced predators. This study has demonstratedthat disease can be added to the suite of threats, andwas probably involved in the species’ originaldecline. Trichomoniasis was recorded in all foursubpopulations of free-living Pink Pigeons, and canbe pathogenic. Pink Pigeons are probably susceptibleto disease because of other stresses, such as environ-mental factors and inbreeding, but management offree-living populations may have reduced the impactof disease. Further research is under way to investigatethe effects of T. gallinae and Leucocytozoon infectionson the productivity and survival of free-living PinkPigeons (N. Bunbury pers. comm.). Understandingthese effects will be important for developing man-agement strategies to ensure the species’ long-termsurvival.
This work was supported by the Durrell Wildlife ConservationTrust and Wildlife Preservation Trust Canada as part of the PinkPigeon conservation programme managed by the Mauritian Wild-life Foundation. The programme is run jointly with the Govern-ment of Mauritius, and we would like to thank the NationalParks and Conservation Service and its Director, Y. Mungroo, inparticular for their financial support of the disease research andveterinary work. We are grateful to R.W. Burn (University ofReading) for his input into the survival analyses and to Dr R. Grif-fiths (DICE), Dr D. Bell and N. Bunbury (UEA), Dr J. Reynolds,B. Coles and one anonymous reviewer for their comments. Wewould like to thank all the Mauritian Wildlife Foundation staffinvolved in Pink Pigeon conservation for their hard work, inparticular those who contributed to this research: L.A. Howes,C. Wheler, M. Nicoll, S. Johnston, B. Vadaparnaiken, S. Paul,S. Green, K. Freeman, D. Djeddour, C. Shaw and C. Cumming.
REFERENCES
Ballou, J.D. 1999. Infectious disease risk assessment in captivepropagation, reintroduction and wildlife conservation. InRoth, T.L., Swanson, W.F. & Blattman, L.K. (eds) Proceed-ings of the 7th World Conference on Breeding EndangeredSpecies: pp 77–86. Cincinnati, OH.
Begon, M. & Bowers, R.G. (eds) 1995. Beyond Host–PathogenDynamics. Ecology of Infectious Diseases in Natural Popula-tions. Cambridge: Cambridge University Press.
Berger, L., Speare, R., Daszak, P., Green, D.E., Cunningham,A.A., Goggin, C.L., Slocombe, R., Ragan, M.A., Hyatt,A.D., McDonald, K.R., Hines, H.B., Lips, K.R., Marantelli,G. & Parkes, H. 1998. Chytridiomycosis causes amphibianmortality associated with population declines in the rainforests of Australia and Central America. Proc. Natl Acad.Sci. USA 95: 9031–9036.

© 2005 British Ornithologists’ Union, Ibis, 147, 772–782
Trichomoniasis and Pink Pigeons 781
Charlton, B.R., Bickford, A.A., Cooper, G.L. & Chiu, H.W.1991. Systemic trichomoniasis in a squab operation. AvianDis. 35: 426–432.
Cohn, J.P. 1991. New focus on wildlife health: tracking andcontrolling animal disease in the wild may be important toconservation efforts. Bioscience 41: 448–450.
Collett, D. 1994. Modelling Survival Data in Medical Research.London: Chapman & Hall.
Conti, J.A. 1993. Diseases, parasites and contaminants. InBaskett, T.S., Sayre, M.W., Tomlinson, R.E. & Mirarchi, R.E.(eds) Ecology and Management of the Mourning Dove:205–224. Harrisburg: Wildlife Management Institute.
Cooper, J.E. 1989. The role of pathogens in threatened popula-tions. An historical review. In Cooper, J.E. (ed.) Disease andThreatened Birds: 51–68. Cambridge: ICBP TechnicalPublication no. 10.
Cooper, J.E., Needham, J.R., Applebee, K. & Jones, C.G.1987. Clinical and pathological studies on the Mauritian PinkPigeon (Columba mayeri ). Ibis 130: 57–64.
Cover, A.J., Harmon, W.M. & Thomas, M.W. 1994. A newmethod for the diagnosis of Trichomonas gallinae infection byculture. J. Wildl. Dis. 30: 457–459.
Cox, D.R. 1972. Regression models and life tables (with discus-sion). J. R. Stat. Soc. B 74: 187–220.
Cunningham, A.A. 1996. Disease risks of wildlife translocations.Conserv. Biol. 10: 349–353.
Daszak, P., Cunningham, A.A. & Hyatt, A.D. 2000. Emerginginfectious diseases of wildlife – threats to biodiversity andhuman health. Science 287: 443–449.
Deem, S.L., Karesh, W.B. & Weisman, W. 2001. Putting theoryinto practice: wildlife health in conservation. Conserv. Biol.15: 1224–1233.
Dobson, A. & Foufopoulos, J. 2001. Emerging infectiouspathogens of wildlife. Phil. Trans. R. Soc. Lond. B 356: 1001–1012.
Flach, E.J. 1984. Observations, Clinical Examinations and Post-Mortem Examinations on Two Mayer’s Pink Pigeons, Nesoenasmayeri, with Locomotor Disorders. Unpublished Report to theJersey Wildlife Preservation Trust.
Flach, E.J. & Cooper, J.E. 1991. Clinical and pathologicalfindings in two Mauritian Pink Pigeons (Columba mayeri).Vet. Record 29: 48–51.
Friend, M., McLean, R.G. & Dein, F.J. 2001. Disease emergencein birds: challenges for the twenty-first century. Auk 118:290–303.
Greenwood, A.G. 1996. Veterinary support for in situ avianconservation programmes. Bird Conserv. Int. 6: 285–292.
Hartup, B.K., Dhondt, A.A., Sydenstricker, K.V., Hochachka, W.M.& Kollias, G.V. 2001. Host range and dynamics of mycoplasmalconjunctivitis among birds in North America. J. Wildl. Dis. 37:72–81.
Johnston, R.F. & Janiga, M. 1995. Feral Pigeons. Oxford:Oxford University Press.
Jones, C.G. 1987. The larger land-birds of Mauritius. In Diamond,A.W. (ed.) Studies of Mascarene Island Birds: 208–300.Cambridge: Cambridge University Press.
Jones, C.G. 1995. Studies on the Biology of the Pink PigeonColumba mayeri. PhD thesis, University of Wales.
Jones, C.G., Swinnerton, K., Hartley, J. & Mungroo, Y.1999. The restoration of the free-living populations of theMauritius Kestrel (Falco punctatus), Pink Pigeon (Columbamayeri ) and Echo Parakeet (Psittacula eques). In Roth, T.L.,
Swanson, W.F. & Blattman, L.K. (eds) Proceedings of the 7thWorld Conference on Breeding Endangered Species: 77–86.Cincinnati, OH.
Jones, C.G., Swinnerton, K.J., Taylor, C.J. & Mungroo, Y.1992. The release of captive-bred Pink Pigeons Columbamayeri in native forest on Mauritius. A progress report July1987–June 1992. Dodo, J. Wildl. Preserv. Trust 28: 92–125.
Jones, C.G., Todd, D.M. & Mungroo, Y. 1989. Mortality,morbidity and breeding success of the Pink Pigeon (Columba(Nesoenas) mayeri ). In Cooper, J.E. (ed.) Disease andThreatened Birds: 89–113. Cambridge: ICBP TechnicalPublication no. 10.
Jones, C.G., Todd, D.M., Swinnerton, K.J. & Mungroo, Y.1988. The release and management at liberty of captive-bredPink Pigeons Columba mayeri on Mauritius. In Dresser, E.L.,Reece, R.W & Maruska, E.J. (eds) Proceedings of the 5thWorld Conference on Breeding Endangered Species inCaptivity: 393–411. Cincinnati, OH.
Kaplan, E.L. & Meier, P. 1958. Nonparametric estimation fromincomplete observations. J. Am. Stat. 53: 457–481.
Laurenson, K., Sillero-Zubiri, C., Thompson, H., Shiefraw, F.,Thirgood, S. & Malcolm, J. 1998. Disease as a threat toendangered species: Ethiopian Wolves, domestic dogs andcanine pathogens. Anim. Conserv. 1: 273–280.
McKelvey, D.S. 1976. A preliminary study of the MauritianPink Pigeon (Nesoenas meyeri ). Mauritius Inst. Bull. 8: 145–175.
McKeon, T., Dunsmore, J. & Raidal, S.R. 1997. Trichomonasgallinae in budgerigars and columbid birds in Perth, WesternAustralia. Aust. Vet. J. 75: 652–655.
Murton, R.K. 1965. The Wood Pigeon. New Naturalist Mono-graph Series. London: Collins.
Narcisi, E.M., Sevoian, M. & Honigberg, B.M. 1991. Pathologicchanges in pigeons infected with a virulent Trichomonas gall-inae strain (Eiberg). Avian Dis. 35: 55–61.
Newton, I. 1998. Population Limitation in Birds. London: AcademicPress.
Ostrand, W.D., Bissonette, J.A. & Conover, M.R. 1995.Trichomoniasis as a factor in Mourning Dove populationdecline in Fillimore, Utah. J. Wildl. Dis. 31: 87–89.
Peirce, M.A., Greenwood, A.G. & Swinnerton, K. 1997.Pathogenicity of Leucocytozoon marchouxi in the PinkPigeon (Columba mayeri ) in Mauritius. Vet. Record 140:155–156.
Richie, T.L. 1988. Interactions between malarial parasitesinfecting the same vertebrate host. Parasitology 96: 607–639.
van Riper, C.I., van Riper, S.G., Goff, M.G. & Laird, M. 1986.The epizootiology and ecological significance of malaria inHawaiian land birds. Ecol. Monogr. 56: 327–344.
Rowan, M.K. 1983. The Doves, Parrots, Louries and Cuckoos ofSouthern Africa. Canberra: Croom-Helm.
Scott, M.E. 1988. The impact of infection and disease on animalpopulations: implications for conservation biology. Conserv.Biol. 2: 40–56.
Spencer, S. 1993. Clinical investigation of the Mauritius PinkPigeon. Anim. Techn. 44: 59–63.
Stabler, R.M. 1951. A survey of Colorado Band-tailed Pigeons,Mourning Doves and wild Common Pigeons for Trichomonasgallinae. J. Parasitol. 37: 471–472.
Stabler, R.M. 1954. Parasitological reviews. Trichomonas galli-nae: a review. Exp. Parasitol. 111: 368–402.

782 K. J. Swinnerton et al.
© 2005 British Ornithologists’ Union, Ibis, 147, 772–782
Steele, K.E., Linn, M.J., Schoepp, R.J., Komar, N., Geisbert, T.W.,Manduca, R.M., Calle, P.P., Raphael, B.L., Clippinger, T.L.,Larsen, T., Smith, J., Lanciotti, R.S., Panella, N.A. &McNamara, T.S. 2000. Pathology of fatal West Nile virusinfections in native and exotic birds during the 1999 out-break in New York City, New York. Vet. Pathol. 37: 208–224.
Swinnerton, K.J. 2001. The Ecology and Conservation of thePink Pigeon Columba mayeri on Mauritius. PhD thesis,University of Kent, Canterbury.
Swinnerton, K.J., Peirce, M.P., Greenwood, A., Chapman,R.E. & Jones, C.G. 2005. Prevalence of Leucocytozoonmarchouxi in the endangered Pink Pigeon Columba mayeri.Ibis in press.
Thorne, E.T. & Williams, E.S. 1988. Disease and endangeredspecies: the Black-footed Ferret as a recent example. Conserv.Biol. 2: 66–74.
Tudor, D.C. 1991. Pigeon Health and Disease. Iowa: Iowa StateUniversity Press.
Viggers, K.L., Lindenmeyer, D.B. & Speatt, D.M. 1993. Theimportance of disease in reintroduction programmes. Wildl.Res. 20: 697–698.
Warner, R.E. 1968. The role of introduced diseases in theextinction of the endemic Hawaiian avifauna. Condor 70:101–120.
Wheler, C., Riddell, C., Swinnerton, K., Chapman, R., Jones,C.G. & Yeung, R.L.S. 1996. Conservation and disease prob-lems of the Mauritius Pink Pigeon (Columba mayeri ). In Pro-ceedings of the Annual Conference of the Association ofAvian Vets: 241–243. Tampa, Flolida.
Wilson, J.E. & Macdonald, J.W. 1967. Salmonella infection inwild birds. Br. Vet. J. 123: 212–219.
Wolff, P.L. & Seal, U.S. 1993. Implications of infectious diseasefor captive propagation and reintroduction of threatenedspecies. J. Zoo Wildl. Med. 24: 229–230.
Woodford, M.H. & Rossiter, P.B. 1994. Disease risks associatedwith wildlife translocation projects. In Olney, P.J.S., Mace, G.M.& Feistner, A.T.C. (eds) Creative Conservation: InteractiveManagement of Wild and Captive Animals: 178–200.London: Chapman & Hall.
Received 10 July 2003; revision accepted 10 June 2005; DOI: 10.1111/j.1474-919x.2005.00466.x.




























