Embed Size (px)
Citation preview

The Role of Prefrontal Cortex in Predictive Fear Learning
Teri M. Furlong, Sindy Cole, Adam S. Hamlin, and Gavan P. McNallyThe University of New South Wales
Pavlovian fear conditioning depends on prediction error, or the discrepancy between actual and expectedoutcomes. We used immunohistochemistry, neuronal tract tracing, and reversible inactivation to study therole of prefrontal cortex and thalamocortical pathways in predictive fear learning. Unexpected, but notexpected, conditioned stimulus (CS)–unconditioned stimulus (US) presentations caused increased c-Fosexpression in the prefrontal cortex (PFC), midline thalamus, lateral amygdala, as well as retrogradelabeled midline thalamic afferents to PFC. Reversible inactivation of dorsomedial PFC, but not infral-imbic PFC, prevented the associative blocking of fear learning. These results suggest a role fordorsomedial PFC (dmPFC), and a thalamic 3 dmPFC pathway, in signaling whether or not aversiveevents are expected or unexpected and so whether they are to be learned about.
Keywords: anterior cingulate, prefrontal cortex, orbitofrontal cortex, thalamus, prediction error
A signature of adaptive behavior is the use of past experience toguide future learning. This has been formalized in error-correctinglearning rules, such as the Rescorla-Wagner model (Rescorla &Wagner, 1972), which state that learning is caused by a discrep-ancy between the actual and expected outcomes of a conditioningtrial, or prediction error. In the case of fear learning, this is thediscrepancy between the aversive unconditioned stimulus (US)that is actually delivered and the US that is expected based on priorconditioned stimulus (CS)–US pairings. Learning occurs preferen-tially to surprising outcomes and their antecedents at the expenseof expected ones. When the expectation matches the outcome (i.e.,there is no prediction error), no learning occurs.
There has been significant progress in understanding the neuralmechanisms of fear memory formation. It is widely accepted thatthe amygdala is critical for the formation and storage of fearmemories produced by conditioning and is likewise important forthe expression of conditioned fear responses (e.g., Kim & Jung,2006; Maren & Quirk, 2004). In contrast, very little is knownabout the neural mechanisms for predictive fear learning—that is,how fear learning about a CS and US on a given trial is influencedby past learning about those stimuli. Indeed, the available evidencehas identified only the midbrain periaqueductal gray (PAG) andthe nucleus accumbens (Acb) in predictive fear learning (forreview, see McNally & Westbrook, 2006). This evidence suggeststhat the PAG determines effectiveness of the US as an aversivereinforcer (e.g., Cole & McNally, 2007), whereas the Acb deter-
mines effectiveness of the CS as a predictor of the US (Iordanova,McNally, & Westbrook, 2006).
Several lines of evidence suggest that the prefrontal cortex(PFC), especially dorsomedial PFC (dmPFC), may also be a crit-ical component of the neural circuit underlying predictive fearlearning. For example, glutamatergic activation of dmPFC pro-duces conditioned place aversion, raising the possibility thatdmPFC might be important for learning about unexpected aversiveUSs (Johansen & Fields, 2004). dmPFC neurons are recruitedfollowing errors in behavioral performance, and the magnitude ofdmPFC neuronal activity during behavioral error is correlated withthe extent of subsequent behavioral adjustment (e.g., Narayanan &Laubach, 2008). In general, rodent, primate, and human PFC havebeen implicated in monitoring and detecting errors in behavior(e.g., Carter & van Veen, 2007; Histed, Pasupathy, & Miller, 2009;Matsumoto, Matsumoto, Abe, & Tanaka, 2007; Miller & Cohen,2001; Taylor, Stern, & Gehring, 2007). Yet whether PFC is im-portant for learning from errors in prediction is unknown.
The midline and intralaminar thalamus, including paraventricu-lar thalamus, centromedial nucleus, rhomboid nucleus, and reuni-ens nucleus may also be important for predictive fear learning.These regions receive projections from the PAG (Krout & Loewy,2000) and, in turn, project extensively to rodent prefrontal cortexwith collaterals to the striatum (including Acb) and amygdala(Berendse & Groenewegen, 1991; Groenewegen & Berendse,1994). The midline and intralaminar thalamus are therefore wellpositioned to modulate cortico-amygdala as well as cortico-striatalinteractions, and thus contribute to predictive fear learning. How-ever, the functions of midline and intralaminar thalamic regionsare only poorly understood (Groenewegen & Berendse, 1994). Ofinterest, recent imaging studies in rats (Holschneider et al., 2006)and humans (Dunsmoor, Bandettini, & Knight, 2008) have shownrecruitment of midline thalamus during fear learning and presen-tations of fear CSs.
In these experiments we studied the role of PFC, midline thalamus,and thalamocortical projections in predictive fear learning using twocomplementary approaches. First, we studied recruitment of PFC, itsthalamic afferents, and other brain regions following expected or
Teri M. Furlong, Sindy Cole, Adam S. Hamlin, and Gavan P. McNally,School of Psychology, The University of New South Wales, Sydney,Australia.
This work was supported by Grant DP0877430 from the AustralianResearch Council to GPM. GPM is an Australian Research Council QEIIFellow (DP0877430).
Correspondence concerning this article should be addressed to Gavan P.McNally, School of Psychology, The University of New South Wales,Sydney, NSW 2052, Australia. E-mail: [email protected]
Behavioral Neuroscience © 2010 American Psychological Association2010, Vol. 124, No. 5, 574–586 0735-7044/10/$12.00 DOI: 10.1037/a0020739
574

unexpected CS–footshock US presentations. To do this, we combineddetection of the activity marker c-Fos with detection of the retrogradeneuronal tracer cholera toxin b subunit (CTb). Then, we studied thecausal role of PFC in predictive fear learning through use of ablocking procedure (Kamin, 1968). Blocking is the principal proce-dure for assessment of predictive learning. In Stage I of a blockingprocedure, CSA is paired with shock. In Stage II, a neutral CSB isconditioned in compound with CSA. Fear learning to CSB is blockedby CSA. There is no prediction error during Stage II: the US isexpected because of Stage I training so CSB is not learned about.Taken together the results reported here show that PFC is critical forpredictive fear learning.
Experiment 1
In Experiment 1, we used immunohistochemistry and neural tracttracing to study the neural correlates of predictive fear learning.Specifically, we studied the neural correlates of expected versusunexpected CS–US presentations using a three-group design. GroupExpected received CS–US pairings each day for 4 days during train-ing and also on test. Group Unexpected was placed in the samecontext but received no stimulus events during training, and receivedCS–US pairings during test. Thus, Groups Expected and Unexpectedreceived physically identical CS–US presentations on test. GroupControl was placed in the same context. Half the rats in this groupreceived CS-alone presentations during training and again on test. Theremaining rats in this group received no stimulus events duringtraining and received CS-alone presentations on test. Hence, half ofGroup Control received the CS at training and test, like GroupExpected, and the other half experienced the CS for the first time attest like Group Unexpected. We used c-Fos immunohistochemistry tostudy the regions recruited by expected versus unexpected CS–USpresentations, and we used retrograde tracing from PFC to studyrecruitment of midline and intralaminar thalamic afferents to PFCduring these presentations.
Method
Subjects. Subjects were experimentally naive male Wistarrats (250–350 g) obtained from Monash Animal Services (Gipps-land, Victoria, Australia). Rats were housed in groups of 8 inplastic cages with ad lib access to food and water. The colonyroom was maintained at 21 °C on a 12 h light/dark cycle (lights on7 a.m.). Rats were handled each day for 3 days to habituate to theexperimenter. The procedures were approved by the Animal Careand Ethics Committee at the University of New South Wales andconducted in accordance with the National Institute of HealthGuide for the Care and Use of Laboratory Animals (PublicationsNo. 80–23) revised 1996. The procedures were designed to min-imize the number of animals used and their suffering.
Apparatus. Rats were trained and tested in a set of fourconditioning chambers (24 � 30 � 21 cm; Med Associates). Thechambers had a grid floor consisting of steel rods 4 mm indiameter spaced 15 mm apart, which were wired to a constant-current generator. Each chamber was located in a sound-attenuating box illuminated by an infrared LED and equipped witha fan (background noise approximately 65dB), speaker and digitalvideo camera connected to a digital multiplexer and DVD recorderin an adjacent laboratory to record each session. Between each use,
the chambers were wiped down with water and the bedding be-neath the grid floor changed.
Tracer microinjection. Half the rats (Group Expected, n � 6;Group Unexpected, n � 6; Group Control, n � 4) received a micro-injection of the retrograde tracer, CTb, into the right dmPFC 10 daysprior to training. Surgery was conducted under anesthesia (Ketamine,100 mg/kg, ip, and Xylazine, 6 mg/kg, ip) combined with an analgesic(Carprofen, 5 mg/kg, sc) and antibiotics (Procaine penicillin, 30mg/kg, im, and Cephazolin sodium, 3.5 mg/kg, im). Rats were placedin a stereotaxic frame, the skull exposed and the incisor bar adjusteduntil the heights of lambda and bregma were equal. A 30-gaugeneedle attached to a 1-�l Hamilton syringe was lowered into dmPFC(anterior-posterior (AP) � 3.72 mm, medial-lateral (ML) � �0.5mm, dorsal ventral (DV) � �3.2 mm) according to the atlas ofPaxinos and Watson (2005). Manual injections of 40 nl of 1% low saltCTb (List Biological Laboratories) were made over 5 min with astereotaxic injector, which was left in place for an additional 2 min topermit diffusion of the injectate. Rats were monitored daily followingsurgery.
Behavioral procedure. There were four daily 10-min trainingsessions. During each session, Group Expected (n � 12) receivedtwo CS–US pairings beginning at 3 min and 8 min into the session.The CS was an 82 dB (A Scale), 750 Hz tone (0.1 s rise and fall)for 30 s, which coterminated with the 0.8 mA, 1-s footshock US.Group Unexpected (n � 12) was exposed to the chamber for equalduration without CS or US presentations. The control group (n �8) received no US presentations at any stage. Half of this group(n � 4) received two CS alone presentations each day duringtraining, while the other was exposed only to the chamber duringtraining. All stimulus events were controlled by a computer (Lab-View, National Instruments).
The test was a single 10-min session. At test, Group Expectedand Group Unexpected received two CS–US pairings beginning at3 min and 8 min into the session in the manner described above.The control group received two CS alone presentations at the sametimes.
Immunohistochemistry and neuronal quantification. Twohours after the commencement of test, rats were deeply anesthe-tized with sodium pentobarbital and transcardially perfused withheparinized saline (0.9%) and paraformaldehyde (4%). The brainswere then removed, postfixed (1 hr), and cytoprotected in sucrose(48 hr).
The entire forebrain and midbrain were sectioned using a cry-ostat (Leica Microsystems). Every fourth section was collected (40�m thick, 160 �m apart) to form a series and processed forc-Fos-immunoreactivity (IR). Sections were prepared by rinsing inalcohol (50%), alcohol containing hydrogen peroxide (3%) andnormal horse serum (5%, NHS) in 0.1M phosphate buffer (pH 7.4,PB) for 30 min each. Incubation in rabbit antic-Fos (1:1000, SantaCruz Biotechnology, 48 hr) was followed by incubation in biotin-ylated donkey antirabbit (1:1000; Jackson Immunoresearch Labora-tories, 24 hr), both in 2% NHS and 0.2% Triton X in PB (at 4 °C).Sections were then incubated in ExtrAvidin peroxidase (1:1000,Sigma-Aldrich, 2.5 hr at RT) and a nickel intensified 3,3-diaminobenzidine (DAB) solution to reveal black IR. After 5 min,glucose oxidase (735 units/ml DAB solution, Sigma-Aldrich, 10min) was added to the DAB solution, which contained 0.05%DAB, 0.2% D-glucose, 0.004% ammonium chloride and 0.004%nickel sulfate in a 0.1M tris buffer (pH � 7.6). The procedure was
575PREFRONTAL CORTEX AND PREDICTING DANGER

the same to reveal brown CTb-IR, except that the sections wereincubated in both the rabbit antic-Fos and goat anti-CTb (1:5000;List Biological Laboratories) and the DAB procedure was repeatedwithout nickel intensification after incubation in donkey antisheep(1:1000; Jackson Immunoresearch Laboratories, 24 hr). Finally,sections were mounted onto gelatin-coated slides, cleared withhistolene, and coverslipped with Entellan.
Counting. c-Fos-IR, CTb-IR, and double labeled CTb-IR/c-Fos-IR were counted using a transmitted light microscope at 10�magnification. Counts were made at the microscope across con-secutive sections of the collected series on one side of the brain ina number of regions. Regions were delineated using clearly visiblelandmarks according to Paxinos & Watson (2005, 2007) unlessotherwise stated. When landmarks did not clearly delineate re-gions, boundaries were created with a 10 � 10 grid reticule (1mm � 1 mm) located in the right eye piece of the microscope. Tengraduations of the grid were equivalent to 0.1 mm on the section.
Counts were made across consecutive sections on one side ofthe brain in a number of regions. Regions were delineated accord-
ing to Paxinos & Watson (2005, 2007) unless otherwise stated.Figure 1 shows the regions counted.
PFC (4 sections, between bregma 3.64 mm and 3.00 mm).Medial: For the anterior part of the cingulate area (Cg), a grid(0.1 mm (h) � 0.05 mm (w)) was aligned with the dorsal-medial edge of the section. For the prelimbic cortex (PL), thegrid was aligned with the dorsal aspect of the forceps minor(fmi) and a 0.12 mm (h) area between the medial edge of thesection and the fmi created. Similarly, for the infralimbic cortex(IL), a 0.08 mm (h) area was created with the grid aligned at thepoint where the fmi and the edge of the section become parallel.Lateral: For the orbital frontal cortex (OFC), the grid (0.1mm � 0.1 mm) was placed against the ventral-medial edge ofthe claustrum, and for the rostral agranular insular cortex(RAIC) against the lateral edge of the section, dorsal to therhinal fissure.
Acb (5 sections, between bregma 1.92 mm and 1.12 mm).The shell could be differentiated from the septum by appear-ance of the tissue. The shell (AcbSh) and core (AcbC) were
Figure 1. Areas counted for c-Fos-immunoreactivity (c-FOS-IR) and delineations used. Squares indicate areascounted via grid reticule (1 mm � 1 mm). Landmarks used to delineate boundaries of counted areas are indicatedas dots. See Method for a full description of c-Fos-IR counting procedure. Figure in parts adapted from “The RatBrain in Stereotaxic Coordinates, 5th ed.,” by George Paxinos and Charles Watson. Copyright 2005 by Paxinosand Watson. Reprinted with permission of Elsevier.
576 FURLONG, COLE, HAMLIN, AND MCNALLY

separated by a line extending from the lateral ventricle (LV),rounding the anterior commisure (ac) by half the width of theac. The boundary of the AcbC (from dorsally located putamen)then continued half the ac laterally and dorsally to the acreaching the LV.
Amygdala (6 sections, between �2.40 mm and �3.36 mm).The lateral part of the amygdala was restricted within the triangu-lar shaped region of the external capsule. The lateral amygdala(La) was then separated from the basolateral amygdala (BLA) bya horizontal line through the center of the central amygdala (CeA).
Thalamus and epithalamus (6 sections, between bregma�2.40 mm and �3.36 mm). Midline: The dense, dark labelingof the paraventricular thalamic nucleus (PVT) and intermediodor-sal nucleus (IMD) appeared distinct from the adjacent medialdorsal thalamus (MD). The PVT and IMD were separated wherethe PVT narrows to meet IMD (PVT: 10 sections, between bregma�1.76 mm and �3.36 mm). The dense/dark labeling of the rhom-boid thalamic nucleus (Rh) was easily recognizable adjacent to thesubmedius thalamic nucleus (Sub). The reuniens (Re) thalamicnucleus was bordered dorsally and laterally by the Rh and Sub, andventrally by the ventral aspect of the mammilliothalamic tract (mt).
Intralaminar (10 sections, between bregma �1.76 mm �3.36and mm). The intralaminar nuclei formed a semicircular patternbordering the MD. The centromedian thalamic nucleus (CM) wasrestricted laterally by a vertical line through the lateral edge of theSub.
Habenula (6 sections, between bregma �2.4 mm and �3.36mm). The lateral habenula (LHb) was restricted to the bound-aries created by the stria medullaris and fasicular retroflexus.
Hypothalamus (6 sections, between bregma �2.4 mm and�3.36 mm). The dorsal part of the hypothalamus was restricteddorsally by the dorsal aspect of the third ventricle (3V), andventrally by the ventral aspect of the f. The dorsomedial (DM)hypothalamus was formed by the area between the 3V and medialedge of the mt, the perifornical hypothalamus (PeF) between themt and lateral to the f (approximately half the width of the f lateralto f), and the lateral hypothalamus (LH), extending to the medialedge of the internal capsule.
Mesencephalon: Periaqueductal gray (PAG). The ventro-lateral PAG (vlPAG) was formed by area bordered approximatelyby the mesencephalon 5 nucleus and trochlear nerve (9 sections,�7.08 mm to �8.52 mm). The lateral (lPAG) and dorsolateral(dlPAG) PAG were separated by a line extending approximately45° from the dorsal-lateral corner of the aqueduct (Aq), and thedlPAG and dorsomedial (dmPAG) separated by a line extendingperpendicularly, approximately half way along the dorsal-lateraledge of the aq (dmPAG/lPAG, 8 sections, �7.08 mm to �8.36mm, dlPAG, 7 sections, �7.08 mm to �8.20 mm).
Ventral tegmental area (VTA). The rostral part of the VTA(5 sections, from �5.16 mm to �5.96 mm) was restricted laterallyby the substantia nigra, dorsally by the red nucleus and ventrallyby the edge of the section.
Caudal pontine reticular oral (PnO; 3 sections, from �7.72mm to �8.20 mm). The grid (0.1 mm � 0.1 mm) was aligneddorsally with the subpeducular tegmentum/superior cerebellar pe-duncle and laterally with the medial edge of the rubrospinal tract.This part of the PnO includes the rostral part of the mesopontinerostromedial tegmental nucleus (RMTg).
Data analysis. The percentages of observations scored asfreezing (defined as the absence of movement other than thatrequired for respiration) were analyzed using a planned orthogonalcontrast testing procedure with the Type I error rate set at 0.05 foreach contrast. A multivariate approach to repeated measures wastaken (O’Brien & Kaiser, 1985). IR counts in each region weresummed across sections. Dual-labeled IR was expressed as rawcounts and as a percentage of the total CTb-IR within each region.These data were analyzed by using planned orthogonal contrasts,with the Type I error rate set at 0.05 for each contrast.
Results
One rat (Group Expected) was excluded because of the absenceof CTb-IR at the injection site. The statistical analyses showed thatthere were no differences between the two control conditions(those that received the tone CS for the first time on test and thosethat had received tone CS alone presentations during training andtest) in either behavior or c-Fos-IR so the data were collapsed intoa single group, “Group Control” for all analyses. Thus, final groupsizes were Group Expected n � 11 (5 rats with CTb), GroupUnexpected n � 12 (6 rats with CTb), and Group Control n � 8(4 rats with CTb).
Behavior. Figure 2A shows the mean (�SEM) levels of freez-ing during training and test. Group Expected acquired conditionedfear to the auditory CS across training. There was significantlymore freezing during the first CS presentation each day thanduring the pre-CS period, F(1, 10) � 102.3, p � .05, and freezingduring the first CS presentation increased significantly across daysof training, F(1, 10) � 48.1, p � .05. No freezing was observed inGroups Unexpected and Control during training.
On test, Group Expected showed overall significantly morefreezing than Groups Unexpected and Control, F(1, 28) � 79.3,p � .05. There was no overall difference between Group Unex-pected and Control, F(1, 28) � 1.7, p � .05. There was morefreezing during CS presentations than during the pre-CS period,F(1, 28) � 13.8, p � .05, and this difference was greatest forGroup Expected, F(1, 28) � 10.5, p � .05. Of interest, GroupUnexpected showed a significantly greater increase in freezing tothe CS between Trial 1 and Trial 2 compared with Groups Controland Expected, F(1, 28) � 6.2, p � .05. These results show thepresence of conditioned fear in Group Expected and the acquisi-tion of fear in Group Unexpected.
Immunohistochemistry. Several cortical regions showedprofiles of c-Fos-IR consistent with modulation by expectancy:Cg, PL, OFC, and RAIC (Figure 2, B–D). In each of these regions,c-Fos-IR was greatest in Group Unexpected compared withGroups Expected and Control, Fs (1, 28) � 9.5, 8.8, 6.1, and 7.1,respectively, all p � .05. In each of these regions, there was nosignificant difference in c-Fos-IR between Groups Expected andControl, Fs (1, 28) � 1.5, 1.1, 0.7, and 0.5, respectively, all p �.05. There were no significant differences between groups inc-Fos-IR in IL, Fs (1, 28) � 1, p � .05.
The midline and intralaminar thalamus, including CM, IMD,Rh, and Re, also showed profiles of c-Fos-IR consistent withmodulation by expectancy (Figure 2, B–D; Table 1). In each ofthese regions c-Fos-IR was greatest in Group Unexpected com-pared with Groups Expected and Control, Fs (1, 28) � 7.5, 4.3,4.3, and 11.5, respectively, all p � .05. In each of these regions,
577PREFRONTAL CORTEX AND PREDICTING DANGER

Figure 2. (A) Mean and SEM levels of freezing during training and test. (B) Representative photomicrographsof c-Fos-immunoreactivity (c-FOS-IR) in cingulate area (Cg), rostral agranular insular cortex (RAIC), andcentromedial thalamus/Rhomboid nucleus (CM/Rh) for Groups Expected and Unexpected. (C) Schematic ofrepresentative distributions of c-Fos-IR in prefrontal cortex and thalamus (each hemisphere shows actualc-Fos-IR of two animals). (D) Mean and SEM counts of c-Fos-IR in prefrontal cortex and midline thalamus fromGroups Expected, Unexpected, and Control. � Unexpected � Expected � Control; p � .05.
578 FURLONG, COLE, HAMLIN, AND MCNALLY

there was no significant difference in c-Fos-IR between GroupsExpected and Control, Fs (1, 28) � 0.1, 0.8, 0.1, and 1.4, respec-tively, all p � .05. However, absolute levels of c-Fos-IR were lowin IMD and Re (Table 1).
The only other brain regions which showed profiles of c-Fos-IRconsistent with modulation by expectancy were lateral amygdalaand lateral hypothalamus (Table 1). For both regions, c-Fos-IRwas significantly greater in Group Unexpected than Groups Ex-pected and Control, Fs (1, 28) � 12.6, 13.2, respectively, p � .05,which did not differ from each other, Fs (1, 28) � 2.1, 3.3,respectively, p � .05. Other regions showed significantly greaterc-Fos-IR in groups receiving CS–US presentations compared withjust CS alone presentations (Table 1). These were habenula, ac-cumbens shell, PeF, rostromedial tegmental nucleus, and ventro-lateral PAG. In each of these regions, Groups Unexpected andExpected did not differ from each other, Fs (1, 28) � 2.8, 0.1, 1.7,0.2, 0.9, respectively, p � .05, but showed significantly greaterc-Fos-IR compared with Group Control, Fs (1, 28) � 6.9, 7.6, 4.4,4.6, 6.6, respectively, p � .05. There were no differences betweengroups in the other brain regions examined (Table 1).
We next examined c-Fos-IR in thalamic projections to PFC.Injection of CTb into dmPFC (Figure 3A) produced retrogradelabeling which was most pronounced and consistent in CM (Figure3B and 3C). There was CTb-IR in PVT, Rh, Re, and CL, but thisretrograde labeling was variable across rats. There was minimalCTb-IR in IMD. For CM, there were no differences between
groups in total number of CTb-IR neurons, Fs (1, 12) � 1, p � .05(Group Expected: M � 129.2, SEM � 16.3; Group Unexpected:M � 122.8, SEM � 11.1; Group Control: M � 121.8, SEM �14.7). This shows equivalent retrograde labeling between groups.Of note, there were differences between groups in dual-labeledCTb-IR/c-Fos-IR. There were significantly more dual-labeled neu-rons in Group Unexpected compared with Groups Expected andControl, F(1, 12) � 8.5, p � .05 (Figure 2D), whereas GroupsExpected and Control did not differ from each other, F(1, 28) �1.2, p � .05. These group differences were also observed whenanalyses were based on the percentages of CTb-IR neurons ex-pressing c-Fos-IR. Group Unexpected had a significantly greaterpercentage of CTb-IR/c-Fos-IR dual-labeled neurons comparedwith Groups Expected and Control, F(1, 12) � 14.8, p � .05,which did not differ from each other, F(1, 28) � 1.7, p � .05(Figure 3D).
Discussion
c-Fos expression in PFC as a consequence of CS–US presenta-tions depended on the subject’s past experience. If CS–US pre-sentations were unexpected, a robust increase of c-Fos-IR wasdetected throughout PFC including in Cg, PL, OFC, and RAIC. Bycontrast, if these same CS–US presentations were expected, therewas no significant increase in c-Fos-IR compared with a groupreceiving the CS alone. The exception to these observations was
Table 1Mean and SEM Numbers of c-Fos-Immunoreactivity (c-Fos-IR) Neurons in Groups Expected,Unexpected, and Control
Group expected Group unexpected Group control
ThalamusHabenulaa 81.7 (9.7) 58.2 (11.5) 33.6 (8.1)PVT 110.5 (13.7) 145.2 (16.9) 115.8 (17.0)IMDb 5.4 (1.5) 15.1 (3.7) 9.6 (3.6)
AccumbensAcbSha 141.9 (20.3) 149.8 (14.2) 81.9 (17.6)AcbC 65.4 (10.8) 83.8 (11.7) 61.9 (12.2)
AmygdalaLAb 40.0 (5.5) 91.6 (10.3) 61.1. (13.3)BLA 84.2 (12.1) 67.6 (11.8) 43.4 (12.2.)CeA 32.8 (4.8) 50.8 (7.0) 49.3 (14.1)
HypothalamusLHb 81.4 (11.1) 112.5 (11.5) 52.9 (6.4)PeF 147.9 (24.4) 179.6 (20.8) 107.3 (11.6)DMH 179.1 (25.4) 183.6 (22.8) 125 (13.6)
MidbraindmPAG 69.7 (7.5) 74.4 (8.9) 71.4 (12.3)dlPAG 65.5 (9.9) 64.9 (8.4) 57.8 (8.2)lPAG 147.5 (16.4) 169.8 (17.8) 120.5 (15.2)vlPAGa 211.8 (25.2) 241.3 (23.6) 148.1 (14.1)PnO/RMTga 42.8 (4.9) 46.8 (7.4) 27.6 (4.8)VTA 43.7 (8.0) 40.7 (6.4) 23.9 (5.3)
Note. PVT � paraventricular thalamic nucleus; IMD � intermediodorsal nucleus; AcbSh � nucleus accum-bens shell; AcbC � nucleus accumbens core; LA � lateral amygdala; BLA � basolateral amygdala; CeA �central amygdala; LH � lateral hypothalamus; PeF � perifornical hypothalamus; DMH � dorsomedialhypothalamus; dmPAG � dorsomedial periaqueductal grey; dlPAG � dorsolateral periaqueductal grey; lPAG �lateral periaqueductal grey; vlPAG � ventrolateral periaqueductal grey; PnO/RMTg � pontine reticular oral/mesopontine rostromedial; VTA � ventral tegmental area.a Structures showing conditioned stimulus–unconditioned stimulus (CS–US) effect (Expected � Unexpected �Control). b Structures showing an expectancy effect (Expected � Control � Unexpected).
579PREFRONTAL CORTEX AND PREDICTING DANGER
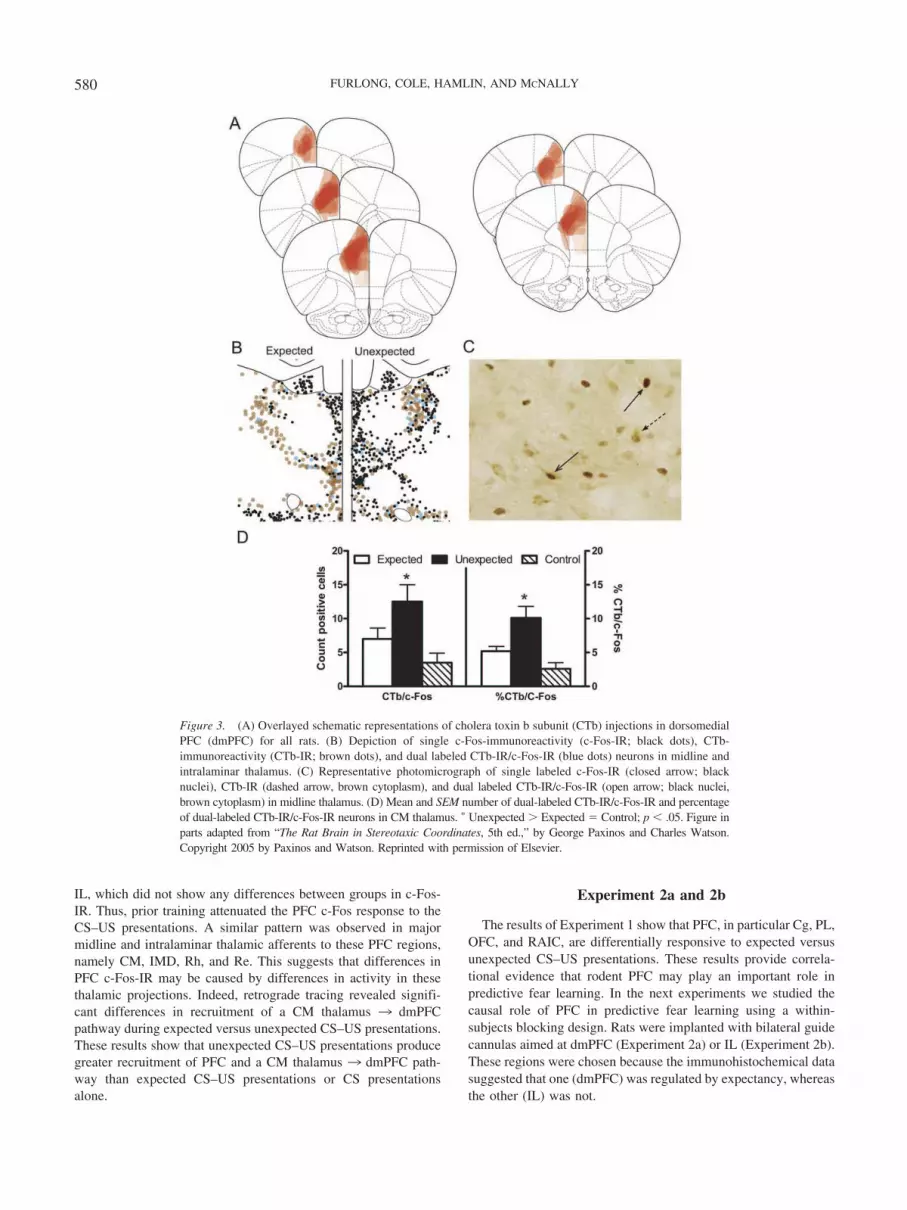
IL, which did not show any differences between groups in c-Fos-IR. Thus, prior training attenuated the PFC c-Fos response to theCS–US presentations. A similar pattern was observed in majormidline and intralaminar thalamic afferents to these PFC regions,namely CM, IMD, Rh, and Re. This suggests that differences inPFC c-Fos-IR may be caused by differences in activity in thesethalamic projections. Indeed, retrograde tracing revealed signifi-cant differences in recruitment of a CM thalamus 3 dmPFCpathway during expected versus unexpected CS–US presentations.These results show that unexpected CS–US presentations producegreater recruitment of PFC and a CM thalamus 3 dmPFC path-way than expected CS–US presentations or CS presentationsalone.
Experiment 2a and 2b
The results of Experiment 1 show that PFC, in particular Cg, PL,OFC, and RAIC, are differentially responsive to expected versusunexpected CS–US presentations. These results provide correla-tional evidence that rodent PFC may play an important role inpredictive fear learning. In the next experiments we studied thecausal role of PFC in predictive fear learning using a within-subjects blocking design. Rats were implanted with bilateral guidecannulas aimed at dmPFC (Experiment 2a) or IL (Experiment 2b).These regions were chosen because the immunohistochemical datasuggested that one (dmPFC) was regulated by expectancy, whereasthe other (IL) was not.
Figure 3. (A) Overlayed schematic representations of cholera toxin b subunit (CTb) injections in dorsomedialPFC (dmPFC) for all rats. (B) Depiction of single c-Fos-immunoreactivity (c-Fos-IR; black dots), CTb-immunoreactivity (CTb-IR; brown dots), and dual labeled CTb-IR/c-Fos-IR (blue dots) neurons in midline andintralaminar thalamus. (C) Representative photomicrograph of single labeled c-Fos-IR (closed arrow; blacknuclei), CTb-IR (dashed arrow, brown cytoplasm), and dual labeled CTb-IR/c-Fos-IR (open arrow; black nuclei,brown cytoplasm) in midline thalamus. (D) Mean and SEM number of dual-labeled CTb-IR/c-Fos-IR and percentageof dual-labeled CTb-IR/c-Fos-IR neurons in CM thalamus. � Unexpected � Expected � Control; p � .05. Figure inparts adapted from “The Rat Brain in Stereotaxic Coordinates, 5th ed.,” by George Paxinos and Charles Watson.Copyright 2005 by Paxinos and Watson. Reprinted with permission of Elsevier.
580 FURLONG, COLE, HAMLIN, AND MCNALLY

In both experiments, rats were subjected to fear conditioning.During Stage I rats received pairings of CSA with the footshockUS (A�). During Stage II rats received pairings of a compound CScomposed of CSA and CSB with shock (AB�) and also compoundpairings of the compound CS CD with shock (CD�). The next dayrats were tested for fear reactions to CSB and CSD. This proceduregenerates distinct prediction errors for AB and CD during Stage II.The prediction error for AB is small because A was paired withshock during Stage I. The presence of CSA generates an expect-ancy of shock which blocks fear learning to CSB during Stage II.By contrast, the prediction error for CD is large and positivebecause neither CSC nor CSD were paired with shock duringStage I. The shock is surprising and conditions fear to CSC andCSD during Stage II. Blocking of fear learning is shown by lessfear to CSB than CSD on test. The question of interest was whetherthis blocking would be prevented by reversible inactivation ofeither dmPFC (Experiment 2a) or IL (Experiment 2b) prior toStage II training.
Method
Subjects and apparatus. Subjects were experimentally naive,male Wistar rats obtained from the same source and maintained underthe same conditions as described for Experiment 1. There were 16 ratsin Experiment 2a and 28 rats in Experiment 2b. Rats were trained andtested in the chambers described for Experiment 1. Two auditorystimuli and two visual stimuli served as CSs throughout. An 82-dB (AScale) 750-sHz tone (0.1-s rise and fall) and an 82-dB (A scale) 20-Hzclicker served as CSB and CSD, respectively, in a counterbalancedfashion. A constant or flashing (4 Hz) presentation of a white fluo-rescent light producing illumination levels of 75 cd/m2 within thechambers served as CSA and CSC, respectively, in a counterbalancedfashion. The light was mounted on the ceiling of each box, immedi-ately above the conditioning chamber. All CSs were 30 s in durationand during Stage I and Stage II training coterminated with the 0.5 mA,1 s footshock US.
Surgery. Bilateral guide cannulas were implanted intodmPFC 5 days prior to training. Surgery was conducted underanesthesia as described for Experiment 1. Rats were placed in astereotaxic frame and a double 26 gauge guide cannula (PlasticsOne, Roanoke, VA) was targeted bilaterally at either dmPFC(AP � 3.1 mm, ML � �0.5 mm, and DV � �2.5 mm) orinfralimbic PFC (IL) (AP � 2.9 mm, ML � �0.5 mm, and DV ��4.5 mm) according to the atlas of Paxinos and Watson (2005).Cannulas were fixed in position with acrylic cement anchored tostainless steel screws fixed to the skull. After surgery, dummycannulas were placed in the guide cannulas and rats were moni-tored daily.
Procedure. Stage I occurred on Days 1–3. During each 21min 10 s session, rats received four presentations of CSA cotermi-nating with the footshock US. The intertrial interval (ITI) wasrandom, ranging from 60 to 360 s, with a mean of 211 s. Stage IIconsisted of a single 22-min session on Day 4. Rats received twocounterbalanced compound presentations each of AB and CDcoterminating with footshock. Immediately prior to this training,0.5 �l of either saline (0.9% wt/vol) or baclofen/muscimol (B/M)(1.0 nM/0.1 nM) was microinjected bilaterally into the dmPFC(Saline, n � 8; B/M n � 8) or IL PFC (Saline, n � 14; B/M n �14). Rats were exposed to the conditioning chamber for 10 min, 4
to 6 hr after conditioning sessions on Days 1–4 to reduce levels ofcontextual conditioning. On Day 5, rats were tested for levels offreezing to CSB and CSD. After a 180-s adaptation period, therewere two presentations each of CSB and CSD in a counterbalancedorder. The ITI between CS presentations was 30 s. Freezingbehavior was scored during each CS presentation every 2 s.
Histology. To confirm cannula placements, rats were over-dosed with sodium pentobarbital and their brains removed. Thebrains were then sectioned coronally (40-�m thick) using a cryo-stat. At the level of dmPFC, every second section was collectedonto a glass slide, stained with cresyl violet and coverslipped.Cannula placements were plotted onto Adobe illustrator templates(Paxinos & Watson, 2005) with the use of a microscope. Rats withplacements outside dmPFC or IL were excluded from the analyses.
Data analysis. The percentages of observation scored asfreezing (defined as the absence of movement other than thatrequired for respiration) were analyzed using a planned orthogonalcontrast testing procedure with the Type I error rate set at 0.05 foreach contrast. A multivariate approach to repeated measures wastaken (O’Brien & Kaiser, 1985).
Results and Discussion
Histology. Figure 4A shows the location of the microinjectioncannula tips in dmPFC and IL. Fourteen rats were excluded fromthe analyses due to misplaced cannulas (1 misplaced from dmPFCand 13 misplaced from IL). Thus, final group sizes were: dmPFCGroup Saline, n � 8, dmPFC Group B/M, n � 7; IL Group Saline,n � 8 and IL Group B/M, n � 7.
Experiment 2a. The results for dmPFC inactivation in Exper-iment 2a are shown in Figure 4B. There were low levels of pre-CSfreezing (defined as freezing during the first 3 min of each con-ditioning session) throughout the experiment. Group Saline didshow significantly more pre-CS freezing on Day 4 than GroupB/M, F(1, 13) � 6.2, p � .05; mean Group Saline � 11%, meanGroup B/M � 4%, but the groups did not differ on any other dayincluding test, Fs (1, 13) � 1, p � .05.
The left panel of Figure 4B shows mean (�SEM) levels offreezing during the first presentation of CSA on each day of StageI training. There were no differences between groups in freezing toCSA during Stage I, F(1, 13) � 1, p � .05, and data are collapsedacross groups for Stage I in Figure 4B. Fear accrued to CSA acrossthe course of training as shown by a significant linear increase infreezing, F(1, 13) � 176.5, p � .05.
The middle panel of Figure 4B shows mean (�SEM) levels offreezing during the two presentations of the AB and CD compoundsin Stage II. From inspection, it is clear that fear was higher to AB thanCD at the start of Stage II training (because CSA had previously beenpaired with shock) and that fear to CD increased across the course ofStage II training, reaching levels identical to those displayed to the ABcompound. There was no overall difference between groups duringStage II, F(1, 13) � 1, p � .05. Thus, B/M infusion into dmPFC didnot affect expression of freezing. There was a significant main effectof stimulus type (AB vs. CD), so that there was more freezing to theAB compound than to CD during Stage II, F(1, 13) � 6.6, p � .05.This did not interact with group, F(1, 13) � 1, p � .05. There was nomain effect of day, F(1, 13) � 2.6, p � .05, but there was a group �day interaction so that the increase in freezing across Stage II wasgreater for B/M infused animals. Finally, there was a significant
581PREFRONTAL CORTEX AND PREDICTING DANGER

Figure 4 A, Location of microinjection cannula tips in dorsomedial PFC (dmPFC; gray) and infralimbic cortex(IL; black dots) for rats receiving infusion of saline or baclofen/muscimol (B/M). (B) Mean and SEM levels offreezing during Stage I, II, and Test after infusion of saline or B/M into dmPFC. (C) Mean and SEM levels offreezing during Stage I, II, and Test after infusion of saline or B/M into IL. � B � D; p � .05.
582 FURLONG, COLE, HAMLIN, AND MCNALLY

interaction between day and CS, F(1, 13) � 17.7, p � .05, so that theincrease in freezing across Stage II was significantly greater for CDcompared with AB which was not different between groups, F(1,13) � 1, p � .05. This confirms the acquisition of fear to CD.
The data of primary interest are those from test presentations ofCSB and CSD and these are shown in the right panel of Figure 4B.There was no overall difference between Group B/M and GroupSaline, F(1, 13) � 4.1, p � .05. This shows that B/M infusions intodmPFC prior to Stage II did not have any overall effect on fearlearning or the consolidation of fear memories. There was overallevidence for blocking of fear learning because there was significantlyless fear to CSB on test than CSD, F(1, 13) � 52.4, p � .05. Of note,the group � CS interaction was significant, F(1, 13) � 11.2, p � .05.This shows that B/M infusions into dmPFC prior to Stage II preventedthe associative blocking of fear learning. To further confirm thisinterpretation, there was significantly more freezing to the blockedCS, CSB, in Group B/M than Group Saline, F(1, 13) � 7.3, p � .05,but there was no difference between groups in freezing to the controlCS, CSD, F(1, 13) � 1, p � .05.
Experiment 2b. The results for IL inactivation in Experiment2b are shown in Figure 4C. Again, there were low levels of pre-CSfreezing throughout the experiment. Group Saline did show sig-nificantly more pre-CS freezing on Day 4 of the experiment thanGroup B/M, F(1, 13) � 5.4, p � .05; mean Group Saline � 14%,mean Group B/M � 7%, but the groups did not differ on any otherday including test, F(1, 13) � 1, p � .05.
There were no differences between groups in freezing to CSAduring Stage I, F(1, 13) � 1, p � .05, and data are collapsed acrossgroups for Stage I in Figure 4C. Fear accrued to CSA across thecourse of Stage I training as shown by a significant linear increase infreezing, F(1, 13) � 123.5, p � .05. The middle panel of Figure 4Dshows mean (�SEM) levels of freezing during the two presentationsof the AB and CD compounds in Stage II. There was no overalldifference between groups during Stage II, F(1, 13) � 1.7, p � .05.There was no significant main effect of CS type (AB vs. CD), F(1,13) � 2.8, p � .05. There was no group � CS Type interaction, F(1,13) � 2.1, p � .05. There was no main effect of day, F(1, 13) � 1.5,p � .05, and there was no group � day interaction (F(1, 13) � 1, p �.05). However, there was a CS Type � day and a CS Type � day �group interaction showing that the increase in freezing across Stage IIwas significantly greater for CD compared with AB, F(1, 13) � 25.2,p � .05, and this was greater for saline than B/M infused animals,F(1, 13) � 5.6, p � .05.
The data of primary interest are those from test presentations ofCSB and CSD and these are shown in the right panel of Figure 4C.There was no overall difference between Group B/M and GroupSaline, F(1, 13) �1, p � .05. This shows that B/M infusions intoIL prior to Stage II did not have any overall effect on fear learningor the consolidation of fear memories. There was overall evidencefor blocking of fear learning because there was significantly lessfear to CSB on test than CSD, F(1, 13) � 79.8, p � .05. There wasno group � CS interaction, F(1, 13) � 2.9, p � .05. This showsthat B/M infusions into IL prior to Stage II did not prevent theassociative blocking of fear learning. To further confirm thisresult, there was no significant difference in freezing betweengroups to either the blocked CS, CSB, F(1, 13) � 1, p � .05, orthe control CS, CSD, F(1, 13) � 1, p � .05.
General Discussion
These experiments studied the role of rodent PFC in predictivefear learning. In Experiment 1 we combined immunohistochemicaldetection of the activity marker c-Fos with the retrograde tracerCTb, to study brain regions differentially responsive to expectedversus unexpected CS–US presentations. The results showed thatCg, PL, OFC, and RAIC all expressed significantly greaterc-Fos-IR in Group Unexpected than Group Expected. The samedifference between groups in c-Fos-IR was detected in amygdala,notably lateral amygdala, a structure critical for fear learning, aswell as in lateral hypothalamus. Midline thalamic nuclei alsoshowed this pattern of group differences in c-Fos-IR, and impor-tantly, these differences were preserved in midline thalamic neu-rons projecting directly to dmPFC. Finally, we confirmed a causalrole for dmPFC, but not IL, in predictive fear learning using awithin-subjects blocking design. Prior fear conditioning of a CSblocked subsequent fear learning to a target CS when the twostimuli were fear conditioned in compound. Reversible inactiva-tion of dmPFC, but not IL, with muscimol and baclofen preventedthis blocking to enable normal fear learning about the target CS.These infusions had no effect on fear learning to a control CS. Thatis, reversible inactivation of dmPFC acted selectively to preventthe associative blocking of fear learning. Taken together, theseresults provide complementary lines of evidence that rodent PFC isimportant for predictive fear learning.
Fear learning occurs preferentially to surprising aversive out-comes. Unexpected aversive USs initiate fear learning whereasexpected aversive USs do not. We suggest that c-Fos expression inmidline thalamus and dmPFC reflects surprisingness of the US:unexpected, but not expected, CS–US presentations increasedc-Fos-IR in dmPFC and its thalamic afferents. Midline and in-tralaminar thalamus project extensively to dmPFC, RAIC, andOFC, and these regions, in turn, project to LA (e.g., Berendse &Groenewegen, 1991; Groenewegen & Berendse, 1994; Jasmin,Burkey, Granato, & Ohara, 2004). All of these regions showeddifferences between groups in c-Fos-IR in the present experimentso that c-Fos-IR was greater in Group Unexpected than GroupExpected. Moreover, we confirmed that these group differenceswere present in thalamic neurons projecting to dmPFC. There wasgreater activation, as indexed by c-Fos-IR, in a thalamocorticalpathway in Group Unexpected compared to Group Expected.
The midline thalamus 3 PFC pathway is part of the “medialpain pathway” conveying information about the aversive affective/motivational quality of noxious stimuli to the forebrain includingthe amygdala (e.g., Price, 2000). The critical neural pathwayswhich convey information about the aversive US to amygdaladuring fear conditioning remain unclear. Initially it was suggestedthat both posterior intralaminar thalamus and parietal insular cor-tex may be important (Shi & Davis, 1999), whereas others reportedconflicting effects of lesions of these structures on the acquisitionof fear learning (e.g., Brunzell & Kim, 2001; Lanuza, Nader, &LeDoux, 2004). Our data suggest that a thalamic and PFC networkmay be important for fear learning because it signals whetheraversive events are expected (low activity in midline thalamus 3PFC pathway) or unexpected (greater activity in a midline thala-mus3 PFC pathway) and so whether they are to be learned about.In the absence of this signal, fear learning occurs but this learningis independent of prediction error.
583PREFRONTAL CORTEX AND PREDICTING DANGER

In human aversive learning, expectation attenuates the US-elicited fMRI bold signal in midline thalamus, cingulate, anddorsolateral PFC (Dunsmoor et al., 2008). This finding is remark-ably similar to that reported here, and is consistent with oursuggestion that a thalamic 3 PFC pathway determines variationsin the effectiveness of the US to support fear learning. Reductionsof activity in a thalamic–dmPFC pathway may be related toreductions in the effectiveness of the shock US as an aversivereinforcer. One candidate structure for causing reductions in thethalamic–dmPFC pathway is the vlPAG. vlPAG contributes topredictive fear learning (e.g., Cole & McNally, 2009; McNally &Cole, 2006). Furthermore, vlPAG receives projections from centralamygdala which are critical for fear CR expression, and vlPAGprojects extensively to the midline and intralaminar thalamus(Krout & Loewy, 2000). We suggest that a vlPAG 3 thalamicpathway imposes negative feedback control over the thalamic 3PFC pathway. In this way the output of the fear system, triggeredby presentation of a feared CS and expressed through vlPAG, mayreduce effectiveness of the aversive US as a teaching signal(Fanselow, 1998; McLaren, 1989; McNally, 2009) by reducingUS-related activity in the thalamic–PFC pathway (Figure 5). Theabsence of differences between Groups Expected and Unexpectedin vlPAG c-Fos-IR is not problematic for this possibility becausedifferences between these groups might only be observed in thesubpopulation of vlPAG neurons projecting to midline and in-tralaminar thalamus.
Current accounts of rodent PFC function during fear condition-ing emphasize a role in regulating expression of conditioned fear.dmPFC is important for sustaining fear responding; whereas ven-tromedial PFC is important for inhibiting fear responding. There isconsiderable evidence for these roles. For example, activity of ratPL neurons correlates with the expression of freezing (Burgos-Robles, Vidal-Gonzalez, & Quirk, 2009), reversible inactivation ofPL prevents the expression of freezing as a conditioned response,whereas microstimulation of this region augments expression ofthis responding (Laurent & Westbrook, 2009; Vidal-Gonzalez,Vidal-Gonzalez, Rauch, & Quirk, 2006). Likewise, fear extinctiontraining increases the excitability of IL neurons (Santini, Quirk, &Porter, 2008), and microstimulation of IL suppresses expression offear (Vidal-Gonzalez et al., 2006; Milad, Vidal-Gonzalez, &Quirk, 2004). These findings have supported a role for PFC inregulating the behavioral expression of already learned associa-tions during fear conditioning. Our results show that dmPFC isequally important for regulating the formation of associationsduring conditioning (see also Takahashi et al., 2009).
It is noteworthy that reversible inactivation of dmPFC using thesodium channel blocker tetrodotoxin does not affect the acquisi-tion of fear learning to a single auditory CS (Corcoran & Quirk,2007). We also found no effect of dmPFC inactivation on learningto the control CS. However, lesions of dmPFC, centered on Cg, doenhance the acquisition of conditioned fear to both auditory andcontextual stimuli (Morgan & LeDoux, 1995). A similar augmen-tation of fear learning was reported following lesions of lateralPFC which included RAIC (Lacroix, Spinelli, Heidbreder, & Fel-don, 2000). These conflicting results are not problematic becauseit is technically difficult to make inferences about predictive learn-ing from the single cue designs used in past research. For example,the augmentation of fear learning after dmPFC lesions is consistentwith the claim that dmPFC is important for predictive fear learn-ing. This follows because past fear learning about a CS limits howmuch is learned about that CS on later trials. Manipulations thatprevent the use of past learning to guide future learning canaugment the acquisition of fear to a single CS (McNally, Pigg, &Weidemann, 2004). These inconsistent results using single cuedesigns underscore the usefulness of blocking and related designsto the study of the brain mechanisms of fear learning.
It is worth commenting on alternate interpretations of these data.In Experiment 1, the groups differed not only in whether theCS–US presentations were consistent or inconsistent with pasttraining and so expected or unexpected, but, they also differed inthe novelty of the stimuli on test. Thus, differences between thesegroups could be linked to variations in effectiveness of the US asa teaching signal supporting fear learning, differences in the nov-elty of the US, or differences in both. Our results do not allowdetermination of whether the thalamic 3PFC pathway is influ-enced by the surprising US or the surprising CS–US pairings. Anunsignalled US (or US alone) condition on test would not havebeen helpful in this regard because US alone presentations are stillsignaled by contextual cues and thus associatively equivalent toour unexpected group (Rescorla & Wagner, 1972). The absence ofdifferences among animals receiving expected versus unexpectedCS alone presentations suggests that the surprisingness of theauditory CS itself was not a critical factor.
These are important issues worthy of future investigation. How-ever, our data clearly show that dmPFC contributions to predictive
Figure 5. Potential circuit level mechanisms for predictive fear learning.A v1PAG midline thalamus3 PFC3 LA pathway signals surprisingnessof the aversive shock US so that neurotransmission in this pathway cor-responds to the discrepancy between actual and expected outcomes of theconditioning trial. Adapted from “The Rat Brain in Stereotaxic Coordi-nates, 5th ed.,” by George Paxinos and Charles Watson. Copyright 2005 byPaxinos and Watson. Reprinted with permission of Elsevier.
584 FURLONG, COLE, HAMLIN, AND MCNALLY

fear learning are independent of any differences in novelty of thestimuli being learned about. The within subjects blocking designsused in Experiments 2a and 2b equate subjects on exposure to allstimuli used in the experiment. Under these conditions, reversibleinactivation of dmPFC, but not IL, prevents the associative block-ing of fear learning. This indicates a causal role for dmPFC inpredictive fear learning. Finally, the Fos protein can be expressedby both neurons and glia. In the absence of double staining forneuron specific (e.g., NeuN) or glia specific (GFAP) markers wecannot exclude the possibility that at least some of the c-Fos-IRdetected in Experiment 1 was expressed in glial cells. Nonetheless,our findings with CTb show that expectancy modulation of c-Fosinduction can be detected in anatomically verified thalamic pro-jection neurons.
In conclusion, dmPFC is a critical component of the neuralcircuit for predictive fear learning. c-Fos-IR in Cg, PL, OFC,RAIC, and LA, as well as a thalamic 3 dmPFC pathway, wasmodulated by past experience, so that only surprising CS–USpresentations increased c-Fos-IR in these regions. dmPFC, but notIL, was causally important for this predictive learning becausereversible inactivation prevented the associative blocking of fearlearning. This pattern of results is consistent with a role fordmPFC, and a thalamic3 dmPFC pathway, in signaling whetheror not aversive events are expected or unexpected and so whetherthey are to be learned about.
References
Berendse, H. W., & Groenewegen. H. J. (1991). Restricted cortical termi-nation fields of the midline and intralaminar thalamic nuclei in the rat.Neuroscience, 42, 73–102.
Brunzell, D. H., & Kim, J. J. (2001). Fear conditioning to tone, but notcontext, is attenuated by lesions of the insular cortex and posteriorextension of the intralaminar complex in rats. Behavioral Neuroscience,115, 365–375.
Burgos-Robles, A., Vidal-Gonzalez, I., & Quirk, G. J. (2009). Sustainedconditioned responses in prelimbic prefrontal neurons are correlatedwith fear expression and extinction failure. The Journal of Neuroscience,29, 8474–8482.
Carter, C. S., & van Veen, V. (2007). Anterior cingulate cortex and conflictdetection: An update of theory and data. Cognitive Affective and Behav-ioral Neuroscience, 7, 367–379.
Cole, S., & McNally, G. P. (2007). Opioid receptors mediate directpredictive fear learning: Evidence from one-trial blocking. Learning &Memory, 14, 229–235.
Cole, S., & McNally, G. P. (2009). Complementary roles for amygdala andperiaqueductal gray in temporal-difference fear learning. Learning &Memory, 16, 1–7.
Corcoran, K. A., & Quirk, G. J. (2007). Activity in prelimbic cortex isnecessary for the expression of learned, but not innate, fears. TheJournal of Neuroscience, 27, 840–844.
Dunsmoor, J. E., Bandettini, P. A., & Knight. D. C. (2008). Neuralcorrelates of unconditioned response diminution during Pavlovian con-ditioning. Neuroimage, 40, 811–817.
Fanselow, M. S. (1998). Pavlovian conditioning, negative feedback, andblocking: Mechanisms that regulate association formation. Neuron, 20,625–627.
Groenewegen, H. J., & Berendse, H. W. (1994). The specificity of the“nonspecific” midline and intralaminar thalamic nuclei. Trends in Neu-roscience, 17, 52–57.
Histed, M. H., Pasupathy, A., & Miller, E. K. (2009). Learning substrates
in the primate prefrontal cortex and striatum: Sustained activity relatedto successful actions. Neuron, 63, 244–253.
Holschneider, D. P., Yang, J., Sadler, T. R., Nguyen, P. T., Givrad, T. K.,& Maarek, J. M. (2006). Mapping cerebral blood flow changes duringauditory-cued conditioned fear in the nontethered, nonrestrained rat.Neuroimage, 29, 1344–1358.
Iordanova, M. D., McNally, G. P., & Westbrook, R. F. (2006). Opioidreceptors in the nucleus accumbens regulate attentional learning in theblocking paradigm. The Journal of Neuroscience, 26, 4036–4045.
Jasmin, L., Burkey, A. R., Granato, A., & Ohara, P. T. (2004). Rostralagranular insular cortex and pain areas of the central nervous system: Atract-tracing study in the rat. The Journal of Comparative Neurology,468, 425–440.
Johansen, J. P., & Fields, H. L. (2004). Glutamatergic activation of anteriorcingulate cortex produces an aversive teaching signal. Nature Neuro-science, 7, 398–403.
Kamin, L. J. (1968). “Attention-like” processes in classical conditioning. InM. R. Jones (Ed.), Miami symposium on the prediction of behavior:Aversive stimulation (pp. 9–33). University of Miami: Miami:
Kim, J. J., & Jung, M. W. (2006). Neural circuits and mechanisms involvedin Pavlovian fear conditioning: A critical review. Neuroscience andBiobehavioral Reviews, 30, 188–202.
Krout, K. E., & Loewy, A. D. (2000). Periaqueductal gray matter projec-tions to midline and intralaminar thalamic nuclei of the rat. The Journalof Comparative Neurology, 424, 111–141.
Lacroix, L., Spinelli, S., Heidbreder, C. A., & Feldon, J. (2000). Differ-ential role of the medial and lateral prefrontal cortices in fear andanxiety. Behavioral Neuroscience, 114, 1119–1130.
Lanuza, E., Nader, K., & LeDoux, J. E. (2004). Unconditioned stimuluspathways to the amygdala: Effects of posterior thalamic and corticallesions on fear conditioning. Neuroscience, 125, 305–315.
Laurent, V., & Westbrook, R. F. (2009). Inactivation of the infralimbic butnot the prelimbic cortex impairs consolidation and retrieval of fearextinction. Learning & Memory, 16, 520–529.
Maren, S., & Quirk, G. J. (2004). Neuronal signalling of fear memory.Nature Reviews Neuroscience, 5, 844–852.
Matsumoto, M., Matsumoto, K., Abe, H., & Tanaka, K. (2007). Medialprefrontal cell activity signaling prediction errors of action values.Nature Neuroscience, 10, 647–656.
McLaren, I. P. L. (1989). The computational unit as an assembly ofneurones: An implementation of an error correcting learning algorithm.In R. Durbin, C. Miall, G. Mitchison (Eds.), The computing neuron (pp.160–179). Amsterdam: Addison Wesley.
McNally, G. P. (2009). The roles of endogenous opioids in fear learning.International Journal of Comparative Psychology, 22, 153–169.
McNally, G. P., & Cole, S. (2006). Opioid receptors in the midbrainperiaqueductal gray regulate prediction errors during Pavlovian fearconditioning. Behavioral Neuroscience, 121, 313–323.
McNally, G. P., Pigg, M., & Weidemann, G. (2004). Blocking, unblockingand overexpectation of fear: Opioid receptors regulate Pavlovian asso-ciation formation. Behavioral Neuroscience, 118, 111–120.
McNally, G. P., & Westbrook, R. F. (2006). Predicting danger: The nature,consequences, and neural mechanisms of predictive fear learning.Learning & Memory, 13, 245–253.
Milad, M. R., Vidal-Gonzalez, I., & Quirk, G. J. (2004). Electrical stim-ulation of medial prefrontal cortex reduces conditioned fear in a tempo-rally specific manner. Behavioral Neuroscience, 118, 389–394.
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of prefrontalcortex function. Annual Review of Neuroscience, 24, 167–202.
Morgan, M. A., & LeDoux, J. E. (1995). Differential contribution of dorsaland ventral medial prefrontal cortex to the acquisition and extinction ofconditioned fear in rats. Behavioral Neuroscience, 109, 681–688.
Narayanan, N. S., & Laubach, M. (2008). Neuronal correlates of post-error
585PREFRONTAL CORTEX AND PREDICTING DANGER

slowing in the rat dorsomedial prefrontal cortex. Journal of Neurophys-iology, 100, 520–525.
O’Brien, R. G., & Kaiser, M. K. (1985). MANOVA method for analysingrepeated measures designs: An extensive primer. Psychological Bulletin,97, 316–333.
Paxinos, G., & Watson, C. (2005). The rat brain in stereotaxic coordinates(5th ed.), San Diego: Elsevier.
Paxinos, G., & Watson, C. (2007). The rat brain in stereotaxic coordinates(6th ed.), San Diego: Elsevier.
Price, D. D. (2000). Psychological and neural mechanisms of the affectivedimension of pain. Science, 288, 1769–1772.
Rescorla, R. A., & Wagner, A. R. (1972). A theory of Pavlovian condi-tioning: Variations in the effectiveness of reinforcement and nonrein-forcement. In A. H. Black & W. F. Prokasy (Eds.), Classical condition-ing II: Current research and theory (pp. 64–99). Appleton-Century-Crofts: New York.
Santini, E., Quirk, G. J., & Porter, J. T. (2008). Fear conditioning andextinction differentially modify the intrinsic excitability of infralimbicneurons. The Journal of Neuroscience, 28, 4028–4036.
Shi, C., & Davis, M. (1999). Pain pathways involved in fear conditioningmeasured with fear-potentiated startle: Lesion studies. The Journal ofNeuroscience, 19, 420–430.
Takahashi, Y. K., Roesch, M. R., Stalnaker, T. A., Haney, R. Z., Calu,D. J., Taylor, A. R., Burke, K. A., et al. (2009). The orbitofrontal cortexand ventral tegmental area are necessary for learning from unexpectedoutcomes. Neuron, 62, 269–280.
Taylor, S. F., Stern, E. R., & Gehring, W. J. (2007). Neural systems forerror monitoring: Recent findings and theoretical perspectives. Neuro-scientist, 13, 160–172.
Vidal-Gonzalez, I., Vidal-Gonzalez, B., Rauch, S. L., & Quirk, G. J.(2006). Microstimulation reveals opposing influences of prelimbic andinfralimbic cortex on the expression of conditioned fear. Learning &Memory, 13, 728–733.
Received March 24, 2010Revision received June 17, 2010
Accepted June 21, 2010
Call for Nominations
The Publications and Communications (P&C) Board of the American PsychologicalAssociation has opened nominations for the editorships of Journal of ExperimentalPsychology: Learning, Memory, and Cognition; Professional Psychology: Researchand Practice; Psychology and Aging; Psychology, Public Policy, and Law; and SchoolPsychology Quarterly for the years 2013–2018. Randi C. Martin, PhD, Michael C.Roberts, PhD, Paul Duberstein, PhD, Ronald Roesch, PhD, and Randy W. Kamphaus,PhD, respectively, are the incumbent editors.
Candidates should be members of APA and should be available to start receivingmanuscripts in early 2012 to prepare for issues published in 2013. Please note that theP&C Board encourages participation by members of underrepresented groups in thepublication process and would particularly welcome such nominees. Self-nominations arealso encouraged.
Search chairs have been appointed as follows:
● Journal of Experimental Psychology: Learning, Memory, and Cognition, LeahLight, PhD, and Valerie Reyna, PhD
● Professional Psychology: Research and Practice, Bob Frank, PhD, and LillianComas-Diaz, PhD
● Psychology and Aging, Leah Light, PhD● Psychology, Public Policy, and Law, Peter Ornstein, PhD, and Brad Hesse, PhD● School Psychology Quarterly, Neal Schmitt, PhD, and Jennifer Crocker, PhD
Candidates should be nominated by accessing APA’s EditorQuest site on the Web.Using your Web browser, go to http://editorquest.apa.org. On the Home menu on the left,find “Guests.” Next, click on the link “Submit a Nomination,” enter your nominee’sinformation, and click “Submit.”
Prepared statements of one page or less in support of a nominee can also be submittedby e-mail to Sarah Wiederkehr, P&C Board Search Liaison, at [email protected].
Deadline for accepting nominations is January 10, 2011, when reviews will begin.
586 FURLONG, COLE, HAMLIN, AND MCNALLY





























