Embed Size (px)
Citation preview

The Origins of the Co. Kildare Dormouse
Deborah G. Glass1, Dawn M. Scott1, Daniel Donoher2, Andrew D.J. Overall1
1Division of Biology, University of Brighton, Brighton, BN2 4GJ, UK. Corre-
sponding author email: [email protected].
2Kildare Animal Foundation, South Green Road, Kildare Town, County Kildare.
1

ABSTRACT
The Hazel dormouse Muscardinus avellanarius has recently been discovered in
Co. Kildare Ireland and is being treated as non-native and, hence, potentially inva-
sive. The mode of arrival and origins are unknown, initial theories included transferal
via hay from Great Britain. The population in Kildare appears to be thriving and
as native populations throughout England are in decline, the success is of broad con-
servation relevance. To gain insight into the recent demography of the Co. Kildare
population, the cytochrome b gene was profiled for individuals from Kildare, Eng-
land and Wales and compared to known haplotypes throughout its European range.
The individuals from Co. Kildare shared the same previously unrecorded haplo-
type, which was placed in a separate clade from the GB haplotype, along with more
southern European populations. The French haplotype was the most recent common
ancestor, albeit with a 3 base pair di↵erence. Inferring either the dormouse has been
present in Ireland for longer than expected, or is derived from a recent source pop-
ulation yet to be sampled. The latter is considered the most likely scenario. Future
work and concerns of the Hazel dormouse in Ireland are discussed.
2

1 Introduction
The hazel dormouse Muscardinus avellanarius has recently been observed in Ireland,
with the first reported sighting located within County Kildare in 2010 (Marnell et al,
2013). How the dormouse arrived in Ireland and from where it derived is currently
unknown. Muscardinus avellanarius is otherwise distributed widely across Europe,
from the Mediterranean to Southern Sweden and eastwards towards Russia, but ex-
cluding Iberia. The range also extends to parts of Asia Minor (Juskaitis, 2007). The
northwest limit of its European range is in Great Britain (GB), where it is found
over much of Southern England, with some isolated populations in northern Wales
(Bright et al. 2006). In parts of its Northern range (UK, Netherlands, Sweden,
Germany and Denmark) populations are declining; as a consequence they are pro-
tected under the European Habitats directive annex IV and the Bern convention
Appendix III (ICUN 2014). There are no historical records of the presence of the
dormouse in Ireland; hence they are considered as non-native and potentially inva-
sive. In addition, Invasive species are recognised as an important element in global
change (Occhipinti-Ambrogi 2007), therefore it is important to closely monitor any
non-native species within their new environment.
Mainland Britain and Ireland have strikingly di↵erent biota, despite being neigh-
bouring islands (Martınkora et al, 2007; McDevitt et al, 2011). Britains flora and
fauna is broadly similar to nearby areas of continental Europe, although it is some-
what restricted (Martınkora et al, 2007). The distinctness between Irish and British
fauna is particularly clear in mammals (Yalden 1999). Native small mammal species
of mainland Britain consist of: four mouse species, four vole species, four shrew
species, one squirrel species and the hazel dormouse (Yalden 1999; Masceretti et al,
2003). Ireland in comparison is species poor, with only five small mammal species
3

in total, the pygmy shrew, the woodmouse, the bank vole, the greater white-toothed
shrew and the red squirrels (Yalden 1999; Masceretti et al, 2003; Martınkora et al,
2007; McDevitt et al, 2011; Montgomery et al, 2011). Although it is extremely
di�cult to establish the status of mammals in Ireland perceived as native, due to
uncertainties of natural reinvaders after the last glaciation (Montgomery et al, 2011),
the pygmy shrew, the woodmouse and the red squirrel are considered to be native
and the rest recent invaders (Yalden 1999; Martınkora et al, 2007; McDevitt et al,
2011; Montgomery et al, 2011).
Whether intentional or accidental, the means of introduction is uncertain. Initial
theories of potential arrival routes include the transfer of individuals via hay, through
the horticultural trade and horse racing industry (Marnell et al, 2013). The extensive
horse-breeding and racing industry, in Co. Kildare results in the regular tra�c of
horse boxes back and forth throughout GB (ibid). There have been further possible
sightings in a number of locations including, Dublin, Laois, Tipperary, Clare, Galway,
Mayo, Kerry and Westmeath (Mooney 2013; Sheehy & Lawton, 2014), although
the only confirmed sightings are of 14 individuals within a 29.6 km2 area of the
Newbridge/Naas region of Co. Kildare (Sheehy & Lawton, 2014).
The hazel dormouse is a nocturnal, arboreal mammal that mainly inhabits areas
of deciduous forest with a thick layer of underbrush, although it has been found in
mixed deciduous-coniferous forests and in hedgerows (Bright et al. 2006). The shy,
arboreal and nocturnal nature of the hazel dormouse in addition to their typically
low population density, makes them notoriously di�cult to observe in their native
range (Bright & Morris 1996; Bright et al. 2006, Naim et al. 2011), thus it may
be possible that the species has been present in the Republic of Ireland for longer
than records indicate. The earliest historical record is of an attempted introduction
by the well-known Irish naturalist R.M. Barrington, who tried to introduce six hazel
4

dormice, sourced from London, into Fassaroe, Co. Wicklow in 1885. However, no
further evidence of them was recorded and it has since been assumed that they failed
to become established (Fitter 1959).
Given that the native populations throughout England are in decline, some appre-
ciation of the apparent recent success of the Irish dormouse is of broad conservation
relevance. To this end it was considered necessary to establish what the current ge-
netic diversity of the Kildare population is, which o↵ers some indication of the small-
est size of the common ancestral population the e↵ective population size. This, in
turn provides some indication of whether the current population has grown up from
few (single introductory event) or many (multiple independent introductions). Ge-
netic profiling involved the analysis of a single variable region of the mitochondrial
DNA, the cytochrome b region, which has been profiled for hazel dormouse popu-
lations across their European range (Mouton et al, 2012). Mutational di↵erences
between cytochrome b haplotypes are understood to correlate with time since they
shared a recent ancestry (Avise, 2004). Consequently, similarity of haplotypes is
expected to identify the most likely source population.
2 Materials & Methods
2.1 Sample Collection
Samples were collected from Kildare, Ireland; from Sussex, the Isle of Wight and
South Devon, England and from north Wales (see table 1). All tissue samples were
provided from animals which died of natural causes. Those from the UK were col-
lected by volunteers involved with the dormouse monitoring scheme (see acknowl-
edgments). The GB hair samples were collected under a Natural England licence by
5

the authors. Hair and tissue samples from the Kildare individuals were provided by
Kildare Animal Foundation, Kildare, Ireland.
DNA was extracted from tissue (ear or tail) samples using the DNeasy Tissue
Kit (QIAGEN) following the manufacturers instructions. Hair sample extraction
required an initial digestion step to maximise DNA yield, following the protocol
by Pfie↵er et al. (2004), before continuing with the DNeasy Kit. A master mix
containing 182.6µl of PCR grade water, 34µl of Tris-HCL (100mM), 34µl of sodium
chloride (100mM), 1.02µl of calcium chloride (3mM), 68µl of 10% SDS, 13.6µl of
40mM DTT and 6.8µl of 250µg/ml Protienase K per sample was added to a 2ml
Eppendorf tube and vortexed for 30 seconds. Each hair sample (>10 hairs with
visible root) was placed in a 1.5ml Eppendorf tube, to which 340µl of the extraction
solution was added and voretexed for 5 seconds. The sample was then transferred to
a water bath set at 56 �C and left to incubate for 2 hours (or until the sample was
fully dissolved), vortexting every half hour. Once the hair was dissolved, the DNeasy
Tissue Kit was used from step 3 of the manufacturers instructions.
2.2 Mitochondrial DNA Analysis
In order to locate the most likely source population for the Kildare dormouse, we
utilized a recent study by Mouton et al, (2012) that provided sequence data from
a 704 base pair (bp) region of the cytochrome b gene. This study identified 33
haplotypes located throughout the M. avellanarius range, excluding Britain. New
sequences were obtained from the Kildare, English and Welsh populations, the latter
two such that the nearest geographical neighbours to Ireland were included in the
analysis.
Table 1
6

Table 1 Geographic locations, corresponding lineages and Genbank accession num-
bers of GB and Irish haplotypes.
A 704 bp fragment of the cytochrome b region of the mtDNA was amplified by
PCR using primers: LMA14255, 5-TGGTGGAATTTCGGTTCTCT 3; RMA15192,
5- GTTGCCTCCAATTCATGTT-3 (Mouton et al, 2012). Amplifications were car-
ried out in 25µl reactions containing 5ng of DNA template and final concentrations
of 15nM 10 x reaction bu↵er, 1.5mM MgCl2, 0.2mM for each dNTP, 10 pmol forward
and reverse primers, 1unit PlatinumTaq DNA polymerase (Invitrogen). Cycling was
performed in Techne TC-4000 Thermal cycler employing 40 cycles (30-45 s at 94 �C,
30 45 s at 50 �C and 45-90 s at 72 �C), with a final extension at 72 �C for 10 minutes.
Sequencing procedures were performed by SourceBioscience (Nottingham, England).
MEGA 6 (Tamura et al, 2013) software was employed to generate sequence align-
ments and pairwise genetic distances. MrBayes (version 3.2.2, Ronquist et al, 2012)
was used to construct Bayesian Markov chain Monte Carlo (MCMC) phylogenetic
trees in order to establish the evolutionary relationships among all haplotypes, for
which we ran 50,000 generations, but otherwise the default priors. The garden dor-
mouse (Eliomys quercinus) and edible dormouse (Glis glis) were used as out-groups,
in keeping with the Mouton et al, (2012) study and the Gliridae molecular phy-
logeny (Montgelard et al, 2003). Minimum spanning networks were generated using
Arlequin (ver. 3.5, Exco�er et al, 2005) to calculate connection values, which were
then input into HapStar (Teacher & Gri�ths, 2011).
7

3 Results
All six individuals from Kildare, Ireland, had identical cytochrome b sequences (Gen-
bank accession number KJ631620). A Nucleotide BLAST search (NCBI) found the
closest match to be haplotype 20 (assession number FN796772), which relates to
the Mouton et al, (2012) French sample. All individuals sampled throughout GB
had identical haplotypes, identical to Polish haplotype 15 within the Central North
Europe lineage (Lithuania, Poland, Germany). A consensus haplotype, GB 15, is
included in the phylogenetic tree to highlight the genetic distance between the geo-
graphic neighbours, GB and Ireland (Fig 1). The phylogenetic analysis placed the
Irish haplotype within Western Europe lineage containing haplotypes from France,
Belgium and Switzerland. The Irish haplotype was 3 mutational steps from the
French haplotype 20 (Fig 1(b)).
Figure 1
Figure 1 (a) Bayesian phylogenetic analysis of Cytochrome b haplotypes of Mus-
cardinus avellanarius, including those from Mouton et al, (2012) and including sup-
port values. Geographic origins are followed by haplotype number. (b) Detail of
minimum spanning network corresponding to the mutational steps between haplo-
types. The dotted lines connect to the remainder of the network, which is identical
to Figure 3 in Mouton et al, (2012).
Genetic distances (table 2) show the Irish haplotype to have the greatest similarity
to the French (0.4%). The genetic distances between the Irish and the Central
Italian and Central - North Europe lineages is considerably higher at 3.9% and 9.5%
respectively.
Table 2
8

Table 2 Cytochrome b genetic distance (%).
4 Discussion
Phylogenetic analysis of the cytochrome b region of the mtDNA produced a number
of revelations. Firstly, the Kildare population shares a recent common ancestry
amongst the Western Europe lineage, whereas the GB population clusters amongst
the Central - North Europe lineage. Our Bayesian tree topology (Fig 1(a)) is identical
to that of Mouton et al, (2012), although our minimum spanning network (Fig 1(b))
connects the Western Europe lineage to the Central Europe lineage via Belgium,
as opposed to the Swiss haplotype 19; otherwise the network is identical to that of
Mouton et al, (2012). The Irish haplotype di↵ers from its closest relative, the French,
at three nucleotide sites. This could suggest that the dormice have been established
in Ireland for a long enough period to accrue this degree of divergence from its closest
neighbours. But, given the lack of observational data, this seems highly unlikely. In
addition, a recent survey study concluded that, based upon dispersal distances, the
hazel dormouse has probably been present for just seven year - since 2007 (Sheehy
& Lawton, 2014). The unique haplotype means that the specific source population
remains unresolved for the time being. However, the phylogenetic analysis places
it amongst the more western, mainland, European populations. The initial theory
that the dormice entered Kildare through bale transport from GB is looking unlikely,
due to the fact that the GB haplotype was so diverse with 68 nucleotide di↵erences.
Lack of di↵erentiation amongst the GB samples (Welsh, South west, South and South
East), makes it unlikely that the source population is from an unsampled region from
GB. Hay importation from other European countries, particularly those of the south
9

and west, may provide the route of entry, but this remains to be established.
Further sampling will be necessary in order to ascertain with less uncertainty the
number of entries into the country. As just six samples from the Irish population
have been sequenced, it would be prudent not to overstate the results. Although
a single founding population from Western Europe is currently supported, further
data may modify this conclusion.
4.1 Future concerns for the hazel dormouse
Our current understanding of the ecology of the hazel dormouse suggests that a
rapid spread throughout Ireland is an unlikely scenario. Muscardinus avellanarius
individuals generally do not disperse very far (Bright & Morris 1991, Bright & Morris
1992, Juskaitis, 2008) and are reluctant to cross open ground (Bright & Morris 1991,
Bright & Morris 1992) and corridor gaps (Bright 1998). Rare inter-patch movement
over 1km has been recorded using genetic techniques (Naim et al, 2011), showing
the occasional long distance movement. Habitat corridor use (such as hedgerows)
is widely recognised to facilitate movement and dispersal between habitat patches
(Baum et al, 2004). Thus, it may be possible that such corridors have facilitated
dormouse dispersal throughout Kildare. Recent sighting-surveys have estimated a
presence in this region for at least seven years (Sheehy & Lawson, 2014).
All information on M. avellanarius comes from their native range, thus it is possi-
ble that the environment within Ireland could give them some competitive advantage
that allows them to be extremely successful in their recipient community. It has been
suggested that the subtle di↵erences between the Irish and British hedgerows may
be responsible for the suggested rapid spread of the dormouse, possibly resulting in
faster breeding than within their native range (Mooney 2013).
10

Invasive species are often defined as those that have an adverse e↵ect on their
recipient community. Life history traits of non-native species are crucial factors in
determining a species ability to become invasive in new environments (Osman &
Whitlatch 2007; Alcarlo et al, 2008). The reproductive biology of invasive species
is often characterised by high fecundity, more than one generation a year, and flex-
ible life history events (Alcarlo et al, 2008). Given the recognised characteristics
of invasive species, M. avellanarius is unlikely to become a pest in Ireland. The
hazel dormouse has a relatively low reproductive output compared to other rodent
species, with a maximum of two broods per year (only occurring when conditions
are optimal), and an average of 2-4 juveniles in each litter within their native range
(Juskaitis 2008). Climate is the main limiting factor of hazel dormice populations.
Hibernation through the winter months (Bright et al, 1996) is essential to their sur-
vival, limiting the time period in which they can reproduce. The need to reach
optimal weights before breeding and hibernation (Juskaitis 2008) combined with use
of torpor during adverse weather conditions (cold and wet) (Bright et al, 1996), fur-
ther restricts the time in which they can forage and breed. Even with the possibility
of a higher food supply in the areas they have been introduced, the climate in Ire-
land is similar to areas in GB. With climate and weather as a limiting factor to the
flexibility of M. avellanarius life history events, a population explosion in Ireland
would be unlikely. However, it is possible that the hazel dormouse could impact neg-
atively on the recipient community through interspecific interactions. Inavasional
meltdown is the process by which the invasion of one non-native species is facilated
by another non-native species, compounding their independant impacts on native
species, communities and ecosystems (Montgomery et al, 2011). This has already
been demonstrated in small mammal communities in Ireland. Montgomery et al,
(2011) found bank vole abundance (first invader) to be positively influenced by the
11

presence of the greater white-toothed shrew (second invader). They also found that
the greater white-toothed shrew had negative and compounding e↵ects on the abun-
dance of wood mice and the occurrence of pygmy shrews (both considered native
species). However, the rapid rate of expansion of the greater white-toothed shrew
over the past seven years is thought to be partly through human-assisted dispersal
(McDevitt et al., 2014), whereas the hazel dormouse is not associated with humans.
Monitoring the dispersal and growth of hazel dormouse over the next few years, even
if a negative impact is unexpected, is going to be crucial in o↵ering a glimpse of the
likelihood of expansion and possibility of interaction with native communities.
AcknowledgmentsWe would like to thank Jen Bousfield, Adam Grogan, Laurie
Jackson, Jenny Macpherson, Stephen Robinson, Janet Rose and Ian White for their
help in collecting samples.
Literature Cited
Alcorlo P., Geiger W. & Otero M. (2008) Reproductive biology and life cycle of
the invasive crayfish Procambarus clarkii (Crustacea: Decapoda) in diverse aquatic
habitats of South-Western Spain: Implications for population control, Fundamental
and Applied Limnology, 173: 197-212
Avise, J.C. 2004. Molecular Markers, Natural History, and Evolution. Sinauer,
USA.
Baum A., Haynes K.J., Dillemuth F.P. & Cronin J.T. (2004) The matrix enhances
the e↵ectiveness of corridors and stepping stones, Etiology 85: 2671-2676
Bright P.W. & Morris P.A. (1991) Ranging and nesting behaviour of the dor-
mouse, Muscardinus avellanarius, in diverse low-growing woodland, The zoological
12

society of London, 224, 177-190
Bright P.W. & Morris P.A. (1992) Ranging and nesting behaviour of the dor-
mouse, Muscardinus avellanarius, in coppice-with-standards woodland, The zoolog-
ical society of London, 226, 589-600
Bright P.W. & Morris P.A. (1996) Why are dormice rare? A case study in con-
servation biology, Mammal Review, 26: 157-187
Bright P.W., Morris P.A. & Mitchell J.T. (2006) The dormouse conservation
handbook, English Nature, Great Britain
Exco�er, L., Laval, G., and Schneider, S. 2005 Arlequin ver. 3.0: An integrated
software package for population genetics data analysis. Evolutionary Bioinformatics
Online 1:47-50.
Freeland J.R (2005) Molecular Ecology, John Wiley & Sons Ltd, Chichester,
England
Fitter, R.S.R. 1959 The Ark in Our Midst: The Story of the Introduced Animals
of Britain. Collins, UK.
Juskaitis R. (2007) peculiarities of habitats of the common dormice (Muscardinus
avellanarius) in Lithuania: a review, Folia Zoologica, 56: 337-348
Juskaitis R. (2008) The common dormouse Muscardinus avellanarius : ecology,
population structure and dynamics, Institute of Ecology of Vilnius University Pub-
lishers, Vilnius.
McDevitt, A.D., Vega, R., Rambau, R.V., Yannic, G., Herman, J.S., Hayden T.J.
and Searle, J.B. 2011 Colonization of Ireland: revisitng the Pygmy shrew syndrome
using mitochondrial, Y chromosomal and microsatellite markers. Heredity 107:548-
557.
McDevitt, A.D., Montgomery, W. I., Tosh, D. G., Lusby, J., Reid, N., White, T.
A., McDevitt, C. D., O’Halloran, J., Searle, J. B., Yearsley, J. M. 2014 Invading and
13

Expanding: Range Dynamics and Ecological Consequences of the Greater White-
Toothed Shrew (Crocidura russula) Invasion in Ireland. PLoS ONE 9: e100403
Marnell F, Donoher D, Sheehy E, Lawton C (2013) Irish Naturalists’ Journal.
33(1):78-79
Martankora, N., McDonald, R.A. and Searle, J.B. 2007. Stoats (Mustela er-
minea) provide evidence of natural overland colonization of Ireland. Proceeding of
the Royal Society B 274: 1387-1393
Mascheretti, S., Rogatcheva M.B., Gndz, I., Fredga, K. and Searle, J.B. 2003.
How did pygmy shrews colonize Ireland? Clues from a phylogenic analysis of mito-
chondrial cytochrome b sequences. Proceeding of the Royal Society B 270: 1593-1599
Montgelard, C., Matthee, CA, and Robinson, TJ. 2003. Molecular systematics of
dormice (Rodentia: Gliridae) and the variation of Graphiurus in Africa. Proceedings
of the Royal Society of London Series B Biological Sciences 270: 1947-1955.
Montgomery, W.I. , Lundy, M.G. and Reid, N. 2011. Invasional meltdown: evi-
dence for unexpected consequences and cumulative impacts of multispecies invasions.
Biological Invasions 14:(6) 1111-1125
Mooney J (2013) Rare UK dormouse moves to Ireland, The Sunday Times, 8
September [http://www.thesundaytimes.co.uk/sto/news/ireland/article1310764.ece]
Mouton A et al. (2012) Evidence of a complex phylogeographic structure in
the common dormouse, Muscardinus avellanarius (Rodentia: Gliridae), Biological
Journal of the Linnean Society. 105: 648-664.
Naim D, Telfer S, Tatman S, Bird S, Kemp S.J., Hughes R & Watts P.C (2011)
Patterns of genetic divergence among populations of the common dormouse, Mus-
cardinus avellanarius in the UK, Molecular Research Notes
Occhipinti-Ambrogi. A. 2007 Global change and marine communities: Alien
species and climate change. Marine Pollution Bulletin 55: 342-352
14

Osman R.W. & Whitlatch R.B. (2007) Variation in the ability of Didemnum
sp. To invade established communities, Journal of Experimental Marine Biology and
Ecology, 342; 40-53
Pfie↵er, I., Vlkel, I., Tubert, H. & Brenig, B. (2004) Forensic DNA-typing of dog
hair: DNA -extraction and PCR amplification, Forensic Science International 141;
149-151
Sheehy, E. and Lawton, C. 2014 Distribution of the non-native Hazel Dormouse
(Muscardinus avellanarius) in Ireland. Irish Naturalists Journal. In Press.
Suchard, M.A. and Huelsenbeck, J.P. 2012 MrBayes 3.2: E�cient Bayesian Phy-
logenetic Inference and Model Choice Across a Large Model Space. Syst Biol. 61:
539-542.
Tamura K, Stecher G, Peterson D, Filipski A, and Kumar S (2013) MEGA6:
Molecular Evolutionary Genetics Analysis Version 6.0. Molecular Biology and Evo-
lution 30: 2725-2729.
15
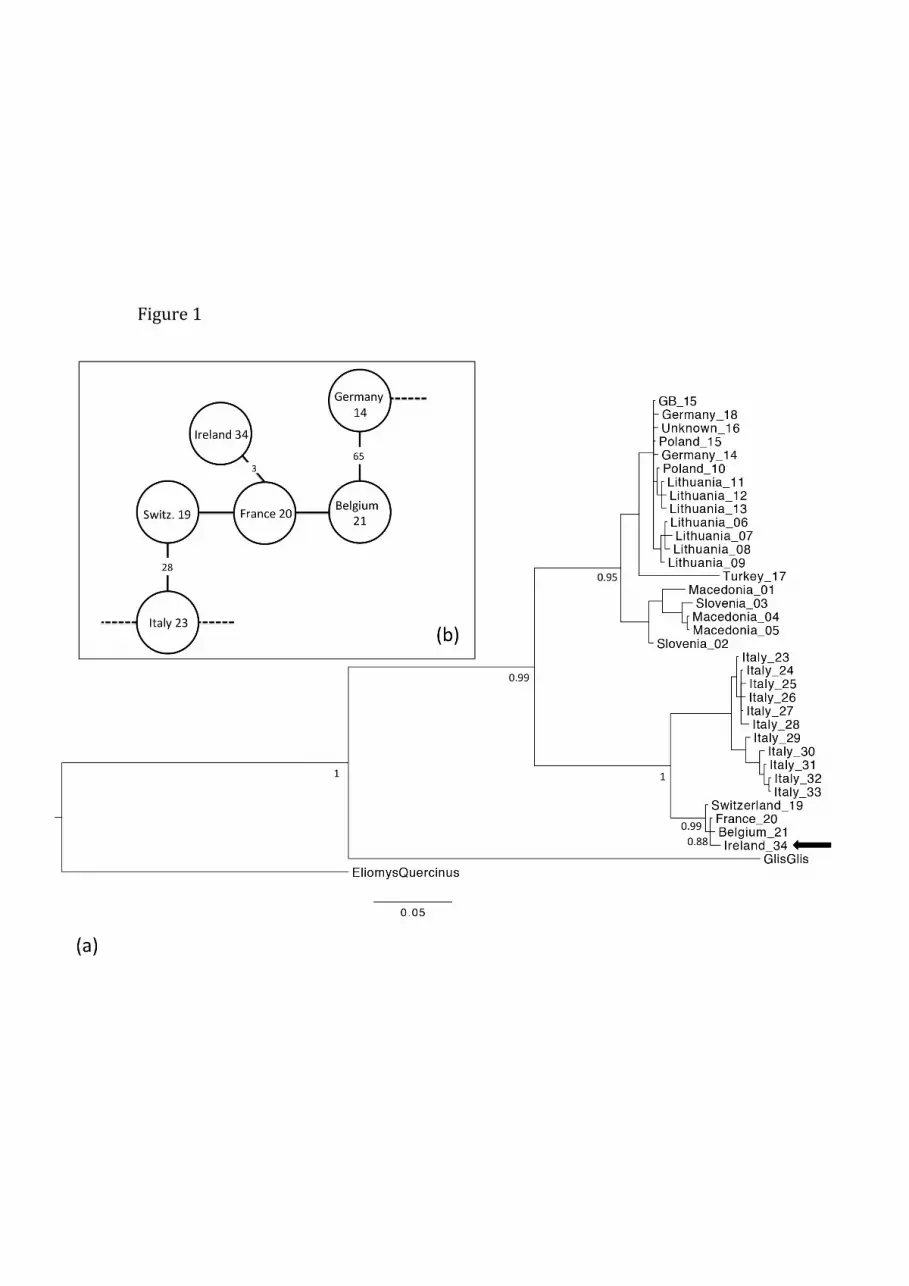
Figure 1

Table 1 Geographic Origin Lineage Number Haplotype Genbank of animals accession Great Britain, South East England (Sussex) Central Northern 15 15 FN796767* Great Britain, Southern England (Isle of Wight) Central Northern 5 15 Great Britain, South West England (Devon) Central Northern 2 15 Great Britain, North Wales (Carmarthen) Central Northern 1 15 Ireland, Co. Kildare Western 6 34 KJ631620 * From Mouton et al, (2012) Table 2 Ireland_34 France_20 Belgium_21 Switz_19 Italy_23 France_20 0.4 Belgium_21 0.5 0.1 Switz_19 0.5 0.1 0.3 Italy_23 3.9 3.8 3.9 3.6 Germany_14 9.5 9.3 9.2 9.2 9.8





























