Embed Size (px)
Citation preview

3158 M. Humbert, G. Raposo, F! Cosson et al. Eur. J. Immunol. 1993. 23: 3158-3166
Martie Humbert., Graga Raposo.0, Pierre CossonoA, Hubert Reggio., Jean Davoust. and Jean Salamero.
Centre d’Immunologie INSERM-CNRS de Marseille-Luminy. and Laboratoire de Biologie Cellulaire de la Differentiation., Marseille
The invariant chain induces compact forms of class I1 molecules localized in late endosomal compartments*
The invariant chain (Ii) binds to newly synthesized major histocompatibility complex (MHC) class11 molecules and is targeted to an acidic compartment where it is degraded.To evaluate its role on the conformation and the subcellular distribution of murine MHC class I1 molecules we have established stable L cell transfectants expressing class I1 IAk heterodimers alone or in conjunction with p31 and p41 Ii chains. In these cells, class I1 molecules were present under three forms: ap heterodimers bearing high mannose carbohydrate moieties, and fully glycosylated aB heterodimers that are sensitive or resistant to sodium dodecyl sulfate dissociation at 20 “C. The latter class I1 molecules called compact heterodimers, were here highly induced in Ii-positive cells. Using in situ iodination of endosomal compartments, class I1 heterodimers were detected in late endosomal compartments essentially as compact forms in Ii-positive cells, and as non-compact forms in Ii-negative cells. Using confocal microscopy, IAk molecules were located in compartments distinct from early endosomes labeled with transferrin, but partially coincident with vesicles containing fluid-phase markers, and highly coincident with compartments containing large amounts of cathepsins B, D, H, and L in Ii-positive and Ii-negative cells. At the ultrastruc- tural level, class I1 molecules were mostly present in multivesicular bodies, even without Ii expression. But Ii chains were needed to induce an efficient presentation of the hen egg lysozyme antigen and were sufficient to promote a major conformational change of the late endosomal, and/or lysosomal resident, class I1 molecules. Ii molecules are presumably playing a chaperoning function favoring the association of peptides with class I1 molecules in endosomal compartments.
1 Introduction
The MHC class I1 heterodimeric molecules involved in the presentation of antigenic peptides to specific CD4+ Tcells [ l , 21 are composed of two chains of 34 kDa and 29 kDa that associate shortly after their synthesis in the rough endoplasmic reticulum (ER), with a transmembrane type I1 glycoprotein called the invariant chain (Ii) [3-51. The murine Ii chains exist under two different forms of 31 kDa
[I 118751
* This work was supported by CNRS, INSERM, ARC (Associa- tion pour la Recherche contre le Cancer) and the Ligue FranGaise Contre le Cancer (ComitC DCpartemental des Bouches du Rh6ne). We thank Leica SARL for their constant help and for their contribution to the color prints presented.
0 Present address: University of Utrecht, Department of Cell Biology, Medical School AZU, H02.314 Heidelberglaan 100, NL-3584 Utrecht, The Netherlands
A Present address: Basel Institute for Immunology, Grenzacher- straBe 487, CH-4005 Basel, Switzerland
Correspondence: Jean Davoust/Jean Salamero, Centre d’Immuno- logie INSERM-CNRS de Marseille-Luminy, Case 906, F-13288 Marseille CCdex 9. France
Abbreviations: ER: Endoplasmic reticulum Ii: Invariant chain HEL: Hen egg lysozyme NaNase: Neuraminidase Endo H: f5-Endoglycosidase H
Key words: Antigen presentation / Confocal microscopy / Endo- somes / Ii chain / Major histocompatibility complex class I1
and 41 kDa [6, 71 resulting from alternative splicing [8]. Once associated in the ER, MHC class 11-Ii complexes are transported through the Golgi with a half-time of 30-60 min [9] . Ii chains are then proteolyticaly cleaved and dissociate from MHC class I1 molecules [lo]. The transient intracellular association of Ii with aB heterodimers led to the hypothesis that Ii could be required for the assembly and/or the transport of MHC class I1 molecules towards endosomal compartments [3]. Recently, Bakke and Dob- berstein have shown that without class I1 molecules, full- length human Ii chains are present in endosomal compart- ments whereas cytoplasmic truncation mutants of Ii are expressed at the cell surface [11].These results suggest that Ii chains have a targeting signal directing the HLA DR-Ii complexes to the endosomal pathway.
In Ii-negative fibroblasts, supertransfection of Ii facilitates the presentation of externally provided antigens [ 12,131 and of some endogenous antigens such as hen egg lysozyme molecules being secreted or ER resident [14]. Ii was not required, however, for the presentation of particular proteins such as RNAse [12, 151. Both types of results have recently been confirmed using splenic B cells derived from mice lacking Ii chains [16]. Several functions of Ii chains have been proposed. First, Ii could protect the MHC class I1 molecules from binding peptides until the ternary ap-Ii complex reaches an endosomal compartment where Ii is degraded leaving ap heterodimers free to bind antigenic peptides. This hypothesis is supported by in vitro studies showing that Ii inhibits peptide binding to class I1 mole- cules [17, 181 and by the detection of Ii-derived peptides that are associated with class I1 molecules and impair the
0014-2980/93/1212-3158$10.00 + ,2510 0 VCH Verlagsgesellschaft mbH, D-69451 Weinheim, 1993

Eur. J. Immunol. 1993.23: 3158-3166 Ii induction of stable class I1 molecules 3159
binding of antigenic peptides [19]. Second, Ii could be involved in the endosomal targeting of class I1 heterodi- mers as indicated by their immunocytochemical localiza- tion in cells transiently expressing the human DR molecules [20]. Moreover, it has recently been shown that the cytoplasmic domain of Ii can redirect a reporter neura- minidase molecule to endosomal compartments [21] .These two functions are presumably linked since the release of Ii from class I1 molecules involves a proteolytic event sensi- tive to weak bases and to thiol protease inhibitors [lo, 22, 231. Furthermore, a proteolytic inhibitor such as leupeptin can affect Ii cleavage and impairs the conversion of newly synthesized DR class I1 molecules to SDS stable forms [24]. In this latter experiment, leupeptin could affect the con- formation of class I1 molecules because of a direct inhibi- tion of Ii cleavage, or because of an indirect inhibition of the transport and the fusion of endosomal vesicles with prelysosomal compartments as recently reported [25].
In this study, we have tested whether in a non lymphoid cell line, Ii chains are sufficient to change the conformation, the intracellular transport and the sites of intracellular accumu- lation of murine class11 molecules. As detected with anti-conformational antibodies, the strict co-localization of class I1 with cathepsins, observed independently of Ii, strongly suggests that intact heterodimeric forms of class I1 molecules are accumulated in intracellular compartments such as the prelysosomes and the lysosomes.
2 Materials and methods
2.1 Materials, cells and antibodies
All chemicals and proteins were purchased from Sigma Chemical Co (St. Louis, MO). Electrophoresis was per- formed with products purchased form Serva (Heidelberg, FRG) . PC31.3 IAk-positive cells [26] were supertransfected with the Ii gene from the cosmid clone H-2k 107 [27] provided by Dr. B. Malissen, using the calcium phosphate precipitation method as described [28]. Transfectants were selected by resistance to G418. PC51.14, IAk and Ii double-positive clones were selected for high expression of Ii chains as tested both by indirect immunofluorescence after fixation and permeabilization, and by immunoprecipitation after 1 h of metabolic labeling and previous class I1 immunopre- cipitation.
Class I (anti-H2Kk 84. 17. 2), transferrin receptor (TfR) (H129-121) and class 11-specific (H39. 146. 1, 39F) and (H40. 481, 40B) mAb, were kindly provided by Dr. M. Pierres. Rabbit and goat antisera directed against rat, mouse and rabbit IgG were purchased from Immuno- tech, (Marseille). Rabbit antisera directed against cathep- sins B, D, H and L were kindly provided by Dr. Bernd Wideranders.
Antigen presentation assays were essentially performed using the soluble hen egg lysozyme (HEL) and the HEL 46-61 specific 3A9 T cell hybridoma [29].
2.2 Biochemical analysis
For pulse-chase radiolabeling cells were incubated in methionine cysteine-free medium (Gibco BRL, NY), for 45 min at 37 "C. They were then washed once with the same medium before incubation for 20 rnin in 3 ml of prewarmed methionine cysteine-free medium containing [35S]methio- nine and [35S]cysteine (0.2 mCi of each/ml) (Amersham). Cells were then incubated at 37 "C for various chase periods with warm complete growth medium supplemented with 2 m~ of both methionine and cysteine.
Cell surface and in situ iodination were performed as described previously [26,30]. Labeled cells were lysed on ice for 45 min in 50 mM Tris pH 7.5, containing 150 mM NaCI, 0.5% Triton X-100, 5mh4 EDTA and protease inhibitors. Lysates were then cleared from the nuclear pellet by centrifugation at 14000 rpm for 20 min. Except when specified, the resulting supernatant was clarified by a 2-h incubation with rabbit anti-mouse or anti-rat IgG preadsorbed on protein A-Sepharose beads (Pharmacia, Uppsala, Sweden) at 4 "C. Extracts were immunoprecipi- tated with specific antibodies followed by anti-IgG-protein A-Sepharose as described [26]. For SDS-PAGE analysis the samples were fractionated as described [31] on 10 to 15% SDS PAGE gradient gels. For neuraminidase (NaNase) treatment, digestion was directly performed on immune complexes adsorbed to protein A-Sepharose. Immunopre- cipitates were incubated for 4 h at 37°C with 100 mU of enzyme in 50 mM sodium acetate, pH 5.8, supplemented with 1mM CaC12. For endoH treatment, immunoprecipi- tated molecules were dissociated from protein A-Sepha- rose by boiling the samples in the presence of 1% SDS.The supernatant were then treated in sodium citrate buffer 0.15 M, pH 5.5, in the presence or absence of 5 mu of endoH for at least 4 h. Gels were fixed and fluorographed with Intensify (New England Nuclear, Boston, MA). They were then dried and exposed to Kodak X-OMAT AR film at -70 "C.
2.3 Fluorescence studies of fluid-phase and receptor-mediated endocytosis
FITC was used to couple fluorescein to iron loaded trans- ferrin: 75 p1 NaHC03, (1 M, pH 9) was added to 300 $ of transferrin (1.67 m) freshly loaded with iron in PBS. A '/ioooo dilution of a 30 mg fluorescein solution in 0.5 ml M NaHC03 was added to allow the conjugation of 50 pg FITC per mg of transferrin at five time points with 10 min of incubation between each addition. Fluorescein-conjugated transferrin (FITC-Tf) was passed over a Sephadex G-25 column (0.5 cm x 15 cm) (Pharmacia) and eluted with PBS.
For transferrin uptake, cells grown for 2 days on glass coverslips were incubated first in 0.5% BSA-PBS++ for 45 rnin and then with 60 n~ of FITC-Tf in the same prewarmed medium for 20 rnin at 37 "C before chilling and fixation. FITC was coupled to 0.5 g of ovalbumin in 9 ml of 250 mM NaHC03 (pH 9) by four additions every 10 rnin of 200 pl of iodoacetamide fluorescein (IAF; 12.5 mg/ml in M HCOJ, pH 9). The reaction was stopped by adding N& CI to a 50 mM final concentration. The labeled ovalbumin (FITC-OVA) was separated from free IAF by gel filtration
3160 M. Humbert, G. Raposo, I? Cosson et al. Eur. J. Immunol. 1993.23: 3158-3166
over a Sephadex G-25 column. Aggregates of labeled protein were retained by anion exchange on a DE52 column and ovalbumin was eluted with 30mM KPO4 buffer, pH 7.4, 150 mM NaCl.
zidine, H202 and pelleted. Pellets were post fixed in 1% osmium tetroxide, dehydrated in ethanol and embedded in Epon.
3 Results 2.4 Intracellular immunofluorescence
Fibroblast cells were cultured for 48 h on glass coverslips. After washing three times with PBS, cells were fixed for 15 min with a 2% paraformaldehyde 0.1% glutaraldehyde solution in PBS. Before permeabilization with a 1% Triton XlOO solution in PBS. Cells were subsequently washed in PBS. Specimens were further incubated in a solution of 1 mg/ml sodium borohydride. The cells were finally washed with 0.2% gelatin in PBS. Antibodies were diluted to a 10 pg/ml concentration in gelatin-PBS. Cells were incu- bated for 20 min with first antibodies at room temperature and then washed three times before incubation with either FITC- or RITC-coupled anti-IgG antiserum (l/200 dilution in PBS gelatin). Slides were mounted in Mowiol medium.
2.5 Confocal microscopy and double-fluorescence analysis
Confocal laser scanning microscopy was performed using a Leica CLSM (Leica, FRG) instrument based on a Leitz Diaplan microscope interfaced with an Argon ion Laser model 2020 (Spectra-Physics, Mt View, CA) adjusted at 488nm and with an HeNe air cooled laser providing a 543 nm line. A focal series of four horizontal planes of section spaced by 0.75pm have been monitored first for fluorescein using 488 nm excitation via a 510 nm dichroic mirror, a FITC band pass 520-560 nm barrier filter and a green sensitive photomultiplier. Second the same four slices were scanned to detect the rhodamine signal using 543 nm excitation via a 580 nm dichroic mirror, a long pass barrier filter above 580 nm and a red sensitive photomultiplier.The two 8 bits-encoded 512 x 512 pixel images from registered horizontal planes of section were combined and visualized with a computer-generated red and green pseudo-color scale adapted for rhodamine and fluorescein and printed using a thermal sublimation color printer UP 3000 (Sony, Japan).To enlighten the coincidence between the two dyes, white color was applied to areas containing simultaneously more than 50 YO of the maximum fluorescein and rhodam- ine signals as indicated on the red and green pseudo-color scale.
2.6 Electron microscopy
Transferrin was coupled to 5-nm colloidal gold particles (Tf-5 nm) as described [32]. For internalization, cells grown on glass coverslips were incubated at 4 "C with Tf-5 nm for 60 min in serum free medium, washed at 4"C, then incubated at 37°C for 20 min before fixation for 30 min with a mixture of 2% paraformaldehyde and of 0.1% glutaraldehyde. Cells were then permeabilized with 0.2% saponin before incubations in the presence of 0.02% saponin with mAb 40B followed by immunolabeling with goat anti-mouse IgG conjugated to HRI! After fixation in 1% glutaraldehyde, they were incubated with diaminoben-
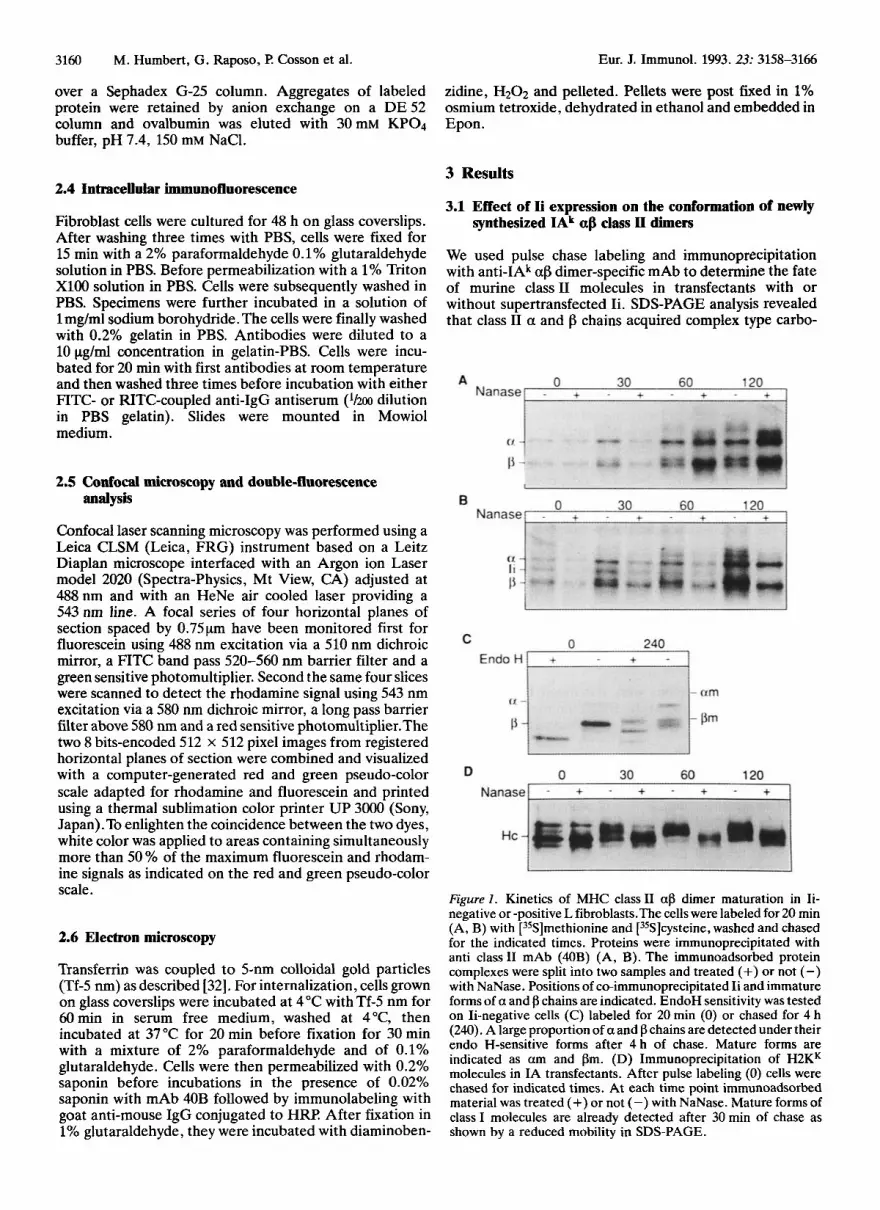
3.1 Effect of Ii expression on the conformatiod of newly synthesized IAk US class II h e r s
We used pulse chase labeling and immunoprecipitation with anti-IAk ap dimer-specific mAb to determine the fate of murine class I1 molecules in transfectants with or without supertransfected Ii. SDS-PAGE analysis revealed that class I1 a and f3 chains acquired complex type carbo-
Figure 1 . Kinetics of MHC class I1 af3 dimer maturation in Ii- negative or -positive L fibroblasts.The cells were labeled for 20 min (A, B) with [35S]methionine and [35S]cysteine, washed and chased for the indicated times. Proteins were immunoprecipitated with anti class I1 mAb (40B) (A, B). The immunoadsorbed protein complexes were split into two samples and treated (+) or not (-) with NaNase. Positions of co-immunoprecipitated Ii and immature forms of a and f3 chains are indicated. EndoH sensitivity was tested on Ii-negative cells (C) labeled for 20 min (0) or chased for 4 h (240). A large proportion of a and p chains are detected under their endo H-sensitive forms after 4 h of chase. Mature forms are indicated as a m and f3m. (D) Immunoprecipitation of H2KK molecules in IA transfectants. After pulse labeling (0) cells were chased for indicated times. At each time point immunoadsorbed material was treated (+) or not (-) with NaNase. Mature forms of class I molecules are already detected after 30 min of chase as shown by a reduced mobility in SDS-PAGE.

Eur. J. Immunol. 1993. 23: 3158-3166 Ii induction of stable class I1 molecules 3161
hydrates in both transfectants as assessed by their reduced mobility and by the appearance of neuraminidase sensitive forms (Fig. 1A and B). Endo H sensitivity was also assayed using a 20-min pulse labeling followed by a chase of 4 h.The proportion of mature class I1 f3 chains after 4 h of chase estimated by scanning densitometry of different SDS- PAGE gels, amounts to 40% of the total f3 chains immu- noprecipitated in Ii-negative transfectants (Fig. 1C). In IA-Ii-positive cells, the maturation of the carbohydrate moieties of class I1 fl chain was correlated with the progres- sive disappearance of the co-immunoprecipitated p31 form of Ii. Even after 2 h of chase, immature afl oligomers were always detected in both cell types (compare Fig. 1 A and lB), supporting previous reports showing that the exit of class I1 molecules from the ER is very slow if not partially defective in mouse fibroblasts [33,34] while newly synthe-
Figure 2. Formation of SDS-resistant class I1 afl dimers. (A) IA, Ii and IA cells were pulse labeled for 30 min and chased for 4 h. Proteins immunoprecipitated using anti-class I1 mAb (40B) were treated with sample buffer containing dithiotreitol at 95 "C (lane B) or at 20°C (lane NB). Compact forms (CF) of afl dimers, a, p, Ii p31 form and a 12-kDa polypeptide only detected in IA, Ii transfec- tants, are indicated. (B) IA, Ii transfectants were pulse labeled (20 min) as above and chased for times indicated. Proteins were immunoprecipitated with anti class I1 mAb. At time 240 min the sample was split and treated (240+) or not (2W) with NaNase. Appearance of a 12-kDa polypeptide associated with class I1 molecules correlates with the dissociation of the p31 form of Ii from ap dimers of classII. (C) (IA) and (D) IA, Ii cells were continuously labeled for 8 h (left two lanes) and further chased for 10 h (right two lanes). Proteins were immunoprecipitated with anti class I1 mAb and were treated with sample buffer containing dithiotreitol at 95 "C (lane B) or at 20°C (lane NB). Compact forms (CF), a, 0, Ii p31 and p41 chains are indicated. Actin present in some of the immunoprecipitated samples is taken as a molecular weight control.
sized class I molecules become sialilated within 30 min (Fig. lD).These results indicate that the presence of Ii does not drastically affect the rate of carbohydrate maturation associated with the transport of class I1 molecules to the Golgi complex in L cells. As shown previously using mouse spleen cells, IAk class IT dimers acquire a particular SDS resistance at 20 "C (compact form), once they dissociate from the p31 Ii chain [35]. To determine whether Ii expression controls the formation of afl compact dimers, the two cell types were pulse labeled for 30 min, chased for 4 h and class I1 immunoprecipitates were treated for SDS- PAGE with or without preheating at 95 "C. Class I1 compact forms were clearly induced by the presence of Ii 4 h after chase, but a weak band corresponding to compact forms was detected without Ii (Fig. 2). During the conversion from non compact to compact af3 dimers, class I1 molecules were associated with a polypeptide having a relative molecular mass of 12 kDa exclusively in Ii-positive cells (Fig. 2A). This p12 protein appears after 1 h of chase and progressively increases until1 2 h of chase, whereas the amount of p31 gradually decreased from 1 to 4 h of chase (Fig. 2B).This is correlated with the appearance of mature and compact class11 dimers. We suggest that this p12 protein corresponds to the p12 degradation product of the human Ii chains. This human p12 fragment represents a C-terminal cleavage product [36], generated in endosomal compartments. It contains the cytoplasmic, transmem- brane and a portion of the exoplasmic domain of Ii. It is likely that Ii degradation shown here in murine fibroblasts with this p12 intermediate product accompanies the con- version of class I1 dimers to compact forms.
The induction of compact forms of class I1 in Ii positive cells was confirmed by examining class I1 dimers continuously labeled for 8 h (Fig. 2C Ii negative, and 2D Ii positive). After a chase of 10 h more than 50% of class I1 dimers were detected as SDS stable compact forms in Ii positive cells (Fig. 2D) whereas without Ii, compact af3 dimers were almost undetectable.
3.2 Effect of Ii expression on the stabTty of IAk afl dimem accumulated in late endosomal compartments
The recent demonstration that leupeptin inhibits the complete degradation of Ii and the conversion of class I1 into an SDS stable forms suggests that compact class I1 molecules would already be present in endosomal compart- ments accessible to endocytic markers before their appea- rance at the cell surface [24]. We have compared the conformation of class I1 molecules immunoprecipitated after in situ iodination of endosomal compartments [30] following a protocol already described for the detection of intracellular h4HC class I1 molecules [26], with those present at the cell surface in Ii-positive and Ii-negative cells. Significant amounts of class I1 molecules were only detect- able after a lag time of 30 to 40min of lactoperoxidase (LPO) uptake (Fig. 3A and B) in both Ii-positive and Ii-negative transfectants. To evaluate the accessibility of endosomal compartments as a function of the internaliza- tion time, we have monitored the kinetics of the iodination of the transferrin receptor (TfR) to LPO endocytosed from the fluid phase in the same specimens. No specific increase in the level of labeled TfR was obtained between 5 and

Eur. J. Immunol. 1993. 23: 3158-3166 3162 M. Humbert, G. Raposo, €? Cosson et al.
A B atically used to avoid the detection of single a and chains. As an early endosomal marker, FITC-Tf was internalized for 20 min at 37 "C before fixation. Immunofluorescence labeling for class I1 was performed using rhodamine cou- pled secondary antibodies. To evaluate the co-localization between the two markers we have superimposed the rhodamine and fluorescein images. There is clearly no overlap between MHC class I1 molecules and the early endosomal labeling with FITC-Tf as assessed by the lack of yellow double labeled Structures. No intracellular Tf/IAk double-labeled structures were detectable in single IA- positive or double IA, Ii-positive cells, while distinct populations of highly labeled dots appear for both markers individually (Fig. 4A, B). Upon incubation of the cells for 20 min with the FITC-OVA fluid-phase marker to label the endocytic pathway, a small subset of the class 11-positive intracellular structures were double positive as assessed in these micrographs by white dots pointing to double- positive structures (Fig. 4C, D). At shorter times of ovalbumin endocytosis (5-8 min), no CQlOCahatiOn was detectable (not shown). After 20 min of uptake of FITC- OVA, we have more frequently detected a partial CO- localization between fluid-phase markers and class I1 mole- cules in IA, Ii double-transfectants than in IA single ones.
Figure 3. In situ iodination of MHC class I1 dimen in endosoma1 compartments in IA and IA, Ii transfectants. (A) IA, Ii- and (B) IA-transfected cells were allowed to internalize lactoperoxidase (LPO) for indicated times. Class I1 molecules (A) transfenin receptor ( C ) and MHC class I molecules (D) were immunopreci- pitated from the cell lysates used in (A). Class 11-immunoprecipi- tated samples were analyzed by SDS-PAGE without heating-at 95 "C. Note that after 30 min of LPO internalization compact forms (A) or unstable forms (B) are detected. Compact (A) unstable forms (B) of ap dimers are present at the cell surface as assessed by cell surface iodination and immunoprecipitation with the 40B mAb (lanes S). CF indicates the position of SDS stable afl dimers.
40min after the initiation of LPO internalization as expected for an early endosomal marker (Fig. 3C). As a negative control (Fig. 3D), the background signal due to cell surface molecules was estimated using class I molecules (H2Kk), which are not located in endosomal compartments [37]. As shown in Fig. 3, ap dimers detected after 30 to 40min of LPO internalization were sensitive to SDS dissociation in single IA transfectants, whereas much of the class I1 ap dimers were SDS stable in double IA, Ii transfectants. Using surface iodination, all class I1 ap dimers were compact in Ii positive cells (Fig. 3A lane S) as already shown for normal spleen cells [35], whereas without Ii, much of the cell surface class I1 molecules were sensitive to SDS dissociation (Fig. 3B lane S). Altogether, class I1 molecules are accumulated in late elements of the endos- omal pathway where SDS stability is induced by co- expression of Ii chains.
3.3 IntraceUular localization of MHC class I1 dimers molecules in Ii-positive and Ii-negative cells
To address the question of whether the invariant chain influences the accumulation of class I1 molecules into a subpopulation of endosomal compartments, we used con- focal microscopy and double immunofluorescence labeling. Anti-conformational anti-class I1 antibodies were system-
Next, we have analyzed the precise localization of the endocytic pool of class I1 molecules by colabeling with lysosomal enzymes or with lgpll0, a constituent of the lysosomal membrane [38]. As shown here, double labeling with anti class I1 and anti-cathepsin D reveals a significant co-localization as supported by a high spatial correlation between the two patterns of staining (Fig. 4E, F). In both Iif and Ii- transfectants only the cathepsin molecules or lgp 110 gave such a high correlation with the distribution of class I1 molecules in intracellular vesicles.The absence of Ii does not significantly affect this particular distribution of class I1 molecules in late elements of the endosomal pathway in transfected L cells. Although we have found no evidence for a colocalization between recycling Tf and intracellular class I1 molecules at the confocal microscope level, a small amount of class I1 molecules could still be present in the early endosomes involved in the recycling of Tf to the cell surface. To test this hypothesis at the ultrastructural level, cells were allowed to internalize 5-nm gold particles conjugated to transfenin (Tf 5 nm), for 20 min at 37 "C. On the electron micrographs, the endocy- tosed Tf 5 nm was present in electron lucent endosomes as well as in tubulo-vesicular structures (Fig. 5) .To localize the class I1 molecules, we have used a highly sensitive immu- noperoxidase detection method that leads to the deposition of an electron-dense reaction product in all class 11-positive compartments even with a low expression. The immunope- roxidase reaction was controlled and the micrographs were carefully printed to detect an eventual superimposition between gold particles and the electron dense reaction product. As shown on the micrographs, the class I1 mole- cules were almost uniquely located in multivesicular bodies both in Ii-positive (Fig. 5A) and in Ii-negative (Fig. 5B) transfectants. Tf gold conjugates were uniquely present in class 11-negative tubulo-vesicular early endosomes.

Eur. J. Immunol. 1993. 23: 3158-3166 Ii induction of stable class 11 molecules 3163
Figure 4. Double-fluorescence confocal microscopy of MHCclass I1 molecules versus markers of the endosomal pathway. (A, B) FITC-Tf (60 nM) was continuously internalized for 20 min at 37°C by transfectants grown on coverslips. FITC-Tf could be recycled after 20 min of chase using an excess of unlabeled Tf. Cells were then fixed and labeled for classII IA molecules with the 4OB mAb and rhodamine-labeled goat anti mouse IgG. Superimposed images of internalized Tf and intracellular class I1 molecules are shown in IA, Ii (A) and IA (B) transfectants. Similarly, FITC-OVA (1.75 mglml) was continuously internalized for 20 min at 37°C by IA, Ii (C) or IA (D) transfectants and intracellular class II molecules were labeled after fixation of the cells. Double immunofluorescence staining of class I1 molecules and cathepsin D in IA, Ii (E) and IA (F) transfectants.

3164 M. Humbert, G. Raposo, P. Cosson et al. Eur. J. Immunol. 1993. 23: 3158-3166
Figure 5. Subcellular localization of class I1 molecules and of Tf-5 nm internalized for 20 min in IA, Ii (A) and IA (B) transfectants. Cells were then fixed and class I1 molecules were revealed by the immunoperoxidase reaction within multivesicular bodies.Tf-5 nm (arrows) are detected in electron-lucent compartments (A) or in tubulo-vesicular structures (B). Bars, 200 nm.

Eur. J. Immunol. 1993. 23: 3158-3166 Ii induction of stable class I1 molecules 3165
4 Discussion
4.1 General considerations
Antigenic peptides bound to MHC class I1 molecules are essentially derived from exogenous proteins that have been internalized [2, 391. During their intracellular transport in APC, newly synthesized class I1 molecules are transiently associated with Ii. Ii is subsequently clipped off by acid proteases [22] presumably at the intracellular site where antigenic peptides are produced [24,35], indicating that newly synthesized class I1 molecules could intersect the endocytic pathway as shown in Ii-positive human EBV transformed B cells [40]. In addition, we showed previously that a pool of intracellular IAk class I1 molecules was accessible to fluid-phase endocytosis in Ii-positive and Ii-negative L cells indicating that IAk molecules can also gain access to the endocytic pathway of transfected fibro- blasts by themselves [26]. The Ii chain could, therefore, serve two important functions needed to favor the presen- tation of exogenous antigens. First, Ii could prevent class I1 molecules from binding peptides until they reach post Golgi compartments in which the dissociation of the complex occurs [19]. Second, Ii has a targeting signal that is sufficient to redirect human DR class I1 molecules toward a subset of endosomal compartments containing exogenous proteins and processed peptides [ l l , 201. Indeed, when coexpressed with cytoplasmic domain deletion mutants of Ii, DR ap dimers can gain access to the cell surface in association with uncleaved Ii chain [41]. Recent studies have shown that Ii expression influences the amount and the conformational stability of cell surface class I1 dimers in spleen cells [16]. Whether Ii expression directly influences the conformation of class I1 molecules located in endoso- ma1 compartments was unknown. Here, we have analyzed whether Ii expression controls the intracellular distribution and the conformational stability of class I1 molecules, in a model APC.
4.2 Conformational stability of class II dimers in late endosomal compartments
By analogy with class I molecules [42,43], the intracellular transport of class11 molecules might be affected by the availability of peptides having a high binding potential [35]. This process could further be controlled by the rate of degradation and dissociation of the associated Ii chain in acidic compartments as suggested before [22, 241. We have shown here that in the presence of Ii, SDS stable class I1 dimers are located in late endosomal Compartments and at the cell surface. Without Ii expression, newly synthesized class I1 molecules are still able to form SDS stable com- plexes but to a much smaller extent compared to Ii-positive cells. Ii is obviously playing a catalyst role during the acquisition of compact dimeric forms since in its absence newly synthesized class I1 dimers that have left the ER in L cells are essentially SDS unstable.
To determine the relationship between the pool of intracel- lular class I1 molecules and endocytic markers, we have characterized the subcellular distribution of IAk molecules in L cell transfectants coexpressing or not Ii chains. At the light microscopy level and using ap dimers-specific mAb, the MHC class I1 molecules have been localized within the
same type of vesicular structures dispersed in the cytoplasm even without Ii expression. At the steady state, the early endosomal compartments are never labeled for ciass I1 molecules, suggesting that Ii is unlikely to target class I1 molecules to early endosomes. However, one cannot exclude a very rapid and undetectable transit of class I1 molecules through early endosomes. We have found here that the intracellular distribution of class I1 molecules was highly correlated with that of cathepsin D. A similar pattern was obtained for the class I1 molecules and the 1gpllO lysosomal protein [38], in Ii+ and Ii- cells (data not shown). The IAk class I1 molecules are thus accumulated together with lysosomal markers independently of Ii expression. Recent investigations on the intracellular distribution of class I1 in different cell types allowed the identification of endosomal Compartments that contain abundant class I1 molecules [36, 37, 40, 441 but no consensus was achieved concerning additional markers. Peters et al. [37] have defined a MHC class 11-specific compartment (MIIC) which is clearly related to lysosomes but is distinct from both the early endosomes and the cation-independent mannose-6-phosphate receptor (CI-M6P-receptor)-rich compartments. In L cells, the distribution of class I1 molecules was indistinguishable from that of all lysosomal markers we used. We do not know yet if lysosomal and/or prelysosomal class I1 molecules are destined to the cell surface. Nevertheless, a p dimeric molecules are clearly accumulated in these compartments indicating that they are resistant to acid protease cleavage in L fibroblasts.
Taken together our results suggest that Ii is not essential for the targeting of IAk ap dimers to lysosomes in fibroblast L cells. Instead of addressing the question of the actual route followed by the aPIi complexes, our experiments are showing for the first time that class II IAk complexes might contain targeting motives that differ from those expressed on the cytoplasmic tail of the invariant chain, but that are sufficient to address ap dimers to the late elements of the endosomal pathwaywe do not know whether Ii and class I1 specific targeting signals are specifying exactly the same subcellular compartment. However, one can envisage that these types of permanent targeting signals, possibly located in the cytoplasmic domains of class I1 dimers, could be redundant with, and complement, specific signals present in the cytoplasmic tail of the Ii chain. Following this assumption, binding of Ii to all class I1 isotypes would ensure a transient targeting of a@Ii complexes to endosomal compartments. A transient delivery of the class I1 mole- cules to the early endosomes or even to the plasma membrane cannot be ruled out.
4.3 Functional implications of Ii expression on antigen presentation
Having the same final destination, the precise intracellular pathway followed by MHC class I1 molecules might still be influenced by Ii. Correspondingly, Ii enhances the presen- tation of some but not of all antigens depending on the sites of processing [12] including the presentation of ER- resident antigens [14]. AS a second possibility, Ii or Ii proteolytic fragments could directly influence the intrinsic capacity of MHC class I1 dimers to bind certain peptides in acidic Compartments as recently pointed out by Riberdy et al. who showed that Ii-derived peptides still associated with

3166 M. Humbert, G. Raposo, P. Cosson et al. Eur. J. Immunol. 1993. 23: 3158-3166
class I1 can inhibit antigen binding in vitro [19]. We have found here that class I1 molecules that were accessible to fluid-phase markers such as the lactoperoxidase in Ii- negative cells, were unstable. To assess whether the Ii- dependent induction of compact forms was functionally relevant in L fibroblasts despite a similar intracellular distribution, we have tested their antigen presentation capability using the HEL antigen, detected with the IAk-restricted, and 46-61 epitope-specific 3A9 T cell hybridoma [14,29]. As expected from previous reports [13,16], the complementation with Ii was here sufficient to shift the dose response curve indicating that a few pg per ml of HEL is needed in the presence of Ii whereas without Ii, 100-fold more HEL is required to achieve a similar T cell response (data not shown). Without Ii, class I1 molecules were equally able to associate with a 46-61 HEL peptide delivered directly to the cell medium as pointed out in previous reports [12, 15, 161. As a third possibility, and in contrast to professional APC expressing antigen-specific B cell receptors andor Fc receptors, the processing of different antigens and their association with class II might occur in various intracellular compartments. In fibroblasts lacking an antigen-specific internalization process, Ii expression could eventually regulate the presentation of some but not of all antigens, depending on their specific degradation pathways.
In conclusion, we have shown that Ii chains are necessary and sufficient to induce a major conformational change of class II ap dimers in fibroblasts. Without Ii, the accumula- tion of SDS unstable af! dimers in late endosomal compart- ments strongly suggests that under normal conditions, Ii dissociation and/or degradation increases the capacity of class II dimers to bind peptides derived from exogenous proteins in endosomal compartments as predicted [19,24] and recently pointed out using cells from mice lacking Ii chains [16]. We think that the intracellular compartments where processed antigens bind to class I1 molecules are close, if not coincident to the sites where Ii dissociates from apIi complexes allowing the formation of SDS-stable af! dimers.
We thank Jolle Henry for her excellent technical help, E Theuillon for his participation in the elaboration of the software used to superimpose two digital images and Drs. M. Moore and I? Golstein for critically reading the manuscript.
Received May21, 1993; in final revised form August13, 1993; accepted August 13, 1993.
5 References
1 Grey, H.M. and Chestnut, R., Immunol. Today 1985. 6: 101. 2 Unanue, E.R., Annu. Rev. Immunol. 1984. 2: 395. 3 Kvist, S., Wimar, K., Claesson, L., Peterson, P. A. and Dob-
4 Machamer, C.E. and Cresswell, P., J. Immunol. 1982. 129:
5 Cresswell, P., Blum, J. S., Kelner, A. N. and Marks, M. S. , CRC
berstein, B., Cell 1982. 29: 61.
2564.
Crit. Rev. Immunol. 1987. 7: 31.
6 Claesson, L., Larhammar, D., Rask, L. and Peterson, PA.,
7 Lipp, J. and Dobberstein, B., J. Cell. Biol. 1986.102: 2162. 8 Strubin, M., Berte, C. and Mach, B., EMBO J. 1986.5: 3483. 9 Long, E. O., Immunol. Today 1989.10: 232.
10 Cresswell, P. and Blum, J. S. in Pernis, B., Silverstein, S. C. and Vogel, H. J. (Eds) Intracellular transport of class I1 antigens in processing and presentation of antigens. Academic Press, San Diego 1988, p43.
Proc. Natl. Acad. Sci USA 1983. 80: 7395.
11 Bakke, 0. and Dobberstein, B., Cell 1990. 63: 707. 12 Nadimi, E, Moreno, J., Momburg, F., Heuser, A., Fuchs, S.,
Adorini, L. and Hammerling, G., Eur. J. Zmmunol. 1991.21: 1255.
13 Stockinger, B., Pessara, U., Lin, R.H., Habicht, J., Orez, M. and Koch, N., Cell 1989. 56: 683.
14 Humbert, M., Bertolino, P., Forquet, F., Rabourdin-Combe, C., Gerlier, D., Davoust, J. and Salamero, J., Eur. J. Zmmunol. 1993. 23: 3167.
15 Peterson, M. and Miller, J., Nature 1990. 345: 172. 16 Viville, S., Neefjes, J., Lotteau,V, Dierich, A., Lemeur, M.,
Ploegh, H., Benoist, C. and Mathis, D., Cell 1993. 72: 635. 17 Roche, PA. and Cresswell, I?, Nature 1990. 345: 615. 18 Teyton, L., OSullivan, D., Dickson, PW., Lotteau,V., Sette,
A., Fink, P. and Peterson, PA., Nature 1990. 348: 39. 19 Riberdy, J.M., Newcomb, J. R., Surman, M. J., Barbosa, J. A.
and Cresswell, P., Nature 1992. 360: 474. 20 Lotteau,V.,Teyton, L., Peleraux, A., Nillson,T., Karlsson, L.,
Schmid, S. L., Quaranta,V. and Peterson, P. A., Nature 1990. 348: 600.
21 Bakke, O., Bremnes, B., Hardersen, H. and Madsen,T., J. Cell. Biochem. 1993. 17C: Abstract H210.
22 Blum, J. S. and Cresswell, I?, Proc. Natl. Acad. Sci. USA 1988. 85: 3975.
23 Nowell, J. and Quaranta,V., J. Exp. Med. 1985. 162: 1371. 24 Neefjes, J.J. and Ploegh, H.L., EMBO J. 1992. 11: 411. 25 Zachgo, S . , Dobberstein, B. and Griffiths, G., J. Cell Sci. 1992.
103: 811. 26 Salamero, J., Humbert, M., Cosson, P. and Davoust, J., EMBO
J. 1990. 9: 3489. 27 Yamamoto, K., Koch, N., Steinmetz, M. and Hammerling,
G.J., J. Immunol. 1985. 134: 3461. 28 Wigler, M., Sweet, R., Sim, G. K. ,Wold, B., Pellicer, A., Lacy,
E., Maniatis,T., Silverstein, S. and Axel, R., CeN1979.16: 777. 29 Allen, P.M. and Unanue, E. R., J. Immunol. 1984.132: 1077. 30 Ludwig, T., Griffiths, G. and Hoflack, B., J. Cell Biol. 1991.
31 Salamero, J., Sztul, E. and Howell, K.H., Proc. Natl. Acad.
32 Slot, J. W. and Geuze, H. J., Eur. J. Cell Biol. 1985. 38: 87. 33 Simonis, S., Miller, J. and Cullen, S. E., J. Immunol. 1989: 143:
34 Anderson, A.P. and Miller, J., Proc. Natl. Acad. Sci. USA
35 Germain, R.N. and Hendrix, L.R., Nature 1991.353: 134. 36 Pieters, J., Hortzman, H., Bakke, O., Griffiths, G. and
37 Peters, P.J., Neefjes, J.J., Oorshot, V., Ploegh, H.L. and
38 Lewis,V., Green, S.A., Marsh, M.,Vihko, P., Helenius, A. and
39 Lanzavecchia, A., Immunol. Rev. 1987. 99: 39. 40 Neefjes, J. J., Stollorz,V., Peters, J. P., Geuze, H. J. and Ploegh,
41 Roche, P. A.,Teletski, C. L., Karp, D. R., Pinet,V., Bakke, 0.
42 Schumacher, T. N., Heemels, M. T., Nefjees, J. J., Kast, W. M.,
43 Townsend, A., OhlCn, C., Bastin, J., Ljunggren, H. G., Foster,
44 Guagliardi, L. E., Koppelman, B., Blum, J.S., Marks, M.S.,
115: 1561.
Sci. USA 1990. 87: 7717.
3619.
1992. 89: 2282.
Lipp, J., J. Cell. Biol. 1991. 115: 1213.
Geuze, H.J., Nature 1991. 349: 669.
Mellman, I., J. Cell Biol. 1985. 100: 1839.
H.L., Cell 1990. 61: 171.
and Long, E.O., EMBO J. 1992. 11: 2841.
Melief, C. J. and Ploegh, H. L., Cell 1990. 62: 563.
L. and Karre, K., Cell 1989.340: 443.
Cresswell, F? and Brodsky, EM., Nature 1990. 343: 133.




























