Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
The neural adhesion molecule L1CAM confers chemoresistancein human glioblastomas
Janka Held-Feindt a,⇑, Sabine Schmelz a, Kirsten Hattermann b, Rolf Mentlein b, H. Maximilian Mehdorn a,Susanne Sebens c
a Department of Neurosurgery, University Medical Center Schleswig-Holstein UKSH, Campus Kiel, Arnold-Heller-Str. 3, Building 41, 24105 Kiel, Germanyb Department of Anatomy, University of Kiel, Olshausenstr. 40, 24098 Kiel, Germanyc Institute for Experimental Medicine, Department of Internal Medicine I, University Medical Center Schleswig-Holstein UKSH, Campus Kiel, Arnold-Heller-Str. 3, Building 6, 24105Kiel, Germany
a r t i c l e i n f o
Article history:Received 16 February 2012Received in revised form 31 July 2012Accepted 18 August 2012Available online 28 August 2012
Keywords:CD171GlioblastomaChemoresistanceTGF-b1Stem cellsL1CAM
a b s t r a c t
Glioblastoma multiforme (GBM) represents the most common and malignant brain tumor. GBM tissuesexhibit elevated expression of the transforming growth factor-beta1 (TGF-b1) and the adhesion moleculeL1CAM. This study investigated the mechanism of L1CAM regulation in GBM cells and its role in themediation of chemoresistance. L1CAM expression levels varied in GBM cells being highest in A172 cellsand low in T98G cells. Inhibition of TGF-b1 signaling in A172 cells reduced L1CAM expression and viceversa stimulation with exogenous TGF-b1 led to upregulation of L1CAM in T98G cells. Additionally,TGF-b1 and L1CAM expression increased during differentiation of glioma stem-like cells. L1CAM express-ing GBM cells and differentiated glioma stem-like cells showed a reduced apoptotic response after treat-ment with the chemotherapeutic drug temozolomide. Accordingly, siRNA-mediated knock-down ofL1CAM in A172 cells and differentiated glioma stem-like cells increased chemosensitivity, whereas over-expression of L1CAM in T98G cells and glioma spheroids diminished the apoptotic response. ElevatedL1CAM expression caused a diminished expression of caspase-8 in GBM and differentiated gliomastem-like cells. These data show that TGF-b1 dependent upregulation of L1CAM expression in GBM cellsleads to the downregulation of caspase-8 and apoptosis resistance pointing to L1CAM as potential targetfor improved therapy of GBM patients.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Gliomas represent the majority of primary brain tumors inadults and the most malignant form, glioblastoma multiforme(GBM), accounts for more than 50% of all intracranial tumors(Davies et al., 2001). Despite recent advances in surgery, radiother-apy and chemotherapy, current treatment regimens have only amarginal impact on patient survival (Nieder et al., 2006; Stuppet al., 2005). One of the primary reasons for this poor prognosisis the remarkably high chemoresistance of GBM. A betterunderstanding of the underlying mechanisms of acquired
chemoresistance is therefore critical for the development ofpotential strategies to overcome chemoresistance.
Interestingly, the neural adhesion molecule L1CAM (CD171) hasbeen found to play a pivotal role in tumorigenesis and chemoresis-tance (Raveh et al., 2009; Sebens Müerköster et al., 2007). L1CAMconsists of a large extracellular part, possessing six Ig-like and fivefibronectin-repeat III domains, which are linked to a short intracel-lular cytoplasmatic domain via a single transmembrane sequence(Schäfer and Altevogt, 2010). Two alternatively spliced exons havebeen identified. In addition, L1CAM can be proteolytically cleavedyielding a 200 kDa ectodomain (Maretzky et al., 2005). Aberrantexpression of L1CAM was documented in various tumors includingneuroblastomas and pancreatic adenocarcinomas (Bergmann et al.,2010; Raveh et al., 2009; Rawnaq et al., 2010), and being correlatedwith poor prognosis and more advanced stages of the disease (Benet al., 2010; Doberstein et al., 2011). For gliomatosis cerebri it wassuggested that L1CAM expression may play a role in the histogen-esis of this extensively infiltrating tumor (Suzuki et al., 2005).Additionally, aberrant expression of L1CAM has been shown topromote proliferation, motility and chemoresistance of cancer cells
0197-0186/$ - see front matter � 2012 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.neuint.2012.08.011
Abbreviations: bFGF, basic fibroblast growth factor; BSA, bovine serum albumin;DMEM, Dulbecco’s Modified Eagle Medium; DMSO, dimethylsulfoxide; EGF,epidermal growth factor; FCS, fetal calf serum; GAPDH, glyceraldehyde-3-phos-phate dehydrogenase; GBM, glioblastoma multiforme; GFAP, glial fibrillary acidicprotein; MAP2, microtubule-associated protein 2; MBP, myelin-basic protein; PBS,phosphate-buffered saline; qPCR, quantitative real-time PCR; SOX2, SRY (sexdetermining region Y)-box2; TGF-b1, transforming growth factor-b1.⇑ Corresponding author. Tel.: +49 (0)431 597 4901; fax: +49 (0)431 597 4941.
E-mail address: [email protected] (J. Held-Feindt).
Neurochemistry International 61 (2012) 1183–1191
Contents lists available at SciVerse ScienceDirect
Neurochemistry International
journal homepage: www.elsevier .com/locate /nci

Author's personal copy
from different tumor entities (Fogel et al., 2003; Gavert et al., 2005;Sebens Müerköster et al., 2007, 2009).
For glioblastomas, it was documented that L1CAM is expressedat high levels (Izumoto et al., 1996; Tsuzuki et al., 1998), mediatingmotility of tumor cells (Yang et al., 2009) via binding of the L1CAMectodomain to integrin receptors followed by activation of thefocal adhesion kinase (Yang et al., 2011). Moreover, L1CAM expres-sion in gliomas was correlated with high expression of the trans-forming growth factor (TGF)-b (Tsuzuki et al., 1998), a cytokinewhich plays a pivotal role in glioblastoma pathology (Golestanehand Mishra, 2005). Interestingly, it was shown that TGF-b1 is apotent inducer of L1CAM expression in human pancreatic ductalepithelial and pancreatic adenocarcinoma cells (Geismann et al.,2009), and that the binding of the transcription factor Slug isessential for TGF-b1-induced L1CAM expression in these cells(Geismann et al., 2011). Recent investigations documented thatL1CAM was differentially expressed in glioblastoma stem cellsrelative to non-stem tumor cells or normal neural progenitors(Bao et al., 2008; Cheng et al., 2011a). Targeting L1CAM using alentiviral-mediated short-hairpin RNA inhibited growth especiallyin glioblastoma stem cells (Bao et al., 2008), and L1CAM regulatesDNA damage checkpoint responses and radiosensitivity ofglioblastoma stem cells through nuclear translocation of theL1CAM intracellular domain (Cheng et al., 2011b).
These results already point to a pivotal role of L1CAM in thepathophysiology of glioblastomas. Nevertheless, up to now nothingis known about the impact of L1CAM on chemoresistance of thesetumors. Thus, the aim of the present study was to investigate inhigh and low L1CAM expressing glioma cells as well as in gliomastem-like cells, whether TGF-b1 is involved in the regulation ofL1CAM expression thereby mediating chemoresistance and to elu-cidate the underlying mechanism.
2. Materials and methods
2.1. Cell lines, tumor specimens and materials
The human glioblastoma cell lines U373, A172 and T98G wereobtained from the DKFZ, Heidelberg, Germany, and the pancreaticductal adenocarcinoma cell line Panc1 was donated by H. Kalthoff,Institute of Experimental Cancer Research, UKSH, Campus Kiel,Kiel, Germany. All cell lines were cultured as reported in Dul-becco’s Modified Eagle Medium (DMEM; Invitrogen, Karlsruhe,Germany) plus 10% fetal calf serum (FCS; Invitrogen; gliomas) orRPMI plus 10% FCS (Panc1) (Geismann et al., 2009, 2011; Held-Feindt and Mentlein, 2002; Held-Feindt et al., 2010; SebensMüerköster et al., 2007). The different cell lines were checked forpurity by immunostaining with cell type-specific markers and forthe absence of Mycoplasma contamination by staining with bis-benzimide as described before (Hattermann et al., 2010; Held-Feindt et al., 2010).
Glioma stem-like cells were generated from 5 glioblastoma pa-tients. From 4 patients (P1–P4), short-term glioma stem-like cellcultures were obtained by dissociation of fresh human glioblas-toma samples and subsequent cultivation in neurosphere medium(Reynolds and Weiss, 1992) plus 10 ng/ml basic fibroblast growthfactor (bFGF; PeproTech; Rocky Hill, NJ, USA) and 20 ng/ml epider-mal growth factor (EGF; ImmunoTools; Friesoythe, Germany)(Hattermann et al., 2010; Yuan et al., 2004). Developing gliomaspheroids were kept for six weeks with trypsinisation processevery second week. Afterwards short-term glioma stem-like cellcultures were divided and cultured for another three weeks –one part was subjected to differentiation (application of 10% FCS;Invitrogen; removing of bFGF and EGF), one part was continuouslycultured in neurosphere medium as described above. From onepatient, the glioma stem-like cell line 25/07 was generated by reg-
ular dissociation of a glioma stem-like cell culture over a period ofone year to enrich cells with self-renewal capacities (Hattermannet al., 2010). Glioma stem-like cells were characterized by the for-mation of neurospheres, the ability to survive and proliferate understem cell conditions and to differentiate into more mature cells(Hattermann et al., 2010), which was proven by qPCR and/or Wes-tern blots as described below. Materials and clinical data were ob-tained in accordance with the Helsinki Declaration of 1975 andwith approval of the ethics committee of the University of Kiel.The diagnosis was established by a neuropathologist. In case of suf-ficient material, matched probes of glioma stem-like cells/gliomacells lines were used for different experiments. Used reagents were:TGF-b1 (RELIATech, Wolfenbüttel, Germany) at 10 ng/ml, SB431542(Calbiochem, via Merck Biosciences, Schwalbach/Ts, Germany) at10 lM and temozolomide (Sigma–Aldrich, Taufkirchen, Germany)at 100 lg/ml or 400 lg/ml (adjusted to stock concentrations of20 mg/ml in dimethylsulfoxide (DMSO)). Stimulation with TGF-b1and SB431542 was performed for 24 h and 48 h in DMEM plus 10%FCS, respectively. Conditions for stimulation with temozolomidesee below (Annexin V/PI staining and caspase-3/7 assay).
2.2. Cell transfection
For L1CAM transfection T98G cells and 25/07 glioma stem-likecells (spheroids) were grown overnight in six or twelve well plates(density: 2 � 105 cells/well or 1 � 105 cells/well) and transfectedwith 20 ll/lg plasmid, Effectene transfection reagent plus 8 ll/lg plasmid enhancer (Qiagen, Hilden, Germany) and 0.4 lg/well(six well) or 0.3 lg/well (twelve well) of the following plasmids:pcDNA 3.1 (mock; Invitrogen) or pcDNA3.1-L1CAM (L1CAM)(Sebens Müerköster et al., 2007). After 8 h, medium was exchangedto DMEM plus 10% FCS (for T98G) or neurosphere medium withoutFCS (25/07 spheroids). For knock-down of L1CAM, A172 cells and25/07 glioma stem-like cells (differentiated cells) were seeded intotwelve well plates (density: 1 � 105 cells/well) followed by trans-fection with 8 ll HiperFect transfection reagent (Qiagen) and0.4 ll negative control siRNA low (Invitrogen) or L1CAM siRNA(siL1CAM; Invitrogen). In all transfection experiments, cells werecultured for additional 24 h to 48 h and transfection efficiencywas controlled for single experiments using qPCR and/or Westernblots (see below).
2.3. Flow cytometry
Cells (A172, T98G, U373 glioma cells; Panc1 cells as positivecontrol) grown in six well plates were detached, washed inphosphate-buffered saline (PBS), incubated with 1% bovine serumalbumin (BSA)/PBS for 30 min at room temperature, followed byincubation with the monoclonal mouse anti-L1CAM antibody L1–11A (subclone UJ127.11; 10 lg/ml in 1% BSA/PBS) (Mechtersheimeret al., 2001) or an isotype-matched control antibody (mousemonoclonal IgG1 10 lg/ml; R&D Systems, Wiesbaden, Germany)for 30 min. Cells were washed, incubated with a goat anti-mouseantibody conjugated with Alexa fluor 488 (Dianova, Hamburg,Germany) in 1% BSA/PBS (1:500) for 30 min, and analysed byflow cytometry (GalaxyArgon Plus, DakoCytomation, Hamburg,Germany) using the Flowmax software as described before (SebensMüerköster et al., 2007).
2.4. Annexin V/PI staining and caspase-3/7 assay
To determine apoptosis regulation after cytotoxic treatment,A172 and T98G glioma cells were stimulated with 100 lg/ml or400 lg/ml temozolomide or an equal volume of DMSO for 24 hand 48 h and the amount of annexin V positive cells as well asthe caspase-3/7 activity was measured. Additionally, L1CAM/mock
1184 J. Held-Feindt et al. / Neurochemistry International 61 (2012) 1183–1191

Author's personal copy
control transfected T98G and 25/07 cells (spheroids) as well assiL1CAM/sicontrol transfected A172 and 25/07 cells (differenti-ated) were stimulated 24 h after transfection with 400 lg/ml tem-ozolomide or DMSO for additional 24 h and caspase-3/7 activitywas measured. Moreover, both 25/07 and P1 untransfected cells(spheroids and differentiated) were chosen for caspase measure-ments. For annexin V/propidium iodide staining, cells were incu-bated with annexin V-FITC and the cell death discriminator(propidium iodide) from Biocarta (Hamburg Germany) and analysedby flow cytometry using the FLOMAX software. Caspase-3/7 activitywas measured using a homogeneous luminescent assay (Caspase-Glo™ 3/7 Assay; Promega, Mannheim, Germany) as describedelsewhere (Sebens Müerköster et al., 2006). To account for treatmentinduced alterations of cell numbers, cells that were not used in thecaspase-3/7 assay, were lysed and the protein content was deter-mined with the proteinassay Dc (Bio-rad, München, Germany).Caspase-3/7 activity was related to the protein content of the samesample and expressed as n-fold caspase-3/7 activity of basal.
2.5. Quantitative real-time PCR (qPCR)
Total RNA of A172, T98G, U373 and Panc1 (unstimulatedcontrols; A172 and T98G stimulated with SB431542 and TGF-b1;L1CAM/mock and siL1CAM/sicontrol transfected T98G and A172cells) was purified with TRIzol Reagent (Invitrogen), treatedwith RNase-free DNase (1 U/ll, Promega), and reversetranscribed by RevertAid™ H Minus M-MuLV Reverse Transcrip-tase (200 U/ll, Fermentas, Vilnius, Lithuania) as described before(Hattermann et al., 2010; Held-Feindt and Mentlein, 2002;Held-Feindt et al., 2010). Quantitative PCR was performed intriplicate using a total reaction volume of 20 ll, containing 1 llof 20� Assays-on-Demand™ Gene Expression Assay Mix(L1CAM, Hs_00240928_m1; TGFb-1, Hs_00171257_m1; caspase-8, Hs01018151_m1; Applied Biosystems, Foster City, CA), 10 ll of2� TaqMan Universal PCR Master Mix and 100 ng or 10 ng of cDNAtemplate (diluted in RNase-free water to 9 ll). After 2 min at 50 �Cand 10 min at 95 �C, 40 cycles of 15 s at 95 �C and 1 min at 60 �Cwere performed. Glyceraldehyde-3-phosphate dehydrogenase(GAPDH, Hs99999905_m1; Applied Biosystems) and TATA-Box binding protein (TBP, Hs_00920494_m1) mRNA was amplifiedin each sample as internal positive control. Each plateincluded at least three ‘‘No Template Controls (NTC)’’. The reactionwas carried out with the MyiQ™ Single Color Real-time PCRDetection System (Bio-Rad, Germany) and fluorescence datawere converted into CT measurements. DCT values of each samplewere calculated as CTgene of interest � CT GAPDH/TBP. DCT = 3.33corresponds to one order of magnitude. Low DCT values indicatehigh expression. Relative gene expression was calculated with2(normalized CT non-stimulated � normalized CT stimulated) = n-fold of control.Additionally, total RNA of 25/07 and P1 to P4 (spheroids,differentiated cells; L1CAM/mock and siL1CAM/sicontroltransfected 25/07 cells) was purified with the PicoPure RNAIsolation Kit (MDS Analytical Technologies, Sunnyvale, CA)according to the manufacturer’s instructions. DNase digestion,cDNA synthesis, and qPCR were performed as described above. Inaddition to L1CAM, TGF-b1 and caspase-8, GFAP (glial fibrillaryacidic protein; Hs_00157674_m1), MAP2 (microtubule-associatedprotein 2; Hs_00258900_m1); MBP (myelin-basic protein;Hs_00175245_m1); Nestin (Hs_00707120_s1) and SOX2 (SRY(sex determining region Y)-box2; Hs_00602736_m1) were ana-lyzed and DCT values determined as described above.
2.6. Western blotting
Cells (unstimulated controls; A172 and T98G stimulated withSB431542 and TGF-b1; L1CAM/mock and siL1CAM/sicontrol
transfected T98G and A172 cells) were lysed with 2� sodiumdodecyl sulfate (SDS) PAGE buffer (128 mmol/l Tris-Base, 4.6%SDS, 10% glycerol) as described before (Sebens Müerköster et al.,2007). Ten microgram of protein were dissolved in SDS–PAGE sam-ple buffer, fractionated by 4–20% SDS–PAGE (ProGel-Tris-glycin gel(Anamed, Darmstadt, Germany)) and immunoblotting was per-formed as described before (Arlt et al., 2001). Used primary anti-bodies were monoclonal mouse anti-L1CAM antibody L1–11A(subclone UJ127.11; 10 lg/ml) (Mechtersheimer et al., 2001), poly-clonal rabbit anti-HSP90 (Santa Cruz Biotechnology, CA, USA;1:1000) and a monoclonal mouse anti-caspase-8 (Cell SignalingTechnology, Billerica, MA, USA; 1:1000). Primary antibodies wereincubated overnight at 4 �C in TBST (50 mM Tris/HCl), pH 7.6,and 150 mM NaCl, 0.05% v/v Tween-20) supplemented with 5%(w/v) non-fat milk power. After extensive washing with Tween-20 in TBS, blots were exposed to the appropriate peroxidase-conju-gated secondary antibodies (Cell Signaling Technology) at 1:1000for 1 h at room temperature. After washing with TBST, blots weredeveloped with SuperSignal West Dura Extended Duration Sub-strate (Perbio Sciences, Bonn, Germany) and L1CAM signal inten-sity was determined in relation to HSP90 signal using aChemiDoc XRS Documentation system and the Quantity One4.5.0 software (both from Bio-Rad).
2.7. Statistical analysis
Values are given as means ± standard deviations (SD). Statisticalsignificance was analyzed by a two-tailed Student’s t-test.⁄p < 0.05, ⁄⁄p < 0.01, ⁄⁄⁄p < 0.001.
3. Results
3.1. L1CAM is expressed in glioblastoma cells and transcriptionallyregulated by TGF-b1
L1CAM expression has been already described in tumor tissuesof glioblastoma patients (Izumoto et al., 1996; Tsuzuki et al., 1998).Thus, in an initial step we measured the expression and transcrip-tional regulation of L1CAM in a panel of human glioblastoma celllines by qPCR, flow cytometry and Western blot analysis (Figs. 1and 2). As demonstrated by flow cytometry, the investigated glio-blastoma cell lines A172, T98G and U373 showed variable amountsof L1CAM – with highest L1CAM expression in A172 (similar to thepancreatic adenocarcinoma cell line Panc1, which served as a posi-tive control) and lowest expression in T98G cells. U373 glioblas-toma cells showed an intermediate L1CAM expression profile(Fig. 1). These findings were in accordance with qPCR results (datanot shown) revealing mean DCT values for L1CAM of �1.96 ± 0.3,10.2 ± 0.7, and 7.4 ± 0.5 in A172, T98G and U373 cells, respectively,and 4.62 ± 0.5 in Panc1 with low DCT (cycle threshold) values indi-cating high expression and DCT = 3.33 corresponding to one orderof magnitude. Protein expression was confirmed by Western blotdemonstrating high basal L1CAM expression in A172 and almostno basal L1CAM expression in T98G cell (unstimulated A172 andT98G samples; Fig. 2, right panels).
Since TGF-b1 has been shown to be a potent inducer of L1CAMexpression in tumor cells (Geismann et al., 2009, 2011), we nextanalysed whether L1CAM expression in the glioblastoma cell linesA172 and T98G is regulated by TGF-b1. Since A172 showed per sehigh L1CAM expression, these cells were treated with the ALK4/ALK5/ALK7 inhibitor SB431542, whereas T98G cells exhibitinglow L1CAM expression were stimulated with TGF-b1. Results forqPCR and Western blots are shown in Fig. 2. For both cell lines aclear TGF-b1 dependent regulation of L1CAM expression was mea-sured. In relation to unstimulated controls (control = 1),
J. Held-Feindt et al. / Neurochemistry International 61 (2012) 1183–1191 1185

Author's personal copy
application of SB431542 for 24 h and 48 h yielded a down-regula-tion of L1CAM mRNA to 0.54 ± 0.2-fold and 0.52 ± 0.1-fold(p < 0.05), respectively, in A172 cells (Fig. 2, upper panel left),and application of TGF-b1 for 24 h and 48 h led to an up-regulationof L1CAM mRNA expression up to 2.8 ± 0.5-fold (p < 0.01) and3.05 ± 0.1-fold (p < 0.05, respectively, in T98G cells (Fig. 2, lowerpanel, left). Same results were obtained by Western blot analysis.Determined L1CAM protein amounts – quantified densitometric-ally in relation to the house-keeping gene HSP90 – were down-regulated in A172 cells after SB431542 application and vice versainduced in T98G cells after TGF-b1 application (Fig. 2, right side).Thus, glioblastoma cell lines exhibit variable L1CAM expressionwhich is highly regulated by TGF-b1.
3.2. Glioma stem-like cell cultures show higher L1CAM and TGF-b1expression levels in differentiated cells than correspondingundifferentiated glioma spheroids
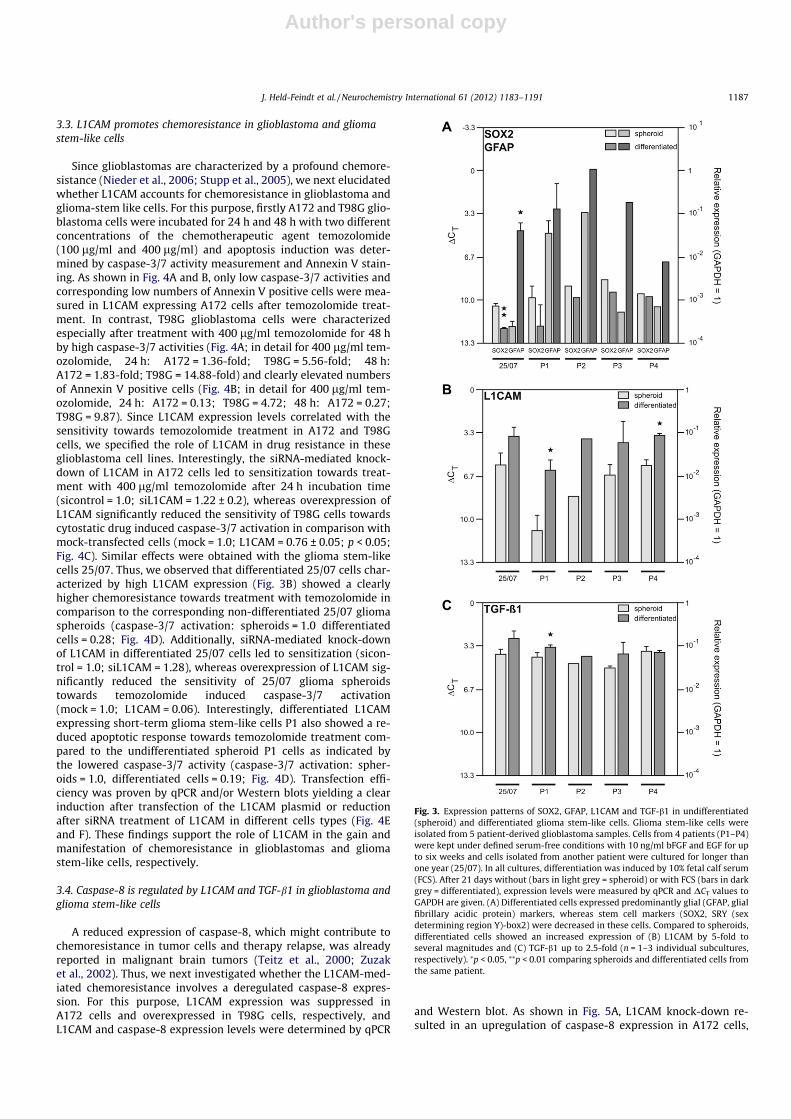
Since recent investigations documented that L1CAM was differ-entially expressed in glioblastoma stem cells relative to non-stemtumor cells or normal neural progenitors (Bao et al., 2008; Chenget al., 2011a), we examined whether L1CAM expression differedbetween undifferentiated glioma spheroids and corresponding dif-ferentiated cells. For this purpose, we used short-term gliomastem-like cell cultures from four patients (P1–P4) and the long-term glioma stem-like cell line 25/07 generated from another pa-tient (Hattermann et al., 2010). Differentiation was induced by10% FCS as described (Hattermann et al., 2010). In the absence ofFCS, all cultures showed the typical free floating spheroid morphol-ogy of stem-like cells. After application of 10% FCS, cell morphologychanged to an adherent, more differentiated phenotype. UsingqPCR we showed that the differentiation process led to a weak in-
crease of the neuronal marker MAP2 and the oligodendroglial mar-ker MBP (data not shown) and to a strong increase mainly of theastroglial marker GFAP (Fig. 3A) in all cell cultures. In contrast,the transcription of the stem cell marker Sox2 and Nestin was de-creased in differentiated cells compared to spheroid cells (Fig. 3A).
Interestingly, irrespective of the duration of the glioma stem-like cultures L1CAM expression was increased after the differenti-ation process in all differentiated cell cultures as determined byqPCR (Fig. 3B, DCT are shown, DCT = 3.33 corresponds to one orderof magnitude, and low DCT values indicate high expression).Accordingly, TGF-b1 expression was also higher in differentiatedglioma stem-like cells than in the corresponding spheroids(Fig. 3C, DCT are shown, DCT = 3.33 corresponds to one order ofmagnitude, and low DCT values indicate high expression). Giventhese DCT values, relative gene expression could be calculated with2(normalized CT non-stimulated � normalized CT stimulated) = n-fold of control.In detail, L1CAM relative gene expression was induced in the rangeof 4.6-fold (25/07) up to 25.1-fold (P1) and TGF-b1 expression wasinduced in the range of 1.7-fold (P1) up to 2.3-fold (25/07) (notshown). Thus, L1CAM and TGF-b1 are expressed in high amountsin differentiated cells arising from spheroid glioma stem-like cells.
Fig. 1. Expression of L1CAM in the glioblastoma cell lines A172, T98G and U373 aswell as the pancreatic adenocarcinoma cell line Panc1, used as positive control, wasanalysed by flow cytometry. A172, T98G and U373 showed variable amounts ofL1CAM – with highest L1CAM expression in A172 and lowest expression in T98Gcells (pale lines). To check binding specificity isotype control antibodies were used(dark lines).
Fig. 2. Transcriptional regulation of L1CAM by transforming growth factor (TGF)-b1. L1CAM expression was measured by qPCR (left sides; mean ± SD from n = 4experiments; unstimulated controls = 1) and Western blot (right sides; onerepresentative experiment from n = 3 is shown) in A172 (upper panel) and T98G(lower panel) glioblastoma cells. After 24 h to 48 h incubation time, 10 lM ALK4/ALK5/ALK7 inhibitor SB431542 mediated a down-regulation of L1CAM mRNA inhigh L1CAM expressing A172 cells, and application of 10 ng/ml TGF-b1 led to an up-regulation of L1CAM mRNA expression in low L1CAM expressing T98G cells. Sameresults were obtained by Western blot, at which signals were quantified densito-metrically in relation to the house-keeping gene HSP90. ⁄p < 0.05, ⁄⁄p < 0.01comparing untreated with treated cells.
1186 J. Held-Feindt et al. / Neurochemistry International 61 (2012) 1183–1191
Author's personal copy
3.3. L1CAM promotes chemoresistance in glioblastoma and gliomastem-like cells
Since glioblastomas are characterized by a profound chemore-sistance (Nieder et al., 2006; Stupp et al., 2005), we next elucidatedwhether L1CAM accounts for chemoresistance in glioblastoma andglioma-stem like cells. For this purpose, firstly A172 and T98G glio-blastoma cells were incubated for 24 h and 48 h with two differentconcentrations of the chemotherapeutic agent temozolomide(100 lg/ml and 400 lg/ml) and apoptosis induction was deter-mined by caspase-3/7 activity measurement and Annexin V stain-ing. As shown in Fig. 4A and B, only low caspase-3/7 activities andcorresponding low numbers of Annexin V positive cells were mea-sured in L1CAM expressing A172 cells after temozolomide treat-ment. In contrast, T98G glioblastoma cells were characterizedespecially after treatment with 400 lg/ml temozolomide for 48 hby high caspase-3/7 activities (Fig. 4A; in detail for 400 lg/ml tem-ozolomide, 24 h: A172 = 1.36-fold; T98G = 5.56-fold; 48 h:A172 = 1.83-fold; T98G = 14.88-fold) and clearly elevated numbersof Annexin V positive cells (Fig. 4B; in detail for 400 lg/ml tem-ozolomide, 24 h: A172 = 0.13; T98G = 4.72; 48 h: A172 = 0.27;T98G = 9.87). Since L1CAM expression levels correlated with thesensitivity towards temozolomide treatment in A172 and T98Gcells, we specified the role of L1CAM in drug resistance in theseglioblastoma cell lines. Interestingly, the siRNA-mediated knock-down of L1CAM in A172 cells led to sensitization towards treat-ment with 400 lg/ml temozolomide after 24 h incubation time(sicontrol = 1.0; siL1CAM = 1.22 ± 0.2), whereas overexpression ofL1CAM significantly reduced the sensitivity of T98G cells towardscytostatic drug induced caspase-3/7 activation in comparison withmock-transfected cells (mock = 1.0; L1CAM = 0.76 ± 0.05; p < 0.05;Fig. 4C). Similar effects were obtained with the glioma stem-likecells 25/07. Thus, we observed that differentiated 25/07 cells char-acterized by high L1CAM expression (Fig. 3B) showed a clearlyhigher chemoresistance towards treatment with temozolomide incomparison to the corresponding non-differentiated 25/07 gliomaspheroids (caspase-3/7 activation: spheroids = 1.0 differentiatedcells = 0.28; Fig. 4D). Additionally, siRNA-mediated knock-downof L1CAM in differentiated 25/07 cells led to sensitization (sicon-trol = 1.0; siL1CAM = 1.28), whereas overexpression of L1CAM sig-nificantly reduced the sensitivity of 25/07 glioma spheroidstowards temozolomide induced caspase-3/7 activation(mock = 1.0; L1CAM = 0.06). Interestingly, differentiated L1CAMexpressing short-term glioma stem-like cells P1 also showed a re-duced apoptotic response towards temozolomide treatment com-pared to the undifferentiated spheroid P1 cells as indicated bythe lowered caspase-3/7 activity (caspase-3/7 activation: spher-oids = 1.0, differentiated cells = 0.19; Fig. 4D). Transfection effi-ciency was proven by qPCR and/or Western blots yielding a clearinduction after transfection of the L1CAM plasmid or reductionafter siRNA treatment of L1CAM in different cells types (Fig. 4Eand F). These findings support the role of L1CAM in the gain andmanifestation of chemoresistance in glioblastomas and gliomastem-like cells, respectively.
3.4. Caspase-8 is regulated by L1CAM and TGF-b1 in glioblastoma andglioma stem-like cells
A reduced expression of caspase-8, which might contribute tochemoresistance in tumor cells and therapy relapse, was alreadyreported in malignant brain tumors (Teitz et al., 2000; Zuzaket al., 2002). Thus, we next investigated whether the L1CAM-med-iated chemoresistance involves a deregulated caspase-8 expres-sion. For this purpose, L1CAM expression was suppressed inA172 cells and overexpressed in T98G cells, respectively, andL1CAM and caspase-8 expression levels were determined by qPCR
and Western blot. As shown in Fig. 5A, L1CAM knock-down re-sulted in an upregulation of caspase-8 expression in A172 cells,
Fig. 3. Expression patterns of SOX2, GFAP, L1CAM and TGF-b1 in undifferentiated(spheroid) and differentiated glioma stem-like cells. Glioma stem-like cells wereisolated from 5 patient-derived glioblastoma samples. Cells from 4 patients (P1–P4)were kept under defined serum-free conditions with 10 ng/ml bFGF and EGF for upto six weeks and cells isolated from another patient were cultured for longer thanone year (25/07). In all cultures, differentiation was induced by 10% fetal calf serum(FCS). After 21 days without (bars in light grey = spheroid) or with FCS (bars in darkgrey = differentiated), expression levels were measured by qPCR and DCT values toGAPDH are given. (A) Differentiated cells expressed predominantly glial (GFAP, glialfibrillary acidic protein) markers, whereas stem cell markers (SOX2, SRY (sexdetermining region Y)-box2) were decreased in these cells. Compared to spheroids,differentiated cells showed an increased expression of (B) L1CAM by 5-fold toseveral magnitudes and (C) TGF-b1 up to 2.5-fold (n = 1–3 individual subcultures,respectively). ⁄p < 0.05, ⁄⁄p < 0.01 comparing spheroids and differentiated cells fromthe same patient.
J. Held-Feindt et al. / Neurochemistry International 61 (2012) 1183–1191 1187

Author's personal copy
whereas vice versa overexpression of L1CAM in T98G cells yielded adown-regulation of caspase-8. Interestingly, same results were ob-tained after inhibition of TGF-b1 signaling in A172 cells bySB431542, and after application of TGF-b1 in T98G cells, respec-
tively (Fig. 5B and C). In detail, after 24 h of treatment withSB431542 caspase-8 expression was increased to 1.4 ± 0.26-foldin A172 cells (control = 1), whereas caspase-8 expression levelwas clearly decreased in T98G cells after 24 h and 48 h TGF-b1
Fig. 4. L1CAM mediated effects on drug-induced apoptosis in the glioma cell lines A172 and T98G as well as in the glioma stem-like cells 25/07 and P1. Apoptosis induced by100 lg/ml or 400 lg/ml temozolomide in A172 and T98G cells as analyzed by quantification of (A) caspase-3/7 activity (n-fold of basal) and (B) number of Annexin V positivecells is clearly elevated in low L1CAM expressing T98G cells but not in A172 cells after 24 h and 48 h incubation time, respectively. (C) Temozolomide treatment (400 lg/ml,24 h) led to increased caspase-3/7 activity after knockdown of L1CAM (siL1CAM) in A172 cells while caspase-3/7 activity was decreased after overexpression of L1CAM(L1CAM) in T98G cells compared to individual controls (sicontrol and mock = 1), respectively (n = 3). (E) Transfection efficiency was proven by L1CAM Western blot (signalswere quantified densitometrically in relation to the house-keeping gene HSP90; control = 1). (D) Differentiated 25/07 and P1 cells characterized by high L1CAM expressionshowed a clearly lower caspase-3/7 activity after treatment with 400 lg/ml temozolomide for 24 h in comparison to 25/07 and P1 glioma spheroids, respectively.Additionally, temozolomide induced caspase-3/7 activity was decreased after overexpression of L1CAM (L1CAM) in 25/07 spheroids and increased after knockdown of L1CAM(siL1CAM) in differentiated 25/07 cells in relation to individual controls (mock and sicontrol = 1), respectively. (F) Transfection efficiency was proven by L1CAM and GAPDHqPCR (controls = 1). ⁄p < 0.05.
1188 J. Held-Feindt et al. / Neurochemistry International 61 (2012) 1183–1191

Author's personal copy
stimulation, respectively. Thus, in both cases expression of L1CAMand caspase-8 was inversely regulated by TGF-b1. To provewhether these results also apply to glioma stem-like cells, spher-oids and differentiated cells of 25/07 as well as of P1–P4 cultureswere analysed by qPCR for L1CAM (Fig. 3B), TGF-b1 (Fig. 3C) andcasapse-8 (Fig. 5D) mRNA expression levels. In all five cases, differ-entiation of glioma spheroids led to a clear up-regulation of L1CAMand TGF-b1 mRNA expression (Fig. 3B and C), while caspase-8mRNA expression was down-regulated in the range of 1.2-fold(P4) up to 7.9-fold (25/07). These data indicate that TGF-b1-medi-ated L1CAM expression contributes to chemoresistance, and thiseffect is accompanied by down-regulation of caspase-8 in glioblas-toma and glioma stem-like cells.
4. Discussion
Glioblastoma multiforme (GBM) is the most common primarybrain tumor. Even though advances in therapy have improvedthe quality of life for GBM patients they have had only little effecton extending survival following diagnosis of the disease. For this,with aggressive surgical resections using state-of-the-art preoper-ative and intraoperative neuroimaging, along with recent advancesin radiotherapy and chemotherapy, the prognosis for GBM patientsremains dismal: survival after diagnosis is about 1 year (Kanu
et al., 2009). The fundamental problem of these tumors is theirhighly infiltrative behavior and their pronounced resistance to-wards conventional radio- and/or chemotherapy. Hence, a betterunderstanding of the precise mechanisms leading to drug resis-tance is a prerequisite for substantial improvement of current ther-apeutical strategies.
In accordance with other groups (Izumoto et al., 1996; Ravehet al., 2009; Tsuzuki et al., 1998) we showed that the cell adhesionmolecule L1CAM is expressed in glioblastoma cell lines. Neverthe-less, in our experimental setting, different glioblastoma cell lineswere characterized by variable L1CAM amounts, e.g. A172 cellsexhibited high L1CAM expression whereas T98G cells only weaklyexpressed L1CAM. Since it has been shown that L1CAM expressioncan be transcriptionally regulated by the cytokine TGF-b1 (in pan-creatic ductal adenocarcinoma cells) (Geismann et al., 2009, 2011)and elevated L1CAM expression could be correlated with TGF-b1 ingliomas (Tsuzuki et al., 1998), we analysed the role of TGF-b1 inL1CAM expression in this tumor entity. The present study demon-strates a clear TGF-b1 dependency of L1CAM expression in glio-blastoma cells. Thus, L1CAM expression was markedly inducedby TGF-b1 in T98G cells whereas in A172 cells the ALK4/ALK5/ALK7 inhibitor SB431542 reduced L1CAM amounts. Regardingthe fact that overexpression or altered signaling of growth factors,including TGF-b1, is a common theme in glioblastomas (Kanu et al.,
Fig. 5. L1CAM and TGF-b1 mediated effects on caspase-8 expression in A172 and T98G glioma cell lines as well as in glioma stem-like cells. (A) After knockdown of L1CAM(siL1CAM) in A172 and overexpression of L1CAM (L1CAM) in T98G cells caspase-8 expression was induced/decreased in relation to individual controls (sicontrol andmock = 1), respectively. Protein integrity was proven by the house-keeping gene HSP90. One representative experiment out of three is shown. (B) Induction of caspase-8mRNA expression (and vice versa reduction of L1CAM, results given in Fig. 2, upper panel) was observed in A172 cells after treatment with 10 lM ALK4/ALK5/ALK7 inhibitorSB431542 for 24 h by qPCR (unstimulated controls = 1; n = 3). (C) In contrast, stimulation of T98G cells with 10 ng/ml TGF-b1 for 24–48 h yielded a L1CAM induction and viceversa caspase-8 reduction as analyzed by Western blot (protein integrity was proven by HSP90; n = 2). (D) Differentiated glioma stem-like cells (25/07 and P1–P4) werecharacterized by clearly lower caspase-8 mRNA expression compared to the respective undifferentiated (spheroid) cells proven by qPCR (glioma spheroids = 1). L1CAM andTGF-b1 mRNA expression determined in parallel are given in Fig. 3B and C.
J. Held-Feindt et al. / Neurochemistry International 61 (2012) 1183–1191 1189

Author's personal copy
2009), these results point overall to a pivotal role of the TGF-b1–L1CAM route in glioblastomas.
Interestingly, in a study with cancer stem cells isolated from gli-oma tumors and cell lines, L1CAM expression co-segregated withthe cancer stem cell population, and levels of L1CAM were higherin glioma stem cells than normal neural progenitors (Bao et al.,2008). Cheng et al. (2011a) described that several invasion-associated proteins including L1CAM, matrix metalloproteinase16, a disintegrin and a metalloproteinase with thrombospondinmotifs 1 and semaphorin3C were, relative to non-stem tumor cells,differentially expressed in glioma stem cells derived from glioblas-toma surgical specimens. Additionally, the group documented thatL1CAM expression was enriched in a population of cancer cells inthe invasive fronts of human primary glioblastomas. Nevertheless,a subpopulation of cancer cells inside the tumors was also L1CAMpositive. In our study we showed that L1CAM expression wassubstantially found in glioma-stem like cells, which were isolatedfrom freshly solid human glioblastomas. Thereby, in our handshighest L1CAM amounts were found in the differentiated cellsderived from the glioma spheroids, characterized by high GFAPand low SOX2 expression in contrast to glioma spheroids. Resultswere obtained by using short- and long-term cultured glioma-stemlike cells, respectively. Additionally, differentiated L1CAM express-ing glioma stem-like cells exhibited high TGF-b1 expression com-pared to corresponding glioma spheroids. However, despite thefact that all analysed patient-derived cells were characterized bya higher expression of TGF-b1 and L1CAM when differentiatedcompared to their corresponding glioma stem-like cells, variableexpression levels were noted among cells of the different patients.One the one hand this can be explained by differences in the basalxpression levels of these factors reflecting the heterogeneity oftumor cells within a tumor and more over among differentpatients. One the other hand it is important to note TGF-b1 isone important inducer of L1CAM but other factors are also ableto induce L1CAM expression, e.g. TNF-a, IL-1b. Thus, differencesin the L1CAM expression (basal as well as induced by differentia-tion) might rely on differences in the autocrine cytokine releaseby these cells.
To attach the importance of L1CAM expression to progression ofglioblastomas, several studies demonstrated a role for L1CAM indifferent processes of tumorigenesis. Thus, Cheng et al. (2011b)described recently that L1CAM regulates DNA damage checkpointresponses and enhances radioresistance of glioma stem cellsthrough nuclear translocation of L1CAM leading to NBS1-mediatedupregulation of c-Myc. In contrast, in matched non-stem tumorcells reduced L1CAM expression showed little or no effect oncheckpoint activation. Targeting L1CAM by RNA interference atten-uated DNA damage checkpoint activation and repair, and sensi-tized glioma stem cells to radiation (Cheng et al., 2011b). Baoet al. (2008) demonstrated that targeting of L1CAM resulted in sig-nificant growth inhibition in CD133+ glioblastoma cells, andL1CAM knock-down decreased expression of the transcription fac-tor Olig2 and upregulated the p21WAF1/CIP1 tumor suppressor. Addi-tionally, the suppression of L1CAM in such cells blocked theirtumorigenic capacity, indicating that L1CAM is a potential thera-peutic target in glioma cancer stem cells (Bao et al., 2008). Otherinvestigations focused on the impact of L1CAM on glioma cellmotility. For this, Yang et al. (2009, 2011) support a mechanismby which L1CAM increases human glioma cell migration and inva-sion by binding to integrin receptors, activating the focal adhesionkinase, and increasing turnover of focal complexes. Additionally,Izumoto et al. (1996) suggested that a short type of L1CAM, L1cs,is expressed on glioma cells and plays an important role in theadhesion and migration of these cells by hemophilic binding, andthat L1cs participates in tumor invasion along neuronal fibers. Incontrast to this, Senner et al. (2002) revealed increased adhesion
in the presence of L1CAM, whereas no L1CAM-dependent effectson proliferation or migration on matrigel or myelin were observed.
However, up to now a role for L1CAM in chemoresistance inglioblastomas has not been investigated. This study is now provid-ing evidence that L1CAM plays an important role in the mediationof drug resistance in gliomas.
By using L1CAM high and low expressing glioblastoma cellsknock-down and overexpression of L1CAM yielded a clearL1CAM-dependent apoptotic response towards the chemothera-peutic drug temozolomide. Same results were obtained in the pa-tient-derived glioma stem-like cells. Supporting these data,progression free survival time after adjuvant combined radiother-apy and chemotherapy with temozolomide (Stupp regimen) ofthe patients from which the short-term glioma stem-like cellswere isolated tended to correlate with the extent of L1CAM expres-sion (data not shown). It is important to note that the differenttherapeutic responses do not rely on differences in the methylationstatus of the O(6)-methylguanine-DNA methyltransferase (MGMT)gene because all patients exhibit an unmethylated (active) MGMTgene status. To draw valid conclusions whether the therapeutic re-sponse to temozolomide treatment correlates with L1CAM expres-sion, a larger cohort of patients has to be analysed which will bethe subject of a clinical study.
Furthermore, we showed that the TGF-b1 regulated L1CAMexpression results in a decreased caspase-8 expression whichmight contribute to the reduced apoptotic response towards tem-ozolomide treatment in glioblastoma cells. Thereby, differentiatedglioma cells being characterized by high L1CAM expressionshowed a clearly higher chemoresistance towards treatment withtemozolomide along with a diminished expression of TGF-b1 andcaspase-8 compared to corresponding glioma stem-like cells. A re-duced expression of caspase-8 in different brain tumors has beenalready reported previously (Teitz et al., 2000; Zuzak et al., 2002).
5. Conclusion
Together these data support the role of TGF-b1-induced L1CAMexpression in the mediation of drug resistance in glioblastoma cellsas it has been demonstrated recently in pancreatic ductal epithelialand carcinoma cells (Geismann et al., 2009; Sebens Müerkösteret al., 2007). Moreover, the observation of L1CAM-dependent mod-ulation of caspase-8 expression provides a plausible explanation ofhow L1CAM might contribute to drug resistance in glioblastomasand underscores the potential of L1CAM as therapeutic target toovercome chemoresistance in these highly malignant braintumors.
Acknowledgments
We thank B. Rehmke, M. Witt-Ramdohr, D. Leisner and J. Krausefor excellent technical assistance. This work was supported bygrants from the University of Kiel. The authors declare no conflictsof interest.
References
Arlt, A., Vorndamm, J., Breitenbroich, M., Fölsch, U.R., Kalthoff, H., Schmidt, W.E.,Schäfer, H., 2001. Inhibition of NF-kappaB sensitizes human pancreaticcarcinoma cells to apoptosis induced by etoposide (VP16) or doxorubicin.Oncogene 20, 859–868.
Bao, S., Wu, Q., Zhizhong, l., Satornsumetee, S., Wang, H., McLendon, R.E.,Hjelmeland, A.B., Rich, J.N., 2008. Targeting cancer stem cells through L1CAMsuppresses glioma growth. Cancer Res. 68, 6043–6048.
Ben, Q.W., Wan, J.C., Li, J., Zhu, Y., Yuan, F., Yao, W.Y., Yuan, Y.Z., 2010. Positiveexpression of L1-CAM is associated with perineural invasion and poor outcomein pancreatic ductal adenocarcinoma. Ann. Surg. Oncol. 17, 2213–2221.
Bergmann, F., Wandschneider, F., Sipos, B., Moldenhauer, G., Schniewind, B., Welsch,T., Schirrmacher, P., Klöppel, G., Altevogt, P., Schäfer, H., Sebens Müerköster, S.,
1190 J. Held-Feindt et al. / Neurochemistry International 61 (2012) 1183–1191

Author's personal copy
2010. Elevated L1CAM expression in precursor lesions and primary andmetastastic tissues of pancreatic ductal adenocarcinoma. Oncol. Rep. 24, 909–915.
Cheng, L., Wu, Q., Guryanova, O.A., Huang, Z., Huang, Q., Rich, J.N., Bao, S., 2011a.Elevated invasive potential of glioblastoma stem cells. Biochem. Biophys. Res.Commun. 406, 643–648.
Cheng, L., Wu, Q., Huang, Z., Guryanova, O.A., Huang, Q., Shou, W., Rich, J.N., Bao, S.,2011b. L1CAM regulates DNA damage checkpoint response of glioblastomastem cells through NSB1. EMBO J. 30, 800–813.
Davies, F.G., Kupelian, V., Freels, S., McCarthy, B., Surawicz, T., 2001. Prevalenceestimates for primary brain tumors in the United States by behavior and majorhistology groups. Neuro Oncol. 3, 152–158.
Doberstein, K., Wieland, A., Lee, S.B., Blaheta, R.A., Wedel, S., Moch, H., Schraml, P.,Pfeilschifter, J., Kristiansen, G., Gutwein, P., 2011. L1-CAM expression in ccRCCcorrelates with shorter patients survival times and confers chemoresistance inrenal cell carcinoma cells. Carcinogenesis 32, 262–270.
Fogel, M., Gutwein, P., Mechtersheimer, S., Riedle, S., Stoeck, A., Smirnov, A., Edler, L.,Ben-Arie, A., Huszar, M., Altevogt, P., 2003. L1 expression as a predictor ofprogression and survival in patients with uterine and ovarian carcinomas.Lancet 362, 869–875.
Gavert, N., Conacci-Sorrell, M., Gast, D., Schneider, A., Altevogt, P., Brabletz, T.,Ben-Ze’ev, A., 2005. L1, a novel target of beta-catenin signaling, transformscells and is expressed at the invasive front of colon cancers. J. Biol. Chem.168, 633–642.
Geismann, C., Morscheck, M., Koch, D., Bergmann, F., Ungefroren, H., Arlt, A., Tsao,M.-S., Bachem, M.G., Altevogt, P., Sipos, B., Fölsch, U.R., Schäfer, H., SebensMüerköster, S., 2009. Up-regulation of L1CAM in pancreatic ductal cells istransforming growth b1- and Slug-dependent: role in malignant transformationof pancreatic cancer. Cancer Res. 69, 4517–4526.
Geismann, C., Arlt, A., Bauer, I., Pfeiffer, M., Schirmer, U., Altevogt, P., SebensMüerköster, S., Schäfer, H., 2011. Binding of the transcription factor Slug to theL1CAM promoter is essential for transforming growth factor-b1 (TGFb)-inducedL1CAM expression in human pancreatic ductal adenocarcinoma cells. Int. J.Oncol. 38, 257–266.
Golestaneh, N., Mishra, B., 2005. TGF-beta, neuronal stem cells and glioblastoma.Oncogene 22, 5722–5730.
Hattermann, K., Held-Feindt, J., Lucius, R., Sebens Müerköster, S., Penfold, M.E.T.,Schall, T.J., Mentlein, R., 2010. The chemokine receptor CXCR7 is highlyexpressed in human glioma cells and mediates anti-apoptotic effects. CancerRes. 70, 3299–3308.
Held-Feindt, J., Mentlein, R., 2002. CD70/CD27 ligand, a member of the TNF family,is expressed in human brain tumors. Int. J. Cancer 98, 352–356.
Held-Feindt, J., Hattermann, K., Sebens Müerköster, S., Wedderkopp, H., Knerlich-Lukoschus, F., Ungefroren, H., Mehdorn, H.M., Mentlein, R., 2010. CX3CR1promotes recruitment of human glioma-infiltrating microglia/macrophages(GIMs). Exp. Cell Res. 316, 1553–1566.
Izumoto, S., Ohnishi, T., Arita, N., Hiraga, S., Taki, T., Hayakawa, T., 1996. Geneexpression of neural cell adhesion molecule L1 in malignant gliomas andbiological significance of L1 in glioma invasion. Cancer Res. 56, 1440–1444.
Kanu, O.O., Mehta, A., Di, C., Lin, N., Bortoff, K., Bigner, D.D., Yan, H., Adamson, D.C.,2009. Glioblastoma multiforme: a review of therapeutic targets. Expert Opin.Ther. Targets 13, 701–718.
Maretzky, T., Schulte, M., Ludwig, A., Rose-John, S., Blobel, C., Hartmann, D.,Altevogt, P., Saftig, P., Reiss, K., 2005. L1 is sequentially processed by twodifferently activated metalloproteases and presenilin/gamma-secretase andregulates neural cell adhesion, cell migration, and neurite outgrowth. Mol. CellBiol. 25, 9040–9053.
Mechtersheimer, S., Gutwein, P., Agmon-Levin, N., Stoeck, A., Oleszewski, M., Riedle,S., Postina, R., Fahrenholz, F., Fogel, M., Lemmon, V., Altevogt, P., 2001.
Ectodomain shedding of L1 adhesion molecule promotes cell migration byautocrine binding to integrins. J. Cell Biol. 155, 661–673.
Nieder, C., Adam, M., Grosu, A.L., 2006. Combined modality treatment ofglioblastoma multiforme: the role of temozolomide. Rev. Recent Clin. Trials 1,43–51.
Raveh, S., Gavert, N., Ben-Ze’ev, A., 2009. L1 cell adhesion molecule (L1CAM) ininvasive tumors. Cancer Lett. 282, 137–145.
Rawnaq, T., Quaas, A., Zander, H., Gros, S.J., Reichelt, U., Blessmann, M., Wilzcak, W.,Schachner, M., Sauter, G., Izbicki, J.R., Kaifi, J.T., 2010. L1 is Highly expressed intumors of the nervous system: a study of over 8000 human tissues. J. Surg. Rev.2010. http://dx.doi.org/10.1016/j.jss.2010.10.029.
Reynolds, B.A., Weiss, S., 1992. Generation of neurons and astrocytes from isolatedcells of the adult mammalian central nervous system. Science 255, 1707–1710.
Schäfer, M.K.E., Altevogt, P., 2010. L1CAM malfunction in the nervous system andhuman carcinomas. Cell. Mol. Life Sci. 67, 2423–2437.
Sebens Müerköster, S., Lust, J., Arlt, A., Hasler, R., Witt, M., Sebens, T., Schreiber, S.,Fölsch, U.R., Schäfer, H., 2006. Acquired chemoresistance in pancreaticcarcinoma cells: induced secretion of IL-1beta and NO lead to inactivation ofcaspases. Oncogene 25, 3973–3981.
Sebens Müerköster, S., Werbing, V., Sipos, B., Debus, M.A., Witt, M., Großmann, M.,Leisner, D., Kötteritzsch, J., Kappes, H., Klöppel, G., Altevogt, P., Fölsch, U.R.,Schäfer, H., 2007. Drug-induced expression of the cellular adhesion moleculeL1CAM confers anti-apoptotic protection and chemoresistance in pancreaticductal adenocarcinoma cells. Oncogene 26, 2759–2768.
Sebens Müerköster, S., Kötteritzsch, J., Geismann, C., Gast, D., Kruse, M.-L., Altevogt,P., Fölsch, U.R., Schäfer, H., 2009. A5-integrin is crucial for L1CAM-mediatedchemoresistance in pancreatic adenocarcinoma. Int. J. Oncol. 34, 243–253.
Senner, V., Kismann, E., Püttmann, S., Hoess, N., Baur, I., Paulus, W., 2002. L1expressed by glioma cells promotes adhesion but not migration. Glia 38, 146–154.
Stupp, R., Mason, W.P., van den Bent, M.J., Weller, M., Fisher, B., Taphoorn, M.J.,Belanger, K., Brandes, A.A., Marosi, C., Bogdahn, U., Curschmann, J., Janzer, R.C.,Ludwin, S.K., Allgeier, A., Lacombe, D., Cairncross, J.G., Eisenhauer, E.,Mirimanoff, R.O., 2005. Radiotherapy plus concomitant and adjuvanttemozolomide for glioblastomas. N. Engl. J. Med. 352, 987–996.
Suzuki, T., Izumoto, S., Fujimoto, Y., Maruno, M., Ito, Y., Yoshimine, T., 2005.Clinicopathological study of cellular proliferation and invasion in gliomatosiscerebri: important role of neural cell adhesion molecule L1 in tumour invasion.J. Clin. Pathol. 58, 166–171.
Teitz, T., Wei, T., Valentine, M.B., Vanin, E.F., Grenet, J., Valentine, V.A., Behm, F.G.,Look, A.T., Lahti, J.M., Kidd, V.J., 2000. Caspase 8 is deleted or silencedpreferentially in childhood neuroblastomas with amplification of MYCN. Nat.Med. 6, 529–535.
Tsuzuki, T., Izumoto, S., Ohnishi, T., Hiraga, S., Arita, N., Hayakawa, T., 1998. Neuralcell adhesion molecule L1 in gliomas: correlation with TGF-beta and p53. J. Clin.Pathol. 51, 13–17.
Yang, M., Adla, S., Temburni, M.K., Patel, V.P., Lagow, E.L., Brady, O.A., Tian, J., Boulos,M.I., Galileo, D.S., 2009. Stimulation of glioma cell motility by expression,proteolysis, and release of the L1 neural cell recognition molecule. Cancer CellInt. 29, 9–27.
Yang, M., Li, Y., Chilukuri, K., Brady, O.A., Boulos, M.I., Kappes, J.C., Galileo, D.S., 2011.L1 stimulation of human cell motility correlates with FAK activation. J.Neurooncol. 105, 27–44.
Yuan, X., Curtin, J., Xiong, Y., Liu, G., Waschsmann-Hogiu, S., Farkas, D.L., Black, K.L.,Yu, J.S., 2004. Isolation of cancer stem cells from adult glioblastoma multiforme.Oncogene 24, 9392–9400.
Zuzak, T.J., Steinhoff, D.F., Sutto, L.N., Phillip, P.C., Eggert, A., Grotzer, M.A., 2002. Lossof caspase-8 mRNA expression is common in childhood primitiveneuroectodermal brain tumor/medulloblastoma. Eur. J. Cancer 38, 83–91.
J. Held-Feindt et al. / Neurochemistry International 61 (2012) 1183–1191 1191






![[Autocrine IL-1beta secretion leads to NF-kappabeta-mediated chemoresistance in pancreatic carcinoma cells in vivo]](https://img.pdfslide.net/doc/110x75/635d10ad095e4caf22058aa8/autocrine-il-1beta-secretion-leads-to-nf-kappabeta-mediated-chemoresistance-in.jpg)






















