Embed Size (px)
Citation preview

T(
MGa
b
c
a
ARR1A
KPCPCEL
1
asrtclla1piot
T
0d
Aquatic Toxicology 108 (2012) 42– 52
Contents lists available at SciVerse ScienceDirect
Aquatic Toxicology
j ourna l ho me pag e: www.elsev ier .com/ locate /aquatox
oxicity of crude oil and pyrene to the embryos of beach spawning capelinMallotus villosus)
arianne Frantzena,∗, Inger-Britt Falk-Petersenb, Jasmine Nahrganga,1, Timothy J. Smitha,ro H. Olsena, Thor Arne Hangstada, Lionel Camusa,c
Akvaplan-niva, Fram Centre, N-9296 Tromsø, NorwayFaculty of Bioscience, Fisheries and Economics, University of Tromsø, N-9037 Tromsø, NorwayThe University Centre in Svalbard N-9171 Longyearbyen, Norway
r t i c l e i n f o
rticle history:eceived 14 June 2011eceived in revised form6 September 2011ccepted 29 September 2011
eywords:olycyclic aromatic hydrocarbonrude oilyreneapelin
a b s t r a c t
Due to a northward shift in oil and gas activities, there is an increasing need to understand the potentialanthropogenic impacts of oil-related compounds on sub-Arctic and Arctic organisms, particularly thosein coastal habitats. Capelin (Mallotus villosus), a key fish species in the Barents Sea ecosystem, undertakesaggregated spawning at both intertidal and subtidal coastal localities in northern Norway. To investigatethe sensitivity of capelin embryos to oil compounds, newly fertilized capelin eggs were collected from aspawning beach and exposed until hatch (32 days) to either the water soluble fraction of crude oil or thesingle PAH compound, pyrene. Threshold levels for egg mortality, development and hatching success weredetermined. Concentrations of 40 �g/L crude oil (
∑26 PAHs) and 55 �g/L pyrene significantly increased
embryonic mortality rates and decreased hatching success, compared with controls, indicating that apotential oil spill in the vicinity of capelin spawning grounds may cause significant impacts. No significant
mbryosC50
incidence of adverse effects such as yolk sac oedema, pericardia oedema, haemorrhages, craniofacialabnormalities, premature hatch or inhibited growth was observed. Histological studies of hatched larvaedid not reveal specific sublethal effects in tissues and organs. Developmental delays and subsequentembryo death were noticed at the period of eye pigmentation in affected groups. Early life-history stagesof capelin are sensitive indicators of PAH impacts, but the mechanisms responsible for the toxic effects
ion.
require further investigat. Introduction
Increasing oil and gas exploration, production, and transportctivities present a risk of accidental oil spills with potential con-equences for marine ecosystems. This environmental threat hasarely been studied in Arctic areas, and it is important to establisholerance limits and analyze possible harmful effects of hydro-arbons on key marine organisms in these waters. Fish eggs andarvae are of particular concern as they are regarded as sensitiveife stages, in part because they are not able to avoid pollutedreas (Rice, 1985). Investigations carried out in Alaska after the989 Exxon Valdez oil spill revealed that Pacific herring (Clupeaallasi) larvae hatching from adhesive demersal eggs at spawn-
ng sites affected by the spill had significantly higher frequenciesf morphological deformities and cytogenetic abnormalities thanhose originating from unaffected sites (Hose et al., 1996). Elevated∗ Corresponding author. Tel.: +47 777 50366; fax: +47 777 50301.E-mail address: [email protected] (M. Frantzen).
1 Pressent address: Faculty of Bioscience, Fisheries and Economics, University ofromsø, N-9037 Tromsø, Norway.
166-445X/$ – see front matter © 2011 Elsevier B.V. All rights reserved.oi:10.1016/j.aquatox.2011.09.022
© 2011 Elsevier B.V. All rights reserved.
mortality was also observed in pink salmon (Oncorhynchus gor-buscha) embryos spawned in intertidal stretches of stream deltasseveral months after the spill (Bue et al., 1996). In addition, exper-imental studies on early life stages in a variety of other fish speciesdemonstrated effects including pericardial and yolk sac oedema,small jaws, body axes defects and inhibited growth in response toexposure to petroleum products (i.e. Carls et al., 1999; Colavecchiaet al., 2004; Couillard, 2002; Heintz et al., 1999; Hendon et al., 2008;Incardona et al., 2004; Marty et al., 1997; Pollino and Holdway,2002). Several of these papers also showed that exposure to dis-solved polycyclic aromatic hydrocarbons (PAHs) are directly toxicto fish embryos at low �g/L aqueous concentrations.
The capelin (Mallotus villosus) stock in the Barents Sea isan important commercial resource, in good years reaching6–8 million tons with annual fishing harvest up to 3 million tons(Gjøsæter, 1998). Capelin represents a major food source for severalpredators in the ecosystem, like cod (Gadus morhua), marine mam-mals and sea birds. Capelin spawns 1 mm demersal eggs during
March and April in subtidal waters outside the north Norway andnorthwestern Russian (Murman) coasts (Gjøsæter, 1998) or inter-tidally (local fjord populations) (Nyholmen and Hopkins, 1988).The eggs are attached to coarse sand or gravel. In some cases thick
ic Toxicology 108 (2012) 42– 52 43
lfc(
stLpsooL(dtadteta
tttsdtoairwdfp
2
2
so(tauoM
2
2
fcmaiawcr
9.0
7.5
8.0
8.5Crude oil experiment
Pyrene expe rimen t
6.0
6.5
7.0
4.5
5.0
5.5
Tem
pera
ture
(°C
)
4.0
Expo sure day1 3 5 7 9 11 13 15 17 25 232119 27 29 31
M. Frantzen et al. / Aquat
ayers of eggs may be spawned. Shore-spawned eggs are exposed toreezing temperatures, sun exposure and droughts during the tidalycles, but are quite resistant to various environmental fluctuationsDavenport, 1989; Davenport and Stene, 1986; Præbel et al., 2009).
Differences in species sensitivity to PAHs have been demon-trated, e.g. Stene and Lønning (1984) ranked fish eggs accordingo increasing sensitivity to 2-methylnaphthalene (1–10 mg/L) as:umpsucker (Cyclopterus lumpus) < capelin < plaice (Pleuronecteslatessa) < flounder (Platichthys flesus) < long rough dab (Hippoglos-oides platessoides) < cod. Effects of the water soluble fraction (WSF)f unweathered Hibernia crude oil on capelin eggs and larvaef the Newfoundland stock was studied by Paine et al. (1992).ethal effects were only noted for embryos at high concentrations2.7–5.4 mg/L; initial concentrations) over long exposures (11–20ays), while larvae were more sensitive, showing increased mor-ality at lower concentrations (0.5–2.7 mg/L; initial concentrations)nd exposures (6 days). Sublethal effects (growth, pigmentation,evelopmental rates, time to hatch) were registered at concen-rations between 10% and 50% of lethal concentrations. Capelinmbryos incubated in oiled sediment showed similar effects, evenhough the water-soluble hydrocarbons in this case were mainlyliphatic compounds (Paine et al., 1991).
Whereas early life-stages of a variety of fish species are knowno be sensitive to PAH compounds, there is a need to further explorehese effects under realistic exposure conditions and on stocks fromhe European Arctic. This paper presents results from experimentaltudies of naturally spawned Barents Sea capelin eggs exposed toifferent concentrations of the WSF of crude oil. The set-up mimicshe natural weathering process of oil in sediment using Kobbe crudeil collected from an exploration well drilled at the Goliat field. Inddition, a model compound, the four-ring PAH pyrene, was testedn order to produce toxicity data for risk assessment models. Vitalates (mortality-, and hatching) and developmental deformationsere monitored throughout the incubation period and at hatch. Aetailed histomorphological study of newly hatched capelin larvaerom control and exposed groups were carried out in order to revealossible sublethal effects in exposed individuals.
. Materials and methods
.1. Egg collection
Fertilized capelin eggs were collected during low tide at thepawning site (69◦ 13′ N, 19◦ 19′ E) in Balsfjord, northern Norwayn the 16 April 2010. Eggs attached to gravel and macro algaeFucus sp.) were transported on ice to the Barents Ecotox Labora-ory, Tromsø where they were placed in particle-filtered (60 �m)nd UV treated running seawater of ambient temperature (4.5 ◦C)ntil the start of the experiments. One crude oil experiment andne pyrene experiment were performed in the period 20 April–21ay 2010.
.2. Experimental design
.2.1. Crude oil experimentCapelin embryos were exposed to the WSF of the Kobbe crude oil
rom the Goliat field in the southwest Barents Sea using oiled rockolumns (Carls et al., 1999) or clean seawater (control). This experi-ental set-up mimicked the release of oil from sediment following
spill, with a decrease in aqueous PAH concentrations and a changen the relative abundance of PAH compounds over time. Gravel of
pproximately 1–8 mm in diameter were washed, dried and mixedith crude oil at concentrations of 3, 6 and 12 g crude oil/kg gravelorresponding to low (L), medium (M) and high (H) oil treatments,espectively. Oiled gravels were dried at room temperature for 24 h
Fig. 1. Measured water temperatures in treatment waters of the crude oil exper-iment (solid line) and the pyrene experiment (dashed line). Water temperaturesfluctuate according to ambient sea water temperatures.
and frozen pending column preparation. The oiled gravels wereinserted (12 kg per column) into 1 m high polypropylene columns(110 mm in diameter), that is one column per treatment. Each oilconcentration and control treatment consisted of 5 replicate glasspetri dishes (diameter 50 mm) containing 20 eggs. Particle-filtered(60 �m) and UV treated seawater (960 ml/min) at ambient seawater temperature (Fig. 1) were pumped upwards through thecolumns, extracting 100 (±10) ml/min of the WSF into each ofthe experimental dishes (pseudoreplicates). Each dish was placedinto a mesh (500 �m) cylinder to avoid any loss of eggs in theflow-through system. The control dishes received water perco-lated through a column containing clean gravel. To ensure thevolatilization of BTEX compounds (benzene, toluene, ethylbenzene,and xylene) and an absence of particulate oil, water was pumpedthrough the columns for 48 h before the exposure of the eggsstarted. In the period 15th of May–21th of May (day 26–32) theset-up was changed from a flow through system to a semi staticsystem to avoid loss of hatched larvae. Exposure waters were thencollected from each column, and 10 ml was added to each dish andchanged every 24-h. Water samples for PAH quantification weretaken from the outlet of each column once per week and frozen at−20 ◦C until analysis.
2.2.2. Pyrene experimentCapelin eggs (three replicates per treatment, 20 eggs per repli-
cate) were exposed to five nominal concentrations of pyrene, acomponent that is known to accumulate for long term in the sedi-ment following an oil spill: 17 (P1), 50 (P2), 150 (P3), 450 (P4) and1350 (P5) �g/L. Solvent control (SC) was prepared by adding thesame amount (0.009% of total volume of sea water) of acetone as atthe highest pyrene concentration. Control (C) embryos were incu-bated in sea water only. The experiment was performed in smallglass petri dishes (diameter 50 mm) into which 10 ml of exposuremedia were added. The experiment was semi-static with treatmentwater changed every 24 h. Prior to water changes, treatment waters
were freshly made by serial dilutions from a pyrene stock solution(50 mg pyrene/ml acetone). Each dilution was stirred for minimum30 min prior to further dilutions, exposure to embryos or collec-tion of water samples for pyrene analysis (collected once a week).
4 c Toxicology 108 (2012) 42– 52
Tcs
2
a(woosswfiuyJ4s
2
(eAaeswmTrsu
2
hebtgSatpC
2
ThwvAmmHpff
80
90
C
50
60
70C
L
M
H
/L)
30
40
50 H
ΣΣ26
PA
H (μ
g/
0
10
20
1 3 7 9 11 13 15 17 19 21 23 25 27 29 315
Expo sure day∑
4 M. Frantzen et al. / Aquati
he experiment was conducted in a small isolated room that wasooled by running sea water, allowing an incubation temperatureimilar to the ambient sea water temperature (Fig. 1).
.2.3. End pointsIn both experiments, developmental abnormalities, hatching
nd mortality were monitored every 2–3 days by stereomicroscopyLeica MZ6 with integrated DFC camera and application soft-are 2.8.1, Leica Microsystems (Switzerland) Ltd.). Development
f eye pigmentation was recorded to quantify possible effectsn the developmental rate of the embryos. At the day of hatch,ub-samples of hatched larvae were photographed (Leica M205Ctereomicroscope with integrated DFC camera and application soft-are 3.6.0, Leica Microsystems (Switzerland) Ltd.) and thereafterxed for later histological studies. The photos of the larvae weresed for morphological analysis (measurement of total length,olk sac length and width) by ImageJ; Processing and Analysis inava. The yolk sac volume was calculated according to the formula/3[(L/2)2 × (H/2)], where L and H are maximum and minimum yolkac diameter, respectively (Hourston et al., 1984).
.3. Chemical analysis
All water analyses were performed by Unilab Analyse ASTromsø, Norway). The water samples from the crude oilxperiment were analyzed for the 16 Environmental Protectiongency (EPA) priority PAHs plus naphthalenes, phenanthrenesnd dibenzothiophenes (NPD). Water samples from the pyrenexperiment were analyzed for pyrene only. Briefly, an internaltandard was added to the water samples that were extractedith dichloromethane. Extracts were analyzed by Gas Chro-atograph/Mass Spectrometry (GC/MS) (Hewlett-Packard/Agilent
echnologies). The quantification of individual components waselated to the internal standard. When concentrations were mea-ured to be below the level of quantification (LOQ), 1/2 LOQ wassed as concentration value.
.4. Histology
Hatched larvae fixed in 2% glutaraldehyde and 2% paraformalde-yde in cacodylate buffer (modified Karnowsky fixation) werembedded in Technovit after general dehydration proceduresefore being sectioned both longitudinally (for overview) andransversely (selected areas: eye/brain, buccal/gill area, oesopha-us area, yolk sac area, liver/pancreas area, urinary bladder area).ections (2–3 �m) from at least 5 larvae from each exposure levelnd the control were cut on a rotation microtome, stained witholuidine blue and mounted on glass slides before being studied andhotographed in the microscope (Leitz Aristoplan with Olympusamedia digital camera).
.5. Statistical analysis and data treatment
All statistical analyses were performed with Statistica 9.1.he data were arcsine square root-transformed (proportionatch/dead) or log-transformed (days to hatch/death), wheneveras necessary for homogenizing variance, but non-transformed
alues are shown in the figures. The tests used were one-wayNOVA (proportion of hatch/dead) and the generalized linearodel (GLM) nested ANOVA (day to hatch/death; nested in treat-ent). When a significant treatment effect was found, the Tukey’s
SD post hoc test (proportion of hatch/dead) or the unequal N HSDost hoc test (day to hatch/death) was applied to distinguish dif-erences. A probability level of p ≤ 0.05 was considered significantor all tests.Fig. 2. Total PAH concentration ( 26 PAH) in treatment waters of the crude oilexperiment at exposure day 1, 8, 15, 22 and 29. C: control, L: low oil treatment, M:medium oil treatment, H: high oil treatment.
3. Results
At the time of collection, eggs were at the 32 blas-tomeres/formation of morula stage and at the start of the exposure(day 1) the eggs were at the blastopore/end of gastrulation stage(Friðgeirsson, 1976).
3.1. Water PAH concentrations
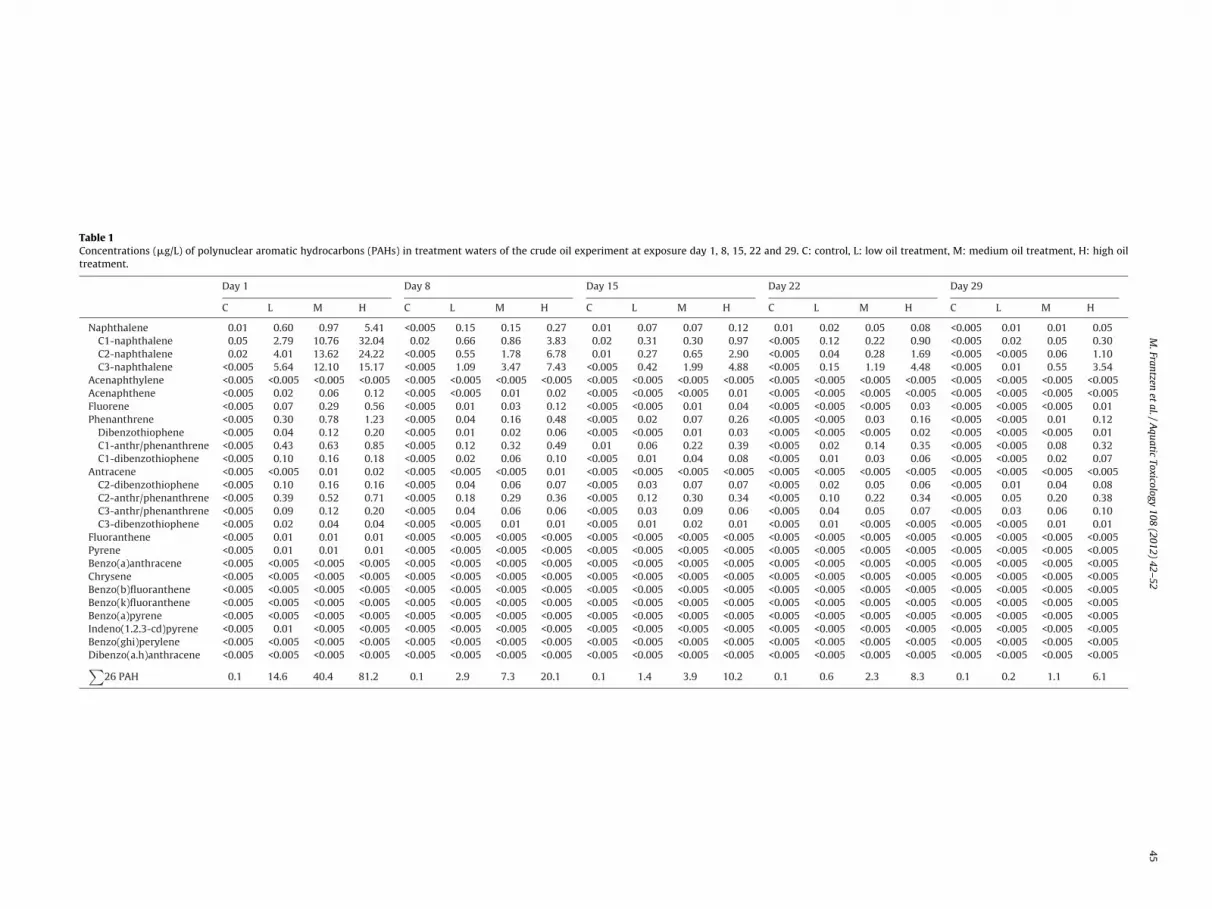
Water samples from day 1, 8, 15, 22 and 29 were analyzedfrom the crude oil experiment (Table 1; Fig. 2). Initial total (
∑26
PAH) concentrations in the water of H, M, L and C treatments were81.2, 40.4, 14.6 and 0.1 �g/L, respectively, and declined over theexperimental period. The predominant PAHs initially present werethe smaller and less substituted naphthalene homologues. RelativePAH abundance progressively shifted towards more substitutednaphthalene homologues and the larger and more substituteddibenzothiophenes and phenanthrenes homologues in the three oiltreatments. Smaller PAHs were exhausted from gravel more rapidlyin the lower oil treatments and, consequently, the proportion ofsmaller PAHs was greater in the higher oil treatments comparedto the low treatment. In all groups anthracene, fluoranthene andpyrene appeared at very low/below detection limit concentrationsthroughout the experiment.
Water samples from day 1, 8, 16 and 29 were analyzed from thepyrene experiment (Table 2). The results from day 1 were inconsis-tent. At day 8 the measured concentrations were 30–66% lower thannominal concentrations, followed by an increase in concentrationsbetween day 8 and 16 to 125–180% of the nominal concentrations.The concentrations remained stable throughout the last part of theexperiment (day 16–29). Average pyrene concentrations from day8, 16 and 29 were used for the calculation of the LC50 values (seeSection 3.4).
3.2. Survival
In the crude oil experiment, there was a significant increase
(n = 5, p < 0.001) in the proportion of dead embryos with increasingtotal PAH (∑26 PAH) concentration in the incubation water (Fig. 3).
The H concentration group was significantly different from the Mconcentration group, and both H and M groups were significantly
M.
Frantzen et
al. /
Aquatic
Toxicology 108 (2012) 42– 52
45
Table 1Concentrations (�g/L) of polynuclear aromatic hydrocarbons (PAHs) in treatment waters of the crude oil experiment at exposure day 1, 8, 15, 22 and 29. C: control, L: low oil treatment, M: medium oil treatment, H: high oiltreatment.
Day 1 Day 8 Day 15 Day 22 Day 29
C L M H C L M H C L M H C L M H C L M H
Naphthalene 0.01 0.60 0.97 5.41 <0.005 0.15 0.15 0.27 0.01 0.07 0.07 0.12 0.01 0.02 0.05 0.08 <0.005 0.01 0.01 0.05C1-naphthalene 0.05 2.79 10.76 32.04 0.02 0.66 0.86 3.83 0.02 0.31 0.30 0.97 <0.005 0.12 0.22 0.90 <0.005 0.02 0.05 0.30C2-naphthalene 0.02 4.01 13.62 24.22 <0.005 0.55 1.78 6.78 0.01 0.27 0.65 2.90 <0.005 0.04 0.28 1.69 <0.005 <0.005 0.06 1.10C3-naphthalene <0.005 5.64 12.10 15.17 <0.005 1.09 3.47 7.43 <0.005 0.42 1.99 4.88 <0.005 0.15 1.19 4.48 <0.005 0.01 0.55 3.54
Acenaphthylene <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005Acenaphthene <0.005 0.02 0.06 0.12 <0.005 <0.005 0.01 0.02 <0.005 <0.005 <0.005 0.01 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005Fluorene <0.005 0.07 0.29 0.56 <0.005 0.01 0.03 0.12 <0.005 <0.005 0.01 0.04 <0.005 <0.005 <0.005 0.03 <0.005 <0.005 <0.005 0.01Phenanthrene <0.005 0.30 0.78 1.23 <0.005 0.04 0.16 0.48 <0.005 0.02 0.07 0.26 <0.005 <0.005 0.03 0.16 <0.005 <0.005 0.01 0.12
Dibenzothiophene <0.005 0.04 0.12 0.20 <0.005 0.01 0.02 0.06 <0.005 <0.005 0.01 0.03 <0.005 <0.005 <0.005 0.02 <0.005 <0.005 <0.005 0.01C1-anthr/phenanthrene <0.005 0.43 0.63 0.85 <0.005 0.12 0.32 0.49 0.01 0.06 0.22 0.39 <0.005 0.02 0.14 0.35 <0.005 <0.005 0.08 0.32C1-dibenzothiophene <0.005 0.10 0.16 0.18 <0.005 0.02 0.06 0.10 <0.005 0.01 0.04 0.08 <0.005 0.01 0.03 0.06 <0.005 <0.005 0.02 0.07
Antracene <0.005 <0.005 0.01 0.02 <0.005 <0.005 <0.005 0.01 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005C2-dibenzothiophene <0.005 0.10 0.16 0.16 <0.005 0.04 0.06 0.07 <0.005 0.03 0.07 0.07 <0.005 0.02 0.05 0.06 <0.005 0.01 0.04 0.08C2-anthr/phenanthrene <0.005 0.39 0.52 0.71 <0.005 0.18 0.29 0.36 <0.005 0.12 0.30 0.34 <0.005 0.10 0.22 0.34 <0.005 0.05 0.20 0.38C3-anthr/phenanthrene <0.005 0.09 0.12 0.20 <0.005 0.04 0.06 0.06 <0.005 0.03 0.09 0.06 <0.005 0.04 0.05 0.07 <0.005 0.03 0.06 0.10C3-dibenzothiophene <0.005 0.02 0.04 0.04 <0.005 <0.005 0.01 0.01 <0.005 0.01 0.02 0.01 <0.005 0.01 <0.005 <0.005 <0.005 <0.005 0.01 0.01
Fluoranthene <0.005 0.01 0.01 0.01 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005Pyrene <0.005 0.01 0.01 0.01 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005Benzo(a)anthracene <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005Chrysene <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005Benzo(b)fluoranthene <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005Benzo(k)fluoranthene <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005Benzo(a)pyrene <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005Indeno(1.2.3-cd)pyrene <0.005 0.01 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005Benzo(ghi)perylene <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005Dibenzo(a.h)anthracene <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005 <0.005∑
26 PAH 0.1 14.6 40.4 81.2 0.1 2.9 7.3 20.1 0.1 1.4 3.9 10.2 0.1 0.6 2.3 8.3 0.1 0.2 1.1 6.1

46 M. Frantzen et al. / Aquatic Toxicology 108 (2012) 42– 52
90
100 ccbc c
90
100 c
b
A B
60
70
80
aa
ab
60
70
80
a
20
30
40
50
20
30
40
50 a
0
10
P5P4P3P2P1SCC0
10
HMLC
Perc
enta
ge d
ead
embr
yos
Treatment
Fig. 3. Average percentage dead embryos in the treatment groups of the crude oil experiment (A; n = 5) and the pyrene experiment (B; n = 3), respectively. Different lowerc ch ext ium c
daintw
ptstd2tti
3
(iTMc
TNe
ase characters indicate treatment groups that are significantly different within eaext for nominal and measured concentrations), L: low crude oil treatment, M: med
ifferent from the L concentration group and the C group. Aver-ge day of death was 27.7 ± 4.8, 26.4 ± 4.4, 24.8 ± 5.9 and 25.2 ± 4.7n the C, L, M and H concentration group, respectively. The GLMested ANOVA indicated a significant difference (p = 0.01) betweenhe groups. The post hoc test however did not reveal which groupsere different from each other.
Also, there was a significant increase (n = 3, p < 0.001) in the pro-ortion of dead embryos with increasing pyrene concentration inhe incubation water (Fig. 3). The P2–P5 concentration groups wereignificantly different from the C and the SC, and the P3–P5 concen-ration groups were also significantly different from P1. Averageay of death was C: 25.8 ± 4.5, SC: 25.7 ± 5.2, P1: 24.6 ± 4.2, P2:2.7 ± 3.7, P3: 23.1 ± 4.8, P4: 20.7 ± 2.7 and P5: 21.0 ± 0.0, respec-ively. Death occurred significantly earlier in P4 and P5 comparedo C and SC, and also in P4 compared to P1. Other pairwise compar-sons were not significant.
.3. Hatch
In the crude oil experiment, there was a significant reductionn = 5, p < 0.001) in the proportion of hatched eggs with increas-∑
ng PAH ( 26 PAH) concentration in the incubation water (Fig. 4).he H concentration group was significantly different from theconcentration group, and both H and M groups were signifi-antly different from the L concentration group and the C group.
able 2ominal and measured concentrations of pyrene in treatment waters of the pyrenexperiment at day 1, 8, 16 and 29.
Treatment Nominal concentration(�g/L)
Measured concentration (�g/L)
Day 1 Day 8 Day 16 Day 29
C 0 <18.7 <12.1 <18.7 <18.7SC 0 <18.7 <12.1 <18.7 <18.7P1 17 <18.7 <12.1 29.8 25.2P2 50 <18.7 26.6 72.0 66.4P3 150 28.2 58.4 247.9 198.6P4 450 336.8 126.4 651.6 638.4P5 1350 367.4 893.2 1710.0 1704.2
periment (p < 0.05). C: control, SC: solvent control, P1–P5: pyrene treatments (seerude oil treatment, H: high crude oil treatment.
Average day of hatch (±1 SD) was 26.3 ± 1.9, 26.2 ± 1.7, 27.0 ± 1.8and 27.8 ± 2.0 in the C, L, M and H concentration group, respec-tively, with no significant difference (p = 0.25) between treatmentpairs.
In the pyrene experiment there was also a significant reduction(n = 3, p < 0.001) in the proportion of hatched eggs with increasingpyrene concentration in the incubation water, and no eggs hatchedin the three highest pyrene treatment groups (Fig. 4). The P2–P5concentration groups were significantly different from the C andthe SC, and the P3–P5 concentration groups were also significantlydifferent from P1. Average day of hatch was 27.0 ± 1.9, 26.8 ± 2.2,25.5 ± 1.8 and 26.1 ± 1.9 in the C, SC, P1 and P2 concentration group,respectively, with no significant difference (p = 0.10) between thegroups.
3.4. Pyrene LC50
Embryo mortality in the different pyrene treatments (averagepyrene concentrations measured on day 8, 16, and 29) was recordedat different time points (0, 48, 96, 168, 216, 360, 432, 504, 576,600, 624, 648, 672, 696, 720 and 768 h). From these observations,the concentration affecting 50% of the test individuals (LC50) atdifferent time points of exposure (time–concentration–responserelationships) was derived by using the Dynamic Energy Budgetmodel for toxic responses (DEBtox) (Kooijman and Bedaux, 1996).This model is based on the DEB theory (Kooijman, 2000) whichdescribes the energy pathways in organisms and is specificallydesigned to decipher effects of toxicants on the allocation and use ofenergy by an organism. The pyrene LC50 values for capelin embryosderived from the model are shown in Table 3.
3.5. Developmental deformities
No yolk sac oedema or other specific types of deformities (e.g.
pericardial oedema, haemorrhages, craniofacial abnormalities, andbody axis defects) were observed in the embryos or hatched lar-vae in any groups of any of the two experiments. The histologicalanalyses supported these findings (see Section 3.7).
M. Frantzen et al. / Aquatic Toxicology 108 (2012) 42– 52 47
70
aa
70 BA
40
50
60
40
50
60
a
a
20
30b
20
30ab
bc
0
10
HMLC
c
Perc
enta
ge h
atch
ed e
ggs
0
10
P5P4P3P2P1SCC
ccc
Treatment
F xperic ch ext ium c
3
mia1twwhfdetitAcwt
TTt
ig. 4. Average percentage hatched eggs in the treatment groups of the crude oil ease characters indicate treatment groups that are significantly different within eaext for nominal and measured concentrations), L: low crude oil treatment, M: med
.6. Developmental rate
No concentration-dependent delay or acceleration in eye pig-entation was observed in any of the two experiments, and the first
ncidence of eye pigmentation was registered on exposure day 9 inll groups of both experiments (Fig. 5). In the crude oil experiment,7–29% of the embryos had reached the pigmentation endpoint onhis day, whereas in the pyrene experiment, the highest incidenceas seen in P3 where 65% of the embryos had pigmented eyes. Thisas significantly higher than in P5 where only 25% of the embryosad reached this stage. No groups, however, were significantly dif-
erent from the controls (45% and 43% in C and SC, respectively). Atay 15 close to 100% of the embryos in all groups had pigmentedyes. However, this showed to be a critical development stage forhe capelin embryos; there was a clear concentration-dependentnhibition of any further development in both experiments, andhe embryos that ceased development died before reaching hatch.t day 21 97–100% of the embryos in the two highest pyrene
oncentrations were dead, and in the crude oil experiment thereas a significant (p = 0.001) concentration-dependent increase inhe proportion of underdeveloped embryos (Fig. 5). There was no
able 3ime dependency of pyrene induced mortality of capelin embryos. LC50 values showhe concentration that kills 50% of experimental animals at specific times.
Day Hour LC50 (�g/L)
2 48 101004 96 51607 168 30909 216 2490
15 360 166018 432 146021 504 132024 576 121025 600 118026 624 116027 648 113028 672 111029 696 109030 720 107031 744 105032 768 1040
ment (A; n = 5) and the pyrene experiment (B; n = 3), respectively. Different lowerperiment (p < 0.05). C: control, SC: solvent control, P1–P5: pyrene treatments (seerude oil treatment, H: high crude oil treatment.
significant difference in larval length or yolk sac volume in hatchedlarvae among treatments in any of the two experiments (data notshown).
3.7. Histology and histopathology of newly hatched larvae
Longitudinal and transverse sections of newly hatched controlgroup capelin larva are shown in Fig. 6a–p. Capelin larvae are welldeveloped at hatch (Friðgeirsson, 1976). They have a differentiatedbuccal apparatus, strongly pigmented eyes, a differentiated gut, asmall but significant yolk sac and large pectoral fins. The longitu-dinal sections of the head in Fig. 6a and b illustrate the presence ofcartilaginous skeletal elements lining the jaws and buccal region.Cartilaginous gill supports were also noted (Fig. 6g). Numerouschloride cells were present in various regions of the body, includingthe gill region. Mucous cells lined the outer surface of the capelinlarvae (Fig. 6f and j). A relatively large olfactory organ was situatedin front of the eyes (Fig. 6a and b); the ciliated sensory cells of theolfactory bulbs were opening towards the exterior. The large eyeswere strongly pigmented and appeared fully functional at hatch(Fig. 6a–c and e), with well-developed retinal layers, eye musclesand nervous connections as noted in the histological sections. Theauditory organ (Fig. 6a, b, e, and f) was composed of large otic cham-bers and sensory maculae, and otoliths were illustrated in severalsections.
The long, straight digestive channel had numerous character-istic mucous cells in the oesophagus area (Fig. 6a and g), theintestine had a lumen (Fig. 6d, h, i, and j) and was lined withmicrovilli. The yolk sac showed a characteristic morphology, ahomogenous/yolk-granule-like area peripherally with numerouslarge, dark-blue staining cellular components (Fig. 6d and i). In theinner part, large yolk-platelets were present (Fig. 6d and i), whilelarge or smaller lipid vacuoles were most often washed out duringthe preparation process leaving empty spaces in the yolk sac.
Pancreatic tissue (with zymogen granules) and liver (with vac-
uolated hepatocytes) were elongated structures mainly locatedbehind the yolk region (Fig. 6i, j, and p). The gall bladder was alsopresent in association with the pancreatic tissue. A small urinarybladder appeared in the hind-gut sections.
48 M. Frantzen et al. / Aquatic Toxicology 108 (2012) 42– 52
Fig. 5. Average percentage of embryos with pigmented eyes (blue bars) and average percentage underdeveloped embryos (red lined bars) in each treatment group at differentdays of exposure. (A) Crude oil experiment (n = 5 dishes per group, n = 17–20 eggs per dish). (B) Pyrene experiment (n = 3 dishes per group, n = 19–21 eggs per dish). Differentl ts witt atmeno rsion
a
(ltaatwta
4
eiie
ower case characters indicate significant differences (p < 0.05) between treatmenreatments (see text for nominal and measured concentrations), L: low crude oil tref the references to colour in this figure legend, the reader is referred to the web ve
Large neuromast cells appeared in some cross-sections laterallylong the body axis.
Larvae that hatched in the hydrocarbon exposed replicatesapparently normal-looking) did not show any obvious histopatho-ogical defects (Fig. 6k–p). The pigmentation and differentiation ofhe eyes appeared similar in control and exposed larvae (Fig. 6knd l). Other sensory organs, like the auditory (Fig. 6m), alsoppeared similar to that of the control larvae. Yolk sac struc-ure was similar to that of controls (Fig. 6o). Skeletal supportsere well developed (Fig. 6l and m), muscle tissue and the diges-
ive channel with associated organs appeared normal (Fig. 6n, o,nd p).
. Discussion
The exposure studies in this investigation showed that chronic
xposures to dissolved PAHs at �g/L concentrations induce mortal-ty in capelin embryos, indicating that capelin is sensitive to PAHsn the same low ppb range as shown for other fish species (e.g. Carlst al., 1999; Heintz et al., 1999).hin each day of each experiment. C: control, SC: solvent control, P1–P5: pyrenet, M: medium crude oil treatment, H: high crude oil treatment. (For interpretation
of the article.)
4.1. PAH toxicity
The composition of PAH mixtures can vary considerably. In gen-eral, petroleum products are enriched with low molecular weightPAHs containing two or three rings, while fuel combustion prod-ucts contain a higher percentage and variety of high molecularweight compounds with four rings and greater. Numerous stud-ies demonstrate that toxicity increases with hydrophobicity, alkylsubstitution, and specific mechanisms of action (i.e. Neff, 1985;Rice et al., 1977). Increased weathering of crude oil enriches thefraction of tricyclic PAHs and their alkylated homologues, mak-ing more weathered crude oil toxic to fish embryos at lowerconcentrations than less weathered crude oil (Carls et al., 1999;Heintz et al., 1999). Paine et al. (1992) exposed capelin embryosto the WSF of unweathered Hibernia crude oil, demonstratinglethal embryo effects only at high exposure concentration of2.7–5.6 mg/L initial total hydrocarbon concentration (THC; mea-
sured by UV spectrometry), and the proportion of hatched larvaewere affected at concentrations 25–50% of the highest exposureconcentration. Aromatics accounted for 96% of the compoundsidentified in the WSF by gas chromatography (∑16 PAH: 860 �g/L;

M. Frantzen et al. / Aquatic Toxicology 108 (2012) 42– 52 49
Fig. 6. (a–l) Histological sections of hatched capelin larvae (a–j: control larvae; k–p: exposed larvae). l.s. = longitudinal section, t.s. = transverse section. (a) Control: Head,buccal and oesophagus region (l.s.) (au = auditory organ, h = heart, oe = oesophagus, ol = olfactory organ). (b) Control: Olfactory organ, eye, auditory organ, mouth, heart (l.s.)(au = auditory organ, h = heart, ol = olfactory organ). (c) Control: Eye with well-developed retinal layers and lens (l.s.). (d) Control: Yolk sac area (l.s.) (i = intestine, ys = yolksac). (e) Control: Head region with brain, eyes, buccal cavity (t.s.). (f) Control: Pharynx region with auditory organs, brain, gills (t.s.) (au = auditory organ, g = gill). (g) Control:Oesophagus area (t.s.) (mc = mucous cells, n = notochord, oe = oesophagus, pec = pectoral fin). (h) Control: Intestinal area between oesophagus and yolk sac (t.s.) (i = intestine,n = notochord). (i) Control: Yolk sac area (t.s.) (i = intestine, k = kidney, p = pancreas, ys = yolk sac). (j) Control: Behind yolk sac (t.s.) (i = intestine, l = liver, mc = mucous cells,p = pancreas). (k) P2: Head region with brain, eyes, buccal cavity (t.s.). (l) H: Head region with brain, eyes, buccal cavity (t.s.). (m) H: Pharynx region with auditory organs, brain,gills (t.s.) (au = auditory organ). (n) H: Oesophagus area (t.s.) (n = notochord, oe = oesophagus, pec = pectoral fin). (o) H: Yolk sac area (t.s.) (i = intestine, l = liver, m = muscletissue; p = pancreas). (p) P2: Hind body (l.s.) (i = intestine, l = liver, m = muscle tissue, p = pancreas) P2: pyrene treatment (50�g/L; nominal concentration), H: high oil treatment.

50 M. Frantzen et al. / Aquatic Toxicology 108 (2012) 42– 52
–l) (Co
cp2ttv4podPaeede
lprisiiaihfscatMbeWt
Fig. 6. (a
orresponding to 5.6 mg/L THC). The dominant identified com-ounds in the WSF were monoaromatics, and naphthalene, 1- and-metyl naphthalene were the only polynuclear PAHs present inhe WSF at concentrations >10 �g/L. In the crude oil experiment ofhe present study, significant effects on hatching success and sur-ival were induced at much lower initial concentration (
∑26 PAH:
0 �g/L; THC not measured). In contrast to Paine et al. (1992) theresent study did allow monoaromatics to volatilize before the startf the exposure and the PAHs identified in the water samples werei- and tricyclic parent- and alkylated compounds, with relativeAH abundance shifting progressively towards more substitutednd bigger PAHs throughout the experiment (Table 1). The differ-nce in PAH composition between these two studies most likelyxplains the difference in effect concentrations identified althoughirect comparison of concentrations are difficult due to differentxperimental set-ups and analytical methods.
In contrast to the present study, Paine et al. (1992) revealed sub-ethal effects such as reduced developmental rate (in terms of eyeigmentation), and reduced size of hatched larvae. Hydrocarbon-elated histopathology in hatched capelin larvae was not detectedn our investigation, which is in agreement with results from atudy on exposed herring larvae (Hose et al., 1996). Histopatholog-cal effects were not detected in herring larvae hatched from eggsn oil-polluted spawning habitats in Alaska after the Exxon Valdezccident, although morphological and genetic damages were reg-stered. In our investigation we investigated possible sublethalistological effects in apparently normal-looking larvae hatching
rom exposed replicates. The fact that organs and tissues appearedimilar to control larvae may indicate that these larvae have a fairhance to survive, but it cannot be excluded that exposure effectsppear at later stages of development. Paine et al. (1992) showedhat embryonic exposure can influence subsequent larval survival.
ortality over 6 days in <50% WSF was greater when exposure
egan before hatch than when exposure began after hatch. Heintzt al. (2000) exposed pink salmon (O. gorbuscha) embryos to theSF of Alaska North Slope (ANS) crude oil through an experimen-al set-up equal to the crude oil experiment in the present study.
ntinued)
The embryos that survived the treatment appeared healthy at hatchbut showed delayed effects on growth and marine survival.
The lack of observed sublethal effects in the present study mayhave been caused by the high mortality in the treatment groupsleaving a too low number of hatched larvae to reveal any significantsublethal effects. Furthermore, some sublethal effects like pericar-dial oedema, yolk sac oedema and skeletal deformities may be lesspronounced in capelin embryos exposed to petrogenic PAH com-pounds compared to other fish species like i.e. herring embryos(Carls et al., 1999; Incardona et al., 2009), as Paine et al. (1992) alsodid not find these effects. In our study no developing embryos weredechorionated for morphological or histological examination priorto hatch. Future studies should include histopathology of embryosduring the period when exposed individuals started to show abnor-mal development compared to controls.
PAHs have generally been thought to act as nonspecific or “base-line” toxicants through the nonpolar narcosis mode of action, orthrough activation of the aryl hydrocarbon receptor (AhR) path-way leading to cytochrome P450 1A (CYP1A) induction (Hintonet al., 2008). However, systematic studies on zebrafish (Daniorerio) embryos exposed to single PAH compounds in combinationwith morpholino gene knockdowns have revealed that differentPAH classes act via distinct toxic mechanisms during fish devel-opment (Incardona et al., 2004, 2005, 2006; Scott et al., 2011).Non-alkylated three-ring PAHs (in particular dibenzothiopheneand phenanthrene) are shown to induce morphological defectsthrough AhR independent cardiac dysfunction, whereas the toxicityof the four-ring non-alkylated PAH pyrene and benz[a]anthraceneare differentially dependent on tissue-specific activation of AhRisoforms or metabolism by CYP1A (Incardona et al., 2004, 2005,2006). In zebrafish embryos, cardiac dysfunction induced by non-alkylated three-ring PAHs was observed 36 h post fertilization(hpf), while the earliest effects of pyrene and benz[a]anthracene
were observed 96 and 48 hpf, respectively (Incardona et al.,2006). Experiments with both zebrafish and herring embryos indi-cate that the toxicity from weathered ANS crude oil are mainlymediated by tricyclic PAHs through AhR independent disruption
ic Toxi
opnedaTAeratozccpepai(hi
cfoiuAltfAthiteit
4s
tchw1tsrttPIlreheac
M. Frantzen et al. / Aquat
f cardiovascular function and morphogenesis, occurring at theharyngula stage (Incardona et al., 2004, 2009). Although no sig-ificant malformations were detected in the developing capelinmbryos in the present study, there was an apparent concentration-ependent cessation of development caused by both crude oilnd pyrene from day 15 (when all embryos had pigmented eyes).his was more pronounced in the pyrene experiment (Fig. 5).ccording to Friðgeirsson (1976), the first heartbeat in capelinmbryos appears at the 40+ somite stage, just prior to occur-ence of eye pigmentation. As such, the stoppage of developmentnd main mortality event observed in our experiments seemo appear at approximately the same stage as the occurrencef AhR independent cardiac disruption reported in herring andebrafish embryos (Incardona et al., 2004, 2009). This may indi-ate that AhR independent tricyclic PAH toxicity is induced inapelin comparable to i.e. herring and zebrafish. AhR dependentyrene toxicity appears to be induced or to be more severe at anarlier developmental stage in capelin than in other species: inyrene-exposed (1000 ppb) zebrafish embryos, mortality mainlyppeared after hatch (120 hpf) (Incardona et al., 2006), and alson sheepshead minnow (Cyprinodon variegatus) pyrene-induced20–150 ppb) mortality was mainly observed during early days postatch (Hendon et al., 2008). The mechanisms causing PAH toxicity
n capelin embryos need to be further investigated.Toxicity information is a key step in risk assessment and toxi-
ity data for local species is needed to determine threshold valuesor local ecological effects. Some information on effects on arcticrganisms exists, but the bulk of available data on arctic speciess not suitable for risk assessment nor do the data provide a goodnderstanding of short and long-term effects of contaminants inrctic ecosystems (Chapman and Riddle, 2005). There is a particu-
ar need for toxicity data on early life stages since they are knowno be more vulnerable than other life stages. The LC50 value (96 h)or pyrene in capelin embryos in this study was 5.16 mg/L (Table 3).ccording to the OECD guideline for the fish embryo toxicity (FET)
est, LC50 (96 h) values above 100 mg/L are considered non-toxic,ence pyrene is toxic to capelin embryos as clearly demonstrated
n this study. LC50 values were also calculated for 14 additionalime points. To our knowledge, no other LC50 values for capelinmbryos exposed to pyrene exist. The information from this studys therefore useful for future implementation in risk assessmentools.
.2. Ecological significance of an oil spill in the vicinity of capelinpawning sites
Total PAH concentrations in the crude oil experiment were inhe range of realistic concentrations of hydrocarbons in the waterolumn after an oil spill: total PAH concentrations of 6–1600 �g/Lave been reported in the water column during the first days oreeks after oil spills (Kingston, 1999; Law, 1978; Reddy and Quinn,
999; Short and Harris, 1996). The results from the present studyherefore indicate that a potential oil spill in the vicinity of capelinpawning sites may have effects on egg/larval survival and furtherecruitment of the exposed population if the oil manages to con-aminate the spawning substrate. The present study also showshat capelin embryo sensitivity (mortality and hatching success) toAHs is in the same range as herring sensitivity (Carls et al., 1999).n 1989, as a consequence of the Exxon Valdez oil spill, the herringarvae productivity in Prince William Sound was calculated to beeduced by 52% (Brown et al. in Carls et al., 1999). Further, sev-ral studies of the shorelines affected by the Exxon Valdez oil spill
ave shown that significant amounts of oil have persisted in thenvironment (especially in the mid-intertidal zone) for more thandecade in toxic forms, sufficiently bioavailable to induce biologi-al long-term impact at the population level (reviewed by Peterson
cology 108 (2012) 42– 52 51
et al., 2003). Animals that inhabit sediments receiving water thathas passed through subsurface oil deposits may be exposed tolow concentrations of PAHs. This process explained the elevatedmortality of incubating pink salmon eggs in oiled rearing streamsfor at least 4 years after the Exxon Valdez oil spill (Bue et al.,1998). Therefore, long-term effects on capelin populations from oilspills in the vicinity of capelin spawning sites may not be ruledout.
Acknowledgements
We thank Knut Forberg for invaluable guidance and partici-pation in collecting the capelin eggs. We also acknowledge FrideTonning, Fredrikke Johansen Musæus and Lies Vansteenbrugge forhistological preparations and morphological measurements, andPaul Renaud for valuable comments to the manuscript draft. Thisstudy was financed by Eni Norge and Eni E&P, through the Arc-tic Seas Biodiversity (ASBD) Project (www.arcticbiodiversity.com;WP3-1.6). Additional support was received from Akvaplan-niva.
References
Bue, B.G., Sharr, S., Moffitt, S.D., Craig, D., 1996. Effects of Exxon Valdez oil spill onpink salmon embryos and pre-emergent fry. Am. Fish. Soc. Symp. 18, 619–627.
Bue, B.G., Sharr, S., Seeb, J.E., 1998. Evidence of damage to pink salmon populationsinhabiting Prince William Sound Alaska, two generations after the Exxon Valdezoil spill. Trans. Am. Fish. Soc. 127, 35–43.
Carls, M.G., Rice, S.D., Hose, J.E., 1999. Sensitivity of fish embryos to weathered crudeoil. Part 1. Low-level exposure during incubation causes malformations, geneticdamage, and mortality in larval Pacific herring (Clupea pallasi). Environ. Toxicol.Chem. 18, 481–493.
Chapman, P.M., Riddle, M.J., 2005. Toxic effects of contaminants in polar marineenvironments. Environ. Sci. Technol. 39, 200A–207A.
Colavecchia, M.V., Backus, S.M., Hodson, P.V., Parrott, J.L., 2004. Toxicity of oil sandsto early life stages of fathead minnows (Pimephales promelas). Environ. Toxicol.Chem. 23, 1709–1718.
Couillard, C.M.A., 2002. Microscale test to measure petroleum oil toxicity to mum-michog embryos. Environ. Toxicol. 17, 195–202.
Davenport, J., 1989. The effects of salinity and low temperature on eggs of the Ice-landic capelin Mallotus villosus. J. Mar. Biol. Assoc. U.K. 69, 1–9.
Davenport, J., Stene, A., 1986. Freezing resistance, temperature and salinity tolerancein eggs, larvae and adults of capelin, Mallotus villosus, from Balsfjord. J. Mar. Biol.Assoc. U.K. 66, 145–157.
Friðgeirsson, E., 1976. Observations on spawning behavior and embryonic devel-opment of the Iselandic capelin, Rit Fiskideildar. J. Mar. Res. Inst. Reykjavik 5,1–35.
Gjøsæter, H., 1998. The population biology and exploitation of capelin (Mallotusvillosus) in the Barents Sea. Sarsia 83, 453–496.
Hendon, L.A., Carlson, E.A., Manning, S., Brouwer, M., 2008. Molecular and devel-opmental effects of exposure to pyrene in the early life-stages of Cyprinodonvariegatus. Comp. Biochem. Physiol. C 147, 205–215.
Heintz, R.A., Short, J.W., Rice, S.D., 1999. Sensitivity of fish embryos to weatheredcrude oil. Part 2. Environ. Toxicol. Chem. 18, 494–503.
Heintz, R.A., Rice, S.D., Wertheimer, A.C., Bradshaw, R.F., Thrower, F.P., Joyce, J.E.,Short, W., 2000. Delayed effects on growth and marine survival of pink salmonOncorhynchus gorbuscha after exposure to crude oil during embryonic develop-ment. Mar. Ecol. Prog. Ser. 208, 205–216.
Hinton, D.E., Segner, H., Au, D.W.T., Kullman, S.W., Hardman, R.C., 2008. Liver toxicity.In: Di Giulio, R.T., Hinton, D.E. (Eds.), The Toxicology of Fishes. CRC Press Taylor& Francis Group, pp. 327–400.
Hourston, A.S., Rosenthal, H., von Westernhagen, H., 1984. Viable hatch from eggsof Pacific herring (Clupea harengus pallasi) deposited at different intensities ona variety of substrates. Technical Report 1274. Department of Fisheries andOceans, Nanaimo, BC, Canada.
Hose, J.E., McGursk, M.D., Marty, G.D., Hinton, D.E., Brown, E.D., Baker, T.T., 1996.Sublethal effects of the Exxon Valdez oil spill on herring embryos and larvae:morphological, cytogenetic, and histopathological assessments, 1989–1991.Can. J. Fish. Aquat. Sci. 53, 2355–2365.
Incardona, J.P., Collier, T.K., Scholz, N.L., 2004. Defects in cardiac function precedemorphological abnormalities in fish embryos exposed to polycyclic aromatichydrocarbons. Toxicol. Appl. Pharmacol. 196, 191–205.
Incardona, J.P., Carls, M.G., Teraoka, H., Sloan, C.A., Collier, T.K., Scholz, N.L., 2005.Aryl hydrocarbon receptor-independent toxicity of weathered crude oil during
fish development. Environ. Health Perspect. 113, 1755–1762.Incardona, J.P., Day, H.L., Collier, T.K., Scholz, N.L., 2006. Developmental toxicity of4-ring polycyclic aromatic hydrocarbons in zebrafish is differentially dependenton AH receptor isoforms and hepatic cytochrome P450 1A metabolism. Toxicol.Appl. Pharmacol. 217, 308–321.

5 c Toxi
I
K
K
K
L
M
N
N
P
P
Short, J.W., Harris, P.M., 1996. Petroleum hydrocarbons in caged mussels deployed
2 M. Frantzen et al. / Aquati
ncardona, J.P., Carls, M.G., Day, H.L., Sloan, C.A., Bolton, J.L., Collier, T.K., Scholz, N.L.,2009. Cardiac arrhythmia is the primary response of embryonic Pacific herring(Clupea pallasi) exposed to crude oil during weathering. Environ. Sci. Technol.43, 201–207.
ingston, P., 1999. Recovery of the marine environment following the Braer Spill,Shetland. In: Proceedings of the 1999 Oil Spill Conference, Washington, DC.American Petroleum Institute, pp. 103–109.
ooijman, S.A.L.M., 2000. Dynamic Energy and Mass Budgets in Biological Systems.Cambridge University Press, Great Britain, ISBN 0-521-78608-8.
ooijman, S.A.L.M., Bedaux, J.J.M., 1996. The Analysis of Aquatic Toxicity Data. VUUniv. Press, Amsterdam, The Netherlands.
aw, R.J., 1978. Determination of petroleum hydrocarbons in water, fish and sedi-ments following the Ekofisk blow-out. Mar. Pollut. Bull. 9, 321–324.
arty, G.D., Hose, J.E., McGurk, M.D., Brown, E.D., Hinton, D.E., 1997. Histopathol-ogy and cytogenetic evaluation of Pacific herring larvae exposed to petroleumhydrocarbons in the laboratory or in Prince William Sound Alaska, after theExxon Valdez oil spill. Can. J. Fish. Aquat. Sci. 54, 1846–1857.
eff, J.M., 1985. Polycyclic aromatic hydrocarbons. In: Rand, G.M., Petrocelli, S.R.(Eds.), Fundamentals of Aquatic Toxicology. Hemisphere Publishing Corpora-tion, New York, pp. 416–454.
yholmen, O., Hopkins, C.C.E., 1988. Some observations on the population biology ofcapelin (Mallotus villosus) from Balsfjord, northern Norway. J. Cons. Int. Explor.Mer. 44, 264–276.
aine, M.D., Legget, W.C., McRuer, J.K., Frank, K.T., 1992. Effects of Hibernia crude
oil on capelin (Mallotus villosus) embryos and larvae. Mar. Environ. Res. 33,159–187.aine, M.D., Leggett, W.C., McRuer, J.K., Frank, K.T., 1991. Effects of incubation inoiled sediment on emergence of capelin (Mallotus villosus) larvae. Can. J. Fish.Aquat. Sci. 48, 2228–2239.
cology 108 (2012) 42– 52
Peterson, C.H., Rice, S.D., Short, J.W., Esler, D., Bodkin, J.L., Ballachey, B.L., Irons, D.B.,2003. Long-term ecosystem response to the Exxon Valdez oil spill. Science 302,2082–2086.
Pollino, C.A., Holdway, D.A., 2002. Toxicity testing of crude oil and related com-pounds using early life stages of the crimson-spotted rainbowfish (Melanotaeniafluviatilis). Ecotoxicol. Environ. Saf. 52, 180–189.
Præbel, K., Christiansen, J.S., Fevolden, S.E., 2009. Temperature and salinity condi-tions in a sub-Arctic intertidal spawning habitat for capelin. Mar. Biol. Res. 5,511–514.
Reddy, C.M., Quinn, J.G., 1999. GC–MS analysis of total petroleum hydrocarbons andpolycyclic aromatic hydrocarbons in seawater samples after the North Cape oilspill. Mar. Pollut. Bull. 38, 126–135.
Rice, S.D., Short, J.W., Karinen, J.F., 1977. Comparative oil toxicity and comparativeanimal sensitivity. In: Wolfe, D.A. (Ed.), Fate and Effects of Petroleum Hydro-carbons in Marine Organisms and Ecosystems. Pergamon Press, New York, pp.78–94.
Rice, S.D., 1985. Effects of oil on fish. In: Engelhardt, F.R. (Ed.), Petroleum Effects inthe Arctic Environment. Elsevier Applied Science Publishers, London, New York,pp. 157–182.
Scott, J.A., Incardona, J.P., Pelkki, K., Shepardson, S., Hodson, P.V., 2011.AhR2-mediated CYP1A-independent cardiovascular toxicity in zebrafish(Danio rerio) embryos exposed to retene. Aquat. Toxicol. 101,165–174.
in Prince William Sound after the Exxon Valdez oil spill. Am. Fish. Soc. Symp. 18,29–39.
Stene, A., Lønning, S., 1984. Effects of 2-methylnaphtalene on eggs and larvae of 6marine fish species. Sarsia 69, 199–203.
















