Embed Size (px)
Citation preview

Transcriptional Targeting to Anterior PituitaryLactotrophic Cells Using Recombinant AdenovirusVectors in Vitro and in Vivo in Normal and Estrogen/Sulpiride-Induced Hyperplasic Anterior Pituitaries*
T. D. SOUTHGATE†‡, S. WINDEATT†§, J. SMITH-ARICA, C. A. GERDES,M. J. PERONE, I. MORRIS, J. R. E. DAVIS, D. KLATZMANN, P. R. LOWENSTEIN¶,AND M. G. CASTRO
Molecular Medicine and Gene Therapy Unit (T.D.S., S.W., J.S.-A., C.A.G., M.J.P., P.R.L., M.G.C.),School of Medicine, Stopford Building; School of Biological Sciences (I.M.); and Endocrine SciencesResearch Group (J.R.E.D.), University of Manchester, Manchester M13 9PT, United Kingdom; andLaboratoire de Biologie et Therapeutique des Pathologies Immunitaires (D.K.), Universite Pierre andMarie Curie, CNRS, Hopital de la Pitie Salpetriere, 75651 Paris, Cedex 13, France
ABSTRACTThe use of pituitary cell type-specific promoters is a powerful mo-
lecular tool to achieve pituitary cell type-specific transcriptional tar-geting of transgenes encoded by viral vectors. It has recently beenproposed that transcriptional targeting of therapeutic genes could beharnessed as a gene therapy strategy for the treatment of pituitarydisease. We describe the successful use of the human PRL promoter(hPrl) encoded within recombinant adenovirus vectors to target trans-gene expression of Herpes Simplex Virus Type 1-Thymidine Kinase(HSV1-TK) or b-galactosidase to lactotrophic cells in vitro and in vivo.Functionally, the restriction of expression of HSV1-TK to lactotrophictumor cells, using the hPrl promoter, resulted in the cell type-specificinduction of apoptosis in the lactotrophic GH3 tumor cell line, in thepresence of ganciclovir (GCV). In the corticotrophic AtT20 cell line, wedetected neither HSV1-TK expression, nor apoptosis in the presenceof GCV. The hPrl promoter encoded within a recombinant adenoviralvector also restricted transgene expression to lactotrophic cells in
primary anterior pituitary (AP) cultures, and importantly, within theanterior pituitary gland in vivo. When the HSV1-TK driven by hPrlpromoter was used in an in vivo model of estrogen/sulpiride lactotrophinduced hyperplasia within the AP in situ, the treatment was noteffective in either reducing the weight of the gland, the number oflactotrophic cells within the transduced area in vivo, or the circulatingPRL levels. This is in contrast to the human cytomegalovirus pro-moter (hCMV) driving expression of HSV1-TK in the same experi-mental paradigm, which was effective in reducing pituitary weightand circulating PRL levels. Our results have important implicationsin the design of gene therapy strategies for pituitary tumors. Wedemonstrate that both the choice of the in vivo animal model, i.e.adenoma in the AP gland in situ, and the particular gene therapystrategy chosen, i.e. use of strong ubiquitous promoters vs. weaker butcell type-specific promoters, determine the experimental therapeuticoutcome. (Endocrinology 141: 3493–3505, 2000)
MANY DISEASES that affect pituitary function remainpoorly treated by currently available therapeutic
strategies. One reason for this is the multiplicity of transcrip-tional, posttranscriptional, hormonal, and neurotransmittercontrols that affect the function of individual pituitary celltypes. Drugs used to regulate anterior pituitary function lackthe capacity to discriminate between different pituitary celltypes, or even between normal and tumoral cells. Further, the
levels of circulating hormones given as replacement for pi-tuitary hormone deficiencies cannot be finely tuned, as en-dogenous hormones normally are. One way of overcomingsome of these limitations is the development of gene therapy(1). The development of pituitary cell type-specific vectorsshould allow endogenous promoters to drive expression ofa variety of therapeutic transgenes, either for hormone re-placement, or to selectively modify the function of a partic-ular cell type (2, 3). The use of cell type-specific promoterswould allow transgene expression to be maintained underthe control of the endogenous physiological regulators. Aparticular implementation of this strategy would be the useof cell type-specific promoters to eliminate tumors originat-ing from a specific pituitary cell type. The pituitary lactotro-phic cell was chosen as a target for this work both becauseof the availability of specific promoter sequences and be-cause of the existence of well characterized in vitro and in vivomodel systems in which to test their function.
A 5,000-bp human PRL promoter (hPrl) has been exten-sively studied and shown to allow pituitary-specific expres-sion (4, 5). The specificity of the rat PRL promoter has beendemonstrated in vivo using transgenic mice (6). The proximal
Received December 28, 1999.Address all correspondence and requests for reprints to: Professor
M. G. Castro, Molecular Medicine and Gene Therapy Unit, School ofMedicine, Room 1.302, Stopford Building; University of Manchester,Manchester, Oxford Road, Manchester M13 9PT, United Kingdom.E-mail: [email protected].
* This work was supported by grants from the BBSRC (UK) and theRoyal Society (to M.G.C.); CRC (UK) (to M.G.C. and P.R.L.) and Euro-pean Union-Biomed program grants Contract No. BMH4-CT98–3277,BMH4-CT98–0297(to P.R.L., M.G.C., and D.K.).
† These authors contributed equally to this work and should beconsidered first authors.
‡Training Fellow supported by Action Research (UK).§ Funded by a BBSRC Studentship.¶ Fellow of The Lister Institute of Preventive Medicine.
0013-7227/00/$03.00/0 Vol. 141, No. 9Endocrinology Printed in U.S.A.Copyright © 2000 by The Endocrine Society
3493
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022

region of the promoter (133bp to 2422 bp) is sufficient toprovide lactotroph specific expression; the distal region(21500 bp to 21800 bp) is required for high levels of ex-pression. For this reason, we have decided to use the 114 bpto 24429 bp sequences containing the proximal element (240bp to 2250 bp), the distal element (21300 bp to 21700 bp),and a portion of the super distal region (23500 bp to 25000bp) of hPrl promoter for our studies.
Castro et al. (1997) (7) were the first to demonstrate thefeasibility of using both adenoviruses and herpes simplextype-1 vectors to transfer genes into anterior pituitary cellsin primary culture (8). We have also recently used herpessimplex type-1 thymidine kinase (HSV1-TK) encoded withina replication defective adenovirus vector under the control ofa strong ubiquitous promoter (hCMV) to successfully inhibitpituitary lactotroph hyperplasia and reduce circulating PRLlevels in an animal model (9).
In this paper, we demonstrate lactotrophic cell type-specific expression of the marker gene b-galactosidase, andthe conditional cytotoxic gene HSV1-TK, both driven by thehPrl promoter and encoded within recombinant adenovirusvectors. Cell type-specific expression was demonstrated inanterior pituitary tumor cell lines, anterior pituitary cells inprimary culture, and the pituitary gland in vivo. Our resultsshow, for the first time, transcriptional targeting of a markertransgene (b-galactosidase), as well as the therapeutic trans-gene HSV1-TK, to a predetermined endocrine cell popula-tion within the AP gland in vivo.
When an adenovirus vector encoding HSV1-TK driven bythe hPrl promoter was used in an in vivo model of estrogen/sulpiride induced lactotroph hyperplasia within the anteriorpituitary (AP) gland in situ, the infection of the AP glandcombined with the administration of ganciclovir, was notable to reduce neither the weight of the gland, the numberof lactotrophic cells in vivo, nor the circulating PRL levels.This is in contrast to the use of an adenovirus expressingHSV1-TK under the transcriptional control of the humancytomegalovirus promoter (hCMV), employed in the sameexperimental paradigm. Such a vector, in combination withganciclovir, was effective in reducing pituitary weight andalso circulating PRL levels. Therefore, our results highlightthat further engineering of these cell type-specific promoterswill be needed to develop effective cell type-specific genetherapy strategies for the treatment of pituitary tumors usingtranscriptional targeted approaches.
Our results also have important implications for the designof gene therapy strategies for pituitary tumors. Our datademonstrate that both the choice of the in vivo animal model(e.g. anterior pituitary adenoma in situ vs. transplantabletumors), as well as the gene therapy strategy chosen (e.g. useof strong ubiquitous promoters vs. weaker but cell type-specific promoters), does influence the therapeutic outcome.
Materials and MethodsCell lines and culture conditions
All cell culture reagents were purchased from Sigma (Poole, Dorset,UK) or Life Technologies, Inc. (Paisley, UK). All tissue culture plastic-ware was purchased from Greiner (Stonehouse, Gloustershire, UK).AtT20 cells (provided by Dr. F. Antoni, Medical Research Council BrainMetabolism Unit, Department of Pharmacology, University of Edin-
burgh, Scotland, UK), and GH3 cells (by Dr. S. Cockle, Department ofBiochemistry and Physiology, University of Reading, Reading, UK),were grown in DMEM supplemented with sodium pyruvate (1 mm),glutamine (2 mm), nonessential amino acids (0.2 mm), 10% (vol/vol)horse serum, and 5% (vol/vol) newborn calf serum at 37 C in a 5% CO2atmosphere. This medium will be referred to as complete growth me-dium. Rat AP primary cell cultures were prepared as described previ-ously (7).
Reagents and antibodies
Different cell types within the AP cultures were identified using thefollowing polyclonal antibodies: guinea-pig antirat b-TSH (1/100), guin-ea-pig antirat PRL (1/500), guinea-pig antirat b-LH (1/100), guinea-pigantihuman GH (1/500), sheep antihuman b-FSH (1/500), and sheepantihuman ACTH (1/500) (provided by NIDDK’s National Hormoneand Pituitary Program, Bethesda, MD). The antibody used to identifyb-galactosidase was a rabbit polyclonal anti-b-galactosidase (1/750) (9);rabbit anti-HSV1-TK antibody (1/1000) was kindly provided by M.Janicot, Rhone-Poulenc-Rorer, France (10, 11).
Secondary antibodies used for either single or double immunolabel-ing were FITC or Texas Red-donkey antirabbit and FITC or Texas Red-goat anti-guinea-pig from Jackson ImmunoResearch Laboratories, Inc.(West Grove, PA).
Generation of recombinant adenoviruses expressingtransgenes under the control of the human PRL promoter
The transgenes, HSV1-TK and b-galactosidase, were excised from theplasmids pMV60/HSV1-TK (10) and pMV12/lacZ (12) by BamHI di-gestion and cloned into a plasmid derived from pGEM9ZF (PromegaCorp., Madison, WI), that contained the neuronal specific enolase (NSE)promoter and the SV40 polyA generating pGEM9ZF/NSE/HSV1-TK/polyA and pGEM9ZF/NSE/lacZ/polyA. The HSV1-TK/polyA andlacZ/polyA cassettes were excised from these plasmids by NspV/XhoIdigestion and the NspV site adapted to NcoI using a linker. The adaptedfragments were then cloned into the p5000 plasmid, containing the 5kbhPRl, in the NcoI/XhoI sites (13). The hPrl/HSV1-TK/polyA cassettewas excised by XbaI/XhoI digestion and cloned into the XbaI/SalI sitein pDE1sp1a (Microbix Biosystems, Toronto, Ontario, Canada). ThehPrl/lacZ/polyA cassette was excised by MunI/XhoI digestion andcloned into the EcoRI/XhoI site in pDE1sp1a.
RAds were generated by cotransfection using the calcium phosphatecoprecipitation method (14) with each shuttle vector and pBHG10 (Mi-crobix Biosystems), and purified using double cesium chloride gradientas previously described (15). Viral DNA was obtained as described byRevah et al., 1996 (16). To confirm the presence of the transgenes, viralDNA digestion with HindIII and subsequent Southern blot hybridiza-tion was performed. The specific probes used were: the 2982 bp lacZfragment excised using BamHI (15033 to 18015) from p5000/hPrl/lacZ,and the 1131 bp HSV1-TK fragment excised using BamHI (15033 to16164) from p5000/hPrl/HSV1-TK. The probes were labeled by ran-dom priming with digoxigenin-dUTP as described by the manufacturers(Roche Molecular Biochemicals, Bell Lane, East Sussex, UK).
The RAd-hCMV/HSV1-TK (RAd128), and RAd-hCMV/b-gal,(RAd35) have been described in detail previously (10–12). RAd-mCMV/b-gal (RAd36) was constructed using the murine CMV promoter (21336to 136 bp) (17) driving the expression of the marker gene b-galactosi-dase. RAd stocks were assayed and shown to be negative, for the pres-ence of replication competent adenovirus or endotoxin (lipopolysac-charide) as previously described (18, 19).
Cell type-specific expression of b-galactosidase in thepituitary tumor cell lines, GH3 and AtT20s: X-galhistochemistry and enzymatic activity from RAdPrl/lacZ,RAd35, and RAd36
GH3 and AtT20 cells were infected at multiplicity of infection (MOI)(number of infectious virus particles/cell) 30 with RAd35, RAd36, orRAd-Prl/lacZ. The cells were incubated for a further 2 days and X-galhistochemistry was performed as described previously (7).
Enzymatic activity was tested by infecting GH3 and AtT20 cells at
3494 IN VIVO TRANSCRIPTIONAL TARGETING Endo • 2000Vol 141 • No 9
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022

FIG. 1. Diagrams of the genomic organization of RAds usingthe hPrl promoter to drive the expression of b-galactosidase(A), and HSV1-TK (B) and the mCMV promoter to drive theexpression of b-galactosidase (C). H, Indicates the location ofHindIII restriction sites in the viral genome. The Southernblots show bands that correspond to the presence of the trans-gene in the viral DNA. A, The 3,257-bp band of lacZ/polyA wasdetected using a labeled lacZ probe; B, the 1,406-bp band ofHSV1-TK/polyA is detected using a labeled HSV1-TK probe;and C, the 5,000-bp band of mCMV/lacZ/polyA was detectedusing a mCMV/lacZ/polyA labeled probe.
IN VIVO TRANSCRIPTIONAL TARGETING 3495
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022

increasing MOIs (0, 3, 10, 30, 100, and 300) with RAd35, RAd36, orRAd-Prl/lacZ. The cells were incubated for 2 days and then harvestedin lysis buffer [25 mm Tris-HCL (pH7.8), 6.7% (vol/vol) glycerol, 10 mmMgCl2, 0.01% (vol/vol) Triton X-100, 1 mm EDTA (pH8)]. The lysateswere incubated with the O-nitrophenol-b-d-galactopyranoside (ONPG)(4 mg/ml) substrate solution at 37 C for appropriate time intervals untilcolor developed within the linear range of the standard curve. Thereactions were stopped by the addition of 510 ml Na2CO3 and sampleswere read on a spectrophotometer (Amersham Pharmacia Biotech, StAlbans, UK) at 420 nm. The enzyme activity was expressed as units ofb-galactosidase produced per cell. Each experimental condition wasdone in quadruplicate, and each experiment was repeated twice.
Cell type-specific expression of HSV1-TK and apoptosis inpituitary tumor cell lines
GH3 and AtT20 cells were infected with RAdPrl/HSV1-TK at an MOI30 for 48 h. The virus was then removed, and fresh medium was added.The cells were then incubated for a further 2 days. HSV1-TK transgeneexpression was assessed using immunofluorescence techniques as de-scribed previously (7). Cellular nuclei were stained using 4,6-diamidino-2-phenylindole (DAPI).
Simultaneous detection of HSV1-TK protein and cellular DNA con-tent within GH3 and AtT20 cells following infection with RAd-Prl/HSV-TK was performed by flow cytometry as described previously (9).Fifty thousand cells were plated in six-well plates and infected withRAdPrl/HSV1-TK at increasing MOI (0, 1, 5, 20, 50, and 100). Experi-ments were performed twice. After 48 h infection, the cells were fed withcomplete growth medium containing 10 mm ganciclovir (GCV; RocheMolecular Biochemicals, Welwyn Garden City, UK) and incubated fora further 72 h. Cells were harvested and then processed for DNA contentand HSV1-TK immunoreactivity as described previously (9, 20, 21).
Infection and detection of HSV1-TK within endocrine cellsin primary AP cultures
AP cells were infected with RAd-Prl/HSV1-TK at an MOI 30, andincubated for 48 h. The virus was removed, fresh complete growth
medium was added, and the primary cultures were incubated for 3additional days. Colocalization of the HSV1-TK transgene within spe-cific cell types present within the primary AP culture was assessed usingimmunofluorescence techniques as described previously (9); the cellularnuclei were stained using DAPI. For the quantification of endocrineand/or transduced cell types in vitro, 10 random fields within each wellwere counted.
Animals
Male 8-week-old Buffalo rats were house bred at the University ofManchester Biological Safety Unit. All animals had free access to foodand water, a 12-h light, 12-h dark cycle, and constant housing temper-ature and humidity. Experiments were conducted according to theUnited Kingdom Animal (Scientific Procedures) Act of 1986.
FIG. 2. Histochemical analysis of b-galactosidase expression in GH3and AtT20 tumor cell lines. Tumor cell lines were infected with RAd-hCMV/lacZ, RAd-mCMV/lacZ, or RAd-Prl/lacZ at MOI 30. Two dayslater, the cells were stained using X-gal histochemistry and analyzedusing light microscopy. Magnification, 310.
FIG. 3. Quantitative analysis of b-galactosidase enzyme activity inpituitary tumor cell lines. AtT20 or GH3 cells were infected withincreasing MOIs of RAd-hCMV/lacZ, RAd-mCMV/lacZ, or RAd-Prl/lacZ. Two days later, the cells were harvested and tested for b-galactosidase activity. These results were then corrected using thenumber of cells present in the cultures and transgene expression isshown as b-galactosidase enzymatic activity (units: U) per cell(mean 6 SEM; n 5 4).
3496 IN VIVO TRANSCRIPTIONAL TARGETING Endo • 2000Vol 141 • No 9
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022

In vivo gene delivery to the anterior pituitary gland
Male 8-week-old Buffalo rats (house bred) were anesthetized withhalothane and placed in a sterotaxic frame. The skull was exposed,bregma was identified, and a hole was drilled posterior to bregma untilrevealing the superior sagittal sinus, and surrounding brain. A thinsurgical suture was passed underneath the vein, and the meningessurrounding the vein were cut to allow the vein to be displaced laterallywithout lesioning the vein wall or the surrounding cortex. Intrapituitaryinjections were made using a 26-gauge Hamilton syringe needle. The tipof the needle had been previously ground until the opening of the needlewas positioned at the base of the tip. Injections were made at the fol-lowing coordinates: antero-posterior from Bregma, 20.57, 20.60, and20.63 cm, and lateral on each side of the midline at 0.05 cm. Thus, wemade a total of six injections per pituitary gland. Previous attempts toinject directly into the pituitary using exclusively stereotaxic coordinatesfailed in reliably delivering directly the RAds into the gland. Thus, wedeveloped a modified strategy as follows: the modified Hamilton needlewas lowered at each coordinate until touching the sphenoidal bone, andmaking contact with the bottom of the rat equivalent of the sella turcica.This leaves the opening of the needle within the pituitary gland, andadequate amounts of recombinant vector were then injected. Underthese conditions of injection, the pituitary was transduced by recombi-nant adenoviruses in 100% of surgical attempts. At each of these sixcoordinates, 1 ml of the recombinant vector [1 3108 (8) pfu] was thendelivered over 1 min per injection site. Animals were then given 10 mlof saline sc and allowed to recover. Forty-eight hours later, animals wereperfused transcardially with Tyrode solution (132 mm NaCl, 1.8 mmCaCl2, 0.32 mm NaH2PO4, 5.56 mm glucose, 11.6 mm NaHCO3, and 2.68mm KCl), pituitary glands were removed and placed in 4% paraformal-dehyde dissolved in 0.1 m PBS for 3 h. Tissue was then paraffin embedded,sectioned using a microtome (5 mm) (Leica Corp.), and mounted onto3-aminopropyltriethoxysilane (APES) (Sigma)-coated glass slides.
Induction of lactotroph hyperplasia and delivery of RAdsinto the AP gland
Male 8-week-old Buffalo rats were implanted with SILASTIC brandpellets (Dow Corning Corp., Midland, MI) containing 15 mg of 17b-estradiol and 50 mg of sulpiride, prepared as previously described (22).The pellets were implanted sc in the lumbar region of each rat underanesthesia. Empty SILASTIC brand pellets were implanted as controls.Three days later, the animals were anesthetized with Fluothane and atotal of 1 3 108 pfu in 6 ml of either RAd35, RAd128 or RAd-PRL/HSV1-TK was delivered to the anterior pituitary gland as describedabove. Rats were given a 10ml sc saline injection post surgery, andglucose was added to their drinking water every 2 days to preventdehydration. One day after surgery, all animals received ip GCV injec-tions at a dose of 25 mg/kg twice daily for 7 days. The animals werekilled using a lethal overdose of pentobarbital and perfused with Ty-rode’s solution. Pituitary, body, and testis weights were recorded. Before
perfusion, trunk blood was collected. Pituitary glands were then placedin 4% paraformaldehyde dissolved in 0.1 m PBS for 3 h. Tissue was thenparaffin embedded, sectioned using a microtome (5 mm), and mountedonto APES-coated glass slides.
Immunohistochemical detection of transgene expressionwithin the anterior pituitary gland in vivo usingfluorescence microscopy
Sections were deparaffinated using xylene for 5 min then rehydratedthrough graded alcohols (100%, 95%, 85%, 70%, 50% ethanol), 3 min eachbefore being washed in saline (0.8% NaCl wt/vol) for 5 min. The block-ing solution was prepared using horse serum (10% vol/vol or 1% vol/vol) diluted in PBS, containing 0.1% Triton X-100. The sections were thenincubated in (1): 10% blocking solution for 2 h at room temp (2); 1%blocking solution for 1 h at room temp (3); primary antibody (diluted in1% blocking solution) for 1 h at room temp (4); five washes in PBScontaining 0.5% Triton X-100 for 5 min (5); secondary antibody (dilutedin 1% blocking solution) for 1 h at room temp; and (6) five washes in PBScontaining 0.5% Triton X-100 for 5 min. The sections were stained withDAPI for 15 min, washed twice in PBS for 5 min, once in dH2O for 5 min,and mounted in Mowiol (Calbiochem Nottingham, UK). Images wereacquired using Openlab software (Improvision, Coventry, UK) on anOlympus Corp. (Tokyo, Japan) Vanox microscope. For the quantificationof transduced or nontransduced pituitary cells, five fields within the areaof the anterior pituitary, which expressed transgenes encoded by viralvectors, were counted.
Determination of hormone levels in peripheral blood
Rat plasma PRL, GH, and TSH-b, concentrations were determinedusing specific RIA kits provided by the National Hormone and PituitaryProgram, NIDDK, and Dr A. F. Parlow (Torrance, CA). Plasma ACTHwas measured using a specific immunoradiometric assay that has beendescribed previously (23).
Statistical analysis
The in vitro and in vivo experimental results were analyzed usingeither ANOVA, followed by the Student’s-Neuman-Keuls multiple com-parison test or the Student’s t test where appropriate, using GraphPadSoftware, Inc. Instat Version 2 (GraphPad Software, Inc., San Diego, CA).
ResultsConstruction of adenoviral vectors expressing transgenesunder the control of the human PRL promoter
Adenoviral vectors were generated by homologous re-combination between the adenoviral genome in pBHG10 and
FIG. 4. Cell-type specific expression ofHSV1-TK under the control of the PRLpromoter in pituitary tumor cell linesGH3 and AtT20. GH3, a PRL/GH-secreting tumor cell line, and AtT20, acorticotrophic tumor cell line, were in-fected with RAd-Prl/HSV1-TK at MOI30. After 48 h infection, the cells werestained for HSV1-TK and the nucleusstained using DAPI. Arrows point to nu-clei of representative cells in HSV1-TKand DAPI panels. Note: cytoplasmicand nuclear distribution of HSV1-TKexpression. Magnification, 3250.
IN VIVO TRANSCRIPTIONAL TARGETING 3497
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022

the plasmid pDE1sp1a containing the transgenes, i.e. HSV1-TK, or b-galactosidase under the control of 114 bp to 24429bp or 114 bp to 24152 bp of the hPrl promoter, respectively,and both followed by the SV40 polyadenylation sequence.The expression construct was inserted within the E1 regionof the adenoviral genome. When DNA from these recombi-nant RAds was digested with HindIII, and probed usingSouthern blot hybridization, a band of the right size, indi-cating the presence of the transgenes within the recombinantvirus genome, was observed, i.e. the 1406 bp HSV1-TK/polyA band, the 3257 bp lacZ/polyA band, and the 5000 bpmCMV/lacZ,polyA band (Fig. 1, A–C).
Cell type-specific expression of b-galactosidase withinpituitary tumor cell lines, GH3 and AtT20
b-galactosidase expression was assessed using X-gal his-tochemical staining, in the GH3 and AtT20 cell lines afterinfection with RAd36 (mCMV/lacZ), RAd35 (hCMV/lacZ),or RAd-Prl/lacZ. When the cells were infected with RAd-Prl/lacZ, only the lactotroph GH3 cells expressed the b-galactosidase enzyme (Fig. 2). No expression of b-galacto-sidase was observed in the corticotrophic AtT20 cell line afterinfection with RAd-Prl/lacZ. These results show that the PRLpromoter can restrict expression of the marker gene b-galactosidase to established lactotroph tumor cell lines invitro.
To quantitate promoter strength from the hCMV, mCMV,and hPrl promoters, within RAds, b-galactosidase enzymeactivity was assessed in the lactotrophic GH3 cells, and thecorticotrophic AtT20 cells following viral infection (Fig. 3).Both in GH3 cells and AtT20 cells, the levels of enzymeactivity increased with increasing MOIs used, of eitherRAd35 (hCMV/lacZ) or RAd36 (mCMV/lacZ). After infec-tion of GH3 cells with RAd-Prl/lacZ, there was an increase inenzyme activity up to MOI 300. After infection of AtT20 cellswith RAd-Prl/lacZ, no enzyme activity above basal levels wasobserved, even at the highest MOI used, i.e. 300. In GH3 cells,at MOIs 100 and 300, there were 13- and 24-fold more enzymeactivity respectively (P , 0.005), in cells infected with RAd36when compared with cells infected with RAd35 (Fig. 3). Levelsof transgene expressed under the control of the mCMV pro-moter in GH3 cells were 46-fold (P , 0.005) and 70-fold (P ,0.005) stronger at MOIs of 100 and 300 MOIs (Fig. 3) comparedwith those expressed under the control of the PRL promoter. InAtT20 cells at MOIs 100 and 300, the mCMV promoter was19-fold (P , 0.05) and 10-fold (P , 0.005) stronger than thehCMV promoter (Fig. 3). In GH3 cells at MOIs 100 and 300, thehCMV promoter was 3-fold (P , 0.005) stronger than the PRLpromoter at both MOIs.
Cell type-specific expression and cytotoxicity of HSV1-TKdriven by the hPrl promoter in pituitary tumor cell lines
The expression of HSV1-TK driven by the hPrl promoterwas also assessed in both GH3 and AtT20 cells using im-munofluorescence microscopy (Fig. 4). Infection of these cellswith RAd-Prl/HSV1-TK resulted in a strong immunocyto-chemical staining in the lactotrophic GH3 cells; there was noincrease, above basal levels, of apoptotic nuclei at the con-centration of virus used (MOI 30) (Fig. 4). In AtT20 cells, weobserved no HSV1-TK transgene expression (Fig. 4).
To quantitate the levels of HSV1-TK expression with increas-ing viral dose in AtT20 and GH3 cells, they were infected for48 h with RAd-Prl/HSV1-TK, using MOIs ranging from 0–100.Figure 5A shows a gradual increase in HSV1-TK immunore-activity, as detected by flow cytometry in the GH3 cell line only.The level of HSV1-TK immunoreactive protein was maximal atMOI 100 in the GH3 cells. HSV1-TK expression remained atbasal levels in AtT20 cells at all MOIs tested (Fig. 5A).
Infection of GH3 cells with RAd-Prl/HSV1-TK in the pres-ence of 10 mm GCV induced apoptosis, which was MOIdependent (Fig. 5B). In the presence and absence of GCV (10mm), even at the highest MOI of RAd-Prl/HSV1-TK used,
FIG. 5. Cell-type specific expression and cytotoxicity of HSV1-TKdriven by the hPrl promoter in GH3 and AtT20 cells. AtT20 and GH3cells were infected with RAd-Prl/HSV1-TK at increasing MOIs andthe percentage of cells expressing HSV1-TK was determined usingfluorescence activated cell sorting analysis (FACS) (A). After infectionwith increasing MOIs of RAd-Prl/HSV1-TK, GH3 (B), and AtT20 (C)cells were incubated with 10 mm GCV for 3 days and analyzed byFACS for apoptosis, using propidium iodide incorporation.
3498 IN VIVO TRANSCRIPTIONAL TARGETING Endo • 2000Vol 141 • No 9
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022

only basal levels of apoptosis were observed in AtT20 cellsindicating there was no HSV1-TK mediated toxicity (Fig. 5C).
Cell type-specific expression of transgenes expressed underthe transcriptional control of the hPrl promoter in ratprimary anterior pituitary cell cultures and in the anteriorpituitary gland in vivo
When rat primary anterior pituitary cultures were infectedwith RAd-Prl/HSV1-TK the expression of HSV1-TK wasmainly restricted to cells expressing PRL (Prl) (89% 6 7 of
cells expressing HSV1-TK also expressed PRL) (Fig. 6 andTable 1), and a subset of GH-producing cells (11% 6 0.8 oftotal HSV1-TK immunoreactive cell population also ex-pressed GH) (Fig. 6 and Table 1). Transgene expression un-der the control of the hPrl promoter was achieved in 60% 66.9 of the total lactotrophic population in anterior pituitarycells in vitro. Expression of the transgene HSV1-TK also ob-served in the 9% 6 0.9 of the total somatotrophic populationpossibly representing the subpopulation of cells that co-express PRL and GH, i.e. mammosomatotrophs. No expres-
FIG. 6. Cell-type specific expressiondriven by the PRL promoter ofHSV1-TK within endocrine anterior pi-tuitary (AP) cells in primary culture.Rat primary AP cultures were infectedwith RAd-Prl/HSV1-TK at MOI 30. Af-ter 48 h infection, the cultures werestained for hormones (PRL, GH, ACTH,TSH, LH, and FSH) and HSV1-TK ex-pression. Arrows point to the represen-tative hormone-producing cells and/orcells expressing HSV1-TK. Magnifica-tion, 3250.
IN VIVO TRANSCRIPTIONAL TARGETING 3499
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022

sion of HSV1-TK was observed in cells synthesizing FSH, LH,TSH, or ACTH. There was no indication of apoptosis asassessed by the nuclear integrity after DAPI staining (Fig. 6).
To determine the specificity of the hPrl promoter in theanterior pituitary gland in vivo, we compared the cell pop-ulations expressing HSV1-TK driven by the hPrl promoter,the hCMV, or the mCMV promoters. All recombinant ade-novirus vectors drove transgene expression in the anteriorpituitary gland in vivo (Figs. 7 and 8 and Table 2). The mCMVand the hCMV promoters drove transgene expression ofeither HSV1-TK or b-gal in all endocrine cell populationsencountered within the anterior pituitary gland (Figs. 7 and8 and Table 2). However, the hPrl promoter restricted ex-pression of HSV1-TK almost exclusively to Prl expressingcells (approximately 50% of the lactotrophic populationwithin the transduced area expressed the transgene) (Fig. 8and Table 2). A small number of GH immunoreactive cellswere also HSV1-TK immunoreactive (0.5–1% of total GHimmunoreactive cells). These cells are likely to be mammo-somatotrophs. There was no indication of apoptosis in thenormal pituitary glands infected with any of the recombinantadenovirus vectors as assessed by the nuclear integrity afterDAPI staining and microscopic examination of all endocrinecell populations (results not shown).
In vivo gene therapy of oestrogen/sulpiride inducedlactotroph hyperplasia using HSV-1 TK under the controlof either the hCMV or the hPrl promoter in combinationwith GCV
In control placebo-treated animals, PRL immunoreactivecells comprised 18 6 1% of the total pituitary cell populationwithin the transduced area (Table 2), whereas in the E/S-implanted animals treated with RAd35/GCV, the popula-tion rose to 23 6 0.5% of the total cell population within thearea of the anterior pituitary, which had been transduced bythe vector (P , 0.005 vs. control). In the E/S-implanted an-imal group treated with RAd128/GCV, we observed a de-crease in the PRL producing cell population to 15 6 1% of thetotal cell population (P , 0.05 vs. E/S implanted, RAd35/GCV-treated group) (Table 2). There was no change in thenumber of PRL positive cells in E/S implanted animals
treated with RAd-Prl/HSV1-TK. There were no significantchanges in GH, FSH, LH, ACTH, or TSH-b cell populationsin any of the treated animal groups with respect to controls(Table 2).
Pituitary weight in control animals averaged 11.3 6 0.5mg. whereas the E/S implanted animals treated withRAd35/GCV had a mean pituitary weight of 22.7 6 1.3 mg(P , 0.0005 vs. controls). The E/S implanted group treatedwith RAd128/GCV had an average pituitary weight of18.7 6 0.5 mg, an 18% reduction when compared with theE/S implanted RAd35/GCV treated group (P , 0.05). TheE/S implanted group treated with RAd-Prl/HSV1-TK/GCVhad pituitary weight of 22.9 6 0.3 mg (not significant vs. E/Simplanted group treated with RAd35/GCV).
Plasma Prl in placebo-implanted control rats was 38 6 4ng/ml, whereas in the E/S-implanted group treated withRAd35 and GCV, circulating Prl levels increased to 660 628.3ng/ml. In the E/S implanted group treated with RAd128and GCV, Prl levels were reduced to 330 6 50.1ng/ml (P ,0.005 vs. E/S implanted RAd35/GCV treated group), areduction of 50% (Fig. 9). In the E/S-implanted group treatedwith RAd-Prl/HSV1-TK and GCV, Prl levels were not sig-nificantly reduced. No changes were observed in circulatingACTH, GH, and TSH-b levels in any of the experimentalgroups (Fig. 9).
Discussion
An advantage of gene therapy, in comparison with clas-sical pharmacology, is the capacity to selectively modulatethe genetic makeup and function of identified pituitary cellsubtypes, while leaving surrounding pituitary cells of otherphenotypes unaffected (24). This will be of particular im-portance when targeting cytotoxic proteins to tumoral celltypes for the purpose of eliminating tumors, while leavingthe surrounding normal pituitary intact. The use of pituitarycell type specific promoters has been proposed to constitutea molecular tool to achieve pituitary cell type-specific tran-scriptional targeting of virally encoded transgenes (2, 3).
Our results indicate that the human PRL promoter en-coded within recombinant adenovirus vectors can restricttransgene expression, e.g. HSV1-TK or b-galactosidase, ex-clusively to lactotrophic cells both in vitro and in vivo. Therestriction of expression of HSV1-TK to lactotrophic cells,using the PRL promoter, resulted in the cell type-specificinduction of apoptosis in the lactotrophic tumor GH3 cellline, in the presence of GCV. In the corticotrophic AtT20 cellline, there was neither HSV1-TK expression, nor apoptosis inthe presence of GCV, even at an MOI 100. Therefore, thehuman PRL promoter is able to target the expression ofHSV1-TK to lactotroph tumor cells.
The measurement of b-galactosidase enzyme activity lev-els enabled us to quantitate and compare the levels of trans-gene expression directed by either the PRL promoter, or theconstitutive mCMV or hCMV promoters. The mCMV pro-moter has been reported to be stronger in a variety of humanand murine immortalized cell lines in vitro (25) or in the brainin vivo (26). Of the three promoters tested, i.e. hCMV, mCMV,and hPrl, the mCMV elicited the highest levels of transgeneexpression in both pituitary tumor cell lines. In GH3 cells, the
TABLE 1. Cell-type specific expression of HSV1-TK driven by thehuman prolactin promoter in primary cultures of the anteriorpituitary gland
Hormone % of total endocrine populationexpressing single hormones
% of hormone (1) cellsexpressing TK (1)immunoreactivity
PRL 31 6 3.3 60 6 6.9GH 23 6 5.5 9 6 0.9LH 3 6 0.7 0ACTH 22 6 0.7 0TSH 4 6 1.4 0FSH 5 6 1.4 0
Hormone-producing cells were stained for the various hormonesand analyzed for the coexpression of HSV1-TK whose expression wasdriven by the human PRL promoter after infection with RAd-PRL/HSV1-TK. Note that out of the total HSV1-TK immunoreactive cellpopulation, 89 6 7% coexpressed HSV1-TK and prolactin and 11 60.8% coexpressed GH and HSV1-TK. Ten fields were counted under340 magnification in the area of transduction.
3500 IN VIVO TRANSCRIPTIONAL TARGETING Endo • 2000Vol 141 • No 9
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022

FIG. 7. Expression of b-galactosidase within endocrine cell populations after in vivo delivery to the anterior pituitary gland of 6 3108 pfu of RAd35or RAd36. After 48 h infection, the pituitaries were removed, embedded in paraffin wax, sectioned, and stained for hormone and b-galactosidaseimmunoreactivity using double-labeling immunofluorescence techniques. RAd35 (left panels), RAd36 (right panels) infection resulted in the expressionof b-galactosidase in all endocrine cell populations. Low magnification images (3100) top panels show widespread distribution of b-galactosidaseexpression within the AP gland and, high magnification images (3500) show colocalization of hormone-producing cells expressing b-galactosidaseas indicated at the bottom of the panels. White arrows indicate cells that are labeled for both the hormone and b-galactosidase.
IN VIVO TRANSCRIPTIONAL TARGETING 3501
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022
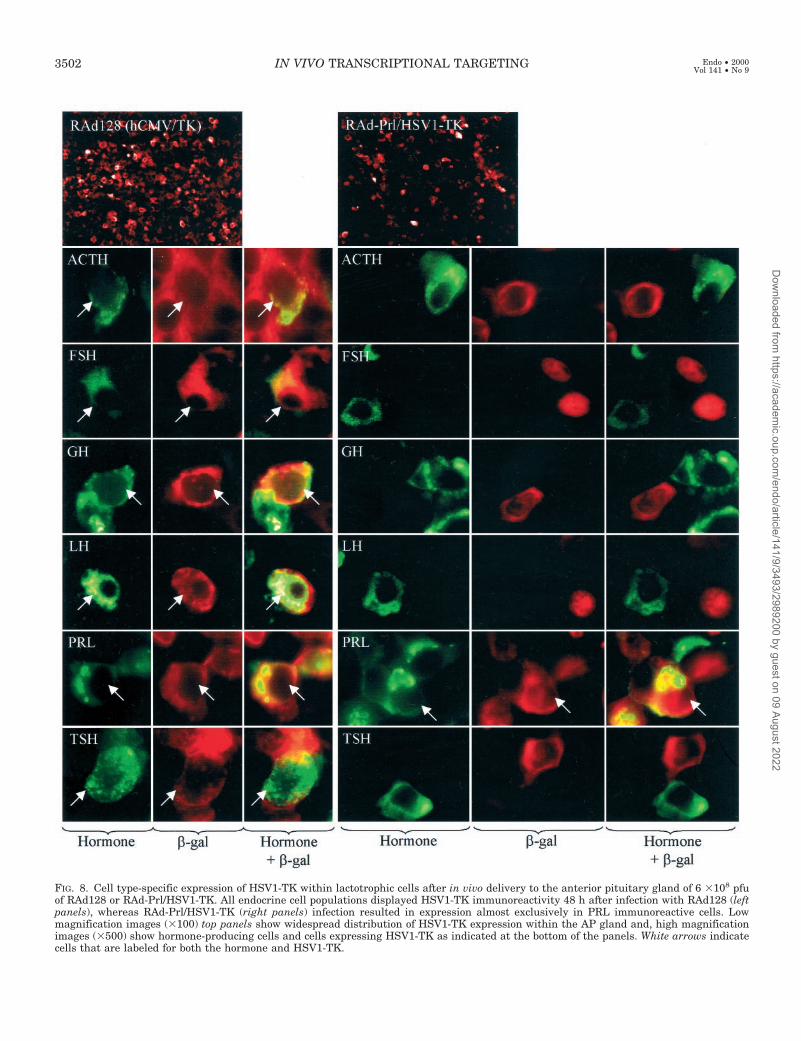
FIG. 8. Cell type-specific expression of HSV1-TK within lactotrophic cells after in vivo delivery to the anterior pituitary gland of 6 3108 pfuof RAd128 or RAd-Prl/HSV1-TK. All endocrine cell populations displayed HSV1-TK immunoreactivity 48 h after infection with RAd128 (leftpanels), whereas RAd-Prl/HSV1-TK (right panels) infection resulted in expression almost exclusively in PRL immunoreactive cells. Lowmagnification images (3100) top panels show widespread distribution of HSV1-TK expression within the AP gland and, high magnificationimages (3500) show hormone-producing cells and cells expressing HSV1-TK as indicated at the bottom of the panels. White arrows indicatecells that are labeled for both the hormone and HSV1-TK.
3502 IN VIVO TRANSCRIPTIONAL TARGETING Endo • 2000Vol 141 • No 9
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022

mCMV promoter elicited approximately 50-fold higher ex-pression when compared with the hPrl promoter.
The strength of the promoter used would determine theamount of virus needed to achieve a therapeutic effect. Byusing a stronger promoter, less virus would be needed toachieve adequate levels of transgene expression (25, 26). Thiswould therefore reduce toxic side effects due to high dosesof viral vector that have been previously shown to causeacute cytotoxicity (27) as well as chronic inflammation in thebrain (10).
Our work has shown that sequences 114 bp to 24152 bpor 24429 bp of the human PRL promoter can restrict trans-gene expression from an adenoviral vector to lactotrophiccells in primary anterior pituitary cultures and importantlywithin the anterior pituitary gland in vivo. However, trans-gene expression was also detected in a subpopulation of GHexpressing cells both in primary culture and in vivo. Thesecells are probably mammosomatotrophs, which are thoughtto be the transitional intermediates of lactotrophs and so-matotrophs (28), and synthesize both GH and PRL.
Lee et al. (1999), (3) have recently shown that the GH and
the a-glycoprotein subunit promoters could restrict the ex-pression of transgenes to somatotroph and null-cell tumorcell lines, respectively. They also showed that the size of sctumors, derived from implanted GH3 cells in nude mice,could be reduced by the delivery of an adenovirus expressingHSV1-TK, under the control of the GH promoter, when com-bined with the administration of GCV (3).
We decided to explore the in vivo delivery of HSV1-TKdriven by the human PRL promoter encoded within recom-binant adenovirus vectors to the anterior pituitary in a ratmodel of estrogen/sulpiride induced pituitary hyperplasia.This enabled us to demonstrate for the first time that, al-though the human PRL promoter is able to restrict transgeneexpression mainly to lactotrophic cells within the anteriorpituitary gland in vivo, the level of expression of the thera-peutic transgene (HSV1-TK) was not sufficient to achieve ameasurable therapeutic outcome.
In contrast, the ubiquitous hCMV promoter driving ex-pression of HSV1-TK, which we used as a positive control forthis experiment, was able to elicit a beneficial therapeuticoutcome. This confirms our previous results showing that
FIG. 9. Changes in circulating hor-mone levels following in vivo gene ther-apy treatment in an estrogen/sulpiride(E/S) implanted in vivo model. Plasmawas analyzed for Prl, GH, and TSH lev-els using specific RIAs (9). The ACTHlevels were analyzed using a specificIRMA (23). The various hormone levelsfor the E/S implanted RAd128/GCV,RAd-PRL/HSV1-TK/GCV and RAd35/GCV-treated groups were compared byfold increase above the control hormonelevels in placebo implanted animals. *,P , 0.005.
TABLE 2. Efficiency of transgene expression and cell ablation within hormone producing anterior pituitary cells in vivo within normaland estrogen/sulpiride induced hyperplastic anterior pituitary glands
Group % of total pituitary cell population PRL GH LH ACTH TSH FSH
Placebo 1 RAd35 1 GCV % Hormone 1ve cells 18 6 1 17 6 2 3 6 1 8 6 1 9 6 1 4 6 125 mg/kg ip twice daily % b-gal 1ve cells 14 6 1.5 13 6 2 11 6 1.5 13 6 1 12 6 1 12 6 1.5(n 5 4) % Both hormone and b-gal 1ve cells 7 6 1 2 6 1 1 6 0.5 2 6 1 1 6 0.5 1 6 0.5E/S 1 RAd35 1 GCV % Hormone 1ve cells 23 6 0.5a 15 6 2.5 3 6 0.5 7 6 0.5 7 6 1 4 6 0.525 mg/kg ip twice daily % b-gal 1ve cells 12 6 1 14 6 1.5 12 6 1 13 6 1 13 6 1 10 6 1(n 5 4) % Both hormone and b-gal 1ve cells 7 6 1.5 2 6 0.5 1 6 0.5 2 6 0.5 2 6 0.5 1 6 0.5E/S 1 RAd128 1 GCV % Hormone 1ve cells 15 6 1b 15 6 2 3 6 0.5 6 6 1 9 6 1 4 6 0.525 mg/kg ip twice daily % TK 1ve cells 27 6 2 29 6 1.5 28 6 2.5 24 6 1 28 6 1.5 26 6 2(n 5 4) % Both hormone and TK 1ve cells 7 6 1 6 6 2 2 6 0.5 2 6 0.5 4 6 1 1 6 0.5E/S 1 RAd-PRL/TK 1 GCV % Hormone 1ve cells 21 6 1.5cd 17 6 3.5 3 6 0.5 7 6 1 8 6 1 3 6 125 mg/kg ip twice daily % TK 1ve cells 11 6 1 12 6 1 13 6 1.5 11 6 1 9 6 1 11 6 1(n 5 4) % Both hormone and TK 1ve cells 10 6 1.5e 1 6 0.5 0 0 0 0
The control animals injected with RAd35/GCV and the E/S implanted animals injected with RAd35/GCV, or RAd128/GCV, or RAd-PRL/HSV-1TK/GCV-treated rats were killed and perfused with Tyrodes solution and processed for double immunocytochemistry as described inMaterials and Methods. Hormone producing anterior pituitary cells were stained for the various hormones and analysed for the coexpressionof HSV1-TK whose expression was driven by hCMV promoter (RAd128) or the human PRL promoter (RAd-PRL/HSV1-TK) or b-galactosidasewhose expression was driven by the hCMV promoter (RAd35). Five fields within the area of the pituitary that had been transduced by the vectorswere counted under 360 magnification. Significant differences between groups are: a P , 0.005 vs. placebo group; b P , 0.05 vs. E/S RAd35/GCVtreated group; c not significant vs. placebo or E/S RAd35/GCV treated groups; d P , 0.05 vs. E/S RAd128/GCV-treated group; e not significantvs. E/S RAd128/GCV-treated group.
IN VIVO TRANSCRIPTIONAL TARGETING 3503
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022

delivery of HSV1-TK driven by the hCMV promoter to ratsbearing estrogen-induced lactotroph hyperplasia, into theAP gland via the transauricular route followed by subse-quent treatment with GCV decreased plasma PRL levels andreduced the mass of the pituitary gland (9). Neither RAd-Prl/HSV1-TK nor RAd-hCMV/HSV1-TK (RAd128) causeddeleterious effects on circulating levels of other anterior pi-tuitary hormones, suggesting that the treatment was non-toxic to the normal endocrine anterior pituitary cells in situ(9), although hormone secretion in response to secretagogueswas not tested. Even if the hCMV promoter is not specific tothe hyperplasic lactotrophic cells, its toxic effects ought to berestricted to those cells, which are actively dividing. In theestrogen/sulpiride model used, only lactotrophic cells areactively dividing. Although the hCMV promoter has no cell-type specificity, in our model only the lactotrophic cells areeliminated, due to the molecular mechanism of cell killing ofHSV1-TK in combination with GCV, which only affects ac-tively dividing cells.
The lessons to be learned from the in vivo experimentdescribed in this paper are very important, not only in termsof the implications for the development of gene therapystrategies for pituitary disease, but also for gene therapy ingeneral. Although cell type-specific promoters are efficient inrestricting transgene expression to predetermined cell types,in this case such a promoter was not strong enough to pro-duce a beneficial therapeutic outcome. Furthermore, a non-specific promoter used in conjunction with a cell killingmechanism that selectively kills actively dividing cells, pro-vided the restricted elimination of hyperplasic lactotrophpituitary cells, without adversely affecting other pituitarycell types.
Our results contrast with those published previouslyshowing tumor regression in a GH transplantable tumormodel in nude mice (3). We believe the different resultsdepend on the particular model used. The transplantablemodel employs a very rapidly growing tumor in whichHSV1-TK would elicit a very strong effect. We believe thatour model represents more closely the situation that wouldbe encountered in a human pituitary tumor in which theproliferative index is low. Because the efficiency of theHSV1-TK plus GCV system depends on both the efficiencyof virus transduction and the rate of cell proliferation, wepredict higher levels of HSV1-TK expression to be requiredto efficiently reduce lactotrophic cells’ proliferation and PRLhypersecretion in our in situ tumor model. This explains, whyHSV1-TK expressed under the control of the hCMV pro-moter, but not, under control of the hPrl promoter, showeda significant therapeutic benefit. Again our results indicatethe importance of properly designed preclinical studies be-fore these therapeutic approaches are taken into the clinic.
Acknowledgments
We are very grateful to Dr. D. Ray for useful comments and discus-sions and to Mrs. R. Poulton and Ms. Tricia Maleniak for expert secre-tarial and technical assistance respectively. We would also like to thankDr. A. F. Parlow (National Hormone and Pituitary Program, Harbor-UCLA Medical Center, Torrance, CA) for the supply of hormone RIAkits and immunocytochemistry antibodies specific for the pituitary hor-mones; A. White, Endocrine Sciences Research Group, School of Med-icine and Biological Sciences, University of Manchester, for the deter-
mination of the plasma ACTH levels; E. Linton, Nuffield Department ofObstetrics and Gynaecology, University of Oxford for the supply ofprecipitating reagents for the RIAs; and Dr. R. Goya, School of Medicine,University of La Plata, Argentina, for help with iodination and RIAprotocols. We also wish to thank Prof A. M. Heagerty, for his continuoussupport and encouragement.
References
1. Castro MG 1999 Gene therapy strategies for the treatment of pituitary tumors.J Mol Endocrinol 22:9–18
2. Castro MG, Windeatt S, Smith-Arica J, Lowenstein PR 1999 Cell-type specificexpression in the pituitary: physiology and gene therapy. Biol Soc Trans27:858–863
3. Lee, EJ, Anderson M, Thimmapaya B, Jameson JL 1999 Targeted expressionof toxic genes directed by pituitary hormone promoters: a potential strategyfor adenovirus-mediated gene therapy of pituitary tumors. J Clin EndocrinolMetab 84:786–794
4. Peers B, Voz ML, Monget P, Mathy-Hartert M, Berwaer M, Belayew A,Martial JA 1990 Regulatory elements controlling pituitary-specific expressionof the human prolactin gene. Mol Cell Biol 10:4690–4700
5. Berwaer M, Monget P, Peers B, Mathy-Hartert M, Bellefroid E, Davis JRE,Belayew A, Martial JA 1991 Multihormonal regulation of the human prolactingene expression from 5000bp of its upstream sequence. Mol Cell Endocrinol80:53–64
6. Crenshaw III EB, Kalla K, Simmons DM, Swanson LW, Rosenfeld MG 1989Cell-specific expression of the prolactin gene in transgenic mice is controlledby synergistic interactions between promoter and enhancer elements. GenesDev 3:959–972
7. Castro MG, Goya RG, Sosa YE, Rowe J, Larregina A, Morelli A, LowensteinPR 1997 Expression of transgenes in normal and neoplastic anterior pituitarycells using recombinant adenoviruses: long term expression, cell cycle depen-dency, and effects on hormone secretion. Endocrinology 138:2184–2194
8. Goya R, Rowe J, Sosa YE, Tomasec P, Lowenstein PR, Castro MG 1998 Useof recombinant herpes simplex virus type 1 vectors for gene transfer into tumorand normal anterior pituitary cells. Mol Cell Endocrinol 138:199–207
9. Windeatt S, Southgate TD, Dewey RA, Bolognani F, Perone MJ, LarreginaAT, Maleniak TC, Morris ID, Goya RG, Klatzmann D, Lowenstein PR,Castro MG 2000 Adenovirus-mediated herpes simplex virus type-1 thymidinekinase (HSV1-TK) gene therapy suppresses oestrogen induced pituitary pro-lactinomas. J Clin Endocrinol Metab 85:1296–1305
10. Dewey RA, Morrissey G, Cowsill CM, Stone D, Bolognani F, Dodd NJF,Southgate TD, Klatzmann D, Lassmann H, Castro MG, Lowenstein PR 1999Chronic brain inflammation and persistent HSV1-TK expression in survivorsof syngeneic glioma treated by adenovirus-mediated gene therapy: implica-tions for clinical trials. Nature Med 5:1256–1264
11. Cowsill C, Southgate TD, Morrissey G, Dewey RA, Morelli AE, MaleniakTC, Forrest Z, Klatzmann D, Wilkinson GWG, Lowenstein PR, Castro MG2000 Central nervous system toxicity of two adenoviral vectors encodingvariants of the herpes simplex virus type 1 thymidine kinase: reduced cyto-toxicity of a truncated HSV1-TK. Gene Ther 7:679–685
12. Shering AF, Bain D, Stewart K, Epstein AL, Castro MG, Wilkinson GWG,Lowenstein PR 1997 Cell-type specific expression in brain cell cultures froma short human cytomegalovirus major immediate early promoter depends onwhether is it inserted into herpesvirus or adenovirus vectors. J Gen Virol78:445–459
13. Takasuka N, White MRH, Wood CD, Robertson WR, Davis JRE 1998 Dy-namic changes in prolactin promoter activation in individual living lactotro-phic cells. Endocrinology 139:1361–1368
14. Castro MG, Rowe J, Morrison E, Tomasec P, Murray CA, Shering AF, Lo-wenstein PR 1996 Calcium-phosphate, DEAE-dextran coprecipitation, andelectroporation to transfer genes into neuronal and glial cell lines. In: Lowen-stein PR, Enquist LW (eds) Protocols for Gene Transfer In Neuroscience To-wards Gene Therapy of Neurological Disorders. John Wiley & Sons, Chich-ester, pp 9–24
15. Lowenstein PR, Shering AF, Bain D, Castro MG, Wilkinson GWG 1996 Theuse of adenovirus vectors to transfer genes to identified target brain cells invitro. In: Lowenstein PR, Enquist LW (eds) Protocols for Gene Transfer InNeuroscience: Towards Gene Therapy of Neurological Disorders. John Wiley& Sons, Chichester, pp 93–114
16. Revah F, Horellou P, Vigne E, Le Gal La Salle G, Robert JJ, Perricaudet M,Mallet J 1996 Gene transfer into the central and peripheral nervous systemusing adenoviral vectors. In: Lowenstein PR, Enquist LW (eds) Protocols forGene Transfer in Neuroscience: Towards Gene Therapy of Neurological Dis-orders. John Wiley & Sons, Chichester, pp 81–92
17. Messerle M, Buhler B, Keil GM, Koszinowski UH 1992 Structural organi-zation, expression, and functional characterization of the murine cytomega-lovirus immediate-early gene 3. J Virol 66:27–36
18. Dion LD, Fang J, Garver Jr RI 1996 Supernatant rescue assay vs. polymerasechain reaction for detection of wild type adenovirus-contaminating recombi-nant adenoviral stocks. J Virol Methods 56:99–107
3504 IN VIVO TRANSCRIPTIONAL TARGETING Endo • 2000Vol 141 • No 9
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022

19. Cotten M, Baker A, Saltik M, Wagner E, Buschle M 1994 Lipopolysaccharideis a frequent contaminant of plasmid DNA preparations and can be toxic toprimary human cells in the presence of adenovirus. Gene Ther 1:239–246
20. Larregina A., Morelli A, Dewey R, Castro MG, Fontana F, Lowenstein PR 1998Fas L induces Fas/Apo1-mediated apoptosis in human embryonic kidney 293cells routinely used to generate E1 deleted adenoviral vectors. Gene Ther 5:563–568
21. Morelli A, Larregina A, Smith J, Dewey R, Southgate T, Fontana A, CastroMG, Lowenstein PR 1999 Neuronal and glial cell type-specific promoterswithin adenovirus recombinants restrict the expression of the apoptosis-inducing molecule Fas ligand to predetermined brain cell types, and abolishperipheral liver toxicity. J Gen Virol 80:571–583
22. Guo YP, Hendry JH, Morris ID, Davis JRE, Beardwell CG 1997 Cell prolif-eration and death in the irradiated pituitary gland and its modification bygrowth stimulants. Int J Radiat Oncol Biol Phys 38:175–181
23. White A, Smith H, Hoadley M, Dobson SH, Ratcliffe JG 1987 Clinical eval-uation of a two-site immunoradiometric assay for adrenocorticotrophin inunextracted human plasma using monoclonal antibodies. Clin Endocrinol(Oxf) 26:41–51
24. Stone D, David A, Bolognani F, Lowenstein PR, Castro MG 2000 Viral vectorsfor gene delivery and gene therapy within the endocrine system. J Endocrinol164:103–118
25. Addison CL, Hitt M, Kunsken D, Graham FL 1997 Comparison of humanversus murine cytomegalovirus immediate early gene promoters for transgeneexpression by adenoviral vectors. J Gen Virol 78:1653–1661
26. Gerdes CA, Castro MG, Lowenstein PR 2000 Stronger and sustained trans-gene expression from a recombinant adenoviral vector under the control of theMCMV promoter in vivo. J Endocrinol 164:P165
27. Thomas CE, Schiedner G, Kochanek S, Castro MG, Lowenstein PR 2000Peripheral infection with adenovirus causes unexpected long-term braininflammation in animals injected intracranially with first generation,but not with high-capacity adenovirus vectors: towards realistic long-termneurological gene therapy for chronic diseases. Proc Natl Acad Sci USA97:7482–7487
28. Frawley LS, Boockfor FR 1991 Mammosomatotropes: presence and functionsin normal and neoplastic pituitary tissue. Endocr Rev 12:337–355
IN VIVO TRANSCRIPTIONAL TARGETING 3505
Dow
nloaded from https://academ
ic.oup.com/endo/article/141/9/3493/2989200 by guest on 09 August 2022





























