Embed Size (px)
Citation preview

Linköping University Medical Dissertations No. 1244
The Norovirus PuzzleCharacterization of human and bovine norovirus
susceptibility patterns
Malin Vildevall
Department of Clinical and Experimental MedicineLinköping University, Sweden
Linköping 2011

© Malin Vildevall, 2011Division of Molecular Virology
Department of Clinical andExperimental MedicineFaculty of Health SciencesLinköping University
Cover: A puzzle illustration by Rada Chakarova.
Published articles have been printed with the permission of the copyright holder. Printed by LIU-Tryck, Linköping, Sweden, 2011
ISBN 978-91-7393-181-6ISSN 0345-0082

To my Children,
You make me realize that everything is
possible
Tonny,
I never could have made it without you
Keep your eyes on the stars but keep your
feet on the ground.
Theodore Roosevelt


Table of contentsAbstract .................................................................................................1
Populärvetenskaplig sammanfattning................................................3
Ett Norovirus Pussel- studier på humana och bovina norovirus och deras koppling till individers mottaglighet för sjukdom .................................. .3
Abbreviations........................................................................................6
List of papers ........................................................................................9
Introduction ........................................................................................ .11
Background .........................................................................................12
Human Noroviruses ...........................................................................12
History ...................................................................................................12Classification and structure ...................................................................13Virus like particles ..................................................................................15Epidemiology and Norovirus seasonality .............................................. .16Symptoms and Transmission of norovirus ............................................ .17Pathogenesis of Norovirus infection ..................................................... .18
Why are some people resistant to winter vomiting disease? ........20
Histo blood group antigens ....................................................................20Secretor status ......................................................................................21Susceptibility to disease ........................................................................23Norovirus Immunology and immune evasion ....................................... .25Norovirus evolution ................................................................................27Treatment, prevention and vaccine studies .......................................... .29
Bovine noroviruses ............................................................................31
Introduction ............................................................................................31Pathogenesis of GIII.2 infection in calves ............................................. .32Zoonotic transmission, are humans at risk? ......................................... .33

What is the receptor for bovine norovirus ............................................. .34
Aim of the Thesis ................................................................................37
Methods ...............................................................................................38
ELISA .....................................................................................................38Hemagglutination Assay ........................................................................39Hemagglutination Inhibition assay ........................................................ .40Pyrosequencing .....................................................................................40Western Blot ..........................................................................................42
Results and Discussion .....................................................................43
Paper I- Antibody Prevalence and Titer to Norovirus (Genogroup II) Cor-relate with Secretor (FUT2) but Not with ABO Phenotype or Lewis (FUT3) Genotype ...............................................................................................43Paper II- Genetic Susceptibility to Symptomatic Norovirus Infection in Ni-caragua ..................................................................................................45Paper III- Human Antibody Responses to Bovine (Newbury-2) Norovirus (GIII.2) and association to Histo-Blood Group Antigens ....................... .47Paper IV- Characterization of the bovine norovirus Hemagglutinin ...... .49
Concluding Remarks..........................................................................53
Acknowledgements ............................................................................56
References ..........................................................................................58

1
Abstract
AbstractWinter vomiting disease is caused by norovirus (NoV) and affects millions of people every year resulting in 200.000 deaths among children in developing countries. It was observed early that not all individuals exposed to the norovirus became ill. The reason for this is now recognized to be dependent upon the secretor status of an individual. The secretor status determines the ability of an individual to express histo-blood group antigens (HBGA) on mucosa and in saliva. A non-secretor is unable to express HBGAs due to a mutation in a gene called FUT2. In this thesis, I have investigated the antibody prevalence and titer in humans in Sweden and Nicaragua to the most common GII NoV and the correlation to secretor status, Lewis status and ABO. I found that secretors had significantly higher antibody prevalence and titer to GII NoV than non-secretors sug-gesting that non-secretors are less prone to be infected by the GII NoV. In Nicaragua, I also found several different NoV strains circulating at the same time. The NoVs have been circulating and evolving in the human population for some time and the same in-dividuals seems to be infected over and over again with the same virus. This suggests that there is no long-term immunity present but possibly short-term immunity, which would make it very difficult to produce a vaccine against NoV. However, recent studies have shown the possibility of using virus like particles as a vaccine candidate and have demonstrated long-term immunity.
The bovine NoV (boNoV) cause gastroenteritis in cattle and are closely related to the human NoV. The possibility of zoonotic transfer to humans is currently being investi-gated. I found that 26% of Swedish blood donors have antibodies to the boNoV sug-gesting that they have been exposed to the virus. The human NoV has been observed to be able to infect and cause disease in cattle, could the boNoV do the same in humans? To date, no boNoV strain has been found in humans. The proposed receptor structure for boNoV is the αGal epitope, which is present in many mammals like cow, pig, horse, sheep and rabbit but not in humans. This indicates that humans are not at risk for boNoV infection because we lack the proper receptor structure. However, recombinations be-tween different NoV strains have been demonstrated and the possibility of more than one receptor being present has been suggested. I found that aa position 365-379 on the boNoV capsid seems to be important for binding to erythrocytes. In this thesis, I hope to add some new pieces to the Norovirus Puzzle.

2
Populärvetenskaplig sammanfattning

3
Populärvetenskaplig sammanfattning
Populärvetenskaplig sammanfattning
Ett Norovirus Pussel- studier på humana och bovina norovirus och deras koppling till individers mottaglighet för sjukdom
Varje år drabbas vi i Sverige av vinterkräksjukan som ofta ger upphov till stora utbrott på förskolor, skolor, äldreboenden och sjukhus. Sjukdomen har ett snabbt förlopp med symptom som kraftiga kaskadkräkningar, diarre, feber och illamående. Orsaken till smittan är ett virus som kallas norovirus. Norovirus kan delas upp i olika undergrupper och i varje undergrupp finns ett flertal olika virustyper som alla ger upphov till vinter-kräksjukan.
Flera tidiga volontärstudier har visat att en del personer tycks vara immuna mot infek-tion med norovirus. En studie visade att när man infekterade 12 individer med norovirus blev 6 st sjuka men 6 st var fortsatt symptomfria. Efter 1 år gav man samma 12 individer samma norovirus igen och samma 6 personer blev sjuka en andra gång. Detta tydde på att vissa personer är resistenta mot norovirus infektion trots upprepad exponering av viruset. Andra personer tycktes kunna drabbas av samma virus flera gånger om och man drog då slutsatsen att dessa personer inte hade fått ett skyddande immunförsvar mot detta virus. Studier av detta slag var starten på att hitta anledningen till att inte alla blev sjuka trots att de utsatts för norovirus. Man började leta hos individerna för att hitta vad som skiljer dem åt. Man fann att vissa personer tycktes sakna förmågan att uttrycka kor-rekta receptorer som viruset behöver för att kunna infektera celler i människokroppen. Dessa receptorer skall normal sett uppvisas på cellernas yta och behövs för att viruset skall kunna binda till och ta sig in i celler och orsaka sjukdom.
På senare år har man visat att uppvisandet av dessa receptorer är beroende av ett enzym som kodas av en gen kallad FUT2 som finns på kromosom 19. Om denna gen har muta-tioner så kan inte enzymet uttryckas och därmed kan inte korrekta receptorer uttryckas på cellens yta. Individer som har denna mutation i dubbel uppsättning, en muterad gen från vardera förälder, kallas för sekretornegativa, och kan inte uttrycka dessa recep-torer i till exempel saliv och på tarmceller. Cirka 20% av sveriges befolkning är sekre-tornegativa och övriga 80% är sekretorpositiva och har då minst en FUT2 gen som fungerar. Man har på senare år upptäckt att vissa virusgrupper inom norovirus faktiskt

4
Populärvetenskaplig sammanfattning
kan infektera även sekretornegativa individer. Så numer kan man inte säga att de har ett totalt skydd mot vinterkräksjukan även om de virus som kan infektera dem tycks vara betydligt färre än de som främst drabbar sekretorpositiva individer.
Det är inte bara människor som drabbas av norovirus infektioner utan även grisar, kor och möss kan bli infekterade. Kor som blir infekterade med norovirus blir det av ett bo-vint norovirus som är mycket likt människans norovirus men som skiljer sig åt på flera sätt, bland annat vilken receptor viruset binder till. Det bovina noroviruset tillhör en egen grupp inom norovirus men är som sagt närbesläktad med det norovirus som drab-bar människan. Kalvar som infekteras med bovint norovirus upplever symtom liknande människans med framför allt diarrer. Man har kunnat visa att människor har antikroppar mot detta bovina norovirus och detta gör att man frågat sig om det finns möjlighet för det bovina noroviruset att sprida sig till människor? Hitills har man inte kunnat påvisa några direkta bevis för att så är fallet men man har sett att det virus som vanligen drab-bar människan också kan infektera och orsaka sjukdom hos både grisar och kalvar. Det finns nu flera studier som visar på att det bovina noroviruset binder till en receptor som inte finns hos människan. Detta borde i så fall göra det omöjligt för det bovina noro-viruset att infektera människor. Dock är det inte alltid så lätt och flera bitar av pusslet saknas fortfarande vad gäller detta. Kanske kan det bovina noroviruset använda sig av mer än en receptor vilket är vanligt för flera olika virusfamiljer. Mer studier behövs för att vi ska kunna avgöra om det finns möjlighet för människor att drabbas av norovirus infektioner från kalvar.
I mina studier som jag presenterar i denna avhandling har jag undersökt bland annat förekomsten av antikroppar mot humant norovirus bland Svenska blodgivare. Jag har också studerat på kopplingen mellan mängden antikroppar och sekretorstatus hos indi-viderna. Jag fann att de individer som var sekretornegativa hade betydligt lägre mängd antikroppar mot norovirus än de sekretorpositiva individerna. Detta tyder på att sekre-torpositiva individer är mer utsatta för infektion med norovirus än sekretornegativa in-divider.
Jag har också tittat på hur förekomsten av sekretornegativa/postiva individer ser ut i Ni-caragua och om det finns kopplingar till resistens mot norovirus sjukdom. I Nicaragua tittade jag på både patienter som hade kräksjuka och friska kontroller. Jag fann att pati-

5
Populärvetenskaplig sammanfattning
enterna var infekterade med flera olika sorters norovirus och att de hade höga mängder antikroppar mor norovirus. De sekretorpositiva individerna hade mer norovirus antik-roppar än de sekretornegativa individerna. I Nicaragua var endast 6% sekretornegativa jämfört med ca. 20% i Sverige. Ingen av de sjuka patienterna var sekretornegativ vilket tyder på att sekretornegativa kan vara skyddade mot sjukdom med norovirus.
I de två senare studierna har jag tittat på det bovina noroviruset och dess koppling till människan. Jag har tittat på förekomst av antikroppar mot bovint norovirus bland svenska blodgivare och fann att ca 26% hade antikroppar. Jag fann en koppling till sekretorstatus som tydde på att sekretorpositiva är mer troliga att infekteras av ett bo-vint norovirus än sekretornegativa. Vidare har jag också tittat på vilken receptor bovint norovirus binder då den tycks skilja sig åt från receptorn för humana norovirus. Jag har också kartlagt platsen på norovirusets yta där den sannolikt binder in till denna receptor.
Sammanfattningsvis kan jag utifrån mina studier se en stark koppling till sekretorstatus vad gäller infektion med den vanligaste humana norovirus stammen. Hurvida männi-skan i framtiden kan bli infekterad av den bovina norovirus stammen behöver utredas ytterligare och svaret på den frågan kommer ge ytterligare bitar i norovirus pusslet.

6
Abbreviations
Abbreviations
Aa amino acid(s)Ab/abs antibody/antibodiesAGE acute gastroenteritisASC antibody secreting cellsboNoV bovine norovirusBSA bovine serum albuminDC dendritic cellsdNTP deoxribonucleotide triphosphateELISA enzyme-linked immunosorbent assayEM electron microscopyFCV feline CalicivirusFuc fucoseFucT-II a1, 2-fucosyltransferase 2FucT-III a1, 3/4-fucosyltransferase 3FUT2 fucosyltransferase 2, the gene enconding FucT-IIFUT3 fucosyltransferase 3, the (Lewis) gene enconding FucT-IIIGal GalactoseGalNAc N-acetylgalactoseamineGI/II/III/IVIV genogroup I, II, III, IV, VGGTA1 gene encoding a1, 3 galactosyltransferaseGlc GlucoseGlcNAc N-acetylglucoseamineGn gnotobiotich hourHA hemagglutination assayHAI hemagglutination inhibition assayHBGA(s) histo blood group antigen(s)HRP horseradish peroxidasehuNoV human norovirusIFN interferonLea/Leb Lewis a/Lewis bMNV murine norovirus (GV)

7
Abbreviations
min minutesmut mutantNoV norovirusNTA N-terminal armNV norwalk virusOD optical densityORF open reading framePBS phosphate buffered salineP region protruding regionPPi pyrophosphateRBC red blood cellRdRp RNA dependent RNA polymeraseRHDV rabbit haemorrhagic disease virusRNA ribonucleic acidRT room temperatureSDS sodium dodecyl sulfateSMV snow mountain virus (GII.2)ssRNA single stranded RNA S region shell regionTh T helperTNF tumor necrosis factorVLP virus like particlesVPg viral-protein genome-linkedWHO world health organizationwt wildtype

8
Abbreviations

9
List of papers
List of papers
Antibody Prevalence and Titer to Norovirus (Genogroup II) Correlate with Secre-tor (FUT2) but not with ABO Phenotype or Lewis (FUT3) Genotype.Larsson M. M, Rydell G, Grahn A, Rodríguez-Díaz J, Åkerlind B, Hutson MA, Estes MK, Larson G and Svensson L. Journal of Infectious Diseases 2006;194:1422-7
Genetic Susceptibility to Symtomatic Norovirus Infection in Nicaragua.Bucardo F, Kindberg E, Paniagua M, Granh A, Larson G, Vildevall M and Svensson L. Journal of Medical Virology 2009;81:728-735
Human Antibody Responses to Bovine (Newbury-2) Norovirus (GIII.2) and Asso-ciation to Histo-Blood Group Antigens.Vildevall M, Grahn A, Oliver SL, Bridger JC, Charpilienne A, Poncet D, Larson G and Svensson L. Journal of Medical Virology 2010;82:1241-1246
Characterization of the Bovine Norovirus Hemagglutinin.Vildevall M, Charpilienne A, Hinkula J, Nasir W, Larson G and Svensson L. In manu-script

10
List of papers

11
Introduction
IntroductionMany of us are familiar with the winter vomiting disease either by experience or through media. The winter vomiting disease is caused by a virus called norovirus (NoV), which is only one of the pathogens causing acute gastroenteritis (AGE) together with bacteria, toxins, parasites and other viruses. The name winter vomiting disease was proposed in 1929 by Zahorsky and coworkers to describe the increase of AGE during the winter months (Zahorsky, 1929). Since then much work has been made to further investigate the role of viruses in AGE. Only in the last two decades a lot of new information has been added to the puzzle. One example is the role of host genetics in relation to NoV ill-ness. Why do some individuals never become sick from NoV infection while others get sick over and over again? The answer to this question seems to lie in the possible genetic pre-disposition to NoV infection. The secretor-status of an individual is now suggested to be the marker for susceptibility to symptomatic NoV illness. However, exceptions have already been reported for several NoV strains and further gives yet another piece of the puzzle.
In recent years the possibility of zoonotic infections from animal to human with bovine norovirus (boNoV) strains has just begun to be investigated. The boNoV infects cattle and cause gastroenteritis. Two studies have demonstrated that humans have antibod-ies (ab) to the boNoV (Vildevall et al., 2010; Widdowson et al., 2004) but what does this mean? So far there is no evidence showing symptomatic infection of humans with bovine NoV. However, studies have also shown that human NoV can infect and trigger disease in cattle. Since this is possible, is it also possible for humans to be infected with the boNoV and what would be the impact of such a zoonotic transmission?
These are just a few questions that need answers and I will address them in this thesis. Some new pieces can be added to the Norovirus Puzzle but many more are needed to complete the puzzle and allow us to see the whole picture.

12
Background
Background
Human Noroviruses
HistoryIn October 1968, 50% of the students and teachers of an elementary school in Norwalk, Ohio, USA developed AGE (Adler and Zickl, 1969). No bacteriological agent could be found and the search to find the etiologic agent began. In 1972, Albert Kapikian us-ing electron microscopy (EM) showed aggregates of 27nm virus particles, which after further investigations were found to be the etiological agent of the Norwalk outbreak (Figure 1). The Norwalk virus (NV) was hence discovered (Kapikian et al., 1972) and was the first virus to be associated with gastroenteritis. During the 1970s other viruses causing gastroenteritis namely rotavirus, enteric adenovirus and astrovirus were identi-fied (Bishop et al., 1973; Kurtz et al., 1977; Richmond et al., 1979). The NV has now been placed into the norovirus genus.
Figure 1. Human NoV particles with a ”star of david” morphology seen by transmission electron microscopy. The virus was visualized by negative staining using phosphotungstic acid. (Photo,
L. Svensson).
Today, NoVs are considered to be the most common cause of acute viral gastroenteritis world wide (Atmar and Estes, 2006) causing approximately 23 million episodes of AGE

13
Human Noroviruses
annually in the US (Mead et al., 1999). The world health organization (WHO) estimates that almost 2 million deaths in children less then 5 years of age are due to diarrhea (Boschi-Pinto et al., 2008; Bryce et al., 2005) and NoVs are now considered to be the second most common cause of severe AGE in children, after Rotavirus (Patel et al., 2008). NoVs are estimated to be responsible for nearly 200.000 deaths among children less then 5 years of age in developing countries (Patel et al., 2008).
Classification and structureNoVs belongs to the family of caliciviridae together with sapovirus, lagovirus, vesi-virus and the newly suggested groups of becovirus or nabovirus (Oliver et al., 2006a) and recoviruses (Farkas et al., 2008) (figure 2). NoVs and sapoviruses cause AGE in humans while lagoviruses includes the rabbit hemorrhagic disease virus (RHDV) which cause severe infection in rabbits often with a fatal outcome (Ohlinger et al., 1990). The vesivirus genus includes viruses infecting pigs and cats (Thiel and Konig, 1999), and the recently suggested nabovirus cause gastroenteritis in cattle (Oliver et al., 2006a). The recovirus (rhesus enteric calicivirus) has been detected in rhesus macaques but the pathogenicity remains to be investigated (Farkas et al., 2008).
SMSV1
GIII.1
GI.3GI.1GI.4
GI.5
GI.2
GV.1
GIV.1
GII.1GII.5
GII.4GII.2
GII.6
GII.
3
ReptilePan-1
VESV
SMSV17
FCV
CaCV
EBHV
RHDV
NB
0.1
TV
GV.1
GIII.2
GIII.1
GI.8GI.7
GI.3
GII.3
GII
GII.7
GII.14
GII.9G
II.8 GII.
6G
II.17
GII.
13 GII.
10
GII.5
GII.2
GII.16GII.12
GII.1GII.18
GII.19
GII.11Bristol (GII.4)
GII.15
GIV.2
GIV.1GI.2
GI.5
GI.4G
I.6N
orw
alk
(GI.1
)
Vesivirus
Sapovirus
Lagovirus
Nebovirus
Recovirus(proposed)
GV.1
GIII.2
GIII.1
GI.8GI.7
GI.3
GII.3
GII
GII.7
GII.14
GII.9G
II.8 GII.
6G
II.17
GII.
13 GII.
10
GII.5
GII.2
GII.16GII.12
GII.1GII.18
GII.19
GII.11Bristol (GII.4)
GII.15
GIV.2
GIV.1GI.2
GI.5
GI.4G
I.6N
orw
alk
(GI.1
)
Norovirus Norovirus
Figure 2. Classification of the caliciviridae family. Caliciviridae consists of 6 different genera of which the norovirus and sapovirus can infect humans. The norovirus genus can be further divided into 5 genogroups where GI, GII and GIV infect humans, GIII infect cattle and GV infect mice. (modified from Prasad., 2010).

14
Human Noroviruses
The NoV genus can then be further divided into five genogroups (GI-GV). GI, II and IV contain viruses that can infect humans. GIII is made up of boNoV and GV murine NoV. In GII, we can also find some viruses that have been detected in healthy pigs (Sugieda and Nakajima, 2002) while GIV NoV has been found in lions (Martella et al., 2007) and dogs (Martella et al., 2008). The NoV genogroups can then be further divided into several genotypes based on sequence diversity. At least 8 genotypes have been described for GI and 19 genotypes or more for GII (Bucardo et al., 2008; Zheng et al., 2006) (Figure 2).
For human infection the most common genotypes found are GI and GII and among these the GII.4 is the most predominant and is also considered to be a globally dominat-ing strain (Blanton et al., 2006; Lopman et al., 2004a; Noel et al., 1999; Widdowson et al., 2004). The GII.4 strain has been estimated to cause up to 80% of all NoV outbreaks (Ramirez et al., 2009) and it was recently reported that GII NoV were found in 86% of small children (median age 17 months) and that the GII.4 was the most prominent strain (Huhti et al., 2011).
The NoVs are small, non-enveloped, icosahedral viruses of approximately 27-38 nm in diameter. The NoVs genome is composed of a positive sense ssRNA (single strand-ed ribonucleic acid) approximately 7.7kb in length. The RNA has three open reading frames (ORFs) encoding the different structural and non-structural proteins. The ORF1 encodes a polyprotein that is cleaved by the viral protease 3CLpro into at least six non-structural proteins, such as the RNA-dependent RNA polymerase (RdRp) and the VPg. ORF2 encodes the capsid protein consisting of the shell (S) region and the protruding (P) region that can be further divided into P1 and P2. The inner S region is involved in forming the icosahedral shell and the P1 and P2 increase stability of the capsid and the hyper variable protruding P2 region is believed to play an important role in receptor binding and immune reactivity. ORF 3 encodes a basic structural protein that may have a role in the packaging of the genome into new virions (Figure 3). The 3´end of the ge-nome contains a poly A tail (Xi et al., 1990) and at the 5´end, the RNA is linked to the viral protein VPg that provides a cap. The N-terminal arms (NTA) of the subunits that form the capsid architecture are involved in a network of interactions possibly associ-ated with the capsid assembly pathway (Prasad, 2010).

15
Human Noroviruses
Figure 3. schematic overview of the NoV particle. The NoV capsid is constructed from the inner shell (S) region and the outer protruding (P) regions. Organisation of the ssRNA of NoV into 3 open reading frames where ORF 1 encodes a polyprotein cleaved by 3CLpro into at least six non-structural proteins. ORF2 encodes the capsid gene with the S and P domains and ORF 3 encodes
a minor strucural protein. NTA: N-terminal arm. (Prasad B.V et al., 1999, with permission).
Virus like particlesSince there is no reliable cell culture system or small animal model available at present for human NoV, virus like particles (VLPs) are commonly used. VLPs constituting the capsid protein can be expressed in yeast or in baculovirus systems and spontaneously self-assemble into a conformation that is similar in morphological and immunological characteristics as authentic NoVs (Han et al., 2005; Jiang et al., 1992). More recently, only the P-dimer of the capsid was expressed in E-coli, yeast or baculovirus and forms P-dimers and P-particles. They show the same binding properties as the slightly bigger VLPs and can also be used instead of authentic NoV particles (Tan et al., 2008a; Tan et al., 2004a; Tan and Jiang, 2005; Tan et al., 2004b).
P2
P1
SNTA
hinge
Nterm NTPase p20 VPg Pro Pol ORF2
Shell domain
ORF3
1 225 279 405 530
Non-structural polyprotein VP1 Capsid VP2
P1 P2 subdomain P1
hinge
16
Human Noroviruses
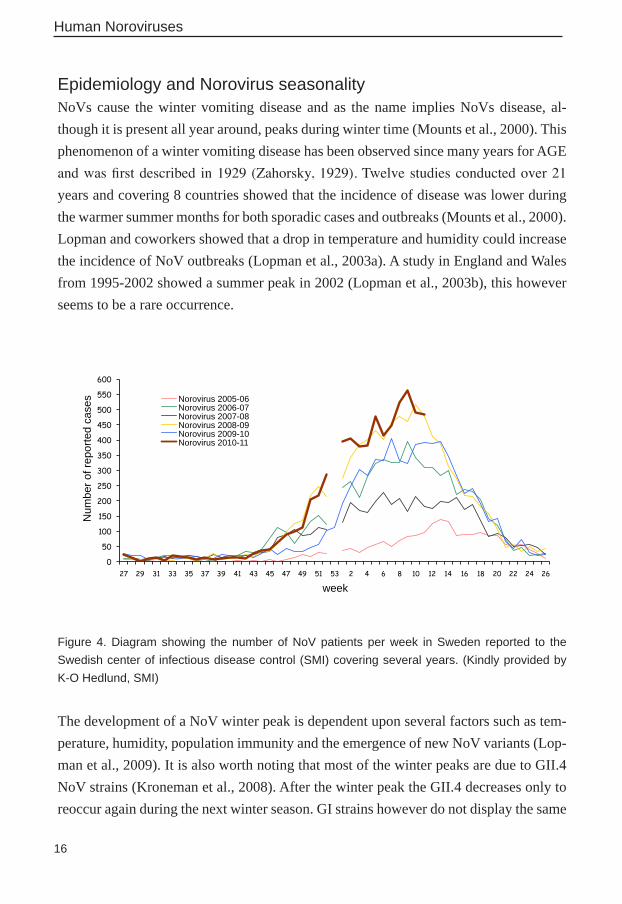
Epidemiology and Norovirus seasonalityNoVs cause the winter vomiting disease and as the name implies NoVs disease, al-though it is present all year around, peaks during winter time (Mounts et al., 2000). This phenomenon of a winter vomiting disease has been observed since many years for AGE and was first described in 1929 (Zahorsky, 1929). Twelve studies conducted over 21 years and covering 8 countries showed that the incidence of disease was lower during the warmer summer months for both sporadic cases and outbreaks (Mounts et al., 2000). Lopman and coworkers showed that a drop in temperature and humidity could increase the incidence of NoV outbreaks (Lopman et al., 2003a). A study in England and Wales from 1995-2002 showed a summer peak in 2002 (Lopman et al., 2003b), this however seems to be a rare occurrence.
0
50
100
150
200
250
300
350
400
450
500
550
600
27 29 31 33 35 37 39 41 43 45 47 49 51 53 2 4 6 8 10 12 14 16 18 20 22 24 26
Num
ber o
f rep
orte
d ca
ses
week
Norovirus 2005-06 Norovirus 2006-07 Norovirus 2007-08 Norovirus 2008-09 Norovirus 2009-10 Norovirus 2010-11
Figure 4. Diagram showing the number of NoV patients per week in Sweden reported to the Swedish center of infectious disease control (SMI) covering several years. (Kindly provided by K-O Hedlund, SMI)
The development of a NoV winter peak is dependent upon several factors such as tem-perature, humidity, population immunity and the emergence of new NoV variants (Lop-man et al., 2009). It is also worth noting that most of the winter peaks are due to GII.4 NoV strains (Kroneman et al., 2008). After the winter peak the GII.4 decreases only to reoccur again during the next winter season. GI strains however do not display the same

17
Human Noroviruses
seasonality as GII and this might be because they are more stable in the environment. GI NoV has been detected in waste water at a low but stable concentration all year around and is mostly associated with food borne outbreaks (Nordgren et al., 2009). In Sweden, NoV cases and outbreaks have been reported for several years and show a clear winter peak pattern (Hedlund et al., 2000) (Figure 4).
Symptoms and Transmission of norovirusThe symptoms for NoV gastroenteritis have been described to include nausea, vomit-ing and diarrhea sometimes accompanied by headache, low grade fever and muscle pain (Atmar et al., 2008; Green et al., 2001; Kaplan et al., 1982; Nordgren et al., 2010). NoVs infect individuals of all ages and vomiting has been seen to occur more frequently among children whereas adults tends to get more diarrhea (Kaplan et al., 1982). The incubation period is 12-48h (Estes et al., 2006; Kaplan et al., 1982) and the disease is generally self-limiting with symptoms resolving within 12-72 h (Atmar et al., 2008; Kaplan et al., 1982). However, in some cases symptoms may last for a longer time espe-cially in young children, elderly or immune compromised individuals (Carlsson et al., 2009; Lopman et al., 2004b; Murata et al., 2007; Nilsson et al., 2003). GII.4 NoV has been associated with more severe AGE episodes in infants than other NoV genotypes. Infants infected with GII.4 had a longer duration of diarrhea than did those infected with other NoV genotypes (Huhti et al., 2011).
NoV infection have also been associated with other conditions than AGE such as en-cephalopathy (Ito et al., 2006) and acute renal failure (Kanai et al., 2010). A few cases of NoV viremia have also been reported (Medici et al., 2010; Takanashi et al., 2009). After infection, the virus is shed in feces in great numbers, which facilitates the transmission. The route of NoV transmission is fecal-oral and by vomits. NoV outbreaks are common in areas like in the homes for elderly, schools, military settings and hospitals (Gallimore et al., 2004a; Gallimore et al., 2004b; Ike et al., 2006; Jiang et al., 1996; Lew et al., 1994; Lyman et al., 2009; Vardy et al., 2007; Wadl et al., 2010). The spread of NoV dif-fer between the different genogroups with GI spreading mainly through contaminated food and water and GII through person-to-person contact (Kroneman et al., 2008). The reason for this is unclear but could be explained by GI being more resistant to environ-mental factors (Nordgren et al., 2009). Viral shedding allows the virus to spread to new

18
Human Noroviruses
individuals and a study has shown that GII.4 strains are shed in higher titers than other strains (Bucardo et al., 2008). The shedding of new NoVs can occur for several weeks after clearing of the symptoms (Atmar et al., 2008; Kirkwood and Streitberg, 2008; Mu-rata et al., 2007; Rockx et al., 2002) and some individuals have even been diagnosed as chronic shedders (Carlsson et al., 2009; Nilsson et al., 2003). The shedding of virus for a long time is one of the factors contributing to the spread of NoV besides the virus ability to withstand disinfectants such as chlorine, freezing and high temperatures and the low viral dose required to cause disease (Duizer et al., 2004; Teunis et al., 2008). Foodborne transmissions and outbreaks have been described to occur due to contaminated shellfish (David et al., 2007; Doyle et al., 2004; Le Guyader et al., 2006; Ng et al., 2005) and ras-berries (Falkenhorst et al., 2005; Hjertqvist et al., 2006; Korsager et al., 2005; Maunula et al., 2009; Ponka et al., 1999).
Pathogenesis of Norovirus infectionSince human NoV (huNoV) cannot be grown in cell culture systems or animal models the knowledge of the pathology of NoV is still limited. To further understand the patho-genesis of gastroenteritis and more specifically of NoV AGE, volunteer studies have been much useful. Diarrhea and vomiting are the two most common symptoms of NoV disease. Diarrhea can have different causes; secretory diarrhea is caused by an influx of anions into the lumen, osmotic diarrhea is caused by impaired digestion of active agents such as disaccharides (Baldi et al., 2009). The altered movement of ions can occur either through transporters or the lateral spaces between cells, which are regulated by tight junctions (Hodges and Gill, 2010). One viral protein that could be responsible for alter-ing the balance of ions and water is the viral 3C like protease that interferes with host translation. Since both ion transporters and tight junctions have a turnover of about 12-18 hours, inhibition of host translation could influence the amount of these regulators of intestinal homeostasis (Hodges and Gill, 2010).
The NoV is believed to infect cells in the upper part of the small intestine, however the exact target cell is not yet identified but cells expressing DC-SIGN (dendritic cells) was recently suggested to be infected by human GI NoV (Bok et al., 2011). By using VLPs and intestinal tissue sections Marionneau and coworkers were able to demonstrate that NoVs bind mainly to intestinal villi and less to cells in the crypts (Marionneau et al.,

19
Human Noroviruses
2002). Several volunteer studies have shown mucosal lesions in the small bowel of in-fected individuals (Meeroff et al., 1980) and shorter and broader villus and hypertrophy of the crypts has also been observed (Schreiber et al., 1973). A study involving authen-tic NoV cases showed increased anion secretion and also immune infiltration of CD8+ lymphocytes (Troeger et al., 2009). Characteristic intestinal lesions could also be seen in asymptomatic volunteers suggesting that they had a subclinical infection (Meeroff et al., 1980). As mentioned earlier vomiting is also one of the main symptoms of NoV disease and is caused by stimuli of the vomiting center in the medulla (Guyton and Hall, 2000). Volunteer studies have shown no histological lesions in the gastric mucosa of infected individuals and no change in gastric secretions of hydrochloric acid, pepsin or intrinsic factor (Meeroff et al., 1980; Widerlite et al., 1975). Viremia of NoV has been found in 15% of NoV gastroenteritis patients by detection of NoV RNA in serum (Takanashi et al., 2009) thereby suggesting extra intestinal spread during NoV gastroenteritis.

20
Why are some people resistant to winter vomiting disease?
Why are some people resistant to winter vomiting disease?
Already in the 1970s it was seen that not all individuals became ill following NoV in-fection. Parrino and coworkers noted that only 6 individuals out of 12 infected with a GI.1 NoV fell ill with gastroenteritis (Parrino et al., 1977). On re-infecting the same 12 individuals 27-42 months later with the same GI.1 isolate, the same 6 individuals devel-oped disease. Further, 4-8 weeks after the second challenge, four of these six individuals were infected again and only 1 displayed symptoms of NoV illness. This study showed that no long-term protection seemed to be present but possibly a short-term protection from illness was observed. Individuals who became ill also displayed high antibody (ab) titers in comparison to the asymptomatic individuals who displayed low titers (Parrino et al., 1977). This suggests that protection to disease was not correlated to a strong ab response.
The possibility of protective immunity was further studied by Johnson and coworkers in 1990 in a volunteer study with 42 individuals (Johnson et al., 1990). After challenge, 15/30 individuals with low pre-existing abs fell ill with gastroenteritis compared to 10/12 among the volunteers with high pre-existing ab. The study did not find any cor-relation between the pre-existing serum ab and protective immunity to NoV disease. Short-term resistance was seen after challenge and some individuals maintained a low ab titer even after multiple challenges.From studies like these the question of why not all individuals become ill in winter vomiting disease arose. During the last decade many studies have been done to try and answer this question and it comes down to the role of acquired immunity as well as innate resistance to infection.
Histo blood group antigensHisto blood group antigens (HBGAs) are present on mucosa and in body fluids like saliva. Blood group antigens are the antigens determining our blood type, for example A, B, AB or O. NoVs have been suggested to use HBGAs as receptors when infecting individuals and studies indicate that individuals having blood group O are most sus-ceptible to disease while blood type B individuals are most resistant (Hennessy et al., 2003; Huang et al., 2003; Hutson et al., 2002; Rockx et al., 2005). Different NoV strains

21
Why are some people resistant to winter vomiting disease?
were shown to have different NoV binding patterns when tested in an enzyme linked immunosorbent assay (ELISA) using saliva with various HBGAs (Huang et al., 2003). Four patterns were described at first with a GII.4 binding to A, B and O individuals. GI showed binding to O and A but not to B and a GII.5 strain showed binding to A and B but not to O. Finally another GII strain had low binding to all blood groups and also to Lewis (Le) positive individuals. Since then more patterns have emerged (Huang et al., 2005). Binding of Norwalk (NV) VLP to HBGA glycosphingolipids revealed binding to A, H and Lewis but not to B (Nilsson et al., 2009). Other studies have reported no association between ABO blood groups and sporadic NoV outbreaks (Fretz et al., 2005; Halperin et al., 2008). Is the blood group antigens the answer or could it be something else considering that not all A and O individuals get infected with NoV?
Secretor statusThe secretor status of an individual determines if the HBGAs can be displayed on in-testinal mucosa and in saliva (Mollicone et al., 1995; Ravn and Dabelsteen, 2000). The FUT2 gene located on chromosome 19 in turn determines the secretor status. The FUT2 gene encodes for the enzyme fucosyltransferase 2 (FucT-II) that is active on the H type 1 precursor by adding a fucose in an a1,2 linkage to a terminal galactose (Figure 5). The enzyme FucT-I (fucosyltransferase I) is involved in the expression of the ABO antigens on red blood cells (RBCs).
A person unable to display HBGAs on mucosa is called a non-secretor. This inability to express HBGAs is caused by a non-functioning FUT2 gene. Approximately, 20% of the Caucasian population are non-secretors and the remaining 80% are secretors (Kindberg et al., 2007; Kindberg et al., 2006; Koda et al., 2001; Larsson et al., 2006; Thorven et al., 2005). Among the non-secretors, the most common mutation causing a non-functioning FUT2 allele is a substitution at nucleotide position 428 (G to A) causing a stop codon to arise and rendering the gene non-functional (Kelly et al., 1995). Two non-functioning FUT2 alleles cannot add a fucose to the precursor making the individual a non-secretor. Secretors can be either homozygous or heterozygous and one functioning FUT2 gene is enough. After the FucT-II has added the fucose to the H-type1 precursor, the H type 1 can then be further extended into A or B antigens by the A or B enzymes depending on the active glycosyltranferases of the individual (Figure 5). The addition of a a1, 4 linked

22
Why are some people resistant to winter vomiting disease?
fucose to the H type 1 precursor forms the Lewis a (Lea) structure. A fucosylation of the H type 1 chain forms the Lewis b (Leb) structure. The fucosyltransferase catalyzing these reactions is the FucT-III enzyme encoded by the FUT3 gene (Figure 5).
Gal GalGlcNAc Glc
Gal GalGlcNAc Glc
Fuc
Gal GalGlcNAc Glc
Fuc
GalNAc
Fuc
Gal GalGlcNAc Glc
Fuc
GalNAc
Fuc
Gal GalGlcNAc Glc
Fuc
Fuc
Gal GalGlcNAc Glc
H type 1 precursor
H-type 1
A B
FucT-II
A
FucT-III
FucT-III
Lea
Leb
LebA
GalGal GalGlcNAc Glc
Fuc
Fuc
GalGal GalGlcNAc Glc
Fuc
FucT-III
LebB
FucT-III
B
Figure 5. Schematic view of the synthesis of ABH and lewis antigens, based on the H type 1 chain. Enzymes are in black boxes. Secretors can express the ABH antigens on mucosa and in saliva due to at least one functional FUT2 allel. Non-secretors however, lacking the FucT-II en-zyme are unable to add a fucose to the H type 1 precursor making them unable to express the H type 1 or the following A or B antigens. The FucT-III enzyme is responsible for the expression of lewis antigens. Gal: galactose, GlcNAc: N-Acetylglucosamine, Glc: Glucose, Fuc: Fucose, Gal-NAc: N-Acetylgalactosamine, FucT: Fucosyltransferase, Le:Lewis
Individuals homozygous for an inactive FUT3 gene lack Lea and Leb structures. Ap-proximately 5% of the Caucasian population lacks the Le antigens and are hence called Le negative (Marionneau et al., 2001). Le negative individuals are much more common in for example Central America (Bucardo et al., 2009). The most common inactivating mutations in FUT3 are T202C, G508A and T1067A (Soejima and Koda, 2005). The H type 2 chain isomers of Lea and Leb are denoted Lex and Ley respectively.

23
Why are some people resistant to winter vomiting disease?
Susceptibility to diseaseHost genetic susceptibility has been suggested for several different viral diseases like tick borne encephalitis virus (Kindberg et al., 2008), human immunodeficiency virus (Blackwell et al., 1991; Dean et al., 1996; Kindberg et al., 2006; Svensson and Kind-berg, 2009) and parvovirus B19 (Brown et al., 1994). Also for NoV, host genetic resis-tance was early suggested as a possible explanation to why not every infected individual displayed disease (Parrino et al., 1977) (figure 6). Secretor status has been associated with increased or decreased susceptibility to urinary tract infection by E.coli (Sheinfeld et al., 1989; Stapleton et al., 1995) and respiratory viruses (Raza et al., 1991). Differ-ent binding patterns have been suggested for different NoV strains as mentioned earlier (Harrington et al., 2004; Huang et al., 2003; Huang et al., 2005; Tan and Jiang, 2008) and several studies have shown that NoV binds only to, or better to, saliva from secre-tor individuals than to saliva from non-secretors (Lindesmith et al., 2003; Marionneau et al., 2005; Marionneau et al., 2002). Marionneau and coworkers (Marionneau et al., 2002) also showed that NV VLPs bind only to duodenal cells of secretors but not to cells of non-secretors suggesting that non-secretors are protected from NoV disease. Hutson and coworkers (Hutson et al., 2005) showed in a volunteer study using GI NoV that 98% of all secretors were infected while none among the non-secretors were infected. Out of the 98% of secretors that were infected, two thirds were symptomatic.
Thorven and coworkers (Thorven et al., 2005) studied three nosocomial outbreaks at three different hospital wards caused by a GII.4 NoV and found that only secretors had symptomatic infection. They also studied community outbreaks caused by GII.2 and GI.6 showing that only secretor positive individuals had been infected (Thorven et al., 2005). They also showed that the outbreak virus only bound to secretor-positive saliva and not to the secretor-negative saliva. This suggests that a viral receptor is not only present in saliva but also in the gastrointestinal tract. Other studies have also shown that non-secretors are protected from infection with GII.4 and GII.3 (Tan et al., 2008b). By transfecting a non-susceptible cell-line with the fucosyltransferase gene one can enable NoV VLP to bind to cells which it did not bind to earlier and that normally do not express the H antigen (Marionneau et al., 2005). This further supports the secretor dependent infection pattern suggested for norovirus infection. Although FUT2 expres-sion is important for VLP and wild type virus binding and attachment, other factors are needed for efficient infection (Guix et al., 2007).

24
Why are some people resistant to winter vomiting disease?
Population
Se-(~20%)
Se+(~80%)
Protected
geneticresistence
secretor-independent
strains
aquiredimmunity
Infected Protected Infected
Figure 6. The non-secretor phenotype has been suggested to be protective for NoV infections in the Caucasian population. Certain NoV strains have been reported to infect also non-secretors. Not all secretor are always infected by NoV probably due to aquired immunity or other host or viral
factors not yet discovered.
However, it is never just so simple, there are exceptions for this secretor dependency among NoVs. In 2005, Rockx and coworkers reported that during a waterborne out-break among school children 4 non-secretors had been infected by a GI.3 NoV (Rockx et al., 2005). It has also been demonstrated elsewhere that GI NoVs bind to Lea, which is an antigen present in non-secretors suggesting that they can get infected (Shirato et al., 2008). Snow Mountain NoV (SMV) has been shown to have a secretor-independent pat-tern since one of the three non-secretors (33%) and eight of twelve secretors (67%) were infected (Lindesmith et al., 2005). This suggests that non-secretors are not completely protected from symptomatic NoV infection. However, it seems to be strain specific. Nordgren and coworkers (Nordgren et al., 2010) investigated an outbreak of gastro-enteritis in Jönköping, Sweden. A total of 83 individuals were included in the study

25
Why are some people resistant to winter vomiting disease?
and among these 33 (40%) were symptomatically infected. Surprisingly, 7/15 (47%) non-secretors were symptomatically infected compared to 26/68 (38%) among the se-cretors. In this outbreak, non-secretors and secretors were equally infected suggesting a secretor independent NoV strain. Nordgren and coworkers could genotype the NoV to GI.3 (Nordgren et al., 2010). Even though non-secretors can be infected by NoV, the numbers of strains that are able to do so are still limited.
In a study in 2006, I screened 105 Swedish blood donors for ab prevalence and titer to GII.4 NoV (Larsson et al., 2006). The study gave indirect evidence that non-secretors can be infected by a GII NoV as seen by the presence of IgG abs. However, the ab titers for secretor positive individuals were significantly higher than for non-secretors. This suggests that non-secretors had been in contact with the GII.4 NoV or a similar strain generating cross-reacting ab to GII.4 NoV. The prevalence of ab cannot however predict whether the individuals have experienced symptomatic NoV disease. The effect of these ab in terms of neutralization has not been investigated as to whether or not the abs could convey protection to winter vomiting disease. All these observations altogether indicate that even if some NoV strains can only infect secretors, the diverse binding patterns and sheer amount of different NoV strains found within the GI and GII NoVs may together be able to infect nearly all individuals.
Norovirus Immunology and immune evasionThe possibility of getting immunity to NoV illness has been debated for many years and the fact that most adult volunteers become ill after exposure suggests that either there is no natural immunity to this virus or that the NoV can evade the immune system. Even after multiple volunteer challenges, Parrino and coworkers could not detect any long-term immunity (Parrino et al., 1977). The ab produced by the individuals did not give protection but rather on the contrary, individuals with high ab titers seemed to be more prone to infection (Parrino et al., 1977). This has also been seen in studies investigating serum and local jejunum ab levels where the highest jejunum and serum ab levels were seen among susceptible individuals compared to those among the resistant individuals in both volunteer studies and authentic outbreaks (Blacklow et al., 1979; Graham et al., 1994; Okhuysen et al., 1995; Taylor et al., 1993). Short-term immunity was however

26
Why are some people resistant to winter vomiting disease?
observed for the Norwalk agent since volunteers did not become ill when re-challenged within 6-14 weeks (Parrino et al., 1977; Wyatt et al., 1974). Not all individuals respond-ed with elevated ab titers after being challenged with NoV (Gary et al., 1987; Parrino et al., 1977). In contrast a study by Farkas and coworkers suggested that pre-existing high ab titers may be protective against NoV illness, as they observed higher ab titers to a specific strain among controls in comparison to among sick patients on a cruise ship (Farkas et al., 2003). A recent study by Reeck and coworkers describe the correlation of pre-existing HBGA blocking ab and protection against gastroenteritis following NV infection (Reeck et al., 2010). Not all secretors become infected with NoV even though they do not have the genetic resistance provided by inactive FUT2 alleles. This could be due to acquired immunity (Figure 6) (Lindesmith et al., 2003).
Antibody prevalence and titer seems to increase with age. A study by Greenberg and co-workers (Greenberg et al., 1979) showed that ab prevalence was lowest among children up to five years of age (20%) and increased to 80-100% among teenagers and adults. In Bangladesh, ab prevalence to NoV was lowest among young children (2-7 months) and rose to approximately 60% among children 20-25 months of age. By 4 years of age all children in the study had ab to NoV (Black et al., 1982). A similar trend has been observed in Sweden where the ab prevalence to NoV GI was 50% in children less than 5 years of age and rose to more than 80% in children older than 10 years (Hinkula et al., 1995). A study recently done in Finland showed that 47.3% of children (2-7 months of age) had an ab prevalence to GII.4 NoV while 91.2% of children above 5 years of age had detectable ab levels (Nurminen et al., 2011). Similarly, in Sweden ab prevalence to NoV GII.4 among adults was as high as 98% (Larsson et al., 2006). Could it be that we are so often exposed to NoV that a long-term immunity is not required? This piece of the puzzle needs to be further investigated.
The individuals infected with NoV develop an ab response but what about the cell me-diated immunity? In 2003, Tacket and coworkers (Tacket et al., 2003) administered Norwalk VLPs to 30 volunteers and determined their immune response. Ninety percent of the challenged volunteers developed an IgG ab response. They could also detect a cell-mediated immune response including lymphoproliferative response and an increase in interferon γ (IFNγ) levels. Since no corresponding increase in IL-4 was detected, results thus confirm a Th1-like response (Ball et al., 1999; Tacket et al., 2003). Another

27
Why are some people resistant to winter vomiting disease?
study investigating SMV GII.2 also demonstrates high IFNγ and IL-2 levels further supporting a Th1 response also against live NoV challenge (Lindesmith et al., 2005). Similar studies for GI NoV have also reported elevated IFNγ secretion (Lindesmith et al., 2010). Additionally, Ko and coworkers have shown that the intestinal mucosal im-munity is also of Th1 type (Ko et al., 2006). Further, immunological studies in mice using the murine GV NoV indicates that CD4+ and CD8+ T cells together with B-cells are required for complete protection (Chachu et al., 2008; Lindesmith et al., 2010).
Norovirus evolutionWithin the GI and GII NoVs infecting humans, the GII.4 strains are most prevalent globally. Over the last two decades several GII.4 NoV epidemic peaks have occurred worldwide in 1996, 2002, 2004 and 2006 (Bull et al., 2006; Fankhauser et al., 2002; Lopman et al., 2004a; Noel et al., 1999; Siebenga et al., 2007a). These epidemic seasons were followed by periods of low NoV activity and after the 1996 pandemic with GII.4, the GII.3 NoV increased from about 4% in 1996 to 25% in 1997 (Noel et al., 1999). But what is it that has made the GII.4 so successful in generating pandemic outbreaks? The GII.4 NoV was first thought to emerge in the population in the mid 1980s but it was un-clear whether it recently emerged during that time or if it had been present in the human population for an extended period of time. A more recent study reported existence of GII.4 NoV prior to the 1980s (Bok et al., 2009). Already in the 1970s they found GII.4 NoV being present in the population and causing the disease. These earlier GII.4 NoVs shared the same binding properties as the Camberwell cluster that was thought to have appeared in the mid 80s.
Recent studies have suggested that GII.4 NoV evolve by epochal evolution (Lindesmith et al., 2008; Siebenga et al., 2007b). Epochal evolution is a process where periods of stasis, defined as no differences in phenotype are followed by periods of rapid evolution where new phenotypes emerge. This is the same type of evolution that is also seen for the influenza virus (Koelle et al., 2006). Lindesmith and coworkers mapped the genetic differences between the different pandemic clusters and observed that most variation could be detected in or near the receptor binding domains and predicted binding pock-ets. They further suggested that GII.4 NoV can persist by evolving new carbohydrate binding domains in response to immune driven selection and that antigenic drift in the

28
Why are some people resistant to winter vomiting disease?
receptor binding regions of the P2 domain can help the virus evade the pre-existing herd immunity (Lindesmith et al., 2008) (Figure 7). One example of this as reviewed by Donaldson and coworkers are the GII.4 strain that emerged in the mid 1980s also called the Camberwell cluster. This cluster in all likelihood encountered a naive population and had been able to infect the susceptible individuals. Herd immunity developed in this population and to avoid total extinction the virus thus evolved. The Grimsby cluster thus emerged 6-8 years later and was able to bind to additional A and B antigens giving an increased host range and the opportunity to infect a new subset of the population (Don-aldson et al., 2008). Just recently, the theory of evolving epitopes has been confirmed and the NoV evolving epitopes are most likely a result of mutations in and around the surface exposed sites (Lindesmith et al., 2011).
Figure 7. Two ways for NoV to evolve and evade herd immunity. To the left the NoV evolves by changing or adding a receptor allowing the virus to infect a larger population. To the right the pres-ence of many different viruses in the population allows a particular virus to avoid herd immunity. Small changes in the NoV genome could also give rise to an antigenic mutant that can escape
herd immunity.
The differences between strains can sometimes be due to a single amino acid (aa) change, but this can be enough to allow the virus to escape pre-existing herd immu-nity or to penetrate a previously naive population. NoVs like the Den Haag and Osaka strains previously infecting secretors have acquired the ability to use Lewis structures as receptors thus increasing their host range and might infect over 95% of the popula-tion (de Rougemont et al., 2011). Only the Lewis negative non-secretors seem to be protected from infection with these strains. De Rougemont and coworkers could also
Herdimmunity

29
Why are some people resistant to winter vomiting disease?
observe that aa not directly involved in HBGA binding contributed to the establishment of new binding profiles (de Rougemont et al., 2011). However, the future evolution of these stains is unclear since they are already able to infect more than 95% of the popu-lation. Increasing their host range further will not allow them to infect a much larger population. Could they evade herd immunity in some other way or will the immune pressure result in their disappearance and give space to other emerging variants? Bok and coworkers suggest the possibility of yet unrecognized virus host interactions apart from ab selection and HBGA binding properties that would allow the GII.4 NoV to persist in the human population (Bok et al., 2009).
Treatment, prevention and vaccine studiesTo date there is no special antiviral treatment available for NoV AGE. Hospitalized pa-tients can receive oral or intravenous rehydration but the fatalities in developing coun-tries are still high (Patel et al., 2008). Several research groups have investigated the possibility of making a vaccine against the NoV using VLPs or the smaller P-particles. In a recent study by Bok and coworkers, 6 chimpanzees were infected with Norwalk GI NoV and they all shed virus for 2 weeks or longer and had an ab response. Biopsies were taken and viral RNA was detected in cells (expressing DC-SIGN) of the jejunum and duodenum (Bok et al., 2011). Chimpanzees were then re-challenged between 2-24 months after initial challenges and they were found to be resistant to re-infection with the same virus. Naïve chimpanzees were then vaccinated with either GI or GII NoV VLP or placebo and then challenged after 18 months. Chimpanzees vaccinated with the GI NoV VLP were protected against infection with NV. However, the chimpanzees re-ceiving the GII VLP were susceptible to infection with NV suggesting that they did not produce cross-reacting ab (Bok et al., 2011). Even though no long-term immunity has been observed previously Bok and coworkers could demonstrate a correlation between the presence of abs in sera and protection from re-infection even 18 months after initial vaccination (Bok et al., 2011).
VLPs have also been given to humans to evaluate the immune response. A GI.1 NoV VLP given by nasal vaccination using an adjuvant showed a good serologic antibody re-sponse. The number of specific antibody-secreting cells (ASC) was reported to be high and cells homed to both gut mucosa and to peripheral lymphoid tissues (El-Kamary et al., 2010). In general the nasal vaccination route has several advantages like ease of ad-

30
Why are some people resistant to winter vomiting disease?
ministration, stimulation of both local and systemic immune responses and ease of stor-age at room temperature. Despite our best efforts there is to date no vaccine available. There are however, promising phase II trials with a novel nasal GI.1 norovirus vaccine based on VLPs. The vaccine was able to reduce norovirus illness by 47% and showed a 26% efficacy against norovirus infection and reduced the severity of illness (Atmar, 2010). To further complicate the norovirus puzzle new animal noroviruses are being investigated. Could these viruses cause any threat to humans? Are noroviruses circulat-ing in both humans and animals and could animals be a reservoir for NoV? Do we need to fear the possibility of new norovirus strains infecting humans or can we be confident that animal noroviruses will stay within the animal kingdom?

31
Bovine noroviruses
Bovine noroviruses
IntroductionSeveral bovine enteric viruses have been identified in calves, and in the 1970s the first reports of a calicivirus-like agent causing gastroenteritis in cattle was described (Woode and Bridger, 1978). In a study by Woode and Bridger, small un-identified viruses were found in feces from calves and were shown to be pathogenic to gnotobiotic (Gn) calves. The authors found what was later named the Newbury agent-1 and the Newbury agent-2 among astroviruses (Woode and Bridger, 1978). The incubation period seemed to be around 1-3 days and the infected calves experienced diarrhea, anorexia and lesions in the small intestine (Hall et al., 1984; Woode and Bridger, 1978). In more recent years the Newbury agent 1 together with the Nebraska strain isolated in the USA (Smiley et al., 2002) has been suggested to make up a new genotype in the calicivirus family called becovirus or nabovirus (Bridger et al., 1984; Oliver et al., 2006a).
The Newbury agent-2 identified in the United Kingdom and the closely related Jena virus identified in Germany is on the other hand more closely related to human GI NoVs than to other animal caliciviruses making it a third genogroup within the NoV genus (Dastjerdi et al., 1999; Liu et al., 1999; Oliver et al., 2003). The bovine Newbury-2 NoV shares a 81 to 85% aa identity to GI NoV and a 74-79% aa identity to GII NoVs (Dastjerdi et al., 1999). Both Jena virus and the Newbury agent-2 have been detected in the feces of newborn calves with diarrhea (van Der Poel et al., 2000). The Jena virus and the Newbury agent-2 are the representatives of the two genotypes of GIII.1 and GIII.2 respectively and the two genotypes are antigenically distinct (Oliver et al., 2006b).
Most of the bovine NoV (boNoV) strains found in cattle have been genetically related to GIII.2 suggesting that this is the most prevalent boNoV strain in cattle worldwide (Han et al., 2004; Ike et al., 2007; Mauroy et al., 2009a; Mauroy et al., 2009b; Milnes et al., 2007; Park et al., 2007; van der Poel et al., 2003; Wise et al., 2004). GIII NoV has also been detected in 18% of cattle in France and was also detected in water samples and in oysters together with GI and GII NoV (Zakhour et al., 2010).

32
Bovine noroviruses
Pathogenesis of GIII.2 infection in calvesInfecting Gn calves with Newbury agent-1 and 2 showed similar patophysiology of the intestine of the calves (Hall et al., 1984; Woode and Bridger, 1978). Infected calves experienced diarrhea, anorexia and intestinal lesions seen as shortening and bluntening of the villi in the first to middle part of the intestine. The most severe lesions were seen 24h after infection and could be detected until day 10 post infection (Hall et al., 1984). Viruses could be detected in intestinal contents and in feces from 1 to 4 days after infec-tion. Gn calves and pigs have also been infected with a human GII.4 NoV causing gas-troenteritis and intestinal lesions (Cheetham et al., 2006; Souza et al., 2008). All of the 5 GII.4 NoV infected Gn calves developed diarrhea. Severe lesions were observed in the duodenum and midjejunum characterized by severe loss of villi (Souza et al., 2008). Diarrhea was detected from day 2-6 after infection and viral shedding was detected dur-ing the same time and viremia was detected in 1/5 (20%) calves as characterized by viral RNA in serum. HuNoV capsid antigens were detected in the enterocytes of the jejunum and also in macrophage-like cells in the lamina propria. In regard to immune responses, the huNoV inoculated calves showed higher levels of TNF-α and INF γ that coincided with the onset of diarrhea. Both Th-1 (IL-12) and Th-2 (IL-4) cytokines were detected in huNoV inoculated calves (Souza et al., 2008) which is similar to findings seen in Gn pigs inoculated with the same strain of human GII.4 NoV (Cheetham et al., 2006).
When a virus invades a cell it can use different endocytic pathways, including macropi-nocytosis, clathrin-mediated endocytosis, caveole/lipid rafts and non-clathrin non-ca-veole pathways (Marsh and Helenius, 2006; Sieczkarski and Whittaker, 2002). A recent study by Mauroy and coworkers, demonstrated that the internalization of boNoV GIII.2 VLP is dependent upon the actin skeleton and that VLP entry into bovine kidney cells is mediated by both the cholesterol-dependent pathway and by macropinocytosis (Mauroy et al., 2011). Their results also suggest that the entry of bovine VLP into cells may re-quire a pH-dependent endocytic pathway, which is in contrast to the earlier study where the entry of GV murine NoV was reported to be pH-independent (Perry et al., 2009).

33
Bovine noroviruses
Zoonotic transmission, are humans at risk?Since the boNoVs are more closely related to huNoVs than to other animal calicviruses the possibility of zoonotic transfer to humans have been investigated. Van der poel and coworkers discussed the possibility of an animal reservoir for human infection based on the close genetic similarities between the strains although they did not find any proof for such zoonotic transmission (van Der Poel et al., 2000). However, natural recombinants have been seen among huNoVs, for example the HU/Snow Mountain/76/US strain (Lo-chridge and Hardy, 2003). Recombination can occur if the host animal is co-infected with two different strains. For boNoV, Han and coworkers, reported a recombinant, Bo/CV521-OH/02/US, with the NoV GIII.I like origin for the RdRp gene and NoV GIII.2 like origin for the capsid gene (Han et al., 2004). Recombination is suggested to have occurred in the ORF1-ORF2 junction region.
One could hypothesize that, since the huNoV can infect and trigger disease in calves, the bovine strain might be able to do the same in humans. Is it possible that a human and boNoV could co-infect a calf and cause a recombinant virus to occur? Could this new NoV be of risk to humans? Could this be a possible way for NoVs to create novel infec-tious strains? The boNoVs have been determined to be genetically and antigenically dis-tinct from human GI and GII NoV suggesting that the risk of infection to humans with a bovine strain is low (Han et al., 2005). However, this piece of the norovirus puzzle is not yet fully understood.
The ab prevalence among pigs to human NV was investigated in the USA and the study demonstrated that 63% were ab positive (Farkas et al., 2005). Of these, 71% were posi-tive for ab to a swine NoV strain (GII) and 52% had abs to the human Hawaii virus (GII). The ab levels were actually higher for GI NV than for the GII Hawaii virus even though the swine virus tested was a GII NoV (Farkas et al., 2005). However, this does not prove that there had been cross-species transmission between humans and pigs. The chances are that some of the seroprevalence observed may be due to the heterolo-gous responses to different antigenic types. Tian and coworkers (Tian et al., 2005) have shown that NV VLP binds to pig gastric mucin and that the synthetic Leb could com-pletely block this binding providing the first evidence that the receptor could be similar or identical to Leb (Tian et al., 2005).

34
Bovine noroviruses
The ab prevalence to bovine GIII.2 NoV among humans has also been investigated (Vildevall et al., 2010; Widdowson et al., 2005). Widdowson and coworkers (Widdow-son et al., 2005) reported that 22% of individuals had specific IgG abs to the boNoV strain. Seroprevalence among veterinarians was significantly higher (p=0,03) than among the general population (28% vs 20%). The possibility of cross-reactive abs being responsible for the ab prevalence to boNoV is unlikely and probably not responsible for most of the seroreactivity seen towards the bovine strain. Among the individuals that were ab positive for bovine GIII NoV, 26% had high reactivity to the bovine strains and low reactivity to the human NV strain (Widdowson et al., 2005). These results were confirmed later by Vildevall and coworkers (Vildevall et al., 2010) showing an ab prev-alence of 26.7% to Newbury-2 VLP among Swedish blood donors. A strong correlation to secretor status has been observed among GII huNoV and this study investigated the possible correlation of ab prevalence and titer to a boNoV GIII.2 NoV and HBGAs. A correlation could be found showing that secretor individuals had higher ab prevalence than non-secretors. Even though two studies have found abs to a bovine NoV strain in humans it does not prove symptomatic infection of humans with a bovine NoV strain.
What is the receptor for bovine norovirusThe suggested receptor structures for human GI and GII NoV are linked to different HBGAs and the secretor status of an individual. The receptor for boNoV is still un-known but is suggested to be the αGal (Gala1-3Galb1-4GlcNAc) epitope, a common carbohydrate structure in mammals (Galili, 2001). Zakhour and coworkers showed that boNoV could bind to epithelial cells in the bovine duodenum (Zakhour et al., 2009). They could also show that the boVLP probably recognizes a carbohydrate structure related to the αGal epitope. The αGal is not expressed in humans due to inactivation of the GGTA1 gene encoding a a1, 3galactosyltransferase. However, αGal is present in almost all other mammalian species (Galili, 2001). Pigs are known to express the αGal epitope and boNoV VLP was shown to bind also to the porcine gut by immunohisto-chemistry (Zakhour et al., 2009) (Figure 8). A study involving hemagglutination assay showed that boNoV can agglutinate RBCs of several species including calves, pigs and rabbits but not of other mammalian species like horse, sheep and goat (Vildevall et al., In Manuscript).

35
Bovine noroviruses
?
Hu
Hu
SwBo
Bo
HuInfection inGn pigs
Infection inGn calves
prevalence of abto BoNoV in humans
BoNoV bindsporcine gut (IHC)
Figure 8. The bovine GIII.2 NoV is more closely related to human noroviruses than to other animal caliciviruses. The human NoVs have been shown to infect both swine and cattle and the boNoV has been shown to bind to gastrointestinal cells from swine. Black arrows indicate infection, grey arrows indicate possible infection.
Mauroy and coworkers have shown that GIII.2 boVLP can bind to structures present on many different cell types like Madin-Darby bovine kidney cells, bovine turbinate cells, embryonic bovine tracheal cells, Georgia bovine kidney cells, embryonic bovine lung cells, bomac cells (macrophagic origin) and jejunocytes (Mauroy et al., 2011). Pre-treating cells with αGalactosidase significantly reduced binding of VLP suggesting that αGal is involved in binding of boVLP to bovine cells. Binding was seen not only to the αGal epitope but also moderately to sialic acid (Mauroy et al., 2011). If sialic acid is involved and can act as an alternative receptor this would increase host spectrum for the boNoV.
In addition, macropinocytosis is considered a non-specific mechanism for internaliza-tion and it does not require a specific ligand-receptor interaction. Macropinocytosis is widely used by antigen-presenting cells and could possibly be an entry portal for NoV. Macropinocytosis could possibly explain the ab prevalence seen in humans towards the boNoV since it does not require a specific receptor-ligand interaction. It is pos-sible that the boNoV can use macropinocytosis when infecting humans even though humans lack the proposed αGal receptor (Figure 8). Viruses have also been observed to bind to alternative ligands when their primary target is inaccessible, which for boNoV

36
Bovine noroviruses
could be sialic acid. However, sialic acid was shown not to be the main receptor in boVLP hemagglutination of bovine RBCs (Vildevall et al., In Manuscript). The feline calicivirus (FCV) belonging to the vesivirus genus enter cells via a clathrin-mediated endocytosis (Stuart and Brown, 2006) and the entry of murine NoV (MNV) belonging to GV is mediated by dynamin and cholesterol (Gerondopoulos et al., 2010; Perry and Wobus, 2010). Human NoV replication has also been demonstrated to be dependent on host cholesterol (Chang, 2009). Altogether, one may hypothesize that there is no clear and common entry path for caliciviruses. However, this needs to be further evaluated to be able to determine the route by which boNoV could infect humans. The Norovirus Puzzle is not yet finished and several more pieces needs to be added to the puzzle.

37
Aim of the Thesis
Aim of the ThesisThe general aims of this thesis were to study host susceptibility patterns of human and bovine NoV and also to further investigate the possible receptor and ligand structures for these viruses. By answering these questions, I hope to be able to add more pieces to the Norovirus Puzzle.
More specifically, the aims of the individual papers were:
Paper I: To analyze the ab prevalence and titer to human GII.4 NoV and its possible association to secretor status, ABO and Le status.
Paper II: To investigate if secretor-status determines susceptibility to NoV infection in Nicaragua and analyze the antibody prevalence and titer to NoV.
Paper III: To study the prevalence and titer of ab towards the bovine GIII.2 NoV in Swedish blood donors and the possible link to secretor status, ABO and Le status.
Paper IV: To characterize the hemagglutinin of bovine GIII.2 NoV.

38
Methods
MethodsDetailed descriptions regarding the materials and methods used can be found in each article. Here I would like to present five of the main methods used in the four papers included in my thesis (ELISA, Hemagglutination assay, Hemagglutination Inhibition assay, Pyrosequencing and Western blot). ELISA was used for ab detection (paper I, II, III and IV) and for binding studies (paper IV). Hemagglutination assay (HA) was used to determine HA activity (paper IV) and Hemagglutination inhibition (HI) was used to determine the ability of different carbohydrates and antibodies to block HA (paper IV). Pyrosequencing was used to determine FUT2 secretor status (paper II) and western blot was used to investigate cross reactivity between human and bovine NoV VLPs (paper III).
ELISATo determine the ab prevalence and titer to human and boNoV VLP, I used this ELISA protocol in paper I, II and III.
Human or boNoV VLPs were diluted in coating buffer (0,05M Sodiumcarbonate, pH 9,5) and coated onto a 96 well plate at a concentration of 0,5-1mg/ml. The plate was then incubated for 2h at 37°C and then overnight at 4°C. The following day, after 5 washes with washing buffer (0.9% NaCl, 0.05% Tween 20) the plate was blocked with 3% BSA (bovine serum albumin)/PBS for 1 h at 37°C. The human serum samples were prepared and diluted in dilution buffer (1% BSA, 0.005% Tween20/PBS) with a starting dilution of 1/100 and then serially diluted 2-fold up to 1/204800. After blocking and follow-ing with another 5 washes, diluted human serum samples were added to the plate and were allowed to bind to the VLPs for 1,5 h at 37°C. To detect the amount of binding, horseradish peroxidase (HRP) conjugated goat-anti-human IgG ab (1/10 000 in dilution buffer) was added after washing the plate. The plate was then incubated for 1 h at 37°C and finally after 5 washes, TMB (tetramethylbenzidine) was added to each well and the reaction was allowed to take place for approximately 10 min in dark at room tempera-ture (RT). The reaction was then stopped using 100ml/well of 2M H2SO4. Subsequently, the plate was read in a microplate ELISA reader at 450 nm. Results were seen as OD (optical density) values for each well.

39
Methods
For human ab titer determination, I also coated the plates with 0,5-1mg/ml BSA instead of VLPs and used the same serially diluted serum samples. The OD results from the BSA wells were then subtracted from the OD results of the VLP wells to account for possible unspecific binding. The negative control wells containing everything but the human serum samples were used to determine the cut off level. Cut off was set as the mean value of the negative controls +2SD. Hence, after the subtraction of the BSA OD values, the VLP OD values were considered positive for a given serum dilution if the OD was more than the cut off. The highest dilution with an OD higher than the cut off was set as the ab titer for that sample.
Hemagglutination AssayThe hemagglutination assay (Hierholzer and Suggs, 1969) can be used to determine the quantification of viruses and also to determine if the virus carries a hemagglutinin. HA takes advantage of the fact that some viruses can bind to the surface of RBCs. In paper IV, I have used HA to characterize the boNoV hemagglutinin.
RBCs were washed in PBS and centrifugated at 600g for 10 min. The supernatant was discarded and the RBCs were stored at 4°C. The VLPs were diluted two-fold in PBS in a V-bottom plate and the RBCs were added at a final concentration of 0.5%. The plate was then left to agglutinate at RT for 1 h and was then read visually. The HA titer was set as the highest dilution of VLPs that was able to cause hemagglutination of the RBCs in 50% of the wells. All the samples and controls were tested in duplicate or quadruple wells. Control wells contained only PBS and RBCs but no VLPs.
To characterize HA properties, RBCs from different species were tested including calf, pig, horse, goat, rabbit, guinea pig, sheep and human. The role of temperature (4-37°C) and pH (4-10) on HA activity was also investigated. Different enzymes including tryp-sin, neuroaminidase and α-galactosidase was incubated with RBCs to determine the characteristics of the receptor present on erythrocytes.

40
Methods
Hemagglutination Inhibition assayThe HAI determines the ability of abs present in a specific serum sample to block HA activity. To start with, an optimal VLP concentration is determined by setting the HAI VLP concentration to two folds below HA titer. For example, if the HA titer is 128 the VLP concentration used in HAI will be set to the concentration correlating to the titer of 32. The VLP concentration will be constant in the HAI assay.
The serum samples that were to be investigated were pre-treated by using a receptor-destroying enzyme, RDE to remove unspecific binding. Two parts of RDE (200ml) was mixed with 1 part serum (100ml) and was then incubated at 37°C for 18h followed by another incubation at 56°C for 30 min to inactivate the enzyme. After addition of 700ml 0,9% NaCl and 25ml washed RBCs to the RDE/serum mix they were incubated at 4°C for 1h with regular vortexing at every 10 min. The samples were centrifuged at 1200 rpm for 10 min and the serum obtained was collected and stored at -20°C. These serum samples were then diluted two-fold in PBS in a 96 well V-bottom plate. After addition of a pre-determined concentration of VLPs to all wells, the VLP and serum sample was incubated for 15-30 min at RT to allow binding. After this incubation, 1% RBCs (to a fi-nal concentration in the well of 0,5%) were added and was allowed to react for approxi-mately 1h at RT. After this, the plates were read visually. The control well was without VLP but with serum sample and RBCs. The HAI titer was set to the highest dilution of serum that was able to completely block agglutination of RBC by VLP.
PyrosequencingPyrosequencing can be used to determine single nucleotide polymophisms (SNP) for example the FUT2 gene which determines the secretor status of an individual. The py-rosequencing method was developed in the mid 1990´s (Ronaghi et al., 1996; Ronaghi et al., 1998) and is a sequencing-by synthesis method that can be monitored in real-time (figure 9). SNPs at position 428 and 571 were investigated by this pyrosequencing pro-tocol.
Prior to pyrosequencing the template was generated by conventional PCR by using a biotinylated forward primer. The dsDNA was immobilized to streptavidin-coated sep-harose beads to isolate the single biotinylated strand, each sample in one well of a 96-

41
Methods
well plate. A sequencing primer was hybridized to the template and incubated with a cocktail of enzymes, namely DNA polymerase, ATP sulfurylase, luciferase and apyrase as well as the substrates, adenosine 5´phosphosulfate (APS) and luciferin. After addi-tion of the first deoxribonucleotide triphosphate (dNTP), DNA polymerase catalyzed the incorporation of the dNTP into the DNA strand and the complimentary strand was synthesized. The principle involved in pyrosequencing is that one dNTP will be added at a time in a known dispension order and if the dNTP is incorporated it is accompanied by release of pyrophosphate (PPi). PPi is then converted into ATP by ATP-sulfurylase in the presence of APS. ATP drives the producion of visible light from luciferin, a re-action driven by luciferase, and the light will be detected by a charge coupled device camera and will be presented as a peak in a pyrogram.
A G G GC
C
C
CC
C
C
C CCAG G G GC C CA A AAA A AT T TTT T T G GA AT TTT C A
polymerase
apyrase
PPi
ATP
3’
3’5’
sulfurylase
luciferase
pyrogram
C G T A T C
Figure 9. Schematic overview of the pyrosequencing reaction. One dNTP at a time is added to a template of ssDNA. A mix of enzymes, namley DNA polymerase, ATP sulfurylase, luciferase and apyrase as well as the substrates, adenosine 5´phosphosulfate (APS) and luciferin will react and if a correct base is inserted visible light will be emitted and registered in a pyrogram as seen to the right. A dNTP that is not incorporated will be degraded by apyrase.

42
Methods
The height of each peak is proportional to the number of nucleotides incorporated. Apy-rase will degrade the unincorporated nucleotides and another one can then be added. Each analysed sample can be classified as homozygous wildtype (wt), heterozygous or homozygous mutant (mut).
Western BlotWestern blot can be used to detect specific proteins in a given sample and I have used it to determine the degree of cross-reactivity between the huNoV (GII.3) and boNoV (GIII.2).
Following separation of the VLP proteins on a 10% sodium dodecyl sulfate (SDS)-polyacrylamide gel, they were then transfered onto a PVDF membrane. Following the transfer of proteins the membrane was blocked with 3% BSA/PBS for 1h. After wash-ing (0.05% Tween 20/PBS) a mouse-anti-bovine VLP ab (1/5000) or a rabbit-anti-hu-man VLP ab (1/1000) was added and incubated at 4°C overnight. Following washing, a HRP-conjugated goat-anti-mouse IgG (1/20 000) or goat-anti-rabbit IgG (1/20 000) was added and incubated for 90 min at RT. Visualization was achieved by 5 min incubation with ECL (electrogenerated chemiluminescence) following exposure of the X-ray film. This resulted in the apperence of bands on the film that were specific to the VLP proteins and showed if there were any cross-reactivity present. I also used human sera samples to act as a primary antibody togheter with HRP-conjugated goat-anti-human IgG (1/10 000) to rule out major cross-reactivity to the different VLPs.

43
Results and Discussion
Results and DiscussionMy first two papers focus on the huNoVs mainly GII NoVs and the possible correla-tion to different HBGAs in both Sweden and Nicaragua. While a correlation to secretor status has been suggested for GII NoV it has not previously been investigated in terms of antibody prevalence and titer, nor has any information about secretor status and sus-ceptibility to NoV been reported from Nicaragua.
The possible zoonotic transmission of NoV between cattle and humans was investigated in Paper III and IV focusing on the bovine GIII.2 newbury-2 NoV. Prevalence and titer of boNoV ab among the Swedish human population was investigated and correlated to HBGAs. In paper IV, I characterized the bovine NoV hemagglutinin.
Paper I- Antibody Prevalence and Titer to Norovirus (Genogroup II) Correlate with Secretor (FUT2) but Not with ABO Phenotype or Lewis (FUT3) Genotype
Secretor status has been suggested to be a susceptibility marker for NoV infection and in this study we analyzed 105 Swedish blood donors for prevalence and titer of abs to a GII.4 NoV. All 105 individuals had been genotyped for secretor status and Le status and phenotyped for ABO and Le status. Results of the ab ELISA showed that 103/105 individuals (98.1%) had abs towards the GII.4 NoV, which is the most common NoV worldwide. This ab prevalence is similar to that previously reported in Sweden to a GI NoV (Hinkula et al., 1995). Table I shows the distribution of abs among different HBGAs and reveals significantly higher ab titers among secretors than among non-secretors (Figure 10).
All the secretor-positive individuals had ab to GII.4, while the only 2 ab negative in-dividuals were non-secretors. We found that non-secretors had significantly lower ab titers than secretors, suggesting that non-secretors are less prone to infection with GII.4 NoV. Three non-secretors however, had high ab titers (>1600) suggesting that they have been recently or repeatedly infected. No significant difference could be found between

44
Results and Discussion
Figure 10, Distribution of NoV GII.4 antibody titers among 105 Swedish blood donors character-
ized according to (A) secretor status, (B) ABO, (C) Lewis phenotype and (D) Lewis genotype.
Le-positive and Le-negative individuals. However, a significant difference in the ab titers was detected between Lea+b- and Lea-b+ where the Lea-b+ had the highest titers. The higher ab prevalence and titer to NoVs among secretors are in agreement with other studies and further supports the evidence that FUT2 plays a role in susceptibility to NoV (Hutson et al., 2005; Lindesmith et al., 2003; Marionneau et al., 2005; Marionneau et al., 2002).
No significant differences could be detected among individuals with different ABO blood groups. A trend could be seen towards lower titers for blood group B and highest titers among blood group O individuals. Other studies have previously suggested that blood group B individuals might be protected from infection (Hutson et al., 2002), but this can not be clearly confirmed by our results.
To summarize, there is a significant difference in ab titers between secretors and non-secretors suggesting that secretors may be more susceptible to GII.4 NoV infection.

45
Results and Discussion
Paper II- Genetic Susceptibility to Symptomatic Norovirus Infection in Nicaragua
Secretor status has been indicated to predict susceptibility to NoV disease based on studies made on the Caucasian populations in Europe and the United States. Limited information is available about the prevalence of non-secretors in Central America and whether secretor status can predict NoV disease. To investigate this, 28 NoV infected patients (median age 1.9 years) and 131 healthy controls (median age 24) were included in the study. Individuals were typed for ABO blood group, Le and secretor status. Serum samples were tested for ab prevalence and titer to GII.3 NoV.
Results showed that 82% (23/28) of patients were infected with a GII NoV, four with a GI and one with both GI and GII (table I). Sequencing revealed that GII.4 was the most common strain, followed by a proposed GII.18 Nica strain, GII.7, GII.17 and GI.4 geno-types. Among the control population, 6% were non-secretors and 25% were Le negative whereas among the NoV infected patients no non-secretors and 26% Le negative cases were detected.
Table I. Relationship between NoV genogroup, genotyps and HBGAs among NoV infected Indi-
viduals in Nicaragua.
No significant correlation could be made between symptomatic infection with NoV and ABO blood groups. However, all patients infected with GI NoV were of blood group O. While the globally dominating GII.4 NoV infected individuals of all blood groups. The prevalence of Le negative (25%) individuals in Nicaragua was high compared to

46
Results and Discussion
that in Sweden (5.7%) (Larsson et al., 2006) and Portugal (10%) (Serpa et al., 2003) but was consistent with the frequency reported from Colombia (22%) (Torrado et al., 1997). Le negative individuals were infected with both GI and GII NoV and Lewis sta-tus (positive vs negative) was not a predictive marker for NoV infection. The frequency of Le negatives may have been high but the number of non-secretors was only 3% (n=4) compared to 20% in Sweden. None of the non-secretors were infected with NoV, which is consistent with NoV protection being provided by the FUT2 inactivation. Four more non-secretors were found by Lewis phenotyping in saliva, by detection of Lea, and none of them had the G428A nonsense mutation in their FUT2 gene, which gives a total of 6% non-secretors.
The ab prevalence and titer to NoV GII.3 VLP was investigated and correlated with pa-tients and controls and also with HBGAs. In this study in Nicaragua 62% (74/120) were ab positive to NoV compared to 98% among Swedes (Larsson et al., 2006). Among secretors the ab prevalence was 63% compared to 33% among non-secretors. A signifi-cantly lower ab prevalence and titer was found among individuals with the AB pheno-type compared to A, B and O blood groups. One of the infected patients was ab negative. This patient was infected with a GII.17 strain suggesting that there is no cross reacting ab between GII.3 and GII.17. In this study similar ab titers were observed towards the GII.3 VLP for both GI and GII infected patients suggesting previous infections with GII strains. Mothers of infected patients revealed high ab prevalence (92%) however some mothers remained ab negative even though the child had a documented NoV infection.
In summary, the non-secretor phenotype was less common in Nicaragua compared to Europe but the Le negatives were much more common. None of the NoV infected pa-tients were non-secretors and the non-secretors also displayed lower ab prevalence and titer compared to secretors. This suggests that secretor status may be a predicator for NoV illness in Nicaragua.

47
Results and Discussion
Paper III- Human Antibody Responses to Bovine (Newbury-2) Nor-ovirus (GIII.2) and association to Histo-Blood Group Antigens
The boNoV forms a third genogroup in the NoV genus and can be further divided into two genotypes. In this study, the GIII.2 Newbury agent-2 VLP was used to detect se-rum abs in humans. Widdowson and coworkers reported that 22% of the population in the Netherlands had ab to bovine NoV with veterinarians having slightly higher ab prevalence than the general population (Widdowson et al., 2005). For human GII NoV infections secretor status has been shown to predict susceptibility to disease. For bovine NoV however, nothing is known about this possible association. ELISA was used to determine ab prevalence and titer to GIII.2 boNoV and to investigate a possible associa-tion with HBGAs (table II).
Table II. Serum antibody prevelence to bovine GIII.2 Newbury agent-2 NoV distributed among the
different ABO, lewis and secretor groups.
Out of 105 Swedish blood donors, the same individuals used in paper I, 26.7% were ab positive which is similar to that seen previously by Widdowson and coworkers (Wid-dowson et al., 2005). The suggested bovine GIII receptor is the αGal epitope, this epit-ope is not present in humans and humans would thus be resistant to infection with GIII NoV. However, the presence of abs towards the GIII.2 NoV suggests the possibility of another receptor being present allowing the boNoV to infect humans. A possible suscep-tibility pattern based on HBGAs was also investigated and revealed that ab titers were generally higher among secretors than among non-secretors suggesting that secretors might be more prone to infection with GIII.2 NoV. However, compared to the geometric mean titers (GMT) seen for GII.4 NoV the titers towards the GIII.2 NoV were gener-ally low (Larsson et al., 2006). A correlation could be seen in the frequency of abs to

48
Results and Discussion
boNoV where Lea+b- had a lower ab frequency than Lea-b+ individuals. This is consistent with susceptibility profiles for huNoV (Bucardo et al., 2009; Kindberg et al., 2007; Larsson et al., 2006; Lindesmith et al., 2003; Tan et al., 2008b; Thorven et al., 2005). No significant difference could be seen in ab prevalence or titer among individuals with different ABO blood groups. Worth noting however, is that when looking at only secre-tors, individuals of blood group B had significantly higher titers than blood group O (p=0.02). Blood group B and the suggested receptor αGal are similar in structure which could explain the higher ab frequency in this group. Among AB individual’s none of the seven had ab titers to boNoV.
Cross reactivity between the bovine GIII.2 strain and the human GII strain was inves-tigated. In this study no cross reactivity could be found using western blot, which is consistent to a previous study showing no cross-reactivity between boNoV GIII.2 and huNoV GI and GII (Han et al., 2005). However, a cross-reactive epitope between hu-NoV GI and boNoV GIII.2 has been reported (Batten et al., 2006; Oliver et al., 2006b). We found individuals with high ab titers to human GII NoV that was ab negative for bovine GIII NoV, for example the 7 AB individuals. This further supports the conclu-sion that no or little cross-reaction exists between GIII.2 and GII.4 NoV.
The results from this study indicate but do not prove that humans may be susceptible to infection by a boNoV strain. A recent study suggests that the boNoV could be inter-nalized by macropinocytosis (Mauroy et al., 2011), which does not require a specific receptor/lignad interaction, and this could explain the presence of antibodies in humans even without clinical symtoms. The observations we have made suggest that the αGal is not the sole receptor and that there may be another receptor used in humans. Sialic acid has been suggested as a possible second receptor structure for boNoV (Mauroy et al., 2011), which would increase the host range for boNoV. However, in paper IV I rule out the sialic acid as a receptor for binding to RBCs (Vildevall et al., In Manuscript), but it
may still be present in binding to cells of the gastrointestinal tract.

49
Results and Discussion
Paper IV- Characterization of the bovine norovirus Hemagglutinin
The NoVs have a capsid protein of approximately 530 aa, which is folded into two major domains, the conserved S domain and the more variable P domain. The P domain has been suggested to have antigenic determinants affecting immunological responses and host specificity. It has been shown previously that human GI NoV can agglutinate human RBCs from humans (Hutson et al., 2003) and that boNov has a hemagglutinin (Zakhour et al., 2009). The receptor for BoNoV is suggested to be the αGal epitope (Zakhour et al., 2009), which is not present in humans.
By using HA and HAI we studied the ability of Newbury agent-2 VLPs to agglutinate RBCs of different species and investigated the biochemical properties of the HA. In this study we have used RBCs of calves (8), pigs (9), rabbits (3), chicken (1), guinea pig (1), sheep’s (3) goat (1), horses (4) and human volunteers (10). HA results showed that boVLP were able to agglutinate RBCs from calves (6/8), pigs (9/9) and rabbits (2/3) but not of any other species tested. The αGal epitope is a common carbohydrate structure found in many mammals like calves, pigs, rabbits, horses, sheep’s and goats (Galili, 2001). Not all of the calves tested were able to agglutinate boVLP, thereby possibly reflecting polymorphisms in the gene associated with the αGal epitope.
Hemagglutination with bovine RBCs occurred at pH between 4-10 and at 4°C and at RT but not at 37°C. This suggests that the hemagglutination of boNoV is pH independent which is in contrast to the human GI NoV which was pH-dependent and was only able to agglutinate RBCs at pH 5-7.2 (Hutson et al., 2003). HuNoV was able to agglutinate RBCs from humans and from chimpanzee. Also, huNoV could only cause HA at 4°C (Hutson et al., 2003).
To investigate which part of the NoV capsid was involved in agglutination, three pep-tides were synthesized from the protruding region of the capsid (table III). These three peptides were used to immunize rabbits to generate peptide specific ab. The peptides themselves were not able to cause agglutination. However, the abs raised against pep-tide 3 was able to inhibit HA suggesting that aa 365-379 of the capsid is involved in agglutination. Modeling was done to show the location of the three different peptides on the capsid structure (Figure 11). The model showed a deletion in the bovine NoV

50
Results and Discussion
compared to the human NoV in the location of a putative binding pocket. This deletion could explain the differences in binding specificities between GI and GIII NoV. The third peptide was located at this position and this could explain the ability of the peptide 3 ab to inhibit agglutination.
Table III. Amino acid sequence and positions of the peptides in the boNoV capsid.
amino acid sequence positions
Peptide 1 (NH2-) FPDFSDCSLTFVVAS (-CONH2) 316-330 Peptide 2 (NH2-) CTVGERTVNTRNPQN (-CONH2) 334-346 Peptide 3 (NH2-) CFRAHLRNLWDPTEHS (-CONH2) 365-379
Twelve glycoconjugates, many of which has been previously suggested to be receptors for viruses including norovirus (Hutson et al., 2002; Rydell et al., 2009; Tan and Jiang, 2010; Zakhour et al., 2009) were used in HAI to investigate their biological relevance. However, none of them were able to inhibit hemagglutination even though the αGal epi-tope showed binding to boVLP in ELISA suggesting that there may be another receptor present. Another explanation is that the synthetic αGal epitope is monovalent but that the αGal receptor when presented in vivo is polyvalent.
Bovine serum samples were investigated for ab prevalence and titer to boNoV showing that 70% (7/10) had ab. Four of these serum samples displayed high ab titers (≥1600) and were able to block binding of boVLP to bovine saliva samples by almost 53% com-pared to 18.3% for the remaining 6 samples. This suggests that these serum antibodies may be able to inhibit infection with boNoV.
To investigate the structure of the virus receptor on the RBCs, cells were treated with different enzymes such as trypsin, neuraminidase and α-galactosidase. Neuraminidase treatment of RBCs did not affect the agglutination suggesting that the receptor is not si-alic acid. However, trypsin-treatment of the RBCs increased the HA titer and this could be due to removal of polypeptides that are presumably causing steric hindrance and blocking the receptor. The fact that trypsin-treatment increased HA titer suggests that

51
Results and Discussion
the receptor is a glycolipid. Treatment with α-galactosidase drastically reduced binding of boVLP to bovine RBCs suggesting that the αGal epitope is important for hemaggluti-nation and possibly connected to a glycolipid. The αGal epitope is closely related to the B blood group antigen and as can be seen by ab studies in humans the individuals car-rying the blood group B antigen had the highest ab prevalence to boVLP in comparison to all other blood groups (Vildevall et al., 2010)
To summarize, the boNoV were able to agglutinate RBCs of calves, pigs and rabbits further demonstrating that it has an agglutinin property. The results also indicate that the αGal epitope is important for binding but does not rule out the possibility of more than one receptor being present.

52
Results and Discussion
Figure 11. (A) Capsid protein P-dimers of Norwalk GI.1 (green) and the modelled Bo/New-bury2/1976/UK GIII.2 (brown) strains with histo-blood group A-trisaccharide in the binding site of Norwalk. Both dimers are presented as side view ribbon models in A and B. (B) A closer look at the Norwalk binding site shows a loop region as indicated by arrows which is a deletion in the modelled GIII.2 strain which could be responsible for the difference in the binding specificities of the two strains. (C) Top view surface model of GIII.2 showing the peptide sequences, used as immunogens, colored blue (peptide 1), purple (peptide 2) and cyan (peptide 3).

53
Concluding Remarks
Concluding RemarksSince I started my PhD, many new pieces have been added to the norovirus puzzle es-pecially concerning the secretor status and susceptibility to disease. Figure 12 attempts to summarize some of my findings regarding both the human and bovine NoVs (Figure 11). Paper I demonstrated that individuals with a secretor-positive phenotype have sig-nificantly higher ab prevalence and titer to the most common norovirus genotype GII.4 than secretor-negative individuals. These results support the hypothesis that noroviruses are more prone to infect secretor positive individuals. However, a few studies have since then observed exceptions to this rule and certain NoV strains infecting secretor-negative individuals have been documented. This further complicates the picture of norovirus susceptibility patterns and suggests that, taken together, all individuals can probably be infected with some NoV strains.
The presence of NoV illness and ab prevalence in the population has been demonstrated in many parts of the world and in paper II the prevalence of NoV disease in Nicaragua was investigated. Paper II clearly showed the diversity of norovirus strains present in patients with AGE and demonstrated the secretor-dependent pattern of infection for these strains since none of the NoV infected patients were secretor-negative. Ab data showed that non-secretor individuals had lower ab prevalence and titer than secretor-positive individuals suggesting repeated infections with NoVs for secretor positive in-dividuals.
Since the discovery of bovine NoV the question of zoonotic transfer has been debated. Human GII.4 NoV has been shown to be able to infect and trigger disease in Gn calves suggesting that the GII.4 NoV may use different receptor structures suitable for infec-tion of several species. Could this also be true for the boNoV? In paper III, I observed that 26.7% of Swedish blood donors had ab to the boNoV GIII.2 strain. The possible correlation to secretor status was also investigated and showed higher ab prevalence for secretor-positive individuals than for secretor-negative individuals.
In paper IV, the boNoV hemagglutinin was characterized and I concluded that the boNoVs carried a hemagglutinin on its surface as can be seen by the boNoVs ability to agglutinate RBCs of calf, pig and rabbit. Previous studies have suggested that the αGal epitope is crucial for the ability of boNoV to bind to the cells (Zakhour et al., 2009).

54
Concluding Remarks
BoHu
α Gal epitope
Hu
HBGA epitope
multiple virusstrains present
secretordependency
α Gal may notbe the only
receptor
BoNoVcarries a
hemagglutinin
aa position365-379 of theNoV capsid is important for
interaction
could BoNoVbind humancells?
Saliva
Figure 12. Summary of results obtained in this thesis. To the left is a schematic view of a human and a cow and the human is infected with a huNoV (yellow/green) and ab to huNoV is present in 98% of the Swedish population (Paper I). Several NoV can be found circulating in the population at the same time (Paper II). The human is also shown to have ab to the boNoV (pink/red) (Pa-per III) and calves has been shown to be infected with both boNoV and huNoV and also display antibodies to both virus types. Both the human and cow possibly display more than one kind of NoV receptor (Paper III, IV). To the right, HA results show that boNoV can bind to and agglutinate bovine RBCs (Paper IV) and it is also suggested that there may be more than one receptor pres-ent apart from the αGal-epitope (red) (paper IV). The location of a possible binding pocket on the boNoV is between aa 365-379 (Paper IV). Human and bovine NoVs have been demonstrated to bind saliva from humans and bovines respectivley.
This epitope is not present in humans but is present in many other mammals like cattle, pigs, horses, sheep’s and rabbits (Galili, 2001). The boNoV VLP were able to aggluti-nate RBC of some of these species but not from all even though they should all have the αGal epitope. Different enzymes were used and trypsin was able to increase agglutina-tion to RBCs but treatment with α-galactosidase abolished agglutination supporting the hypothesis that αGal is a necessary receptor but does not rule out the possibility of more than one receptor being present.

55
Concluding Remarks
By making models of the boNoV, I was able to find a position between aa 365-379 that appears to be a binding pocket for boNoV. The possibility of the boNoV to infect humans is still unclear but recombinations have been reported and the possibility of the bovine NoV evolving and extending its host range is not impossible. I believe that my studies have contributed to some new pieces of the Norovirus Puzzle but many more are still needed.

56
Acknowledgements
AcknowledgementsI would like to start by saying that there are so many persons that I would like to thank and I hope you all know how grateful I am for everything. Thank you!
A special thanks to my supervisor, Lennart Svensson, for your guidance and help during these years that I have been a PhD student.
I would also like to thank Jorma Hinkula for input on papers, help with methods and for answering questions all the time.
I would like to send a big thank you to Britt Åkerlind who was the first to get me inter-ested in virology and to Lena Serrander and Marie Abrahamsson in clinical virology for your help in finding me good samples to work with.
And to my friend and office mate, Carolin, what would I have done whithout you? Thank you for always being there!
To Elin, a big Thank you for always taking the time to answer stupid questions and for nice times together in the lab and away.
Big thank you also to everybody in Lennart´s group; Johan, Beatrice, Marie, Filemon and Sumit for your help and friendship all of these years. I wish you all the best.
A big thank you to Marie Larsson and her group, my fellow PhD students Veronica, Veegard, Karl and Rada for fun and challenching times at level 13. A special thanks to Rada who made most of the nice pictures in this thesis, I am so grateful to you.
Thank you also to my friends and collegues Tina, Josefin, Camilla, Lina and Johanna for all nice fikas and after work activities. Good luck to all of you with everything you decide to do.
Thanks to Pia and Lotta for all your help and company. And to Lena svensson, I have really missed you at level 13.

57
Acknowledgements
Thanks also to everybody at level 13 for making this a great place to work! Thanks for nice fika-times and discussions!
Thank you also to Göran Larson and Gustaf Rydell in Gothenburg for good collabora-tions and nice discussions during the years.
Thank you also to my good friends Heli and Henrik for many nice times together and for listening to all my strange lab issues. I look forward to our vacation this summer!
Thank you to my mother Åsa and Anders for keeping us busy during our visits at Kull-bol and for providing us with good food and farmlife adventures. Thanks also to my brothers Niklas and Samir, I wish you all the best.
To my father Benny, marita and my siblings Patrik, Frida (lill), Felicia, Frida and Tobi-as for nice times and happy holidays together. Good luck with your studies and future adventures!
To my children, you are the stars of my life. Thank you for all your hugs and kisses all those days when I really needed them.
And last but not least, my husband Tonny. If it hadn’t been for you this would have never been possible. You have always been by my side, supporting me every step of the way. I am finally at the end of this journey and I owe it all to you. I´m Yours.

58
References
ReferencesAdler, J.L., and R. Zickl. 1969. Winter vomiting disease. J Infect Dis 119:668-673.Atmar, R. 2010. Intranasal Norovirus VLP Vaccine Demonstrates Clinical Efficacy
When Compared to Placebo in Phase I/II Trial. In Infectious Diseases Society of Amerika. Vancouver, British Colombia.
Atmar, R.L., and M.K. Estes. 2006. The epidemiologic and clinical importance of noro-virus infection. Gastroenterol Clin North Am 35:275-290, viii.
Atmar, R.L., A.R. Opekun, M.A. Gilger, M.K. Estes, S.E. Crawford, F.H. Neill, and D.Y. Graham. 2008. Norwalk virus shedding after experimental human infec-tion. Emerg Infect Dis 14:1553-1557.
Baldi, F., M.A. Bianco, G. Nardone, A. Pilotto, and E. Zamparo. 2009. Focus on acute diarrhoeal disease. World J Gastroenterol 15:3341-3348.
Ball, J.M., D.Y. Graham, A.R. Opekun, M.A. Gilger, R.A. Guerrero, and M.K. Estes. 1999. Recombinant Norwalk virus-like particles given orally to volunteers: phase I study. Gastroenterology 117:40-48.
Batten, C.A., I.N. Clarke, S.L. Kempster, S.L. Oliver, J.C. Bridger, and P.R. Lambden. 2006. Characterization of a cross-reactive linear epitope in human genogroup I and bovine genogroup III norovirus capsid proteins. Virology 356:179-187.
Bishop, R.F., G.P. Davidson, I.H. Holmes, and B.J. Ruck. 1973. Virus particles in epi-thelial cells of duodenal mucosa from children with acute non-bacterial gastro-enteritis. Lancet 2:1281-1283.
Black, R.E., H.B. Greenberg, A.Z. Kapikian, K.H. Brown, and S. Becker. 1982. Acqui-sition of serum antibody to Norwalk Virus and rotavirus and relation to diar-rhea in a longitudinal study of young children in rural Bangladesh. J Infect Dis 145:483-489.
Blacklow, N.R., G. Cukor, M.K. Bedigian, P. Echeverria, H.B. Greenberg, D.S. Sch-reiber, and J.S. Trier. 1979. Immune response and prevalence of antibody to Norwalk enteritis virus as determined by radioimmunoassay. J Clin Microbiol 10:903-909.
Blackwell, C.C., V.S. James, S. Davidson, R. Wyld, R.P. Brettle, R.J. Robertson, and D.M. Weir. 1991. Secretor status and heterosexual transmission of HIV. BMJ 303:825-826.
Blanton, L.H., S.M. Adams, R.S. Beard, G. Wei, S.N. Bulens, M.A. Widdowson, R.I. Glass, and S.S. Monroe. 2006. Molecular and epidemiologic trends of calici-viruses associated with outbreaks of acute gastroenteritis in the United States, 2000-2004. J Infect Dis 193:413-421.
Bok, K., E.J. Abente, M. Realpe-Quintero, T. Mitra, S.V. Sosnovtsev, A.Z. Kapikian, and K.Y. Green. 2009. Evolutionary dynamics of GII.4 noroviruses over a 34-year period. J Virol 83:11890-11901.
Bok, K., G.I. Parra, T. Mitra, E. Abente, C.K. Shaver, D. Boon, R. Engle, C. Yu, A.Z. Kapikian, S.V. Sosnovtsev, R.H. Purcell, and K.Y. Green. 2011. Chimpanzees as an animal model for human norovirus infection and vaccine development. Proc Natl Acad Sci U S A 108:325-330.

59
References
Boschi-Pinto, C., L. Velebit, and K. Shibuya. 2008. Estimating child mortality due to diarrhoea in developing countries. Bull World Health Organ 86:710-717.
Bridger, J.C., G.A. Hall, and J.F. Brown. 1984. Characterization of a calici-like virus (Newbury agent) found in association with astrovirus in bovine diarrhea. Infect Immun 43:133-138.
Brown, K.E., J.R. Hibbs, G. Gallinella, S.M. Anderson, E.D. Lehman, P. McCarthy, and N.S. Young. 1994. Resistance to parvovirus B19 infection due to lack of virus receptor (erythrocyte P antigen). N Engl J Med 330:1192-1196.
Bryce, J., C. Boschi-Pinto, K. Shibuya, and R.E. Black. 2005. WHO estimates of the causes of death in children. Lancet 365:1147-1152.
Bucardo, F., E. Kindberg, M. Paniagua, A. Grahn, G. Larson, M. Vildevall, and L. Svensson. 2009. Genetic susceptibility to symptomatic norovirus infection in Nicaragua. J Med Virol 81:728-735.
Bucardo, F., J. Nordgren, B. Carlsson, M. Paniagua, P.E. Lindgren, F. Espinoza, and L. Svensson. 2008. Pediatric norovirus diarrhea in Nicaragua. J Clin Microbiol 46:2573-2580.
Bull, R.A., E.T. Tu, C.J. McIver, W.D. Rawlinson, and P.A. White. 2006. Emergence of a new norovirus genotype II.4 variant associated with global outbreaks of gastroenteritis. J Clin Microbiol 44:327-333.
Carlsson, B., A.M. Lindberg, J. Rodriguez-Diaz, K.O. Hedlund, B. Persson, and L. Svensson. 2009. Quasispecies dynamics and molecular evolution of human norovirus capsid P region during chronic infection. J Gen Virol 90:432-441.
Chachu, K.A., D.W. Strong, A.D. LoBue, C.E. Wobus, R.S. Baric, and H.W.t. Virgin. 2008. Antibody is critical for the clearance of murine norovirus infection. J Virol 82:6610-6617.
Chang, K.O. 2009. Role of cholesterol pathways in norovirus replication. J Virol 83:8587-8595.
Cheetham, S., M. Souza, T. Meulia, S. Grimes, M.G. Han, and L.J. Saif. 2006. Pathogen-esis of a genogroup II human norovirus in gnotobiotic pigs. J Virol 80:10372-10381.
Dastjerdi, A.M., J. Green, C.I. Gallimore, D.W. Brown, and J.C. Bridger. 1999. The bovine Newbury agent-2 is genetically more closely related to human SRSVs than to animal caliciviruses. Virology 254:1-5.
David, S.T., L. McIntyre, L. MacDougall, D. Kelly, S. Liem, K. Schallie, A. McNabb, A. Houde, P. Mueller, P. Ward, Y.L. Trottier, and J. Brassard. 2007. An outbreak of norovirus caused by consumption of oysters from geographically dispersed harvest sites, British Columbia, Canada, 2004. Foodborne Pathog Dis 4:349-358.
de Rougemont, A., N. Ruvoen-Clouet, B. Simon, M. Estienney, C. Elie-Caille, S. Aho, P. Pothier, J. Le Pendu, W. Boireau, and G. Belliot. 2011. Qualitative and quan-titative analysis of the binding of GII.4 norovirus variants onto human blood group antigens. J Virol
Dean, M., M. Carrington, C. Winkler, G.A. Huttley, M.W. Smith, R. Allikmets, J.J.

60
References
Goedert, S.P. Buchbinder, E. Vittinghoff, E. Gomperts, S. Donfield, D. Vlahov, R. Kaslow, A. Saah, C. Rinaldo, R. Detels, and S.J. O’Brien. 1996. Genetic restriction of HIV-1 infection and progression to AIDS by a deletion allele of the CKR5 structural gene. Hemophilia Growth and Development Study, Mul-ticenter AIDS Cohort Study, Multicenter Hemophilia Cohort Study, San Fran-cisco City Cohort, ALIVE Study. Science 273:1856-1862.
Donaldson, E.F., L.C. Lindesmith, A.D. Lobue, and R.S. Baric. 2008. Norovirus patho-genesis: mechanisms of persistence and immune evasion in human populations. Immunol Rev 225:190-211.
Doyle, A., D. Barataud, A. Gallay, J.M. Thiolet, S. Le Guyaguer, E. Kohli, and V. Vail-lant. 2004. Norovirus foodborne outbreaks associated with the consumption of oysters from the Etang de Thau, France, December 2002. Euro Surveill 9:24-26.
Duizer, E., P. Bijkerk, B. Rockx, A. De Groot, F. Twisk, and M. Koopmans. 2004. Inac-tivation of caliciviruses. Appl Environ Microbiol 70:4538-4543.
El-Kamary, S.S., M.F. Pasetti, P.M. Mendelman, S.E. Frey, D.I. Bernstein, J.J. Treanor, J. Ferreira, W.H. Chen, R. Sublett, C. Richardson, R.F. Bargatze, M.B. Sztein, and C.O. Tacket. 2010. Adjuvanted intranasal Norwalk virus-like particle vac-cine elicits antibodies and antibody-secreting cells that express homing recep-tors for mucosal and peripheral lymphoid tissues. J Infect Dis 202:1649-1658.
Estes, M.K., B.V. Prasad, and R.L. Atmar. 2006. Noroviruses everywhere: has some-thing changed? Curr Opin Infect Dis 19:467-474.
Falkenhorst, G., L. Krusell, M. Lisby, S.B. Madsen, B. Bottiger, and K. Molbak. 2005. Imported frozen raspberries cause a series of norovirus outbreaks in Denmark, 2005. Euro Surveill 10:E050922 050922.
Fankhauser, R.L., S.S. Monroe, J.S. Noel, C.D. Humphrey, J.S. Bresee, U.D. Parashar, T. Ando, and R.I. Glass. 2002. Epidemiologic and molecular trends of “Nor-walk-like viruses” associated with outbreaks of gastroenteritis in the United States. J Infect Dis 186:1-7.
Farkas, T., S. Nakajima, M. Sugieda, X. Deng, W. Zhong, and X. Jiang. 2005. Serop-revalence of noroviruses in swine. J Clin Microbiol 43:657-661.
Farkas, T., K. Sestak, C. Wei, and X. Jiang. 2008. Characterization of a rhesus monkey calicivirus representing a new genus of Caliciviridae. J Virol 82:5408-5416.
Farkas, T., S.A. Thornton, N. Wilton, W. Zhong, M. Altaye, and X. Jiang. 2003. Homol-ogous versus heterologous immune responses to Norwalk-like viruses among crew members after acute gastroenteritis outbreaks on 2 US Navy vessels. J Infect Dis 187:187-193.
Fretz, R., P. Svoboda, D. Schorr, M. Tanner, and A. Baumgartner. 2005. Risk factors for infections with Norovirus gastrointestinal illness in Switzerland. Eur J Clin Microbiol Infect Dis 24:256-261.
Galili, U. 2001. The alpha-gal epitope (Gal alpha 1-3Gal beta 1-4GlcNAc-R) in xeno-transplantation. Biochimie 83:557-563.
Gallimore, C.I., M.A. Barreiros, D.W. Brown, J.P. Nascimento, and J.P. Leite. 2004a. Noroviruses associated with acute gastroenteritis in a children’s day care facil-

61
References
ity in Rio de Janeiro, Brazil. Braz J Med Biol Res 37:321-326.Gallimore, C.I., D. Cubitt, N. du Plessis, and J.J. Gray. 2004b. Asymptomatic and symp-
tomatic excretion of noroviruses during a hospital outbreak of gastroenteritis. J Clin Microbiol 42:2271-2274.
Gary, G.W., L.J. Anderson, B.H. Keswick, P.C. Johnson, H.L. DuPont, S.E. Stine, and A.V. Bartlett. 1987. Norwalk virus antigen and antibody response in an adult volunteer study. J Clin Microbiol 25:2001-2003.
Gerondopoulos, A., T. Jackson, P. Monaghan, N. Doyle, and L.O. Roberts. 2010. Mu-rine norovirus-1 cell entry is mediated through a non-clathrin-, non-caveolae-, dynamin- and cholesterol-dependent pathway. J Gen Virol 91:1428-1438.
Graham, D.Y., X. Jiang, T. Tanaka, A.R. Opekun, H.P. Madore, and M.K. Estes. 1994. Norwalk virus infection of volunteers: new insights based on improved assays. J Infect Dis 170:34-43.
Green, K., R.M. Chanock, and A.Z. Kapikian. 2001. Human Caliciviruses. In Fields Virology, 4th ed. D. Knipe, and P. Howley, editors. Lippincott Williams & Wilkins, Philadephia. 841-874.
Greenberg, H.B., J. Valdesuso, A.Z. Kapikian, R.M. Chanock, R.G. Wyatt, W. Szmu-ness, J. Larrick, J. Kaplan, R.H. Gilman, and D.A. Sack. 1979. Prevalence of antibody to the Norwalk virus in various countries. Infect Immun 26:270-273.
Guix, S., M. Asanaka, K. Katayama, S.E. Crawford, F.H. Neill, R.L. Atmar, and M.K. Estes. 2007. Norwalk virus RNA is infectious in mammalian cells. J Virol 81:12238-12248.
Guyton, A., and J. Hall. 2000. Textbook of Medical Physiology. W.B. Saunders Com-pany, Philadelphia, US.
Hall, G.A., J.C. Bridger, B.E. Brooker, K.R. Parsons, and E. Ormerod. 1984. Lesions of gnotobiotic calves experimentally infected with a calicivirus-like (Newbury) agent. Vet Pathol 21:208-215.
Halperin, T., H. Vennema, M. Koopmans, G. Kahila Bar-Gal, R. Kayouf, T. Sela, R. Ambar, and E. Klement. 2008. No association between histo-blood group an-tigens and susceptibility to clinical infections with genogroup II norovirus. J Infect Dis 197:63-65.
Han, M.G., J.R. Smiley, C. Thomas, and L.J. Saif. 2004. Genetic recombination be-tween two genotypes of genogroup III bovine noroviruses (BoNVs) and capsid sequence diversity among BoNVs and Nebraska-like bovine enteric calicivi-ruses. J Clin Microbiol 42:5214-5224.
Han, M.G., Q. Wang, J.R. Smiley, K.O. Chang, and L.J. Saif. 2005. Self-assembly of the recombinant capsid protein of a bovine norovirus (BoNV) into virus-like particles and evaluation of cross-reactivity of BoNV with human noroviruses. J Clin Microbiol 43:778-785.
Harrington, P.R., J. Vinje, C.L. Moe, and R.S. Baric. 2004. Norovirus capture with histo-blood group antigens reveals novel virus-ligand interactions. J Virol 78:3035-3045.
Hedlund, K.O., E. Rubilar-Abreu, and L. Svensson. 2000. Epidemiology of calicivirus

62
References
infections in Sweden, 1994-1998. J Infect Dis 181 Suppl 2:S275-280.Hennessy, E.P., A.D. Green, M.P. Connor, R. Darby, and P. MacDonald. 2003. Norwalk
virus infection and disease is associated with ABO histo-blood group type. J Infect Dis 188:176-177.
Hierholzer, J.C., and M.T. Suggs. 1969. Standardized viral hemagglutination and hem-agglutination-inhibition tests. I. Standardization of erythrocyte suspensions. Appl Microbiol 18:816-823.
Hinkula, J., J.M. Ball, S. Lofgren, M.K. Estes, and L. Svensson. 1995. Antibody preva-lence and immunoglobulin IgG subclass pattern to Norwalk virus in Sweden. J Med Virol 47:52-57.
Hjertqvist, M., A. Johansson, N. Svensson, P.E. Abom, C. Magnusson, M. Olsson, K.O. Hedlund, and Y. Andersson. 2006. Four outbreaks of norovirus gastroen-teritis after consuming raspberries, Sweden, June-August 2006. Euro Surveill 11:E060907 060901.
Hodges, K., and R. Gill. 2010. Infectious diarrhea: Cellular and molecular mechanisms. Gut Microbes 1:4-21.
Huang, P., T. Farkas, S. Marionneau, W. Zhong, N. Ruvoen-Clouet, A.L. Morrow, M. Altaye, L.K. Pickering, D.S. Newburg, J. LePendu, and X. Jiang. 2003. Noro-viruses bind to human ABO, Lewis, and secretor histo-blood group antigens: identification of 4 distinct strain-specific patterns. J Infect Dis 188:19-31.
Huang, P., T. Farkas, W. Zhong, M. Tan, S. Thornton, A.L. Morrow, and X. Jiang. 2005. Norovirus and histo-blood group antigens: demonstration of a wide spectrum of strain specificities and classification of two major binding groups among multiple binding patterns. J Virol 79:6714-6722.
Huhti, L., E.D. Szakal, L. Puustinen, M. Salminen, H. Huhtala, O. Valve, V. Blazevic, and T. Vesikari. 2011. Norovirus GII-4 Causes a More Severe Gastroenteritis Than Other Noroviruses in Young Children. J Infect Dis
Hutson, A.M., F. Airaud, J. LePendu, M.K. Estes, and R.L. Atmar. 2005. Norwalk vi-rus infection associates with secretor status genotyped from sera. J Med Virol 77:116-120.
Hutson, A.M., R.L. Atmar, D.Y. Graham, and M.K. Estes. 2002. Norwalk virus infec-tion and disease is associated with ABO histo-blood group type. J Infect Dis 185:1335-1337.
Hutson, A.M., R.L. Atmar, D.M. Marcus, and M.K. Estes. 2003. Norwalk virus-like particle hemagglutination by binding to h histo-blood group antigens. J Virol 77:405-415.
Ike, A.C., S.O. Brockmann, K. Hartelt, R.E. Marschang, M. Contzen, and R.M. Oehme. 2006. Molecular epidemiology of norovirus in outbreaks of gastroenteritis in southwest Germany from 2001 to 2004. J Clin Microbiol 44:1262-1267.
Ike, A.C., B.N. Roth, R. Bohm, A.J. Pfitzner, and R.E. Marschang. 2007. Identifica-tion of bovine enteric Caliciviruses (BEC) from cattle in Baden-Wurttemberg. Dtsch Tierarztl Wochenschr 114:12-15.
Ito, S., S. Takeshita, A. Nezu, Y. Aihara, S. Usuku, Y. Noguchi, and S. Yokota. 2006.

63
References
Norovirus-associated encephalopathy. Pediatr Infect Dis J 25:651-652.Jiang, X., E. Turf, J. Hu, E. Barrett, X.M. Dai, S. Monroe, C. Humphrey, L.K. Picker-
ing, and D.O. Matson. 1996. Outbreaks of gastroenteritis in elderly nursing homes and retirement facilities associated with human caliciviruses. J Med Vi-rol 50:335-341.
Jiang, X., M. Wang, D.Y. Graham, and M.K. Estes. 1992. Expression, self-assembly, and antigenicity of the Norwalk virus capsid protein. J Virol 66:6527-6532.
Johnson, P.C., J.J. Mathewson, H.L. DuPont, and H.B. Greenberg. 1990. Multiple-chal-lenge study of host susceptibility to Norwalk gastroenteritis in US adults. J Infect Dis 161:18-21.
Kanai, T., S. Yotsumoto, and M.Y. Momoi. 2010. Norovirus-associated renal acute renal failure with nephrotic syndrome. Pediatr Int 52:e23-25.
Kapikian, A.Z., R.G. Wyatt, R. Dolin, T.S. Thornhill, A.R. Kalica, and R.M. Chanock. 1972. Visualization by immune electron microscopy of a 27-nm particle associ-ated with acute infectious nonbacterial gastroenteritis. J Virol 10:1075-1081.
Kaplan, J.E., G.W. Gary, R.C. Baron, N. Singh, L.B. Schonberger, R. Feldman, and H.B. Greenberg. 1982. Epidemiology of Norwalk gastroenteritis and the role of Norwalk virus in outbreaks of acute nonbacterial gastroenteritis. Ann Intern Med 96:756-761.
Kelly, R.J., S. Rouquier, D. Giorgi, G.G. Lennon, and J.B. Lowe. 1995. Sequence and expression of a candidate for the human Secretor blood group alpha(1,2)fu-cosyltransferase gene (FUT2). Homozygosity for an enzyme-inactivating non-sense mutation commonly correlates with the non-secretor phenotype. J Biol Chem 270:4640-4649.
Kindberg, E., B. Akerlind, C. Johnsen, J.D. Knudsen, O. Heltberg, G. Larson, B. Bot-tiger, and L. Svensson. 2007. Host genetic resistance to symptomatic norovirus (GGII.4) infections in Denmark. J Clin Microbiol 45:2720-2722.
Kindberg, E., B. Hejdeman, G. Bratt, B. Wahren, B. Lindblom, J. Hinkula, and L. Svens-son. 2006. A nonsense mutation (428G-->A) in the fucosyltransferase FUT2 gene affects the progression of HIV-1 infection. AIDS 20:685-689.
Kindberg, E., A. Mickiene, C. Ax, B. Akerlind, S. Vene, L. Lindquist, A. Lundkvist, and L. Svensson. 2008. A deletion in the chemokine receptor 5 (CCR5) gene is as-sociated with tickborne encephalitis. J Infect Dis 197:266-269.
Kirkwood, C.D., and R. Streitberg. 2008. Calicivirus shedding in children after recov-ery from diarrhoeal disease. J Clin Virol 43:346-348.
Ko, G., Z.D. Jiang, P.C. Okhuysen, and H.L. DuPont. 2006. Fecal cytokines and mark-ers of intestinal inflammation in international travelers with diarrhea due to Noroviruses. J Med Virol 78:825-828.
Koda, Y., M. Soejima, and H. Kimura. 2001. The polymorphisms of fucosyltransfer-ases. Leg Med (Tokyo) 3:2-14.
Koelle, K., S. Cobey, B. Grenfell, and M. Pascual. 2006. Epochal evolution shapes the phylodynamics of interpandemic influenza A (H3N2) in humans. Science 314:1898-1903.

64
References
Korsager, B., S. Hede, H. Boggild, B.E. Bottiger, and K. Molbak. 2005. Two outbreaks of norovirus infections associated with the consumption of imported frozen raspberries, Denmark, May-June 2005. Euro Surveill 10:E050623 050621.
Kroneman, A., L. Verhoef, J. Harris, H. Vennema, E. Duizer, Y. van Duynhoven, J. Gray, M. Iturriza, B. Bottiger, G. Falkenhorst, C. Johnsen, C.H. von Bonsdorff, L. Maunula, M. Kuusi, P. Pothier, A. Gallay, E. Schreier, M. Hohne, J. Koch, G. Szucs, G. Reuter, K. Krisztalovics, M. Lynch, P. McKeown, B. Foley, S. Coughlan, F.M. Ruggeri, I. Di Bartolo, K. Vainio, E. Isakbaeva, M. Poljsak-Prijatelj, A.H. Grom, J.Z. Mijovski, A. Bosch, J. Buesa, A.S. Fauquier, G. Her-nandez-Pezzi, K.O. Hedlund, and M. Koopmans. 2008. Analysis of integrated virological and epidemiological reports of norovirus outbreaks collected within the Foodborne Viruses in Europe network from 1 July 2001 to 30 June 2006. J Clin Microbiol 46:2959-2965.
Kurtz, J.B., T.W. Lee, and D. Pickering. 1977. Astrovirus associated gastroenteritis in a children’s ward. J Clin Pathol 30:948-952.
Larsson, M.M., G.E. Rydell, A. Grahn, J. Rodriguez-Diaz, B. Akerlind, A.M. Hutson, M.K. Estes, G. Larson, and L. Svensson. 2006. Antibody prevalence and titer to norovirus (genogroup II) correlate with secretor (FUT2) but not with ABO phenotype or Lewis (FUT3) genotype. J Infect Dis 194:1422-1427.
Le Guyader, F.S., F. Bon, D. DeMedici, S. Parnaudeau, A. Bertone, S. Crudeli, A. Doyle, M. Zidane, E. Suffredini, E. Kohli, F. Maddalo, M. Monini, A. Gallay, M. Pommepuy, P. Pothier, and F.M. Ruggeri. 2006. Detection of multiple noro-viruses associated with an international gastroenteritis outbreak linked to oyster consumption. J Clin Microbiol 44:3878-3882.
Lew, J.F., A.Z. Kapikian, X. Jiang, M.K. Estes, and K.Y. Green. 1994. Molecular char-acterization and expression of the capsid protein of a Norwalk-like virus re-covered from a Desert Shield troop with gastroenteritis. Virology 200:319-325.
Lindesmith, L., C. Moe, J. Lependu, J.A. Frelinger, J. Treanor, and R.S. Baric. 2005. Cellular and humoral immunity following Snow Mountain virus challenge. J Virol 79:2900-2909.
Lindesmith, L., C. Moe, S. Marionneau, N. Ruvoen, X. Jiang, L. Lindblad, P. Stewart, J. LePendu, and R. Baric. 2003. Human susceptibility and resistance to Norwalk virus infection. Nat Med 9:548-553.
Lindesmith, L.C., E. Donaldson, J. Leon, C.L. Moe, J.A. Frelinger, R.E. Johnston, D.J. Weber, and R.S. Baric. 2010. Heterotypic humoral and cellular immune re-sponses following Norwalk virus infection. J Virol 84:1800-1815.
Lindesmith, L.C., E.F. Donaldson, and R.S. Baric. 2011. Norovirus GII.4 strain anti-genic variation. J Virol 85:231-242.
Lindesmith, L.C., E.F. Donaldson, A.D. Lobue, J.L. Cannon, D.P. Zheng, J. Vinje, and R.S. Baric. 2008. Mechanisms of GII.4 norovirus persistence in human popula-tions. PLoS Med 5:e31.
Liu, B.L., P.R. Lambden, H. Gunther, P. Otto, M. Elschner, and I.N. Clarke. 1999. Mo-lecular characterization of a bovine enteric calicivirus: relationship to the Nor-

65
References
walk-like viruses. J Virol 73:819-825.Lochridge, V.P., and M.E. Hardy. 2003. Snow Mountain virus genome sequence and
virus-like particle assembly. Virus Genes 26:71-82.Lopman, B., B. Armstrong, C. Atchison, and J.J. Gray. 2009. Host, weather and viro-
logical factors drive norovirus epidemiology: time-series analysis of laboratory surveillance data in England and Wales. PLoS One 4:e6671.
Lopman, B., H. Vennema, E. Kohli, P. Pothier, A. Sanchez, A. Negredo, J. Buesa, E. Schreier, M. Reacher, D. Brown, J. Gray, M. Iturriza, C. Gallimore, B. Bot-tiger, K.O. Hedlund, M. Torven, C.H. von Bonsdorff, L. Maunula, M. Poljsak-Prijatelj, J. Zimsek, G. Reuter, G. Szucs, B. Melegh, L. Svennson, Y. van Dui-jnhoven, and M. Koopmans. 2004a. Increase in viral gastroenteritis outbreaks in Europe and epidemic spread of new norovirus variant. Lancet 363:682-688.
Lopman, B.A., G.K. Adak, M.H. Reacher, and D.W. Brown. 2003a. Two epidemiologic patterns of norovirus outbreaks: surveillance in England and wales, 1992-2000. Emerg Infect Dis 9:71-77.
Lopman, B.A., M. Reacher, C. Gallimore, G.K. Adak, J.J. Gray, and D.W. Brown. 2003b. A summertime peak of “winter vomiting disease”: surveillance of noro-viruses in England and Wales, 1995 to 2002. BMC Public Health 3:13.
Lopman, B.A., M.H. Reacher, I.B. Vipond, J. Sarangi, and D.W. Brown. 2004b. Clini-cal manifestation of norovirus gastroenteritis in health care settings. Clin Infect Dis 39:318-324.
Lyman, W.H., J.F. Walsh, J.B. Kotch, D.J. Weber, E. Gunn, and J. Vinje. 2009. Prospec-tive study of etiologic agents of acute gastroenteritis outbreaks in child care centers. J Pediatr 154:253-257.
Marionneau, S., F. Airaud, N.V. Bovin, J. Le Pendu, and N. Ruvoen-Clouet. 2005. Influ-ence of the combined ABO, FUT2, and FUT3 polymorphism on susceptibility to Norwalk virus attachment. J Infect Dis 192:1071-1077.
Marionneau, S., A. Cailleau-Thomas, J. Rocher, B. Le Moullac-Vaidye, N. Ruvoen, M. Clement, and J. Le Pendu. 2001. ABH and Lewis histo-blood group antigens, a model for the meaning of oligosaccharide diversity in the face of a changing world. Biochimie 83:565-573.
Marionneau, S., N. Ruvoen, B. Le Moullac-Vaidye, M. Clement, A. Cailleau-Thomas, G. Ruiz-Palacois, P. Huang, X. Jiang, and J. Le Pendu. 2002. Norwalk virus binds to histo-blood group antigens present on gastroduodenal epithelial cells of secretor individuals. Gastroenterology 122:1967-1977.
Marsh, M., and A. Helenius. 2006. Virus entry: open sesame. Cell 124:729-740.Martella, V., M. Campolo, E. Lorusso, P. Cavicchio, M. Camero, A.L. Bellacicco, N.
Decaro, G. Elia, G. Greco, M. Corrente, C. Desario, S. Arista, K. Banyai, M. Koopmans, and C. Buonavoglia. 2007. Norovirus in captive lion cub (Panthera leo). Emerg Infect Dis 13:1071-1073.
Martella, V., E. Lorusso, N. Decaro, G. Elia, A. Radogna, M. D’Abramo, C. Desario, A. Cavalli, M. Corrente, M. Camero, C.A. Germinario, K. Banyai, B. Di Martino, F. Marsilio, L.E. Carmichael, and C. Buonavoglia. 2008. Detection and mo-

66
References
lecular characterization of a canine norovirus. Emerg Infect Dis 14:1306-1308.Maunula, L., M. Roivainen, M. Keranen, S. Makela, K. Soderberg, M. Summa, C.H.
von Bonsdorff, M. Lappalainen, T. Korhonen, M. Kuusi, and T. Niskanen. 2009. Detection of human norovirus from frozen raspberries in a cluster of gastroenteritis outbreaks. Euro Surveill 14:
Mauroy, A., L. Gillet, E. Mathijs, A. Vanderplasschen, and E. Thiry. 2011. Alternative attachment factors and internalisation pathways for GIII.2 bovine noroviruses. J Gen Virol
Mauroy, A., A. Scipioni, E. Mathijs, C. Saegerman, J. Mast, J.C. Bridger, D. Ziant, C. Thys, and E. Thiry. 2009a. Epidemiological study of bovine norovirus infection by RT-PCR and a VLP-based antibody ELISA. Vet Microbiol 137:243-251.
Mauroy, A., A. Scipioni, E. Mathijs, C. Thys, and E. Thiry. 2009b. Molecular detection of kobuviruses and recombinant noroviruses in cattle in continental Europe. Arch Virol 154:1841-1845.
Mead, P.S., L. Slutsker, V. Dietz, L.F. McCaig, J.S. Bresee, C. Shapiro, P.M. Griffin, and R.V. Tauxe. 1999. Food-related illness and death in the United States. Emerg Infect Dis 5:607-625.
Medici, M.C., L.A. Abelli, I. Dodi, G. Dettori, and C. Chezzi. 2010. Norovirus RNA in the blood of a child with gastroenteritis and convulsions--A case report. J Clin Virol 48:147-149.
Meeroff, J.C., D.S. Schreiber, J.S. Trier, and N.R. Blacklow. 1980. Abnormal gastric motor function in viral gastroenteritis. Ann Intern Med 92:370-373.
Milnes, A.S., S.H. Binns, S.L. Oliver, and J.C. Bridger. 2007. Retrospective study of noroviruses in samples of diarrhoea from cattle, using the Veterinary Laborato-ries Agency’s Farmfile database. Vet Rec 160:326-330.
Mollicone, R., A. Cailleau, and R. Oriol. 1995. Molecular genetics of H, Se, Lewis and other fucosyltransferase genes. Transfus Clin Biol 2:235-242.
Mounts, A.W., T. Ando, M. Koopmans, J.S. Bresee, J. Noel, and R.I. Glass. 2000. Cold weather seasonality of gastroenteritis associated with Norwalk-like viruses. J Infect Dis 181 Suppl 2:S284-287.
Murata, T., N. Katsushima, K. Mizuta, Y. Muraki, S. Hongo, and Y. Matsuzaki. 2007. Prolonged norovirus shedding in infants <or=6 months of age with gastroenteri-tis. Pediatr Infect Dis J 26:46-49.
Ng, T.L., P.P. Chan, T.H. Phua, J.P. Loh, R. Yip, C. Wong, C.W. Liaw, B.H. Tan, K.T. Chiew, S.B. Chua, S. Lim, P.L. Ooi, S.K. Chew, and K.T. Goh. 2005. Oyster-associated outbreaks of Norovirus gastroenteritis in Singapore. J Infect 51:413-418.
Nilsson, J., G.E. Rydell, J. Le Pendu, and G. Larson. 2009. Norwalk virus-like par-ticles bind specifically to A, H and difucosylated Lewis but not to B histo-blood group active glycosphingolipids. Glycoconj J 26:1171-1180.
Nilsson, M., K.O. Hedlund, M. Thorhagen, G. Larson, K. Johansen, A. Ekspong, and L. Svensson. 2003. Evolution of human calicivirus RNA in vivo: accumulation of mutations in the protruding P2 domain of the capsid leads to structural changes

67
References
and possibly a new phenotype. J Virol 77:13117-13124.Noel, J.S., R.L. Fankhauser, T. Ando, S.S. Monroe, and R.I. Glass. 1999. Identification
of a distinct common strain of “Norwalk-like viruses” having a global distribu-tion. J Infect Dis 179:1334-1344.
Nordgren, J., E. Kindberg, P.E. Lindgren, A. Matussek, and L. Svensson. 2010. Norovi-rus gastroenteritis outbreak with a secretor-independent susceptibility pattern, Sweden. Emerg Infect Dis 16:81-87.
Nordgren, J., A. Matussek, A. Mattsson, L. Svensson, and P.E. Lindgren. 2009. Preva-lence of norovirus and factors influencing virus concentrations during one year in a full-scale wastewater treatment plant. Water Res 43:1117-1125.
Nurminen, K., V. Blazevic, L. Huhti, S. Rasanen, T. Koho, V.P. Hytonen, and T. Vesi-kari. 2011. Prevalence of norovirus GII-4 antibodies in Finnish children. J Med Virol 83:525-531.
Ohlinger, V.F., B. Haas, G. Meyers, F. Weiland, and H.J. Thiel. 1990. Identification and characterization of the virus causing rabbit hemorrhagic disease. J Virol 64:3331-3336.
Okhuysen, P.C., X. Jiang, L. Ye, P.C. Johnson, and M.K. Estes. 1995. Viral shedding and fecal IgA response after Norwalk virus infection. J Infect Dis 171:566-569.
Oliver, S.L., E. Asobayire, A.M. Dastjerdi, and J.C. Bridger. 2006a. Genomic charac-terization of the unclassified bovine enteric virus Newbury agent-1 (Newbury1) endorses a new genus in the family Caliciviridae. Virology 350:240-250.
Oliver, S.L., C.A. Batten, Y. Deng, M. Elschner, P. Otto, A. Charpilienne, I.N. Clarke, J.C. Bridger, and P.R. Lambden. 2006b. Genotype 1 and genotype 2 bovine noroviruses are antigenically distinct but share a cross-reactive epitope with human noroviruses. J Clin Microbiol 44:992-998.
Oliver, S.L., A.M. Dastjerdi, S. Wong, L. El-Attar, C. Gallimore, D.W. Brown, J. Green, and J.C. Bridger. 2003. Molecular characterization of bovine enteric calicivi-ruses: a distinct third genogroup of noroviruses (Norwalk-like viruses) unlikely to be of risk to humans. J Virol 77:2789-2798.
Park, S.I., C. Jeong, H.H. Kim, S.H. Park, S.J. Park, B.H. Hyun, D.K. Yang, S.K. Kim, M.I. Kang, and K.O. Cho. 2007. Molecular epidemiology of bovine norovi-ruses in South Korea. Vet Microbiol 124:125-133.
Parrino, T.A., D.S. Schreiber, J.S. Trier, A.Z. Kapikian, and N.R. Blacklow. 1977. Clini-cal immunity in acute gastroenteritis caused by Norwalk agent. N Engl J Med 297:86-89.
Patel, M.M., M.A. Widdowson, R.I. Glass, K. Akazawa, J. Vinje, and U.D. Parashar. 2008. Systematic literature review of role of noroviruses in sporadic gastroen-teritis. Emerg Infect Dis 14:1224-1231.
Perry, J.W., S. Taube, and C.E. Wobus. 2009. Murine norovirus-1 entry into permissive macrophages and dendritic cells is pH-independent. Virus Res 143:125-129.
Perry, J.W., and C.E. Wobus. 2010. Endocytosis of murine norovirus 1 into murine mac-rophages is dependent on dynamin II and cholesterol. J Virol 84:6163-6176.
Ponka, A., L. Maunula, C.H. von Bonsdorff, and O. Lyytikainen. 1999. An outbreak of

68
References
calicivirus associated with consumption of frozen raspberries. Epidemiol Infect 123:469-474.
Prasad, B.V., M.E. Hardy, T. Dokland, J. Bella, M.G. Rossman, M.K Estes. 1999. X-ray Crystallographic Structure of the Norwalk Virus Capsid. Science 8;286: 287-290
Prasad, B.V. 2010. Structural biology of calicivirus capsids. In Fourth International Conference on Caliciviruses. Santa Cruz, Chile.
Ramirez, S., G.M. Giammanco, S. De Grazia, C. Colomba, V. Martella, and S. Arista. 2009. Emerging GII.4 norovirus variants affect children with diarrhea in Pal-ermo, Italy in 2006. J Med Virol 81:139-145.
Ravn, V., and E. Dabelsteen. 2000. Tissue distribution of histo-blood group antigens. APMIS 108:1-28.
Raza, M.W., C.C. Blackwell, P. Molyneaux, V.S. James, M.M. Ogilvie, J.M. Inglis, and D.M. Weir. 1991. Association between secretor status and respiratory viral illness. BMJ 303:815-818.
Reeck, A., O. Kavanagh, M.K. Estes, A.R. Opekun, M.A. Gilger, D.Y. Graham, and R.L. Atmar. 2010. Serological correlate of protection against norovirus-in-duced gastroenteritis. J Infect Dis 202:1212-1218.
Richmond, S.J., E.O. Caul, S.M. Dunn, C.R. Ashley, S.K. Clarke, and N.R. Seymour. 1979. An outbreak of gastroenteritis in young children caused by adenoviruses. Lancet 1:1178-1181.
Rockx, B., M. De Wit, H. Vennema, J. Vinje, E. De Bruin, Y. Van Duynhoven, and M. Koopmans. 2002. Natural history of human calicivirus infection: a prospective cohort study. Clin Infect Dis 35:246-253.
Rockx, B.H., H. Vennema, C.J. Hoebe, E. Duizer, and M.P. Koopmans. 2005. Associa-tion of histo-blood group antigens and susceptibility to norovirus infections. J Infect Dis 191:749-754.
Ronaghi, M., S. Karamohamed, B. Pettersson, M. Uhlen, and P. Nyren. 1996. Real-time DNA sequencing using detection of pyrophosphate release. Anal Biochem 242:84-89.
Ronaghi, M., M. Uhlen, and P. Nyren. 1998. A sequencing method based on real-time pyrophosphate. Science 281:363, 365.
Rydell, G.E., J. Nilsson, J. Rodriguez-Diaz, N. Ruvoen-Clouet, L. Svensson, J. Le Pen-du, and G. Larson. 2009. Human noroviruses recognize sialyl Lewis x neogly-coprotein. Glycobiology 19:309-320.
Schreiber, D.S., N.R. Blacklow, and J.S. Trier. 1973. The mucosal lesion of the proxi-mal small intestine in acute infectious nonbacterial gastroenteritis. N Engl J Med 288:1318-1323.
Serpa, J., R. Almeida, C. Oliveira, F.S. Silva, E. Silva, C. Reis, J. Le Pendu, G. Oliveira, L.M. Ribeiro, and L. David. 2003. Lewis enzyme (alpha1-3/4 fucosyltransfer-ase) polymorphisms do not explain the Lewis phenotype in the gastric mucosa of a Portuguese population. J Hum Genet 48:183-189.
Sheinfeld, J., A.J. Schaeffer, C. Cordon-Cardo, A. Rogatko, and W.R. Fair. 1989. Asso-

69
References
ciation of the Lewis blood-group phenotype with recurrent urinary tract infec-tions in women. N Engl J Med 320:773-777.
Shirato, H., S. Ogawa, H. Ito, T. Sato, A. Kameyama, H. Narimatsu, Z. Xiaofan, T. Miyamura, T. Wakita, K. Ishii, and N. Takeda. 2008. Noroviruses distinguish between type 1 and type 2 histo-blood group antigens for binding. J Virol 82:10756-10767.
Siebenga, J.J., H. Vennema, E. Duizer, and M.P. Koopmans. 2007a. Gastroenteritis caused by norovirus GGII.4, The Netherlands, 1994-2005. Emerg Infect Dis 13:144-146.
Siebenga, J.J., H. Vennema, B. Renckens, E. de Bruin, B. van der Veer, R.J. Siezen, and M. Koopmans. 2007b. Epochal evolution of GGII.4 norovirus capsid proteins from 1995 to 2006. J Virol 81:9932-9941.
Sieczkarski, S.B., and G.R. Whittaker. 2002. Dissecting virus entry via endocytosis. J Gen Virol 83:1535-1545.
Smiley, J.R., K.O. Chang, J. Hayes, J. Vinje, and L.J. Saif. 2002. Characterization of an enteropathogenic bovine calicivirus representing a potentially new calicivirus genus. J Virol 76:10089-10098.
Soejima, M., and Y. Koda. 2005. Molecular mechanisms of Lewis antigen expression. Leg Med (Tokyo) 7:266-269.
Souza, M., M.S. Azevedo, K. Jung, S. Cheetham, and L.J. Saif. 2008. Pathogenesis and immune responses in gnotobiotic calves after infection with the genogroup II.4-HS66 strain of human norovirus. J Virol 82:1777-1786.
Stapleton, A., T.M. Hooton, C. Fennell, P.L. Roberts, and W.E. Stamm. 1995. Effect of secretor status on vaginal and rectal colonization with fimbriated Escherichia coli in women with and without recurrent urinary tract infection. J Infect Dis 171:717-720.
Stuart, A.D., and T.D. Brown. 2006. Entry of feline calicivirus is dependent on clathrin-mediated endocytosis and acidification in endosomes. J Virol 80:7500-7509.
Sugieda, M., and S. Nakajima. 2002. Viruses detected in the caecum contents of healthy pigs representing a new genetic cluster in genogroup II of the genus “Norwalk-like viruses”. Virus Res 87:165-172.
Svensson, L., and E. Kindberg. 2009. Human Genetic Factors Involved in Viral Patho-genesis. In Cellular Signaling and Innate Immune Responses to RNA Virus INfection. A.L. Brasier, editor Washington. 177-193.
Tacket, C.O., M.B. Sztein, G.A. Losonsky, S.S. Wasserman, and M.K. Estes. 2003. Humoral, mucosal, and cellular immune responses to oral Norwalk virus-like particles in volunteers. Clin Immunol 108:241-247.
Takanashi, S., S. Hashira, T. Matsunaga, A. Yoshida, T. Shiota, P.G. Tung, P. Khamrin, S. Okitsu, M. Mizuguchi, T. Igarashi, and H. Ushijima. 2009. Detection, genet-ic characterization, and quantification of norovirus RNA from sera of children with gastroenteritis. J Clin Virol 44:161-163.
Tan, M., P. Fang, T. Chachiyo, M. Xia, P. Huang, Z. Fang, W. Jiang, and X. Jiang. 2008a. Noroviral P particle: structure, function and applications in virus-host

70
References
interaction. Virology 382:115-123.Tan, M., R.S. Hegde, and X. Jiang. 2004a. The P domain of norovirus capsid protein
forms dimer and binds to histo-blood group antigen receptors. J Virol 78:6233-6242.
Tan, M., and X. Jiang. 2005. The p domain of norovirus capsid protein forms a subviral particle that binds to histo-blood group antigen receptors. J Virol 79:14017-14030.
Tan, M., and X. Jiang. 2008. Association of histo-blood group antigens with suscepti-bility to norovirus infection may be strain-specific rather than genogroup de-pendent. J Infect Dis 198:940-941; author reply 942-943.
Tan, M., and X. Jiang. 2010. Norovirus gastroenteritis, carbohydrate receptors, and animal models. PLoS Pathog 26;6(8)
Tan, M., M. Jin, H. Xie, Z. Duan, X. Jiang, and Z. Fang. 2008b. Outbreak studies of a GII-3 and a GII-4 norovirus revealed an association between HBGA pheno-types and viral infection. J Med Virol 80:1296-1301.
Tan, M., W. Zhong, D. Song, S. Thornton, and X. Jiang. 2004b. E. coli-expressed re-combinant norovirus capsid proteins maintain authentic antigenicity and recep-tor binding capability. J Med Virol 74:641-649.
Taylor, M.B., C.I. Schildhauer, S. Parker, W.O. Grabow, J. Xi, M.K. Estes, and W.D. Cubitt. 1993. Two successive outbreaks of SRSV-associated gastroenteritis in South Africa. J Med Virol 41:18-23.
Teunis, P.F., C.L. Moe, P. Liu, S.E. Miller, L. Lindesmith, R.S. Baric, J. Le Pendu, and R.L. Calderon. 2008. Norwalk virus: how infectious is it? J Med Virol 80:1468-1476.
Thiel, H.J., and M. Konig. 1999. Caliciviruses: an overview. Vet Microbiol 69:55-62.Thorven, M., A. Grahn, K.O. Hedlund, H. Johansson, C. Wahlfrid, G. Larson, and L.
Svensson. 2005. A homozygous nonsense mutation (428G-->A) in the human secretor (FUT2) gene provides resistance to symptomatic norovirus (GGII) in-fections. J Virol 79:15351-15355.
Tian, P., M. Brandl, and R. Mandrell. 2005. Porcine gastric mucin binds to recombi-nant norovirus particles and competitively inhibits their binding to histo-blood group antigens and Caco-2 cells. Lett Appl Microbiol 41:315-320.
Torrado, J., B. Ruiz, J. Garay, A. Cosme, J.I. Arenas, J.C. Bravo, E. Fontham, and P. Correa. 1997. Lewis, secretor, and ABO phenotypes, and sulfomucin ex-pression in gastric intestinal metaplasia. Cancer Epidemiol Biomarkers Prev 6:287-289.
Troeger, H., C. Loddenkemper, T. Schneider, E. Schreier, H.J. Epple, M. Zeitz, M. Fromm, and J.D. Schulzke. 2009. Structural and functional changes of the duo-denum in human norovirus infection. Gut 58:1070-1077.
van der Poel, W.H., R. van der Heide, F. Verschoor, H. Gelderblom, J. Vinje, and M.P. Koopmans. 2003. Epidemiology of Norwalk-like virus infections in cattle in The Netherlands. Vet Microbiol 92:297-309.
van Der Poel, W.H., J. Vinje, R. van Der Heide, M.I. Herrera, A. Vivo, and M.P. Koop-

71
References
mans. 2000. Norwalk-like calicivirus genes in farm animals. Emerg Infect Dis 6:36-41.
Vardy, J., A.J. Love, and N. Dignon. 2007. Outbreak of acute gastroenteritis among emergency department staff. Emerg Med J 24:699-702.
Vildevall, M., A. Charpilienne, J. Hinkula, W. Nasir, G. Larson, and L. Svensson. In Manuscript. Characterization of Bovine Norovirus Hemagglutinin.
Vildevall, M., A. Grahn, S.L. Oliver, J.C. Bridger, A. Charpilienne, D. Poncet, G. Lar-son, and L. Svensson. 2010. Human antibody responses to bovine (Newbury-2) norovirus (GIII.2) and association to histo-blood group antigens. J Med Virol 82:1241-1246.
Wadl, M., K. Scherer, S. Nielsen, S. Diedrich, L. Ellerbroek, C. Frank, R. Gatzer, M. Hoehne, R. Johne, G. Klein, J. Koch, J. Schulenburg, U. Thielbein, K. Stark, and H. Bernard. 2010. Food-borne norovirus-outbreak at a military base, Ger-many, 2009. BMC Infect Dis 10:30.
Widdowson, M.A., E.H. Cramer, L. Hadley, J.S. Bresee, R.S. Beard, S.N. Bulens, M. Charles, W. Chege, E. Isakbaeva, J.G. Wright, E. Mintz, D. Forney, J. Massey, R.I. Glass, and S.S. Monroe. 2004. Outbreaks of acute gastroenteritis on cruise ships and on land: identification of a predominant circulating strain of norovi-rus--United States, 2002. J Infect Dis 190:27-36.
Widdowson, M.A., B. Rockx, R. Schepp, W.H. van der Poel, J. Vinje, Y.T. van Duyn-hoven, and M.P. Koopmans. 2005. Detection of serum antibodies to bovine norovirus in veterinarians and the general population in the Netherlands. J Med Virol 76:119-128.
Widerlite, L., J.S. Trier, N.R. Blacklow, and D.S. Schreiber. 1975. Structure of the gastric mucosa in acute infectious bacterial gastroenteritis. Gastroenterology 68:425-430.
Wise, A.G., S.S. Monroe, L.E. Hanson, D.L. Grooms, D. Sockett, and R.K. Maes. 2004. Molecular characterization of noroviruses detected in diarrheic stools of Michigan and Wisconsin dairy calves: circulation of two distinct subgroups. Virus Res 100:165-177.
Woode, G.N., and J.C. Bridger. 1978. Isolation of small viruses resembling astroviruses and caliciviruses from acute enteritis of calves. J Med Microbiol 11:441-452.
Wyatt, R.G., R. Dolin, N.R. Blacklow, H.L. DuPont, R.F. Buscho, T.S. Thornhill, A.Z. Kapikian, and R.M. Chanock. 1974. Comparison of three agents of acute infec-tious nonbacterial gastroenteritis by cross-challenge in volunteers. J Infect Dis 129:709-714.
Xi, J.N., D.Y. Graham, K.N. Wang, and M.K. Estes. 1990. Norwalk virus genome clon-ing and characterization. Science 250:1580-1583.
Zahorsky, J. 1929. Hyperemesis hiemis or the winter vomiting disease. Arch Pediatr 46:391-395.
Zakhour, M., H. Maalouf, I. Di Bartolo, L. Haugarreau, F.S. Le Guyader, N. Ruvoen-Clouet, J.C. Le Saux, F.M. Ruggeri, M. Pommepuy, and J. Le Pendu. 2010. Bovine norovirus: carbohydrate ligand, environmental contamination, and po-

72
References
tential cross-species transmission via oysters. Appl Environ Microbiol 76:6404-6411.
Zakhour, M., N. Ruvoen-Clouet, A. Charpilienne, B. Langpap, D. Poncet, T. Peters, N. Bovin, and J. Le Pendu. 2009. The alphaGal epitope of the histo-blood group antigen family is a ligand for bovine norovirus Newbury2 expected to prevent cross-species transmission. PLoS Pathog 5:e1000504.
Zheng, D.P., T. Ando, R.L. Fankhauser, R.S. Beard, R.I. Glass, and S.S. Monroe. 2006. Norovirus classification and proposed strain nomenclature. Virology 346:312-323.














![[CiliciaJP 2015] The Puzzle of Consciousness](https://img.pdfslide.net/doc/110x75/63320f0a8d2c463a5800b02c/ciliciajp-2015-the-puzzle-of-consciousness.jpg)














